
Specific enumeration of the probiotic strain Enterococcus Faecium NCIMB 10415 in the intestinal...
10
SPECIFIC ENUMERATION OF THE PROBIOTIC STRAIN ENTEROCOCCUS FAECIUM NCIMB 10415 IN THE INTESTINAL TRACT AND IN FAECES OF PIGLETS AND SOWS M. MACHA a, *, D. TARAS a , W. VAHJEN a , A. ARINI b and O. SIMON a a Institute for Animal Nutrition, Free University of Berlin, Germany; b Cerbios-Pharma SA, Barbengo, Switzerland (Received 20 July 2004, accepted 20 September 2004) The intestinal bacterium Enterococcus faecium NCIMB 10415 (E. faecium SF68) has been used for more than a decade as a probiotic strain in animal nutrition as well as in the prevention and treatment of diarrhoea in humans. Beneficial effects have been shown in feeding and clinical trials. However, the strain has no selective growth markers and monitoring in the intestinal tract is impossible by cultivation. Using specific nucleotide sequences, in this study a probe for colony hybridization was constructed in order to quantify this probiotic strain in feed and intestinal and faecal samples from piglets and sows. The probiotic strain showed almost constant amounts in sow faeces (1.8×10 5 cfu/g wet weight), while contents in digesta and piglet faeces varied on a lower level depending on gut section and piglet age. The ratio of specific probiotic counts and total enterococci was much lower than in sow faeces however the strain could be detected reliably in faeces already on the 14th day of life. The application of the colony hybridization method enables for the first time the selective detection of the widely used probiotic E. faecium NCIMB 10415 strain among total Enterococcus spp. counts of digesta, faeces and feed. It is now possible to monitor the presence of the probiotic in the intestinal tract and faeces. Results of this study have implications for the proposed modes of action of probiotics in animal nutrition. Keywords: Probiotics; Enterococcus faecium; Enumeration; DNA hybridisation; Pigs 1. INTRODUCTION Probiotics have been used to treat diarrhoea in humans (Marteau et al., 2001) and as feed additives in animal production, especially in pigs and poultry (Jadamus et al., 2001; Reuter, 2001; Jadamus et al., 2002; Vahjen et al., 2002). Proposed probiotic effects in animal nutrition include improved performance and hygiene control, but underlying principles are still a matter of discussion (Tannock, 2002). A prerequisite for probiotics is its viability in the intestinal tract (Fuller, 1989; Saarela et al., 2000). To elucidate the modes of action of probiotic feed additives it is essential to acquire precise data about viable cell numbers within the digestive tract. *Corresponding author: M. Macha, Institute for Animal Nutrition, Free University of Berlin, Bru¨mmerstraße 34, D-14195 Berlin, Germany Archives of Animal Nutrition, December 2004, Vol. 58(6), pp. 443 – 452 ISSN 1745-039X print; ISSN 1477-2817 online # 2004 Taylor & Francis Ltd DOI: 10.1080/00039420400020058
-
Upload
boehringer-ingelheim -
Category
Documents
-
view
0 -
download
0
Transcript of Specific enumeration of the probiotic strain Enterococcus Faecium NCIMB 10415 in the intestinal...

SPECIFIC ENUMERATION OF THE PROBIOTIC
STRAIN ENTEROCOCCUS FAECIUM NCIMB 10415 IN
THE INTESTINAL TRACT AND IN FAECES OF
PIGLETS AND SOWS
M. MACHAa,*, D. TARASa, W. VAHJENa, A. ARINIb and O. SIMONa
aInstitute for Animal Nutrition, Free University of Berlin, Germany; bCerbios-Pharma SA,Barbengo, Switzerland
(Received 20 July 2004, accepted 20 September 2004)
The intestinal bacterium Enterococcus faecium NCIMB 10415 (E. faecium SF68) has been used for more thana decade as a probiotic strain in animal nutrition as well as in the prevention and treatment of diarrhoea inhumans. Beneficial effects have been shown in feeding and clinical trials. However, the strain has no selectivegrowth markers and monitoring in the intestinal tract is impossible by cultivation. Using specific nucleotidesequences, in this study a probe for colony hybridization was constructed in order to quantify this probioticstrain in feed and intestinal and faecal samples from piglets and sows. The probiotic strain showed almostconstant amounts in sow faeces (1.8�105 cfu/g wet weight), while contents in digesta and piglet faeces varied ona lower level depending on gut section and piglet age. The ratio of specific probiotic counts and totalenterococci was much lower than in sow faeces however the strain could be detected reliably in faeces alreadyon the 14th day of life. The application of the colony hybridization method enables for the first time theselective detection of the widely used probiotic E. faeciumNCIMB 10415 strain among total Enterococcus spp.counts of digesta, faeces and feed. It is now possible to monitor the presence of the probiotic in the intestinaltract and faeces. Results of this study have implications for the proposed modes of action of probiotics inanimal nutrition.
Keywords: Probiotics;Enterococcus faecium; Enumeration; DNAhybridisation; Pigs
1. INTRODUCTION
Probiotics have been used to treat diarrhoea in humans (Marteau et al., 2001) and asfeed additives in animal production, especially in pigs and poultry (Jadamus et al., 2001;Reuter, 2001; Jadamus et al., 2002; Vahjen et al., 2002). Proposed probiotic effects inanimal nutrition include improved performance and hygiene control, but underlyingprinciples are still a matter of discussion (Tannock, 2002). A prerequisite for probioticsis its viability in the intestinal tract (Fuller, 1989; Saarela et al., 2000). To elucidate themodes of action of probiotic feed additives it is essential to acquire precise data aboutviable cell numbers within the digestive tract.
*Corresponding author: M. Macha, Institute for Animal Nutrition, Free University of Berlin,Brummerstraße 34, D-14195 Berlin, Germany
Archives of Animal Nutrition, December 2004, Vol. 58(6), pp. 443 – 452
ISSN 1745-039X print; ISSN 1477-2817 online # 2004 Taylor& Francis LtdDOI: 10.1080/00039420400020058

The ease to obtain information about the viability of a bacterial strain depends on theorganism studied. Allochthonous bacteria like Bacillus spp. or yeasts and fungi aregenerally easier to detect. Spore forming probiotics (Bacillus spp.) can easily bequantified, because the intestine harbours no aerobic spore forming bacteria. A simpleheating step during the extraction procedure allows quantification of Bacillus spp.strains (Jadamus et al., 2001).However, autochthonous bacteria like Escherichia coli, lactobacilli, bifidobacteria or
enterococci require more sophisticated methods for their quantification on the strainlevel (Festl et al., 1986; Datta et al., 1987; Betzl et al., 1990; Goverde et al., 1993; Durantet al., 1997; Duez et al., 2000; Numata et al., 2000; Lynch et al., 2002; Blum-Oehler etal., 2003; Brigidi et al., 2003). Thus, molecular biology methods must complement oreven replace conventional cultivation and identification techniques in order tospecifically quantify a single bacterial strain among a multitude of other bacterialstrains present in a sample (Anderson, 2003). Although other approaches have beenestablished like ELISA techniques or immunoblotting (Durant et al., 1997; Duez et al.,2000), molecular methods based on nucleic acid sequence diversity became thecommonly applied method of choice.The application of specific nucleid acid probes for example enables the direct
differentiation on the level of bacterial strains or even only presence of specific traits, ifspecific sequences are known. Colony hybridization is a hybrid of conventional andmolecular microbiology methods (Buluwela et al., 1989), as it combines both cultivationand subsequent hybridization of lysed colonies with a specific probe. The method is wellestablished and allows direct enumeration of bacteria within mixed bacterialpopulations (Datta et al., 1987; Betzl et al., 1990; Numata et al., 2000; Lynch et al.,2002; Radhika et al., 2002). Probes are often directed against rRNA or correspondingrDNA sequences, because these more or less conserved sequences allow phylogeneticdifferentiation down to the species level (Festl et al., 1986; Betzl et al., 1990; Lynch etal., 2002). In order to detect strains within a species, probes are designed that targetcertain genes which encode for specific enzymes, toxins or other cellular componentsequences that are specific to the strain in question (Datta et al., 1987; Goverde et al.,1993; Radhika et al., 2002). Nevertheless, cultivation on selective media is necessary andthus sensitivity is limited for strains that occur in cell counts below the resolution of theselective medium (Duez et al., 2000; Numata et al., 2000).This study was carried out in order to establish a specific detection procedure for the
probiotic strainE. faeciumNCIMB10415,which is an authorised feed additive for variousfarm animals according to the ‘‘List of the authorised additives in feedingstuffs publishedin application of Article 9t (b) of Council Directive 70/524/EEC concerning additives infeedingstuffs’’. Furthermore, the viability of the probiotic strain was studied in thedigestive tract of suckling and weaned piglets as well as in mother sow faeces and feed.
2. MATERIALSANDMETHODS
2.1. Animals and feeding conditions
Pregnant commercial Duroc6German Landrace hybrid sows were fed a barley/wheatbased diet (control group) or the same diet supplemented with 1.6 (+ 1.2)�106 E.faecium NCIMB 10 415 viable cells per gram feed (trial group) starting at 90 days antepartum and continuously throughout the feeding trial. After birth, piglets from control
444 M. MACHA et al.

sows were used to create a control group without supplementation of the probiotic,while piglets from probiotic fed mother sows were allocated to the trial group thatreceived the supplemented feed throughout the trial. Suckling piglets had free access tostarter feed from day 14 up to day 28, where they were weaned and further kept inflatdeck boxes. Starter feed intake was not taken into account, because differences inindividual feed intake prohibited meaningful statistical analysis.
2.2. Sampling and cultivation
Faecal samples from five sows were taken 10 days ante partum as well as 14 days postpartum. Faeces from two piglets per litter were taken on days 14, 28, 35 and 70,respectively. From each litter of the trial group one piglet was euthanized on days 14,28, 35, 56 and digesta samples were taken from stomach, jejunum, ileum and colon. Allpiglet had free access to feed until sedation. Samples were diluted in phosphate-buffered-saline (NaCl 8 g�l71, NA2HPO4 10 mM, NAH2PO4 2 mM, pH 7.4) andcultivated aerobically on Slanetz-Bartley-Agar (Oxoid Ltd., Basingstoke, Hampshire,UK) at 378C for 48 h.
2.3. Probe preparation
The probe was prepared by PCR using specific primers targeting a 1.2 kb sequence on aunique plasmid of E. faecium NCIMB10415 and Digoxigenin labelled nucleotides(Holtke et al., 1995) according to the instructions of the manufacturer (PCR DIGLabeling Mix, Roche, Mannheim, Germany). Primer sequences were: SFB: 5’-AACGTG CTT TTC TCG TTC C-3’ and SFC: 5’-TAGCTTGTCGCCTATCCG-3’. PCRwas carried out in T1 Thermocycler (Biometra, Gottingen, Germany) under thefollowing conditions: 948C denaturation for 30 s, 548C annealing temperature for 50 s,728C elongation for 90 s; 25 cycles, final elongation 2 min at 728C. Specificity of theprobe was evaluated with respective primers in a PCR against a range of Enterococcusspp. reference strains (Table I).
2.4. Colony hybridization
For colony hybridization, colonies fromSlanetz-Bartley agar plateswere transferred ontopositively charged nylon membranes (Roche, Mannheim, Germany). Cell lysis wasachieved by incubation with a lysozyme solution (100 ml solution contained: 1 glysozyme, 0.5 MTris-HCl, 50 mMEDTA,pH 8.0) for 15 minat 378Cfollowedby10 minincubation with 10% SDS solution. DNA was denatured for 15 min in 0.5 M NaOH.After washing in 26 SSC solution (17.5 g�l71 NaCl, 8.8 g�l71 tri-sodium-citrate�2H2O,pH 7.0) DNA was fixed on nylon membranes by baking for 30 min at 1208C.Hybridization was carried out in a hybridization oven (Biometra, Gottingen,
Germany) using a commercial hybridization kit (DIG easyHyb Granules, Roche,Mannheim, Germany). After prehybridization for 30 min at 428C, membranes and1.5 ml PCR generated DNA probe per ml hybridization solution were incubatedovernight at 428C and subsequently washed two times with 26 SSC, 0.1% SDSsolution at room temperature, followed by washes with 0.56 SSC, 0.1% SDS at 688C.Membranes were kept in washing buffer (11.6 g�l71 maleic acid, 8.8 g�l71 NaCl,3 ml�l71 Tween 20, pH 7.5) until detection. Positive colonies were detected by
445SPECIFIC ENUMERATION OF ENTEROCOCCUS FAECIUM

chemoluminescence using the Digoxigenin System (DIG Luminescent Detection Kit,Roche, Mannheim, Germany) with CSDP as substrate and a sensitive charge coupleddensity camera (SensiCam QE 12bit, PCO Computer Optics, software version Camware1.2). An example of the colony hybridization is shown in Figure 1.
3. RESULTS
3.1. Probe specificity
The specificity of the 1.2 kb PCR generated E. faecium NCIMB 10415 probe wasevaluated against a range of reference strains. OnlyE. faeciumNCIMB 10415 gave a PCRproduct of the expected size except one reference strain (DSMZ 2918), which produced aweak band of similar size after PCR amplification (Table I). The origin of this PCRartefact was not further studied, as the respective strain and all other tested referencestrains did not produce positive signals after colony hybridization. Similarly, colonyhybridization of intestinal or faecal samples from control animals of the feeding trialnever showed positive signals. Thus, it can be concluded that the employed primer and therespective PCR generated probe are specific for the E. faecium NCIMB 10415 plasmid.
3.2. Feed
Feed content of added E. faecium NCIMB 10415 was monitored continuouslythroughout the feeding trial. High total enterococci colony counts were not expected or
TABLE I Reference strains and PCR results with E. faecium NCIMB 10415 specific primers
Species Strain1 Remarks2 PCR Colony hybridization
Enterococcus faecium NCIMB 10415 + +Enterococcus faecium DSMZ 6177 7 7Enterococcus faecium DSMZ 20477 7 7Enterococcus faecium DSMZ 2146 =ATCC 6057 7 7Enterococcus faecium DSMZ 2918 7/+ 7Enterococcus faecium NCIMB 700583 7 7Enterococcus faecium NCIMB 8842 =ATCC6057 7 7Enterococcus faecium NCIMB 2702 7 7Enterococcus faecium NCIMB 702822 7 7Enterococcus faecium NCIMB 702711 7 7Enterococcus faecium NCIMB 12494 =ATCC19635 7 7Enterococcus faecium NCIMB 12412 =ATCC19634 7 7Enterococcus faecium NCIMB 11181 7 7Enterococcus faecalis DSMZ 6134 7 7Enterococcus faecalis DSMZ 8629 =ATCC 19433, NCIMB775 7 7Enterococcus faecalis DSMZ 20478 7 7Enterococcus faecalis DSMZ 20376 7 7Enterococcus faecalis DSMZ 13519 7 7Enterococcus faecalis DSMZ 20380 7 7Enterococcus hirae DSMZ 20160 =ATCC 8043, , NCIMB 6459 7 7Enterococcus durans DSMZ 20633 =ATCC 19432 7 7Enterococcus cecorum DSMZ 20682 =ATCC 43198 7 7Enterococcus villorum DSMZ 15688 =ATCC700913, NCIMB13634 7 7
1DSMZ: German Collection of Microorganisms and Cell Cultures, 38124 Braunschweig, Germany; NCIMB: NationalCollections of Industrial and Marine Bacteria Ltd., Aberdeen, Aberdeen, Scotland, UK.2ATCC: American Type Culture Collection, Manassas, Virginia, USA.
446 M. MACHA et al.

evidenced in feed and thus hybridization was carried out on random samples ofprobiotic supplemented feed, which all reacted positive and control feed samples, whichall reacted negative (data not shown).
3.3. Faeces
Faeces of sows fed the supplemented diet showed a nearly constant probiotic colonycount with an average of 1.8�105 cfu/g wet weight 10 days before as well as 14 days afterbirth. In contrast to sow faeces, E. faeciumNCIMB 10415 colony counts in piglet faeceswas only in the range of 103 – 105 cfu/g wet weight (Figure 2). Surprisingly, colonycounts were higher in suckling piglets before receiving creep feed (0 – 14 days) than inweaned piglets with a constant supply of probiotic supplemented feed.
3.4. Intestine
Although trial groups consisted of five animals per sampling day the number of sampleswas sometimes lower due to missing intestinal content, which occurred in the upperjejunum and ileum, or either excessive or no growth on the employed selective agar.The probiotic strain could be detected in the ileum and colon of three of four 14-day
old suckling piglets (Figure 3). Detection in the upper alimentary tract (i.e. stomach andupper jejunum) was only possible in one of four animals. Colony counts varied between102 and 107 cfu/g wet weight. One week after weaning (35 days of age), the strain wasonly detectable in the stomach of three of five animals, the upper jejunum was onlypositive for one sample. On the 28th and the 56th day of life, the strain could bedetected throughout the intestine in various concentrations and frequencies.The ratio of E. faecium NCIMB 10415 to total enterococci in the intestinal tract is
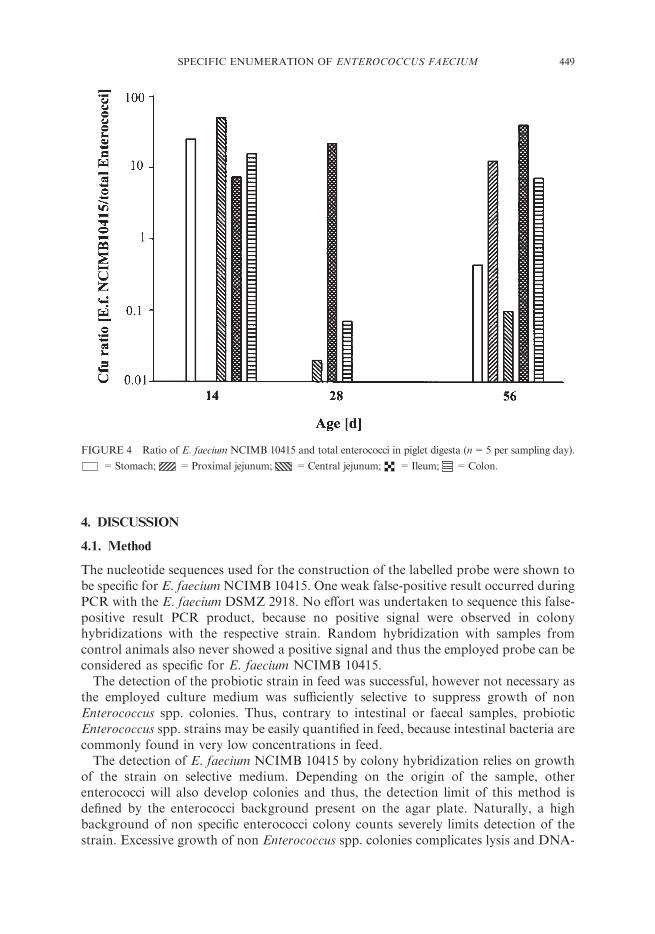
shown in Figure 4. Day 35 was omitted, because only three of a total of 25 samples werepositive for the probiotic strain. The ratio varied between 0.03 to 50% from stomach tocolon. Ileal samples always showed constantly high ratios on all sampling days.
FIGURE 1 Example of a colony hybridization. A=Agar plate with Enterococcus spp. colonies; B=Nylonmembrane after hybridization with the E. faecium NCIMB10415 specific probe. ~=E. faecium NCIMB10415colony.
447SPECIFIC ENUMERATION OF ENTEROCOCCUS FAECIUM
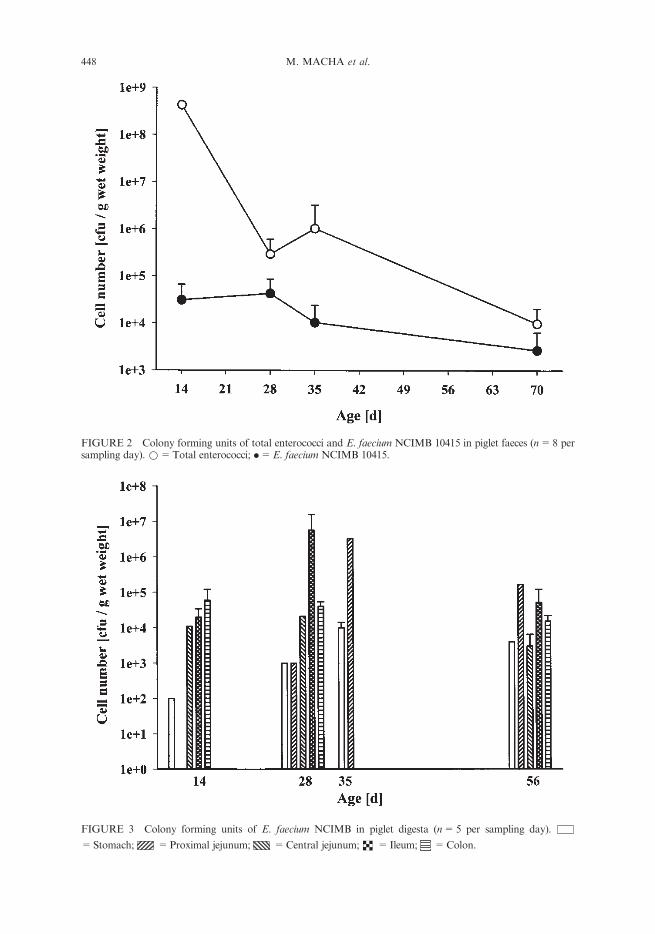
FIGURE 2 Colony forming units of total enterococci and E. faecium NCIMB 10415 in piglet faeces (n=8 persampling day). *=Total enterococci; .=E. faecium NCIMB 10415.
FIGURE 3 Colony forming units of E. faecium NCIMB in piglet digesta (n=5 per sampling day).
=Stomach; =Proximal jejunum; =Central jejunum; =Ileum; =Colon.
448 M. MACHA et al.
4. DISCUSSION
4.1. Method
The nucleotide sequences used for the construction of the labelled probe were shown tobe specific for E. faeciumNCIMB 10415. One weak false-positive result occurred duringPCR with the E. faecium DSMZ 2918. No effort was undertaken to sequence this false-positive result PCR product, because no positive signal were observed in colonyhybridizations with the respective strain. Random hybridization with samples fromcontrol animals also never showed a positive signal and thus the employed probe can beconsidered as specific for E. faecium NCIMB 10415.The detection of the probiotic strain in feed was successful, however not necessary as
the employed culture medium was sufficiently selective to suppress growth of nonEnterococcus spp. colonies. Thus, contrary to intestinal or faecal samples, probioticEnterococcus spp. strains may be easily quantified in feed, because intestinal bacteria arecommonly found in very low concentrations in feed.The detection of E. faecium NCIMB 10415 by colony hybridization relies on growth
of the strain on selective medium. Depending on the origin of the sample, otherenterococci will also develop colonies and thus, the detection limit of this method isdefined by the enterococci background present on the agar plate. Naturally, a highbackground of non specific enterococci colony counts severely limits detection of thestrain. Excessive growth of non Enterococcus spp. colonies complicates lysis and DNA-
FIGURE 4 Ratio of E. faecium NCIMB 10415 and total enterococci in piglet digesta (n=5 per sampling day).
=Stomach; =Proximal jejunum; =Central jejunum; =Ileum; =Colon.
449SPECIFIC ENUMERATION OF ENTEROCOCCUS FAECIUM

fixation procedure due to very high amounts of cell debris and non specific DNA. Fromour own experience, a maximum of 100 colonies per selective plate can be used forcolony hybridization. Thus, the detection of a single Enterococcus spp. colony by colonyhybridization is limited to 1% of total enterococci.Colony appearance of indigenous enterococci differed largely between agar plates
from faecal and intestinal samples. Faecal samples displayed typical growth ofenterococcal colonies with only minor background of non Enterococcus spp. colonies,but growth inhibition was poor for not further characterized presumptive Lactobacillusspp. (Vitek-Analyser results) especially in small intestinal samples. Furthermore, uptakeof the discriminating tetrazolium dye into Enterococcus spp. cells was not alwayscomplete and hindered visual evaluation of colony counts on some plates.With regard to total enterococci count, it must be taken into account that enterococci
as well as lactobacilli are able to produce bacteriocin-like substances which often inhibitclosely related species (Park et al., 2003). Therefore, results could be biased becausegrowth of some strains may be suppressed by background populations present on theagar plate.
4.2. Detection and viability
This study confirms again, as shown in earlier studies with a bacillus probiotic (Jadamuset al., 2001), that bacteria are transferred from sow to suckling piglet by contact tomaternal faeces before starter feed is available. However, as in the study with thebacillus probiotic, colony counts in the stomach and the small intestine of sucklingpiglets were low. Only two of five 14-day old suckling piglets yielded specific probioticcfu in stomach samples and detection was not possible in any sample from the proximaljejunum.Although colony counts of the probiotic Enterococcus strain in the stomach were very
low (50 – 120 cfu/g), a drastic increase occurred to about 5�104 cfu/g in colon and faecalsamples. Even if methodological bias on specific cfu like high background growth instomach samples or higher dry matter in terminal sections are taken into consideration,these results can be viewed as a true colonization of the probiotic strain in the ileum andthe hind gut of suckling piglets.Weaning imposes a remarkable stress upon the animal. Diet changes lead to deep
modifications of the intestinal epithelium and consequently, intestinal bacterialpopulations adapt to new substrate conditions, i.e. new bacterial communities andphysiological functions develop. The established succession of intestinal bacterialcommunities changes/collapses during the first days after weaning (Savage 1977;Simpson et al., 1999; Hooper et al., 2002) and as a direct result many piglets suffer fromintestinal bacterial or viral infections (Nabuurs, 1998). In this study the probioticappeared not to be sufficiently competitive to prolong its colonization ability of thesuckling period, because 1 week after weaning the probiotic could only be detected instomach and proximal jejunum samples. No detection was possible in ileum or hindgutsamples, but faecal contents again showed specific probiotic cfu. This is the reversepicture of the suckling period, in which only ileum and hind gut samples showedsignificant probiotic colonization. The relatively high amount of probiotic cfu instomach samples doubtlessly originated from feed intake, whereas faecal cfu probablyoriginated from probiotic cells that only passed the intestine.
450 M. MACHA et al.

The continous intake of the probiotic for three weeks however showed a comparativelyeven colonization in intestinal and faecal samples of 103 – 104 probiotic cfu/g wet weight.Compared to stomach samples, only a 10 – 15-fold increase in cfu was observed in ileumand hind gut samples. Preweaning piglets displayed 100- or more fold cfu increase fromstomach to colon/faecal samples. These results indicate that the colonization of theprobiotic strain was not as successful after weaning. It may be speculated that probioticcells entering the suckling piglet via maternal faeces were already adapted to the sowsintestine and its bacterial inhabitants. Thus, transfer of the probiotic E. faecium strain asa part of an functioning microbiota into an uncolonized habitat may enhance theprobiotic colonization, as cells of this strain are already competitive against existingbacteria. Finally, the results regarding the ratio of probiotic to total enterococci cfuindicate that regardless of initial amount, with some exceptions most intestinal samplesdisplayed a ratio of 5 – 25% probiotic cfu to total enterococci. These numbers have to beviewed as low within the group of enterococci, they are insignificant regarding totalbacterial cfu. From an ecological point of view it is very acceptable that a single bacterialstrain will not be able to dominate a diverse habitat like the intestine. The theory ofclimax communities predicts a maximal colonization limit for each member of anecosystem (Savage, 1977) and our results may support this view.This study also allows the deduction of some important implications for the modes of
action of Enterococcus spp. probiotics. First, excess amounts of probiotic cells are notable to enhance the colonization of the strain, because maximum colonization capacityis defined by the intestinal environment and its bacterial populations (Savage, 1977).Second, absolute probiotic cell numbers are insignificant compared to the total amountof bacterial cells present in the gastrointestinal tract. Therefore, a probiotic effect viaproduction of metabolites (i.e. lactate) is very unlikely. Furthermore, specific traits likeenterocins, host immune response activating substances or other modulating substancesmust be very potent in order to induce the observed changes in bacterial composition.Thirdly, a long term priming effect of the probiotic on bacterial populations of themother sow is proposed, whereby the contact with already modified bacterialpopulations enhances probiotic colonization in the suckling piglet.
5. CONCLUSIONS
The application of the colony hybridization method enables for the first time theselective detection of the widely used probiotic Enterococcus faecium NCIMB 10415strain among total Enterococcus spp. counts of digesta, faeces and feed. It is nowpossible to monitor the presence of the probiotic in the intestinal tract and faeces.Results of this study have implications for the proposed modes of action of probiotics inanimal nutrition.
Acknowledgement
This study was funded by the Deutsche Forschungsgemeinschaft (DFG-FOGR 438-1).
451SPECIFIC ENUMERATION OF ENTEROCOCCUS FAECIUM

References
Anderson, K.L. (2003) The complex world of gastrointestinal bacteria. Can. J. Anim. Sci., 83, 409 – 427.Betzl, D., Ludwig, W. and Schleifer, K.H. (1990) Identification of lactococci and enterococci by colony
hybridization with 23S rRNA-targeted oligonucleotide probes. Appl. Environ. Microbiol., 56, 2927 – 2929.Blum-Oehler, G., Oswald, S., Eiteljorge, K., Sonnenborn, U., Schulze, J., Kruis, W. and Hacker, J. (2003)
Development of strain-specific PCR reactions for the detection of the probiotic Escherichia coli strain Nissle1917 in fecal samples. Res. Microbiol., 154, 59– 66.
Brigidi, P., Swennen, E., Vitali, B., Rossi, M. andMatteuzzi, D. (2003) PCR detection of Bifidobacterium strainsand Streptococcus thermophilus in feces of human subjects after oral bacteriotherapy and yogurt consumption.Int. J. Food Microbiol., 81, 203 – 209.
Buluwela, L., Forster, A., Boehm, T. and Rabbitts, T.H. (1989) A rapid procedure for colony screening usingnylon filters. Nucleic Acids Res., 17, 452.
Datta, A.R., Wentz, B.A. and Hill, W.E. (1987) Detection of hemolytic listeria monocytogenes by using DNAcolony hybridization. Appl. Environ. Microbiol., 53, 2256 – 2259.
Duez, H., Pelletier, C., Cools, S., Aissi, E., Cayuela, C., Gavini, F., Bouquelet, S., Neut, C. and Mengaud, J.(2000) A colony immunoblotting method for quantitative detection of a Bifidobacterium animalis probioticstrain in human faeces. J. Appl. Microbiol., 88, 1019 – 1027.
Durant, J.A., Young, C.R., Nisbet, D.J., Stanker, L.H. and Ricke, S.C. (1997) Detection and quantification ofpoultry probiotic bacteria in mixed culture using monoclonal antibodies in an enzyme-linked immunosorbentassay. Int. J. Food Microbiol., 38, 181 – 189.
Festl, H., Ludwig, W. and Schleifer, K.H. (1986) DNA hybridization probe for the Pseudomonas fluorescensgroup. Appl. Environ. Microbiol., 52, 1190 – 1194.
Fuller, R. (1989) Probiotics in man and animals. J. Appl. Bacteriol., 66, 365– 378.Goverde, R.L., Jansen, W.H., Brunings, H.A., Huis in ‘t Veld, J.H. and Mooi, F.R. (1993) Digoxigenin-labelled
inv- and ail-probes for the detection and identification of pathogenic Yersinia enterocolitica in clinicalspecimens and naturally contaminated pig samples. J. Appl. Bacteriol., 74, 301– 313.
Holtke, H.J., Ankenbauer, W., Muhlegger, K., Rein, R., Sagner, G., Seibl, R. and Walter, T. (1995) Thedigoxigenin (DIG) system for non-radioactive labelling and detection of nucleic acids – an overview. Cell. Mol.Biol. (Noisy-le-grand), 41, 883 – 905.
Hooper, L.V., Midtvedt, T. and Gordon, J.I. (2002) How host-microbial interactions shape the nutrientenvironment of the mammalian intestine. Annu. Rev. Nutr., 22, 283 – 307.
Jadamus, A., Vahjen, W., Schafer, K. and Simon, O. (2002) Influence of the probiotic strain Bacillus cereus var.toyoi on the development of enterobacterial growth and on selected parameters of bacterial metabolism indigesta samples of piglets. J. Anim. Physiol. Anim. Nutr., 86, 42 – 54.
Jadamus, A., Vahjen, W. and Simon, O. (2001) Growth behaviour of a spore forming probiotic strain in thegastrointestinal tract of broiler chicken and piglets. Arch. Anim. Nutr., 54, 1 – 17.
Lynch, P.A., Gilpin, B.J., Sinton, L.W. and Savill, M.G. (2002) The detection of Bifidobacterium adolescentis bycolony hybridization as an indicator of human faecal pollution. J. Appl. Microbiol., 92, 526– 533.
Marteau, P.R., de Vrese, M., Cellier, C.J. and Schrezenmeir, J. (2001) Protection from gastrointestinal diseaseswith the use of probiotics. Am. J. Clin. Nutr., 73, 430S– 436S.
Nabuurs, M.J. (1998) Weaning piglets as a model for studying pathophysiology of diarrhea.Vet. Q., 20(Suppl 3),S42 –S45.
Numata, N., Ushimizu, M., Ohtomo, M., Chida, K., Fujita, H., Saito, T., Suto, D., Oguro, M., Hayakawa, Y.,Sasaki, K., Arakawa, E., Shimada, T. andWatanabe, H. (2000) The use of colony hybridization in the isolationof thermostable direct hemolysin-producing Vibrio parahaemolyticus from foods implicated in an incidence offood poisoning. Jpn. J. Infect. Dis., 53, 75– 77.
Park, S.H., Itoh, K. and Fujisawa, T. (2003) Characteristics and identification of enterocins produced byEnterococcus faecium JCM 5804T. J. Appl. Microbiol., 95, 294 – 300.
Radhika, B., Padmapriya, B.P., Chandrashekar, A., Keshava, N. and Varadaraj, M.C. (2002) Detection ofBacillus cereus in foods by colony hybridization using PCR-generated probe and characterization of isolatesfor toxins by PCR. Int. J. Food Microbiol., 74, 131 – 138.
Reuter, G. (2001) Probiotics- possibilities and limitations of their application in food, animal feed, and inpharmaceutical preparations for men and animals. Berl. Munch. Tierarztl. Wochenschr., 114, 410– 419.
Saarela, M., Mogensen, G., Fonden, R., Matto, J. and Mattila-Sandholm, T. (2000) Probiotic bacteria: safety,functional and technological properties. J. Biotechnol., 84, 197 – 215.
Savage, D.C. (1977) Microbial ecology of the gastrointestinal tract. Annu. Rev. Microbiol., 31, 107 – 133.Simpson, J.M., McCracken, V.J., White, B.A., Gaskins, H.R. andMackie, R.I. (1999) Application of denaturant
gradient gel electrophoresis for the analysis of the porcine gastrointestinal microbiota. J. Microbiol. Methods,36, 167– 179.
Tannock, G.W. (2002) Probiotics and Prebiotics – Where are we going? In: Tannock, G.W. (Ed) Probiotics andPrebiotics – Where are we going? Caister Academic Press, Wymondham, UK, pp. 1 – 39.
Vahjen, W., Jadamus, A. and Simon, O. (2002) Influence of a probiotic Enterococcus faecium strain on selectedbacterial groups in the small intestine of growing turkey poults. Arch. Anim. Nutr., 56, 419– 429.
452 M. MACHA et al.
















