
S-adenosylmethionine Inhibits Ubiquitin-Proteasome System In Vitro and on Rat Vascular Smooth Muscle...
15
1 S-adenosylmethionine Inhibits Ubiquitin-Proteasome System In Vitro And On Rat Vascular Smooth Muscle Cells *F. D’Anselmi 1 , A. Cucina 1 , G. Cavallaro 1 , M. Bizzarri 2 , R. A. Cavallaro 1 , A. Fuso 1 ; A. Cavallaro 1 , S. Scarpa 1 1 Department of Surgery “P.Valdoni”, University of Rome “La Sapienza” 2 Department of Experimental Medicine, University of Rome “La Sapienza”
Transcript of S-adenosylmethionine Inhibits Ubiquitin-Proteasome System In Vitro and on Rat Vascular Smooth Muscle...

1
S-adenosylmethionine Inhibits Ubiquitin-Proteasome System In Vitro And On Rat Vascular
Smooth Muscle Cells
*F. D’Anselmi1, A. Cucina
1, G. Cavallaro
1, M. Bizzarri
2, R. A. Cavallaro
1, A. Fuso
1; A.
Cavallaro1, S. Scarpa
1
1Department of Surgery “P.Valdoni”, University of Rome “La Sapienza”
2Department of Experimental Medicine, University of Rome “La Sapienza”

2
ABSTRACT
S-adenosylmethionine is a metabolite regulating many biological processes; S-adenosylmethionine
effect on ubiquitin-proteasome system (UPS) has not been studied yet. We investigated S-
adenosylmethionine effects on UPS activity both in vitro, by inhibitor screening assay, and in rat
vascular smooth muscle cells, by Western Blot of proteasomal targets. We found that S-
adenosylmethionine inhibited UPS activity.
Keywords: ubiquitin-proteasome system, 20S proteasome, S-adenosylmethionine, vascular smooth
muscle cells, atherosclerosis, Sulfur-containing aminoacids cycle.
Abbreviations: UPS, ubiquitin-proteasome system; SAM, S-adenosylmethionine; Hcy,
Homocysteine; rVSMCs, rat vascular smooth muscle cells; SCAC, Sulfur-containing aminoacids
cycle; MTHFR, Methylenetetrahydrofolate reductase; CBS, Cystationine beta synthase; SAH, S-
adenosylhomocysteine; GSH, Glutathione; DMEM, Dulbecco’s Modified Eagle’s Medium; FCS,
Foetal Calf Serum; MS, Methionine synthase; THF, Tetrahydrofolate; MET, Methionine; ATP,
Adenosine triphosphate; MAT, Methionine Adenosyl transferase; CYS, cysteine.

3
INTRODUCTION
VSMCs (vascular smooth muscle cells) hyperproliferation is a key process in atherosclerosis and
restenosis [1]. The ubiquitin-proteasome system (UPS) is the major intracellular protein degradation
pathway and it accounts for the degradation of 80-90% of all intracellular proteins [2]. In the UPS
pathway, proteins are first covalently tagged with poly-ubiquitin chains and then are degraded by
the 26S proteasome. The 26S proteasome is a multicatalytic protease system that consists of a 20S
catalytic core and two 19S regulatory complexes [3]. The 20S proteolytic core cleaves ubiquitinated
substrates by three different proteolytic activities: caspase-like, trypsin like and chymotripsin-like
activities .
Inhibition of UPS leads to antiproliferative and proapoptotic effects in rat VSMCs (rVSMCs) in
vitro, as well as on atherosclerotic plaques formation and on vascular restenosis in vivo [4-7].
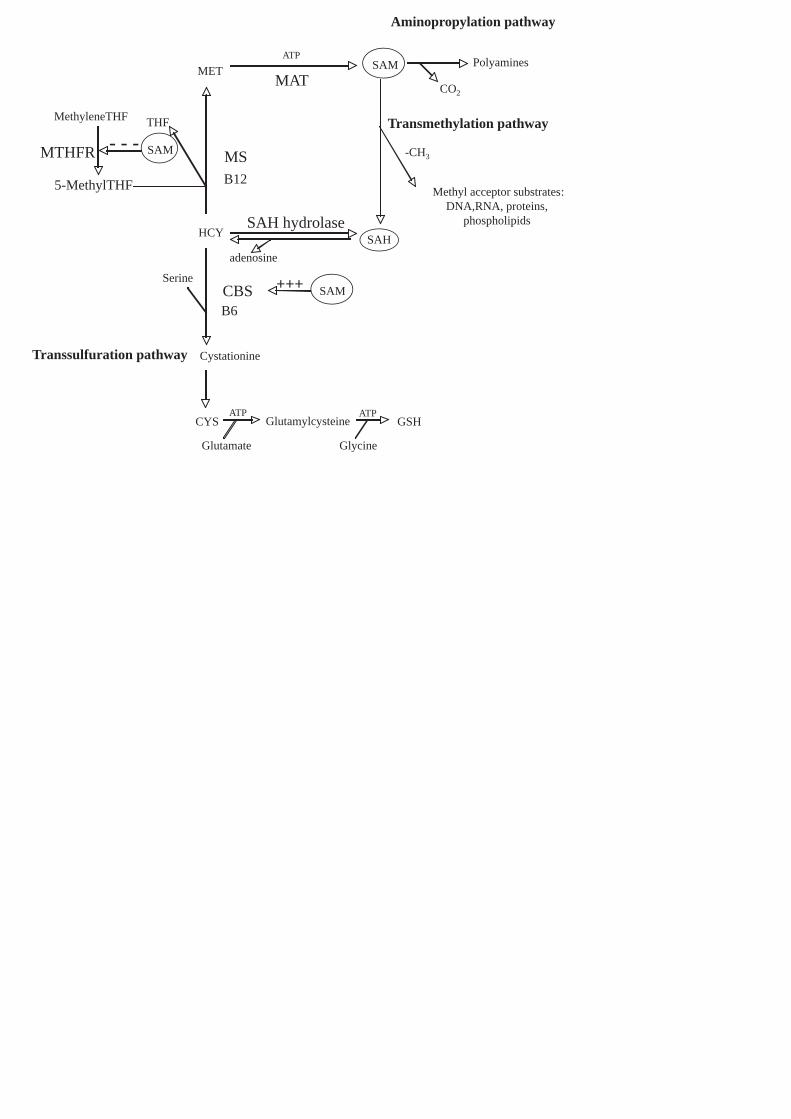
VSMCs hyperproliferation may be caused by the deregulation of Sulfur-containing aminoacids
cycle (SCAC), the metabolic pathway depicted in Figure (1) [8]. This deregulation causes the
increase of the aminoacid homocysteine (Hcy) in the cell and then in the plasma. This condition,
also known as hyperhomocysteinemia, is the major clinical marker of SCAC metabolic
dysfunctions and it is a risk factor for atherosclerosis. Both genetic and nutritional factors may
account for abnormalities in SCAC: mutations in Methylenetetrahydrofolate reductase (MTHFR)
and Cystationine beta synthase (CBS) genes are the most frequent genetic causes of Hcy
accumulation; on the other hand the clearance of Hcy depends on folates, vitamins B12 and B6, and
on S-adenosyl-L-methionine (SAM) bioavailability.
SAM is the key regulator of SCAC: it plays a pivotal role in transmethylation, aminopropylation
and transsulfuration pathways [9]. Through transmethylation reactions, SAM is converted to S-
adenosylhomocysteine (SAH), donating the methyl group to a large variety of substrates including
DNA, RNA, proteins and phospholipids. The transfer of methyl group is catalyzed by substrate-
specific methyltransferases, interacting with SAM through a specific domain. The transmethylation
pathway may regulate many cellular processes, from gene silencing [10] to membrane fluidity [11].

4
In the aminopropylation pathway, SAM undergoes decarboxylation to produce polyamines,
fundamental molecules for normal cell growth. In transsulfuration pathway, the Sulfur atom of Hcy
is converted to cysteine and finally to glutathione (GSH), the major cellular antioxidant [12]. In this
pathway, SAM strongly enhances Hcy clearance: in fact SAM acts as allosteric activator of CBS,
increasing its activity about three fold [13,14]. In normal conditions, SAM controls its own
biosynthesis: low SAM concentrations result in low CBS activity, thereby addressing Hcy to
transmethylation pathway toward SAM formation; on the contrary, high SAM concentrations allow
the clearance of Hcy into the transsulfuration pathway, leading to cysteine and GSH biosynthesis
[13]. As shown above, SAM is a pleiotropic molecule regulating many biological processes within
SCAC; nevertheless, in spite of the well-known involvement of SCAC deregulation and UPS
activity in atherosclerosis, SAM effect on UPS has not been investigated yet. In this work we tested
SAM effect on UPS activity in VSMCs.
MATERIALS AND METHODS
Reagents
SAM was prepared as 50 mMol/L stock solution in sterile tetradistilled water from a pharmaceutical
preparation containing SAM 1,4-butanedisulfonate salt. Antibodies against ubiquitin (sc-8017), c-
myc (sc-40), p53 (sc-6243), were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Monoclonal antibody against tubulin (clone B-5-1-2) was obtained from Sigma-Aldrich (St.
Louis, MO, USA).
Proteasome Activity Assay
The proteasome activity was determined using the 20S proteasome activity assay kit from
CHEMICON (Temecula, CA, USA). This assay is based on detection of the fluorophore 7-Amino-
4-methylcoumarin (AMC) after cleavage from the labelled substrate LLVY-AMC. This substrate is
specific to measure the chymotrypsin-like activity of 20S proteasome. To test proteasome activity
in an in vitro inhibitor screening we used purified proteasome enzyme to draw the activity curve of

5
20S proteasome alone and with 100 μMol/L and 500 μMol/L of SAM. To test proteasome activity
on VSMCs, cells were lysed by incubation in lysis buffer without protease inhibitors (50mM
HEPES, 5mM CHAPS, 0.5 mM EDTA, 0.035% SDS, pH 7.5) for an hour on ice. After
centrifugation (14,000 rpm for 10 minutes) supernatants were isolated and protein concentration
was determined using Bradford assay. After incubation for 120 minutes at 37°C, the fluorescence
of free AMC dye was monitored at an excitation wavelength of 380 nm and emission of 460 nm on
a fluorescence spectrometer LS 50B (Perkin Elmer, MA, USA).
Cell Culture
Primary rVSMCs were obtained from thoracic aorta of adult Lewis rats by collagenase type II
digestion and cultivated in Dulbecco’s Modified Eagle’s Medium (DMEM, Sigma-Aldrich, St.
Louis, MO, USA) supplemented with 10% of Foetal Calf Serum (FCS, Euroclone Wetherby, UK)
and antibiotics (Penicillin 100 IU/mL, Streptomycin 100 μg/mL, Gentamycin 200 μg/mL), all from
Sigma-Aldrich (St. Louis, MO, USA). Purity of rVSMCs was assessed to be 95% by
immunostaining with anti-smooth muscle -actin antibody (Sigma-Aldrich, St. Louis, MO, USA).
VSMCs were treated with SAM 100 μMol/L but not with SAM 500 μMol/L: the former
concentration resulted non-toxic, on the contrary the latter resulted toxic in previous viability
Trypan Blue tests (data not shown).
Preparation of Cellular Extracts and Western Blot Analysis
Cytoplasmic extracts were prepared as described elsewhere [10]. Cellular extracts were separated
on SDS-PAGE gels with a concentration of acrylamide specific for the protein studied. Proteins
were blotted onto nitrocellulose membranes (BIO-RAD, Bio-Rad Laboratories, Hercules, CA,
USA), probed with the specific antibodies and detected with enhanced chemoluminescence (ECL)
kit from Amersham (Amersham Biosciences, Little Chalfont Buckingamshire, England).
Statistics
Data are expressed as mean + SD of at least three independent experiments. Differences between
experimental conditions were determined by ANOVA (Analysis of Variance) test with Bonferroni

6
post test and by Student’s T test. An error probability of p<0.05 was considered as significant. The
GraphPad InStat software (GraphPad Software, Inc. San Diego, CA, USA) was used for statistical
calculations.
RESULTS
Specificity of 20S Proteasome Inhibition
In an in vitro inhibitor-screening assay, performed on purified proteasome specimens (20S enzyme),
we found that SAM inhibited chymotrypsin-like proteasome activity by 11,4% (p<0,01) with SAM
100 μMol/L and 37% (p<0,001) with SAM 500 μMol/L, with respect to control (Figure 2).
Experiment done with another preparation containing SAM p-toluenesulfonate salt gave similar
results (data not shown).
Specificity of Proteasome Inhibition in Cell Extracts
In protein extracts from rVSMCs treated for one day with SAM 100 μMol/L we observed a
significant inhibition (29,6%, p<0,0001) of the chymotrypsin-like proteasome activity (Figure 3). In
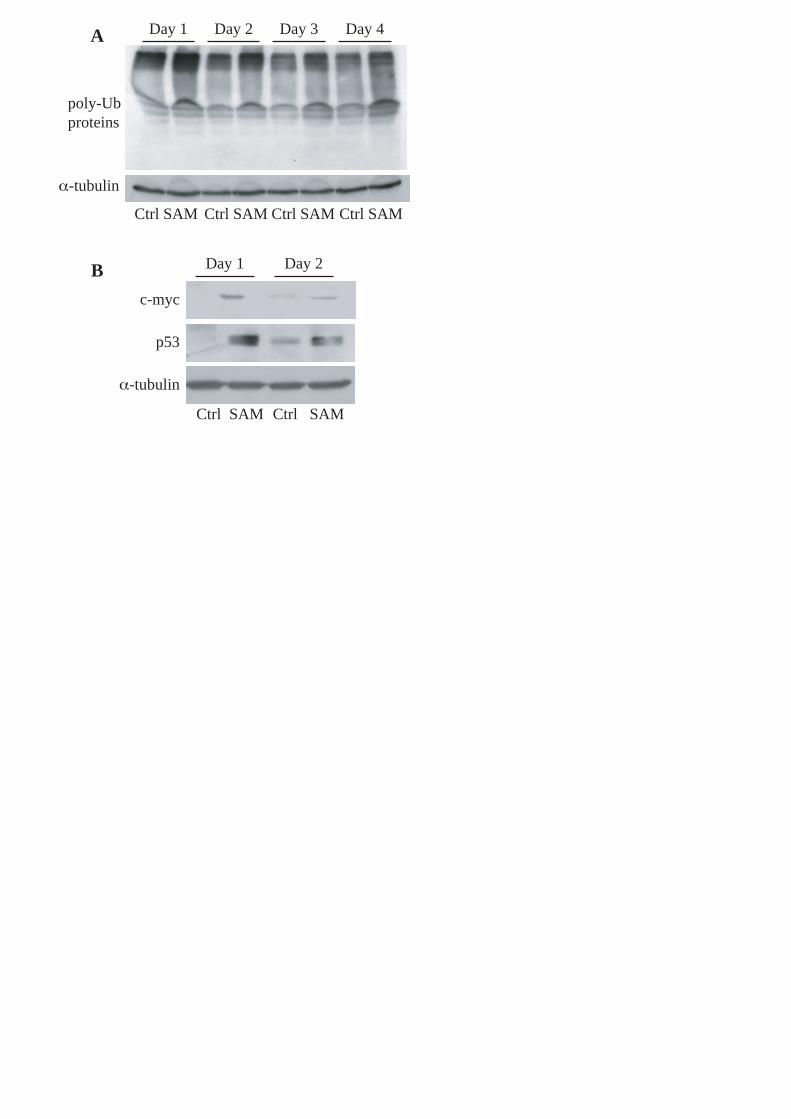
rVSMCs, SAM 100 μMol/L caused the accumulation of poly-ubiquitinated proteins (Figure 4A)
and of specific substrates of proteasome such as c-myc and p53 (Figure 4B).
DISCUSSION
Since 2002 the UPS hyperactivity has been linked to VSMCs hyperproliferation and the UPS has
been considered a target in the treatment of atherosclerosis and restenosis [2,4,5]. However, recent
clinical study supported the existence of a connection between proteasome inhibition, oxidative
stress and plaque instability in atherosclerosis [15]. Therefore the role of UPS activity in
atherosclerosis and specifically in VSMCs metabolism still remains to be clarified. The metabolic
deregulation of SCAC may lead to hyperhomocysteinemia, a pathological condition which
contributes to the pathogenesis of atherosclerosis through a combination of multiple damaging
effects on the arterial wall and particularly through the VSMCs hyperproliferation [8,16,17].

7
SAM is a pivotal regulator of SCAC. In vitro SAM inhibited the chymotrypsin-like activity of
purified proteasome enzyme 20S in an inhibitor-screening assay (Figure 2). Further studies are in
progress in order to clarify the mechanism of inhibition: particularly it remains to be verified if
SAM inhibits the UPS through its methylating activities or as an allosteric inhibitor. In fact in our
test no methyltransferases were present in the reaction mixture, but spontaneous methyl-group
transfer from SAM to a protein may happen [18]; on the other hand SAM is already known to
regulate enzymatic activities as an allosteric activator or inhibitor [13,19].
SAM inhibitory effect was further confirmed in rVSMC protein extracts where the chymotrypsin-
like activity of proteasome was downregulated (Figure 3). This inhibition may be the sum of direct
inhibitory effect on proteasome and other pleiotropic effects exerted by SAM. The decrease of UPS
activity has been of 29,6% in rVSMCs treated with SAM 100μM; this concentration [10,20] and
even higher [21] concentrations are routinely used in cell culture studies to investigate SAM
properties.
A recent review classifies the proteasome inhibitors according to their cellular effects: they may be
defined as poisons, which induce apoptosis, or as remedies, which modulate cellular functions and
protect cell from death; moreover, this review postulates that the distinction of a poison from a
remedy depends on cell type and on the degree of UPS inhibition [3]. The degree of UPS inhibition
obtained with SAM in rVSMCs is in agreement with the concept that a strong and chronic
inhibition of proteasome activity could not be beneficial in atherosclerosis [15,22].
It is well-known that a specific feature of proteasome inhibition is the accumulation of poly-
ubiquitinated proteins [4]. For the first time we demonstrated that SAM induced the accumulation
of poly-ubiquitinated proteins (Figure 4A), and increased the intracellular levels of c-myc and p53
(Figure 4B), two substrates of UPS. These data confirm the inhibition of proteasome measured by
20S activity assay kit.
In according to our results, we can state that SAM inhibited the chymotrypsin-like activity of UPS
in rVSMCs. This finding may be the start point for future investigations aimed to clarify the role of

8
SAM and other molecules involved in SCAC in VSMCs hyperproliferation and on UPS activity
during the neointimal hyperplasia.
ACKNOWLEDGMENTS
This work was supported by University of Rome “La Sapienza”. We thank to Dr. Gabriella
Dobrowolny for the excellent scientific and technical suggestions and Dr. Luisa Di Renzo for
reviewing the English in the manuscript.
REFERENCES
[1] Dzau, V.J., Braun-Dullaeus, R.C., Sedding, D.G. (2002) Nat Med, 8:1249-1256.
[2] Herrmann, J., Ciechanover, A., Lerman, L.O., Lerman, A. (2005) Int J Cardiovasc
Intervent., 7:5-13.
[3] Meiners, S., Ludwig, A., Stangl, V., Stangl, K (2007) Med Res Rev. [Epub ahead of
print]
[4] Meiners, S., Laule, M., Rother, W., Guenther, C., Prauka, I., Muschick, P., Baumann,
G., Kloetzel, P.M., Stangl, K. (2002) Circulation, 105:483-489.
[5] Herrmann, J., Ciechanover, A., Lerman, L.O., Lerman, A. (2004) Cardiovasc Res.,
61:11-21.
[6] Herrmann, J., Lerman, L.O., Mukhopadhyay, D., Napoli, C., Lerman, A. (2006)
Arterioscler Thromb Vasc Biol.,26:1948-1957.
[7] Marfella, R., D'Amico, M., Esposito, K., Baldi, A., Di Filippo, C., Siniscalchi, M.,
Sasso, F.C., Portoghese, M., Cirillo, F., Cacciapuoti, F., Carbonara, O., Crescenzi, B.,

9
Baldi, F., Ceriello, A., Nicoletti, G.F., D'Andrea, F., Verza, M., Coppola, L., Rossi, F.,
Giugliano, D. (2006) Diabetes., 55:622-632.
[8] Tsai, J.C., Perrella, M.A., Yoshizumi, M., Hsieh, C.M., Haber, E., Schlegel, R., Lee,
M.E. (1994) Proc Natl Acad Sci U S A, 91:6369-6373.
[9] Bottiglieri, T. (2002) Am J Clin Nutr., 76:1151S-1157S.
[10] Scarpa, S., Fuso, A., D'Anselmi, F., Cavallaro, R.A.(2003) FEBS Lett., 541:145-148.
[11] Turchetti, V., Bellini, M.A., Leoncini, F., Petri, F., Trabalzini, L., Guerrini, M., Forconi,
S. (2000) Clin Hemorheol Microcirc., 22:215-221.
[12] Lu, S.C. (2000) Int J Biochem Cell Biol., 32:391-395.
[13] Ignoul, S., Eggermont, J. (2005) Am J Physiol Cell Physiol., 289:C1369-C1378.
[14] Finkelstein, J.D., Kyle, W.E., Martin, J.L., Pick, A.M. (1975) Biochem Biophys Res
Commun., 66:81-87.
[15] Versari, D., Herrmann, J., Gossl, M., Mannheim, D., Sattler, K., Meyer, F.B., Lerman,
L.O., Lerman, A. (2006) Arterioscler Thromb Vasc Biol., 26:2132-2139.
[16] Tang, L., Mamotte, C.D., Van Bockxmeer, F.M., Taylor, R.R. (1998) Atherosclerosis,
136:169-173.
[17] Carmody, B.J., Arora, S., Avena, R., Cosby, K., Sidawy, A.N. (1999) J Vasc Surg.,
30:1121-1128.
[18] Kimzey, A.L., McFadden, P.N. (1994) J Protein Chem.,13(6):537-46
[19] Selhub, J. (1999) Annu Rev Nutr.;19:217-246.

10
[20] Fuso, A., Seminara, L., Cavallaro, R.A., D'Anselmi, F., Scarpa, S. (2006) Mol Cell
Neurosci. Aug;32(4):419.
[21] Ou, X., Yang, H., Ramani, K., Ara, A.I., Chen,H., Mato, J.M., Lu, S.C. (2007) Biochem
J. Jan 1;401(1):87-96.
[22] Herrmann, J., Saguner, A.M., Versari, D., Peterson, T.E., Chade, A., Olson, M.,
Lerman, L.O., Lerman, A. (2007) Circ Res., [Epub ahead of print]
FIGURE LEGENDS
Figure 1. Biosynthesis and metabolic pathways of SAM in the SCAC.
SAM increases (+++) CBS activity and decreases (---) MTHFR activity in allosteric manner.
MTHFR (Methylenetetrahydrofolate reductase), MS (Methionine synthase), THF
(Tetrahydrofolate), MET (Methionine), ATP (Adenosine triphosphate), MAT (Methionine
Adenosyl transferase), SAH (S-adenosylhomocysteine), HCY (Homocysteine), CBS (Cystationine
beta synthase), CYS (cysteine), GSH (glutathione), B12 and B6 (vitamins B12 and B6).
Figure 2. 20S proteasome chymotrypsin-like activity in an in vitro inhibitor screening assay.
Experimental conditions were: 20S proteasome as control (Ctrl); 20S proteasome plus 100 μMol/L
(SAM100) or 500 μMol/L (SAM 500) of SAM. The average fluorescence in control was counted as
100%. Inhibition was expressed as a percentage of the normalized value to control (n=6, *p<0,01
SAM100 vs ctrl; **p<0,001 SAM 500 vs ctrl and vs SAM100 by ANOVA and Bonferroni post-
test).
Figure 3. Proteasome chymotrypsin-like activity in rVSMCs. Cells were kept after one day of
culture in DMEM 10%FCS (Ctrl) and in DMEM 10%FCS with 100 μMol/L of SAM (SAM 100).
The average fluorescence in control was counted as 100%. Inhibition was expressed as a percentage
of the normalized value to control (n=6, *p<0,0001 SAM100 vs ctrl by Student’s T test).

11
Figure 4. Western blot of poly-ubiquitinated proteins and of specific proteasomal targets. A,
Representative western blot of accumulated poly-ubiquitinated proteins and housekeeping gene -
tubulin in rVSMCs cultured in DMEM 10%FCS (Ctrl) and in DMEM 10%FCS with 100 μMol/L of
SAM (SAM) for four days. B, Western blot of c-myc, p53 and housekeeping gene -tubulin in
rVSMCs cultured for two days in DMEM 10%FCS (Ctrl) and in DMEM 10%FCS with 100
μMol/L of SAM (SAM).
Aminopropylation pathway
Polyamines
CO2
Transmethylation pathway
Transsulfuration pathway
B6
B12
CBS SAM+++
ATP
HCY
MET
Cystationine
CYS
SAM
SAH
MS
Methyl acceptor substrates:
DNA,RNA, proteins,
phospholipids
-CH3
MethyleneTHF
5-MethylTHF
MTHFR
THF
Serine
Glutamylcysteine GSH
Glutamate Glycine
ATP ATP
MAT
SAH hydrolase
adenosine
SAM- - -

0
20
40
60
80
100
120
ctrl SAM 100 SAM 500
*
**

*
0
20
40
60
80
100
120
ctrl SAM 100
Day 1 Day 4Day 3Day 2
Day 2Day 1
Ctrl SAM Ctrl SAMCtrl SAMCtrl SAM
Ctrl SAM Ctrl SAM
poly-Ub
proteins
α-tubulin
c-myc
α-tubulin
p53
A
B




















