
In vitro inhibition of Citrobacter freundii, a red-leg syndrome associated pathogen in raniculture,...
9
In vitro inhibition of Citrobacter freundii, a red-leg syndrome associated pathogen in raniculture, by indigenous Lactococcus lactis CRL 1584 Sergio E. Pasteris a , Marcos G. Guidoli a , Marı ´a C. Otero a,b , Marta I. Bu ¨ hler a , Marı ´a E. Nader-Macı ´as b, * a Instituto Superior de Investigaciones Biolo ´gicas (INSIBIO-CONICET) - Instituto de Biologı´a ‘‘Dr. Francisco D. Barbieri’’, Facultad de Bioquı´mica, Quı´mica y Farmacia - Universidad Nacional de Tucuma ´n, Chacabuco 461, CP: T4000ILI, San Miguel de Tucuma ´n, Argentina b Centro de Referencia para Lactobacilos (CERELA-CONICET), Chacabuco 145, CP: T4000ILC, San Miguel de Tucuma ´n, Argentina 1. Introduction Raniculture represents a growing commercial aqua- culture activity widely distributed throughout the world, Lithobates catesbeianus (Frost et al., 2006) being the selected species. The Food and Agriculture Organization (2001) has indicated that bullfrog meat has a high proportion of essential amino acids, a large degree of absorbable proteins and low contents of sodium chloride and lipids (mainly cholesterol). These commercial require- ments have resulted in an intensive L. catesbeianus production process that increases the risk of bacterial disease outbreaks (Mauel et al., 2002). Red-leg syndrome (RLS) or bacterial dermatosepticemia is one of the main infectious diseases that affect captive bullfrog, causing high economic losses (Texeira et al., 2002; Densmore and Earl Green, 2007). The etiological agents involve Citrobacter freundii, and the widespread therapies include the use of antibiotics (Glorioso et al., 1974; Mauel et al., 2002, Schadich and Cole, 2009; Zhen-Yu et al., 2009). Thus, the Veterinary Microbiology 151 (2011) 336–344 A R T I C L E I N F O Article history: Received 13 November 2010 Received in revised form 18 March 2011 Accepted 23 March 2011 Keywords: Lithobates catesbeianus Raniculture Infectious diseases Lactic acid bacteria Probiotics A B S T R A C T Red-leg syndrome (RLS) is one of the main infectious diseases that cause economic losses in Lithobates catesbeianus hatcheries, Citrobacter freundii being an etiological agent. Treatment or prevention with therapeutics or chemicals results in modifications of the indigenous microbiota, development of antibiotic resistance, presence of their residues in food and enhancement of production costs. Thus, probiotics could be used as an alternative therapy. Lactic acid bacteria are part of the indigenous microbiota of healthy frogs and can prevent pathogen colonization by different mechanisms, including the production of antagonistic substances. In this work, the evaluation and characterization of the inhibition of C. freundii CFb by Lactococcus lactis subsp. lactis CRL 1584, a potentially probiotic candidate, were carried out. This strain produced lactic acid, H 2 O 2 and bacteriocin in static and shaken conditions and inhibited pathogen growth in associative cultures, with an earlier inhibition under agitated conditions. The elimination of each of the antimicrobial metabolites partially abolished the inhibition of the pathogen, suggesting that the inhibitory effect could be attributed to a combined action of the three antagonistic molecules. Electron microphotographs revealed the damage caused by L. lactis CRL 1584 supernatants to C. freundii cells. The addition of pure lactic acid, H 2 O 2 and bacteriocin to the culture media showed that each metabolite caused different morphological modifications in C. freundii, in agreement with the effect on viable cell counts. The results support the possibility that L. lactis CRL 1584 might be considered as a probiotic to be used in the prevention of RLS in raniculture. ß 2011 Elsevier B.V. All rights reserved. * Corresponding author. Tel.: +54 381 4310465; fax: +54 381 4311720. E-mail address: [email protected] (M.E. Nader-Macı ´as). Contents lists available at ScienceDirect Veterinary Microbiology jou r nal h o mep ag e: w ww .els evier .co m/lo c ate/vetm ic 0378-1135/$ – see front matter ß 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.vetmic.2011.03.025
Transcript of In vitro inhibition of Citrobacter freundii, a red-leg syndrome associated pathogen in raniculture,...

In vitro inhibition of Citrobacter freundii, a red-leg syndrome associatedpathogen in raniculture, by indigenous Lactococcus lactis CRL 1584
Sergio E. Pasteris a, Marcos G. Guidoli a, Marıa C. Otero a,b, Marta I. Buhler a,Marıa E. Nader-Macıas b,*a Instituto Superior de Investigaciones Biologicas (INSIBIO-CONICET) - Instituto de Biologıa ‘‘Dr. Francisco D. Barbieri’’, Facultad de Bioquımica, Quımica y Farmacia -
Universidad Nacional de Tucuman, Chacabuco 461, CP: T4000ILI, San Miguel de Tucuman, Argentinab Centro de Referencia para Lactobacilos (CERELA-CONICET), Chacabuco 145, CP: T4000ILC, San Miguel de Tucuman, Argentina
1. Introduction
Raniculture represents a growing commercial aqua-culture activity widely distributed throughout the world,Lithobates catesbeianus (Frost et al., 2006) being theselected species. The Food and Agriculture Organization(2001) has indicated that bullfrog meat has a highproportion of essential amino acids, a large degree of
absorbable proteins and low contents of sodium chlorideand lipids (mainly cholesterol). These commercial require-ments have resulted in an intensive L. catesbeianus
production process that increases the risk of bacterialdisease outbreaks (Mauel et al., 2002). Red-leg syndrome(RLS) or bacterial dermatosepticemia is one of the maininfectious diseases that affect captive bullfrog, causinghigh economic losses (Texeira et al., 2002; Densmore andEarl Green, 2007). The etiological agents involve Citrobacter
freundii, and the widespread therapies include the use ofantibiotics (Glorioso et al., 1974; Mauel et al., 2002,Schadich and Cole, 2009; Zhen-Yu et al., 2009). Thus, the
Veterinary Microbiology 151 (2011) 336–344
A R T I C L E I N F O
Article history:
Received 13 November 2010
Received in revised form 18 March 2011
Accepted 23 March 2011
Keywords:
Lithobates catesbeianus
Raniculture
Infectious diseases
Lactic acid bacteria
Probiotics
A B S T R A C T
Red-leg syndrome (RLS) is one of the main infectious diseases that cause economic losses
in Lithobates catesbeianus hatcheries, Citrobacter freundii being an etiological agent.
Treatment or prevention with therapeutics or chemicals results in modifications of the
indigenous microbiota, development of antibiotic resistance, presence of their residues in
food and enhancement of production costs. Thus, probiotics could be used as an alternative
therapy. Lactic acid bacteria are part of the indigenous microbiota of healthy frogs and can
prevent pathogen colonization by different mechanisms, including the production of
antagonistic substances. In this work, the evaluation and characterization of the inhibition
of C. freundii CFb by Lactococcus lactis subsp. lactis CRL 1584, a potentially probiotic
candidate, were carried out. This strain produced lactic acid, H2O2 and bacteriocin in static
and shaken conditions and inhibited pathogen growth in associative cultures, with an
earlier inhibition under agitated conditions. The elimination of each of the antimicrobial
metabolites partially abolished the inhibition of the pathogen, suggesting that the
inhibitory effect could be attributed to a combined action of the three antagonistic
molecules. Electron microphotographs revealed the damage caused by L. lactis CRL 1584
supernatants to C. freundii cells. The addition of pure lactic acid, H2O2 and bacteriocin to
the culture media showed that each metabolite caused different morphological
modifications in C. freundii, in agreement with the effect on viable cell counts. The
results support the possibility that L. lactis CRL 1584 might be considered as a probiotic to
be used in the prevention of RLS in raniculture.
� 2011 Elsevier B.V. All rights reserved.
* Corresponding author. Tel.: +54 381 4310465; fax: +54 381 4311720.
E-mail address: [email protected] (M.E. Nader-Macıas).
Contents lists available at ScienceDirect
Veterinary Microbiology
jou r nal h o mep ag e: w ww .e ls evier . co m/lo c ate /vetm i c
0378-1135/$ – see front matter � 2011 Elsevier B.V. All rights reserved.
doi:10.1016/j.vetmic.2011.03.025

utilization of probiotics mainly represented by lactic acidbacteria (LAB) has been proposed as an alternative therapyinstead of antibiotics for the treatment or prevention ofinfectious diseases in aquaculture (Nikoskelainen et al.,2001; Irianto and Austin, 2002; Balcazar et al., 2006;Farzanfar, 2006; Gatesoupe, 2008; Ringø et al., 2010).However, in raniculture there is no evidence concerningthe use of autochthonous probiotic LAB strains from L.
catesbeianus. Previous results allowed us to select Lacto-
coccus lactis subsp. lactis CRL 1584 with beneficial proper-ties (surface characteristics and antagonistic activityagainst Listeria and Pseudomona species) (Pasteris et al.,2009a). Therefore, the aim of our research group is theformulation of probiotic products for veterinary applica-tion to prevent bacterial infectious diseases in raniculturethrough the restoration of indigenous microbiota. Theseproducts could reduce the need for antibiotics, improvingthe health status of animals and the economic profitabilityof livestock. The purpose of the present research was toevaluate and further study the inhibitory effects of L. lactis
subsp. lactis CRL 1584, an indigenous resident in a bullfroghatchery and a potential probiotic candidate (Pasteriset al., 2009a), on the growth of C. freundii, an RLS-relatedpathogen in raniculture.
2. Materials and methods
2.1. Bacterial strains and culture conditions
L. lactis subsp. lactis CRL 1584 was selected from amonga group of indigenous LAB strains isolated from an L.
catesbeianus hatchery on the basis of its beneficialproperties and identified by phenotypic and genotypicapproaches (Pasteris et al., 2006, 2008, 2009a). C. freundii
CFb was isolated from skin ulceration in animal displayingRLS (Pasteris et al., 2006). The strains were grown on LAPTgbroth (Raibaud et al., 1963) for 7 h for the LAB strain and8 h for the pathogenic bacteria at 37 8C in static conditions.All microorganisms were stored in milk-yeast extract (13%skim milk, 1% yeast extract) at �20 8C.
2.2. L. lactis CRL 1584 growth and determination of
antagonistic metabolites
The LAB strain was grown in the conditions statedabove in both static and agitated (120 cycles/min) culturesfor 7 h. Samples were taken at different time intervals forgrowth (O.D. 540 nm and CFU/ml) and pH determinations.Cell-free supernatants of LAB cultures were obtained bycentrifugation at 3000 � g at 4 8C for 20 min and used toquantify bacteriocin, lactic acid and H2O2 production.Bacteriocin was determined using Listeria monocytogenes
Scott A as an indicator strain (Pasteris et al., 2009a), whilelactic acid was quantified by HPLC. Thus, 400 ml of LABstrain supernatants taken at different incubation timeswere treated with 200 ml trichloroacetic acid (4% w/v) for2 min and centrifuged at 14000 � g at 4 8C for 5 min. 20 mltreated supernatants were analyzed using a Knauerchromatography system attached to a K-2301 refractionindex detector and a ZC 90 (Zeltex) oven. Differentcompounds were separated in a Biorad Aminex HPX-
87H (300 � 7.8 mm) column at 41 8C using 5 mM H2SO4
with an isocratic gradient at a 0.6 ml/min flux speed. 5 mMH2SO4 pattern solutions were used for calibration. The L.
lactis strain was grown in LAPTg broth in static and shakenconditions to study the kinetics of H2O2 production. Atdifferent time intervals, samples were taken and H2O2 wasdetermined according to Juarez Tomas et al. (2004).
2.3. Associative cultures of L. lactis CRL 1584 and C. freundii
CFb
Co-culture assays were carried out in LAPTg brothunder static and agitated conditions as described above.The strains were inoculated to obtain 108 CFU/ml of L. lactis
CRL 1584 and 107 CFU/ml of C. freundii CFb. Bacterialgrowth was quantified by the plate dilution method usingselective culture media: Mac Conkey agar for C. freundii
and MRS agar (de Man et al., 1969) for the LAB strain. Purecultures were performed in LAPTg broth at 37 8C under thesame conditions.
2.4. Determination of the chemical nature of the antagonistic
activity of L. lactis CRL 1584 supernatant on C. freundii CFb
To determine the chemical nature of the antagonisticsubstances produced by L. lactis on C. freundii, cell-freesupernatants from 5 h LAB cultures were treated asfollows: (a) neutralized with 2 N NaOH; (b) treated with0.5 mg/ml catalase 1 h at 25 8C; (c) treated with 1 mg/ml a-chymotrypsin prepared in 0.05 M Tris–HCl buffer + 1 MCaCl2, pH 8.0, 1 h at 37 8C; (d) treated with catalase and a-chymotrypsin as stated above; (e) untreated supernatant.C. freundii cells from 5 h cultures were centrifuged, washedtwice with sterile physiological solution and suspended inall the L. lactis supernatants and in the C. freundii spentsupernatant. All cell suspensions were incubated in staticand agitated conditions for 5 and 7 h at 37 8C. CFU/ml weredetermined and ultrastructural cell damages evaluated bytransmission electron microscopy.
2.5. Effect of the different antagonistic substances produced
by L. lactis CRL 1584 in pure form on C. freundii CFb viability
To evaluate the effect of each of the antagonisticsubstances produced by L. lactis on C. freundii, experimentswere performed by adding the pure reagents (lactic acidand H2O2) or bacteriocin (as concentrated supernatant) atsimilar levels to those produced by the LAB strain. C.
freundii cells were harvested from 5 h cultures, washedtwice with sterile physiological solution and suspended inmodified LAPTg broth prepared as follows: (a) LAPTg + 5 g/llactic acid; (b) LAPTg + 0.01 mM H2O2; (c) LAPTg + 2200 AU(Arbitrary Units)/ml bacteriocin; (d) LAPTg + 5 g/l lacticacid + 0.01 mM H2O2; (e) LAPTg + 5 g/l lacticacid + 2200 AU/ml bacteriocin; (f) LAPTg + 0.01 mMH2O2 + 2200 AU/ml bacteriocin; (g) LAPTg + 5 g/l lacticacid + 0.01 mM H2O2 + 2200 AU/ml bacteriocin. LAPTg withno additions was used as control. The bacteriocin-contain-ing medium was prepared by concentration (10�) of theLAB strain supernatant (60 8C, 10 min in a Buchi R110rotavap), neutralized and treated with catalase. Control
S.E. Pasteris et al. / Veterinary Microbiology 151 (2011) 336–344 337

cultures were carried out in LAPTg broth withoutmodifications. All suspensions were incubated for 5 and7 h at 37 8C in static and shaken conditions. CFU/ml weredetermined and ultrastructural cell damage was evaluated.
All chemicals used were purchased from Sigma–AldrichChemical Co., St. Louis, MO (USA) with the exception ofH2SO4 and H2O2, which were obtained from Ciccarellilaboratories (Buenos Aires, Argentina). The components forLAPTg preparation were supplied by Britania laboratories(Buenos Aires, Argentina).
2.6. Transmission electron microscopy (TEM)
Cells obtained under each of the above experimentalconditions were harvested, suspended in LAPTg brothsupplemented with 3% glutaraldehyde (1:1 v/v) andincubated for 30 min at room temperature; then theywere centrifuged at 14000 � g at 4 8C for 5 min andsuspended in 3% glutaraldehyde. Finally, they were fixedin osmium tetroxide and observed by TEM.
2.7. Statistical analysis
All experiments were performed by triplicate. Thenumber of microorganisms in the associative and purecultures under different conditions was subjected to amultivariate analysis of variance (General ANOVA). TheLSD Fisher test was used to compare the results fromdifferent cultures incubated during the same time periodand the same treatments at different times. A probability ofP < 0.05 was considered statistically significant. A MINI-TAB (version 15) software was used.
3. Results
3.1. L. lactis CRL 1584 growth and production of antimicrobial
compounds
L. lactis in pure cultures reached the stationary phase5 h after inoculation while the number of viable cellsincreased more than 3 log10 units. The pH values decreased2 units after 7 h of incubation (Fig. 1). The statisticalanalysis indicated that agitation did not induce anysignificant effects (P < 0.05) on LAB strain growth.
Bacteriocin was produced during L. lactis growth withno significant differences in production between static andagitated cultures. However, highest bacteriocin concen-
tration in both static (2315 AU/ml) and agitated (2225 AU/ml) cultures was similar, although in static conditionsthese values were obtained at 5 h while in agitated cultures1 h later.
Lactic acid production also increased with the LABstrain growth and depended on incubation conditions. Instatic cultures, organic acid concentration increased up to7 h of incubation with a maximum of 5.23 g/l, while inagitated conditions a maximum of 4.30 g/l was reachedafter 6 h growth.
Although H2O2 could not be detected by the quantita-tive assay, the qualitative method of TMB + peroxidase(Pasteris et al., 2009a) showed the presence of bluecolonies, indicating that L. lactis was able to produce H2O2.
3.2. Associative cultures of L. lactis CRL 1584 and C. freundii
CFb
The growth of L. lactis and C. freundii in associativecultures under static conditions showed a significantreduction (P < 0.05) in C. freundii viable cell counts after6 h, decreasing 3 log10 units compared to control (Fig. 2A).No viable cells of the pathogen were detected after 8 h ofco-culture. A similar behavior was observed in agitatedconditions, where the number of viable cells of C. freundii
was lower than in the control culture and the reductionbecame significant (P < 0.05) after 6 h of co-incubation. Atthis time, there was a difference of 5.5 log10 units withrespect to the control, with no viable cell counts at 7 h ofincubation, which means an earlier decrease in the numberof viable pathogen cells (Fig. 2B). In both static and shakenconditions, a higher decrease in pH values in the
Fig. 1. Growth and pH values of L. lactis CRL 1584 under static conditions.
Fig. 2. Growth of C. freundii CFb in pure and associative cultures with L.
lactis CRL 1584 in (A) static and (B) shaken conditions. (*) indicates
significant differences (P < 0.05) between the number of viable cells of the
individual and associative cultures at the same incubation times.
S.E. Pasteris et al. / Veterinary Microbiology 151 (2011) 336–344338
associative cultures was detected when compared with thepure cultures, with no significant differences between thetwo incubation conditions. With respect to the LAB strain,the number of viable cells increased from 7.026 � 0.03 to8.92 � 0.01 log10 CFU/ml after 4 h of incubation in staticculture and remained stable during the assay. Similar resultswere observed under shaken conditions.
3.3. Effect of the antimicrobial substances produced by L.
lactis CRL 1584 on C. freundii CFb viability
Analysis of the variance of the number of C. freundii
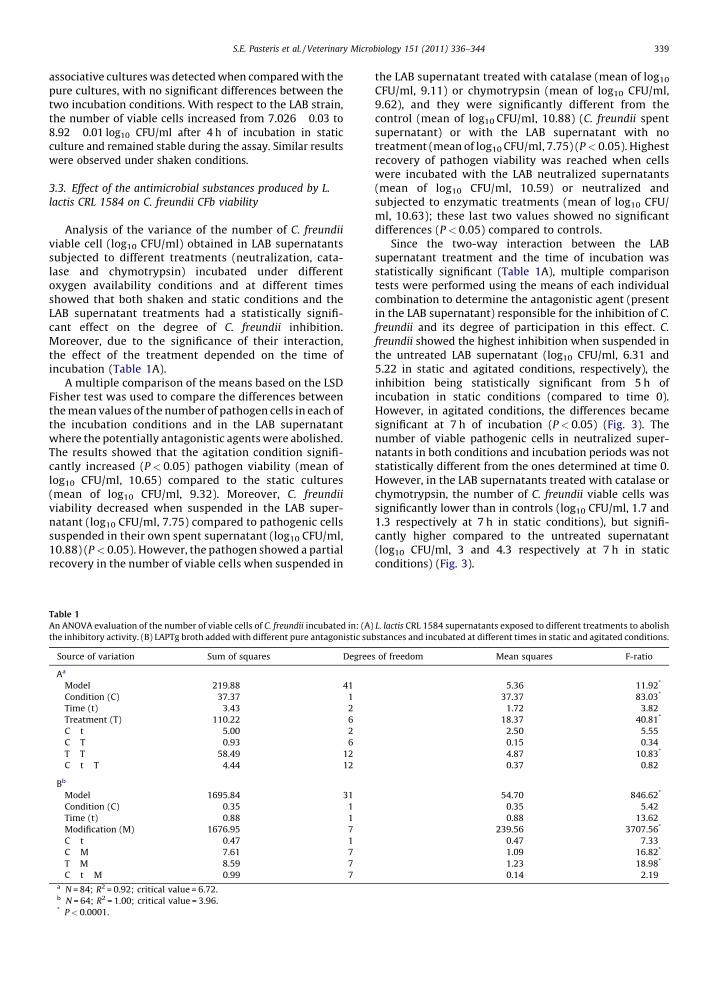
viable cell (log10 CFU/ml) obtained in LAB supernatantssubjected to different treatments (neutralization, cata-lase and chymotrypsin) incubated under differentoxygen availability conditions and at different timesshowed that both shaken and static conditions and theLAB supernatant treatments had a statistically signifi-cant effect on the degree of C. freundii inhibition.Moreover, due to the significance of their interaction,the effect of the treatment depended on the time ofincubation (Table 1A).
A multiple comparison of the means based on the LSDFisher test was used to compare the differences betweenthe mean values of the number of pathogen cells in each ofthe incubation conditions and in the LAB supernatantwhere the potentially antagonistic agents were abolished.The results showed that the agitation condition signifi-cantly increased (P < 0.05) pathogen viability (mean oflog10 CFU/ml, 10.65) compared to the static cultures(mean of log10 CFU/ml, 9.32). Moreover, C. freundii
viability decreased when suspended in the LAB super-natant (log10 CFU/ml, 7.75) compared to pathogenic cellssuspended in their own spent supernatant (log10 CFU/ml,10.88) (P < 0.05). However, the pathogen showed a partialrecovery in the number of viable cells when suspended in
the LAB supernatant treated with catalase (mean of log10
CFU/ml, 9.11) or chymotrypsin (mean of log10 CFU/ml,9.62), and they were significantly different from thecontrol (mean of log10 CFU/ml, 10.88) (C. freundii spentsupernatant) or with the LAB supernatant with notreatment (mean of log10 CFU/ml, 7.75) (P < 0.05). Highestrecovery of pathogen viability was reached when cellswere incubated with the LAB neutralized supernatants(mean of log10 CFU/ml, 10.59) or neutralized andsubjected to enzymatic treatments (mean of log10 CFU/ml, 10.63); these last two values showed no significantdifferences (P < 0.05) compared to controls.
Since the two-way interaction between the LABsupernatant treatment and the time of incubation wasstatistically significant (Table 1A), multiple comparisontests were performed using the means of each individualcombination to determine the antagonistic agent (presentin the LAB supernatant) responsible for the inhibition of C.
freundii and its degree of participation in this effect. C.
freundii showed the highest inhibition when suspended inthe untreated LAB supernatant (log10 CFU/ml, 6.31 and5.22 in static and agitated conditions, respectively), theinhibition being statistically significant from 5 h ofincubation in static conditions (compared to time 0).However, in agitated conditions, the differences becamesignificant at 7 h of incubation (P < 0.05) (Fig. 3). Thenumber of viable pathogenic cells in neutralized super-natants in both conditions and incubation periods was notstatistically different from the ones determined at time 0.However, in the LAB supernatants treated with catalase orchymotrypsin, the number of C. freundii viable cells wassignificantly lower than in controls (log10 CFU/ml, 1.7 and1.3 respectively at 7 h in static conditions), but signifi-cantly higher compared to the untreated supernatant(log10 CFU/ml, 3 and 4.3 respectively at 7 h in staticconditions) (Fig. 3).
Table 1
An ANOVA evaluation of the number of viable cells of C. freundii incubated in: (A) L. lactis CRL 1584 supernatants exposed to different treatments to abolish
the inhibitory activity. (B) LAPTg broth added with different pure antagonistic substances and incubated at different times in static and agitated conditions.
Source of variation Sum of squares Degrees of freedom Mean squares F-ratio
Aa
Model 219.88 41 5.36 11.92*
Condition (C) 37.37 1 37.37 83.03*
Time (t) 3.43 2 1.72 3.82
Treatment (T) 110.22 6 18.37 40.81*
C � t 5.00 2 2.50 5.55
C � T 0.93 6 0.15 0.34
T � T 58.49 12 4.87 10.83*
C � t � T 4.44 12 0.37 0.82
Bb
Model 1695.84 31 54.70 846.62*
Condition (C) 0.35 1 0.35 5.42
Time (t) 0.88 1 0.88 13.62
Modification (M) 1676.95 7 239.56 3707.56*
C � t 0.47 1 0.47 7.33
C � M 7.61 7 1.09 16.82*
T � M 8.59 7 1.23 18.98*
C � t � M 0.99 7 0.14 2.19a N = 84; R2 = 0.92; critical value = 6.72.b N = 64; R2 = 1.00; critical value = 3.96.* P < 0.0001.
S.E. Pasteris et al. / Veterinary Microbiology 151 (2011) 336–344 339
3.4. Effect of different antagonistic substances added to the
culture media on C. freundii CFb viability
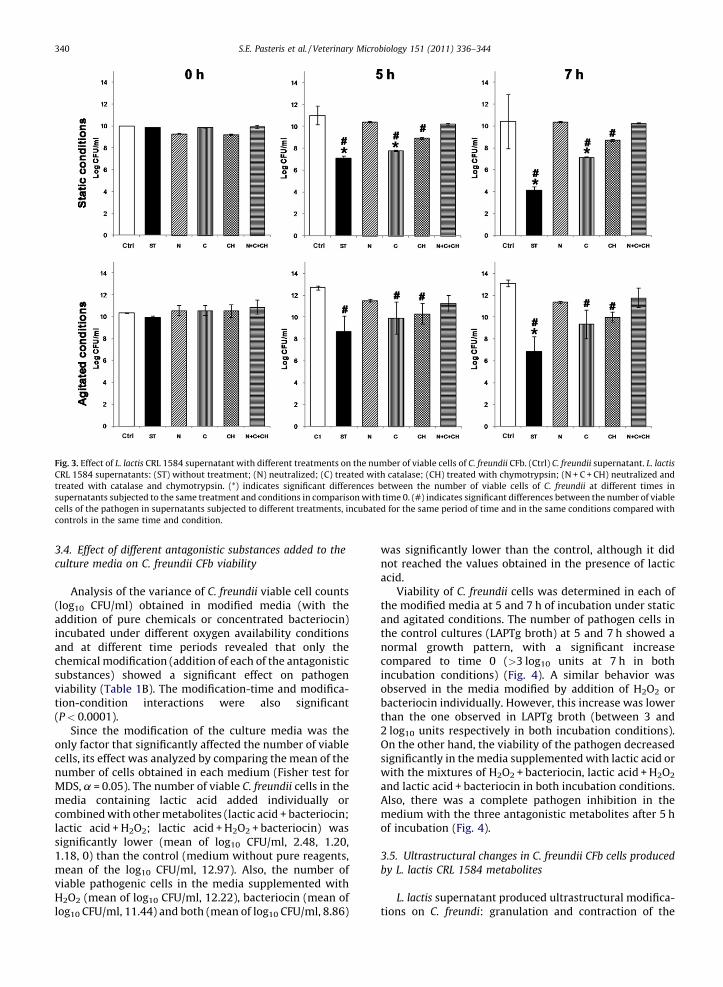
Analysis of the variance of C. freundii viable cell counts(log10 CFU/ml) obtained in modified media (with theaddition of pure chemicals or concentrated bacteriocin)incubated under different oxygen availability conditionsand at different time periods revealed that only thechemical modification (addition of each of the antagonisticsubstances) showed a significant effect on pathogenviability (Table 1B). The modification-time and modifica-tion-condition interactions were also significant(P < 0.0001).
Since the modification of the culture media was theonly factor that significantly affected the number of viablecells, its effect was analyzed by comparing the mean of thenumber of cells obtained in each medium (Fisher test forMDS, a = 0.05). The number of viable C. freundii cells in themedia containing lactic acid added individually orcombined with other metabolites (lactic acid + bacteriocin;lactic acid + H2O2; lactic acid + H2O2 + bacteriocin) wassignificantly lower (mean of log10 CFU/ml, 2.48, 1.20,1.18, 0) than the control (medium without pure reagents,mean of the log10 CFU/ml, 12.97). Also, the number ofviable pathogenic cells in the media supplemented withH2O2 (mean of log10 CFU/ml, 12.22), bacteriocin (mean oflog10 CFU/ml, 11.44) and both (mean of log10 CFU/ml, 8.86)
was significantly lower than the control, although it didnot reached the values obtained in the presence of lacticacid.
Viability of C. freundii cells was determined in each ofthe modified media at 5 and 7 h of incubation under staticand agitated conditions. The number of pathogen cells inthe control cultures (LAPTg broth) at 5 and 7 h showed anormal growth pattern, with a significant increasecompared to time 0 (>3 log10 units at 7 h in bothincubation conditions) (Fig. 4). A similar behavior wasobserved in the media modified by addition of H2O2 orbacteriocin individually. However, this increase was lowerthan the one observed in LAPTg broth (between 3 and2 log10 units respectively in both incubation conditions).On the other hand, the viability of the pathogen decreasedsignificantly in the media supplemented with lactic acid orwith the mixtures of H2O2 + bacteriocin, lactic acid + H2O2
and lactic acid + bacteriocin in both incubation conditions.Also, there was a complete pathogen inhibition in themedium with the three antagonistic metabolites after 5 hof incubation (Fig. 4).
3.5. Ultrastructural changes in C. freundii CFb cells produced
by L. lactis CRL 1584 metabolites
L. lactis supernatant produced ultrastructural modifica-tions on C. freundi: granulation and contraction of the
Fig. 3. Effect of L. lactis CRL 1584 supernatant with different treatments on the number of viable cells of C. freundii CFb. (Ctrl) C. freundii supernatant. L. lactis
CRL 1584 supernatants: (ST) without treatment; (N) neutralized; (C) treated with catalase; (CH) treated with chymotrypsin; (N + C + CH) neutralized and
treated with catalase and chymotrypsin. (*) indicates significant differences between the number of viable cells of C. freundii at different times in
supernatants subjected to the same treatment and conditions in comparison with time 0. (#) indicates significant differences between the number of viable
cells of the pathogen in supernatants subjected to different treatments, incubated for the same period of time and in the same conditions compared with
controls in the same time and condition.
S.E. Pasteris et al. / Veterinary Microbiology 151 (2011) 336–344340
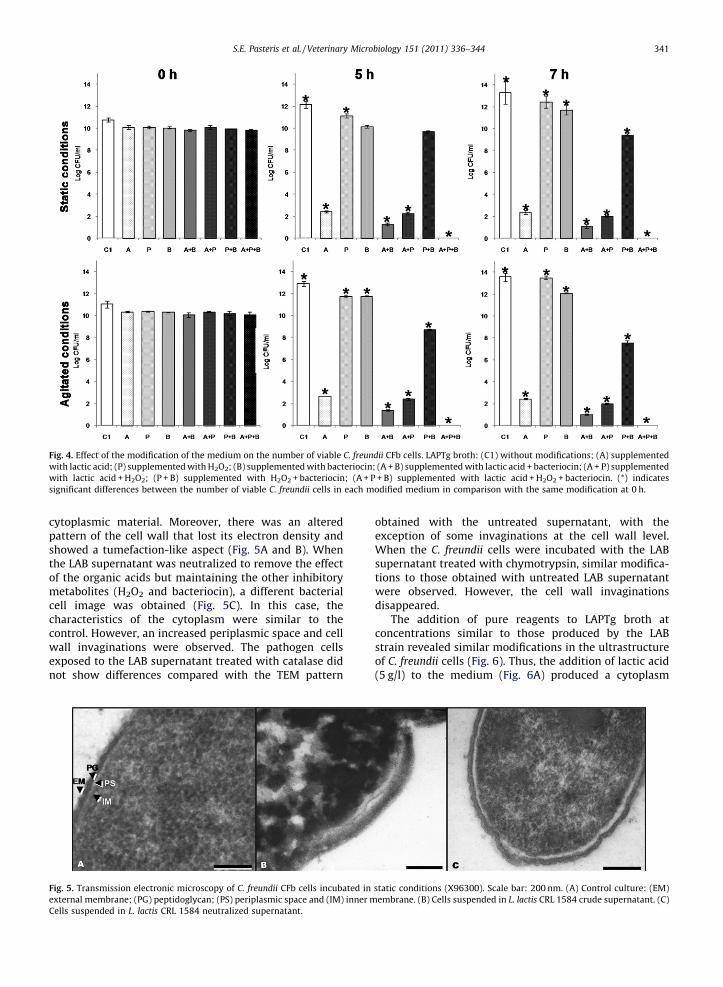
cytoplasmic material. Moreover, there was an alteredpattern of the cell wall that lost its electron density andshowed a tumefaction-like aspect (Fig. 5A and B). Whenthe LAB supernatant was neutralized to remove the effectof the organic acids but maintaining the other inhibitorymetabolites (H2O2 and bacteriocin), a different bacterialcell image was obtained (Fig. 5C). In this case, thecharacteristics of the cytoplasm were similar to thecontrol. However, an increased periplasmic space and cellwall invaginations were observed. The pathogen cellsexposed to the LAB supernatant treated with catalase didnot show differences compared with the TEM pattern
obtained with the untreated supernatant, with theexception of some invaginations at the cell wall level.When the C. freundii cells were incubated with the LABsupernatant treated with chymotrypsin, similar modifica-tions to those obtained with untreated LAB supernatantwere observed. However, the cell wall invaginationsdisappeared.
The addition of pure reagents to LAPTg broth atconcentrations similar to those produced by the LABstrain revealed similar modifications in the ultrastructureof C. freundii cells (Fig. 6). Thus, the addition of lactic acid(5 g/l) to the medium (Fig. 6A) produced a cytoplasm
Fig. 4. Effect of the modification of the medium on the number of viable C. freundii CFb cells. LAPTg broth: (C1) without modifications; (A) supplemented
with lactic acid; (P) supplemented with H2O2; (B) supplemented with bacteriocin; (A + B) supplemented with lactic acid + bacteriocin; (A + P) supplemented
with lactic acid + H2O2; (P + B) supplemented with H2O2 + bacteriocin; (A + P + B) supplemented with lactic acid + H2O2 + bacteriocin. (*) indicates
significant differences between the number of viable C. freundii cells in each modified medium in comparison with the same modification at 0 h.
Fig. 5. Transmission electronic microscopy of C. freundii CFb cells incubated in static conditions (X96300). Scale bar: 200 nm. (A) Control culture: (EM)
external membrane; (PG) peptidoglycan; (PS) periplasmic space and (IM) inner membrane. (B) Cells suspended in L. lactis CRL 1584 crude supernatant. (C)
Cells suspended in L. lactis CRL 1584 neutralized supernatant.
S.E. Pasteris et al. / Veterinary Microbiology 151 (2011) 336–344 341

granulation of the pathogen cells, while the periplasmicspace and the ultrastructure of the cell wall werepreserved. When LAPTg was supplemented with0.01 mM H2O2, the C. freundii cells showed cytoplasmiccontractions and therefore an enhancement of theperiplasmic space (Fig. 6B). However, there were nomodifications in either cytoplasm or cell wall ultrastruc-tures. When C. freundii cells were exposed to 2200 AU/mlbacteriocin, invaginations along the whole cellular surfacewere observed (Fig. 6C). Moreover, when C. freundii cellswere incubated with LAPTg supplemented with 0.01 mMH2O2 and 2200 AU/ml bacteriocin, cytoplasm contractions,enhancement of the periplasmic space and invaginationsalong the cell wall were evidenced (Fig. 6D).
4. Discussion
LAB are known to inhibit pathogens by the productionof antagonistic molecules such as organic acids, H2O2,bacteriocins and diacetyl (Verschuere et al., 2000). Theinhibitory effect of beneficial microorganisms on specificpathogens in aquaculture (Balcazar et al., 2006, 2007;Ringø et al., 2005, 2010) and the species specificity(Zoetendal et al., 2008) are two of the suggested criteriafor the selection and inclusion of specific bacteria in aprobiotic product. Our research group works in the designof probiotics for raniculture and therefore has previouslystudied the main cultivable microorganisms of theautochthonous microbiota of an L. catesteianus hatchery.The LAB genera with beneficial properties include Lacto-
coccus, Pediococcus and Lactobacillus (Pasteris et al., 2009a,
2009b). From these results, L. lactis CRL 1584 was selectedto determine the active inhibitory mechanisms on C.
freundii CFb, a pathogen associated with RLS outbreaks inraniculture, and to analyze the effect of oxygen availabilityon the inhibitory metabolite production by the LAB strain.Thus, an in vitro co-culture experimental model wasapplied to evaluate the interactions between both micro-organisms. During the first stage of this work, the ability ofL. lactis to produce lactic acid, H2O2 and bacteriocin wasdemonstrated. Lactic acid production reached its highestlevel at 6 and 7 h in agitated and static cultures,respectively, being higher in static conditions, as reportedby Shimizu et al. (1999). These results can be explained bythe fact that LAB are aerotolerant anaerobic microorgan-isms. Bacteriocin production reached its highest level at 5and 6 h in static and agitated cultures, respectively, andfrom this point onwards production decreased with time.Similar results were published by Hernandez de Rojas et al.(2004) for an L. lactis strain isolated from cheese. Althoughthere were no significant differences in bacteriocinproduction between shaken and static cultures, specificproduction at the time of maximum concentration washigher in agitated conditions (32.9 AU/107 CFU), asreported for some LAB strains by Vazquez et al. (2005).
There are no reports on the effect of culture conditions(oxygen availability) of LAB strains or further expression oftheir beneficial properties when included in a probioticproduct. Thus, it is important to increase knowledge of thein vitro expression of the beneficial properties of LABstrains by studying the best incubation conditions for theproduction of antagonistic molecules and, in the near
Fig. 6. Transmission electronic microscopy of C. freundii CFb cells incubated in static conditions (X96300). Scale bar: 200 nm. Cells were suspended in LAPTg
broth supplemented with: (A) 5 g/l lactic acid; (B) 0.01 mM H2O2; (C) 2200 AU/ml bacteriocin; (D) 2200 AU/ml bacteriocin + 0.01 mM H2O2.
S.E. Pasteris et al. / Veterinary Microbiology 151 (2011) 336–344342

future, to evaluate their effect during the administration ofL. lactis to frogs in hatchery conditions. Therefore, shakenconditions were also assayed in this work to stimulateH2O2 production since some LAB strains express thisproperty in agitated cultures (Juarez Tomas et al., 2003).Although H2O2 was not detected by the quantitative assay,the qualitative method showed that L. lactis produces thismetabolite. Moreover, treatment of the LAB supernatantswith catalase showed an increase in the number of viable C.
freundii cells (Fig. 3). Thus, these results allowed us tosuppose that H2O2 is released at lower concentrations thanthe detection limit of the method (0.05–0.1 mM).
It is important to emphasize that L. lactis CRL 1584produces three different antagonistic metabolites thatcould inhibit RLS-associated pathogens and food-bornebacteria (Pasteris et al., 2009a). Other L. lactis strains wereproposed as probiotics in fish aquaculture according totheir in vitro competitive adhesion and inhibitory activityagainst pathogens (Balcazar et al., 2007; Sequeiros et al.,2010).
There are no previous reports on the microbialinteractions between LAB strains and specific pathogensassociated with amphibians or related environments. Theresults showed that C. freundii was inhibited earlier whenco-cultured with L. lactis in agitated cultures. Thisinhibition could be due to an increase in H2O2 productionin shaken conditions, as described by Juarez Tomas et al.(2003). Hydrogen peroxide could participate in thesensitivity of bacterial cells to the action of other inhibitorycompounds produced by the LAB strain.
The decrease in the pH values of the associative cultureswas higher in static conditions. Lower variations in pHtogether with a faster pathogen inhibition, as observed inagitated conditions, would indicate that lactic acid was notthe only antagonistic agent involved in the decrease inpathogen viability.
To assess the effect of each antagonist metaboliteproduced by L. lactis on the growth inhibition of C. freundii,assays were performed to eliminate the effect of H2O2 orbacteriocin. The results showed that the number of viablecells of the pathogen decreased in both cases, with valueshigher that the ones for the untreated LAB supernatant.This would indicate that both H2O2 and bacteriocinparticipate in the inhibition of C. freundii. However, whenthe LAB supernatants were neutralized, the number ofviable C. freundii cells was similar to the controls in spite ofthe presence of H2O2 and bacteriocin. Thus, it is possible topostulate that the presence of organic acids in the mediumcould affect some membrane proteins, causing a modifica-tion in selective permeability that increases sensitivity tothe other antimicrobial metabolites that dissipate protonmotive force, as evidenced by Bender et al. (1986).Moreover, it has been reported that lactic acid permea-bilizes the outer membrane of Escherichia coli, increasingits sensitivity towards other antimicrobials (Alakomi et al.,2000).
Experiments were performed adding the pure com-mercial reagents or bacteriocin (as concentrated super-natant) to the fresh culture media to determine theindividual effect of the three metabolites detected in theLAB supernatant on pathogen viability. The highest
inhibitory effect on C. freundii was exerted by lactic acid.However, it was necessary to add the three antimicrobialmetabolites together to obtain the complete inhibition ofthe pathogenic bacteria, as reported for Staphylococcus
aureus by Otero and Nader-Macias (2006).The damages detected in the ultrastructure of C. freundii
cells were correlated with the decrease in the number ofviable cells. Thus, extreme structural modifications weredetected in the pathogenic bacterium in contact with thecrude LAB supernatant for 7 h that were reverted when thepathogen was suspended in neutralized, catalase- orchymotrypsin-treated LAB supernatants. These resultssupport the idea that the inhibition of C. freundii is dueto a synergistic effect between the three inhibitorymetabolites produced by L. lactis.
All the ultrastructural alterations evidenced in thepathogenic cells resuspended in the LAB supernatant werealso observed in the assays performed with LAPTg brothsupplemented with pure reagents. Thus, the addition oflactic acid modified the pathogenic cells by generatingcytosolic agglomerations without apparent changes in theultrastructure of the cell wall or the plasma membrane.Similar results were observed in S. aureus cells co-culturedwith Lactobacillus gasseri (Otero and Nader-Macias, 2006).When LAPTg broth was supplemented with H2O2, thepathogenic cells showed a larger periplasmic space. Theseobservations agree with the results of Otero and Nader-Macias (2006), who studied the effect of lactic acid andH2O2 on the ultrastructure of S. aureus cells. This effectcould be due to the oxidations that H2O2 causes on plasmamembrane components, mainly on fatty acids and someproteins (Watts et al., 2003). The addition of bacteriocin tothe medium with C. freundii cells produced an alteration inthe cell wall, which presents periodic waves along itssurface. Similar results were published by Hyde et al.(2006) for Bacillus subtilis cells exposed to purified nisin Z.
5. Conclusion
This work describes for the first time the microbialinteractions and the cellular modifications that occurduring inhibition of an RLS-associated pathogen by L. lactis
subsp. lactis CRL 1584, an autochthonous microorganismisolated from an L. catesbeianus hatchery. These findingsand the beneficial properties of this specific LAB strain(Pasteris et al., 2009a) indicate the need for further studiesto determine if could be included in a probiotic productaimed at increasing raniculture productivity. Besides,additional studies are required to determine whether L.
lactis will be used as a pure or multi strain probiotic,depending on its behavior against other LAB strainspreviously selected on the basis of their beneficialproperties.
References
Alakomi, H.L., Skytta, E., Saarela, M., Mattila-Sandholm, T., Latva-Kala, K.,Helander, I.M., 2000. Lactic acid permeabilizes gram-negative bac-teria by disrupting the outer membrane. Appl. Environ. Microbiol. 66,2001–2005.
S.E. Pasteris et al. / Veterinary Microbiology 151 (2011) 336–344 343

Balcazar, J.L., de Blas, I., Ruiz-Zarzuela, I., Cunningham, D., Vendrell, D.,Muzquiz, J.L., 2006. The role of probiotic in aquaculture. Vet. Micro-biol. 114, 173–186.
Balcazar, J.L., Vendrell, D., de Blas, I., Ruiz-Zarzuela, I., Girones, O., Muz-quiz, J.L., 2007. In vitro competitive adhesion and production ofantagonistic compounds by lactic acid bacteria against fish patho-gens. Vet. Microbiol. 122, 373–380.
Bender, G.R., Sutton, S.W., Marquis, R.W., 1986. Acid tolerance, protonpermeabilities, and membrane ATPases of oral streptococci. Infect.Immun. 53 (2), 331–338.
de Man, J.C., Rogosa, M., Sharpe, E., 1969. A medium for cultivation oflactobacilli. J. Appl. Bacteriol. 23, 130–145.
Densmore, C.L., Earl Green, D., 2007. Diseases of amphibians. ILAR J. 48 (3),235–254.
FAO/WHO., 2001. Health and nutritional properties of probiotics in foodincluding powder milk with live lactic acid bacteria. Report of ajoint FAO/WHO expert consultation. www.who.int/foodsafety/publications/fs_management/en/probiotics.pdf
Farzanfar, A., 2006. The use of probiotics in shrimp aquaculture. FEMSImmunol. Med. Microbiol. 48, 149–158.
Frost, D.R., Grant, T., Faivovich, J., Bain, R.H., Haas, A., Haddad, C.F.B., de Sa,S.O., Channing, A., Wilkinson, M., Donnellan, S.C., Raxworthy, C.J.,Campbell, J.A., Blotto, B.L., Moler, P., Drewes, R.C., Nussbaum, R.A.,Lynch, J.D., Green, D.M., Wheeler, W.C., 2006. The amphibian tree oflife. Bull. Am. Mus. Nat. Hist. 297.
Gatesoupe, F.J., 2008. Updating the importance of lactic acid bacteria infish farming: natural occurrence and probiotic treatments. J. Mol.Microbiol. 14 (1–3), 107–114.
Glorioso, J.C., Amborski, R.L., Larkin, J.M., Amborski, G.F., Culley, D.C.,1974. Microbiological studies on septicemic bullfrogs (Rana catesbei-ana). Am. J. Vet. Res. 35 (3), 447–450.
Hernandez de Rojas, A., Martınez, B., Suarez, J.E., Rodrıguez, A., 2004.Enhanced production of lactococcin 972 in chemostat cultures. Appl.Microbiol. Biotechnol. 66, 48–52.
Hyde, A.J., Parisot, J., McNichol, A., Bonev, B.B., 2006. Nisin-inducedchanges in Bacillus morphology suggest a paradigm of antibioticaction. Proc. Natl. Acad. Sci U.S.A. 103 (52), 19896–19901.
Irianto, A., Austin, B., 2002. Probiotics for aquaculture: a review. J. Fish Dis.25, 633–642.
Juarez Tomas, M.S., Bru, E., Nader-Macıas, M.E., 2003. Comparison of thegrowth and H2O2-production by vaginal probiotic lactobacilli underdifferent culture conditions. Am. J. Obstet. Gynecol. 188, 35–44.
Juarez Tomas, M.S., Otero, M.C., Ocana, V.S., Nader-Macıas, M.E., 2004.Production of antimicrobial substances in lactic acid bacteria. Deter-mination of hydrogen peroxide. In: Spencer, J.F.T., Ragout de Spencer,A.L. (Eds.), Methods in Molecular Biology. Public Health Microbiology:Methods and Protocols, 268. Humana Press, Inc., Totowa, pp. 337–346.
Mauel, M.J., Miller, D.L., Frazier, K.S., Hines, M.E., 2002. Bacterial patho-gens isolated from cultured bullfrog (Rana catesbeiana). J. Vet. Diagn.Invest. 14 (5), 431–433.
Nikoskelainen, S., Ouwehand, A., Salminen, S., Bylund, G., 2001. Protectionof rainbow trout (Oncorhynchus mykiss) from furunculosis by Lacto-bacillus rhamnosus. Aquaculture 198, 229–236.
Otero, M.C., Nader-Macias, M.E., 2006. Inhibition of Staphylococcus aureusby H2O2-producing Lactobacillus gasseri isolated from the vaginal tractof cattle. Anim. Reprod. Sci. 96 (1), 35–46.
Pasteris, S.E., Buhler, M.I., Nader-Macıas, M.E., 2006. Microbiological andhistological studies in farmed-bullfrog (Rana catesbeiana) displayingred-leg syndrome. Aquaculture 251, 11–18.
Pasteris, S.E., Gonzalez, A., Van Schoor, A., Buhler, M.I., Nader-Macıas, M.E.,Vandamme, P., DeVuyst, L., 2008. Genotypic identification of lacticacid bacteria from a Rana catesbeiana hatchery. In: Abstr. TEC 13 p. 88.V Congreso Argentino de Microbiologıa General (SAMIGE), Santa Fe,Argentina.
Pasteris, S.E., Roig Babot, G., Otero, M.C., Nader-Macıas, M.E., 2009a.Beneficial properties of lactic acid bacteria isolated from a Ranacatesbeiana hatchery. Aquac. Res. 40, 1605–1615.
Pasteris, S.E., Vera Pingitore, E., Roig Babot, G., Otero, M.C., Buhler, M.I.,Nader-Macıas, M.E., 2009b. Characterization of the beneficial proper-ties of lactobacilli isolated from bullfrog (Rana catesbeiana) hatchery.Antonie van Leeuwenhoek 95, 375–385.
Raibaud, P., Gapin, J.V., Ducluzeau, R., Mocquot, G., Oliver, G., 1963. Legenre Lactobacillus dans le tube digestif du rat. II Caracteres desouches heterofermentaires isolees de rats ‘‘Holo’’ et ‘‘Gnotoxeni-ques’’ Ann. Microbiol. (Annales de L’Institut Pasteur) 124, 2223–2235.
Ringø, E., Løvmo, L., Kristiansen, M., Bakken, Y., Salinas, I., Myklebust, R.,Olsen, R.E., Mayhew, T.M., 2010. Lactic acid bacteria vs. pathogens inthe gastrointestinal tract of fish. Aquac. Res. 41 (4), 451–467.
Ringø, E., Schillinger, U., Holzalpfel, W., 2005. Antimicrobial activity oflactic acid bacteria isolated from aquatic animals and the use of lacticacid bacteria in aquaculture. In: Pierzynowski, S.G., Zabielski, R.(Eds.), Biology in Growing Animals Series. Elsevier, Oxford,England, pp. 408–443.
Schadich, E., Cole, A.L.J., 2009. Inhibition of frog antimicrobial peptides byextracellular products of the bacterial pathogen Aeromonas hydro-phila. Lett. Appl. Microbiol. 49, 384–387.
Sequeiros, C., Vallejo, M., Marguet, E.R., Olivera, N.L., 2010. Inhibitoryactivity against the fish pathogen Lactococcus garvieae produced byLactococcus lactis TW34, a lactic acid bacterium isolated from theintestinal tract of a Patagonian fish. Arch. Microbiol. 192, 237–245.
Shimizu, H., Mizuguchi, T., Tanaka, E., Shioya, S., 1999. Nisin production bya mixed-culture system consisting of Lactococcus lactis and Kluyver-omyces marxianus. Appl. Environ. Microbiol. 65 (7), 3134–3141.
Texeira, R.D., Pereira Mello, S.C.R., Lima dos Santos, C.A.M., 2002. Theworld market for frog legs. Globefish 68, 1–44.
Vazquez, J.A., Miron, J., Gonzalez, M.P., Murado, M.A., 2005. Effects ofaeration on growth and on production of bacteriocins and othermetabolites in cultures of eight strains of lactic acid bacteria. Bio-technol. Appl. Biochem. 127 (2), 111–124.
Verschuere, L., Rombaut, G., Zorruelos, P., Verstraete, W., 2000. Probioticbacteria as biological control agents in aquaculture. Microbiol. Mol.Biol. Rev. 64 (4), 655–671.
Watts, R.J., Washington, D., Howsawkeng, J., Loge, F.J., Teel, A.L., 2003.Comparative toxicity of hydrogen peroxide, hydroxyl radicals, andsuperoxide anion to Escherichia coli. Adv. Environ. Res. 7, 961–968.
Zhen-Yu, X., Yong-Can, Z., Shi-Feng, W., Bing, M., Xian-Dong, X., Wan-Yao,W., Yong-Qin, F., 2009. First isolation and identification of Elizabeth-kingia meningoseptica from cultured tiger frog, Rana tigerina rugulosa.Vet. Microbiol. 138, 140–144.
Zoetendal, E.G., Rajilic-Stojanovic, M., de Vos, W.M., 2008. High-through-put diversity and functionality analysis of the gastrointestinal tractmicrobiota. Gut 57 (11), 1605–1615.
S.E. Pasteris et al. / Veterinary Microbiology 151 (2011) 336–344344












![Public Input No. 11-NFPA 1584-2012 [ Section No. 3.3.21 ]](https://static.fdokumen.com/doc/165x107/632e2442adaa8ed39e01a710/public-input-no-11-nfpa-1584-2012-section-no-3321-.jpg)







