
Effects of late introduction of sows to two farrowing environments on the progress of farrowing and...
33
L. J. Pedersen and T. Jensen farrowing and maternal behavior Effects of late introduction of sows to two farrowing environments on the progress of published online May 9, 2008 J ANIM SCI http://jas.fass.org/content/early/2008/05/09/jas.2007-0749.citation the World Wide Web at: The online version of this article, along with updated information and services, is located on www.asas.org by guest on October 13, 2011 jas.fass.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Effects of late introduction of sows to two farrowing environments on the progress of farrowing and...

L. J. Pedersen and T. Jensenfarrowing and maternal behavior
Effects of late introduction of sows to two farrowing environments on the progress of
published online May 9, 2008J ANIM SCI
http://jas.fass.org/content/early/2008/05/09/jas.2007-0749.citationthe World Wide Web at:
The online version of this article, along with updated information and services, is located on
www.asas.org
by guest on October 13, 2011jas.fass.orgDownloaded from

1
1
Running head: Introduction of sows to farrowing pens 1
2
Effects of late introduction of sows to two farrowing environments on the progress of farrowing 3
and maternal behavior. 4
5
L. J. Pedersen1 and T. Jensen. 6
7
Institute of Animal Health, Welfare and Nutrition., Faculty of Agricultural Sciences, University of 8
Aarhus, 8830 Tjele, Denmark. 9
10
11
1Corresponding author: [email protected] 12
13
Page 1 of 31 Journal of Animal Science
Published Online First on May 9, 2008 as doi:10.2527/jas.2007-0749 by guest on October 13, 2011jas.fass.orgDownloaded from

2
2
14
ABSTRACT: To evaluate the effect of late introduction to farrowing pens on the progress of farrowing 15
and maternal behavior, 20 primiparous and 20 multiparous sows were allocated randomly to 1 of 2 16
treatments 1) early introduction to pen (EP, n = 20) and 2) late introduction to pen (LP, n = 20). To 17
evaluate the difference between loose housed sows and crated sows when introduced late to the 18
farrowing environment a third treatment was included 3) late introduction to farrowing crate (LC, n =19
20). Sow behavior and piglet birth intervals were recorded using video recordings from 16 h before the 20
birth of the first piglet (BFP) until 48 h after BFP. Behavioral data were analyzed using Proc Mixed in 21
SAS whereas the percentage of stillborn piglets as well as the response of the sow to piglet scream was 22
analyzed using Proc Genmod in SAS. Before farrowing (16 h to 3 h before BFP) sows introduced late 23
to pens had more postural changes per hour than sows introduced early to pens (LP = 12.7, EP = 8.9; P24
= 0.04), whereas there were no differences between sows introduced late to crates and sows introduced 25
late to pens (LC = 14.2, LP = 12.7; P = 0.53). Inter-birth interval (P = 0.04), variation in the inter-birth 26
interval (P = 0.01), and percentage of stillborn piglets (P = 0.003) were affected by an interaction 27
between parity and treatment. In multiparous sows there were no differences between treatments (P >28
0.18) either in the progress of farrowing or in the percentage of stillborn piglets. For primiparous sows, 29
there were no differences (P > 0.22) between sows that were introduced late to pens and sows that were 30
introduced early to pens. Primiparous sows that were introduced late to crates compared to pens had 31
longer inter-birth intervals (LC = 29 ± 4.9 min, LP = 16 ± 2.9 min; P = 0.02), a greater variation of 32
these intervals (LC = 35 ± 8.3 min, LP = 16 ± 3.6 min; P = 0.006), and a greater percentage of stillborn 33
piglets (LC = 21 %; 95 % confidence interval ranging 14 to 30 %, LP = 5 %; 95 % confidence interval 34
ranging 2 to 12 %; P = 0.004). After farrowing, neither postural changes, time spent in lateral lying, 35
Page 2 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

3
3
number of near crushing situations, nor the response to piglet scream test were affected by treatment (P36
> 0.09). When sows and gilts were introduced late to farrowing pens, neither progress of farrowing nor 37
maternal behavior of importance for piglet crushing were influenced. However, crating primiparous 38
sows that were introduced late to the farrowing environment compared to pen housing had detrimental 39
effects on the progress of farrowing and the percentage of still born piglets. 40
41
Keywords: environment, farrowing pens, maternal behavior, sows, stillborn piglets 42
43
INTRODUCTION 44
In intensive pig production sows are often moved to the farrowing unit close to the time of farrowing. 45
This is due to a combination of an increased weaning age and increased litter size leading to more use 46
of foster sows that nurse 2 litters in succession. Thus some farrowing crates are occupied for a longer 47
period and consequently there will not be room for the next batch of sows. To move sows immediately 48
before farrowing may be stressful, with potential negative effects on the progress of farrowing and 49
maternal behavior (Lawrence et al., 1997). Because maternal behavior and neonatal piglet viability is 50
considered of great importance for early piglet survival when sows are kept loose, the production 51
success may be particularly sensitive to such management procedures in loose-housed sows. When kept 52
under natural conditions, sows will seek isolation and initiate nest-building 2 to 3 d before farrowing 53
(Jensen, 1986). Thus sows introduced late to the farrowing housing begin seeking isolation while still 54
in the gestation unit. The lack of an outlet for this strong motivation may also constitute a stressor, 55
interfering with aspects of maternal behavior of importance for early piglet survival such as nesting 56
behavior and carefulness when lying down (Andersen et al., 2005; Pedersen et al., 2006). Present 57
knowledge indicates that moving sows shortly before farrowing may interfere with the progress of 58
Page 3 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

4
4
farrowing and other aspects of maternal behavior thereby reducing piglet survival. Such a 59
relationship may particularly be important when sows are housed loose in pens where the impact of 60
piglet viability and maternal behavior on piglet survival seems greater. The aim of the present study 61
was 1) to investigate the effect of late introduction to farrowing pens on the progress of farrowing and 62
maternal behavior and 2) to evaluate differences between penned and crated sows in maternal 63
behaviors. 64
65
MATERIALS AND METHODS 66
Animals, Housing and Treatment 67
All procedures involving animals were approved by the Danish Animal Experiments Inspectorate in 68
accordance with the Danish Ministry of Justice Law no. 382 (June 10, 1987) and Acts 333 (May 19, 69
1990), 726 (September 9, 1993) and 1016 (December 12, 2001). 70
71
Sixty sows (Landrace x Yorkshire) were used in a randomized complete block design with 2 blocks. In 72
block 1 the sows were primiparous and in block 2 the sows were multiparous (2.2 ± 1.1 litters, mean ±73
SD). Within blocks sows were randomly distributed to 3 times of introduction to the farrowing unit. To 74
evaluate the effect of late introduction to farrowing pens on the progress of farrowing and maternal 75
behavior, 20 primiparous and 20 multiparous sows were allocated randomly to 1 of 2 treatments 1) 76
early introduction to pen (EP; n = 20) and 2) late introduction to pen (LP; n = 20). To evaluate the 77
difference between loose-housed sows and crated sows when introduced late to the farrowing 78
environment a third treatment was included 3) late introduction to farrowing crate (LC; n = 20). In the 79
late introduction to crates and pens, sows were moved from the gestation pen to the farrowing 80
Page 4 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

5
5
environment on d 114 of pregnancy while in the early introduction to pens the sows were moved from 81
the gestation pen to the farrowing pen on d 95 of pregnancy, except the first 6 sows that were moved on 82
d 105 of pregnancy. The reason for changing the day of introduction from d 105 to d 95 was that it 83
allowed for additional sampling of the time taken to adjust to the new environment (data not reported 84
here). Initial analysis showed no differences in any response variables between the 2 d of introduction. 85
Thus the 2 treatments were treated as 1 throughout the paper. 86
87
The experiment was conducted run from February to October 2005 at the experimental herd of 88
Research Center Foulum. Sows were walked a distance of 100 m from the gestation unit to the 89
farrowing unit in batches of 6 sows approximately weekly. They were moved between 1000 and 1100. 90
Each farrowing unit comprised between 20 and 28 farrowing pens or crates in an environmentally 91
controlled building. The room temperature was kept constant around 20°C throughout the experiment. 92
Farrowing pens for loose-housed sows (Figure 1) measured 2.7 m x 3.2 m and were divided into a 93
resting area (2.7 m x 1.5 m) with concrete floor and an activity area with slatted floor (2.7 m x 1.7 m). 94
A moulded piglet box of plastic with an integral plastic flooring measuring 1 m in length x 0.5 m in 95
width x 0.45 m in height was situated in the center of the pen. The box had a single opening of 31 cm x 96
33 cm facing towards the resting area of the pen and an infrared heating lamp was connected to each 97
box. The day before expected farrowing the piglet box was filled with a 10 cm layer of chopped straw 98
and the heating lamp was turned on. At the back wall of the resting area a 2.5 m long farrowing rail of 99
galvanized iron (4 cm in diameter) was positioned 15 cm from the wall and 22 cm above the floor. The 100
back wall and the sidewalls were made of solid plastic. The front wall that was facing the alley from 101
where the sows were fed and inspected consisted of iron tubes. The feed trough, a water nipple with an 102
Page 5 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

6
6
open water bowl below, and a straw rack were positioned in the front of the pen. Pens for crated sows 103
(Figure 1) consisted of a total area measuring 2.35 m in length and 2 m in width. Two thirds of the 104
floor was concrete whereas the remaining third at the hind part of the sow had slatted metal floor. 105
Within a pen the sow was crated by spindle tubing. The crates measured 2 m in length and were 106
adjustable in width from 0.65 cm to 0.80 cm. 107
108
During gestation (from 4 wk after mating) all sows were housed in dynamic groups of 30 to 35 sows 109
with a space allowance of approximately 3 m2 per sow and with access to 1 electronic sow feeding 110
station per pen. Primiparous sows and multiparous sows were kept separate in different groups. Pens 111
were divided into a resting area with concrete floor and an activity area with slatted floor also made of 112
concrete. The feeding station was placed in the slatted floor area. Sows got a daily ration of 2.6 kg feed 113
containing 7.62 MJ net energy/kg. The feeding cycle started at 0500 and access to the feeding station 114
was closed during nighttime from 0900 to 0500. Approximately 200 g chopped straw per sow was 115
delivered daily by hand in the solid concrete area. 116
117
In the farrowing units sows were fed twice daily with a standard sow meal containing 8.40 MJ net 118
energy/kg. Before farrowing sows were fed a ration of 2.6 kg daily and after farrowing the sows feed 119
ration was gradually increased over the next 2 wk until sows were fed ad libitum. In the farrowing pens 120
long straw was available ad libitum from a straw rack. Crated sows got approximately 500 g long straw 121
twice daily delivered in front of their feed trough. If soiled, the pen was cleaned once daily, but fresh 122
straw was left untouched. During the previous farrowing and lactation all multiparous sows had been 123
housed in the same type of pen or crate as the experimental housing. Thus older multiparous sows may 124
have had experience with both types of farrowing housing. 125
Page 6 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

7
7
126
Behavioral Observations and Tests 127
A digital video camera (TVCCD-14IR, Monacor, Bremen, Germany) connected to a standard PC was 128
placed above each pen/crate. Recordings were carried out from d 114 of pregnancy until 48 h after the 129
birth of first piglet (BFP) using a digital surveillance system (MSH-Video, M.Shafro & Co., Riga, 130
Latvia). The time of birth of each individual piglet was recorded and the mean birth interval as well as 131
the variation (calculated as SD) in the birth intervals within each litter was calculated. Nest building 132
behavior was observed from 16 h before BFP until 8 h after. The duration of nest building within each 133
hour was recorded (for the applied definition of nest building see Table 1). The number of postural 134
changes per hour, defined as the number of shifts between the 4 postures (lateral lying, sternal lying, 135
sitting, and standing and walking combined) in Table 1 and the duration of lateral lying per hour were 136
observed from 16 h before, and 48 h after, BFP. Initial data inspection showed that around 80 % of the 137
sows (both crated and penned) did not show extensive nest building earlier than 16 h before BFP. Very 138
little nest building behavior was observed after BFP and therefore only the last 16 h before BFP was 139
included when analyzing the duration of nest building behavior. The 16 h were divided into 2 periods: 140
P1 was 16 h to 3 h before BFP and P2 was 2 h before BFP until BFP. After BFP the 48 h observations 141
of postural changes and lateral lying were divided into 2 periods: P3 was 0 to 8 h after BFP and P4 was 142
9 to 48 h after BFP. 143
144
From BFP until 48 h after, the total number of incidents, where piglets were either crushed or at risk of 145
being crushed (defined as a piglet being trapped underneath the sow’s body but surviving, or the piglet 146
being hit, and knocked out of the way by the sow during a postural change (Marchant et al., 2001) was 147
observed. 148
Page 7 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

8
8
149
Between 18 and 36 h after BFP the sow’s response to the playback sound of a screaming piglet was 150
tested (Thodberg et al., 1998). The test was repeated at 4 d after farrowing. The scream of an alien 151
piglet that was held by a person who firmly pressed the body of the piglet was recorded on a tape 152
recorder (AV30 MkII, Dan-sound Educational, Denmark) and a standardized 2-min sequence was used 153
during testing. The playback recorder was placed on the roof of the piglet box in the pens and on the 154
bars at the back of the crates. The test was not started until at least 5 min after a nursing bout. The 155
observer remotely switched on the playback recorder while the sow was lying laterally. Sow behavior 156
was observed from a remote video. The sound of the screaming piglet was played until the sow was 157
standing up or for a maximum of 2 min. The latency until the sow responded either by rolling over to 158
sternal lying, by sitting, or standing up was recorded during the test. 159
160
Other Recordings 161
The length of pregnancy was recorded by calculating the number of days from first insemination until 162
the day of BFP. The total number of piglets born was counted and all dead piglets were necropsied to 163
count the number of stillborn piglets and the number of crushed piglets. A piglet was categorized as 164
being stillborn when the piglet had not been breathing and a piglet was defined as crushed when 165
traumas were visible as internal or external lacerations (for methodology see Damm et al., 2005b). 166
167
Statistical Analysis 168
The following basic generalized linear model was used to analyze data: 169
Model (1): Yijk =µ + αi + βj + αβij + Xijk + εijk 170
Page 8 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

9
9
In model (1) µijk is the expected value of the response variable, αi is the effect of the ith treatment (LC, 171
LP, EP), βj is the effect of the jth parity (primiparous sows, multiparous sows), Xijk is the number of 172
total piglets born in the kth sow, and ε is the error term. The experimental unit for treatment and parity 173
effects was sows. 174
175
In the analysis of the length of gestation, the average inter-birth interval as well as the variation in the 176
inter-birth interval, the total number of near-crushing situations, the number of postural changes per 177
hour, the time spent in lateral lying per hour and the duration of nest building behavior per hour, the 178
response value Yijk was normally distributed with N(µijk,σ2). Proc Mixed in SAS (SAS Inst. Inc, Cary, 179
NC) version 9.1 was used to analyze these data. 180
181
For time spent nest building the basic model (1) was extended to include periods (P1 and P2) as a fixed 182
effect and sows as a normally distributed random effect against which the effects of treatment and 183
parity were tested. A similar model was used when analyzing the number of postural chances and time 184
spent in lateral lying, where data from 16 h before BFP until 48 h after BFP was used. In conjunction 185
with P1 and P2 two additional periods were included in the fixed period effect, P3 and P4. 186
187
Data on mortality were not normally distributed and were thus analyzed in a generalized linear model 188
for Poisson distributed data ( McCullagh and Nelder, 1989) using the Proc Genmod in SAS. Therefore, 189
when analyzing data on mortality (still born, crushed and dead of liveborn), Yijk was set to logit (Pijk)190
where Pijk is the expected number of dead piglets in the ijkth litter. The response value Yijk was 191
Page 9 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

10
10
approximately binominal distributed with Bin(Pijk, Nijk), where Nijk is the number of total piglets born 192
in the ijkth litter. Due to a tendency for over dispersion, the PSCALE option in Proc Genmod in SAS. 193
194
In analysis of the piglet scream test it turned out that more than 40 % of the sows did not respond to the 195
test. Therefore data were dichotomized into two categories: sows that responded within 2 min and sows 196
that did not. When analyzing the probability that sows were responding to the piglet scream test on d 1 197
and d 4, Yijk was set to logit (Pijk), where Pijk is the expected probability of sows responding in the 198
piglet scream test. The observed response of the sow to the piglet scream test in the ijkth litter is Yijk ∼199
Bin (Pijk). 200
201
Pairwise comparisons of treatment effects for each parity class were made using the contrast statement 202
in the respective procedures of SAS. Estimated LS means ± SE are shown in the text, figures, and 203
tables for the normally distributed data whereas data on mortality and responsiveness to the piglet 204
scream test is shown as LS means with 95 % confidence intervals. 205
206
RESULTS 207
Two primiparous sows on the LP treatment and 1 primiparous sow on the LC treatment were omitted 208
from the analysis, due to various technical problems with the video recordings of these animals. 209
210
Page 10 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

11
11
Farrowing 211
Length of gestation was affected by parity (P = 0.03), with longer gestation length for primiparous 212
sows than for multiparous sows. There was no effect of treatment (P = 0.20) on length of gestation 213
(Table 2). 214
215
The mean inter-birth-interval (IBI) was affected by an interaction between treatment and parity (P =216
0.04). The IBI was not different between the EP sows and the LP sows either in primiparous (P = 0.83) 217
or in multiparous sows (P = 0.35). In contrast, the IBI of primiparous sows was longer in the LC sows 218
than in the LP sows (P = 0.02), whereas there was no difference (P = 0.27) in the IBI between the 219
multiparous LC and LP sows (Table 2). Furthermore the IBI increased with increasing number of total 220
piglets born (F1,50 = 4.7, P = 0.03). There were no differences in IBI between primiparous and 221
multiparous sows either in LC sows (P = 0.14), LP sows (P = 0.09) or EP sows (P = 0.40). 222
223
A similar interaction between treatment and parity was found for the variation in the IBIs (P = 0.01). 224
The variation in the IBI was greater for the primiparous LC sows than for the primiparous LP (P =225
0.006) whereas there was no difference (P = 0.18) between multiparous LC sows and multiparous LP 226
sows (Table 2). The primiparous LC sows had a greater variation than multiparous LC sows (P = 0.06) 227
whereas primiparous LP sows had less variation in IBI than multiparous LP sows (P = 0.03; Table 2). 228
Also the percentage of stillborn piglets was affected by the same interaction (P = 0.003), in that 229
primiparous LC sows had greater percentage of stillborn piglets than primiparous LP sows (P = 0.004). 230
There was no difference between LC and LP sows (P = 0.26) and between LP and EP sows (P = 0.60) 231
for the multiparous sows. Primiparous LC sows also had greater percentage of stillborn piglets than 232
multiparous LC sows (P < 0.001). The back transformed estimated mean percentages of stillborn 233
Page 11 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

12
12
piglets are shown in Table 2 along with the significances level of the chi-square test of the contrasts 234
between the treatments within parity class and between parity class within treatment (Table 2). Using 235
these estimates the odds ratio of giving birth to a stillborn piglet in primiparous LC sows was 236
calculated to be 5 times greater than for primiparous LP sows (95 % confidence interval was ranging 237
1.6 to 14.7; X2 = 8, P = 0.004). The odds ratio for primiparous LP sows was not different from that of 238
primiparous EP sows (2.7 with a 95 % confidence interval ranging 0.5 to 13.0) (P = 0.20). Treatments 239
were not different in the percentage of crushed piglets (P = 0.68) or in the percentage of liveborn 240
piglets that died (P = 0.28; Table 2). 241
242
Nesting Behavior and Postural Changes 243
The time spent nest building tended to be different between treatments (F2,53 = 2.6, P = 0.08). However, 244
the EP and LP sows (LP = 101 ± 48 s/h, EP = 142 ± 50 s/h; P = 0.50) were not different and neither 245
were the LP and LC sows (LC = 46 ± 17 s/h, LP = 101 ± 48 s/h; P = 0.13). Time spent nest building 246
was influenced by an interaction between period and parity (P = 0.002) in that primiparous sows and 247
multiparous sows spent an equal amount of time nest building during P1 (primiparous = 221 ± 84 s/h; 248
multiparous = 387 ± 140 s/h) whereas primiparous sows spent less time nest building than multiparous 249
sows during P2 (primiparous = 7 ± 3 s/h, multiparous = 92 ± 33 s/h; P = 0.002). 250
251
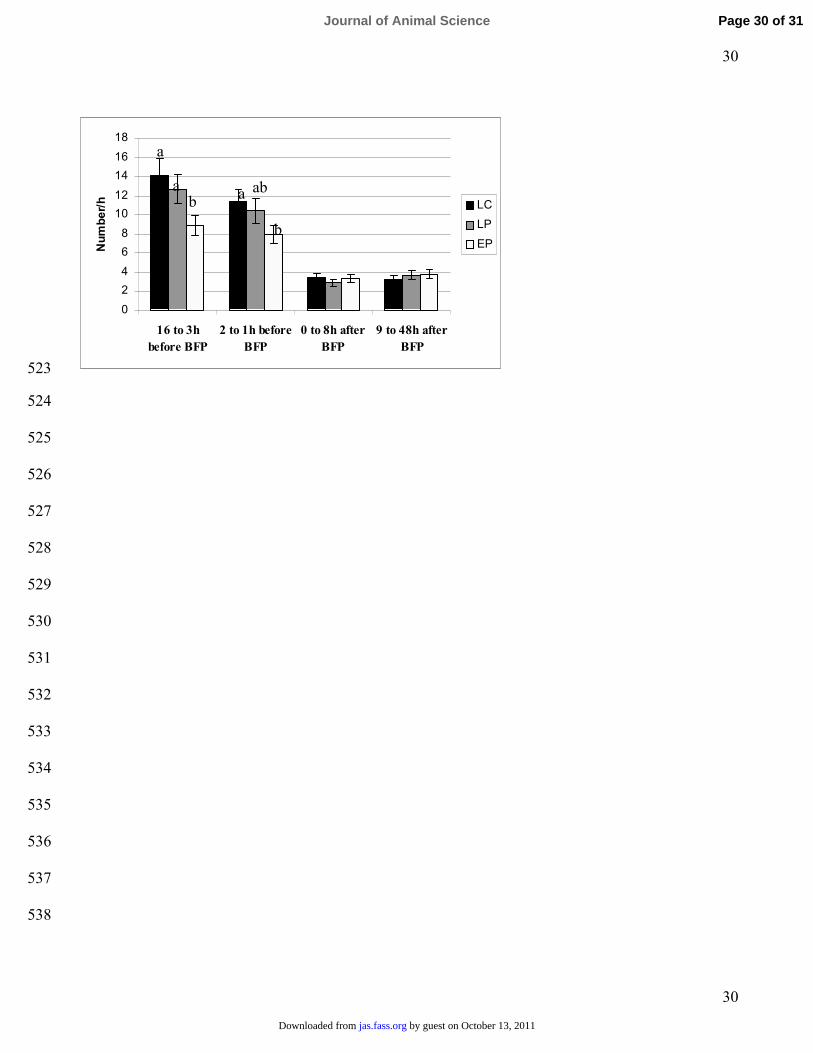
The number of postural changes was affected by an interaction between period and treatment (P =252
0.02). During P1 and P2, respectively, EP sows had less postural changes than LP sows (P = 0.04 and 253
P = 0.12), whereas EP sows and LP sows did not differ (P > 0.40) during P3 and P4. There were no 254
Page 12 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

13
13
differences (P > 0.33) between LP and LC sows in the number of postural changes during any of the 255
periods (Figure 2). 256
257
Time spent lying lateral was not affected by treatment (P = 0.68). Time spent lying lateral was 258
influenced by an interaction between period and parity (P = 0.002). Primiparous sows compared to 259
multiparous sows spent more time in lateral lying during P1 (primiparous = 1,138 ± 127 s/h-1,260
multiparous = 762 ± 121 s/h-1; P = 0.03) and P2 (primiparous = 2601 ± 127 s/h, multiparous = 1,856 ±261
121 s/h; P = 0.001) whereas there were no difference in P3 (primiparous = 3,241 ± 127 s/h; 262
multiparous = 3,220 ± 121 s/h) and P4 (primiparous = 2,873 ± 127 s/h; multiparous = 2,658 ± 121 s/h). 263
264
Near-Crushing Incidents and Odds Ratio for Responding to the Sound of a Screaming Piglet 265
The number of near-crushing incidents was affected by parity (P = 0.03) and total number of piglets 266
born (P = 0.03). Primiparous sows had more near-crushing situations during the 48-h observation 267
period than multiparous sows (2.25 ± 1.37 vs. 0.69 ± 0.74) and the number of near crushing situations 268
increased with increasing number of total piglets born (estimated regression coefficient = 0.11 ± 0.05). 269
The number of near-crushing situations tended to differ between treatments (P = 0.08), in that LC sows 270
tended to have fewer near-crushing situations than LP (LC = 0.52 ± 0.71, LP = 1.82 ± 1.35; P = 0.08). 271
Crushing incidences were not different for LP and EP sows (LP = 1.82 ± 1.35, EP = 2.04 ± 1.40). 272
273
The odds ratio for sows to respond to the sound of a screaming piglet did not differ between treatments 274
and between parity on d 1 or d 4. On d 1, 59 % responded to the scream test whereas on d 4, 56 %275
responded. 276
Page 13 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

14
14
277
DISCUSSION 278
Effects during the Nest Building Phase 279
Results showed that introducing sows to farrowing pens close to the time of farrowing increased the 280
number of postural changes but had no influence on the time spent nest building. This may indicate that 281
while the motivation to nest build is unaffected by the late move, the sows were still responding to the 282
new environment by general restlessness. Restlessness was not further increased in crated sows 283
introduced late compared to penned sows introduced late. Earlier studies have shown that being crated 284
compared to being penned on introduction to the farrowing unit increases restlessness during the nest 285
building phase (Jarvis et al., 2001) and induces increased HPA–axis activity (Cronin et al., 1991; 286
Lawrence et al., 1994; Jarvis et al., 1997, 2001). It is thus surprising that we did not find a similar effect 287
of crating in the present study. The effect on restlessness of introducing the sow late to the farrowing 288
unit may be greater than the effect of crating. 289
290
Even though crated sows performed a similar number of postural changes they tended to perform less 291
nest building behavior than penned sows moved late. An effect of environment on nesting behavior has 292
been found in several earlier studies (Thodberg et al., 2002; Damm et al., 2003 Jarvis et al., 2001) 293
where sows in farrowing pens spent more time nest building than sows in farrowing crates. In addition, 294
the nature and the amounts of substrate directed behavior differ between the housing systems (Jarvis et 295
al., 2001). Primiparous sows in the present experiment appeared to start their nest building later than 296
multiparous sows, which has also been found in earlier studies (Jarvis et al., 2001; Thodberg et al., 297
2002). 298
299
Page 14 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

15
15
Effects on the Progress of Farrowing and Stillborn Piglets 300
Being introduced late to pens did not influence the progress of farrowing, and the percentage of 301
stillborn and crushed piglets either in primiparous sows or multiparous sows. This indicated that the 302
stress of being introduced to a novel environment in itself is not large enough to interfere with the 303
progress of farrowing. 304
305
In contrast, crating sows compared to penning them upon late introduction affected both the progress of 306
farrowing and the number of stillborn piglets. The effect was however only present in the primiparous 307
sows. Several authors have shown that crating of primiparous sows is stressfull and induces changes in 308
the HPA-axis during the peri-parturient period even when the sows were introduced 5 d before 309
expected farrowing (Lawrence et al., 1994; Jarvis et al, 1997, 2001). An additional activation of the 310
HPA-axis and or increased opioid secretion caused by the stress of being crated in primiparous sows 311
may contribute to the longer IBI, increased variation in the IBIs and greater percentage of stillborn 312
piglets seen in the present study. Effects of crating primiparous sows on the progress of farrowing and 313
the percentage of stillborn piglets, are likely to be the combined effects of the novelty of confinement 314
(Cronin et al., 1991; Lawrence et al., 1994), restriction of the highly motivated nest-building behavior 315
(Lawrence et al., 1994) and farrowing in a novel environment (Lawrence et al., 1992). It seems less 316
likely that being in a crate during farrowing in itself is stressful because neither Gilbert et al. (1997) nor 317
Jarvis et al. (1998) found any effect of crating on the HPA- axis activity during farrowing. This is 318
possibly due to the fact that sows spent the majority of farrowing lying still anyway. Another 319
explanation may be, that the natural physiological increase in cortisol is so high during farrowing that it 320
overrides any additional stress increased elevation of cortisol. 321
322
Page 15 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

16
16
The difference between the crated primiparous sows and the crated multiparous sows in the progress of 323
farrowing and the percentage of stillborn piglets, that were not evident in the penned sows, further 324
strengthens the importance of the novelty of confinement. Primiparous sows were crated for the first 325
time in their life when introduced to the farrowing crate and they may thus respond stronger to the 326
stress of crating due to lack of experience with such a restrictive environment. This is supported by 327
Jarvis et al. (2002) and Thodberg et al. (2002) who found that sows in their second parity showed some 328
signs of adaptation to crating in response to previous experience, in both nest building behavior and 329
progress of farrowing. 330
331
A change in the housing method from the gestation period to the farrowing period has been shown to 332
affect both the peri-parturient behavior (Beattie et al., 1995; Harris and Gonyou, 1998; Boyle et al., 333
2000, 2002) and the percentage of stillborn piglets (Cronin et al. 1996). This supports the concept that 334
the novelty of being crated for the first time in the farrowing unit is stressful to an extent that it affects 335
both behavior and the birth process. The difference in the percentage of stillborn piglets for the 336
primiparous sows moved late to crates compared to the primiparous sows moved late to pens was 337
greater in the present study than what was found by Cronin et al. (1996) where primiparous sows were 338
moved earlier to either crates or pens. Beside the more long lasting negative effect of crating on the 339
birth process (Biensen et al., 1996; Cronin et al., 1996) there may be an additive effect of the novelty of 340
crating. 341
342
Due to the current EU legislation the use of confinement during gestation will be banned from 2014. 343
However, crating is still allowed and widely used during farrowing. Seen in the light of the negative 344
effect this shift in housing system has on the progress of farrowing and the percentage of stillborn 345
Page 16 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

17
17
piglets as emphasized by both the present and previous studies it would be beneficial to hasten 346
development of farrowing pens for loose sows. 347
348
Effects on Posture and Maternal Behavior after Birth 349
The majority of crushing and death in general occur within the first 1 to 2 d of the piglets’ life. During 350
the first 8 h after BFP the sows are mainly in lateral lying (Pedersen et al., 2003). Because crushing 351
occurs during postural changes, a high level of restlessness during this particular period may constitute 352
a potential risk situation for newborn piglets (Damm et al., 2005a,b). However, late introduction to the 353
farrowing pens did not increase restlessness during this period, or during the next 9 to 48 h, indicating 354
that sows were not influenced by the late introduction to an extent that increase restlessness. 355
356
The number of postural changes seems important for piglets’ risk of being crushed. In addition, it may 357
be equally important that the sow perform pre-lying behavior that effectively clusters the piglets before 358
the sow lies down. Such behaviors have been shown to reduce the number of near crushing situations 359
(Marchant et al., 2001). Sows that are disturbed for example due to novelty of the environment or due 360
to restriction in their movement may be less attentive during lying down and the risk of near crushing 361
situations may thus be greater. The present study did, however not find increased number of near 362
crushing situations in the late introduced sows compared to the early introduced sows indicating that 363
sows readily adapted to the new environment. Penned sows tended to have more near-crushing 364
incidents than crated sows, however without a parallel increase in percentage of piglets that died from 365
crushing. This may indicate that even though the crate helps reduce the number of near crushing 366
situations it may not effectively reduce the risk of crushing. Weber et al. (2007) found in a large survey 367
on crated and penned sows that a greater percentage of piglets died from crushing in penned sows than 368
Page 17 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

18
18
in crated sows, whereas the total mortality did not differ between the 2 systems. Perhaps weak piglets 369
are more easily crushed in a pen than in a crate because weak piglets are unable to respond properly to 370
pre-lying behavior. On the other hand weak piglets may die from other causes such as hypothermia, 371
lack of milk or disease in a crate system. Even though primiparous and multiparous sows had the same 372
number of postural changes after BFP, the primiparous sows had more near-crushing incidents than 373
multiparous sows. An explanation for this could be that primiparous sows are less experienced in 374
performing pre-lying behavior and in clustering the piglets before lying down. However this has never 375
been studied in detail. 376
377
Even though both a large number of postural changes and a large number of near crushing situations 378
may be considered a risk to piglet survival, data on crushing does not always support this (Pedersen et 379
al., 2006). How the sow responds when a piglet is trapped underneath it may be equally important. A 380
trapped piglet will scream if possible and it will have a better chance to escape if the sow responds to 381
its scream by changing posture than if the sow remains lying (Weschler and Hegglin, 1997). Previous 382
research has shown much variability in the way sows respond to the scream of a trapped piglet 383
(Grandinson et al., 2003). Although the trait seems slightly heritable (Grandinson et al., 2003), 384
environmental disturbances may also affect the response. For example novelty of the environment may 385
take the sows attention away from the newborn piglets or make them more responsive. However, no 386
such effects were found in the present study by the late introduction to pens. 387
388
The present study showed that sows introduced late to farrowing pens did not respond to the new 389
environment to an extent that affected the progress of farrowing and behaviors of importance for piglet 390
survival. Therefore, late introduction to the farrowing environment does not seem to be a problem 391
Page 18 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

19
19
when sows are housed in farrowing pens on introduction. In contrast, late introduction to crates 392
compared to pens severely affected the progress of farrowing and the percentage of stillborn piglets in 393
primiparous sows. Whether this effect was due to the novelty of crating or crating in itself cannot be 394
verified from the present study because no sows were introduced early to crates. 395
396
LITERATURE CITED 397
Andersen, I. L., S. Berg, and K. E. Boe. 2005. Crushing of piglets by the mother sow (Sus scrofa) - 398
purely accidental or a poor mother? Appl. Anim. Behav. Sci. 93:229-243. 399
Beattie, V. E., N. Walker, and I. A. Sneddon. 1995. Effect of rearing environment and change of 400
environment on the behavior of gilts. Appl. Anim. Behav. Sci. 46:57-65. 401
Biensen, N. J., E. H. von Borell, and S. P. Ford. 1996. Effects of space allocation and temperature on 402
periparturient maternal behaviors, steroid concentrations, and piglet growth rates. J. Anim. Sci. 403
74:2641-2648. 404
Boyle, L. A., F. C. Leonard, P. B. Lynch, and P. Brophy. 2000. Influence of housing system during 405
gestation on the behavior and welfare of gilts in farrowing crates. Anim. Sci. 71:561-570. 406
Boyle, L. A., F. C. Leonard, P. B. Lynch, and P. Brophy. 2002. Effect of gestation housing on behavior 407
and skin lesions of sows in farrowing crates. Appl. Anim. Behav. Sci. 76:119-134. 408
Cronin, G. M., J. L. Barnett, F. M. Hodge, J. A. Smith, and T. H. McCallum. 1991. The Welfare of 409
Pigs in 2 Farrowing Lactation Environments - Cortisol Responses of Sows. Appl. Anim. Behav. 410
Sci. 32:117-127. 411
Page 19 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

20
20
Cronin, G. M., G. J. Simpson, and P. H. Hemsworth. 1996. The effects of the gestation and farrowing 412
environments on sow and piglet behavior and piglet survival and growth in early lactation. 413
Appl. Anim. Behav. Sci. 46:175-192. 414
Damm, B. I., B. Forkman, and L. J. Pedersen. 2005a. Lying down and rolling behavior in sows in 415
relation to piglet crushing. Appl. Anim. Behav. Sci. 90:3-20. 416
Damm, B. I., L. Lisborg, K. S. Vestergaard, and J. Vanicek. 2003. Nest-building, behavioral 417
disturbances and heart rate in farrowing sows kept in crates and Schmid pens. Livest. Prod. Sci. 418
80:175-187. 419
Damm, B. I., L. J. Pedersen, T. Heiskanen, and N. P. Nielsen. 2005b. Long-stemmed straw as an 420
additional nesting material in modified Schmid pens in a commercial breeding unit: effects on 421
sow behavior, and on piglet mortality and growth. Appl. Anim. Behav. Sci. 92:45-60. 422
Gilbert, C. L., M. I. Boulton, M. L. Forsling, J. A. Goode, and T. J. McGrath. 1997. Restricting 423
maternal space during farrowing in the pig. Effects on oxytocin, vasopressin and cortisol 424
secretion following vagino-cervical stimulation and administration of naloxone. Anim. Reprod. 425
Sci. 46:245-259. 426
Grandinson, K., L. Rhydmer, E. Strandberg, and K. Thodberg. 2003. Genetic analysis of on-farm tests 427
of maternal behaviour in sows. Livest. Prod. Sci. 83:141-151. 428
Harris, M. J., and H. W. Gonyou. 1998. Increasing available space in a farrowing crate does not 429
facilitate postural changes or maternal responses in gilts. Appl. Anim. Behav. Sci. 59:285-296. 430
Page 20 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

21
21
Jarvis, S., S.K. Calvert, J. Stevenson, N. vanLeeuwen, A.B. Lawrence. 2002. Pituitary-adrenal 431
activation in pre-parturient pigs (sus scrofa) is associated with behavioural restriction due to 432
lack of space rather than nesting substrate. Anim. Welfare, 11:371-384. 433
Jarvis, S., A.B. Lawrence, K.A. McLean, J. Chirnside, and S.K. Calvert. 1997. The effect of the 434
environemnet on behavioural activity, ACTH, beta-endorpines and cortisol in pre-parturient 435
gilts. Anim. Sci. 65:465-472. 436
Jarvis, S., A. B. Lawrence, K. A. McLean, J. Chirnside, L. A. Deans, and S. K. Calvert. 1998. The 437
effect of environment on plasma cortisol and [beta]-endorphin in the parturient pig and the 438
involvement of endogenous opioids. Anim. Reprod. Sci. 52:139-151. 439
Jarvis, S., B. J. Van der Vegt, A. B. Lawrence, K. A. McLean, L. A. Deans, J. Chirnside, and S. K. 440
Calvert. 2001. The effect of parity and environmental restriction on behavioral and 441
physiological responses of pre-parturient pigs. Appl. Anim. Behav. Sci. 71:203-216. 442
Jensen, P. 1986. Observations on the Maternal-Behavior of Free-Ranging Domestic Pigs. Appl. Anim. 443
Behav. Sci. 16:131-142. 444
Lawrence, A.B., K.A. McLean, S. Jarvis, C.L. Gilbert, and J.C. Petherick. 1997. Stress and parturition 445
in the pig. Reprod. Dom. Anim. 32:231-236. 446
Lawrence, A. B., J. C. Petherick, K. A. McLean, L. A. Deans, J. Chirnside, A. Gaughan, E. Clutton, 447
and E. M. C. Terlouw. 1994. The effect of environment on behavior, plasma cortisol and 448
prolactin in parturient sows. Appl. Anim. Behav. Sci. 39:313-330. 449
Page 21 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

22
22
Lawrence, A. B., J. C. Petherick, K. Mclean, C. L. Gilbert, C. Chapman, and J. A. Russell. 1992. 450
Naloxone Prevents Interruption of Farrowing and Increases Plasma Oxytocin Following 451
Environmental Disturbance in Parturient Sows. Physiol. & Behav. 52:917-923. 452
Marchant, J. N., D. M. Broom, and S. Corning. 2001. The influence of sow behavior on piglet mortality 453
due to crushing in an open farrowing system. Anim. Sci. 72:19-28. 454
McCullagh, P., and J.A. Nelder. 1989. Generalized Linear Models. 2nd Edn. London: Chapman & Hall. 455
Pedersen, L.J., B.I. Damm, J.M. Marchant, and K.H. Jensen. 2003. Effects of feed-back from the nest 456
on maternal responsiveness and postural changes in primiparous sows during the first 24 h after 457
farrowing on set. Appl. Anim. Behav. Sci. 83:109-124. 458
Pedersen, L. J., E. Jorgensen, T. Heiskanen, and B. I. Damm. 2006. Early piglet mortality in loose-459
housed sows related to sow and piglet behavior and to the progress of farrowing. Appl. Anim. 460
Behav. Sci. 96:215-232. 461
Rushen, J., J. Ladewig, and A. M. B. Depassille. 1995. A Novel Environment Inhibits Milk Ejection in 462
the Pig But Not Through Hpa Activity. Appl. Anim. Behav. Sci. 45:53-61. 463
Thodberg, K., M. S. Herskin, and K. H. Jensen. 1998. Developement of a test for maternal behavior in 464
sows - "the screaming piglet test". Page 140 in Procedings of the 32nd International Congress 465
of the International Society for Applied Ethology, Institut National de la Recherche 466
Agronomique, France. 467
Page 22 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

23
23
Thodberg, K., K. H. Jensen, and M. S. Herskin. 2002. Nest building and farrowing in sows: relation to 468
the reaction pattern during stress farrowing environment and experience. Appl. Anim. Behav. 469
Sci. 77:21-42. 470
Weber, R., N.M. Keil, M. Fehr, and R. Horat. 2007. Piglet mortality on farms using farrowing systems 471
with and without crates. Anim. Welfare 16:277-279. 472
Wechsler, B., and D. Hegglin. 1997. Individual differences in the behaviour of sows at the nest-site and 473
the crushing of piglets. Appl. Anim. Behav. Sci. 51:39-49. 474
Page 23 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

24
24
475Table 1. Ethogram for the behavioral elements observed from 16 h before birth of
first piglet (BFP) until 48 h after BFP
Behavior Description
Lateral lying Sow is lying on the side with one shoulder touching the floor.
Sternal lying Sow is lying on the belly without a shoulder touching the floor.
Sitting Both feet on the sow’s front legs as well as the sow’s posterior
are touching the floor.
Standing/walking Sow stands on all four legs.
Nest building Sow stands on all four legs engaged in either of the following
behaviors: Rooting (repeated snout movements on the floor with
or without straw), pawing, carrying straw in the mouth and
arranging straw in the nest (up and down head movements
directed towards straw on the ground or in a pile).
476
Page 24 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

25
25
477
Table 2. Progress of farrowing and mortality for primiparous and multiparous sows moved day 114 of
pregnancy to pens (LP) or crate (LC) or moved day 109 of pregnancy to pens (EP) for farrowing
Primi parous sows Multi parous sows
Treatments
(n)
LC
(9)
LP
(8)
EP
(10)
LC
(10)
LP
(10)
EP
(10)
Length of
gestation, d1
117.5 ± 0.4 116.3 ± 0.4 117.3 ± 0.4 115.8 ± 0.4 116.3 ± 0.4 116.8 ± 0.4
Inter-birth
interval, min1
29 ± 4.9 a 16 ± 2.9b 17 ± 2.7b 21 ± 3.4 b 27 ± 4.3 b 22 ± 3.6 b
Variation in the
inter-birth
interval, min1
35 ± 8.3ax 16 ± 3.6 bx 18 ± 4.2b 22 ± 5.1 by 26 ± 6.9by 26 ± 6.1 b
Total piglets
born2
13.4 ±3.8 13.3 ±3.6 14.5 ± 2.7 16.5 ± 2.27 15.8 ±2.8 15.3 ±2.3
Stillborn ,% 3 21 (14 to 30)ax 5 (2 to 12)b 2 (1 to 7)b 4 (2 to 8) by 7 (4 to 13)b 5 (2 to12) b
Total dead of
liveborn,% 3
6 (2 to 18) 19 (9 to 34) 18 (9 to 32) 14 (6 to 30) 18(9 to 35) 22 (12 to36)
Crushed,% 3 5 (1 to 15) 15 (7 to 29) 12 (5 to 24) 4 (1 to 17) 8 (3 to 23) 13 (6 to 25)
a,b Row values within parity with different superscripts differ (P < 0.05). x,y Row values within treatment different superscripts differ (P < 0.05). 1 Lsmeans (± SE). 2 The number given is the raw mean ± std because no statistical calculation was carried out. 3 The estimated mean as percentage of total born piglets with the 95 % confidence interval shown in bracket.
478
Page 25 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

26
26
Figure 1. Schematic drawing and picture of the farrowing environment for (A) sows in pens and (B) 479
sows in crates 480
481
Page 26 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

27
27
482A)
B)
483
Straw rack
Feed for sow
Water for sow and piglets
Feed for piglets
Creep area
3,27 m
2,68 m
Straw rack
Feed for sow
Water for sow and piglets
Feed for piglets
Creep area
3,27 m
2,68 m 2,35 m
0,60 m
2,00 m0,83 m0,65 m
Feed for piglets
Creep area
0,70 m
Heat lamp
Water for piglets
0,5 m
Feed for sow
2,35 m
0,60 m
2,00 m0,83 m0,65 m
Feed for piglets
Creep area
0,70 m
Heat lamp
Water for piglets
0,5 m
2,35 m
0,60 m
2,00 m0,83 m0,65 m
Feed for piglets
Creep area
0,70 m
Heat lamp
Water for piglets
0,5 m
Feed for sow
Figure 1 E-2007-0749
Page 27 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

28
28
Figure 2. The number of postural changes per h in different periods relative to the birth of the first 484
piglet (BFP) for sows moved d 114 to farrowing crates (LC), sows moved d 114 to farrowing pens (LP) 485
and sows moved d 95 to farrowing pens (EP). a,b Means with different superscript indicate significant 486
difference between columns within periods (P < 0.05)487
488
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
505
506
Page 28 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

29
29
507
508
509
510
511
512
513
514
515
516
517
518
519
520
521
522
Page 29 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from
30
30
523
524
525
526
527
528
529
530
531
532
533
534
535
536
537
538
02468
1012
141618
16 to 3hbefore BFP
2 to 1h beforeBFP
0 to 8h afterBFP
9 to 48h afterBFP
Num
ber/h LC
LP
EP
a
ab a
b
ab
Page 30 of 31Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

31
31
539
540
541
542
543
544
545
546
547
548
549
550
551
552
Figure 2 E-2007-0749
Page 31 of 31 Journal of Animal Science
by guest on October 13, 2011jas.fass.orgDownloaded from

Citations
ticleshttp://jas.fass.org/content/early/2008/05/09/jas.2007-0749.citation#otherarThis article has been cited by 1 HighWire-hosted articles:
by guest on October 13, 2011jas.fass.orgDownloaded from




















