
Detection of Bifidobacterium animalis subsp. lactis (Bb12) in the Intestine after Feeding of Sows...
11
Published Ahead of Print 8 August 2008. 10.1128/AEM.00309-08. 2008, 74(20):6338. DOI: Appl. Environ. Microbiol. Andrews, Bryan Vinyard and Joseph F. Urban Jr. Gloria Solano-Aguilar, Harry Dawson, Marta Restrepo, Kate Feeding of Sows and Their Piglets (Bb12) in the Intestine after lactis subsp. Bifidobacterium animalis Detection of http://aem.asm.org/content/74/20/6338 Updated information and services can be found at: These include: REFERENCES http://aem.asm.org/content/74/20/6338#ref-list-1 at: This article cites 54 articles, 22 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on March 13, 2014 by guest http://aem.asm.org/ Downloaded from on March 13, 2014 by guest http://aem.asm.org/ Downloaded from
Transcript of Detection of Bifidobacterium animalis subsp. lactis (Bb12) in the Intestine after Feeding of Sows...

Published Ahead of Print 8 August 2008. 10.1128/AEM.00309-08.
2008, 74(20):6338. DOI:Appl. Environ. Microbiol. Andrews, Bryan Vinyard and Joseph F. Urban Jr.Gloria Solano-Aguilar, Harry Dawson, Marta Restrepo, Kate Feeding of Sows and Their Piglets
(Bb12) in the Intestine afterlactissubsp. Bifidobacterium animalisDetection of
http://aem.asm.org/content/74/20/6338Updated information and services can be found at:
These include:
REFERENCEShttp://aem.asm.org/content/74/20/6338#ref-list-1at:
This article cites 54 articles, 22 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on March 13, 2014 by guest
http://aem.asm
.org/D
ownloaded from
on M
arch 13, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Oct. 2008, p. 6338–6347 Vol. 74, No. 200099-2240/08/$08.00�0 doi:10.1128/AEM.00309-08
Detection of Bifidobacterium animalis subsp. lactis (Bb12) in theIntestine after Feeding of Sows and Their Piglets�
Gloria Solano-Aguilar,1* Harry Dawson,1 Marta Restrepo,1 Kate Andrews,2Bryan Vinyard,3 and Joseph F. Urban, Jr.1
Diet, Genomics, and Immunology Laboratory, Beltsville Human Nutrition Research Center, Agricultural Research Service, U.S. Department ofAgriculture, Beltsville, Maryland1; Food and Feed Safety Research Unit, Southern Plains Agricultural Research Center, U.S. Department of
Agriculture, College Station, Texas2; and Biometrical Consulting Service, Beltsville Agricultural Research Center,U.S. Department of Agriculture, Beltsville, Maryland3
Received 5 February 2008/Accepted 4 August 2008
A real-time PCR method has been developed to distinguish Bifidobacterium animalis subspecies in thegastrointestinal tracts of pigs. Identification of a highly conserved single-copy tuf gene encoding the elongationfactor Tu involved in bacterial protein biosynthesis was used as a marker to differentiate homologous Bi-fidobacterium animalis subsp. lactis (strain Bb12) from Bifidobacterium animalis subsp. animalis, as well asBifidobacterium suis, Bifidobacterium breve, Bifidobacterium longum, several species of Lactobacillus, and Entero-coccus faecium. Real-time PCR detection of serially diluted DNA extracted from a pure culture of Bb12 waslinear for bacterial numbers ranging from 10 to 10,000 tuf gene copies per PCR (r2 � 0.99). Relative differencesin Bb12 bacterial numbers in pigs fed daily with Bb12 were determined after detection of Bb12 tuf gene copiesin DNA extracted from the intestinal contents. Piglets treated with Bb12 immediately after birth maintaineda high level of Bb12 in their large intestines with continuous daily administration of Bb12. Piglets born toBb12-treated sows during the last third of their gestation and also treated with Bb12 at birth (T/T group) hada higher number of Bb12 organisms per gram of intestinal contents compared to placebo-treated piglets bornto placebo-treated sows (C/C group), Bb12-treated sows (T/C group), or piglets born to placebo sows buttreated with Bb12 immediately after birth (C/T group). In addition, there was a significant increase in geneexpression for Toll-like receptor 9 (TLR9) in piglets from the T/T group, with no change in TLR2 and TLR4.These findings suggest that the tuf gene represents a specific and functional marker for detecting Bifidobac-terium animalis subsp. lactis strain Bb12 within the microbiota of the intestine.
Bifidobacteria are anaerobic, gram-positive, non-spore-forming, non-motile bacilli commonly found in the gastroin-testinal tracts (GITs) of animals, including humans (1). Bi-fidobacteria are the predominant bacterial species in the GITsof infants; they represent about 3% of the total microbiota inthe intestine of healthy adult humans (16) and are associatedwith beneficial health effects (15, 16, 30, 32, 36). Despite thegeneral acceptance of bifidobacteria as a probiotic, and theiruse in health-promoting foods such as fermented milks, infantformula, cheese, and ice cream, there is little definitive infor-mation to support a mechanism of action. Stimulation of hostresistance, immune modulation, and competitive exclusion ofpathogens, however, have been proposed as likely mechanisms(40).
One of the Bifidobacterium species commonly used in thefood industry is Bifidobacterium animalis subsp. lactis strainBb12, which is marketed around the world under a variety oflabels in dairy products and infant formulas (37, 40). Thetaxonomy of B. animalis subsp. lactis has been controversialsince its original description by Meile et al. in 1997 (27), and
several studies have investigated its similarity with the closelyrelated species Bifidobacterium animalis subsp. animalis (41).New genotypic evidence reported by Ventura et al. (46–48),Zhu and Dong (54), Masco et al. (23), and Kwon et al. (19)indicate that B. animalis subsp. lactis and B. animalis subsp.animalis should be considered two separate taxonomic entitiesat the subspecies level. B. animalis subsp. lactis exhibits prop-erties such as elevated oxygen tolerance (34), differentialgrowth in milk-based media (46), and hydrolysis of milk pro-teins (13); these properties differ from B. animalis subsp. ani-malis and facilitate its growth in commercial products undernonanaerobic conditions. Traditional bacteriological and bio-chemical identification techniques, such as selective growth ofspecies in differential media, cannot be routinely used to dif-ferentiate Bifidobacterium species. These methods are timeconsuming and limited by low sensitivity and reproducibilitydue to the multitudes of species that grow and require furtheridentification (24). In addition, the information obtained byculture-based growth methods provides only a fragmented pic-ture of the relative distribution of species within the GITbecause a significant part of its microbiota cannot be grown invitro (43, 55). Culture-independent methods have been devel-oped in recent years as an alternative to characterize wholebacterial communities by direct extraction of DNA from fecalsamples without prior cultivation (6). These methods includefluorescent in situ hybridization, dot blot hybridizations, andDNA arrays and fingerprinting methods such as terminal re-striction fragment length polymorphism and denaturating or
* Corresponding author. Mailing address: Diet, Genomics, and Im-munology Laboratory, Beltsville Human Nutrition Research Center,Agricultural Research Service, U.S. Department of Agriculture, 10300Baltimore Avenue, BARC-East, Bldg. 307C, Rm. 225, Beltsville, MD20705. Phone: (301) 504-8068. Fax: (301) 504-9062. E-mail: [email protected].
� Published ahead of print on 8 August 2008.
6338
on March 13, 2014 by guest
http://aem.asm
.org/D
ownloaded from

temperature gradient gel electrophoresis (26). Conserved andvariable regions within the 16S ribosomal gene are widely usedas markers to study bacterial diversity by PCR with sensitivitythat is approximately 100 times greater than that of traditionalculture-based and fluorescent in situ hybridization methods(25). The 16S ribosomal gene variable regions may be utilizedfor genus or species differentiation if species-specific probescan be designed (19, 25, 39). Highly homologous species like B.animalis subsp. lactis, however, are not generally distinguishedby 16S ribosomal gene-based differentiation (28, 38, 53) orquantitative assessment. In addition, the use of 16S ribosomalgene-based probes for quantitative real-time PCR remains dif-ficult because the copies of ribosomal DNA per genome canvary (5). An alternative to 16S ribosomal gene-based analysisof Bifidobacterium species is comparing conserved protein cod-ing sequences of bacterial genes, such as those for transaldo-lase (35), recA (17, 47), hsp60 (14, 54), groEL (49), groES (49),tuf (48), atpD (49), dnak (50), or xfp (52). After comparing thediscriminating properties of each of these sequences, we se-lected the highly conserved and ubiquitous tuf gene encodingthe elongation factor Tu that facilitates the elongation ofpolypeptides from the ribosome and aminoacyl tRNA duringtranslation and that has been used as a phylogenetic marker foreubacteria (31, 33). The tuf gene is universally distributed inBifidobacterium and Lactobacillus species, and only one tufgene per bacterial genome has been found (7, 48). It was theonly gene that was able to discriminate closely related B. ani-malis isolates at the subspecies level. The objective of this workwas to validate the use of a real-time PCR assay to detect thesingle-copy tuf gene of B. animalis subsp. lactis as a marker forBb12 in the GITs of pigs orally treated with Bb12. In addition,the effect of different dietary exposures to Bb12 on the numberof organisms detected in the intestinal contents of pigs and onthe host innate immune response was assessed by measuringthe localized gene expression of Toll-like receptors (TLRs).TLR2, -4, and -9 were chosen because of their abilities to bindto bacterial products that activate the innate immune systemand influence the course of acquired immunity (2). We dem-onstrate, for the first time, the use of the tuf gene to quanti-tatively detect Bb12 in the GITs of Bb12-treated piglets andthe in vivo effect on host TLR expression. There is also asignificant maternal effect on Bb12-treated piglets from Bb12-treated sows that results in higher and more persistent levels inthe proximal colon. The sensitivity of the molecular assay usedto detect B. animalis subsp. lactis should prove useful insupporting health benefits associated with the probiotic ef-ficacy of Bb12.
MATERIALS AND METHODS
Reference strains and culture conditions. The bacterial strains used to designand validate the quantitative assay for B. animalis subsp. lactis (strain Bb12) arelisted in Table 1. All bifidobacteria were cultured on Bifidobacterium-selectivemedium (Bifido; Anaerobe Systems, Morgan Hill, CA). Lactobacilli were cul-tured on Lactobacillus-selective MRS agar (Difco Laboratories, Detroit, MI),and enterococci were cultured on Enterococcus-selective agar (Difco Laborato-ries, Detroit, MI). The plates were incubated at 37°C for 3 days in a Bactron IVanaerobic chamber (Sheldon Manufacturing, Inc., Cornelius, OR). Colony for-mation was examined with a stereoscopic microscope, and bacterial character-istics were determined by Gram staining. Bacteria were swabbed from the plate,resuspended in 1 ml of sterile phosphate-buffered saline (PBS), and stored at�20°C until required for further processing for DNA extraction. Serial 10-folddilutions of 1 g of lyophilized B. animalis subsp. lactis strain Bb12 organisms,
provided by Chr. Hansen (Milwaukee, WI), were plated on anaerobic Brucellablood agar (BRU; Anaerobe Systems) and Bifido plates to test purity. The plateswere subsequently incubated at 37°C for 3 days in an anaerobic chamber with agas mixture of 5% CO2, 5% hydrogen, and 90% nitrogen. Bacteria were iden-tified by Gram staining and with an API 20A anaerobe identification strip(bioMerieux, Hazelwood, MO). CFU were determined in duplicate. The highestdilution showing growth of bacteria was used for the final CFU determination.
Study design and sample collection. The presence of Bb12 was evaluated inintestinal contents or fecal samples taken from Bb12-treated or placebo-treatedpigs from three independent experiments using pregnant sows and their litters.In the first experiment, four pregnant sows were given an oral daily dose of 5 g(3.7 � 1010 CFU/sow/day) of freeze-dried Bb12 during the last third of theirpregnancy. Four additional sows were given an oral daily dose with 5 g of theplacebo containing only the vehicle used in the probiotic product. Immediatelyafter birth, piglets born from each sow received either a daily probiotic treatmentof 1.5 g (1.05 � 1010 CFU/day) or an equivalent amount of placebo containingonly the vehicle for 32 days. This experimental design gave the following fourdifferent experimental groups of pigs: (i) Bb12-treated sows and Bb12-treatedpiglets (T/T) (n � 18); (ii) Bb12-treated sows and placebo control-treated piglets(T/C) (n � 16); (iii) placebo control-treated sows and Bb12-treated piglets (C/T)(n � 21); and (iv) placebo control-treated sows and placebo control-treatedpiglets (C/C) (n � 19). An additional fifth litter (n � 9) of untreated piglets froman untreated sow located in a different region of the farrowing barn was used asa negative control for probiotic and placebo treatment. All 83 piglets wereweaned at day 21 and euthanized on day 32 after birth. Five grams of intestinalcontents from the proximal colon was taken immediately after necropsy and keptfrozen at �20°C until further processing for DNA extraction. Both products(probiotic and placebo) were microbiologically tested throughout the experimentfor purity and stability of the bacteria in the probiotic product and for theabsence of bacterial growth in the placebo product.
In a second replicate experiment, fecal samples were collected from a total of20 piglets born to sows treated with either Bb12 (3.7 � 1010 CFU/sow/day) orplacebo during the last third of gestation. Piglets also received a daily dose (1.05 �1010 CFU/day) of lyophilized Bb12 or the equivalent amount of placebo startingat birth and given until day 23 (the weaning date). This experimental designassigned five piglets per experimental group (T/T, T/C, C/T, and C/C). Freshfecal samples were aseptically collected in a sterile 50-ml conical tube after pigswere manually stimulated to defecate. At least 1 g of feces was collected at 10,23, 32, and 45 days of age. Samples were immediately stored at �20°C and keptfrozen until DNA extraction.
In a third replicate experiment, three sows were orally inoculated daily duringthe last third of their pregnancy and through weaning of their piglets at 19 daysafter birth with 2.2 g of freeze-dried Bb12 (3.52 � 1010 CFU/sow/day). Threeadditional sows were inoculated with the same amount of a placebo preparation.Piglets within each litter were randomly divided at birth into two groups, wherehalf of the litter received a daily probiotic treatment of 1.1 g (1.76 � 1010
CFU/pig/day) and the other half received an equivalent amount of placebo
TABLE 1. Bacterial strains examined
Species Straina Sourceb
BifidobacteriumB. animalis subsp. lactis Bb12 Chr. Hansen
27536 ATCCB. animalis subsp. animalis 25527T ATCCB. suis 27531 ATCCB. breve 15700T ATCCB. longum 15707T ATCC
BB46 Chr. Hansen
OtherEnterococcus faecium SF273 Chr. HansenLactobacillus paracasei LC-01 Chr. HansenLactobacillus bulgaricus LBA40 Chr. HansenLactobacillus acidophilus 53544 ATCC
53545 ATCCLA05 Chr. Hansen
a T, T type strain.b Chr. Hansen strains were obtained from the Chr. Hansen Collection. All
other strains were obtained from ATCC.
VOL. 74, 2008 DETECTION OF Bb12 IN INTESTINES OF SOWS AND PIGLETS 6339
on March 13, 2014 by guest
http://aem.asm
.org/D
ownloaded from

through weaning at 19 days after birth and for an additional 72 days postweaning.This experimental design gave four groups of pigs that included the following: (i)T/T piglets (n � 14); (ii) T/C piglets (n � 17); (iii) C/T piglets (n � 13); and (iv)C/C piglets (n � 13). Five grams of intestinal contents from the proximal colonwas collected at necropsy and stored at �20°C until needed for further process-ing for DNA extraction. Sets of three to five piglets were euthanized at 7, 19, 32,and 91 days after birth, and intestinal samples were collected from the proximalcolon along with a 2-cm2 tissue section of intestinal mucosa. All animal proce-dures were approved by the Beltsville Area Animal Care and Use Committee.
DNA extraction. Reference bacteria cell suspensions (Table 1) were centri-fuged for 10 min at 5,000 � g. The bacterial pellet was resuspended and incu-bated for 30 min at 37°C with enzymatic lysis buffer (20 mM Tris-Cl [pH 8.0], 2mM sodium EDTA, 1.2% Triton X-100, and 20 mg/ml lysozyme [Sigma, MO]),followed by an incubation with proteinase K and buffer for 30 min at 70°C. Afterenzymatic lysis was carried out, bacterial DNA was isolated from the samplesusing the DNeasy tissue kit (Qiagen, Valencia, CA) according to the instructionsof the manufacturer. The DNA was eluted in TE buffer (10 mM Tris-HCl, 1 mMEDTA [pH 8.0]).
Similarly, DNA from the fecal contents of animals was isolated using theQIAamp DNA stool mini kit (Qiagen, Valencia, CA). Briefly, 1 g of homoge-nized contents from different intestinal sites was thawed, weighed, and resus-pended with lysis buffer. After heating the suspension at 95°C for increased DNAyield, removal of inhibitors and proteinase K digestion were done before DNAwas bound to a column, washed, and eluted in TE buffer. DNA concentrationwas determined by spectrophotometry. An aliquot of 100 ng of DNA from eachextraction was used as a template for all bacterial quantifications.
Primers specific for Bifidobacterium animalis subsp. animalis. Complete andpartial sequences of the 16S to 23S intergenic spacer region of several Bifidobac-terium species and Enterococcus faecium were retrieved from GenBank (Table 2)to develop primers and probes for 5� nuclease assays. All published sequences inGenBank for the dnaK (50), groES, groEL, atpD (49), tuf (47), recA (47), hsp60(14), and transaldolase (35) genes of Bifidobacterium species were aligned usingthe Clustal program (44), and the overall nonconserved regions of these se-quences were used to design primers and probes for the detection of B. animalissubsp. animalis. To increase the specificity and sensitivity of the assay, TaqManminor groove binding probes were used. The primers and probes were designedusing Primer Express software (Applied Biosystems, CA). The oligonucleotideprobes designed were labeled with the 5� reporter dye 5�-tetrachloro-fluoresceinphosphoramidite and the 3� quencher BHQ1 (Biosource, CA) (Table 3). Fordetermination of the total bacterial load, a previously described set of primersand probes for the 16S rRNA genes of eubacteria labeled with the 5� reporterdye 6-carboxyfluorescein and the 3� quencher NFQ-MGB (Applied Biosystems,CA) was used (29). Similarly, the primers and probes used to detect totalBifidobacterium spp. (10) and Lactobacillus spp. (12) were used as describedpreviously. All primers were tested for specificity using the Clustal alignment tool(44) and bacterial reference strains as templates for real-time PCR analysis. Theassays were performed with a 25-�l PCR amplification mixture containing 1�Thermo-Start QPCR master mix with ROX (ABgene, Rochester, NY), 50 to 300nM of forward and reverse primer, 100 to 200 nM of probe, and an equivalent of100 ng of bacterial DNA. The bacterial DNA concentration was determined byspectrophotometer (Beckman Coulter DU640, Fullerton, CA). The amplifica-tion conditions were 50°C for 2 min, 95°C for 10 min, and 40 cycles at 95°C for15 s and 60°C for 1 min. Fluorescent signals measured during amplification wereprocessed postamplification and were considered positive if the fluorescenceintensity was �20-fold of the standard deviation of the baseline fluorescence.This level was defined as the threshold cycle (CT) value and is inversely corre-lated to the amount of nucleic acid in the original sample (CT value of 40 � noamplification). CT values for each assay were compared among reference strainsto establish discriminatory properties between homologous Bifidobacteriumspecies.
Specific primers for Bifidobacterium animalis subsp. lactis (strain Bb12). Theamount of Bb12 in culture or in intestinal samples was determined using theprimer-probe set for the tuf gene (Table 3). A genomic tuf gene fraction wasamplified and used as an internal control and to generate a standard curve. Thesize of the fragment (117 bp) and its molecular mass were quantitatively deter-mined on the DNAChip (Agilent, Waldbronn, Germany) using Bioanalyzer(Agilent Technologies, Wilmington, DE). The molecular mass was used to cal-culate the copy number of the target amplicon used to generate a series of sixserial dilutions of known concentrations using a range that matched the expectedconcentration range of the unknown samples. Serial dilutions were analyzed intriplicate by real-time PCR in separate sample wells but within the same run, andthe resulting CT values were recorded. The unknown samples were evaluatedwithin the testing interval known to be linear. A plot of CT versus the logarithm
of the copy number corresponding to that CT resulted in a straight line standardcurve. The number of target gene copies was then extrapolated from the stan-dard curve equation. All dilutions of unknown samples were run in triplicate, andsamples were diluted in TE buffer using salmon sperm as a carrier (Invitrogen,CA) at a concentration of 60 �g/ml.
Sensitivity of the tuf gene assay for detection of Bb12. The sensitivity of the tufgene duplex 5� nuclease assay was assessed by testing for a series of dilutedsamples of exogenous Bb12 added to fecal samples of control pigs (those notexposed to probiotic treatment). One gram of lyophilized Bb12 was cultured onMRS agar and incubated anaerobically for 3 days at 37°C to verify purity and forquantitative determination of CFU per gram. Ten different 1-gram aliquots ofhomogenized feces of pigs not previously exposed to the probiotic were spikedwith either nine 10-fold dilutions of 1 gram of lyophilized Bb12 previouslydissolved in PBS or with 10-fold dilutions of a pure Bb12 broth culture. Aftertotal DNA extraction from spiked feces, serial 10-fold dilutions were analyzed intriplicate by real-time PCR in separate sample wells but within the same run, andthe resulting CT was compared to the standard curve analysis to estimate thebacterial copy number in each serial dilution. The efficiency of the PCR ampli-fication for the Bb12 tuf assay was calculated for different matrices (PBS andfeces) using the following formula as stated by Bustin and Nolan (4): efficiency �10(�1/slope)�1. Similarly, samples from intestinal contents of pigs that receivedBb12 orally for different periods of time were used to evaluate the detection levelof the Bb12 tuf gene assay.
Host mRNA gene expression by quantitative real-time PCR. The mRNA levelsof the TLR2, TLR4, and TLR9 genes were detected in the proximal colon ofpiglets derived from experiment 3. Tissue sections (2 cm2) were dissected fromthe proximal colon mucosa of pigs from the four treatment groups at days 7, 19,32, and 91 and were immediately frozen in liquid nitrogen and stored at �70°C.Tissue RNA was extracted after homogenization in Trizol reagent (Invitrogen,Gaithersburg, MD), and quantitative real-time PCR was performed on cDNAsynthesized from each sample using 10 �g of total RNA (42). RNA was treatedwith DNase in the presence of RNA inhibitor (Ambion, CA). The absence ofgenomic DNA contamination was confirmed after no signal was detected whenrunning a PCR using a non-exon-spanning probe for the RPL32 housekeepinggene. DNase-treated RNA was quantified using Bioanalyzer 2100 and the RNA6000 Labchip kit (Agilent Technologies, Palo Alto, CA) (9). Briefly, cDNA wassynthesized with Superscript RT (Invitrogen), and oligo(dT) and 50 ng of thiscDNA were used for real-time PCR amplifications using a Thermo-Start DNApolymerase master mix (ABgene, Rochester, NY) and the ABI Prism 7700sequence detector system (Applied Biosystems, Foster City, CA). Amplificationconditions were as follows: 50°C for 2 min; 95°C for 10 min; 40 cycles of 95°C for15 s; and 60°C for 1 min. All probes and primers selected for real-time PCR weredesigned using the Primer Express software package (Applied Biosystems, Fos-ter City, CA), and nucleotide sequences were obtained from GenBank or theTIGR porcine EST database. The TLR2, TLR4, and TLR9 genes were selectedto evaluate innate immune response activation by bacterial ligand. The sequenceinformation for genes assayed can be found in the Porcine Immunology andNutritional Database (http://www.ars.usda.gov/Services/docs.htm?docid�6065).
Fluorescence signals were processed after amplification and were consideredpositive if the fluorescence intensity was 20-fold more or greater than the stan-dard deviation of the baseline fluorescence. Gene expression was normalizedbased upon a constant amount of amplified RNA and cDNA (3, 8, 9). Relativequantification of target gene expression was evaluated by comparing linear re-gression lines constructed using CT values from cDNA processed at differenttimes for control pigs and pigs given different probiotic treatments after normal-ization with the RPL32 housekeeping gene. Changes in gene expression overtime are determined by comparing differences in the slope of the regression linesthat represented each treatment. A negative slope will indicate an upregulationof the gene since the CT value is inversely correlated to the amount of geneexpressed in the original sample.
Statistical analysis. Bb12 tuf copy numbers in the fecal samples or intestinalcontents collected from the proximal colon were determined by quantitativedetection of tuf gene copies at different times posttreatment. CT values generatedafter real-time PCR amplification with the Bb12 tuf gene assay were transformedto log10 base to represent bacterial copy numbers of Bb12 using the tuf gene asthe bacterial marker (mean � standard error [SE]). Contrast statements wererun to compare the effect of the treatment group within a time interval oftreatment. Statistical results are noted for mean values from each treatmentgroup and at each time interval evaluated, and significant differences werereported at a P value of 0.05. The analysis was performed using an SASsoftware package (Statview 5.0 for Macintosh Abacus Concepts, Berkeley, CA).Gene expression changes in TLR across time in proximal colon were done by atwo-way analysis of covariance. Linear regression lines representing each treat-
6340 SOLANO-AGUILAR ET AL. APPL. ENVIRON. MICROBIOL.
on March 13, 2014 by guest
http://aem.asm
.org/D
ownloaded from

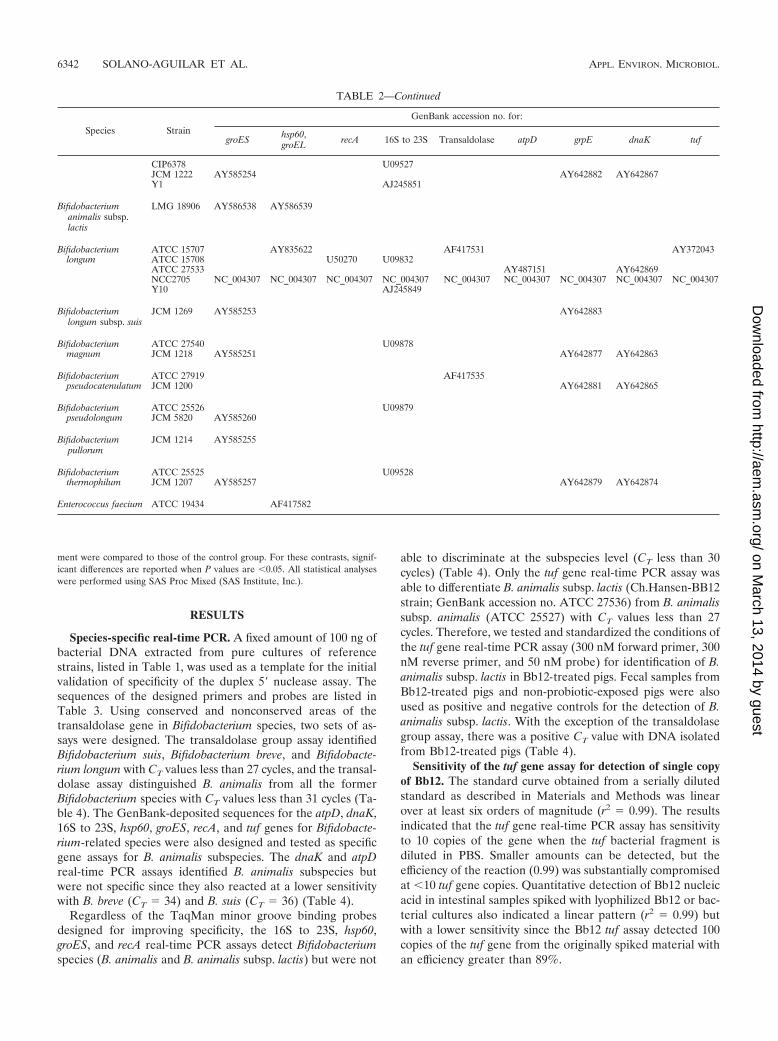
TABLE 2. GenBank sequence used to design real-time PCR assays
Species Strain
GenBank accession no. for:
groES hsp60,groEL recA 16S to 23S Transaldolase atpD grpE dnaK tuf
Bifidobacterium ATCC 15703 AY487144 AY372045adolescentis CIP6459 U09512
CIP6460 U09513CIP6461 U09514DSM 20083 AF417530JCM 1275 AY585248 AY642885 AY642864
Bifidobacterium ATCC 27535 AF417536angulatum JCM 7096 AY585256 AY642880 AY642870
Bifidobacterium LMG 11615 AY642876animalis ATCC 25527 AY585250 AY488178 AY370929 U09858 AY487152 AY642878 AY642871 AY370920
ATCC 27536 AY488181 AY370923 L36967 AY491983 AY370912ATCC 27672 AY488183 AY372028 AY491986 AY370921ATCC 27673 AY372029 AY370917ATCC 27674 AY488179 AY372030 AY491988 AY370918LMG 11615 AY642887 AY642876LMG 18900 AY642890 AY642872LMG 18906 AY642889 AY642873NCC 239 AY488182 AY370924 AY491984 AY370913NCC 311 AY370925 AY370914NCC 330 AY491985NCC 363 AY488176 AY370928 AY370915NCC 383 AY370926 AY370922NCC 402 AY488177 AY370927 AY491987 AY370916
Bifidobacteriumanimalis subsp.lactis
DSM 10140 AY488180 AY372031 X89513 AF417537 AY487153 AY667066 AY642912 AY370919
Bifidobacterium ATCC 15696 U50267bifidum ATCC 29521 AY487145 AY372041
CIP567 U09517CTP6465 U09831DSM 20456 AF417538DSM 20456 AF417533JCM 1255 AY585252 AY642888 AY642868
Bifidobacterium ATCC 15698 U09518 AF417532breve ATCC 15700 AY487154 AY372046
ATCC 15701 U50268CIP6468 U09519CIP6469 U09520CIP6470 U09521NCFB 2258 AF094756UCC2003 AY585262 AY585261Y8 AJ245850
Bifidobacterium ATCC 27539 U09522 AY487146catenulatum DSM 20103 AF417534 AY372044
JCM 1194 AY585249 AY642884 AY642875
Bifidobacteriumchoerinum
ATCC 25911 AY487148
Bifidobacterium ATCC 25911 U09523coryneforme ATCC 27686 AY487147
JCM 5819 AY585258
Bifidobacteriumcuniculi
ATCC 27916 U09790
Bifidobacterium ATCC 27534 U10434 AY487149dentium JCM 1195 AY585247 AY642886 AY642866
Bifidobacteriumglobosum
ATCC 25865 U09524
Bifidobacteriumindicum
ATCC 25912 U09791
Bifidobacterium ATCC 15697 U50269 U09792 AF417529 AY487150infantis ATCC 15702 AF417540
ATCC 25962 U09525
Continued on following page
VOL. 74, 2008 DETECTION OF Bb12 IN INTESTINES OF SOWS AND PIGLETS 6341
on March 13, 2014 by guest
http://aem.asm
.org/D
ownloaded from
ment were compared to those of the control group. For these contrasts, signif-icant differences are reported when P values are 0.05. All statistical analyseswere performed using SAS Proc Mixed (SAS Institute, Inc.).
RESULTS
Species-specific real-time PCR. A fixed amount of 100 ng ofbacterial DNA extracted from pure cultures of referencestrains, listed in Table 1, was used as a template for the initialvalidation of specificity of the duplex 5� nuclease assay. Thesequences of the designed primers and probes are listed inTable 3. Using conserved and nonconserved areas of thetransaldolase gene in Bifidobacterium species, two sets of as-says were designed. The transaldolase group assay identifiedBifidobacterium suis, Bifidobacterium breve, and Bifidobacte-rium longum with CT values less than 27 cycles, and the transal-dolase assay distinguished B. animalis from all the formerBifidobacterium species with CT values less than 31 cycles (Ta-ble 4). The GenBank-deposited sequences for the atpD, dnaK,16S to 23S, hsp60, groES, recA, and tuf genes for Bifidobacte-rium-related species were also designed and tested as specificgene assays for B. animalis subspecies. The dnaK and atpDreal-time PCR assays identified B. animalis subspecies butwere not specific since they also reacted at a lower sensitivitywith B. breve (CT � 34) and B. suis (CT � 36) (Table 4).
Regardless of the TaqMan minor groove binding probesdesigned for improving specificity, the 16S to 23S, hsp60,groES, and recA real-time PCR assays detect Bifidobacteriumspecies (B. animalis and B. animalis subsp. lactis) but were not
able to discriminate at the subspecies level (CT less than 30cycles) (Table 4). Only the tuf gene real-time PCR assay wasable to differentiate B. animalis subsp. lactis (Ch.Hansen-BB12strain; GenBank accession no. ATCC 27536) from B. animalissubsp. animalis (ATCC 25527) with CT values less than 27cycles. Therefore, we tested and standardized the conditions ofthe tuf gene real-time PCR assay (300 nM forward primer, 300nM reverse primer, and 50 nM probe) for identification of B.animalis subsp. lactis in Bb12-treated pigs. Fecal samples fromBb12-treated pigs and non-probiotic-exposed pigs were alsoused as positive and negative controls for the detection of B.animalis subsp. lactis. With the exception of the transaldolasegroup assay, there was a positive CT value with DNA isolatedfrom Bb12-treated pigs (Table 4).
Sensitivity of the tuf gene assay for detection of single copyof Bb12. The standard curve obtained from a serially dilutedstandard as described in Materials and Methods was linearover at least six orders of magnitude (r2 � 0.99). The resultsindicated that the tuf gene real-time PCR assay has sensitivityto 10 copies of the gene when the tuf bacterial fragment isdiluted in PBS. Smaller amounts can be detected, but theefficiency of the reaction (0.99) was substantially compromisedat 10 tuf gene copies. Quantitative detection of Bb12 nucleicacid in intestinal samples spiked with lyophilized Bb12 or bac-terial cultures also indicated a linear pattern (r2 � 0.99) butwith a lower sensitivity since the Bb12 tuf assay detected 100copies of the tuf gene from the originally spiked material withan efficiency greater than 89%.
TABLE 2—Continued
Species Strain
GenBank accession no. for:
groES hsp60,groEL recA 16S to 23S Transaldolase atpD grpE dnaK tuf
CIP6378 U09527JCM 1222 AY585254 AY642882 AY642867Y1 AJ245851
Bifidobacteriumanimalis subsp.lactis
LMG 18906 AY586538 AY586539
Bifidobacterium ATCC 15707 AY835622 AF417531 AY372043longum ATCC 15708 U50270 U09832
ATCC 27533 AY487151 AY642869NCC2705 NC_004307 NC_004307 NC_004307 NC_004307 NC_004307 NC_004307 NC_004307 NC_004307 NC_004307Y10 AJ245849
Bifidobacteriumlongum subsp. suis
JCM 1269 AY585253 AY642883
Bifidobacterium ATCC 27540 U09878magnum JCM 1218 AY585251 AY642877 AY642863
Bifidobacterium ATCC 27919 AF417535pseudocatenulatum JCM 1200 AY642881 AY642865
Bifidobacterium ATCC 25526 U09879pseudolongum JCM 5820 AY585260
Bifidobacteriumpullorum
JCM 1214 AY585255
Bifidobacterium ATCC 25525 U09528thermophilum JCM 1207 AY585257 AY642879 AY642874
Enterococcus faecium ATCC 19434 AF417582
6342 SOLANO-AGUILAR ET AL. APPL. ENVIRON. MICROBIOL.
on March 13, 2014 by guest
http://aem.asm
.org/D
ownloaded from

Quantitative detection of Bb12 in GITs of neonatal pigs. Forexperiment 1, proximal colon contents were aseptically col-lected from all piglets on day 32 after birth. Bb12 tuf genecopies in 100 ng of DNA extracted per gram of proximal coloncontents were detected at a significantly higher number (P 0.05) in piglets from the T/T (n � 18) and C/T (n � 21) groups(1.40 � 0.0.14 and 1.05 � 1.3, respectively) than the T/C (n �16) (0.36 � 0.08) and C/C (n � 19) (0.03 � 0.08) groups (Fig.1). Bb12 tuf gene copies were undetectable in the intestinalcontents from a separate group of control untreated pigletsmaintained in a different section of the farrowing barn (n � 9)(data not shown).
For experiment 2, fecal samples were collected from fivepiglets in each of the experimental groups (C/C, C/T, T/C, and
T/T) at days 10, 23, 32, and 45 after birth. All pigs were weanedon day 23, and no further Bb12 or placebo treatments weregiven to any of the pigs. Ten days after birth and after contin-uous daily treatment with either placebo or Bb12, pigs from theT/T and C/T groups had a significantly higher number of Bb12tuf gene copies per gram of feces (2.32 � 1.04 and 2.48 � 1.05,respectively) compared to pigs from the T/C (0.71 � 0.39) andC/C (0.036 � 0.080) groups. There was, however, a significantreduction of at least 1 log in Bb12 tuf gene copies in the T/T(0.9 � 0.6) and C/T (1.36 � 0.70) pigs at day 23 after birth evenwith continuous treatment with the probiotic (P 0.05), whileBb12-tuf gene copies in the C/C or T/C groups were lower thanthe linear detection limit of the assay (0.7 � 0.89 and 0.2 � 0.4,respectively). At day 32 after birth and 9 days after termination
TABLE 3. Primers and probes used in the duplex 5� nuclease assays
Target gene Primer/probe Sequence (5� 3 3�)Amplicon
length(bp)
Transaldolase gene Forward CGA CAA GAA GCT CGA GGA GAT 116Reverse CGG ATC CTC GGC GAA CTProbe CCT TGC CTT CGA GAC CCT TGG CCT
Transaldolase group gene Forward GCG TCC GCT GTG GGC 106Reverse CTT CTC CGG CAT GGT GTTProbe TCC ACC GGC ACC AAG AAC GC
atpD Forward GAT GTT ACC AAG GGC CAT GTG 83Reverse CGC TCC TTG ATC ACG ATC TTC TProbe CGA CGT TTC CGG CCA CAT TCT CABGB probe TTC CGG CCA CAT TC
dnaK Forward GCA GCT CTG GCC TAC GGT 189Reverse ATA ATG CGC TGG TCC CAA TCProbe CCC GAC GTA GCC TGC ACC TGGBGB probe AGG CTA CGT CGG GCG
groES Forward TTG GCC CAG GTC GTC GT 119Reverse AGG TAT TCC TCG CCC TTG AAG TProbe AGG GCG AGC GTG TTC CCA TGG ABGB probe CGT GTT CCC ATG GAC
16S to 23S Forward TTT GCC GAG TGC GAT GGT 104Reverse GTG GCG GCC AGG GAA CProbe CCT GGC TTG CTG GCG TGG AAG AG
groEL, hsp60 Forward CCA AGT GGG TAA GCA TGA ATT TC 109Reverse GGT ATC GGC CAG CTT ATC CAProbe CCT GAC GAG CTT CCT CAT CGT ATT CAA TGBGB probe CAA AGA TCA TTG AAT ACG ATG AG
recA Forward GAA GGC GAT ATG GGT GAC AG 133Reverse GCC GAT CTT CTC TCG CAA CTBGB probe CAC AGG CGA ACA CGA
tuf Forward GTG TCG AGC GCG GCA A 117Reverse CTC GCA CTC ATC CAT CTG CTTBGB probe ATC AAC ACG AAC GTC GAG A
B. breve groEL, hsp60 Forward ATG TTG ACG GCG AGG CTC 85Reverse AAC CCG GGT GCT TTG ACAProbe CCC TGA TTC TGA ACA ACA T
E. faecium groEL, hsp60 Forward GAA ACG ACG GTG TCA TCA 126Reverse TCC ATT TTG TCG TTA TCT GProbe CCA CGA TCA AAC TGC ATA C
VOL. 74, 2008 DETECTION OF Bb12 IN INTESTINES OF SOWS AND PIGLETS 6343
on March 13, 2014 by guest
http://aem.asm
.org/D
ownloaded from

of treatment with Bb12, the level of Bb12 tuf gene copies waslower than the linear detection limit of the assay for all treat-ment groups (Fig. 2). No Bb12 tuf gene signal was detected 45days after birth or 22 days after termination of Bb12 treatment.For experiment 3, piglets from each of the treatment groups(T/T, T/C, C/T, and C/C) were euthanized on days 7, 19, 32, or91 after continuous treatment with either Bb12 (1.76 � 1010
CFU/pig/day) or the placebo. Bb12 tuf gene copies per gram ofintestinal contents were detected in the highest numbers (amean log range from 2.01 to 2.4/gram) in piglets from the T/Tgroup compared to those from the C/C group throughout theentire period of examination (P 0.05) (Fig. 3). Higher Bb12
tuf gene copies (a mean log range from 1.50 to 1.79/gram) weredetected in piglets from the C/T group at days 19 and 91 thanin the control (C/C) group (P 0.05) (Fig. 3). In contrast,Bb12 tuf gene copy numbers were marginally above the level ofdetection in the proximal colon contents of piglets from theT/C group (a mean log range from 0.58 to 1.12/gram) or werenot present in the C/C group (a mean log range from 0.08 to0.9/gram). No difference in total bacterial load, measured bythe 16S to 23S gene against all eubacteria or Lactobacillus spp.,were detected in the proximal colon contents among the var-ious treatment groups throughout the 91 days of the study(data not shown). The total number of Bifidobacterium spp.,however, was significantly increased only in the T/T group (CT
TABLE 4. Average CT values obtained with 100 ng of extracted DNA
Source of DNA sample Strain
CT value for:
Transaldolasegroup gene
Transaldolasegene atpD dnaK 16S to 23S hsp60 groES recA tuf
B. animalis subsp. lactis Ch.Hansen-Bb12 40 27 24 24 29 26 28 26 27B. animalis subsp.
lactisaATCC 27536 40 26 23 24 29 25 27 24 26
B. animalis ATCC 25527 40 31 25 40 30 27 27 26 40B. suis ATCC 27531 27 40 36 40 40 40 40 40 40B. breve ATCC 15700 25 40 33 34 40 40 40 40 40B. longum ATCC 15707 27 40 40 40 40 40 40 40 40E. faecium Ch.Hansen-SF273 40 40 40 40 40 40 40 40 40L. paracasei Ch.Hansen-Lc-01 40 40 40 40 40 40 40 40 40L. bulgaricus Ch.Hansen-LBA40 40 40 40 40 40 40 40 40 40L. acidophilus ATCC 53544 40 40 40 30 40 40 40 40 40Treated pig 40 32 29 30 37 31 29 31 32Control pig 40 40 35 40 40 40 40 40 40
a Originally identified as B. animalis.
FIG. 1. Bifidobacterium animalis subsp. lactis (strain Bb12) ex-pressed as a tuf gene copy number detected in piglets. Intestinal con-tents were taken from the proximal colon of piglets from experiment 1at day 32 after birth and following treatment with either Bb12 (1.05 �1010 CFU/day) or an equivalent volume of placebo. Bacterial copynumbers were extrapolated from a standard curve for tuf gene copynumber run in parallel with DNA extracted from intestinal contents asdescribed in Materials and Methods. Data were transformed to log10base to represent bacterial copy numbers of Bb12 using the tuf gene asthe bacterial marker (mean � SE). Gray bars, control placebo-treatedsow and control placebo-treated piglet (n � 19) (C/C); diagonal bars,control placebo-treated sow and Bb12-treated piglet (n � 21) (C/T);horizontal bars, Bb12-treated sow and control placebo-treated piglet(n � 16) (T/C); solid bars, Bb12-treated sow and Bb12-treated piglet(n � 18) (T/T); and empty bars, control untreated sows and untreatedpiglet (n � 9). Statistically significant results (P 0.05) are notedabove mean values (asterisk) of tuf gene copy number in differenttreatment groups.
FIG. 2. Bifidobacterium animalis subsp. lactis (strain Bb12) in fecalsamples from pigs following termination of oral treatment with Bb12.Bb12 expressed as a quantitative tuf gene copy number was determinedin fecal samples collected at 10, 23, 32, and 45 days after birth. Pigletsin the C/T and T/T groups from experiment 2 received Bb12 (1.05 �1010 CFU/day) from birth until weaning at day 23 after birth. Bacterialcopy numbers were extrapolated from a standard curve for tuf genecopy number run in parallel with DNA extracted from fecal samples asdescribed in Materials and Methods. Gray bars, control placebo-treated sow and control placebo-treated piglet (n � 5) (C/C); diagonalbars, control placebo-treated sow and Bb12-treated piglet (n � 5)(C/T); horizontal bars, Bb12-treated sow and control placebo-treatedpiglet (n � 5) (T/C); and solid bars, Bb12-treated sow and Bb12-treated piglet (n � 5) (T/T). Statistically significant results (P 0.05)are noted above mean values (asterisk) of tuf gene copy number indifferent treatment groups.
6344 SOLANO-AGUILAR ET AL. APPL. ENVIRON. MICROBIOL.
on March 13, 2014 by guest
http://aem.asm
.org/D
ownloaded from

value � 24.9 � SE 1.3) compared to the C/C group (30.2 � SE1.74) at day 91 after continuous Bb12 treatment (P 0.05).
Local host immune response. Mucosal tissue sections fromthe proximal colon were collected from all pigs in experiment3. The levels of mRNA (CT value) for TLR2, TLR4, and TLR9were compared among treatment groups. Linear regressionlines were constructed with the CT values obtained at differenttimes (days 7, 19, 32, and 91), and slopes of lines were com-pared among treatment groups. A negative slope indicatedupregulation of the gene. Among the TLR molecules evalu-ated, only TLR9 was significantly upregulated over time (slope ��0.89; R2 � 0.55) in pigs from the T/T group (P 0.01)(Table 5).
DISCUSSION
To detect the presence of Bifidobacterium animalis subsp.lactis strain Bb12 in the intestinal contents of young pigs afteroral ingestion, a duplex 5� nuclease assay using the tuf gene wasdesigned, optimized, and validated. The tuf gene is uniqueamong the bacterial housekeeping genes tested because it isexpressed as a single copy in the bacterial genome and is usefulfor differentiating the closely related B. animalis subsp. anima-lis from B. animalis subsp. lactis strain Bb12 (Table 4). Withthis highly specific and sensitive assay, it was possible to detectmore than 100 bacterial copies of the tuf gene per 100 ng ofDNA extracted from 1 g of intestinal contents from the prox-imal colon and feces of pigs fed viable Bb12. The number ofBb12 organisms detected using the tuf gene assay was depen-dent on the dietary treatment regimen and the time of sam-pling. Piglets treated with Bb12 after birth (C/T group) (log1.05 � 1.3) and those from Bb12-treated sows that were alsotreated from birth (T/T group) (log 1.40 � 0.14) had thehighest number of tuf gene copies in the proximal colon con-
tents after 32 days of continuous daily feeding compared toplacebo-treated pigs (C/C group) (log 0.3 � 0.08) and placebo-treated pigs from Bb12-treated sows (T/C group) (log 0.04 �0.08) (Fig. 1). Similar differences were evident in fecal samplesanalyzed 10 days after birth (Fig. 2); Bb12 tuf gene copiesdetected in pigs from the C/T group (log 2.48 � 1.05) and T/Tgroup (log 2.32 � 1.04) were 1 and 2 bacterial logs higher thanthose detected at day 23 (C/T group � 1.36 � 0.7; T/T group �0.85 � 0.57). Fecal samples taken 11 (32 days after birth) and24 (45 days after birth) days after Bb12 feeding ceased werebelow the detection limit, suggesting that a continuous admin-istration of Bb12 is needed to maintain detectable levels ofBb12 in the GITs of growing pigs.
The experimental design used in these studies provided ascheme to evaluate the contribution of the sow to Bb12 num-bers found in the GITs of its offspring. On days 7, 19, 32, and91 after birth, piglets from the T/T group had significantlyhigher numbers of Bb12 in their proximal colons than piglets inthe control group (P 0.05) (Fig. 3). A significant differencein bacterial log numbers between piglets in the C/T group andthe C/C group was only observed at day 19 after birth (1.46 �SE 0.45) and 72 days after weaning (day 91 after birth) (1.79 �0.06) (P 0.05), with a slight difference among treatmentgroups C/T and T/T at 91 days after treatment (P 0.1). TheBb12 feeding regimens tested did not influence the total num-ber of Eubacteria and Lactobacillus species (data not shown) inthe proximal colon over 91 days of feeding with or withoutfeeding Bb12 to the sow (data not shown); however, the totalnumber of Bifidobacterium species in the T/T group at day 91was increased. This suggests that daily feeding of greater than1010 CFU/pig/day of Bb12 did not significantly alter otherrepresentative bacterial populations in the intestine, but Bb12through the mother can influence the bacterial load of Bi-fidobacterium species in the offspring later in life. Similar ob-servations of an influence of maternal Bifidobacterium countson infants’ fecal Bifidobacterium levels have been recently de-scribed in humans (11).
The impact of Bb12 treatment on localized host tissue re-sponses was evaluated in the proximal colon mucosa. TheTLRs are type I transmembrane proteins expressed by manycell types, including intestinal epithelial cells and classical im-mune cells, that function as recognition receptors for the in-nate immune system through binding of ligands associatedlargely with microbial, viral, and certain protozoan and meta-
TABLE 5. Gene expression analysis in proximal colon mucosa ofpigs treated with Bb12
TLRSlope for treatment groupa:
C/C C/T T/C T/T
2 �0.03 0.12 0.09 �0.284 �0.03 0.04 �0.006 �0.169 0.26 0.12 �0.04 �0.89b
a Slopes of linear regression lines constructed with CT values obtained formRNA expression at different times (days 7, 19, 32, and 91). Changes in geneexpression over time are determined by comparing differences in the slope of theregression lines that represented each treatment. A negative slope will indicatean upregulation of the gene since the CT value is inversely correlated to theamount of gene expressed in the original sample.
b Statistical significance is noted when P was 0.05.
FIG. 3. Bifidobacterium animalis subsp. lactis (strain Bb12) in pigsafter continuous treatment with Bb12. Bb12 tuf gene copy number wasdetermined in proximal colon contents of pigs of experiment 3 at 7, 19,32, and 91 days after birth and continuous treatment with either 1.76 �1010 CFU/pig/day of Bb12 or an equivalent amount of placebo. Bac-terial copy numbers were extrapolated from a standard curve for tufgene copy number run in parallel with DNA extracted from proximalcolon contents as described in Materials and Methods. Gray bars,control placebo-treated sow and control placebo-treated piglet (n �13) (C/C); diagonal bars, control placebo-treated sow and Bb12-treated piglet (n � 13) (C/T); horizontal bars, Bb12-treated sow andcontrol placebo-treated piglet (n � 17) (T/C); and solid bars, Bb12-treated sow and Bb12-treated piglet (n � 14) (T/T). Statistically sig-nificant (P 0.05) results are noted above mean values (asterisk) of tufgene copy number in different treatment groups.
VOL. 74, 2008 DETECTION OF Bb12 IN INTESTINES OF SOWS AND PIGLETS 6345
on March 13, 2014 by guest
http://aem.asm
.org/D
ownloaded from

zoan pathogens (2, 51). Gene expression analysis on proximalcolon mucosa indicated a significant upregulation of TLR9only in pigs from the T/T group that maintained the highestnumber of Bb12 tuf gene copies throughout the experiment(T/T group) (P 0.01) compared to pigs from all other groups(C/T, T/C, and C/C group) (Table 5; Fig. 3) No significantchanges in mRNA expression were seen with either TLR2 orTLR4, suggesting a selective induction of TLR9 by the higherlevels of Bb12. TLR9 plays a key role in the detection ofbacterial DNA to induce innate immunity that is linked to theactivation of various cell types for development of an acquiredimmune response (18, 20–22). Tohno et al. (45) reported theexpression of TLR2 and TLR9 in the mesenteric lymph nodesand ileal Peyer’s patches of suckling piglets and that isolatedlymphocytes were induced to proliferate and release cytokinesafter in vitro stimulation with CpG 2006, a TLR9 ligand, zy-mosan, and heat-killed lactic acid-forming bacteria. Our resultssupport these findings and add the dimension of in vivo acti-vation of TLR9 by feeding live Bb12.
Data generated from these experiments suggest that detec-tion of Bb12 using a single-copy Bb12 tuf gene real-time PCRassay is a useful tool to study the relationship between local-ized accumulation of Bb12 in the GIT and modulation of thehost innate immune response. Oral feeding of live Bb12 tosows during gestation and to their piglets at the day of birthand for 91 days after birth established higher numbers of Bb12in the intestinal contents that induced significant upregulationof mRNA expression for TLR9. These changes were not ob-served in pigs that did not receive Bb12 or in those whosemothers were treated only with Bb12 during the last trimesterof pregnancy (T/C group). Piglets from untreated sows thatreceived daily oral treatment with Bb12 from the day of birththrough 91 days after birth also failed to express significantchanges in TLR9 (C/T group). This suggests that exposure ofthe mother to Bb12 influenced both Bb12 load in the piglet inthe presence of continuous daily feeding and significantly af-fected the host innate immune system development in theambient environment. It is not clear if these differences arebased on an early inoculation of the piglet as it transversesthe birth canal and perianal area of the Bb12-treated sow or ifthe sow provides Bb12-supportive lactogenic colostrum andmilk during nursing, as has recently been suggested for humansfor other Bifidobacterium species (11). One could also considerepigenetic factors that could program the piglet for greaterresponsiveness to Bb12 during development. The pattern ofupregulation of TLR9 may suggest that Bb12 provides a “prim-ing signal” when piglets are inoculated by the sow and through-out early development. Future studies will evaluate the level ofinnate and acquired immunity and the degree of inflammationwhen these pigs are challenged with a proinflammatory stim-ulus.
ACKNOWLEDGMENTS
This work was supported by funds from USDA CRIS no. 1235-52000-054 and a trust agreement with Nestle.
Probiotic bacteria were kindly provided by Chr. Hansen (UnitedStates and Denmark).
REFERENCES
1. Banerjee, A., and S. Gerondakis. 2007. Coordinating TLR-activated signal-ing pathways in cells of the immune system. Immunol. Cell Biol. 85:420–424.
2. Bochud, P., M. Bochud, A. Telent, and T. Calandra. 2007. Innate immuno-genetics: a tool for exploring new frontiers of host defence. Lancet Infect.Dis. 7:531–542.
3. Bustin, S. A. 2002. Quantification of mRNA using real-time reverse tran-scription PCR (RT-PCR): trends and problems. J. Mol. Endocrinol. 29:23–39.
4. Bustin, S. A., and T. Nolan. 2004. Data analysis and interpretation, p.439–491. In Stephen A. Bustin (ed.), A-Z of quantitative PCR. InternationalUniversity Line Publishers, La Jolla, CA.
5. Candela, M., B. Vitali, D. Matteuzzi, and P. Brigidi. 2004. Evaluation of therrn operon copy in Bifidobacterium using real-time PCR. Lett. Appl. Micro-biol. 38:229–232.
6. Carey, C. M., J. L. Kirk, S. Ojha, and M. Kostrzynska. 2007. Current andfuture uses of real-time polymerase chain reaction and microarrays in thestudy of intestinal microbiota, and probiotic use and effectiveness. Can. J.Microbiol. 53:537–550.
7. Chavagnat, F., M. Haueter, J. Jimeno, and M. G. Casey. 2002. Comparisonof partial tuf gene sequences for the identification of lactobacilli. FEMSMicrobiol. Lett. 217:177–183.
8. Dawson, H. D., A. R. Royaee, S. Nishi, D. Kuhar, W. M. Schnitzlein, F.Zuckermann, J. Urban, Jr., and J. K. Lunney. 2004. Identification of keyimmune mediators regulating T helper 1 responses in swine. Vet. Immunol.Immunopathol. 100:105–111.
9. Dawson, H. D., E. Beshah, S. Nishi, G. Solano-Aguilar, M. Morimoto, A.Zhao, K. B. Madden, T. K. Ledbetter, J. P. Dubey, T. Shea-Donohue, J. K.Lunney, and J. Urban, Jr. 2005. Localized multigene expression patternssupport an evolving Th1/Th2-like paradigm in response to infections withToxoplasma gondii and Ascaris suum. Infect. Immun. 73:1116–1128.
10. Delroisse, J. M., A. L. Boulvin, I. Parmentier, R. D. Dauphin, M. Vandenbol,and D. Portetelle. 2006. Quantification of Bifidobacterium spp. and Lacto-bacillus spp. in rat fecal samples by real-time PCR. Microbiol. Res. doi:10.1016/j.micres.2006.09.004.
11. Gronlund, M.-M., M. Gueimonde, K. Laitinen, G. Kociubinski, T. Gronroos,S. Salminen, and E. Isolauri. 2007. Maternal breast-milk and intestinalbifidobacteria guide the compositional development of the Bifidobacteriummicrobiota in infants at risk of allergic disease. Clin. Exp. Allergy 37:1764–1772.
12. Haarman, M., and J. Knol. 2006. Quantitative real-time PCR analysis offecal Lactobacillus species in infants receiving a probiotic infant formula.Appl. Environ. Microbiol. 72:2359–2365.
13. Janer, C., F. Arigoni, B. H. Lee, C. Pelaez, and T. Requena. 2005. Enzymaticability of Bifidobacterium animalis subsp. lactis to hydrolyze milk proteins:identification and characterization of endopeptidase O. Appl. Environ. Mi-crobiol. 71:8460–8465.
14. Jian, W., L. Zhu, and X. Doug. 2001. New approach to phylogenetic analysisof the genus Bifidobacterium based on partial HSP60 gene sequences. Int. J.Syst. Evol. Microbiol. 51:1633–1638.
15. Karimi, O., and A. S. Pena. 2003. Probiotics: isolated bacteria strain ormixtures of different strains? Two different approaches in the use of probi-otics as therapeutics. Drugs Today 39:565–597.
16. Klijn, A., A. Mercenier, and F. Arigoni. 2005. Lessons from the genomes ofbifidobacteria. FEMS Microbiol. Rev. 29:491–509.
17. Kullen, M. J., L. J. Brady, and D. J. O’Sullivan. 1997. Evaluation of using ashort region of the recA gene for rapid and sensitive speciation of dominantbifidobacteria in the human large intestine. FEMS Microbiol. Lett. 154:377–383.
18. Kumagai, Y., O. Takeuchi, and S. Akira. 2008. TLR9 as a key receptor forthe recognition of DNA. Adv. Drug Deliv. Rev. 60:795–804.
19. Kwon, H. S., E. H. Yang, S. H. Lee, S. W. Yeon, B. H. Kang, and T. Y. Kim.2005. Rapid identification of potentially probiotic Bifidobacterium species bymultiplex PCR using species specific primers based on the region extendingfrom 16S rRNA through 23S rRNA. FEMS Microbiol. Lett. 250:55–62.
20. Lee, J., D. Rachmilewitz, and E. Raz. 2006. Homeostatic effects of TLR9signaling in experimental colitis. Ann. N. Y. Acad. Sci. 1072:351–355.
21. Lee, J., J. Mo, K. Katakura, I. Alkalay, A. N. Rucker, Y. Liu, H. Lee, C. Shen,G. Cojocaru, S. Shenouda, M. Kagnoff, L. Eckmann, Y. Ben-Neriah, and E.Raz. 2006. Maintenance of colonic homeostasis by distinctive apical TLR9signalling in intestinal epithelial cells. Nat. Cell Biol. 8:1327–1336.
22. Lee, J., J. Mo, C. Shen, A. N. Rucker, and E. Raz. 2007. Toll-like receptorsignaling in intestinal epithelial cells contributes to colonic homeostasis.Curr. Opin. Gastroenterol. 23:27–31.
23. Masco, L., M. Ventura, R. Zink, G. Huys, and J. Swings. 2004. Polyphasictaxonomic analysis of Bifidobacterium animalis and Bifidobacterium lactisreveals relatedness at the subspecies level: reclassification of Bifidobacteriumanimalis as Bifidobacterium animalis subsp. animalis subsp. nov. and Bi-fidobacterium lactis as Bifidobacterium animalis subsp. lactis subsp. nov. Int.J. Syst. Evol. Microbiol. 54:1137–1143.
24. Masco, L., G. Huys, E. De Brandt, R. Temmerman, and J. Swings. 2005.Culture dependent and culture independent qualitative analysis of probioticproducts claimed to contain bifidobacteria. Int. J. Food Microbiol. 102:221–230.
25. Matsuki, T., K. Watanabe, and R. Tanaka. 2003. Genus- and species-specific
6346 SOLANO-AGUILAR ET AL. APPL. ENVIRON. MICROBIOL.
on March 13, 2014 by guest
http://aem.asm
.org/D
ownloaded from

PCR primers for the detection and identification of bifidobacteria. Curr.Issues Intest. Microbiol. 4:61–69.
26. McCartney, A. L. 2002. Application of molecular biological methods forstudying probiotics and the gut flora. Br. J. Nutr. 88:S29–S37.
27. Meile, L., W. Ludwig, U. Rueger, C. Gut, P. Kaufmann, G. Dasen, S. Wenger,and M. Teuber. 1997. Bifidobacterium lactis. sp. nov., a moderately oxygentolerant species isolated from fermented milk. Syst. Appl. Microbiol. 20:57–64.
28. Miyake, T., K. Wanatabe, T. Wanatabe, and H. Oyaizu. 1998. Phylogeneticanalysis of the genus Bifidobacterium and related genera based on 16S rDNAsequences. Microbiol. Immunol. 42:661–667.
29. Nadkarni, M. A., E. F. Martin, N. A. Jacques, and N. Hunter. 2002. Deter-mination of bacterial load by real-time PCR using a broad-range (universal)probe and primers set. Microbiology 148:257–266.
30. Ouwehand, A. C., S. Salminen, and E. Isolauri. 2002. Probiotics: an overviewof beneficial effects. Antonie van Leeuwenhoek 82:279–289.
31. Paradis, S., M. Boissinot, N. Paquette, S. D. Belanger, E. Martel, D. K.Boudreau, F. J. Picard, M. Ouellette, P. H. Roy, and M. G. Bergeron. 2005.Phylogeny of the Enterobacteriaceae based on genes encoding elongationfactor Tu and F-ATPase -subunit. Int. J. Syst. Evol. Microbiol. 55:2013–2025.
32. Parracho, H., A. L. McCartney, and G. Gibson. 2007. Probiotics and prebi-otics in infant nutrition. Proc. Nutr. Soc. 66:405–411.
33. Picard, F. J., K. Danbing, D. K. Boudreau, M. Boissinot, A. Huletsky, D.Richard, M. Ouellette, P. H. Roy, and M. G. Bergeron. 2004. Use of the tufsequences for genus-specific PCR detection and phylogenetic analysis of 28streptococcal species. J. Clin. Microbiol. 42:3686–3695.
34. Prasad, J., H. Gill, J. Smart, and P. K. Gopal. 1998. Selection and charac-terisation of Lactobacillus and Bifidobacterium strains for use as probiotics.Int. Dairy J. 8:993–1002.
35. Requena, T., J. Burton, T. Matsuki, K. Munro, M. A. Simon, R. Tanaka, K.Watanabe, and G. Tannock. 2002. Identification, detection, and enumerationof human Bifidobacterium species by PCR targeting the transaldolase gene.Appl. Environ. Microbiol. 68:2420–2427.
36. Saavedra, J. M. 2007. Use of probiotics in pediatrics: rationale, mechanismsof action, and practical aspects. Nutr. Clin. Pract. 22:351–365.
37. Saavedra, J. M., A. Abi-Hanna, N. Moore, and R. H. Yolken. 2004. Long-term consumption of infant formulas containing live probiotic bacteria: tol-erance and safety. Am. J. Clin. Nutr. 79:261–267.
38. Sakata, S., C. S. Ryu, M. Kitahara, M. Sakamoto, M. Hayashi, M.Fukuyama, and Y. Benno. 2006. Characterization of the genus Bifidobacte-rium by automated ribotyping and 16S rRNA gene sequences. Microbiol.Immunol. 50:1–10.
39. Satokari, R. M., E. E. Vaughan, H. Smidt, M. Saarela, J. Matto, and V. M.de Vos. 2003. Molecular approaches for the detection and identification ofbifidobacteria and lactobacilli in the human gastrointestinal tract. Syst. Appl.Microbiol. 26:572–584.
40. Saxelin, M., S. Tynkkynen, T. Mattila-Sandholm, and W. M. de Vos. 2005.Probiotic and other functional microbes: from markets to mechanisms. Curr.Opin. Biotechnol. 16:204–211.
41. Scardovi, V., and L. D. Trovatelli. 1974. Bifidobacterium animalis (Mitsuoka)comb. nov. and the “minimum” and “subtile” groups of new bifidobacteriafound in sewage. Int. J. Syst. Bacteriol. 24:21–28.
42. Solano-Aguilar, G. I., E. Beshah, K. Vengroski, L. Gasbarre, D. Junker, M.Cochran, C. Weston, D. Valencia, C. Chiang, H. D. Dawson, J. F. Urban, andJ. K. Lunney. 2002. Limited effect of recombinant porcine interleukin-12 onporcine lymphocytes due to a low level of IL-12 beta2 receptor. Vet. Immu-nol. Immunopathol. 89:133–148.
43. Suau, A., R. Bonnet, M. Sutren, J. J. Godon, G. R. Gibson, M. D. Collins,and J. Dore. 1999. Direct analysis of the genes encoding 16S rRNA fromcomplex communities reveals many novel molecular species within the hu-man gut. Appl. Environ. Microbiol. 65:4799–4807.
44. Thompson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin, and D. G.Higgins. 1997. The Clustal_X windows interface: flexible strategies for mul-tiple sequence alignment aided by quality analysis tools. Nucleic Acids Res.25:4876–4882.
45. Tohno, M., T. Shimosato, M. Moue, H. Aso, K. Watanabe, Y. Kawai, T.Yamaguchi, T. Saito, and H. Kitazawa. 2006. Toll-like receptor 2 and 9 areexpressed and functional in gut associated lymphoid tissues of presucklingnewborn swine. Vet. Res. 37:791–812.
46. Ventura, M., and R. Zink. 2002. Rapid identification, differentiation andproposed new taxonomic classification of Bifidobacterium lactis. Appl. Envi-ron. Microbiol. 68:6429–6434.
47. Ventura, M., and R. Zink. 2003. Comparative sequence analysis of the tufand recA genes and restriction fragment length polymorphism of the internaltranscribed spacer region sequences supply additional tools for discriminat-ing Bifidobacterium lactis from Bifidobacterium animalis. Appl. Environ. Mi-crobiol. 69:7517–7522.
48. Ventura, M., C. Canchaya, V. Meylan, T. R. Klaenhammer, and R. Zink.2003. Analysis, characterization, and loci of the tuf genes in Lactobacillus andBifidobacterium species and their direct application for species identification.Appl. Environ. Microbiol. 69:6908–6922.
49. Ventura, M., C. Canchaya, R. Zink, G. F. Fitzgerald, and D. van Sinceren.2004. Characterization of the groEL and groES loci in Bifidobacterium breveUCC 2003: genetic, transcriptional, and phylogenetic analyses. Appl. Envi-ron. Microbiol. 70:6197–6209.
50. Ventura, M., R. Zink, G. F. Fitzgerald, and D. van Sinderen. 2005. Genestructure and transcriptional organization of the dnaK operon of Bifidobac-terium breve UCC 2003 and application of the operon in bifidobacterialtracing. Appl. Environ. Microbiol. 71:487–500.
51. Watson, J. L., and D. M. McKay. 2006. The immunophysiological impact ofbacterial CpG DNA on the gut. Clin. Chim. Acta 364:1–11.
52. Yin, X., J. R. Chambers, K. Barlow, A. S. Park, and R. Wheatcroft. 2005. Thegene encoding xylulose-5-phosphate/fructose-6-phosphate phosphoketolase(xfp) is conserved among Bifidobacterium species within a more variableregion of the genome and both are useful for strain identification. FEMSMicrobiol. Lett. 246:251–257.
53. Youn, S. Y., J. M. Seo, and G. E. Ji. 2008. Evaluation of the PCR method foridentification of Bifidobacterium species. Lett. Appl. Microbiol. 46:7–13.
54. Zhu, L., and X. Dong. 2003. Species identification of genus Bifidobacteriumbased on partial HSP60 gene sequences and proposal of Bifidobacteriumthermacidophilum subsp. porcinum subsp. nov. Int. J. Syst. Evol. Microbiol.53:1619–1623.
55. Zoetendal, E. G., C. T. Collier, S. Koike, R. I. Mackie, and H. R. Gaskins.2004. Molecular ecological analysis of the gastrointestinal microbiota: areview. J. Nutr. 134:465–472.
VOL. 74, 2008 DETECTION OF Bb12 IN INTESTINES OF SOWS AND PIGLETS 6347
on March 13, 2014 by guest
http://aem.asm
.org/D
ownloaded from




















