
Dimensions of maternal behaviour characteristics in domestic and wild×domestic crossbred sows
16
Dimensions of maternal behaviour characteristics in domestic and wilddomestic crossbred sows Marek S ˇ pinka a,* , Gudrun Illmann a , Francien de Jonge b , Maria Andersson c , Teun Schuurman d , Per Jensen c a Group of Ethology, Research Institute of Animal Production, CZ-104 01 Prague-Uhr ˇı ´ne ˇves, Czech Republic b Department of Ecological Agriculture and Society, Wageningen University and Research Centre, Haarweg 333, 6709 RZ Wageningen, Netherlands c Department of Animal Environment and Health, Faculty of Veterinary Medicine, Section of Ethology, Swedish University of Agricultural Sciences, POB 234, S-532 23 Skara, Sweden d Human and Animal Physiology Group, Wageningen Institute of Animal Sciences, Wageningen University, Haarweg 10, 6709 PJ Wageningen, Netherlands Accepted 29 May 2000 Abstract We examined the maternal behaviour of seven domestic and seven wilddomestic primiparous sows during 10 days post partum to investigate two questions: (1) Did maternal behaviour change during domestication? (2) Can the interindividual variability of maternal behaviour be subsumed into a few dimensions of maternal temperament? We recorded: (a) willingness to leave the nest for food on Day 2; (b) reaction to a playback of squeezed piglet distress vocalisation on Day 2; (c) spontaneous nursing behaviour and spontaneous lying-down behaviour on Day 5 (from an overnight video recording); (d) reactions to playbacks of various piglet distress vocalisations on Day 6 and (e) reactions to a human in the ‘nest’ with piglets on Day 9. Moreover, data on baseline cortisol saliva concentration and its increase during a brief transportation period and novel environment challenge at the age of 5 months were available. Crossbred sows did not differ from domestic ones in any aspect of maternal behaviour except for a higher tendency to terminate final massage during nursings and a higher frequency of changing posture from lying to standing and back during the night. Factor analysis (based on correlation matrix of 11 behaviour and cortisol variables calculated for all 14 sows after removing the effect of breed) indicated that 82% of the variability in the data could be explained by three factors: first, ‘calmness’ on which low night time frequency of major posture changes, carefulness of lying-down behaviour and high propensity to remain in nursing position after milk ejection loaded positively while cortisol concentrations during challenge loaded negatively; second, ‘protectiveness’ with high loadings of the reaction scores to the playbacks of piglet distress calls and the human presence near the piglets; and third, ‘nursing activity’ which was Applied Animal Behaviour Science 70 (2000) 99–114 * Corresponding author. Tel.: 420-2-6771-0713; fax: 420-2-6771-0779. E-mail address: [email protected] (M. S ˇ pinka). 0168-1591/00/$ – see front matter # 2000 Elsevier Science B.V. All rights reserved. PII:S0168-1591(00)00151-9
Transcript of Dimensions of maternal behaviour characteristics in domestic and wild×domestic crossbred sows

Dimensions of maternal behaviour characteristics indomestic and wild�domestic crossbred sows
Marek SÏpinkaa,*, Gudrun Illmanna, Francien de Jongeb,Maria Anderssonc, Teun Schuurmand, Per Jensenc
aGroup of Ethology, Research Institute of Animal Production, CZ-104 01 Prague-UhrÏõÂneÏves, Czech RepublicbDepartment of Ecological Agriculture and Society, Wageningen University and Research Centre,
Haarweg 333, 6709 RZ Wageningen, NetherlandscDepartment of Animal Environment and Health, Faculty of Veterinary Medicine, Section of Ethology,
Swedish University of Agricultural Sciences, POB 234, S-532 23 Skara, SwedendHuman and Animal Physiology Group, Wageningen Institute of Animal Sciences, Wageningen University,
Haarweg 10, 6709 PJ Wageningen, Netherlands
Accepted 29 May 2000
Abstract
We examined the maternal behaviour of seven domestic and seven wild�domestic primiparous
sows during 10 days post partum to investigate two questions: (1) Did maternal behaviour change
during domestication? (2) Can the interindividual variability of maternal behaviour be subsumed
into a few dimensions of maternal temperament? We recorded: (a) willingness to leave the nest for
food on Day 2; (b) reaction to a playback of squeezed piglet distress vocalisation on Day 2; (c)
spontaneous nursing behaviour and spontaneous lying-down behaviour on Day 5 (from an overnight
video recording); (d) reactions to playbacks of various piglet distress vocalisations on Day 6 and (e)
reactions to a human in the `nest' with piglets on Day 9. Moreover, data on baseline cortisol saliva
concentration and its increase during a brief transportation period and novel environment challenge
at the age of 5 months were available. Crossbred sows did not differ from domestic ones in any
aspect of maternal behaviour except for a higher tendency to terminate ®nal massage during
nursings and a higher frequency of changing posture from lying to standing and back during the
night. Factor analysis (based on correlation matrix of 11 behaviour and cortisol variables calculated
for all 14 sows after removing the effect of breed) indicated that 82% of the variability in the data
could be explained by three factors: ®rst, `calmness' on which low night time frequency of major
posture changes, carefulness of lying-down behaviour and high propensity to remain in nursing
position after milk ejection loaded positively while cortisol concentrations during challenge loaded
negatively; second, `protectiveness' with high loadings of the reaction scores to the playbacks of
piglet distress calls and the human presence near the piglets; and third, `nursing activity' which was
Applied Animal Behaviour Science 70 (2000) 99±114
* Corresponding author. Tel.: �420-2-6771-0713; fax: �420-2-6771-0779.
E-mail address: [email protected] (M. SÏpinka).
0168-1591/00/$ ± see front matter # 2000 Elsevier Science B.V. All rights reserved.
PII: S 0 1 6 8 - 1 5 9 1 ( 0 0 ) 0 0 1 5 1 - 9

strongly positively associated with nursing frequency, and negatively with the proportion of
nutritive nursings and baseline cortisol values. The results indicate that most aspects of pig maternal
behaviour have not been signi®cantly changed by domestication and that substantial variability in
maternal behaviour exists between sows, perhaps in the form of several behaviour characteristics
which encompass both behaviour and endocrine pro®les of the sows. # 2000 Elsevier Science B.V.
All rights reserved.
Keywords: Maternal behaviour; Pigs; Domestication; Nursing; Cortisol; Vocalisation; Temperament
1. Introduction
Maternal behaviour is important for pig husbandry since it partly determines mortality
and growth of the piglets. About 15% of the live born piglets die before weaning (McKay,
1993; Blackshaw et al., 1994), and among them 70±80% is attributable to the sow
behaviour, directly or indirectly. During the ®rst day of life, piglets get trapped under
the sow while she is lying-down or rolling from her belly to her side (Dyck and Swiestra,
1987; Weary et al., 1996a; Herskin et al., 1998). Individual dams differ in the frequency of
postural changes, and in how often they change posture in a way that is dangerous for the
piglets (i.e. ¯opping straight down to one side versus descending to lying posture in a
vertical plane; lying down when piglets are nearby). Even after a piglet is trapped, the sow's
behaviour is important. Just 20% of piglets trapped under the sow's body are actually
crushed. If a trapped piglet starts screaming, the sow may stand up immediately and release
it. In cases when the sow stood up within 1 min, only 5% of piglets died; in contrast, 67%
of the piglets trapped for longer than 4 min were lost (Weary et al., 1996a). Two aspects of
the sow's behaviour, thus, affect the probability of crushing: ®rst, the sow's general
restlessness and her lying-down behaviour; second, the sow's individual reactivity to a
screaming piglet (Wechsler and Hegglin, 1997).
Another problem in piglet production is low milk intake in some piglets or in whole
litters which results in low or too variable weaning weights. This may even lead to piglet
losses, either directly due to starvation or indirectly due to the weak piglets being crushed
by the sow (Hutson et al., 1993; Weary et al., 1996b). Low frequency of nursings (SÏpinka
et al., 1997; SÏpinka et al., 1999), and a high proportion of non-nutritive nursings (nursings
without milk ejection, CastreÂn et al., 1989; Illmann and Madlafousek, 1995; Illmann et al.,
1999) are probably the aspects of nursing behaviour which can most signi®cantly reduce
milk intake. A tendency to terminate the udder massage by the piglets soon after milk
ejection might also suppress future milk production in the glands, although the evidence is
equivocal (Algers and Jensen, 1991; SÏpinka and Algers, 1995; Jensen et al., 1998; Illmann
et al., 1998).
These undesired aspects in the maternal behaviour of domestic sows may have arisen
during the process of domestication, especially during the intense selective breeding
regimes prevalent in last few decades. Information about whether this was the case can be
useful in judging the likelihood that maternal characteristics of sows could be improved
through selective breeding programmes. The present study was conducted in order to
assess whether sows carrying different proportions of genes from domestic and wild boar
100 M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114

stock vary in biologically signi®cant aspects of maternal behaviour. In order to answer this
question, we collected data on maternal behaviour in seven domestic and seven crossbred
wild�domestic primiparous sows. During the data analysis, we found that the breeding
effect could explain only a small proportion of the variation in the data. Therefore,
we investigated the interindividual variation through a post-hoc correlational and factor
analysis. We focused on questions of how various aspects of maternal behaviour
relate to each other and whether they could be subsumed into a few `maternal behaviour
traits'.
2. Methods
2.1. Animals
We used 14 primiparous sows which were born to Yorkshire�Dutch Landrace mothers.
Seven of them were sired by various Yorkshire fathers (`domestic' sows) and seven by one
male wild boar (`crossbred' sows). The sows were born and reared in the outdoor housing
system described below. At the age of 20±21 months, all experimental sows were
inseminated with Great Yorkshire sperm. Hence, the offspring interacting with the
experimental mothers in the current study were either 100% domestic or 25% wild boar.
2.2. Housing conditions
The experiment was carried out at the Wageningen Agricultural University, The
Netherlands in April±June 1997. All sows were loose housed during pregnancy. Two
days before the scheduled farrowing induction, the sows were locked in half-covered
concrete-¯oored rectangular outdoor pens (2.2 m�4.5 m). About 5 kg of straw was
provided in the covered half of the farrowing pen, the open half was provided with
drinking nipples for both the sows and the piglets and also served as dunging area. At the
back of the farrowing pens, half-open doors allowed behavioural observations, without
disturbances to sow or litter. At 8 a.m. and 1:30 p.m., sows were allowed to leave the pens in
order to feed in separate feeding crates positioned about 20 m from the pens. Sows which
were to be tested in the same batch (see below) were not housed in adjacent pens in order to
reduce the possibility of habituation to acoustic stimuli in the playbacks tests.
2.3. Procedure
2.3.1. Farrowing and cross-fostering
In order to enable early and balanced cross-fostering between litters, farrowings were
induced on about Day 112 of pregnancy by a prostaglandin analogue (PGF2a) injection.
The induction was scheduled in four batches of four, four, four and two sows. All sows in a
batch farrowed within a time period of 24 h (Day 0). Within 36 h after parturition
(Day 0 or 1), all piglets were weighed, ear tagged and cross-fostered within each batch.
The newly composed litters were balanced with respect to litter size, number of piglets
of each genotype (pure domestic or 25% wild boar), average piglet weight and sex
M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114 101

ratio (in that order of priority). The resulting litter sizes varied between seven and
nine piglets among which two±four piglets were the sow's own progeny while the rest
originated from the other litters in the batch. Mortality of the piglets was assessed by
noting any losses occurring between cross-fostering and video recording 4 days later (see
below).
2.3.2. Nest-leaving and playback experiment on Day 2
On Day 2 after parturition (Day 2), each sow was tested for her willingness to leave the
nest and the young for food and also for her reactivity to squeezed piglet distress
vocalisation. At the time of the morning feeding, the non-tested sows were ®rst allowed
to feed in a normal way. After they were locked back again in their farrowing pens, the pen
of the ®rst sow to be tested was opened and the latency of her emergence from the
farrowing pen was noted. If she did not leave the pen within 5 min, the gate was closed and
latency was set equal to 300 s. We then waited for the moment when the sow was lying
down in the farrowing pen for the ®rst time. When her body touched the ground, a 30 s
recording of piglet distress vocalisation was played (at about natural volume) from a
loudspeaker through an expanded-metal-covered opening in the outer wall of the farrowing
pen. The recording was the vocalisation of a 2-days-old piglet, unrelated to any animal in
the experiment. During the recording, vocalisations were stimulated by restraining the
piglet on the ground by an experimenter using both hands. The reaction of the sow to the
playback was video recorded and later scored using the following scale: 0 Ð no reaction,
1 Ð head movement, 2 Ð body movement, 3 Ð sits, 4 Ð stands up, 5 Ð makes contact
with the expanded metal covering the loudspeaker, 6 Ð bites or paws at the expanded
metal. Two extra points were added if the sow stood up within 5 s since the beginning of the
playback or one point if she did so within 30 s. After the testing of the ®rst sow was
®nished, we continued with testing of the next sow until all sows from a batch (up to four)
were tested. The same vocal stimulus was used for all sows.
2.3.3. Undisturbed nursing behaviour
During the night between Days 4 and 5 (or between Days 5 and 6) post partum, each sow
was video recorded either between 18:00 and 24:00 or between 00:00 and 06:00 h while
there were no people present near the animal facilities. The videotapes were later analysed
to quantify the following parameters of the undisturbed nursing behaviour: intervals
between nursings, proportion non-nutritive nursings, duration of pre-ejection and post-
ejection udder massage, and proportion of nursings terminated by the sow. Non-nutritive
nursings (nursings without milk ejection) were identi®ed by the absence of synchronised
rapid sucking activity of the litter (Illmann and Madlafousek, 1995). A nursing was
classi®ed as terminated by the sow if she rolled over onto her belly or stood up while some
of the piglets were still active at the udder.
2.3.4. Undisturbed lying-down behaviour
The videotaped six night time hours of undisturbed behaviour were also used to assess
the frequency and carefulness of the sows' lying-down behaviour. Each occurrence of the
sow's descent from a standing to a lying posture was counted and given a carefulness score
as follows: one point for snif®ng the ¯oor before lying-down, one point for rooting, one
102 M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114

point if the sow descended in a vertical plane (as opposed to ¯opping over to one side),
one point if there were, at that moment, no piglets near her on the side on which she lay
down.
2.3.5. Playback experiment on Day 5 or 6
These tests were conducted on Day 5 or 6 post partum either in the morning or in the
afternoon. The behaviour of the sow was videotaped in her pen while she was subjected to
three different tests of her reactivity to piglet distress vocalisations. The tests were:
Trapped piglet call ± A 35 s playback of a piglet's distress vocalisation which had been
recorded as described in the section on playback experiment on Day 2. However, different
recordings were used from those on Day 2. The playback was started while the sow was
descending to a lying posture at the moment her body touched the ground.
Isolated piglet call ± A 60 s playback of a piglet's grunting and squeaking which had been
recorded when a 7-day old piglet had been isolated from her mother and littermates. The
playback was started at any moment between 20 and 35 min after the last milk ejection
when both the sow and the litter was resting.
Teat fighting calls ± A 50 s playback of loud vocalisations recorded while two piglets had
been fighting for a teat during nursing. The playback was started during a nursing about
15 s after the pre-ejection udder massage by the piglets had begun.
Each of the three tests was repeated twice using vocalisations from different piglets to
those used for the playback stimuli. Hence, each sow was presented with a total of six
playback stimuli during the experimental session. The playbacks were always separated by
at least 15 min. The order of the tests was variable in individual sows as it was dependent on
the spontaneous nursing and resting behaviour of the animals. The behavioural reactions of
the sow were scored using the same scale as in the playback experiment on Day 2. A
preliminary analysis showed that the scores from the three types of tests were highly
intercorrelated and displayed similar results in the domestic-crossbred comparison. There-
fore, the score reactions from the six tests (reactions to trapped-piglet, isolated-piglet and
teat-®ghting calls, each tested twice) were combined into a ®nal score.
2.3.6. The human-in-the-nest test
On Day 8 or 9 post partum, the response of the sow to a human being present near her
piglets was tested. First, the non-tested sows were allowed to feed in a normal way. After
they were locked back again in their farrowing pens, the ®rst sow to be tested was allowed
to leave the pen, enter the feeding crate and was locked there for 10 min while she was
feeding. In the meanwhile, an experimenter entered the pen with her piglets, closed the bar
gate behind him/her and stood quietly about 1 m behind the bars of the pen gate. The sow
was released from the feeding pen and once she approached her pen, her behaviour was
scored. Every 20 s for 5 min, we recorded her location, using instantaneous sampling, as
follows: nose contact with the bar gate Ð 2 points, within 1 m in front of the gate Ð 1
point, other place Ð 0 points. We also scored the average frequency of her calls during each
20 s period, using the following scale: frequent vocalisation (approx. >1 call/s) Ð 2 points,
moderate vocalisation (<1 call/s) Ð 1 point, not vocalising Ð 0 points. Third, a additional
point was given to the sow within the current 20 s interval if she bit the bars of the gate or
M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114 103

jumped on it with her front legs. The average number of points for a 20 s interval was taken
as a measure of the sow's reaction to the human presence near her young.
2.4. Comparison with daytime nursing behaviour data
In order to see whether the individual characteristics of nursing behaviour were stable
over the diurnal period, we compared the nursing data with the data collected on the same
animals at 4±7 days post partum through direct daytime 4 h observations by Gustafsson
et al. (1999).
2.5. Cortisol data
In the post-hoc analysis (see Section 2.6), the set of behavioural scores was comple-
mented with data on cortisol baseline concentration and cortisol concentrations under a
complex challenge in the same sows. These data were collected about 15 months earlier,
when the animals were 5 months old. The gilts were housed in pairs in small pens and
trained to chew on cottonwool plugs for saliva collection. The ®rst saliva sample was
collected in the gilt's home pen just before the test started (t�0 min). At t�10 min, the gilt
was loaded to a carrier cage and, using a tractor, transported to another building. At the end
of transportation, 20 min after the ®rst sample, a second sample was taken and the gilt was
released to a 3 m�7 m pen with concrete ¯ooring. At 40 min, the third saliva sample was
taken and a gate was opened which connected this pen to a second pen provided richly with
straw. The fourth to sixth samples were collected at 55, 65 and 75 min after the ®rst one. All
tests were performed between 8:00 and 11:30 h in the morning in order to reduce cortisol
concentration differences caused by the circadian rhythm (Ekkel et al., 1996; Ruis et al.,
1997).
The saliva samples were immediately put on ice and centrifuged after the ®nal one was
collected. Plasma was stored atÿ208C until the assay. 0.5 ml of the thawn, undiluted saliva
sample was extracted with 2.5 ml dichloromethane. After evaporation of the solvent the
extract was subjected to a cortisol assay described by Janssens et al. (1994). The cortisol-
antibody used was a rabbit antiserum (K7348) raised against a cortisol-bovine-serum-
albumin-conjugate. Sensitivity of the assay was 0.5 ng/ml. The intra- and inter-assay
variances were 8 and 15%, respectively.
Two variables were extracted from the cortisol data. The ®rst was equalled to the pre-test
sample at time 0 and was labelled `cortisol baseline' concentration. The second was an
average of the second to sixth sample and was named `cortisol under challenge'
concentration.
2.6. Statistical analysis
2.6.1. Domestic�crossbred differences
Data on nest-leaving on Day 2 were treated as a binary variable (sow left the nest within
5 min or did not), and the domestic�crossbred comparison was made using the Fisher
exact probability test. In all other comparisons between the domestic and crossbred sows,
Mann±Whitney nonparametric U-tests were applied.
104 M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114

2.6.2. Night�daytime nursing behaviour comparison
In order to see whether the nursing behaviour was different over night compared to
daytime, and whether the sows kept their individual nursing style over the diurnal period,
we pooled the data from daytime direct observation (Gustafsson et al., 1999) and night
video recording. The data set was then subjected to an ANOVAwith breed, time (day versus
night), breed�time interaction, and sow nested within breed as the main effects. For the
breed effect, the between-pig MS was used as the error term.
2.6.3. Association of piglet mortality with maternal traits
The dependence of piglet mortality on maternal characteristics was assessed by
computing the Spearman rank correlation between the proportion of piglets dying from
Days 0 to 4 within a given `litter' (the group of piglets composed through cross fostering
and suckled by the given sow) and the scores of their foster mother in the tests and
observations. The breed effect was removed prior to computing the correlations.
2.6.4. Post-hoc correlational and factor analysis
Correlational post-hoc analysis was used to investigate whether the results from the three
tests, nursing and posture change behaviour scores from the night time video recording and
the cortisol data (11 variables in total) related to each other. A factor analysis was carried
out in an attempt to describe the complex results in terms of a few factors. In order to make
the results easier to interpret and present in a ®gure, we inverted some of the variables such
that the higher the level of the variable, the better (presumably) the survival and growth of
the piglets. Instead of proportion of nursings terminated by the sow, we took the proportion
of nursing not terminated by the sow and labelled it `massage allowance'; instead of
nursing interval, the nursing frequency (mean number of nursings per hour) was used; and
instead of proportion of non-nutritive nursings, the proportion of nutritive nursings was
used. The duration of pre-ejection udder massage was changed to `speed of milk ejection'
by converting the number of seconds into negative values. Similarly, the night time number
of standing-up acts was rendered negative and labelled `night time inactivity'. The effect of
breed (domestic versus crossbred) was removed from this modi®ed set of variables by a
GLM procedure with breed as the only main effect. Next, the residual values were used to
construct a matrix of product-moment correlations between the variables. Finally, the
matrix was subjected to factor analysis with the procedure Factor of the SAS software.
Squared multiple correlations were used for the prior communality estimates. Factors with
eigenvalues larger than 1 were retained and subjected to orthogonal varimax prerotation
followed by promax oblique rotation. This procedure was chosen in order to separate out
original variables on the resulting factors as well as possible, and, at the same time, show
whether the factors are correlated to each other or not.
3. Results
3.1. Crossbred�domestic differences
Crossbred sows were more likely to terminate the post-ejection massage during nursings
(57% versus 17% of cases, Mann±Whitney U-test, U�1, p<0.001) which resulted in
M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114 105

shorter post-ejection udder massage (268 versus 428 s, Mann±Whitney U-test, U�6,
p<0.05). Second, the crossbred sows tended to stand up, on average, three times as often
during the 6 h of night time video recording than the domestic ones (4.7 versus 1.7 times,
Mann±Whitney U-test, U�9, p<0.1). There were no other signi®cant differences between
the two breeds in any of the recorded aspects of maternal reactivity, posture changing
behaviour or nursing behaviour.
No difference was detected in the saliva unbound cortisol baseline concentrations at 5
months of age (Fig. 1: crossbred sows 2.2 ng/ml versus domestic sows 1.8 ng/ml, Mann±
Whitney U-test, U�15, NS). However, the `cortisol under challenge' concentration, i.e. the
average concentration in ®ve samples collected after the transport and during the new
environment challenge (Fig. 1) was more than twice higher in the crossbred sows (5.5
versus 2.7 ng/ml, Mann±Whitney U-test, U�4, p<0.01).
The mortality of piglets during the 4 days following cross-fostering was not different
between those suckled by the crossbred and domestic mothers (10% versus 13%, Mann±
Whitney U-test, U�20, NS; Table 1).
3.2. Comparison with daytime nursing behaviour data
Of the ®ve variables in the pooled day-and-night data set (duration of pre- and post-
ejection udder massage, nursing interval, proportion of non-nutritive nursings, and
proportion of nursings terminated by the sow), the ANOVA model with time, breed,
time�breed interaction and the sow identity as main effects was not signi®cant for the
nursing interval and the proportion of non-nutritive nursings. The proportion of nursings
terminated by the sow was lower (ANOVA, F1, 12�10.67, p<0.01) and post-ejection udder
massage was longer (ANOVA, F1, 12�13.49, p<0.01) in the night than during daytime
(Table 2). Crossbred sows terminated more nursings (ANOVA, F1, 12�86.37, p<0.0001),
and allowed shorter post-ejection massage (ANOVA, F1, 12�7.97, p<0.05). The interaction
between time of day and breed was not signi®cant for any variable. Sow identity in¯uenced
the proportion of sow-terminated nursings (ANOVA, F12, 12�4.55, p<0.01) and the
Fig. 1. Saliva cortisol concentrations of domestic (solid line) and crossbred (hatched line) sows at 5 months of
age during a challenge test. See text for details. Means�S.E. are depicted.
106 M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114

duration of pre-ejection massage (ANOVA, F12, 12�3.02, p<0.05) but not the nursing
interval, percentage of non-nutritive nursings or the post-ejection massaging duration.
3.3. Piglet losses and maternal traits
Piglet mortality within a given litter between Days 0 and 4 tended to be correlated
negatively with the number of lying-down acts during the 6 h night time recording
(rS�ÿ0.51, p<0.1). Mortality did not correlate signi®cantly with the carefulness of the
sow's lying-down behaviour (rS�ÿ0.41, p�0.15) or with any other of the recorded
maternal behaviours or reaction scores (all rS between ±0.30 and 0.30, p>0.20). Neither
was mortality related to any of the Factors 1±3 (rS�±0.23, 0.36, and ±0.07, respectively).
3.4. Post-hoc correlational and factor analysis of maternal traits
In the correlational matrix of the 11 quantitative behaviour and cortisol variables, there
were 11 signi®cant and three marginally signi®cant correlations, which formed three
interconnected groups of behaviours (Fig. 2). The factor analysis based on the correlation
matrix revealed that ®rst the three factors had eigenvalues of 3.19, 2.98 and 2.06,
explaining 32, 30 and 20% (82% in total) of the variation in the matrix. The fourth factor
had an eigenvalue of only 0.64 and hence, the ®rst three were extracted and subjected to
varimax orthogonal and then promax oblique rotation (Fig. 3). After promax rotation, the
Table 1
Maternal reactivity, posture changing behaviour and nursing behaviour in domestic and crossbred sowsa
Variable Breed U value Significance
Domestic Crossbred
Nest leaving on Day 2 (number of
sows with latency <300 s)
Three out
of seven
Four out
of seven
± NS
Reaction to trapped piglet vocalisations
on Day 2 (arbitrary score)
4.9�1.4 5.9�1.4 22 NS
Reaction to three types of piglet call
on Day 5 (arbitrary score)
27.6�7.2 31.9�7.6 19 NS
Reaction to a human in the piglet
nest on Day 8 (arbitrary score)
22.8�5.0 19.6�4.1 22 NS
Internursing interval (min) 39.7�2.5 35.4�1.6 15 NS
Non-nutritive nursings (%) 13.7�5.4 17.0�3.0 22 NS
Nursings terminated by the sow (%) 16.7�2.7 56.9�19.6 1 ***
Pre-ejection massage duration (s) 96�12 111�10 18 NS
Post-ejection massage duration (s) 428�53 268�32 6 **
Number of standing-ups during six night time hours 1.7�0.6 4.7�1.2 9 *
Carefulness of lying-down (arbitrary score) 3.5�0.3 3.1�0.1 12 NS
Piglet mortality (% of pigs dying between Days 0 and 4) 9.9�2.9 12.9�3.8 20 NS
a With the exception of the variable `nest leaving' on Day 2, means�S.E. are given. Significances as
resulting from the Mann±Whitney U-tests (Fisher exact probability test in the case of nest leaving): NS; P>0.1;
*P< 0.1; **P<0.05; ***P<0.001.
M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114 107
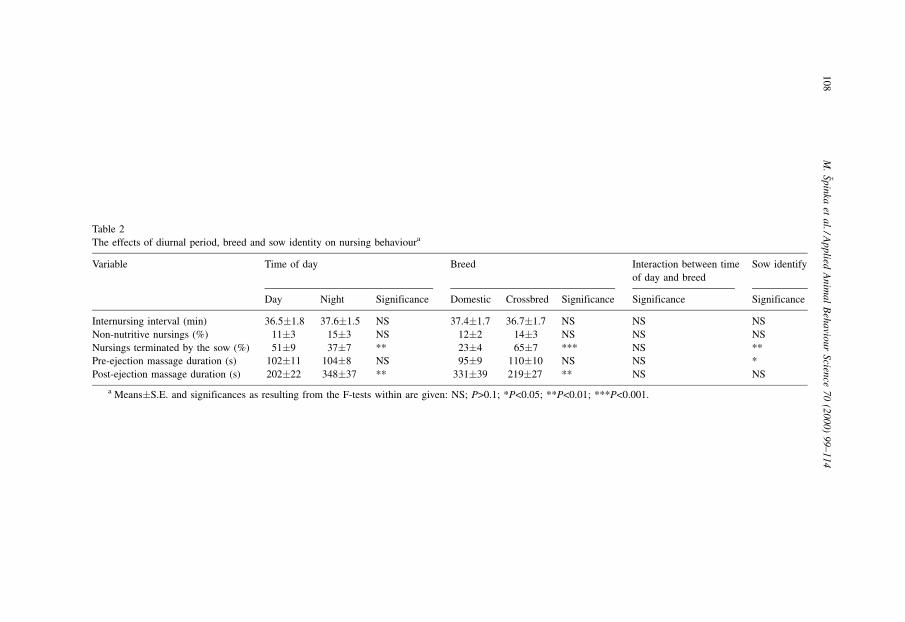
Table 2
The effects of diurnal period, breed and sow identity on nursing behavioura
Variable Time of day Breed Interaction between time
of day and breed
Sow identify
Day Night Significance Domestic Crossbred Significance Significance Significance
Internursing interval (min) 36.5�1.8 37.6�1.5 NS 37.4�1.7 36.7�1.7 NS NS NS
Non-nutritive nursings (%) 11�3 15�3 NS 12�2 14�3 NS NS NS
Nursings terminated by the sow (%) 51�9 37�7 ** 23�4 65�7 *** NS **
Pre-ejection massage duration (s) 102�11 104�8 NS 95�9 110�10 NS NS *
Post-ejection massage duration (s) 202�22 348�37 ** 331�39 219�27 ** NS NS
a Means�S.E. and significances as resulting from the F-tests within are given: NS; P>0.1; *P<0.05; **P<0.01; ***P<0.001.
10
8M
.S Ïp
inka
eta
l./Ap
plied
An
ima
lB
eha
viour
Scien
ce70
(2000)
99±114

Fig. 2. Clusters of correlated variables from the correlational matrix of 11 behavioural and cortisol variables.
Pearson correlation values are given. Dotted arrows: p<0.1; hatched arrows: p<0.05; solid arrows: p<0.01.
Variable labels: LY, carefulness of lying-down; AM, allow massage, i.e. proportion of nursings which the sows
did not terminate; NI, night inactivity, i.e. negatively taken number of body position changes during 6 h; CC,
cortisol concentration during challenge; RH, reaction to human near piglets; RT, reaction to trapped piglet
screaming on Day 2; RC, reaction of various distress calls by the piglets on Day 6; ES, ejection speed, i.e. the
briefness of pre-ejection massage; NF, nursing frequency; CB, cortisol baseline; %N, proportion of nutritive
nursings.
Fig. 3. Results of the factor analysis: loadings of the 11 behaviour and cortisol variables on the three factors
with eigenvalues >1. Variable labels: %N, proportion of nutritive nursings; AM, allow massage, i.e. proportion
of nursings which the sows did not terminate; CB, cortisol baseline; CC, cortisol concentration during challenge;
ES, ejection speed, i.e. the briefness of pre-ejection massage; LY, carefulness of lying-down; NF, nursing
frequency; NI, night inactivity, i.e. negatively taken number of body position changes during 6 h; RH, reaction
to human near piglets; RT, reaction to trapped piglet screaming on Day 2; RC, reaction of various distress calls
by the piglets on Day 6.
M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114 109

three factors remained almost orthogonal (uncorrelated) with the correlations between the
factors being at ÿ0.03, 0.01 and ÿ0.16. Three groups of behaviour variables were
suggested by the factor pattern. High propensity to allow post-ejection udder massage,
carefulness of lying-down behaviour and night time inactivity loaded highly on Factor 1.
Reaction scores in the three stimuli tests (to trapped piglet vocalisation on Day 2, to piglet
distress vocalisations on Day 5 or 6, and to human intrusion into the `nest' on Day 8 or 9)
and the speed of milk release loaded highly on Factor 2. Finally, nursing frequency was
strongly positively, and the proportion of nutritive nursings strongly negatively, associated
with Factor 3. Cortisol concentration under challenge loaded highly negatively on Factor 1,
and cortisol baseline negatively on Factor 3. Factor 2 was not associated with cortisol
concentrations at all.
4. Discussion
In this study, we documented striking similarities in maternal behaviour of domestic and
wild boar�domestic pigs. This study complements a previous paper by Gustafsson et al.
(1999) which was based on parallel observations on the same animals and which focused
on prefarrowing nest building behaviour and on daytime nursing and general proximity
behaviours. Since all 14 experimental sows were born to and raised by mothers of the same
breed and kept until testing in the same type of environment, we have controlled for other
than genetic factors. The studies were performed on the assumption that behavioural traits,
especially the quantitative ones like frequency and duration of behaviours, are in¯uenced
by many genes. For such traits, the behavioural phenotype of a hybrid should be about
halfway between that of the two parental genotypes (Price, 1998). A lack of difference
between one of the parental genotypes and the hybrid suggests that the other parent's
genotype is not differing grossly from the hybrid either. This is exactly what was found in
the two studies. The results thus might suggest that domestic sows retain the maternal
behaviour which has evolved under natural selection. This conclusion is in agreement with
previous observations of semi-naturally kept domestic sows (Jensen, 1986, 1988) and
female wild boar (Horrell, 1997). However, our conclusion must be regarded as pre-
liminary for two reasons. First, the sample size was small. Second, all seven crossbred sows
were sired by the same wild boar. The sows within this group were, thus, half-siblings
which may somewhat weaken the genetic comparison.
The present study and that of Gustafsson et al. (1999) found only two behavioural
differences between domestic and crossbred sows. First, the domestic sows had a lower
tendency to terminate nursings early after milk ejection. This may be interpreted as an
enhanced tendency in domestic mothers to invest more into the current litter as opposed to
saving resources for future reproduction (Jensen and Gustafsson, 1997; Gustafsson et al.,
1999). This interpretation is based on the assumption that longer post-ejection massage
increases milk production. Support for such a relationship was found in some studies
(Algers and Jensen, 1991; Jensen et al., 1998) but not in others (SÏpinka and Algers, 1995;
Illmann et al., 1998). An alternative explanation arises from the second behavioural
difference found in the two studies: the domestic sows were generally less active as
manifested in a higher proportion of lying during the day at 3 weeks postpartum
110 M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114

(Gustafsson et al., 1999) and in less frequent interruptions of the night time rest through
standing-up acts (this study). Lower activity saves energy and might, thus, be a result of the
intense selection for fast growth, ef®cient food conversion, heavier live weight and larger
body size (Robert et al., 1987). As a side effect, the less active domestic sows may remain
in the nursing position longer after milk ejection. A third interpretation is that higher agility
of crossbred sows is due to hybrid vigour, since, presumably, a higher heterozygosity was
achieved in the wild boar�Landrace hybrids than in the Yorkshire�Landrace crosses.
We found a close similarity in the duration of pre-ejection udder massage, nursing
interval, and the proportion of non-nutritive nursings observed by a human observer during
the day and the same variables scored from the night time video recordings. This
demonstrates, ®rst, that these aspects of nursing behaviour were not in¯uenced by daytime
human activity, and second, that the nursing effort was evenly distributed over the diurnal
period. Equal nursing frequency over day and night was also found in domestic sows by
Hartman et al. (1962). During the night, sows terminated fewer nursings and, consequently,
post-ejection udder massage lasted longer than during the day. This was probably
connected to the fact that sows rested in a recumbent posture for most of the night, as
reported previously by Bùe (1991, 1994).
In the ANOVA analysis of the pooled day-and-night nursing data, the sows' identity as a
factor signi®cantly in¯uenced the proportion of nursings terminated by the sow and the pre-
ejection massage duration (Table 2). This documents that these nursing characteristics
were retained in their nursing behaviour throughout the diurnal period. On the other hand,
the internursing interval and the proportion of non-nutritive nursings were not signi®cantly
affected by sow identity, showing that the frequency of nursing and the related frequency of
failures to release milk were not stable over the diurnal period.
Several previous studies found piglet mortality to be in¯uenced by sow reactivity to
piglet distress calls, and to carefulness and frequency of postural changes (McGlone and
Blecha, 1987; Hutson et al., 1993; Wechsler and Hegglin, 1997). In our study, the
frequency of postural changes was moderately related to piglet mortality, but carefulness
of lying-down and the reactivity to vocal stimuli was not. This failure to con®rm the
previously reported relationships may be due to the fact that we followed the mortality only
starting with the moment of cross-fostering, about 12±36 h after the birth of the piglets.
Since more than a half of the live-born piglet mortality occurs during the parturition and
within 24 h after birth (Weary et al., 1996a), we probably missed the period when the
maternal behaviour is most in¯uential on piglets' survival.
The post-hoc correlational analysis indicated three groups of interdependent variables
(Fig. 2). Subsequent factor analysis, although based on low ratio of replicates (14) to
original variables (11), succeeded in visualising this structure by suggesting three factors
which explained more than 80% of the variation and on which the variables separated out
well (Fig. 3). Factor 1 might be tentatively termed `calmness' as it was associated with the
sow remaining in a nursing position for a long time after milk ejection, with long periods of
uninterrupted resting with the piglets in the night, and with careful lying-down behaviours
which preclude trapping piglets under the sow's body. Factor 2 could be perhaps best called
`protectiveness' as it is connected with quick and intense reactions of the sows to stimuli
which signal danger to the piglets. The association of this factor with short duration of pre-
ejection massage might indicate that it represents a general speed and/or intensity of the
M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114 111

sows' reactions to any piglet-related signals including tactile ones. Factor 3 could be
labelled `nursing activity' as it is associated with high nursing frequency which is, in turn,
known to be connected with high proportion of non-nutritive nursings (this study; Fraser,
1975; CastreÂn et al., 1989; SÏpinka et al., 1997).
The three factors remained almost orthogonal after the oblique promax rotation,
demonstrating that they are independent of each other. This excludes the possibility that
differences in pig maternal style scale along just one dimension of `emotionality' or
`temperament' in the narrow sense (Wilson et al., 1994; Clarke and Boinski, 1995; Gosling
and John, 1999). An important area of future research is how sows' maternal behaviour
characteristics relate to their emotionality in other biologically signi®cant situations, such
as food competition, meeting strangers, novel environment, or behaviour towards humans
(Lawrence et al., 1991; Forkman et al., 1995; Erhard and Mendl, 1999; Erhard et al., 1999;
Thodberg et al., 1999).
The factor analysis also indicated that corticosteroid pro®les of individual sows were
related to their maternal behaviour. Speci®cally, cortisol concentrations under challenge
loaded strongly negatively on Factor 1. This may suggest a common factor underlying
`calm care' behaviour towards the piglets and `calm' physiological reaction to a challenge
(low adrenal cortex reactivity). Factor 3, describing `nursing activity', was negatively
related to cortisol baseline values. However, high or low cortisol pro®les at 5 months were
not predictive for protective reactions to piglets in danger as revealed in Factor 2. The
cortisol data were collected in juvenile animals long before the maternal behaviour was
expressed. However, it is known that cortisol pro®les remain individually stable during
ontogeny for periods of months in pigs (von Borell and Ladewig, 1992; Ladewig et al.,
1993). We, therefore, assume that the correlations between cortisol concentrations and later
maternal behaviour re¯ect a real relationship. To our knowledge, there are no other data on
the relationship between sows' pituitary-adrenocortical (PAC) pro®les and their styles of
maternal behaviour. It would be important to know whether exogenous ACTH or cortisol
will alter maternal behaviour or vice versa, whether maternal experience has an effect on
PAC functioning.
5. Conclusions
Two main conclusions follow from this study.
First, we found no differences in most aspects of maternal behaviour between domestic
and crossbred domestic�wild boar sows. This might indicate that maternal behaviour has
not changed signi®cantly during domestication and/or under modern breeding pro-
grammes.
Second, our post-hoc analysis suggests that pig maternal behaviour may be affected by
three underlying behaviour characteristics: `calmness', scaling from slow and careful
behaviour in the presence of piglets to quick and unrestful; `protectiveness', contrasting
intense and quick versus weak and slow responding to stimuli from the piglets, especially
those signalling distress or danger; and `nursing activity', in which the frequency of nursing
plays a prominent role. These behaviour characteristics or temperament traits may be
correlated with individual differences in pituitary-adrenocortical functioning.
112 M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114

Acknowledgements
This study was funded by grants No. 523/98/0789 and 523/99/0985 from the grant
Agency of the Czech Republic and grant No. M02/99/03 from the Czech Ministry of
Agriculture. Suzanne Held kindly helped with the English in two versions of the text.
References
Algers, B., Jensen, P., 1991. Teat stimulation and milk production during early lactation in sows Ð effects of
continuous noise. Can. J. Anim. Sci. 71, 51±60.
Blackshaw, J.K., Blackshaw, A.W., Thomas, F.J., Newman, F.W., 1994. Comparison of behaviour patterns of
sows and litters in a farrowing crate and a farrowing pen. Appl. Anim. Behav. Sci. 39, 281±295.
Bùe, K., 1991. The process of weaning in pigs Ð when the sow decides. Appl. Anim. Behav. Sci. 30, 47±59.
Bùe, K., 1994. Variation in maternal behaviour and production of sows in integrated loose housing systems in
Norway. Appl. Anim. Behav. Sci. 41, 53±62.
CastreÂn, H., Algers, B., Jensen, P., 1989. Occurrence of unsuccessful sucklings in newborn piglets in a semi-
natural environment. Appl. Anim. Behav. Sci. 23, 61±73.
Clarke, A.S., Boinski, S., 1995. Temperament in nonhuman-primates. Am. J. Primatol. 37, 103±125.
Dyck, G.W., Swiestra, E.E., 1987. Causes of piglet death from birth to weaning. Can. J. Anim. Sci. 67, 543±547.
Ekkel, E.D., Dieleman, S.J., Schouten, W.G.P., Portela, A., Cornelissen, G., Tielen, M.J.M., Halberg, F., 1996.
The circadian rhythm of cortisol in the saliva of young pigs. Physiol. Behav. 60, 985±989.
Erhard, H.W., Mendl, M., 1999. Tonic immobility and emergence time in pigs Ð more evidence for behavioural
strategies. Appl. Anim. Behav. Sci. 61, 227±237.
Erhard, H.W., Mendl, M., Christiansen, S.B., 1999. Individual differences in tonic immobility may reflect
behavioural strategies. Appl. Anim. Behav. Sci. 64, 31±46.
Forkman, B., Furuhaug, I.L., Jensen, P., 1995. Personality, coping patterns and aggression in piglets. Appl.
Anim. Behav. Sci. 45, 31±42.
Fraser, D., 1975. The nursing and suckling behaviour of pigs. III. Behaviour when milk ejection is elicited by
manual stimulation of the udder. Brit. Vet. J. 131, 416±426.
Gosling, S.D., John, O.P., 1999. Personality dimensions in nonhuman animals: a cross-species review. Psychol.
Sci. 8, 69±75.
Gustafsson, M., Jensen, P., de Jonge, F.H., Illmann, G., SÏpinka, M., 1999. Maternal behaviour of domestic sows
and crosses between domestic sows and wild boar. Appl. Anim. Behav. Sci. 65, 29±42.
Hartman, D.A., Ludwick, T.M., Wilson, R.F., 1962. Certain aspects of lactation performance in sows. J. Anim.
Sci. 21, 883±886.
Herskin, M.S., Jensen, K.H., Thodberg, K., 1998. Influence of environmental stimuli on maternal behaviour
related to bonding, reactivity and crushing of piglets in domestic sows. Appl. Anim. Behav. Sci. 58, 241±254.
Horrell, I., 1997. The characterisation of suckling in wild boar. Appl. Anim. Behav. Sci. 53, 271±277.
Hutson, G.D., Dickenson, L.G., Wilkinson, J.L., Luxford, B.G., 1993. The response of sows to novel visual,
olfactory, auditory and tactile stimuli. Appl. Anim. Behav. Sci. 35, 255±266.
Illmann, G., Madlafousek, J., 1995. Occurrence and characteristics of unsuccessful nursings in minipigs during
the first week of life. Appl. Anim. Behav. Sci. 44, 9±18.
Illmann, G., SÏpinka, M., SÏ teÏtkovaÂ, Z., 1998. Influence of massage during simulated non-nutritive nursings on
piglets' milk intake and weight gain. Appl. Anim. Behav. Sci. 55, 279±289.
Illmann, G., SÏpinka, M., SÏ teÏtkovaÂ, Z., 1999. Predictability of nursings without milk ejection in domestic pigs.
Appl. Anim. Behav. Sci. 61, 303±311.
Janssens, C.J.J.G., Helmond, F.A., Wiegant, V.M., 1994. Increased cortisol response to exogenous
adrenocorticotropic hormone in chronically stressed pigs: influence of housing conditions. J. Anim. Sci.
72, 1771±1777.
Jensen, P., 1986. Observations on the maternal behaviour of free-ranging domestic pigs. Appl. Anim. Behav. Sci.
16, 131±142.
M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114 113

Jensen, P., 1988. Maternal behaviour and mother-young interactions during lactation in free-ranging domestic
pigs. Appl. Anim. Behav. Sci. 20, 297±308.
Jensen, P., Gustafsson, M., 1997. Towards a functional view on domestication. In: Proceedings of the 31st ISAE
Congress, Prague, 13±16 August 1997, Polygrafia SAV, Praha, pp. 43±44.
Jensen, P., Gustafsson, M., Augustsson, H., 1998. Teat massage after milk ingestion in domestic piglets: an
example of honest begging? Anim. Behav. 55, 779±786.
Ladewig, J., de PassilleÂ, A.M., Rushen, J., Schouten, W., Terlouw, E.M.C., von Borell, E., 1993. Stress and the
physiological correlates of stereotypic behaviour. In: Lawrence, A.B., Rushen, J. (Eds.), Stereotypic Animal
Behaviour: Fundamentals and Applications to Welfare, CAB International, Wallingford, pp. 97±118.
Lawrence, A.B., Terlouw, E.M.C., Illius, A.W., 1991. Individual differences in behavioural responses of pigs
exposed to non-social and social challenges. Appl. Anim. Behav. Sci. 30, 73±86.
McGlone, J.J., Blecha, F., 1987. An examination of behavioral, immunological and productive traits in four
management systems for sows and piglets. Appl. Anim. Behav. Sci. 18, 269±289.
McKay, R.M., 1993. Preweaning losses of piglets as a result of index selection for reduced backfat thickness and
increased growth rate. Can. J. Anim. Sci. 73, 437±442.
Price, E.O., 1998. Behavioral genetics and the process of domestication. In: Grandin, T. (Ed.), Genetics and the
Behavior of Domestic Animals, Academic Press, New York, pp. 31±65.
Robert, S., Dancosse, J., Sallaire, A., 1987. Some observations on the role of environment and genetics in
behaviour of wild and domestics forms of Sus scrofa (European wild boars and domestic pigs). Appl. Anim.
Behav. Sci. 17, 253±262.
Ruis, M.A.W., Brake, J.H.A.T., Engel, B., Ekkel, E.D., Buist, W.G., Blokhuis, H.J., Koolhaas, J.M., 1997. The
circadian-rhythm of salivary cortisol in growing pigs Ð effects of age, gender, and stress. Physiol. Behav. 62,
623±630.
SÏpinka, M., Algers, B., 1995. Functional view on udder massage after milk let-down in pigs. Appl. Anim. Behav.
Sci. 43, 197±212.
SÏpinka, M., Illmann, G., Algers, B., SÏ teÏtkovaÂ, Z., 1997. The role of nursing frequency in milk production in
domestic pigs. J. Anim. Sci. 75, 1223±1228.
SÏpinka, M., Illmann, G., SÏ teÏtkovaÂ, Z., KrejcÏõÂ, P., TomaÂnek, M., SedlaÂk, L., LidickyÂ, J., 1999. Prolactin and
insulin levels in lactating sows in relation to nursing frequency. Domestic Anim. Endocrinol. 17, 53±64.
Thodberg, K., Jensen, K.H., Herskin, M.S., 1999. A general reaction pattern across situations in prepubertal
gilts. Appl. Anim. Behav. Sci. 63, 103±119.
von Borell, E., Ladewig, J., 1992. Relationship between behaviour and adrenocortical response pattern in
domestic pigs. Appl. Anim. Behav. Sci. 34, 195±206.
Weary, D.M., Pajor, E.A., Fraser, D., Honkanen, A.M., 1996a. Sow body movements that crush piglets: a
comparison between two types of farrowing accommodation. Appl. Anim. Behav. Sci. 49, 149±158.
Weary, D.M., Pajor, E.A., Thompson, B.K., Fraser, D., 1996b. Risky behaviour by piglets: a trade off between
feeding and risk of mortality by maternal crushing? Anim. Behav. 51, 619±624.
Wechsler, B., Hegglin, D., 1997. Individual differences in the behaviour of sows at the nest-site and the crushing
of piglets. Appl. Anim. Behav. Sci. 51, 39±49.
Wilson, D.S., Clark, A.B., Coleman, K., Dearstyne, T., 1994. Shyness and boldness in humans and other
animals. Trends Ecol. Evol. 9, 442±446.
114 M. SÏpinka et al. / Applied Animal Behaviour Science 70 (2000) 99±114




















