
l(+)Lactic acid production from starch by a novel amylolytic Lactococcus lactis subsp. lactis B84
8
L(+)-Lactic acid production from starch by a novel amylolytic Lactococcus lactis subsp. lactis B84 Kaloyan Petrov a , Zoltan Urshev b , Penka Petrova c, a Institute of Chemical Engineering, Bulgarian Academy of Sciences, 103 Acad. G. Bonchev Street, 1113 Sofia, Bulgaria b LB Bulgaricum Plc., R&D Center, 12A Malashevska Street, Sofia 1202, Bulgaria c Institute of Microbiology, Bulgarian Academy of Sciences, 26 Acad. G. Bonchev Street,1113 Sofia, Bulgaria article info Article history: Received 15 August 2007 Received in revised form 22 February 2008 Accepted 23 February 2008 Available online 29 February 2008 Keywords: Lactococcus lactis subsp. lactis Lactic acid production Starch hydrolysis Amylase activity abstract A new Lactococcus lactis subsp. lactis B84, capable of utilizing starch as a sole carbon source and producing L(+)-lactate, was isolated from spontaneously fermented rye sourdough. Aiming at maximum lactic acid productivity, the components of the media and the cultivation conditions were varied. In MRS-starch medium (with absence of yeast and meat extracts), at 33 1C, agitation 200 rpm and pH 6.0 for 6 days complete starch hydrolysis occurred and 5.5 g l 1 lactic acid were produced from 18 g l 1 starch. The identification of strain B84 was based on genetic criteria. Amplified ribosomal DNA restriction analysis (ARDRA), PCR with species-specific primers and sequencing of the 16S rDNA proved its species affiliation. Four genes for enzymes, involved in starch degradation were detected in B84 genome: amyL, amyY, glgP and apu, coding cytoplasmic and extracellular a-amylases, glycogen phosphorylase and amylopullulanase, respectively. Reverse transcription PCR experiments showed that both genes, encoding a-amylases (amyL and amyY) were expressed into mRNAs, whereas apu and glgP were not. Amylase activity assay was performed at different pH and temperatures. The cell-bond amylase proved to be the key enzyme, involved in the starch hydrolysis with maximum activity at 45 1C and pH 5.4. & 2008 Elsevier Ltd. All rights reserved. 1. Introduction Lactic acid is a valuable product with many practical applica- tions: as a preservative, pH regulator and taste-enhancer in food industry, for implants and suture in the medical practice, as a reagent for polylactic and polyacrylic acids synthesis for biode- gradable polymers (Wee et al., 2006). Many efforts have been made to develop biotechnologies for lactate generation from such a cheap, abundant and renewable substrate as the starch. The main problem in starch utilization by LAB, however, is the starch saccharification—a step that could be performed separately prior to fermentation or in parallel with the LA synthesis (Hofvendahl et al., 1999). One-step process of lactic acid production from starch was achieved by the use of LAB, possessing extracellular amylase activity, such as Lactobacillus plantarum A6 (Giraud et al., 1991), Lactobacillus manihotivorans (Morlon-Guyot et al., 1998), Lactobacillus amylophilus (Vishnu et al., 2002), Lactobacillus amylovorus (Zhang and Cheyran, 1991) and Streptococcus bovis (Narita et al., 2004). Lactococcus lactis is quite desirable for LA industry, because it is homofermentative, secretes only L(+)-lactic acid and is highly productive (Hofvendahl and Hahn-Ha ¨gerdal, 1997), but at present its use in starch fermentations is restrained by the absence of amylolytic properties. Most usual overcoming of this problem is the preliminary treatment of the starchy material with a-amylase/ amyloglucosidase mixture (Hofvendahl et al., 1999) or hydrolysis by commercial amylolytic enzyme products (Nolasco-Hipolito et al., 2002; A ˚ kerberg et al., 1998). Other approach is co-cultivation of lactococci together with amylolytic strains, such as Aspergillus awamori (Roble et al., 2003). In fact, this idea was ‘‘borrowed’’ from the natural microbial communities in the starchy foods, in which the metabolites, produced by the starch-hydrolyzing Bacillus spp. may create enough alternative energy sources for LAB (Østergaard et al., 1998). Third solution of the problem is the direct improve- ment of the Lactococcus strain by transformation with vectors, bearing cloned a-amylase gene (Satoh et al., 1997; Okano et al., 2007). Therefore, the isolation of natural lactococci, able to utilize the starch directly for LA production is of special interest. L. lactis is one of the most used lactic acid bacterium by the dairy industry. It participates in the manufacture of fresh and ARTICLE IN PRESS Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/fm Food Microbiology 0740-0020/$ - see front matter & 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.fm.2008.02.005 Abbreviations: ARDRA, amplified ribosomal DNA restriction analysis; LA, lactic acid; LAB, lactic acid bacteria; RT-PCR, reverse transcription polymerase chain reaction. Corresponding author. Tel.: +359 2 9793182; fax: +35928700109. E-mail address: [email protected] (P. Petrova). Food Microbiology 25 (2008) 550– 557
Transcript of l(+)Lactic acid production from starch by a novel amylolytic Lactococcus lactis subsp. lactis B84

ARTICLE IN PRESS
Food Microbiology 25 (2008) 550– 557
Contents lists available at ScienceDirect
Food Microbiology
0740-00
doi:10.1
Abbre
acid; LA
reaction� Corr
E-m
journal homepage: www.elsevier.com/locate/fm
L(+)-Lactic acid production from starch by a novel amylolyticLactococcus lactis subsp. lactis B84
Kaloyan Petrov a, Zoltan Urshev b, Penka Petrova c,�
a Institute of Chemical Engineering, Bulgarian Academy of Sciences, 103 Acad. G. Bonchev Street, 1113 Sofia, Bulgariab LB Bulgaricum Plc., R&D Center, 12A Malashevska Street, Sofia 1202, Bulgariac Institute of Microbiology, Bulgarian Academy of Sciences, 26 Acad. G. Bonchev Street, 1113 Sofia, Bulgaria
a r t i c l e i n f o
Article history:
Received 15 August 2007
Received in revised form
22 February 2008
Accepted 23 February 2008Available online 29 February 2008
Keywords:
Lactococcus lactis subsp. lactis
Lactic acid production
Starch hydrolysis
Amylase activity
20/$ - see front matter & 2008 Elsevier Ltd. A
016/j.fm.2008.02.005
viations: ARDRA, amplified ribosomal DNA r
B, lactic acid bacteria; RT-PCR, reverse transc
.
esponding author. Tel.: +359 2 9793182; fax:
ail address: [email protected] (P. Petro
a b s t r a c t
A new Lactococcus lactis subsp. lactis B84, capable of utilizing starch as a sole carbon source and
producing L(+)-lactate, was isolated from spontaneously fermented rye sourdough. Aiming at maximum
lactic acid productivity, the components of the media and the cultivation conditions were varied. In
MRS-starch medium (with absence of yeast and meat extracts), at 33 1C, agitation 200 rpm and pH 6.0
for 6 days complete starch hydrolysis occurred and 5.5 g l�1 lactic acid were produced from 18 g l�1
starch.
The identification of strain B84 was based on genetic criteria. Amplified ribosomal DNA restriction
analysis (ARDRA), PCR with species-specific primers and sequencing of the 16S rDNA proved its species
affiliation.
Four genes for enzymes, involved in starch degradation were detected in B84 genome: amyL, amyY,
glgP and apu, coding cytoplasmic and extracellular a-amylases, glycogen phosphorylase and
amylopullulanase, respectively. Reverse transcription PCR experiments showed that both genes,
encoding a-amylases (amyL and amyY) were expressed into mRNAs, whereas apu and glgP were not.
Amylase activity assay was performed at different pH and temperatures. The cell-bond amylase proved
to be the key enzyme, involved in the starch hydrolysis with maximum activity at 45 1C and pH 5.4.
& 2008 Elsevier Ltd. All rights reserved.
1. Introduction
Lactic acid is a valuable product with many practical applica-tions: as a preservative, pH regulator and taste-enhancer in foodindustry, for implants and suture in the medical practice, as areagent for polylactic and polyacrylic acids synthesis for biode-gradable polymers (Wee et al., 2006). Many efforts have beenmade to develop biotechnologies for lactate generation from sucha cheap, abundant and renewable substrate as the starch. Themain problem in starch utilization by LAB, however, is the starchsaccharification—a step that could be performed separately priorto fermentation or in parallel with the LA synthesis (Hofvendahlet al., 1999).
One-step process of lactic acid production from starch wasachieved by the use of LAB, possessing extracellular amylaseactivity, such as Lactobacillus plantarum A6 (Giraud et al.,1991), Lactobacillus manihotivorans (Morlon-Guyot et al., 1998),
ll rights reserved.
estriction analysis; LA, lactic
ription polymerase chain
+359 2 8700109.
va).
Lactobacillus amylophilus (Vishnu et al., 2002), Lactobacillus amylovorus
(Zhang and Cheyran, 1991) and Streptococcus bovis (Narita et al.,2004). Lactococcus lactis is quite desirable for LA industry, because it ishomofermentative, secretes only L(+)-lactic acid and is highlyproductive (Hofvendahl and Hahn-Hagerdal, 1997), but at presentits use in starch fermentations is restrained by the absence ofamylolytic properties. Most usual overcoming of this problem is thepreliminary treatment of the starchy material with a-amylase/amyloglucosidase mixture (Hofvendahl et al., 1999) or hydrolysis bycommercial amylolytic enzyme products (Nolasco-Hipolitoet al., 2002; Akerberg et al., 1998). Other approach is co-cultivationof lactococci together with amylolytic strains, such as Aspergillus
awamori (Roble et al., 2003). In fact, this idea was ‘‘borrowed’’ fromthe natural microbial communities in the starchy foods, in whichthe metabolites, produced by the starch-hydrolyzing Bacillus spp.may create enough alternative energy sources for LAB (Østergaardet al., 1998). Third solution of the problem is the direct improve-ment of the Lactococcus strain by transformation with vectors,bearing cloned a-amylase gene (Satoh et al., 1997; Okano et al.,2007). Therefore, the isolation of natural lactococci, able to utilizethe starch directly for LA production is of special interest.
L. lactis is one of the most used lactic acid bacterium by thedairy industry. It participates in the manufacture of fresh and

ARTICLE IN PRESS
K. Petrov et al. / Food Microbiology 25 (2008) 550–557 551
semi-hard cheeses, cream, butter and sour milks, giving thespecific taste and texture of the products (Samarzija et al., 2001).Widespread in fermented foods, it could be isolated also fromcertain sources, containing starch: fermented cereal beverages,such as maize pozol, non-alcoholic drink bushera and sorghumbeer (Dıaz-Ruiz et al., 2003; Muyanja et al., 2003; Sawadogo-Lingani et al., 2007); sorghum, maize, or wheat sourdough(Hamad et al., 1997; Rocha and Malcata, 1999; Corsetti et al.,2001), cooked fish products and the starchy meal poi, made fromthe taro plant (Østergaard et al., 1998; Brown and Valiere, 2004).In spite of the starch abundance in these inhabitant niches, veryfew of the isolated lactococci were found to possess amylolyticproperties (Østergaard et al., 1998; Dıaz-Ruiz et al., 2003;Muyanja et al., 2003).
On the other hand, the genes encoding several putativeamylolytic enzymes (amyL, amyY, apu, glgP) were found in thefully sequenced genome of L. lactis subsp. lactis IL 1403 (Bolotinet al., 2001). Although it has been proven that this strain wasunable to produce amylase enzymes (Satoh et al., 1997), the fullsequence enables the investigation of the genetic basis of thelactococcal amylolytic activity in general.
In the present study, we report the isolation of a natural strainL. lactis subsp. lactis B84, which is capable to accomplish fullstarch saccharification and to produce lactic acid. The purpose ofthis work is to study the gene expression of the key enzymes,responsible for the starch hydrolysis by Lactococcus, usingthe methods of reverse transcription polymerase chain reaction(RT-PCR).
2. Materials and methods
2.1. Bacterial strains, media and cultivation conditions
L. lactis subsp. lactis B84 was isolated from spontaneouslyfermenting sourdough, prepared from rye flour (Balvan mill,Bulgaria). It was deposited in the Bulgarian National Collection forMicroorganisms and Cell Cultures (http://www.nbimcc.org/en/about.htm), under registration no. 8618. The strain was propa-gated in MRS medium (Scharlau, Spain) and maintained at 4 1C(subcultured every week), or frozen at �80 1C with 15% (w/w)glycerol added.
Reference strains were supplied from the Belgian Collection ofMicroorganisms (LMG) and the German Collection of Microorgan-isms and Cell Cultures (DSMZ): Enterococcus faecium LMG 11423,Enterococcus faecalis LMG 2937, Enterococcus durans LMG 10746,Leuconostoc mesenteroides subsp. mesenteroides LMG 6893, Leuco-
nostoc lactis LMG 8894, Streptococcus thermophilus DSM 20617,L. lactis subsp. cremoris DSM 20069; L. lactis subsp. lactis DSM20481. Strain L. lactis subsp. lactis IL 1403 was provided byT. Sasaki (Meiji Dairies Co., Japan) with the kind permission ofA. Chopin (Laboratoire de Genetique Microbienne, INRA, France).The reference strains were cultivated in M17 broth (Merck) orGM17 medium (M17, supplemented with 2% glucose), at optimaltemperatures (30 or 37 1C) for 48 h.
2.2. Phenotypic and biochemical characteristics
The strain L. lactis subsp. lactis B84 was observed for its cellmorphology by immersed microscopy and Gram-reaction by theKOH method (Gregersen, 1978). Catalase activity was tested using20% H2O2. Quantitatively L,D-lactic acid isomers were determinedusing a commercial enzymatic kit (Roche Diagnostics GmbH,Mannheim, Germany).
2.3. Starch fermentation by L. lactis subsp. lactis B84
A number of nutrient media were prepared and the influenceof every constituent on the lactic acid production and the cellgrowth was estimated. Batch cultures were grown in derivativesof Elliker, MRS (Elliker et al., 1956; de Man et al., 1960) and M17(Oxoid) media with soluble potato starch (Fluka) as a sole carbonsource. Three of the fermentation media were found as mostuseful: M1 had the MRS content, with 20 g l�1 starch instead ofglucose; M2 medium was M1 without the meat and yeastextracts; M2A contained the components of M2, where the meatpeptone was substituted with peptone from casein (Scharlau).
The kinetic experiments without pH control were performed in500-ml flasks, containing 200 ml medium, inoculated with 2 ml of24-h precultures, grown in M1 broth, at 33 1C and vigorousagitation (200 rpm), using rotary shaker, New Brunswick Scien-tific, USA.
Experiments with pH control were carried out in a ‘‘NewBrunswick Scientific—BIOFLO’’ fermentor with magnetic stirrerand working volume 450 ml, at 33 1C, stirring at 200 rpm. The pHadjustment was carried out using 5 M NaOH.
Mean values from three independent batches were shown.
2.4. Analytical methods
Cell growth was monitored by a measurement of opticaldensity of the broth (OD620), using a spectrophotometer (UV/visSpectrophotometer HElIOS b, UNICOM). Cell dry weight wascalculated from the optical density using calibration curve for thestrain.
The quantification of glucose and lactic acid concentrationswas carried out after filtration of cultures through a membranefilter (0.45mm pore size, Boeco, Germany) by a high-performanceliquid chromatography (HPLC) system equipped with a refracto-metric detector (Perkin-Elmer chromatograph, series 10). A HPLCcolumn (Aminex HPX-87H, Bio-Rad, Richmond, CA, USA) was usedwith 5 mM sulfuric acid as the mobile phase at an elution flowrate of 0.6 ml min�1.
The residual starch content was determined by measuring thelight absorption of the iodine-starch complex color at wavelength580 nm (Nakamura, 1981).
2.5. Amylase activity assay
The amylase activity was detected by measurement of theiodine complexing ability of starch according the method ofGiraud et al. (1993) at different pH values (4.8–6.4; 0.1 mol l�1
citrate-phosphate buffer), and temperatures (33–55 1C). Theextracellular and the cell-bond enzymatic activities were deter-mined at the beginning of the stationary phase of the culture, asdescribed by Agati et al. (1998). One enzyme unit was defined asthe amount of enzyme hydrolyzing 10 mg starch in 30 min.
2.6. DNA and RNA isolations
Total genomic DNA was isolated from 48-h-old cells of L. lactis
subsp. lactis strains B84 and IL 1403, grown in MRS or M1 media,using GFX Genomic blood DNA purification kit (AmershamBiosciences), following manufacturer’s recommendations.
The pure total RNA from the same cultures was obtained from0.1 g harvested biomass, washed twice in sterile, RNase free bi-distilled water. The microscale procedure of InvisorbsRNA kit I(AJ Roboscreen GmbH) was used. The quantity and purity of theRNA was measured by spectrophotometer at wavelengths of 260and 280 nm. DNase (Fermentas) treatment was carried out under

ARTICLE IN PRESS
K. Petrov et al. / Food Microbiology 25 (2008) 550–557552
the following conditions: mixture of total RNA in 100ml RNase freewater, 20ml of 10� DNase buffer, 10 ml DNaseI (1 unit ml�1) and70ml deionized water were incubated at 37 1C for 1 h, followed byextraction with phenol/chloroform. RNA was then precipitated at�20 1C for 1 h using 0.1 vol. of 3 M Sodium acetate (pH 5.0) and2.5 vol. of 100% ethanol.
2.7. PCR amplification of 16S rRNA genes and ARDRA analysis
All polymerase chain reactions (PCR), presented here wereprepared with PuReTaqTM Ready to GoTM PCR beads (AmershamBiosciences), according to the manufacturer’s instructions, in atotal volume 50ml and final concentrations of primers 0.5 pmolml�1 (all purchased from MWG-Biotech, Germany. The PCRamplifications were done in a Progene termocycler (Techne, UK).
The ARDRA (amplified ribosomal DNA restriction analysis) wasperformed with primer pair SacI-POmod (50-CCGAGCTCAACA-GAGTTTGATCCTGGCTCAG-30) and SalI-T7-PC5 (50-GGTCGACCGT-TAATACGACTCACTATGGGATACCTTGTTACGACTTA-30), according toRodtong and Tannock (1993). One mililiter of 48-h cultures ofeach strain was harvested, the cells were washed and resuspendedin 100 ml in ddH2O; 1ml of the suspension was used as template.The following program was used: 1 cycle of 95 1C for 5 min (celllysis); 35 cycles of 93 1C for 1 min (denaturation), 53 1C for 1 min(annealing) and 72 1C for 30 s; final extension at 72 1C for 7 min.
The enzymatic digest of the amplified DNA was done using theendonuclease HhaI (New England BioLabs), in a final volume of30ml, at 37 1C. Restriction fragments received were separated in 3%agarose gel (Sigma), in Tris–borate buffer (0.089 M Tris, 0.089 Mboric acid, 0.002 M EDTA), and stained with Ethidium bromide(Sambrook and Russel, 2001).
The amplification of the 16S rRNA gene for the sequenceanalysis was performed with universal eubacterial primer pair:forward primer fD1: 50-AGAGTTTGATCCTGGCTCAG-30 and reverseprimer rD1: 50-AAGGAGGTGATCCAGCC-30 (Weisburg et al., 1991).The final volume of the template DNA was 1–2 ng ml�1, thetemperature profile was: 95 1C for 5 min, 35 cycles consisting of94 1C for 1 min, 55 1C for 1 min, 72 1C for 2 min, followed by finalelongation at 72 1C for 5 min. The 1.5 kb PCR products werevisualized in 1% agarose gel.
2.8. L. lactis (sub)species-specific PCR
Second approach for identification at sub-species level of thenewly isolated lactococcal strain was the application of themethod of Nomura et al. (2002), based on the length polymorph-ism of the PCR fragments, obtained with primer pair, targeting thegene, coding glutamate—decarboxylase. For the PCR amplificationwas used the primer pair gadB21: 50-CGTTATGGATTTGATGGATA-
Table 1Oligonucleotide primers, targeting the genes, putatively involved in the starch hydroly
Gene Primer Sequence (50–30) Annealing
temperature
amyL amyLF CCCTCAGATTCTCAGCACT
amyLR CATAAAATTCAGCTCCACCAT 58
amyY amyYF GTAACTTTACTGGCGACGT
amyYR CGGTGCTGTAAGCTGGAA 56
apu apuF CAACCTTTTGGGGCCATCAA
apuR CCACTAATCACTTTTTCTGTC 59
glgP glgPF GCAAGATTTCGAAGAGCGTC
glgPR GTCATCAGCATATTCCCTAAC 60
TAAAGC-30 and GAD7: 50-ACTCTTCTTAAGAACAAGTTTAACAGC-30.The same cell suspension as for the ARDRA analysis was applied astemplate. The PCR conditions were: 3 min at 95 1C (cell lysis anddenaturation), followed by 40 cycles: 30 s at 92 1C, 30 s at 45 1C,30 s at 72 1C and final elongation for 7 min at 72 1C. The amplifiedfragments were analyzed in 4.0% agarose gel, using 100 bp PCR-Low (Sigma) as a molecular sizes standard.
2.9. DNA sequencing and phylogenetic analysis
The 16S rDNA amplification products were purified using GFXPCR DNA and gel band purification kit (Amersham Biosciences).The nucleotide sequences were determined by MWG-BiotechCompany (Germany) using automated sequencer (Applied Bio-systems 3700) and the dye-deoxy termination procedure. Primerpair, used for the sequencing was the described above fD1/rD1.The sequence analysis was performed using Chromas and CAP3Sequence Assembly Programs (http://genome.cs.mtu.edu/cap).Sequence comparison was done using BLAST (Altschul et al.,1997) and Clustal-W programs (Thompson et al., 1994). Thecomplete 16S rRNA gene sequence of the strain L. lactis subsp.lactis B84, obtained in this study was deposited in the NCBIGenbank database under accession number EF589778.
2.10. PCR amplification of the genes, putatively involved in starch
hydrolysis by L. lactis subsp. lactis
The detection of genes, most possibly taking part in the starchhydrolysis, was carried out using primer pairs, designed on thebasis of the full sequence of the strain L. lactis subsp. lactis IL 1403(Table 1). Single colonies or total DNA were used as a templates, inthe following PCR program: 1 cycle of 95 1C; 35 cycles: 1 min at95 1C, 1 min at the annealing temperature, 2 min at 72 1C; and finalelongation for 7 min at 72 1C.
2.11. Reverse transcription analysis
First strand copy-DNA synthesis was performed mixing 0.3 mgof the DNase treated total RNA as template, 0.2 mg Exo-resistantrandom primer (Fermentas), 4 ml 5� reaction buffer, dNTP mix tofinal concentration 1 mM, 200 units RevertAidTM M-MuLV ReverseTranscriptase (Fermentas) and DEPC-treated water to final volume20ml. The mixture was incubated at 25 1C for 10 min and then at42 1C for 1 h. Heating at 70 1C for 10 min stopped the reaction.
RT-PCR, targeting different genes, putatively involved inthe starch hydrolysis was performed with primer pairs, listed inTable 1.
sis by Lactococcus lactis subsp. lactis
(1C)
Amplicon
size (bp)
Positions in L. lactis subsp.
lactis IL1403 chromosome
Sequence accession
no. (NCBI)
2156–2174 AE006357
1362 3518–3498
11,306–11,288 AE006398
1530 9776–9793
6497–6516 AE006303
1732 8229–209
3995–4014 AE006303
2346 6341–6321

ARTICLE IN PRESS
0 100 200 3000
1
2
3
4
5
30°C33°C37°C
Agitation (rpm)
Lac
tic
acid
(g/
l)
Fig. 2. Effect of the temperature and agitation on the lactic acid production by
Lactococcus lactis subsp. lactis B84 in M2 medium, without pH control.
0 50 100 150 2000
2
4
6
8
10
12
14
16
18
20
Lactic acid
Dry cell
Starch
Glucose
Time (h)
Con
cent
rati
on (
g/l)
Fig. 3. Batch lactic acid fermentation from potato starch by Lactococcus lactis
subsp. lactis B84 under non-controlled pH.
K. Petrov et al. / Food Microbiology 25 (2008) 550–557 553
3. Results
3.1. Starch fermentation by Lactococcus lactis subsp. lactis B84
3.1.1. Optimization of the media content
A variety of nutritive media with different content wasinvestigated in batch processes with duration 165 h, at 33 1C andstirring at 200 rpm. The media composition effect on the finallactate yield, biomass accumulation and starch hydrolysis degreeare presented in Fig. 1. Summarily, the medium componentsshould be quite different according to the purpose: the increasedlactic acid formation or, to the contrary, the vigorous biomassgrowth. The highest lactate production was obtained in M2 (brothwithout meat and yeast extracts), gaining 5.10 g l�1 LA, whereas inM1 (containing extracts) it reached only 3.50 g l�1. In the opposite,M1 medium was more appropriate than M2 for the cellspropagation (1.9 g l�1) and also for starch hydrolysis, achieving7.5 g l�1 digested starch at the end of the process. In M2 and M2A,the strain B84 grew in weaker degree (0.75 and 0.52 g l�1,respectively) and the starch hydrolysis attained was lower �6.4and 6.0 g l�1 digested starch. Therefore, the high accumulation ofbiomass in M1 affected in proportional increase in the starchhydrolysis, but it diminished lactic acid secretion.
Peptone was another very important medium constituent. Itsexclusion from the media led to drastic decrease in LA output. Thecomparison between LA, produced in media containing differentpeptones, revealed the influence of peptone’s origin on the lactateyield. Meat peptone was evaluated as most suitable (M2 broth,final lactate concentration 5.10 g l�1), next was the peptone fromcasein (M2A broth, 4.66 g l�1 lactate), followed by soy peptone(2.2 g l�1).
The use of original content of Elliker or M17 (with starchinstead of glucose), resulted in lower amounts of lactate obtained(o2 g l�1), independently of the strain good growth.
The presence of salts in the media was essential for the lactateformation, as in M2 with all salts excluded; only 3.43 g l�1 lactatewas synthesized.
Aiming at maximum lactic acid productivity, all further datawere obtained using M2 medium.
3.1.2. Optimization of the cultivation conditions without pH control
Further, the influence of other cultivation factors on the LAsynthesis was observed. LA concentration, reached at different
0
1
2
3
4
5
6
7
8
Dry cell
Lactic acid
Starch hydrolysed
M1
Con
cent
rati
ons
(g/l)
M2 M2A
Fig. 1. Effect of the nutrient media composition on the lactic acid, biomass and
hydrolyzed starch concentrations by Lactococcus lactis subsp. lactis B84 in batch
process without pH control. Media contents are described in Section 2.
temperatures and agitation was shown in Fig. 2. The optimalincubation was at temperature 33 1C and strong aeration. Agita-tion at more than 200 rpm did not affect the process. Thedynamics of lactic acid and biomass production, as well as thestarch degradation at these conditions, without pH control, wasillustrated in Fig. 3. The starch amount decreased to 11.5 g l�1,corresponding to 36% hydrolyzed starch at the end of the process.During the fermentation, pH of the media decreased from 6.5to 4.2.
The analysis of the intermediate products by HPLC showed theremaining dextrin and glucose. The concentration of glucose,formed by the autoclaving was unchangeable (�2 g l�1) during thecultivation, indicating that glucose residues released from thestarch were immediately consumed for growth or for lactateformation by B84 cells. No peak of maltose was detected.
In order to evaluate the general capacity of the strain B84 toconvert sugars into lactic acid, the glucose utilization kineticswas also observed. The comparison between the maximalvalues of LA, produced from 20 g l�1 glucose and 20 g l�1 starch(8 and 5.1 g l�1, respectively) indicated that the conversionpossibilities of the strain were at medium level (28–40%). Thus,the amount of lactate secreted is a strain-specific feature and theimprovement of the cultivation conditions is effective in certainlimits.
ARTICLE IN PRESS
0 50 100 150 2000
2
4
6
8
10
12
14
16
18
20 Lactic acid
Dry cell
Starch
Glucose
Time (h)
Con
cent
rati
on (
g/l)
Fig. 4. Batch lactic acid fermentation from potato starch by Lactococcus lactis
subsp. lactis B84 under pH, maintained at 6.0.
1000 bp 900 bp 800 bp
700 bp
600 bp
500 bp
400 bp
300 bp
200 bp
100 bp
1 62 3 4 5 7 8 9 10 11
Fig. 5. Amplified ribosomal DNA restriction analysis (ARDRA) of B84 and referent
strains with HhaI. No. of lanes and samples: 1, molecular weight marker 100 bp; 2,
Enterococcus faecium LMG 11423; 3, Enterococcus faecalis LMG 2937; 4, Enterococcus
durans LMG 10746; 5, Leuconostoc mesenteroides subsp. mesenteroides LMG 6893; 6,
Leuconostoc lactis LMG 8894; 7, Streptococcus thermophilus DSM 20617; 8,
Lactococcus lactis subsp. cremoris DSM 20069; 9, Lactococcus lactis subsp. lactis
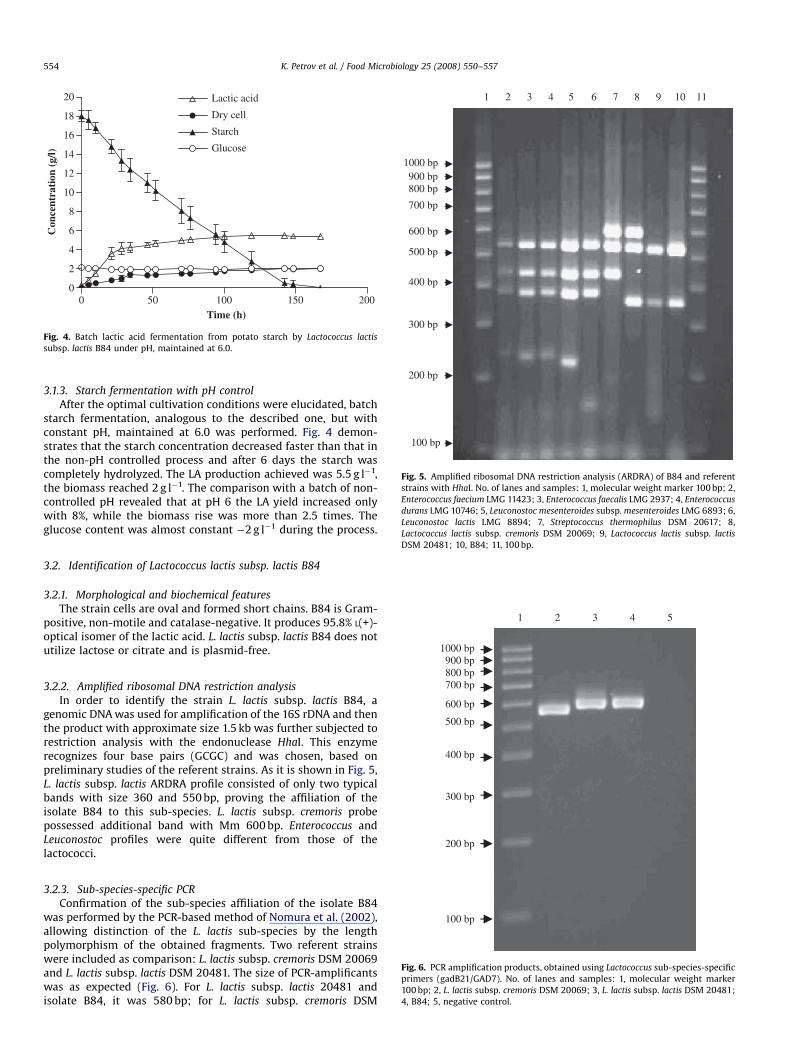
K. Petrov et al. / Food Microbiology 25 (2008) 550–557554
3.1.3. Starch fermentation with pH control
After the optimal cultivation conditions were elucidated, batchstarch fermentation, analogous to the described one, but withconstant pH, maintained at 6.0 was performed. Fig. 4 demon-strates that the starch concentration decreased faster than that inthe non-pH controlled process and after 6 days the starch wascompletely hydrolyzed. The LA production achieved was 5.5 g l�1,the biomass reached 2 g l�1. The comparison with a batch of non-controlled pH revealed that at pH 6 the LA yield increased onlywith 8%, while the biomass rise was more than 2.5 times. Theglucose content was almost constant �2 g l�1 during the process.
DSM 20481; 10, B84; 11, 100 bp.
1
1000 bp
2 3 4 5
3.2. Identification of Lactococcus lactis subsp. lactis B84
3.2.1. Morphological and biochemical features
The strain cells are oval and formed short chains. B84 is Gram-positive, non-motile and catalase-negative. It produces 95.8% L(+)-optical isomer of the lactic acid. L. lactis subsp. lactis B84 does notutilize lactose or citrate and is plasmid-free.
900 bp800 bp700 bp
600 bp
500 bp
400 bp
300 bp
200 bp
3.2.2. Amplified ribosomal DNA restriction analysis
In order to identify the strain L. lactis subsp. lactis B84, agenomic DNA was used for amplification of the 16S rDNA and thenthe product with approximate size 1.5 kb was further subjected torestriction analysis with the endonuclease HhaI. This enzymerecognizes four base pairs (GCGC) and was chosen, based onpreliminary studies of the referent strains. As it is shown in Fig. 5,L. lactis subsp. lactis ARDRA profile consisted of only two typicalbands with size 360 and 550 bp, proving the affiliation of theisolate B84 to this sub-species. L. lactis subsp. cremoris probepossessed additional band with Mm 600 bp. Enterococcus andLeuconostoc profiles were quite different from those of thelactococci.
100 bp
Fig. 6. PCR amplification products, obtained using Lactococcus sub-species-specific
primers (gadB21/GAD7). No. of lanes and samples: 1, molecular weight marker
100 bp; 2, L. lactis subsp. cremoris DSM 20069; 3, L. lactis subsp. lactis DSM 20481;
4, B84; 5, negative control.
3.2.3. Sub-species-specific PCR
Confirmation of the sub-species affiliation of the isolate B84was performed by the PCR-based method of Nomura et al. (2002),allowing distinction of the L. lactis sub-species by the lengthpolymorphism of the obtained fragments. Two referent strainswere included as comparison: L. lactis subsp. cremoris DSM 20069and L. lactis subsp. lactis DSM 20481. The size of PCR-amplificantswas as expected (Fig. 6). For L. lactis subsp. lactis 20481 andisolate B84, it was 580 bp; for L. lactis subsp. cremoris DSM

ARTICLE IN PRESS
1 52 3 4 6 7 8 9
K. Petrov et al. / Food Microbiology 25 (2008) 550–557 555
20069—540 bp (a result of deletion in the glutamate—decarbox-ylase gene; Nomura et al., 2002).
10 000 bp 8000 bp 6000 bp 5000 bp 4000 bp 3500 bp 3000 bp 2500 bp2000 bp
1500 bp
1000 bp
750 bp
500 bp
Fig. 8. RT-PCR amplification products, showing the transcription of amyL and
amyY in L. lactis subsp. lactis B84, cultured in M2 medium (lanes 4 and 7,
respectively). No. of lanes and samples: 1, molecular weight marker 1 kb; 2, IL 1403
3.2.4. Sequencing of the 16S rRNA gene
The complete sequence of the L. lactis subsp. lactis B84 16SrRNA gene also confirmed the affiliation of the strain to the sub-species lactis. Sequence comparison with the GenBank data (NCBI)showed 99% identity with the 16S rRNA gene of the type strainL. lactis subsp. lactis NCDO 604T (Accession no. AB100803; Moriet al., 2004), as well as with L. lactis subsp. lactis IL 1403(Accession no. X64887; Chiaruttini and Milet, 1993).
The opportunity for B84 to belong to biovar diacetylactis wasrejected for two reasons: first, the strain was plasmid-free,whereas the citrate utilization was known as a plasmid-encodedproperty (Kempler and McKay, 1979); and second, a PCR productwith primers, targeting the lactococcal citrate-permease gene,using total genomic DNA was not obtained (data not shown).
(amyL); 3, DSM 20481 (amyL); 4, B84 (amyL); 5, IL 1403 (amyY); 6, DSM 20481
(amyY); 7, B84 (amyY); 8, negative control (amyL); 9, negative control (amyY).
3.3. Gene expression of the starch-hydrolyzing enzymes
RT-PCR experiments were carried out on the base of thepreliminary studies of the starch metabolism (GenomeNet, KyotoUniversity Bioinformatics Center, www.genome.ad.jp) and on thecomplete genome sequence of L. lactis subsp. lactis IL 1403. Fourprimer pairs (Table 1), targeting genes for enzymes, taking part instarch degradation, were generated.
In the first place, PCR amplification experiments with chro-mosomal DNA, proving the presence of amyL, amyY, apu and glgP
(encoding cytoplasmic and extracellular a-amylases, amylopullu-lanase and glycogen phosphorylase) in the genome of L. lactis
subsp. lactis B84 were performed. Genomic DNA of L. lactis subsp.lactis IL 1403 was used as control. All genes tested were present inboth B84 and IL 1403 chromosomes (Fig. 7). Next, total RNA fromthe strains L. lactis subsp. lactis IL 1403, DSM 20481 and B84,cultivated in MRS-starch medium M1, was isolated and subjectedto reverse transcription. The obtained copy-DNA was used as atemplate for PCR with the described primer pairs. The resultsshowed that two of the genes—amyL and amyY were transcribedinto mRNA in the strain B84, but not in IL 1403. In the type strainL. lactis subsp. lactis DSM 20481, amyL and amyY were also presentin the chromosome (data not shown), but both were nottranscribed (Fig. 8).
1 6
10 000 bp 8000 bp 6000 bp 5000 bp 4000 bp 3500 bp
3000 bp 2500 bp 2000 bp
1500 bp
1000 bp
750 bp
500 bp
250 bp
2 3 4 5 7 8 9
Fig. 7. PCR detection of the genes, putatively involved in the starch hydrolysis by
Lactococcus lactis subsp. lactis, using chromosomal DNA, as a template. No. of lanes
and samples: 1, molecular weight marker 1 kb; 2, B84 (amyL); 3, IL 1403 (amyL); 4,
B84 (amyY); 5, IL 1403 (amyY); 6, B84 (apu); 7, IL 1403 (apu); 8, B84 (glgP); 9, IL
1403 (glgP).
The gene glgP was not expressed in both strains L. lactis subsp.lactis B84 and IL 1403 in the presence of starch, while the gene apu
was transcribed in IL 1403, but not in B84. Although the splittingof starch by a plasmid-encoded pullulanase of L. lactis strainIBB500 has been reported (Doman et al., 2000), the amylopullu-lanase, encoded by apu gene had no relation to the starchhydrolysis by B84.
All described experiments were duplicated with cells, grownon MRS (containing 20 g l�1 glucose). It was found that the genesamyL and amyY of B84 were inducibly transcribed into mRNA onlyif the cells were cultured in the starch-containing medium M1. Inthis medium, the constant low level of glucose (2 g l�1), main-tained during the fermentation, did not repress the expression ofamyL and amyY in B84, in contrast to the high concentration of20 g l�1 glucose in MRS.
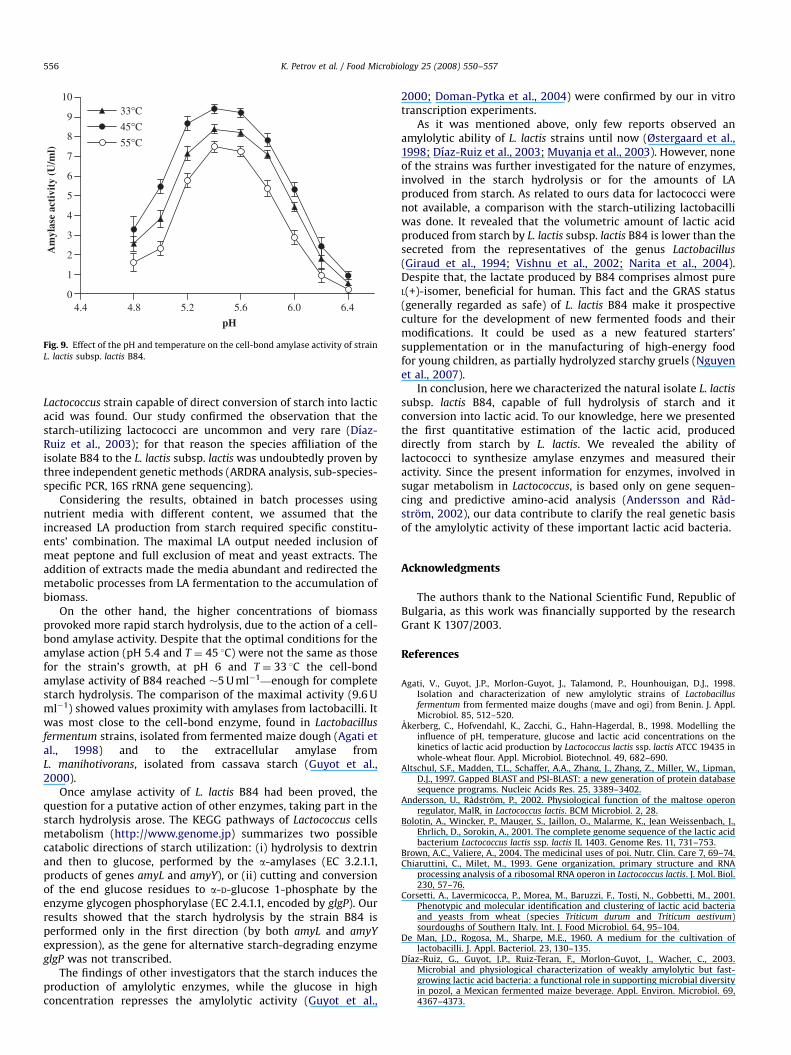
3.4. Amylase activity assay
Since the RT-PCR experiments proved the expression of thegenes, encoding cytoplasmic and extracellular amylases, anamylase activity assay, able to distinguish the share of eachenzyme, was performed. The cultures were grown in M1 mediumto the beginning of the stationary phase. The activities ofharvested cells (cell-bond amylase) and of the cell-free liquidsupernatants (extracellular amylase) were measured in parallel, asdescribed by Agati et al. (1998).
The results of the tests, performed at different pH andtemperatures revealed that the activity of the harvested cellswas much higher than that of the liquid supernatant. Varying thetrial conditions, the maximum cell-bond amylase activity reached9.6 U ml�1, while the extracellular remained at less than 1 U ml�1.The assay strongly suggested that the cell-bond enzyme wasmainly responsible for the starch hydrolysis.
In a consequence of experiments, illustrated in Fig. 9, theinfluence of the pH and temperature on the cell-bond amylaseactivity was examined. The enzyme required acidic values of pHfor its action and possessed maximum activity at pH around pH5.4. The temperature optimum was found to be 45 1C.
4. Discussion
In the presented work, the microbial content of 11 types offlours and sourdoughs was tested for amylolytic LAB and only one
ARTICLE IN PRESS
4.4 4.8 5.2 5.6 6.0 6.40
1
2
3
4
5
6
7
8
9
1033°C
45°C
55°C
pH
Am
ylas
e ac
tivi
ty (
U/m
l)
Fig. 9. Effect of the pH and temperature on the cell-bond amylase activity of strain
L. lactis subsp. lactis B84.
K. Petrov et al. / Food Microbiology 25 (2008) 550–557556
Lactococcus strain capable of direct conversion of starch into lacticacid was found. Our study confirmed the observation that thestarch-utilizing lactococci are uncommon and very rare (Dıaz-Ruiz et al., 2003); for that reason the species affiliation of theisolate B84 to the L. lactis subsp. lactis was undoubtedly proven bythree independent genetic methods (ARDRA analysis, sub-species-specific PCR, 16S rRNA gene sequencing).
Considering the results, obtained in batch processes usingnutrient media with different content, we assumed that theincreased LA production from starch required specific constitu-ents’ combination. The maximal LA output needed inclusion ofmeat peptone and full exclusion of meat and yeast extracts. Theaddition of extracts made the media abundant and redirected themetabolic processes from LA fermentation to the accumulation ofbiomass.
On the other hand, the higher concentrations of biomassprovoked more rapid starch hydrolysis, due to the action of a cell-bond amylase activity. Despite that the optimal conditions for theamylase action (pH 5.4 and T ¼ 45 1C) were not the same as thosefor the strain’s growth, at pH 6 and T ¼ 33 1C the cell-bondamylase activity of B84 reached �5 U ml�1—enough for completestarch hydrolysis. The comparison of the maximal activity (9.6 Uml�1) showed values proximity with amylases from lactobacilli. Itwas most close to the cell-bond enzyme, found in Lactobacillus
fermentum strains, isolated from fermented maize dough (Agati etal., 1998) and to the extracellular amylase fromL. manihotivorans, isolated from cassava starch (Guyot et al.,2000).
Once amylase activity of L. lactis B84 had been proved, thequestion for a putative action of other enzymes, taking part in thestarch hydrolysis arose. The KEGG pathways of Lactococcus cellsmetabolism (http://www.genome.jp) summarizes two possiblecatabolic directions of starch utilization: (i) hydrolysis to dextrinand then to glucose, performed by the a-amylases (EC 3.2.1.1,products of genes amyL and amyY), or (ii) cutting and conversionof the end glucose residues to a-D-glucose 1-phosphate by theenzyme glycogen phosphorylase (EC 2.4.1.1, encoded by glgP). Ourresults showed that the starch hydrolysis by the strain B84 isperformed only in the first direction (by both amyL and amyY
expression), as the gene for alternative starch-degrading enzymeglgP was not transcribed.
The findings of other investigators that the starch induces theproduction of amylolytic enzymes, while the glucose in highconcentration represses the amylolytic activity (Guyot et al.,
2000; Doman-Pytka et al., 2004) were confirmed by our in vitrotranscription experiments.
As it was mentioned above, only few reports observed anamylolytic ability of L. lactis strains until now (Østergaard et al.,1998; Dıaz-Ruiz et al., 2003; Muyanja et al., 2003). However, noneof the strains was further investigated for the nature of enzymes,involved in the starch hydrolysis or for the amounts of LAproduced from starch. As related to ours data for lactococci werenot available, a comparison with the starch-utilizing lactobacilliwas done. It revealed that the volumetric amount of lactic acidproduced from starch by L. lactis subsp. lactis B84 is lower than thesecreted from the representatives of the genus Lactobacillus
(Giraud et al., 1994; Vishnu et al., 2002; Narita et al., 2004).Despite that, the lactate produced by B84 comprises almost pureL(+)-isomer, beneficial for human. This fact and the GRAS status(generally regarded as safe) of L. lactis B84 make it prospectiveculture for the development of new fermented foods and theirmodifications. It could be used as a new featured starters’supplementation or in the manufacturing of high-energy foodfor young children, as partially hydrolyzed starchy gruels (Nguyenet al., 2007).
In conclusion, here we characterized the natural isolate L. lactis
subsp. lactis B84, capable of full hydrolysis of starch and itconversion into lactic acid. To our knowledge, here we presentedthe first quantitative estimation of the lactic acid, produceddirectly from starch by L. lactis. We revealed the ability oflactococci to synthesize amylase enzymes and measured theiractivity. Since the present information for enzymes, involved insugar metabolism in Lactococcus, is based only on gene sequen-cing and predictive amino-acid analysis (Andersson and Rad-strom, 2002), our data contribute to clarify the real genetic basisof the amylolytic activity of these important lactic acid bacteria.
Acknowledgments
The authors thank to the National Scientific Fund, Republic ofBulgaria, as this work was financially supported by the researchGrant K 1307/2003.
References
Agati, V., Guyot, J.P., Morlon-Guyot, J., Talamond, P., Hounhouigan, D.J., 1998.Isolation and characterization of new amylolytic strains of Lactobacillusfermentum from fermented maize doughs (mave and ogi) from Benin. J. Appl.Microbiol. 85, 512–520.
Akerberg, C., Hofvendahl, K., Zacchi, G., Hahn-Hagerdal, B., 1998. Modelling theinfluence of pH, temperature, glucose and lactic acid concentrations on thekinetics of lactic acid production by Lactococcus lactis ssp. lactis ATCC 19435 inwhole-wheat flour. Appl. Microbiol. Biotechnol. 49, 682–690.
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W., Lipman,D.J., 1997. Gapped BLAST and PSI-BLAST: a new generation of protein databasesequence programs. Nucleic Acids Res. 25, 3389–3402.
Andersson, U., Radstrom, P., 2002. Physiological function of the maltose operonregulator, MalR, in Lactococcus lactis. BCM Microbiol. 2, 28.
Bolotin, A., Wincker, P., Mauger, S., Jaillon, O., Malarme, K., Jean Weissenbach, J.,Ehrlich, D., Sorokin, A., 2001. The complete genome sequence of the lactic acidbacterium Lactococcus lactis ssp. lactis IL 1403. Genome Res. 11, 731–753.
Brown, A.C., Valiere, A., 2004. The medicinal uses of poi. Nutr. Clin. Care 7, 69–74.Chiaruttini, C., Milet, M., 1993. Gene organization, primary structure and RNA
processing analysis of a ribosomal RNA operon in Lactococcus lactis. J. Mol. Biol.230, 57–76.
Corsetti, A., Lavermicocca, P., Morea, M., Baruzzi, F., Tosti, N., Gobbetti, M., 2001.Phenotypic and molecular identification and clustering of lactic acid bacteriaand yeasts from wheat (species Triticum durum and Triticum aestivum)sourdoughs of Southern Italy. Int. J. Food Microbiol. 64, 95–104.
De Man, J.D., Rogosa, M., Sharpe, M.E., 1960. A medium for the cultivation oflactobacilli. J. Appl. Bacteriol. 23, 130–135.
Dıaz-Ruiz, G., Guyot, J.P., Ruiz-Teran, F., Morlon-Guyot, J., Wacher, C., 2003.Microbial and physiological characterization of weakly amylolytic but fast-growing lactic acid bacteria: a functional role in supporting microbial diversityin pozol, a Mexican fermented maize beverage. Appl. Environ. Microbiol. 69,4367–4373.

ARTICLE IN PRESS
K. Petrov et al. / Food Microbiology 25 (2008) 550–557 557
Doman, M., Czerniec, E., Targonski, Z., Bardowski, J., 2000. Production and geneticregulation of an amylase in Lactococcus lactis. In: Bielecki, S., Tramper, J., Polak,J. (Eds.), Food Biotechnology—Progress in Biotechnology, vol. 17. ElsevierScience, Amsterdam, pp. 67–72.
Doman-Pytka, M., Renault, P., Bardowski, J., 2004. Gene-cassette for adaptation ofLactococcus lactis to a plant environment. Lait 84, 33–37.
Elliker, P.R., Anderson, A.W., Hannesson, G., 1956. An agar culture medium for lacticacid streptococci and lactobacilli. J. Dairy Sci. 39, 1611–1612.
Giraud, E., Brauman, A., Kekele, S., Lelong, B., Raimbault, M., 1991. Isolation andphysiological study of an amylolytic strain of Lactobacillus plantarum. Appl.Microbiol. Biotechnol. 36, 379–383.
Giraud, E., Gosselin, L., Marin, B., Parada, J.L., Raimbault, M., 1993. Purification andcharacterization of an extracellular amylase from Lactobacillus plantarum strainA6. J. Appl. Bacteriol. 75, 276–282.
Giraud, E., Champailler, A., Raimbault, M., 1994. Dagradation of starch by a wildamylolytic strain of Lactobacillus plantarum. Appl. Environ. Microbiol. 60,4319–4323.
Gregersen, T., 1978. Rapid method for distinction of Gram-negative from Gram-positive bacteria. Eur. J. Appl. Microbiol. Biothechnol. 5, 123–127.
Guyot, J.P., Calderon, M., Morlon-Guyot, J., 2000. Effect of pH control on lactic acidfermentation of starch by Lactobacillus manihotivorans LMG 18010T. J. Appl.Microbiol. 88, 176–182.
Hamad, S.H., Dieng, M.C., Ehrmann, M.A., Vogel, R.F., 1997. Characterization of thebacterial flora of the Sudanese sorghum flour and sorghum sourdough. J. Appl.Microbiol. 83, 764–770.
Hofvendahl, K., Hahn-Hagerdal, B., 1997. L-lactic acid production from whole wheatflour hydrolysate using strains of Lactobacilli and Lactococci. Enzyme Microb.Technol. 20, 301–307.
Hofvendahl, K., Akerberg, C., Zacchi, G., Hahn-Hagerdal, B., 1999. Simultaneousenzymatic wheat starch saccharification and fermentation to lactic acid byLactococcus lactis. Appl. Microbiol. Biotechnol. 52, 163–169.
Kempler, G.M., McKay, L.L., 1979. Characterization of plasmid deoxyribonucleicacid in Streptococcus lactis subsp. diacetylactis: evidence for plasmid-linkedcitrate utilization. Appl. Environ. Microbiol. 37, 316–323.
Mori, S., Mori, K., Suzuki, I., Kasumi, T., 2004. Phylogenetic analysis of Lactococcuslactis subspecies based on decoding the sequence of the pepT tripeptidasegene, the pepV dipeptidase gene and 16S rRNA. Syst. Appl. Microbiol. 27,414–422.
Morlon-Guyot, J., Guyot, J.P., Pot, B., Jacobe de Haut, I., Raimbault, M., 1998.Lactobacillus manihotivorans sp. nov., a new starch-hydrolysing lactic acidbacterium isolated during cassava sour starch fermentation. Int. J. Syst.Bacteriol. 48, 1101–1109.
Muyanja, C.M., Narvhus, J.A., Treimo, J., Langsrud, T., 2003. Isolation, characterisa-tion and identification of lactic acid bacteria from bushera: a Ugandantraditional fermented beverage. Int. J. Food Microbiol. 80, 201–210.
Nakamura, L.K., 1981. Lactobacillus amylovorus, a new starch-hydrolyzing speciesfrom cattle waste-corn fermentations. Int. J. Syst. Bacteriol. 31, 56–63.
Narita, J., Nakahara, S., Fukuda, H., Kondo, A., 2004. Efficient production of L-(+)-lactic acid from raw starch by Streptococcus bovis 148. J. Biosci. Bioeng. 97,423–425.
Nguyen, T.T.T., Loiseau, G., Icard-Verniere, C., Rochette, I., Treche, S., Guyot, J.P.,2007. Effect of fermentation by amylolytic lactic acid bacteria, in processcombinations, on characteristics of rice/soybean slurries: a new method forpreparing high energy density complementary foods for young children. FoodChem. 100, 623–631.
Nolasco-Hipolito, C., Matsunaka, T., Kobayashi, G., Sonomoto, K., Ishizaki, A., 2002.Synchronized fresh cell bioreactor system for continuous L-(+)-lactic acidproduction using Lactococcus lactis IO-1 in hydrolysed sago starch. J. Biosci.Bioeng. 93, 281–287.
Nomura, M., Kobayashi, M., Okamoto, T., 2002. Rapid PCR-based method which candetermine both phenotype and genotype of Lactococcus lactis subspecies. Appl.Environ. Microbiol. 68, 2209–2213.
Okano, K., Kimura, S., Narita, J., Fukuda, H., Kondo, A., 2007. Improvement in lacticacid production from starch using a-amylase secreting Lactococcus lactis cellsadapted to maltose or starch. Appl. Microbiol. Biotechnol. 75, 1007–1013.
Østergaard, A., Embarek, P.K.B., Wedell-Neergaard, C., Huss, H.H., Gram, L., 1998.Characterization of anti-listerial lactic acid bacteria isolated from Thaifermented fish products. Food Microbiol. 15, 223–233.
Roble, N.D., Ogbonna, J.C., Tanaka, H., 2003. L-lactic acid production from rawcassava starch in a circulating loop bioreactor with cells immobilized in loofa(Luffa cylindrica). Biotechnol. Lett. 25, 1093–1098.
Rocha, J.M., Malcata, F.X., 1999. On the microbiological profile of traditionalPortuguese sourdough. J. Food Prot. 62, 1416–1429.
Rodtong, S., Tannock, G.W., 1993. Differentiation of Lactobacillus strains byribotyping. Appl. Environ. Microbiol. 59, 3480–3484.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTAL W: improving thesensitivity of progressive multiple sequence alignment through sequenceweighting, positions-specific gap penalties and weight matrix choice. NucleicAcids Res. 22, 4673–4680.
Samarzija, D., Antunac, N., Havranek, J.L., 2001. Taxonomy, physiology and growthof Lactococcus lactis: a review. Mljekarstvo 51, 35–48.
Sambrook, J., Russel, D.W., 2001. In: Irwin, N. (Ed.), Molecular Cloning:A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
Satoh, E., Ito, Y., Sasaki, Y., Sasaki, T., 1997. Application of the extracellular alpha-amylase gene from Streptococcus bovis 148 to construction of a secretion vectorfor yogurt starter strains. Appl. Environ. Micribiol. 63, 4593–4596.
Sawadogo-Lingani, H., Lei, V., Diawara, B., Nielsen, D.S., Møller, P.L., Traore, A.S.,Jakobsen, M., 2007. The biodiversity of predominant lactic acid bacteria in doloand pito wort for the production of sorghum beer. J. Appl. Microbiol. 103,765–777.
Vishnu, C., Seenayya, G., Reddy, G., 2002. Direct fermentation of pure and crudestarchy substrates to L(+)-lactic acid using Lactobacillus amylophilus CV6. WorldJ. Microbiol. Biotechnol. 18, 429–433.
Wee, Y.-J., Kim, J.-N., Ryu, H.-W., 2006. Biotechnological production of lactic acidand its recent applications. Food Technol. Biotechnol. 44, 163–172.
Weisburg, W.G., Barns, S.M., Pelletier, D.A., Lane, D.J., 1991. 16S ribosomal DNAamplification for phylogenetic study. J. Bacteriol. 173, 697–703.
Zhang, D.X., Cheyran, M., 1991. Direct fermentation of starch to lactic acid byLactobacillus amylovorus. Biotechnol. Lett. 13, 733–738.




















