
Toxigenic fungi and mycotoxin production in Maldive fish ...
Upload
khangminh22Category
view
0download
0
Coprophilous Fungi from Koala Faeces:
A Novel Source of Antimicrobial
Compounds
Elisa Hayhoe
A thesis submitted for the degree of
Doctor of Philosophy
Department of Chemistry and Biotechnology
Faculty of Science, Engineering and Technology
Swinburne University of Technology
Melbourne, Australia
2016

Abstract
i
Abstract
An urgent need for novel antimicrobial compounds is driven by the increased resistance
of pathogens to current drugs and the rising incidence of opportunistic infections in
immunosuppressed individuals. Natural products and their derivatives have long been
exploited for their pharmaceutical potential, and fungi have provided numerous
chemically and biologically diverse secondary metabolites. Coprophilous fungi remain
relatively unexplored compared with fungi from other substrata and biological niches,
despite the fact that they are prime candidates for the discovery of antimicrobials due to
their ubiquity and their dominance in a highly competitive environment. This research
presents, for the first time, the screening of coprophilous fungi from koala faeces for
antibacterial, antifungal and anti-quorum sensing activity.
Fungi were isolated from the faeces of koalas living in Boho South and French Island in
Victoria, Australia. The 31 fungal isolates were identified by DNA sequencing and
submitted to the National Center for Biotechnology Information, where they represent
only the second set of coprophilous fungi to have been isolated from koala faeces. All
but one of the isolates were members of the phylum Ascomycota, a weighted diversity
that is common in Ascomyceteous-dominated coprophilous collections in the literature.
Extracts were prepared by lyophilisation and liquid extraction of the fermentation
liquors and mycelial biomass. Antibacterial activity was assessed against one Gram-
positive bacterium Staphylococcus aureus and three Gram-negative bacteria:
Escherichia coli, Pseudonomas aeruginosa and Klebsiella pneumoniae. In plate-hole
diffusion assays, 54.8% of the fungi produced extracts that were capable of inhibiting at
least one test bacterium. The lowest minimum inhibitory concentration values were
attributed to the extracts from a Fusarium oxysporum isolate (S6W2, 1.56 mg/mL
against S. aureus and K. pneumoniae and 0.78 mg/mL against E. coli) and a
Sordaria alcina isolate (F14P1, 3.16 mg/mL against P. aeruginosa). Antifungal activity
against Candida albicans and Drechslera brizae was tested, and 8.6% and 38.7%,
respectively, of the coprophilous fungi produced extracts that inhibited the test fungi in

Abstract
ii
agar assays. An extract from isolate S6W2 demonstrated the lowest minimum inhibitory
concentration against C. albicans (0.52 mg/mL) and an extract from an
Aspergillus niger isolate (S5P3) demonstrated the lowest minimum inhibitory
concentration against D. brizae (6.25 mg/mL). In disk diffusion assays against the
indicator strain Chromobacterium violaceum, 12.9% of the fungi produced extracts that
exhibited anti-quorum sensing activity.
The liquor extract from isolate S6W2 was separated by activity-guided fractionation
using XAD-16 resin and analytical and preparative reversed-phase high-performance
liquid chromatography in conjunction with plate-hole diffusion and microdilution
assays. Following analysis with nuclear magnetic resonance spectroscopy and mass
spectrometry, the bioactive compound was identified as fusaric acid.
This research suggests that coprophilous fungi from koala faeces may represent a source
of novel antimicrobials that warrant further exploration, especially given the paucity of
research on this particular source.

Memorial
iii
Memorial
In memory of Sydney Melbourne Brisbane
(02.05.1915–25.02.2009)
with whom I shared a curious mind.
See you in the Spring.

Dedication
iv
Dedication
I dedicate this thesis to my parents, Julie and Douglas,
for their unwavering support, love and belief in me.

Acknowledgements
v
Acknowledgements
I would like to thank my supervisor, Enzo Palombo, for fostering my interest in
research during my undergraduate and honours studies. I have always appreciated his
optimistic and enthusiastic approach to research and academia. With his open-door
policy, he created an environment where I felt encouraged to discuss all matters of
science. No question was considered too far-fetched or ingenuous, and his good sense of
humour made it all the more enjoyable. I wish to thank my associate supervisor, Qi
Yang, for her support and advice, particularly regarding the mass spectography and
nuclear magnetic resonance data. Although we did not see each other very often during
my candidature, I am grateful for her help and very kind words. I would like to express
my appreciation for the time and assistance provided by Noel Hart. Retirement did not
hold Noel back, and along with completing his PhD, he generously went out of his way
to help me in the chemistry lab, read my thesis chapters and assist with Chapter Five. I
would like to acknowledge the assistance and advice provided by the CSIRO instrument
specialists, Stuart Littler, Jo Cosgriff, Carl Braybrook and Roger Mulder. I also
acknowledge Kath Handasyde for the collection of koala faeces for this study.
Many thanks are given to Ngan Nguyen, Chris Key, Soula Mougos, Savitri Galappathie
and Andrea Chisholm for sharing their technical expertise with me. I would also like to
show my appreciation to the Swinburne Higher Degree Research personnel for their
support. It was a pleasure to teach with Daniel Eldridge, and I thank him for sharing his
chemistry knowledge as well as his positive outlook and his love of teaching. The
completion of my thesis would not have been possible without the support of Alison
Miller, and I am truly thankful for her help in my overcoming writer’s block.
Professional editor Dr Gillian Dite provided copyediting and document formatting
services according to standards D and E of the Australian Standards for Editing
Practice and the Guidelines for Editing Research Theses from the Institute of
Professional Editors. Gillian was my formatting angel, but in addition to her editing
prowess, she was an unexpected and much appreciated source of encouragement.

Acknowledgements
vi
My PhD was brighter for having met Sarah, and I thank her for her support, advice and
fantastic sense of humour. I also extend appreciation to all my other Swinburne friends
for their camaraderie, Bita, Shanthi, Kaylass, Liz, Jun, Shahanee, Jiawey, Jafar, Rue,
Azadeh, Dhivya, Snehal, Shaku, Vanu, Rashida and Rohan. I thank Justin for the
encouragement, advice and time that he has given me since our very first class together
in the undergraduate science program. I am very lucky to feel the love and support of
many school friends, but I extend a special thank you to Ella for her unwavering belief
in me and words of wisdom during challenging times and to Georgie for her life-long
friendship and ability to bring me back to non-study life with our uncontrollable
laughter. I would also like to acknowledge the quiet support from my two fluffy thesis
companions, Smokey and Pepe.
Jacob arrived late in the PhD journey, making a dynamic and energetic entrance, and an
overwhelmingly positive impact on my life and subsequently the progress of my thesis.
I will forever cherish the love, patience, honesty and support he has provided,
particularly during tough times. I cannot wait to start the next part of our lives together.
Finally, I am sincerely thankful to my family for encouraging me to pursue my goals. I
dedicate this thesis to my Mum and Dad who have gone above and beyond over my
(many) years of study, and without whom I would never have made it. All that I have
achieved and all that I am is a testament to their love, sacrifices and belief in me. There
have been some challenges along the way but I believe that they have brought us closer
together, and for that I am truly grateful. I cannot thank them enough.

Declaration
vii
Declaration
I, Elisa Hayhoe, declare that this thesis is original work and contains no material that
has been accepted for the award any other degree or diploma, except where due
reference is made.
To the best of my knowledge, this thesis contains no material previously published or
written by any other person except where due reference is made.
Where the work is based on joint research or publications, the relative contributions of
the respective workers or authors has been disclosed.
Signature: ............................................

Table of Contents
viii
Table of Contents
Abstract ......................................................................................................................... i
Memorial ..................................................................................................................... iii Dedication ................................................................................................................... iv
Acknowledgements ....................................................................................................... v
Declaration ................................................................................................................. vii
List of Tables .............................................................................................................. xii List of Figures ............................................................................................................ xiv
Abbreviations ........................................................................................................... xvii
CHAPTER 1 Introduction ............................................................................................. 1
1.1 Introduction ....................................................................................................... 2
1.2 Aim ................................................................................................................... 3
1.3 Thesis outline .................................................................................................... 3
CHAPTER 2 Literature Review .................................................................................... 5
2.1 Natural products and secondary metabolites ...................................................... 6
2.2 History of natural products ................................................................................ 6
2.2.1 Traditional medicine ................................................................................. 6
2.2.2 The golden era .......................................................................................... 7
2.2.3 A natural product decline .......................................................................... 8
2.2.4 Rekindled interest today ........................................................................... 8
2.3 The need for new antimicrobial compounds ..................................................... 10
2.3.1 Antibiotics .............................................................................................. 10
2.3.2 Antifungal agents ................................................................................... 13
2.4 Bioactive metabolites isolated from microorganisms ....................................... 14
2.4.1 Actinomycetes ........................................................................................ 16
2.4.2 Bacteria .................................................................................................. 16
2.4.3 Fungi ...................................................................................................... 17
2.5 Filamentous fungi as a source of natural products ............................................ 17
2.6 Enhancing natural product discovery from fungi .............................................. 18
2.6.1 Genome mining and engineering ............................................................ 18
2.6.2 Analytical chemistry platforms for metabolomics ................................... 19
2.6.3 Bioprospecting ....................................................................................... 21
2.7 Coprophilous fungi .......................................................................................... 23
2.7.1 Life in the dung environment .................................................................. 23

Table of Contents
ix
2.7.2 Succession of fungi on dung ................................................................... 24
2.7.3 Distribution and diversity ....................................................................... 25
2.7.4 Antimicrobial compounds from coprophilous fungi ................................ 25
2.8 Koala faeces: a unique environment for fungi .................................................. 28
CHAPTER 3 Isolation and Identification of Coprophilous Fungi ................................ 31
3.1 Introduction ..................................................................................................... 32
3.1.1 Chapter aims .......................................................................................... 33
3.2 Materials and methods ..................................................................................... 33
3.2.1 Isolation media ....................................................................................... 33
3.2.2 Collection of faeces ................................................................................ 33
3.2.3 Isolation of fungi from faeces ................................................................. 34
3.2.4 Cultivation of fungal isolates .................................................................. 35
3.2.5 Storage of fungal isolates........................................................................ 36
3.2.6 Identification of fungal isolates ............................................................... 36
3.3 Results and discussion ..................................................................................... 40
3.3.1 Collection of koala faeces ....................................................................... 40
3.3.2 Incubation of koala faeces ...................................................................... 40
3.3.3 Isolation of fungi .................................................................................... 42
3.3.4 Identification of fungi by ITS sequencing ............................................... 44
3.3.5 Comparison of the ITS regions of the fungal isolates .............................. 51
3.4 Summary ......................................................................................................... 54
CHAPTER 4 Screening of Coprophilous Fungi for Antimicrobial Activity ................. 55
4.1 Introduction ..................................................................................................... 56
4.1.1 Chapter aims .......................................................................................... 57
4.2 Materials and methods ..................................................................................... 57
4.2.1 Bacterial cultures .................................................................................... 57
4.2.2 Fungal cultures ....................................................................................... 58
4.2.3 Fungal fermentation for preparation of crude extracts ............................. 59
4.2.4 Antibacterial testing ............................................................................... 60
4.2.5 Antifungal testing ................................................................................... 63
4.2.6 Anti-QS testing....................................................................................... 66
4.3 Results and discussion ..................................................................................... 66
4.3.1 Production of mycelial extracts ............................................................... 66
4.3.2 Production of EtOAc and aqueous extracts from liquor .......................... 67
4.3.3 Antibacterial activity of coprophilous fungi extracts ............................... 68

Table of Contents
x
4.3.4 Antifungal activity of coprophilous fungi extracts .................................. 86
4.3.5 Anti-QS activity of coprophilous fungi extracts ...................................... 96
4.4 Summary ....................................................................................................... 101
CHAPTER 5 Bioactivity-Guided Separation of Extract S6W2 .................................. 104
5.1 Introduction ................................................................................................... 105
5.1.1 Chapter aim .......................................................................................... 105
5.2 Materials and methods ................................................................................... 106
5.2.1 Liquid fermentation of fungi ................................................................. 106
5.2.2 Amberlite XAD-16 resin ...................................................................... 106
5.2.3 Analytical and preparative reversed-phase HPLC ................................. 108
5.2.4 Evaporation and drying of fractions ...................................................... 108
5.2.5 Bioassays ............................................................................................. 109
5.2.6 Identification of active compound......................................................... 110
5.3 Results and discussion ................................................................................... 111
5.3.1 Selection of extract for bioactivity-guided separation............................ 111
5.3.2 Bioactivity-guided separation of S6W2 ................................................ 112
5.4 Summary ....................................................................................................... 135
CHAPTER 6 Conclusion .......................................................................................... 136
6.1 Introduction ................................................................................................... 137
6.2 Summary of findings ..................................................................................... 137
6.2.1 Chapter Three ....................................................................................... 137
6.2.2 Chapter Four ........................................................................................ 138
6.2.3 Chapter Five ......................................................................................... 139
6.3 Scope for further research .............................................................................. 140
6.3.1 Immediate source of additional study.................................................... 140
6.3.2 Broadening of the fungal survey ........................................................... 140
6.3.3 Optimisation of fermentation conditions ............................................... 141
6.4 Conclusion .................................................................................................... 142
References ................................................................................................................ 143
Appendices ............................................................................................................... 179
Appendix 1: 1H NMR spectrum (400 MHz) of F12–14 in D2O ............................. 180
Appendix 2: 13C NMR spectrum (100 MHz) of F12–14 in D2O ............................ 181
Appendix 3: COSY Spectrum of F12–14 in D2O .................................................. 182
Appendix 4: HSQC Spectrum of F12–14 in D2O .................................................. 183
Appendix 5: 1H NMR spectrum (400 MHz) of fusaric acid in D2O/NaOD ............ 184

Table of Contents
xi
Appendix 6: 13C NMR spectrum (100 MHz) of fusaric acid in D2O/NaOD ........... 185
Appendix 7: Publications arising from this thesis ................................................. 186

List of Tables
xii
List of Tables
Table 2.1 Structural classes of antibiotics* ................................................................. 11
Table 2.2 Resistance among bacteria that commonly cause infections* ...................... 12
Table 2.3 Approximate number of bioactive compounds of microbial origin* ............ 15
Table 2.4 Antibacterial (AB) and antifungal (AF) compounds isolated from coprophilous fungi* .................................................................................................... 27
Table 3.1 PCR primers ............................................................................................... 38
Table 3.2 Thermal cycle for PCR ............................................................................... 39
Table 3.3 Identification of fungi isolated from koala faeces determined by BLAST .... 49
Table 4.1 Antibacterial activity of coprophilous fungal extracts .................................. 70
Table 4.2 MIC and MBC of coprophilous fungal extracts against Gram-positive and Gram-negative bacteria* ....................................................................................... 75
Table 4.3 Summary of the most effective extracts against each test bacteria ............... 77
Table 4.4 Summary of extract concentrations used in time-course assay (mg/mL) ...... 79
Table 4.5 Inhibition of C. albicans by coprophilous fungi extracts ............................. 87
Table 4.6 Inhibition of D. brizae ................................................................................ 90
Table 4.7 MIC of coprophilous fungal extracts against D. brizae ................................ 95
Table 4.8 Anti-QS activity of coprophilous fungi extracts against C. violaceum ......... 98
Table 5.1 Summary of the antimicrobial activity of the liquor extract from the F. oxysporum isolate S6W2 (100 mg/mL)* ............................................................... 112
Table 5.2 Confirmation of bioactivity in scaled-up S6W2 liquor (12.5 mg/mL)* ...... 113
Table 5.3 Bioactivity of XAD-16 fractions from the separation of S6W2 liquor and aqueous extracts (12.5 mg/mL)* ............................................................................... 116
Table 5.4 Bioactivity of the effluent and eluate fractions from the separation of S6W2 liquor using XAD-16 resin (12.5 mg/mL)* ..................................................... 119
Table 5.5 Bioactivity of fractions 12 and 13 from the Stage 2 Separation of S6W2 liquor using preparative reversed-phase HPLC* ........................................................ 121
Table 5.6 Bioactivity of F12–14 from the preparative chromatography of S6W2* .... 123

List of Tables
xiii
Table 5.7 Summary of the 1H (400 MHz) and 13C (100 MHz) NMR spectral data of F12–14 and FA–Na+ in D2O .................................................................................. 130

List of Figures
xiv
List of Figures
Figure 2.1 Source of small-molecule drugs approved from 01/01/1981 to 31/12/2010 (Sourced from Newman and Cragg (2012).................................................. 9
Figure 2.2 The mallee moth, Telanepsia stockeri, feeds on koala faeces and was named in honour of Australia’s chief scientist, Dr John Stocker. Photo: Van Dugteren (1999). ......................................................................................................... 24
Figure 2.3 Koalas feed exclusively on Eucalyptus leaves. Photo: E. Hayhoe. ............. 28
Figure 2.4 Koala faeces present a unique substrate for fungal colonisation, consisting of highly fibrous remnants of Eucalyptus foliage (www.abc.net.au/science/scibblygum). ....................................................................... 29
Figure 3.1 The faeces were collected in Boho South and French Island in Victoria, Australia. .................................................................................................................... 34
Figure 3.2 Schematic representation of the ITS regions of fungal DNA and the binding sites of the ITS1 forward primer (ITS1 fwdpr) and ITS4 reverse primer (ITS revpr). ................................................................................................................. 38
Figure 3.3 A koala faecal pellet (a) intact and (b) cut in half, showing the dense and dry interior. .......................................................................................................... 40
Figure 3.4 a) An isolate later identified as Sordaria alcina dominated this faeces-in-agar plate. The faecal pellet in this instance was able to be cut in half so that one larger piece was present for fungal growth and isolation; b) this faeces-in-agar plate contains fragments of a faecal pellet that broke apart upon slicing. .............................. 42
Figure 3.5 Images of a selection of fungi isolated from koala faeces, growing on PDA plates. ................................................................................................................. 43
Figure 3.6 Agarose gel electrophoresis of PCR products following the amplification of the ITS regions of rDNA of a selection of fungi isolated from koala faeces. ............................................................................................................... 45
Figure 3.7 A section of the DNA sequence chromatogram showing the ITS2 region and adjacent 28S rDNA of the isolate S4P4, KT071741, Trichoderma pseudokoningii. ........................................................................................................... 46
Figure 3.8 Pairwise alignment of the ITS1 and ITS2 regions and adjacent rDNA of the isolate S4P4, KT071741, Trichoderma pseudokoningii, isolated from koala faeces to that of the sequence with the greatest similarity in the NCBI database (at the time of analysis) with the accession number X93970. ............................................ 48
Figure 3.9 Jalview representation of ClustalW multiple alignment of ITS1, 5.8S rDNA, ITS2 and adjacent 18S rDNA and 28S rDNA of four fungi isolated from koala faeces. ............................................................................................................... 52

List of Figures
xv
Figure 3.10 Phylogenetic position of fungi isolated from koala faeces. ....................... 53
Figure 4.1 S. aureus plate-hole diffusion assay, following incubation. Clear zones indicate inhibition of S. aureus by liquor extracts from the fungi S4P4 and F14P1 and by the positive control CHX. The extracts from fungi F13P1 and F14W1 did not inhibit growth of the bacterium. ............................................................................ 69
Figure 4.2 Time-course growth assay of E. coli in the presence of S6W2 liquor extract. Data points represent mean ± standard deviation............................................. 80
Figure 4.3 Time-course growth assay of S. aureus in the presence of S6W2 liquor extract. Data points represent mean ± standard deviation. ............................................ 81
Figure 4.4 Time-course growth assay of E. coli in the presence of F14P1 liquor extract. Data points represent mean ± standard deviation............................................. 82
Figure 4.5 Time-course growth assay of S. aureus in the presence of F14P1 liquor extract. Data points represent mean ± standard deviation. ............................................ 83
Figure 4.6 C. albicans plate-hole diffusion assay following incubation. Clear zones indicate inhibition of the yeast by the liquor extract from the fungus S5P3 (left) and by the positive control CHX (right). ............................................................................ 87
Figure 4.7 Example of a hyphal extension assay where the positive control (CHX) disk demonstrated only very minimal inhibition of D. brizae. ..................................... 93
Figure 4.8 Spore germination plate following incubation and treatment with MTT. A clear zone surrounds the S6W2 aqueous extract. ..................................................... 95
Figure 4.9 Anti-QS activity using C. violaceum in a disk diffusion assay. Disks were impregnated with 2 mg of liquor extracts from fungi S4P4, F13P1, F14W1 and F14P1. .................................................................................................................. 97
Figure 4.10 Example of results obtained in anti-QS disk diffusion assays. In each photo the disk is 6 mm: a) colourless, turbid anti-QS halo, b) clear antibacterial halo and c) no halo. ..................................................................................................... 98
Figure 4.11 Magnified view of the clear zone produced by the gentamicin antibiotic disk. .......................................................................................................... 100
Figure 4.12 Colourless cells were able to grow upon subculture and incubation and regained QS directed production of violacein (right). A sample of purple cells from the same plate was also subcultured for comparison (left). ........................................ 101
Figure 5.1 Bioactivity-guided separation and chromatography of S6W2 liquor from the scaled-up fermentation. ............................................................................... 114
Figure 5.2 Fractions collected from the separation of the aqueous extract of isolate S6W2 on XAD-16 resin: a) Initial column effluent that passed through the XAD-16 resin without interaction; b) first water wash; c) second water wash; d) methanol eluate; e) final methanol eluate. ................................................................................. 115

List of Figures
xvi
Figure 5.3 Preliminary bioactivity-guided separation of S6W2 fermentation liquor. . 117
Figure 5.4 HPLC chromatograms of methanol eluate fractions from the XAD-16 separation of S6W2 liquor and aqueous extracts, in 5–80% ACN/H2O/0.1% trifluoroacetic acid (TFA), peak detection λ 254 nm. Isocratic elution to 15 minutes followed by a 30 minute gradient to 80% ACN. ........................................................ 118
Figure 5.5 An overlay of analytical HPLC chromatograms of fractions 12, 13 and 14 from the preparative reversed-phase HPLC of S6W2 liquor, at λ 215 nm. Isocratic elution with 5%ACN/H2O/0.1% TFA for 15 minutes followed by a 30 minute gradient to 80% ACN/H2O/0.1% TFA. The three fractions were combined and designated F12–14. ............................................................................................. 122
Figure 5.6 Analytical reversed-phase HPLC chromatogram of F12–14 from the preparative reversed-phase HPLC of S6W2 liquor, using 10% ACN/H2O/0.1% TFA at λ 254 nm. ...................................................................................................... 124
Figure 5.7 Diode array UV profile (λ 190–400 nm) of the main peak of F12–14 at 20.15 minutes and of the first shoulder at 21.9 minutes. ............................................ 124
Figure 5.8 The effect of different mobile phase pH and injection volumes on the elution of F12–14 at λ 272 nm................................................................................... 126
Figure 5.9 The EIMS of F12–14 and of the top match chemical structure 3-butylpyridine in the National Institute of Standards and Technology spectral database. ................................................................................................................... 127
Figure 5.10 The positive and negative ESI mass spectra from the LC-MS analysis of F12–14. ................................................................................................................ 128
Figure 5.11 Structure of fusaric acid with assigned atom numbering. ....................... 129
Figure 5.12 Comparison of 1H NMR spectra (400 MHz) of (a) F12–14 in D2O and (b) Fusaric acid in D2O/NaOD. .................................................................................. 131
Figure 5.13 Comparison of 13C NMR spectra (100 MHz) of (a) F12–14 in D2O and (b) Fusaric acid in D2O/NaOD. .................................................................................. 132

Abbreviations
xvii
Abbreviations
δ chemical shift (ppm)
ACN acetonitrile
BLAST basic local alignment search tool
cfu colony-forming units
CHX chlorohexidine
COSY correlated spectroscopy (homonuclear)
D2O deuterium oxide
EIMS electron ionisation mass spectrometry
ESI electrospray ionisation
EtOAc ethyl acetate
GC-MS gas chromatography mass spectrometry
HPLC high-performance liquid chromatography
HSQC heteronuclear single quantum coherence
ITS internal transcribed spacer
J coupling constant
LC-MS liquid chromatography mass spectrometry
MBC minimum bactericidal concentration
MFC minimum fungicidal concentration
MHA Mueller–Hinton agar
MHB Mueller–Hinton broth
MIC minimum inhibitory concentration
MLST multilocus sequence typing
MS mass spectrometry
MTT thiazolyl blue tetrazolium bromide
NaOD sodium deuteroxide
NCBI National Center for Biotechnology Information
NMR nuclear magnetic resonance
PCR polymerase chain reaction
PDA potato dextrose agar

Abbreviations
xviii
PDB potato dextrose broth
QS quorum sensing
rDNA ribosomal DNA
sd standard deviation
TFA trifluoroacetic acid
UV ultraviolet
WHO World Health Organization

Coprophilous Fungi from Koala Faeces: A Novel Source of Antimicrobial Compounds
1
CHAPTER 1
Introduction

Chapter 1: Introduction
2
1.1 Introduction
Humans have a long history of exploring and exploiting the natural world for the
treatment and management of disease. But it was not until the 1940s, with the
development of penicillin, that the enormous potential of microorganisms as a source of
novel bioactive compounds was realised (Florey et al., 1949). Although the following
three decades were very productive and saw the discovery of almost all groups of
important antibiotics, pharmaceutical firms began to reduce or eliminate their natural
product research programs believing that they were not compatible with recently
developed technologies, such as high-throughput screening (Lam, 2007). What followed
was a steady decline in the output of the pharmaceutical industry, suggesting that the
dismissal of natural product research may have been misguided (Newman and Cragg,
2012).
New antimicrobial compounds are urgently needed to respond to increasing resistance
to antibiotics and antifungal agents (WHO, 2014). There is renewed belief that the
structural diversity found in nature, coupled with advances in technology, will provide
new drug candidates for the treatment of many diseases (Bérdy, 2012; Cragg and
Newman, 2013; Li and Vederas, 2009). Fungi harbour a tremendous capacity to
produce diverse and complex secondary metabolites, and the vast majority of these
remain unexplored (Bérdy, 2005; Brakhage, 2013; Chapman, 2009; Strobel, 2003). One
group of fungi that has recently piqued the interest of researchers is the coprophilous
fungi that thrive, if not dominate, in the highly competitive environment of faeces.
In the past, the majority of studies of coprophilous fungi have focused on their
taxonomy and ecology. Recently, however, the potential of coprophilous fungi for
natural product discovery has been demonstrated by a high frequency of bioactive
secondary metabolites being isolated from relatively limited efforts (Bills et al., 2013;
Essig et al., 2014; Ganesh Kumar et al., 2010; Jayanetti et al., 2015). The koala
(Phascolarctos cinereus) has an unusual diet consisting exclusively of leaves from
Eucalyptus spp., which are low in nutrients and are toxic to most animals (Moyal,

Chapter 1: Introduction
3
2008). Koala faeces contain undigested cellulose, highly lignified fibre and tannin, and
there are very few studies of the inhabiting fungi (Bell, 2005; Cribb, 1997; Peterson et
al., 2009). The current study is the first to explore the capacity for the fungi found in
koala faeces to produce antimicrobial compounds.
1.2 Aim
The major aims of this thesis were to:
i. isolate and identify fungi from koala faeces
ii. assess the antimicrobial activity of extracts prepared from the fungi
iii. identify the active compounds within the most active extract.
1.3 Thesis outline
Chapter Two provides a comprehensive review of the literature to discuss the major
concepts relevant to the project. First, the history of natural product research and the
need for new antimicrobial compounds is described. Second, the role of microorganisms
in drug discovery, with a specific focus on filamentous fungi, is discussed. The
literature review concludes with an overview of coprophilous fungi and an introduction
to koala faeces as an unusual micro-environment for fungal colonisation.
Chapter Three outlines the isolation of fungi from the faeces of koalas living in Boho
South and French Island in Victoria, Australia. The isolates were identified via DNA
sequencing and comparison with the National Center for Biotechnology Information
(NCBI) nucleotide database.
Chapter Four describes the preparation of extracts from the isolates and their screening
against Staphylococcus aureus, Escherichia coli, Pseudonomas aeruginosa, Klebsiella
pneumoniae, Candida albicans and Drechslera brizae. The extracts were also assessed
for their ability to quench quorum sensing (QS) signals, using the indicator strain

Chapter 1: Introduction
4
Chromobacterium violaceum. The premise behind this work was that the fitness of
coprophilous fungi is challenged by competing and invading organisms within koala
faeces and, therefore, survival is enhanced by the production of chemicals to mediate
these interactions.
Chapter Five presents the findings from the bioactivity-guided separation of the most
active extract and the ultimate identification of the active constituent.
Finally, Chapter Six presents a summary of this project’s major findings as well as a
brief discussion of the scope for further research.

Coprophilous Fungi from Koala Faeces: A Novel Source of Antimicrobial Compounds
5
CHAPTER 2
Literature Review

Chapter 2: Literature Review
6
2.1 Natural products and secondary metabolites
The term natural product, in its broadest sense, refers to a chemical that is produced by
a living organism and exerts a biological effect on other organisms (Colegate and
Molyneux, 2008). This includes beneficial bioactivity, such as therapeutic activity for
diseases of humans, plants and animals, as well as toxic bioactivity that is responsible
for causing some diseases. In most cases, natural products appear to be non-essential for
the organism’s everyday metabolism (Vicente et al., 2003). Because of this, the term
natural product is often used interchangeably with secondary metabolite even though
their starting materials invariably come from the major biosynthetic pathways of
primary metabolism (Madigan et al., 2003).
Secondary metabolites were once hypothesised to be waste or detoxification products
and, therefore, of little importance to an organism’s survival (Paech, 1950). However, it
has become clear that such views were dismissive and inaccurate because many
secondary metabolites (such as those involved in defence, mutualism and
communication) are key components of complex mechanisms that contribute to the
fitness of an organism (Bills et al., 2013; Cragg and Newman, 2013). The use of natural
product as a simple descriptor was suggested almost 50 years ago (Zenk, 1967) to
remove bias against any class of metabolite and to avoid the negative implications of
the word secondary. Despite this, the use of secondary metabolite has continued (in
addition to natural product) and it has become entrenched in the scientific lexicon. Thus,
in the current work, both terms will be used.
2.2 History of natural products
2.2.1 Traditional medicine
Humans have a long history of exploring and exploiting the natural world for the
treatment and management of disease. Almost all indigenous cultures have, at some
time, made use of naturally derived medicines. Examples include the well-known

Chapter 2: Literature Review
7
traditional Chinese medicine and Indian (Ayurvedic) medicine, which both date back to
approximately 3,000 BC (Ng, 2005). While traditional medicines were prepared as
crude extracts, tinctures and dried plant matter, the prescription of a therapeutic
compound would have the obvious advantage of a quantified dosage. In 1805, the first
pure pharmaceutical compound was isolated from a traditional medicinal plant:
morphine from opium produced by cut seed pods of the poppy, Papaver somniferum
(Sneader, 2005). This led to increased interest in phytochemical studies and the
isolation of natural products (such as atropine, quinine and colchicine) from plants that
were historically uses as medicines (Cragg and Newman, 2013).
2.2.2 The golden era
It was not until the late 1940s that the true potential of microorganisms as a source of
novel bioactive compounds was realised. In 1928, Sir Alexander Fleming observed the
accidental contamination of a bacterial culture (Staphylococcus spp.) by the filamentous
fungus Penicillium notatum (Fleming, 1929). Later work by Howard Florey and his
team resulted in the isolation of penicillin as the active compound as well as
characterisation of its structure and observations of its broad therapeutic use (Florey et
al., 1949). Research into microbial derived natural products was accelerated by the
onset of World War II during which infectious diseases were a major problem. In
Australia, the Commonwealth Serum Laboratories manufactured penicillin to supply the
Australian forces and some of the American forces serving in the Pacific in 1944, and,
in the same year, Australia became the first country in the world to supply penicillin
freely to civilians (CSL, 2015). The following three decades are often referred to as the
golden era of natural product research, which saw the discovery of almost all groups of
important antibacterial antibiotics: tetracyclines, aminoglycosides, cephalosporins and
macrolides (Bérdy, 2005).

Chapter 2: Literature Review
8
2.2.3 A natural product decline
By 1990, about 80% of drugs were either natural products or analogues inspired by
natural products (Li and Vederas, 2009). However, the rate of discovery and the
introduction of new natural product drugs into clinical use began to decline. Many
pharmaceutical firms began to reduce or eliminate their natural product research
programs, believing that the protocols were not compatible with the high-throughput
screening and combinatorial chemistry technologies that had been developed over this
period (Lam, 2007). These new high-throughput screening techniques were not as
successful as anticipated, with only one approved drug derived from combinatorial
chemistry over the 20 years to 2010, namely the Bayer anti-tumour drug sorafenib.
According to the review by Newman and Cragg (2012), there has been a steady decline
in the output of the pharmaceutical industry since the late 1980s. At this time, over 60
small molecule new chemical entities were being described every year by
pharmaceutical companies’ research and development programs but this reduced to an
average of 23 new chemical entities per year over the decade from 2001 to 2010. It is
noteworthy that this downturn coincided with the period of reduced interest in natural
products by the major pharmaceutical companies, and suggests that their reliance on
other techniques may have been misguided.
2.2.4 Rekindled interest today
Despite the reduction in natural product research by pharmaceutical companies, the
majority of drugs used today are either natural products, semi-synthetically produced
from natural products or chemically synthesised based on natural products. In the period
from 1981 to 2010, a total of 1,073 small molecule drugs were approved, with only 387
(36%) being classified as truly synthetic and devoid of natural inspiration (Figure 2.1)
(Newman and Cragg, 2012). Approximately half of the top-selling pharmaceuticals are
derived from natural products; these include cholesterol lowering statins and many anti-
cancer drugs and antibiotics (Demain 2009). In 2001, the worldwide market for

Chapter 2: Literature Review
9
antibiotics was US$32 billion (Projan and Youngman, 2002) and the majority of these
were (and are today) derived from natural products (Demain, 2009).
Figure 2.1 Source of small-molecule drugs approved from 01/01/1981 to 31/12/2010 (Sourced from Newman and Cragg (2012). Note: The categories of sources are as follows: S, totally synthetic drug, often found by random screening/modification of an existing agent; S/NM, synthetic/natural product mimic; S*, made by total synthesis, but the pharmacophore is from a natural product; S*/NM, made by total synthesis, but the pharmacophore is from natural product mimic; N, natural product; NB, natural product botanical.
The overall decline in the number of new chemical entities in drug development
pipelines and the ongoing need for new pharmaceuticals has rekindled interest in natural
product research. There is renewed belief that the structural diversity offered by nature,
coupled with advances in technology, will provide new drug candidates for many
diseases (Cragg et al., 2014; Cragg and Newman, 2013; Li and Vederas, 2009;
Milshteyn et al., 2014; Mishra and Tiwari, 2011).
S, 387, 36%S/NM, 146, 14%
S*, 55, 5%
S*/NM, 122, 11%
N, 59, 6%NB, 5, 0%ND, 299, 28%
S S/NM S* S*/NM N NB ND

Chapter 2: Literature Review
10
2.3 The need for new antimicrobial compounds
New antimicrobial compounds are urgently needed to respond to increased resistance to
antibiotics and antifungal agents.
2.3.1 Antibiotics
Most antibiotic classes were discovered before 1970, and a large majority of the
compounds that have been approved since then are based on chemical modifications of
existing scaffolds (Table 2.1) (Genilloud, 2014). All classes of antibiotics have seen the
emergence of resistant bacteria. Combined with the lack of new antibiotics, this
resistance represents a serious public health threat given that infectious diseases
continue to be one of the leading causes of death worldwide (Livermore, 2009).
One of the major concerns is the increased prevalence of resistance among
K. pneumoniae, S. aureus and E. coli. These bacteria cause infections that are common
in hospitals and in the community, and their resistance to current antibiotics means that
infections are becoming increasingly difficult to control. A recent report by the World
Health Organization (WHO) states that, in many settings, more than 50% of these
bacteria demonstrated resistance to commonly used antibacterial drugs (Table 2.2)
(WHO, 2014). Another concerning finding was that all WHO regions reported the
presence of K. pneumoniae that were resistant to carbapenems, which are usually the
last line of available treatment. In the current research, antibacterial activity of the
extracts was screened against these three bacteria and P. aeruginosa, which has also
demonstrated multi-drug resistance (Yuhico and Blair, 2011).

Chapter 2: Literature Review
11
Table 2.1 Structural classes of antibiotics*
Antibiotic class Antibiotic Year of discovery
Analogues developed after 2000
Penicillin (β-lactam) Penicillin G 1928
Aminoglycosides Streptomycin 1943 Plazomycin (ACHN-490) Chloramphenicols Chloramphenicol 1946
Cyclopeptides Polymixin B 1947 NAB739
Tetracyclines Chlortetracycline 1948 Tigecyclines (Omadacycline, Eravacycline)
β-lactams Cephalosporin C 1948 Macrolides Erythomycin 1948 Telithromycin
Pleuromutilin Pleuromutilin 1952 Retapamulin
Glycopeptides Vancomycin 1953 Dalbavancin, Oritavancin, Telavancin
Streptogramins Streptogramin B 1953
Rifamycins Rifampicin 1957
Lincomycins Lincomycin 1961
Quinolones Fluoroquinolones 1962 Delafloxacin, Nemonoxacin Macrolide Fidaxomicin 1975
Carbapenem (β-lactam) Imipenem 1976 Ertapenem, Doripenem
Monobactam (β-lactam) Aztreonam 1981 Lipopeptides Daptomycin 1986 CB-183,315
Oxazolidinone Linezolid 1995 Radezolid, Tedizolid * Sourced from Genilloud (2014)

Chapter 2: Literature Review
12
Table 2.2 Resistance among bacteria that commonly cause infections*
Bacterium Resistance/
Examples of typical disease
Incidencea ≥50%
resistance nationallyb
E. coli Cephalosporinsc 86 5/6
Fluoroquinolones Urinary tract infections, blood stream infections
92 5/6
K. pneumoniae Cephalosporinsc 87 6/6
Carbapenemsc Pneumonia, urinary tract infections, blood stream infections
71 2/6
S. aureus Methicillin Wound infections, blood stream infections
85 5/6
* Adapted from the WHO global report on antimicrobial resistance and surveillance [WHO 2014] a Reported by a WHO Member State (out of 194 providing data) b The number of WHO regions (out of six) with national reports of 50% resistance or more c 3rd generation drugs
2.3.1.1 QS inhibition
Bacteria are able to sense their population’s density through an intercellular QS
communication system. This system regulates gene expression in response to cell
density through the constant production and detection of signalling molecules. When
the population density reaches a certain threshold, or quorum, specific sets of genes are
collectively expressed, allowing the bacteria to act en masse. QS systems have been
shown to coordinate a variety of physiological processes in bacteria including the
production of antibiotics, bioluminescence, the release of virulence factors and biofilm
production (Kociolek, 2009). Many pathogenic bacteria rely on this communication
system for infection of their hosts and it is, therefore, a potential target for the discovery
and development of new antibacterial compounds (Helman and Chernin, 2015).
Furthermore, disruption of QS, rather than killing bacteria, may also reduce the
development of resistant strains (Hentzer and Givskov, 2003). In the current study,

Chapter 2: Literature Review
13
extracts were tested for anti-QS activity using a disk diffusion assay against the
screening bacterium C. violaceum. In this wild type strain, the production of the purple
pigment violacein is under N-acylhomoserine lactone QS control (McClean et al.,
1997). When QS signals are quenched, non-pigmented bacteria grow and can be
observed as a turbid halo of viable but colourless cells surrounding the extract
impregnated disk. Growth inhibition by an extract results in a zone of inhibition similar
to that observed in the conventional antibacterial plate-hole diffusion assay.
2.3.2 Antifungal agents
2.3.2.1 Candida
Over 20 species of the yeast Candida are capable of causing infection, making the
genus the most common cause of fungal infection worldwide (WHO, 2014). Candida
cause superficial candidiasis, such as oral thrush, as well as invasive candidiasis, such
as the bloodstream infection known as candidaemia. One common risk factor for
candidiasis is the prior use of antibiotics because it alters the normal microbiota and can
lead to dominance by Candida species (Ben-Ami et al., 2012). Therefore, candidiasis
(particularly the invasive forms) is a major problem among patients in intensive care
and in those receiving immunosuppressive therapy (Pfaller and Diekema, 2007).
Currently, there are only three classes of antifungal agents available to treat serious
Candida infections: the azoles, the echinocandins and the polyenes (e.g.,
amphotericin B). Formulations of amphotericin B are available in many countries but it
has higher toxicity compared with the other two agents and a few Candida species have
been reported to develop resistance during the treatment period (WHO, 2014). The
newer class of antifungals, the echinocandins, are the treatment of choice for Candida
infections in developed countries, but they are not yet available for standard treatment in
many developing countries and there has been an emergence of species not susceptible
to the therapy (Kale-Pradhan et al., 2012; Pfaller et al., 2011). Azoles remain the most
frequently prescribed antifungal class and are often the only therapy available. The

Chapter 2: Literature Review
14
azole fluconazole is classified as a fungistatic agent and is listed on the WHO’s current
Model List of Essential Medicines (WHO, 2015). Unfortunately, Candida species are
also rapidly acquiring resistance to fluconazole and it is therefore imperative that new
compounds with novel mechanisms of action are identified to attenuate Candida
infections. C. albicans is both a member of the healthy human microbiome and a major
pathogen in immunocompromised individuals. C. albicans was therefore chosen for use
as a test fungus in the current study in screening for antifungal activity.
2.3.2.2 Phytopathogenic fungi
Phytopathogenic fungi are becoming increasingly problematic for important food crops,
particularly in developing countries (Vurro et al., 2010). These plant diseases cause a
reduced harvest yield (or post-harvest rot), which has serious implications in terms of
food security and a sustainable economy (Fears et al., 2014). Resistance to current
agrochemicals is an ongoing challenge, and concerns relating to the safety and
environmental impact of agrochemicals have led to more stringent regulatory processes
(Lamberth et al., 2013). Thus, there is now a demand for new, safer and more selective
compounds, and it is important that natural products continue to be studied for their
potential application in crop protection (Dayan et al., 2009; Olufolaji, 2010). The
filamentous fungus D. brizae is a phytopathogen and was chosen for antifungal
screening in the current study.
2.4 Bioactive metabolites isolated from microorganisms
Secondary metabolites are produced by almost all types of living organisms. They are
produced by prokaryotes and eukaryotes of the Plant, Animal and Fungi Kingdoms,
although there are variations in their production ability among the groups (Bérdy,
2005). Historically, plants have been a major focus for natural product discovery, owing
to their structurally diverse secondary metabolites and their use in traditional medicine.
Higher plants have provided a number of important drugs, such as the anti-cancer agent
paclitaxel that was originally isolated in small quantities from the bark of the Pacific
Chapter 2: Literature Review
15
yew tree, Taxus brevifolia. Fortunately, a paclitaxel precursor was also discovered to be
produced in the needles of a different Taxus species; otherwise, an estimated 12,000 of
the slow-growing Pacific yew tree would have needed to be destroyed to produce
enough material to complete the clinical trials (Cragg and Boyd, 1996). Today, most of
the clinical paclitaxel is produced either by semisynthesis or by plant cell fermentation
using a Taxus cell line, both of which are expensive and time consuming processes
(Gond et al., 2014). Microorganisms can provide a more ecologically friendly and
potentially cheaper alternative to higher order plants because they can be artificially
cultivated and require substantially less time for growth. For example, a number of
fungi also produce paclitaxel and its precursors, and these fungi are being explored as a
potential alternative to the reliance on Taxus trees (Gond et al., 2014; Zhou et al., 2010)
A recent review by Bérdy (2012) summarises the past, current and future aspects of
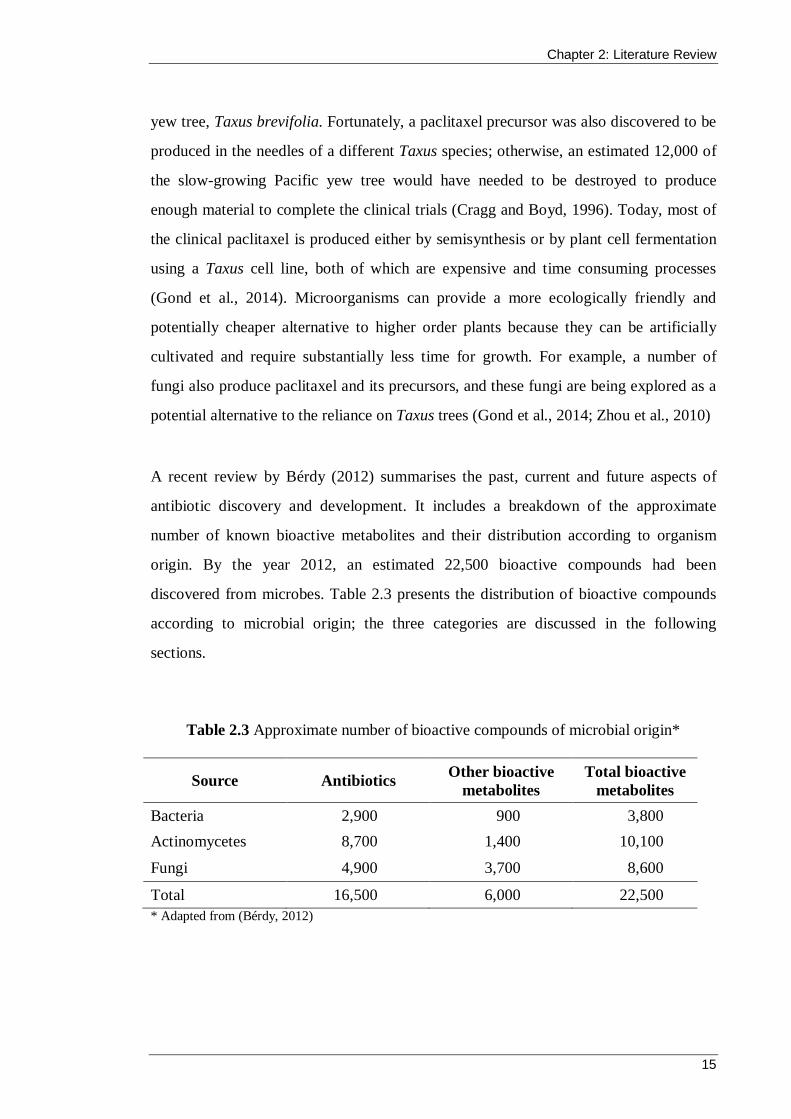
antibiotic discovery and development. It includes a breakdown of the approximate
number of known bioactive metabolites and their distribution according to organism
origin. By the year 2012, an estimated 22,500 bioactive compounds had been
discovered from microbes. Table 2.3 presents the distribution of bioactive compounds
according to microbial origin; the three categories are discussed in the following
sections.
Table 2.3 Approximate number of bioactive compounds of microbial origin*
Source Antibiotics Other bioactive metabolites
Total bioactive metabolites
Bacteria 2,900 900 3,800 Actinomycetes 8,700 1,400 10,100
Fungi 4,900 3,700 8,600
Total 16,500 6,000 22,500 * Adapted from (Bérdy, 2012)

Chapter 2: Literature Review
16
2.4.1 Actinomycetes
In terms of microorganisms, the actinomycetes represent the largest producers of
bioactive compounds. Actinomycetes are commonly isolated from soil and are closely
related to bacteria in size and physiology but are similar in structure to fungi
(Goodfellow, 2010). In the late 1930s, actinomycetes were the focus of the first
established systematic screening protocols to detect antibiotics produced by
microorganisms. This research by Selman Waksman and colleagues led to the discovery
of many new antibiotics (a word that Waksman coined), including the first effective
treatment for tuberculosis (streptomycin) for which he was awarded the Nobel Prize in
1952 (Waksman, 1954). It has been estimated that the actinomycetes produce over
10,000 bioactive compounds and account for about 45% of all microbial bioactive
secondary metabolites, with 7,600 (80%) of these compounds being produced by
Streptomyces species (Table 2.3) (Bérdy, 2012; Kurtböke, 2012). Predictive modelling
has suggested that a further 150,000 bioactive compounds are yet to be discovered from
Streptomyces species (Watve et al., 2001).
2.4.2 Bacteria
The total number of known bioactive compounds from bacteria is about 3,800, which
represents 17% of all microbial metabolites (Table 2.3) (Bérdy, 2012). The
Pseudonomas and Bacillus genera are the most well-known producers, but in recent
years, myxobacteria and cyanobacteria species have garnered interest for their prolific
production of structurally interesting natural products. For example, 16-membered ring
macrolides, known as epothilones, that constitute a novel class of antimicrotubule-
targeting agents have been isolated from various myxobacteria (Wang et al., 2009).
These compounds have a mechanism of action similar to that of paclitaxel and are being
extensively studied for their potential use in chemotherapy for tumours (Cragg and
Newman, 2013; Hofle and Reichenbach, 2012).

Chapter 2: Literature Review
17
2.4.3 Fungi
Fungi harbour a tremendous capacity to produce diverse and complex secondary
metabolites. There are approximately 8,600 known bioactive secondary metabolites,
representing 38% of all microbial products (Table 2.3) (Bérdy, 2012). Secondary
metabolism in the fungi is almost exclusively associated with the filamentous fungi of
the dikaryomycota – the Basidiomycetes and Ascomycetes. Filamentous ascomycete
genomes encode a plethora of enzymes that synthesise secondary metabolites, including
non-ribosomal peptide synthetases, polyketide synthases and terpene synthases. In
contrast, the Basidiomycetes favour terpenoid biosynthesis for their secondary
metabolite production, with polyketide synthases and non-ribosomal peptide synthetases
occurring to a lesser extent (Bills et al., 2013; Wawrzyn et al., 2012). These enzymes
build the core structural scaffolds of most fungal secondary metabolites and the genes
that encode them are generally located in clusters. Genome mining of secondary
metabolite gene clusters has demonstrated that many pathways remain silent under
standard cultivation conditions, indicating that the capability of fungi to produce
secondary metabolites is far greater than first thought (Brakhage, 2013; Spraker and
Keller, 2014).
2.5 Filamentous fungi as a source of natural products
Fleming’s observation of antimicrobial activity in the filamentous fungus P. notatum
and the subsequent development of the antibiotic penicillin by Florey and colleagues is
arguably the most significant discovery of a bioactive compound from fungi. Penicillin
was the first available broad spectrum antibiotic and its discovery set up the paradigm
for future drug discovery research from fungi. Since then, thousands of compounds
have been discovered that inhibit the growth of bacteria, fungi, protozoa, parasites,
insects, viruses and even human tumour cells. Many other molecules with cytotoxic,
mutagenic, carcinogenic, teratogenic, immunosuppressive, enzyme inhibitory,
allelopathic and other biological effects have been found (Keller et al., 2005). The most
famous fungal metabolites in clinical use today include the β-lactam antibiotics (e.g.,

Chapter 2: Literature Review
18
penicillins G and V), the cholesterol lowering statin lovastatin and its derivatives, the
immunosuppressant cyclosporin, and the migraine medication ergotamine. The statin
drugs (such as lovastatin, mevinolin, compactin, pravastatin and atorvastatin) are the
biggest selling pharmaceutical group today, providing enormous revenue to
pharmaceutical companies and extending the lives of millions of people (Endo, 2010).
Filamentous Ascomycetes are the most prolific producers of bioactive compounds
among the fungal species, with approximately 6,400 compounds having been isolated.
The three common genera Aspergillus, Penicillium and Fusarium have produced
approximately 950, 900 and 350 compounds, respectively, and several hundred have
been isolated from species of Trichoderma, Phoma, Alternaria and Acremonium
(Bérdy, 2005).
With the return to natural product drug discovery, there has been a recent increase in
interest towards secondary fungal metabolites, and the belief that they may provide the
leads and scaffolds for the development of desperately needed drugs for a multitude of
diseases (Bills et al., 2013; Brakhage, 2013; Prakash, 2015).
2.6 Enhancing natural product discovery from fungi
Fungal natural product research is most effective when it takes a multidisciplinary
approach that respects the complexity and diversity of nature and also embraces new
technologies and approaches (Cragg et al., 2014; Demain, 2009). The renewed interest
in compounds derived from natural products has led to rapid development in diverse
research disciplines. The following sections describe three key areas of research that are
enhancing and accelerating natural product discovery from fungi.
2.6.1 Genome mining and engineering
Advances in microbial genomics have led to a greater understanding of the biosynthetic
gene clusters in fungi that are responsible for the production of important secondary

Chapter 2: Literature Review
19
metabolites (Milshteyn et al., 2014; Zotchev et al., 2012). Since the first genome of a
filamentous fungus (Neurospora crassa) was published in 2003 (Galagan et al., 2003),
approximately 1,204 full genome sequences of fungi have become available (Genilloud,
2014). The number of sequenced genomes is expected to grow dramatically in the near
future, and with automated gene prediction technology, this will provide a preview of
the biosynthetic pathways within a fungus, for both known and unknown natural
products (Bills et al., 2013). Activation of silent or unproductive biosynthetic pathways
can result in the isolation of novel bioactive compounds, superior analogues of
previously known compounds or increase the yield of natural products (Brakhage,
2013). Furthermore, manipulation of the pathways via metabolic engineering (also
known as combinatorial biosynthesis) can allow microbes to produce compounds that
would not normally exist in nature (Krivoruchko and Nielsen, 2015; Wong and Khosla,
2012; Wu et al., 2012). For example, Fisch et al. (2011) used combinatorial biosynthesis
of fungal polyketide synthases to produce bassianin in Aspergillus oryzae. This
metabolite had already been isolated but the original strain no longer exists and the
production of bassianin had not been observed since. Thus, bassianin was effectively an
extinct metabolite. Rational domain substitutions between polyketide synthases that
encode the biosynthesis of closely related compounds was used to create hybrid
synthetases. Combined with the coexpression of two cytochrome P450 encoding genes,
the biosynthesis pathway for bassianin was resurrected.
2.6.2 Analytical chemistry platforms for metabolomics
The improvement of analytical chemical platforms combined with the benefits of high-
throughput screening appears to be one area of focus for future success in natural
product discovery from fungi. Bioactivity-guided separation is a technique commonly
used in natural product drug discovery and was the methodology employed in the
current study. The aim of the process is to isolate compounds with particular biological
activities from crude extracts. To achieve this, the abundant and often diverse
compounds in the extract must be separated into fractions that are then screened for the
biological activity of interest. The fractions without bioactivity are disregarded and

Chapter 2: Literature Review
20
those that demonstrate bioactivity are further separated, often using a variety of
separation techniques, until the bioactive constituent has been isolated and can be
identified.
Given the complexity of the initial natural extract, bioactivity-guided separation is often
a complicated, labour-intensive and time-consuming process. The complexity of the
starting material does not make it amenable for the rapid high-throughput screening
protocols that are favoured by pharmaceutical research and development programs. An
ideal solution is automated separation of the crude extract into individual components,
coupled with full spectroscopic identification prior to high-throughput screening (Li and
Vederas, 2009). Alternatively, the crude extract can undergo partial purification by pre-
treatment (to remove promiscuous materials such as tannins) or pre-fractionation prior
to high-throughput screening (Cragg and Newman, 2013). The development of modern
hyphenated chemical analytical techniques, such as high-performance liquid
chromatography (HPLC) coupled online to nuclear magnetic resonance (NMR)
spectroscopy, has helped to address this challenge by enabling the rapid separation and
identification of potential drug leads from an extract (Brkljača and Urban, 2011).
Techniques such as this often include high-resolution mass spectrometry analysis and
comparison of hits to a compound database that provides an assessment of a secondary
metabolite’s novelty and its potential as a drug candidate (El-Elimat et al., 2013;
Nielsen and Larsen, 2015).
Recently, a new method was described by Sica et al. (2015) that eliminates the need for
traditional extraction processes of a fungus, and subsequent activity-guided separation.
The in situ technique achieves identification of secondary metabolites directly on the
surface, or surrounding, a fungal culture in a Petri dish. Continued development of
analytical chemistry platforms for metabolomics studies will contribute to the
improvement of drug discovery from fungi and aid researchers in finding novel
compounds.

Chapter 2: Literature Review
21
2.6.3 Bioprospecting
The two previous approaches are concerned with enhancing natural product discovery,
but once a producer has been established, bioprospecting efforts aim to expand the
diversity of fungi available for exploitation. The number of described fungal species in
the world is varyingly estimated from 45,000 to up to 300,000. This represents only a
small proportion of the predicted total diversity, with several studies estimating that
there may be as many as 1.5 million species (Chapman, 2009). Therefore, there is a
largely untapped reservoir of fungi awaiting discovery, and given their proven potential
for producing structurally complex and diverse secondary metabolites, isolation of new
species will increase the likelihood of discovering novel bioactive compounds.
In the search for bioactive compounds, biodiverse environments are more likely to play
host to increasingly diverse microbiota, which in turn, may produce chemically novel
metabolites (Bérdy, 2005). Australia has been described as a megadiverse nation,
placing it in a group of countries that have less than 10% of the global surface but
support more than 70% of the biological diversity on earth (Kumar et al., 2015).
Although this statement may have been in response to the myriad of flora and fauna in
Australia, the diversity of macroorganisms in an environment is likely mirrored by the
diversity of microorganisms (Strobel, 2006). Thus, the various biotopes within Australia
represent an ideal location to search for new species of fungi.
2.6.3.1 Biological niches of interest
Further to exploring biodiverse geographical locations for fungi, interest has grown in
three unique biological niches that have emerged as natural product hot spots. First, the
intracellular spaces of many higher plants support the growth of a group of fungi known
as endophytes. These usually exist as symptomless inhabitants, suggesting a long-
standing and complicated symbiotic relationship with the plant (Strobel and Daisy,
2003). Adaptation to the micro-environment has included the uptake of some plant
DNA into their genomes, which has allowed certain endophytes to synthesise

Chapter 2: Literature Review
22
phytochemicals originally associated with the host (Zaferanloo et al., 2012). For
example, Stierle et al. (1993) reported that an endophytic fungus (Taxomyces
andreanae) isolated from T. brevifolia was capable of producing the same paclitaxel
compound for which the host plant was well known. Since then, many endophytic fungi
have been found to produce paclitaxel, both from Taxus species and non-Taxus species
(Kumaran et al., 2009; Liu et al., 2009; Strobel et al., 1996; Zhou et al., 2010). The
relatively low yield from these fungi needs to be improved before they can be
considered a viable alternative for commercial production (Gond et al., 2014).
Numerous compounds with antimicrobial activity have also been isolated from
endophytes, including those of the alkaloid, peptide, steroid, terpenoid, phenol, quinone,
flavonoid and aliphatic chemical classes (Zaferanloo et al., 2012).
The second biological niche that has recently garnered interest is faeces, particularly
from herbivorous mammals, that support the growth of coprophilous fungi. These fungi
are the focus of the present study and will be discussed in greater depth in Section 2.7.
Finally, marine fungi have recently emerged as a biological niche that is capable of
producing a diverse range of natural products. Although the majority of the world’s
surface is oceans, the marine environment has not been studied as extensively as the
terrestrial environment due to the difficulty in collecting samples. Early research into
marine natural products was limited to the study of organisms from shallow waters until
improvements were made in the areas of scuba diving and trawling (Cragg and
Newman, 2005). One study conducted by Giuseppe Brotzu in 1945 investigated
seawater samples from a sewage outlet in Sardinia and led to the discovery of the
β-lactam antibiotic cephalosporin C from Acremonium chrysogenum (Abraham and
Loder, 1972). Since then, marine fungi have been isolated from various sources
including sponges, algae, wood, tunicates, sediment, molluscs, coral, plants and fish,
and have led to the discovery of over 1,000 natural products (Gomes et al., 2015; Rateb
and Ebel, 2011).

Chapter 2: Literature Review
23
2.7 Coprophilous fungi
Coprophilous fungi are fungi that inhabit or are associated with the faeces of animals,
including soil contaminated with faeces. Coprophilous fungi play an important role as
recyclers in the ecosystem by degrading the contents of faeces and subsequently
returning micronutrients into the ecosystem. The majority of studies of coprophilous
fungi have focused on their taxonomy and ecology. Recently, however, the potential of
coprophilous fungi for natural product discovery has been supported by a high
frequency of bioactive secondary metabolites isolated from relatively limited efforts
(Bills et al., 2013).
2.7.1 Life in the dung environment
The spores of some coprophilous taxa will only germinate on faeces if they have
previously travelled through the digestive track of the animal, whereas other species are
introduced to the faeces via air dispersal of spores or contact with other surfaces (Misra
et al., 2014). Following growth on the faeces, the fungi are dispersed to nearby
vegetation by rain, arthropods, mammals, or by their spores being forcefully discharged
into the air. The latter method forms part of a cyclic relationship that some species have
developed, whereby their spores are mucilaginous and adhere to the vegetation that is
eaten by the animal whose faeces they colonise. The spores are then ingested by the
animal and germination, growth and sporulation occur on the freshly excreted faeces
(Krug et al., 2004). Other coprophilous fungi, such as Mucor hiemalis, produce a sticky
droplet around their spores that sticks to the bodies of insects that visit the dung. The
insects then inadvertently disperse the spores to a new environment. A similar method is
employed by some cleistothecial Ascomycetes whose fruiting bodies have modified
appendages for attachment to fur. The structures are particularly useful for fungi that
inhabit the faeces of rodents because they deposit dung at the entrance to their burrow
and thus act as a vector between different dung deposits (Krug et al., 2004). In
Australia, some species of Mallee moth feed on koala faeces and lay their eggs on the
surface. The caterpillars then complete their development in a single dung pellet, spin

Chapter 2: Literature Review
24
their cocoons and emerge as adult moths Figure 2.2 (Van Dugteren, 1999). These moths
help to recycle nutrients back into Australia’s nutrient-poor soils, and their ingestion of
the faeces may also aid in disseminating coprophilous fungi.
Figure 2.2 The mallee moth, Telanepsia stockeri, feeds on koala faeces and was named in honour of Australia’s chief scientist, Dr John Stocker. Photo: Van Dugteren (1999).
2.7.2 Succession of fungi on dung
The succession of fungi that appear on dung at various stages of degradation appears to
follow a generalised pattern (Richardson, 2002). Fast-growing Zygomycetes are
frequently observed early in succession, especially on fresh dung, and use the easily
metabolised simple sugars, starch and protein. Ascomycetous species are the most
commonly identified coprophilous fungi. These appear when the simple carbon sources
are depleted because they can use the hemicellulose and cellulose material.
Basidiomycetes usually appear later in the succession and are able to metabolise both
cellulose and lignin (Krug et al., 2004; Sarrocco et al., 2015a; Wicklow et al., 1980).
The order of appearance is also likely to be affected by the time taken to form fruiting
bodies. The simple sporangia of the Zygomycetes can develop more quickly than the
more complex Ascomycete fruiting body, which requires less energy than the even

Chapter 2: Literature Review
25
larger Basidiomycete fructification (Krug, 2004). Furthermore, the activities of
competitors such as bacteria, insects, worms and other fungi can influence succession
(Krug et al., 2004). The challenge that these competitors pose to the fitness of
coprophilous fungi is met, at times, with the production of bioactive compounds by the
fungi, and this forms the premise behind bioprospecting of coprophilous fungi for
natural products.
2.7.3 Distribution and diversity
The majority of research on coprophilous fungi is concerned with identification, usually
by morphology, and a high proportion of the studies are from Europe and North
America. There are approximately 52 genera of Zygomycetes, 169 genera of
Ascomycetes and 33 genera of Basidiomycetes that are known to use dung as a
substrate (Misra et al., 2014) and new species continue to be discovered (Fukiharu et al.,
2015; Goh et al., 2013; Kruys, 2015). The literature on coprophilous fungi describes
species from a large variety of animal dung, including sheep, cow, horse, camel, llama,
deer, tapir, rabbit, hare, dormouse, hedgehog, chamois, grouse, goose, goat, buffalo,
zebra, elephant, giraffe, rhinoceros, tiger, kangaroo, wombat and koala (Bell, 2005;
Bills and Polishook, 1993; Cribb, 1988, 1989, 1997; Doveri et al., 2012; Doveri et al.,
2010; Ganesh Kumar et al., 2010; Goh et al., 2013; Gupta, 2010; Melo et al., 2012;
Pandey, 2009; Richardson, 2001c; Sarrocco et al., 2015c; Watling and Richardson,
2010). In almost all surveys of coprophilous fungi, species of the phylum Ascomycota
dominate the collection. For example, Watling and Richardson (2010) studied the fungi
from dung samples of sheep, cattle, horse, rabbit and goose living in the Southern
Atlantic archipelago of the Falkland Islands. They isolated 97 taxa, 60 of which were
Ascomycetous species, 28 were Basidiomycetes and five were Zygomycetes.
2.7.4 Antimicrobial compounds from coprophilous fungi
The fitness of coprophilous fungi in the dung environment is challenged by the presence
of bacteria, protists, invertebrates and other fungi competing for a nutrient-rich,

Chapter 2: Literature Review
26
transient resource. Within the dung microcosm there are antagonistic and defensive
interactions that may require the production of bioactive compounds that affect
prospective competitors or predators (Gloer, 1995). Although competitive interactions
had long been observed among fungi (Ikediugwa and Webster, 1970; Wicklow, 1981),
it was the formative research of J. B. Gloer and colleagues into the chemistry associated
with these interactions that exemplified their potential for producing bioactive
compounds. Since their isolation of an antifungal diphenyl ether from Preussia
fleischhakii in 1988 (Weber and Gloer, 1988), Gloer has co-authored at least 23 papers
that describe the identification of one or more novel bioactive metabolites from
coprophilous fungi (see references in Table 2.4).
In comparison with the fungal endophytes, reports of novel antimicrobial compounds
from coprophilous fungi have been limited and come mainly from a few laboratories.
However, the efforts that have been undertaken have shown that coprophilous fungi are
indeed a rich source of structurally diverse metabolites, with several demonstrating
antibacterial activity and even more inhibiting fungi. The increased occurrence of
antifungal activity may be biased because of the dependence on fungal phenotypic
assays in the initial screening process (Bills et al., 2013). It is therefore important to also
test extracts and compounds for antibacterial activity.
Table 2.4 lists some of the known species of coprophilous fungi that are reported to
have produced antifungal or antibacterial compounds. Compounds from all major
biosynthetic classes are represented, and all but one of the examples (Coprinopsis
cinerea) are members of the Ascomycota phylum. The coprophilous ascomycetes have
emerged as the prominent phylum for producing antimicrobial compounds due to their
higher frequency of isolation and their broader array of biosynthesis enzymes (Bills et
al., 2013).

Chapter 2: Literature Review
27
Table 2.4 Antibacterial (AB) and antifungal (AF) compounds isolated from coprophilous fungi*
Genus Species Compounds Biological
activity (AF/AB)*
Reference
Podospora araneosa Sordarins AF (Hauser and Sigg, 1971a; Odds, 2001)
Pseudeurotium ovalis Pseurotins AB (Bloch and Tamm, 1981; Breitenstein et al., 1981)
Pseudoarachniotus roseus Aranorosins AB
(Fehlhaber et al., 1988; Mukhopadhyay et al., 1998; Roy et al., 1992)
Preussia isomera Preussomerins AF (Weber et al., 1990) Sporormiella similis Similins AF (Weber et al., 1992) Podospora appendiculata Appendolides AF (Wang et al., 1993)
Scopulariopsis Unknown Restricticins AF (O'Sullivan et al., 1993)
Apiospora montagnei Apiosporamide AF (Alfatafta et al., 1994)
Coniochaeta saccardoi Coniochaetones AF (Wang et al., 1995a)
Petriella sordida Petriellins AF (Lee et al., 1995) Polytolypa hystricis Polytolypin AF (Gamble et al.,
1995) Sporormiella australis Australifungin AF (Hensens et al.,
1995) Sporormiella intermedia Zaragozic acid B AF (Bergstrom et al.,
1995) Sporormiella teretispora Terezines AF (Wang et al.,
1995b) Cercophora areolate Cercophorins AF (Whyte et al., 1996) Cercophora sordarioides Arthrinones
Cerdarin AF AF
(Whyte et al., 1997)
Podospora anserine Anserinones AF/AB (Wang et al., 1997) Ascodesmis sphaerospora Arugosin F AF (Hein et al., 1998) Preussia fleischhakii Diphenyl ethers AF (Weber and Gloer,
1988) Sporormiella vexans Sporovexins AF/AB (Soman et al.,
1999) Bombardioidea anartia Bombardolides AF/AB (Hein et al., 2001) Nigrosabulum globosum Pseudodestruxins
Ascochlorin AB AF
(Che et al., 2001)
Stilbella aciculosa Fusidic acid AB (Kuznetsova et al., 2001)
Podospora decipiens Decipenin A Decipenolides
AF/AB AB
(Che et al., 2002)
Coniochaeta ellipsoidea Coniosetin AF/AB (Segeth et al., 2003)
Podosordaria tulasnei Tulasnein AF (Ridderbusch et al.,

Chapter 2: Literature Review
28
Genus Species Compounds Biological
activity (AF/AB)*
Reference
2004) Podospora communis Communiols AB (Che et al., 2005;
Che et al., 2004a) Podospora curvicolla Curvicollides AF (Che et al., 2004b) Sporormiella minimoides Sporminarins AF (Mudur et al.,
2006) Stilbella erythrocephala Antiamoebins AF (Lehr et al., 2006) Coprinopsis cinerea Copsin AB (Essig et al., 2014) Hypocopra rostrata Hypocoprins AB (Jayanetti et al.,
2015) * Modified from Bills et al. (2013)
2.8 Koala faeces: a unique environment for fungi
The koala (Figure 2.3) is an arboreal marsupial inhabiting the bushland of eastern and
south-eastern Australia. The koala has an unusual diet consisting exclusively of the
leaves from Eucalyptus spp., which are low in nutrients and are toxic to most animals
due to the presence of phenolics (principally tannin) and essential oils (Cork and Foley,
1997).
Figure 2.3 Koalas feed exclusively on Eucalyptus leaves. Photo: E. Hayhoe.

Chapter 2: Literature Review
29
Due to their nutrient-poor diet, koalas spend most of their time sleeping to conserve
energy (Cork and Hume, 1983). Koalas are able to obtain nutrition from Eucalyptus
leaves by hindgut fermentation using their highly developed caecum, which is
proportionately the largest of all animals. The finer, more nutritious leaves are retained
in the caecum and colon (the hindgut) and fermented by anaerobic bacteria to detoxify
the material and release enough nutrition to maintain the koala’s slow metabolism
(Moyal, 2008; Osawa et al., 1993). The larger particles, consisting of undigested
cellulose, hemicellulose, highly lignified fibre and tannin, are compressed in the rectum
and excreted rapidly as firm, dry and fibrous faecal pellets (Figure 2.4) (Moyal, 2008;
Tyndale-Biscoe, 2005).
Figure 2.4 Koala faeces present a unique substrate for fungal colonisation, consisting of highly fibrous remnants of Eucalyptus foliage (www.abc.net.au/science/scibblygum).
The diet and digestive system of an animal can influence the diversity of coprophilous
fungi in the dung microcosm (Kruys and Ericson, 2008). The unusual feeding habits and
hindgut digestion of the koala lends weight to the hypothesis that interesting
coprophilous fungi will be isolated from their faeces. There has been very little research
into the fungi that grow on koala faeces, and the early studies were conducted from a
taxonomic perspective only (Bell, 2005; Cribb, 1997). Recently, Peterson et al. (2009)
screened fungi from koala faeces for the production of hydrolytic enzymes on the
premise that, to survive, they must be capable of degrading the lignocellulose-rich
material that the faeces contain. They isolated 37 fungi: 18 Ascomycetous genera, three
Basidiomycetes and one Zygomycete. Their research represents the one other instance,

Chapter 2: Literature Review
30
besides the current study, in which fungi from koala faeces were identified by internal
transcribed spacer (ITS) sequencing. The following chapters present, for the first time,
the systematic isolation of fungi across a succession period from koala faeces and their
identification by ITS sequencing for the principal purpose of screening their secondary
metabolites for antimicrobial activity.

Coprophilous Fungi from Koala Faeces: A Novel Source of Antimicrobial Compounds
31
CHAPTER 3
Isolation and Identification of Coprophilous Fungi

Chapter 3: Isolation and Identification of Coprophilous Fungi
32
3.1 Introduction
Fungi are an important but often overlooked member of the Australian biota.
Estimates of the number of described species in Australia range from 5,672 to
12,500, and the total diversity is varyingly estimated from 50,000 up to 250,000
(Chapman, 2009). Studies of coprophilous (dung-inhabiting) fungi often focus on
their taxonomy and their ecological aspects rather than investigating their secondary
metabolites for bioactivity. Coprophilous fungi have shown promise as a source of
antimicrobial compounds (see Section 2.7.4), but the majority of research has taken
place in the northern hemisphere, leaving Australian coprophilous fungi as an
underexplored arena primed for discovery (Bell, 2005; Krug et al., 2004).
The koala has an unusual diet consisting solely of eucalyptus leaves, which are
extremely toxic to most animals. The koala is able to extract enough nutrients from
eucalyptus leaves to maintain metabolism and excretes faeces consisting of
undigested cellulose, highly lignified fibre and phenolic compounds (Peterson et al.,
2009). The digestive system of an animal may influence the species composition and
richness of the dung microcosm (Kruys and Ericson, 2008), and therefore, the
unusual feeding habits and hindgut digestion of the koala lends weight to the
hypothesis that interesting coprophilous fungi will be isolated from their faeces.
To the best of my knowledge, this is the first time that fungi have been
systematically isolated across a succession period from koala faeces and identified by
ITS sequencing for the principal purpose of screening their secondary metabolites for
antimicrobial activity. The ITS sequences obtained in this research have been
submitted to the NCBI and represent only the second collection of coprophilous
fungi from koala faeces in their database (Peterson et al., 2009). This chapter
describes the isolation and identification of coprophilous fungi from the faeces of
koalas living in Boho South and French Island in Victoria, Australia.

Chapter 3: Isolation and Identification of Coprophilous Fungi
33
3.1.1 Chapter aims
The major aims of this chapter were to:
i. isolate coprophilous fungi from a collection of koala faeces
ii. identify fungal isolates via sequencing of the ITS rDNA regions
iii. determine the phylogenetic relationship between the isolates.
3.2 Materials and methods
3.2.1 Isolation media
Culture media were purchased from Becton Dickinson (NJ, USA). All media were
prepared using distilled water and sterilised by autoclaving at 121 °C for 15 minutes
prior to use. Potato dextrose agar (PDA) consisted of 4 gL−1 potato starch, 20 gL−1
glucose and 15 gL−1 agar with a natural pH of 5.6 ± 0.2. Potato dextrose broth (PDB)
was used for liquid cultivation of isolates and consisted of 4 gL−1 potato starch and
20 gL−1 glucose. Water agar consisted of 15 gL−1 agar. Agar plates were prepared by
aseptically by pouring 15–20 mL of molten (~50 °C), sterilised media into circular
(80 mm diameter) Petri dishes (Techno-Plas, Australia).
3.2.2 Collection of faeces
Koala faeces were obtained in Victoria, Australia from areas surrounding Boundary
Hill Rd in Boho South and Bayview Rd on French Island (Figure 3.1). The faeces
were collected by Dr Kath Handasyde (Department of Zoology, University of
Melbourne, Australia) within 15 minutes of falling to the ground using clean
disposable gloves and they were then lightly brushed to remove any materials
adhering to the surface. This was to reduce the isolation of organisms that had
originated in the soil and leaf debris adjacent to the faeces rather than being present
in and on the faeces themselves. The faeces were then stored in clean paper
envelopes where they were allowed to dry at room temperature until further use. This

Chapter 3: Isolation and Identification of Coprophilous Fungi
34
drying process minimised the number of bacteria that may grow when the faeces are
later incubated (Krug, 2004).
Figure 3.1 The faeces were collected in Boho South and French Island in Victoria, Australia.
3.2.3 Isolation of fungi from faeces
Faeces-in-agar and ethanol-faeces-agar (pour-plate and spread-plate) plates were
prepared from surface sterilised koala faeces. All plates were incubated in the dark at
25 °C for up to 6 weeks. The plates were inspected daily and fungi were isolated by
either transferring hyphae as they appeared or by using a sterile inoculating loop and
blade to transfer small slices of agar to a separate PDA plate. The isolates were then
incubated at 25 °C and monitored to ensure the absence of contamination.
3.2.3.1 Surface sterilisation
To sterilise the surface of the koala faeces, the pellets were soaked in a weak bleach
solution (0.01% v/v NaClO) for 1 minute, rinsed twice with MilliQ water and blotted
dry with sterile filter paper (Bradner et al., 2000).
This image cannot currently be displayed.

Chapter 3: Isolation and Identification of Coprophilous Fungi
35
3.2.3.2 Faeces-in-agar plates
Using a sterile blade and forceps, the surface-sterilised faecal pellets were cut in half
lengthways and each half was placed in the centre of an empty Petri dish. Then,
20 mL of cooled, molten water agar was poured into one Petri dish to surround the
pellet and 20 mL of PDA was poured into the other Petri dish.
3.2.3.3 Ethanol-faeces-agar plates
Faecal pellets were cut into small pieces (approximately 2 mm3) with a sterile blade
and then placed in 60% ethanol and shaken by hand for 4 minutes. First, 1 mL of the
faeces suspension was removed and added to a sterile Petri dish. Then 20 mL of
PDA was poured into the dish and moved gently to evenly mix the suspension and
the agar, thus achieving a pour-plate (Bills and Polishook, 1993). A further 1 mL of
the suspension was spread evenly onto the surface of a PDA plate with a sterile glass
spreader to produce a spread-plate for isolation of fungi.
3.2.4 Cultivation of fungal isolates
All fungal subculture and inoculation processes were carried out in a Class 1
ultraviolet biological safety cabinet (Email Westinghouse Pty Ltd.) with adherence to
aseptic techniques.
Fungal isolates were grown on PDA plates after inoculation from agar slant stock
cultures. A small portion of the mycelium that was growing at the slant perimeter
was removed and transferred to the centre of a PDA plate. The cultures were
incubated at 25 °C until radial growth approached the Petri plate edges.

Chapter 3: Isolation and Identification of Coprophilous Fungi
36
3.2.5 Storage of fungal isolates
Medium-term storage of fungal isolates was achieved using agar slants. McCartney
bottles were filled with 15 mL of PDA then sterilised and allowed to solidify on an
angle of approximately 30° to produce a slant. A segment of mycelia from an isolate
growing on a Petri plate was placed on the surface of the agar slant and incubated
until growth had extended approximately 30 mm. Agar slants were then stored
upright at 2 °C to 8 °C.
For long-term storage of fungal isolates, glycerol stocks were prepared.
Microcentrifuge tubes containing 180 µL of liquid fungal culture and 20 µL of 80%
glycerol (at a final concentration of 8%) were stored at −80 °C for up to 24 months.
Deep freeze stocks for the purpose of DNA extraction were also prepared. Slices of
fungi were removed from the surface of the PDA plates and then transferred to
microcentrifuge tubes and stored at −80 °C without the addition of a glycerol
solution. Preliminary experiments suggested that this was an adequate form of
storage for the later extraction of DNA.
3.2.6 Identification of fungal isolates
Pure cultures of each fungal isolate were established on PDA and photos were taken
and general observations were recorded. Identification of the isolates was performed
via amplification and direct sequencing of the ITS1, 5.8S and ITS2 regions of the
fungal rDNA and comparison with the NCBI nucleotide database
(http://blast.ncbi.nlm.nih.gov/Blast.cgi).
3.2.6.1 Extraction of fungal DNA
Genomic DNA was isolated from the fungi using the ZR Fungal/Bacterial DNA
MiniPrep kit (Zymo Research Corp., USA) with the following minor adjustments to

Chapter 3: Isolation and Identification of Coprophilous Fungi
37
the manufacturer’s protocol. Each sample was introduced to a lysis tube in the form
of a slice (approximately 5 mm × 15 mm) taken from the perimeter of actively
growing fungi on a PDA plate. Prior to DNA extraction, these mycelia samples had
been stored at −80 °C according to the long-term storage protocol (see Section 3.2.5).
After the addition of the DNA elution buffer, the column matrix was allowed to rest
for 4 minutes. The isolated DNA was assessed, in terms of quantity and quality, by
1% (w/v) agarose gel electrophoresis.
3.2.6.2 Agarose gel electrophoresis
Agarose gel electrophoresis was used to analyse extracted DNA and fragments
generated by polymerase chain reaction (PCR). A 50 × stock solution of Tris-acetate-
EDTA was prepared with (per 1 L) 2 M Tris base and 6.5 M EDTA disodium salt at
a pH of 8. Ethidium bromide (at a final concentration of 0.5 µg/mL) was
incorporated into the 1% (w/v) agarose gels to allow for detection of DNA bands. A
loading dye was added to each sample lane and a 1 Kb ladder was added to one lane
for estimation of DNA length. Samples were electrophoresed in a Mini-Sub cell GT
tank with an attached PowerPac Mini (Bio-rad) in 1 × Tris-acetate-EDTA for 40
minutes at 120 V and 30–40 mAmps.
3.2.6.3 PCR amplification and direct sequencing of fungal ITS
regions
PCR was performed to amplify the ITS1, 5.8S and ITS2 regions of the extracted
fungal DNA. Table 3.1 lists the primers used in the PCR (White et al., 1990) and
Figure 3.2 shows the location of the two primers on nuclear ribosomal DNA. The
ITS1 forward primer (ITS1 fwdpr) locates upstream (5’) from the ITS1 region and
the ITS4 reverse primer (ITS4 revpr) locates immediately downstream (3’) from the
ITS2 region (White et al., 1990).

Chapter 3: Isolation and Identification of Coprophilous Fungi
38
Table 3.1 PCR primers
Primer Sequence (5’–3’)
ITS1 TCC GAT GGT GAA CCT GCG G
ITS4 TCC TCC GCT TAT TGA TAT GC
Figure 3.2 Schematic representation of the ITS regions of fungal DNA and the binding sites of the ITS1 forward primer (ITS1 fwdpr) and ITS4 reverse primer (ITS revpr). Note: PCR amplification using these two primers results in an amplicon spanning the 5.8S rDNA, ITS regions 1 and 2 and immediately adjacent areas of the 18S and 28S rDNA.
Each 50 μL reaction contained 25 μL of GoTaq Green Master Mix (Promega, NSW,
Australia), 10 µmol of each primer (Sigma, NSW, Australia), 100 ng of DNA
template and 21 μL of nuclease-free water. PCR was performed in a MyCycler™
thermal cycler (Bio-Rad, Hercules, USA) as described in Table 3.2.
Amplified DNA was purified using the centrifugation procedure of the Wizard SV
Gel and PCR Clean-up System kit (Promega, Madison, USA) in accordance with the
manufacturer’s protocol. Purified samples were visualised via electrophoresis to
assess the quality and quantity for preparation of samples for direct sequencing.
This image cannot currently be displayed.
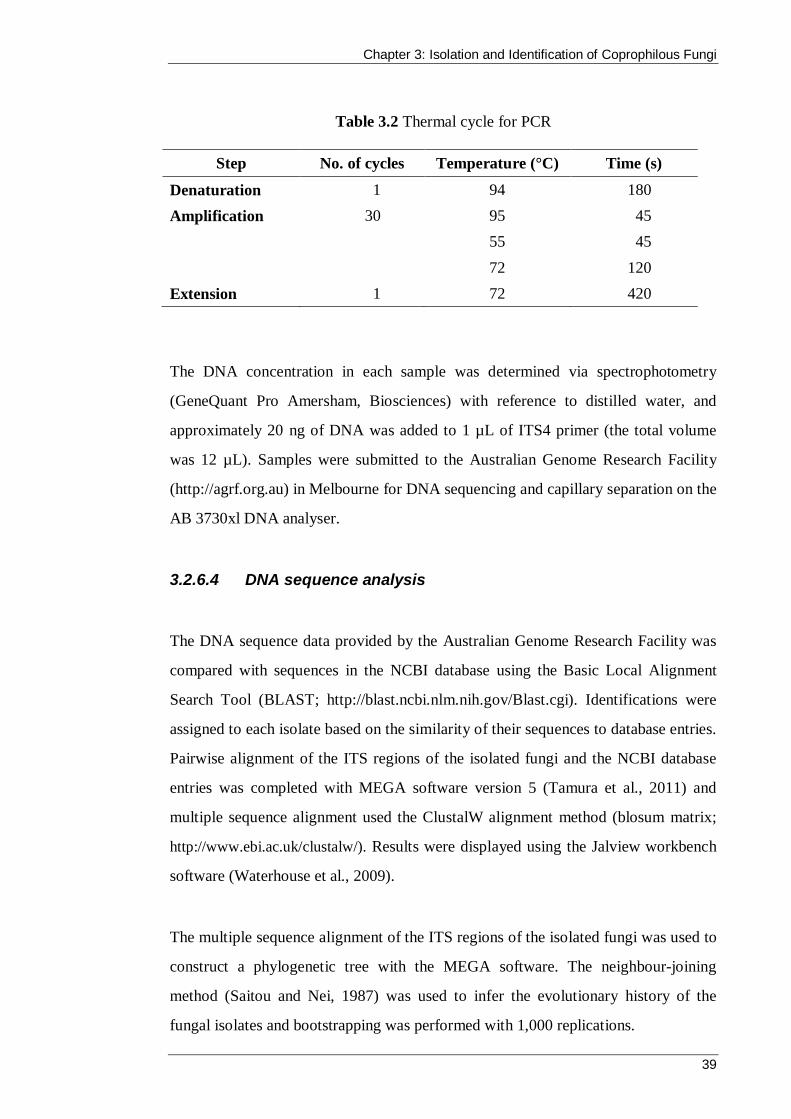
Chapter 3: Isolation and Identification of Coprophilous Fungi
39
Table 3.2 Thermal cycle for PCR
Step No. of cycles Temperature (°C) Time (s)
Denaturation 1 94 180 Amplification 30 95 45 55 45
72 120 Extension 1 72 420
The DNA concentration in each sample was determined via spectrophotometry
(GeneQuant Pro Amersham, Biosciences) with reference to distilled water, and
approximately 20 ng of DNA was added to 1 µL of ITS4 primer (the total volume
was 12 µL). Samples were submitted to the Australian Genome Research Facility
(http://agrf.org.au) in Melbourne for DNA sequencing and capillary separation on the
AB 3730xl DNA analyser.
3.2.6.4 DNA sequence analysis
The DNA sequence data provided by the Australian Genome Research Facility was
compared with sequences in the NCBI database using the Basic Local Alignment
Search Tool (BLAST; http://blast.ncbi.nlm.nih.gov/Blast.cgi). Identifications were
assigned to each isolate based on the similarity of their sequences to database entries.
Pairwise alignment of the ITS regions of the isolated fungi and the NCBI database
entries was completed with MEGA software version 5 (Tamura et al., 2011) and
multiple sequence alignment used the ClustalW alignment method (blosum matrix;
http://www.ebi.ac.uk/clustalw/). Results were displayed using the Jalview workbench
software (Waterhouse et al., 2009).
The multiple sequence alignment of the ITS regions of the isolated fungi was used to
construct a phylogenetic tree with the MEGA software. The neighbour-joining
method (Saitou and Nei, 1987) was used to infer the evolutionary history of the
fungal isolates and bootstrapping was performed with 1,000 replications.

Chapter 3: Isolation and Identification of Coprophilous Fungi
40
3.3 Results and discussion
3.3.1 Collection of koala faeces
Observations of the faecal pellets were consistent across the collection with lengths
ranging from 11 mm to 29 mm, medium to dark brown colour, pleasant eucalypt-like
scent, very dry and dense consistency, visible undigested fibre, and in some of the
samples, the presence of a small amount of animal fur. Figure 3.3 shows a
representative faecal pellet from the French Island collection.
Figure 3.3 A koala faecal pellet (a) intact and (b) cut in half, showing the dense and dry interior.
There were no discernible differences in the appearance of the faeces from koalas
residing in French Island and the faeces from koalas in Boho South. Individual faecal
pellets were chosen at random to be incubated for the isolation of fungi.
3.3.2 Incubation of koala faeces
The koala faeces were incubated using two techniques: Faeces-in-agar (water agar
and PDA) and ethanol-faeces-agar (pour and spread) plates. The faeces-in-agar
method required the faecal pellet to be sliced in half so that one half could be
a) b) 1 cm 1 cm

Chapter 3: Isolation and Identification of Coprophilous Fungi
41
incubated in the presence of water agar and the other with PDA. However, due to the
very dry and densely packed nature of the faecal pellets, it was difficult to cut them
evenly in half. Therefore, some plates had one larger piece while other plates
contained smaller segments (see Figure 3.4b). It is possible that the degree of
fragmentation of the sample may have affected the quantity or variety of fungi
isolates because it changed the internal surface area of the faeces that was in contact
with the agar.
Faecal fragment size was more consistent on the ethanol-faeces-agar plates because
the samples were first cut into small pieces and then shaken in ethanol. Ethanol was
included because it is a recognised method for reducing the growth of rapidly
growing zygomycetous and ascomycetous species, and to maximise the isolation of
ascomycetous and basidiomycetous species that usually appear later in the
succession (Krug et al., 2004; Peterson et al., 2009; Warcup, 1950). No
zygomycetous species were isolated from the koala faeces, which could be due to the
ethanol or because they prefer fresh dung as a substrate and may have been affected
by the drying period (Bell, 2005).
In some studies, antibiotics such as chloramphenicol and ampicillin are included in
the PDA to reduce or eliminate bacterial competition (Krug, 2004; Peterson et al.,
2009). Antibiotics were not used in this study, and no bacterial growth was observed
at any point during incubation of the koala faeces. Although there was no bacterial
competition, some of the faeces-in-agar plates resulted in the dominance of one
fungal species, which made isolation of emerging species difficult. One such fungus,
later identified as Sordaria alcina, belongs to an almost exclusively coprophilous
genus and was identified on six occasions in this project. Many of the fungi
belonging to the order Sordariales (although not exclusively) are capable of secreting
antifungal compounds such as sordarin and its derivatives. This may have been
produced during incubation in this study and inhibited the growth of competing
fungal species (Bell, 2005; Bills et al., 2009; Hauser and Sigg, 1971b), as shown in
Figure 3.4a.
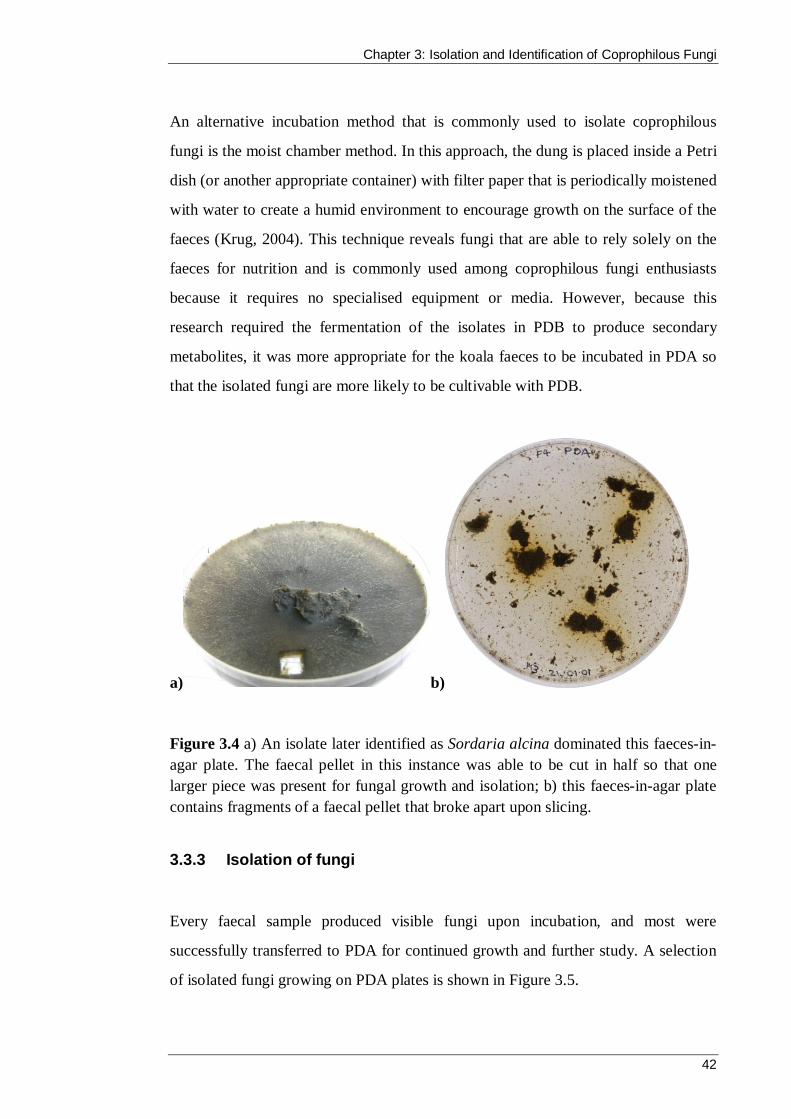
Chapter 3: Isolation and Identification of Coprophilous Fungi
42
An alternative incubation method that is commonly used to isolate coprophilous
fungi is the moist chamber method. In this approach, the dung is placed inside a Petri
dish (or another appropriate container) with filter paper that is periodically moistened
with water to create a humid environment to encourage growth on the surface of the
faeces (Krug, 2004). This technique reveals fungi that are able to rely solely on the
faeces for nutrition and is commonly used among coprophilous fungi enthusiasts
because it requires no specialised equipment or media. However, because this
research required the fermentation of the isolates in PDB to produce secondary
metabolites, it was more appropriate for the koala faeces to be incubated in PDA so
that the isolated fungi are more likely to be cultivable with PDB.
a) b)
Figure 3.4 a) An isolate later identified as Sordaria alcina dominated this faeces-in-agar plate. The faecal pellet in this instance was able to be cut in half so that one larger piece was present for fungal growth and isolation; b) this faeces-in-agar plate contains fragments of a faecal pellet that broke apart upon slicing.
3.3.3 Isolation of fungi
Every faecal sample produced visible fungi upon incubation, and most were
successfully transferred to PDA for continued growth and further study. A selection
of isolated fungi growing on PDA plates is shown in Figure 3.5.

Chapter 3: Isolation and Identification of Coprophilous Fungi
43
Some isolates did not survive subculturing beyond the original faecal agar plate,
which suggests the presence of symbiotic relationships among the mixed culture or
dependence by the isolate on the presence of the koala faeces. Some isolates that
survived subculturing exhibited difficulty in producing confluent growth. In these
cases, PDA may not have been the ideal nutrient medium and 25 °C may not have
been the optimum temperature for incubation.
Figure 3.5 Images of a selection of fungi isolated from koala faeces, growing on PDA plates.
It is likely that there were fungal species present in the faeces that were unable to be
cultured under laboratory conditions. These species may be identified by sequencing
the DNA extracted directly from the faeces using a method that is usually employed
when the objective of the research is to report on the distribution and diversity of
specific fungi taxa within a substratum (Anderson and Cairney, 2004; Bastias et al.,
2006; Herrera et al., 2011). For example, researchers in Australia have used this

Chapter 3: Isolation and Identification of Coprophilous Fungi
44
technique by sampling DNA from soil to study the effects of fire on fungal
communities in eucalypt forests in New South Wales and Victoria (Chen and
Cairney, 2002).
This study is not an exhaustive survey of the coprophilous fungal community found
in koala faeces. The fungi that were isolated were those that survived the drying
process, were able to undergo sporulation or spore germination under the prescribed
incubation method, and could be grown on PDA. In addition, these isolates were not
significantly restricted by antagonistic behaviour of competing fungi, such as the
production of diffusible antibiotics or hyphal interference (Wicklow, 1981).
The collection of fungi in this research represents an unknown fraction of the species
present in a collection of faeces from 12 individual koalas living in two locations at
one time of the year. Given these considerations, an expanded survey of the
coprophilous fungi population could address experimental variables, including
adjustment of the drying time for faeces, changes to incubation temperature, the
introduction of light and dark cycles, and the use of a different medium such as malt
extract agar. In terms of variables relating to the collection of faeces, some studies
have suggested that season and location are likely to affect the diversity of the
coprophilous fungal community (Krug et al., 2004; Kruys and Ericson, 2008; Melo et
al., 2012; Richardson, 2001c). Therefore, although time intensive, a more
comprehensive survey would include many faecal pellets collected at different times
of the year from numerous koalas residing in various locations. This approach may
identify fungal species that were not found in this research and therefore increase the
opportunities for discovery of novel bioactive compounds produced as secondary
metabolites.
3.3.4 Identification of fungi by ITS sequencing
Genomic DNA was extracted from the isolated fungi using the Zymo Fungal DNA
MiniPrep after the isolates had been stored as agar slices at −80 °C. The

Chapter 3: Isolation and Identification of Coprophilous Fungi
45
manufacturer’s protocol suggests that fungal cells be added in the form of a
suspension in water or isotonic buffer; however, the addition of frozen mycelia on a
very thin slice of agar directly to the lysis solution did not appear to negatively affect
the extraction and sufficient DNA yields were achieved from all isolates.
Amplification of the ITS regions of the fungal isolates resulted in PCR products
ranging from 513–603 base pairs in length. Figure 3.6 shows the gel electrophoresis
image of PCR products from 20 isolates that were later identified as belonging to the
phyla Ascomycota.
Figure 3.6 Agarose gel electrophoresis of PCR products following the amplification of the ITS regions of rDNA of a selection of fungi isolated from koala faeces. Note: 5 µl of DNA was loaded per lane. Lane M is the 1kb DNA ladder; lanes 1–20 are samples: F14P1, S5P5, S4P1, F14W1, F3P1, F13P1, S4P2, F14W2, F3W3, S5P3, S3P1, S3P2, S3P3, F11EP1, F16EP2, F16EP1, F16ES1, F4P1, F2P1 and F2P2.
The PCR products were sequenced at the Australian Genome Research Facility and
resulted in DNA sequences spanning the 5.8S rDNA, ITS regions 1 and 2, and the
adjacent 18S and 28S rDNA. The sequence data chromatogram for each isolate was
analysed with particular attention paid to the beginning and end of the sequences
where peaks can be of poor resolution. Sequences were edited at residues where the
peaks were incongruent with nucleotide allocation (see Figure 3.7). These sequences
were searched against the NCBI database using BLAST and identifications were
considered based on their similarity to sequences in the database (see Table 3.3).

Chapter 3: Isolation and Identification of Coprophilous Fungi
46
Pairwise alignment of the ITS regions of the isolated fungi and their NCBI database
matches revealed points of difference between the sequences (see Figure 3.8).
Figure 3.7 A section of the DNA sequence chromatogram showing the ITS2 region and adjacent 28S rDNA of the isolate S4P4, KT071741, Trichoderma pseudokoningii. Note: Comparison of the chromatogram with a pairwise alignment of the sequence with its top BLAST match (X93970) highlighted two residues (cytosine, blue and adenosine, green) that were apparent in the peaks but not the sequence. The sequence was edited at these two points. Sequence chromatograms from the Australian Genome Research Facility show reverse complement sequences as the reverse primer, ITS4 was used for sequencing.
The ITS sequences were submitted to GenBank (www.ncbi.nlm.nih.gov/
genbank/) and were assigned the accession numbers KT071713–KT071744
(KT071743 has been omitted). To date, this submission was only the second
collection of ITS sequences from fungi isolated from koala faeces present in the
Genbank database. The sequences form the GenBank PopSet (population sequence
data) with the identifier 914704626. All of the isolates returned sequence matches of
at least 95% (E = 0) with known fungal species, and 19 of the 31 isolates were
identified with matches of at least 99%. Identification based on comparison with
GenBank accessions requires that other users submit accurate sequencing results
with correct assignment of taxonomic groups (Bellemain et al., 2010). Moreover, the
non-coding ITS regions are subject to relatively fast evolutionary rates that may lead

Chapter 3: Isolation and Identification of Coprophilous Fungi
47
to difficulties in achieving perfect sequence alignment at high taxonomic levels
(Huang et al., 2009). The GenBank database has numerous environmental isolates
that are listed as unidentified fungi or have not been identified at the species level,
which can further impede identification (McMullan-Fisher et al., 2011). For
example, isolate F5A4 (HQ406811) was identified as Penicillium spinulosum in this
work, but it also shared maximum sequence homology with an accession listed as
“Unverified – Uncultured Penicillium clone” (HQ211519) in the database (Deslippe
et al., 2012).
When research such as this requires the comparison of sample ITS sequences with
GenBank accessions, there is no agreed upon percentage of sequence similarity that
definitively indicates that the two sequences represent the same species (Bruns et al.,
2007; Hughes et al., 2009). However, a number of studies have used ≤3% ITS
sequence divergence to indicate conspecificity in fungi (Morris et al., 2008; O'Brian
et al., 2005; Ryberg et al., 2008; Smith et al., 2007; Walker et al., 2008). All but two
of the fungi isolated from the koala faeces (S4W1, Neurospora intermedia,
KT071742, 96% and F16EP1, Anthostomella leucospermi, KT071725, 95%; Table
3.3) meet this cut-off. An alternative method which may provide a more robust
identification at the species level is multilocus sequence typing (MLST). MLST
measures the variation in DNA sequences of a number of genes such as those
encoding beta-tubulin and calmodulin (Hong et al., 2014; Perrone et al., 2014).
Almost all of the fungi isolated from the koala faeces were identified as species
belonging to the phylum Ascomycota. One isolate (F3W1, KT071734) identified as
Bjerkandera adusta is from the phylum Basidiomycota. No members of the phylum
Zygomycota were isolated. The prevalence of ascomycetous species in the collection
of fungi is consistent with previous studies of coprophilous fungi, in which members
of the phylum Ascomycota have dominated the dung mycota (Bell, 2005; Doveri et
al., 2013; Gupta, 2010; Melo et al., 2012; Pandey, 2009; Peterson et al., 2009;
Richardson, 2001a, c; Watling and Richardson, 2010).

Chapter 3: Isolation and Identification of Coprophilous Fungi
48
Figure 3.8 Pairwise alignment of the ITS1 and ITS2 regions and adjacent rDNA of the isolate S4P4, KT071741, Trichoderma pseudokoningii, isolated from koala faeces to that of the sequence with the greatest similarity in the NCBI database (at the time of analysis) with the accession number X93970. Note: The two edited residues (Figure 3.7) are indicated in yellow and the one discrepancy between the two sequences appears in red.
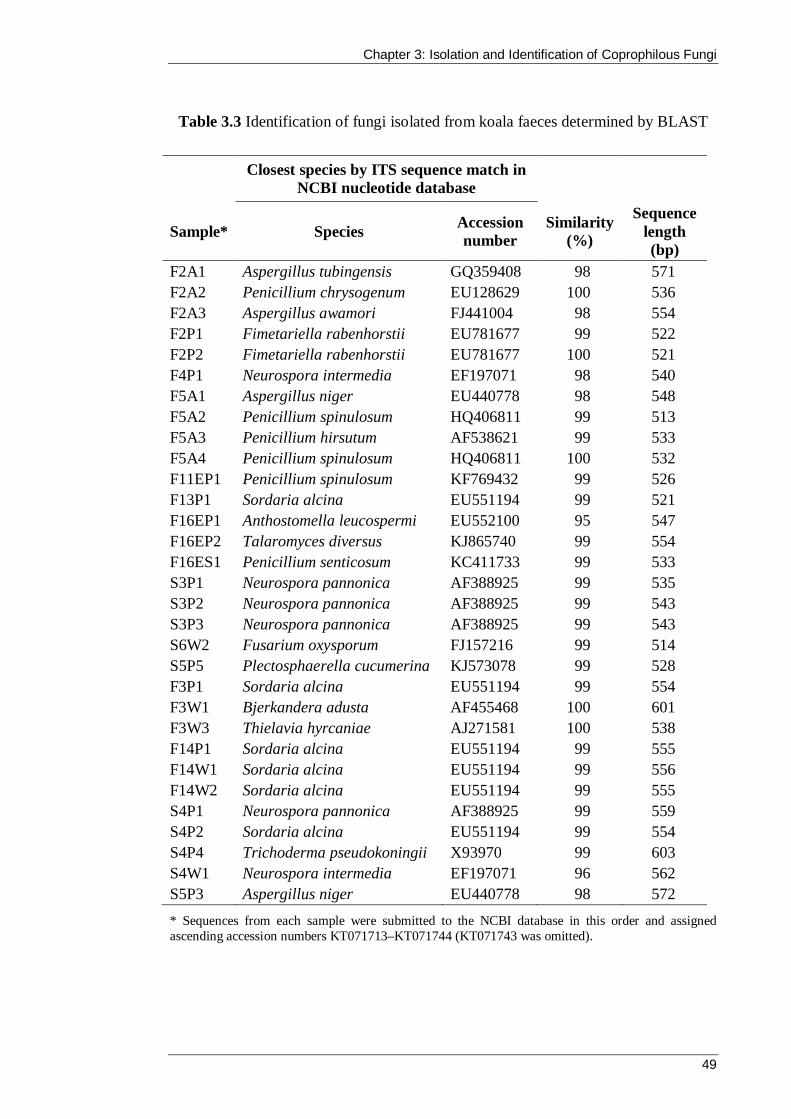
Chapter 3: Isolation and Identification of Coprophilous Fungi
49
Table 3.3 Identification of fungi isolated from koala faeces determined by BLAST
* Sequences from each sample were submitted to the NCBI database in this order and assigned ascending accession numbers KT071713–KT071744 (KT071743 was omitted).
Closest species by ITS sequence match in NCBI nucleotide database
Sample* Species Accession number
Similarity (%)
Sequence length (bp)
F2A1 Aspergillus tubingensis GQ359408 98 571 F2A2 Penicillium chrysogenum EU128629 100 536 F2A3 Aspergillus awamori FJ441004 98 554 F2P1 Fimetariella rabenhorstii EU781677 99 522 F2P2 Fimetariella rabenhorstii EU781677 100 521 F4P1 Neurospora intermedia EF197071 98 540 F5A1 Aspergillus niger EU440778 98 548 F5A2 Penicillium spinulosum HQ406811 99 513 F5A3 Penicillium hirsutum AF538621 99 533 F5A4 Penicillium spinulosum HQ406811 100 532 F11EP1 Penicillium spinulosum KF769432 99 526 F13P1 Sordaria alcina EU551194 99 521 F16EP1 Anthostomella leucospermi EU552100 95 547 F16EP2 Talaromyces diversus KJ865740 99 554 F16ES1 Penicillium senticosum KC411733 99 533 S3P1 Neurospora pannonica AF388925 99 535 S3P2 Neurospora pannonica AF388925 99 543 S3P3 Neurospora pannonica AF388925 99 543 S6W2 Fusarium oxysporum FJ157216 99 514 S5P5 Plectosphaerella cucumerina KJ573078 99 528 F3P1 Sordaria alcina EU551194 99 554 F3W1 Bjerkandera adusta AF455468 100 601 F3W3 Thielavia hyrcaniae AJ271581 100 538 F14P1 Sordaria alcina EU551194 99 555 F14W1 Sordaria alcina EU551194 99 556 F14W2 Sordaria alcina EU551194 99 555 S4P1 Neurospora pannonica AF388925 99 559 S4P2 Sordaria alcina EU551194 99 554 S4P4 Trichoderma pseudokoningii X93970 99 603 S4W1 Neurospora intermedia EF197071 96 562 S5P3 Aspergillus niger EU440778 98 572

Chapter 3: Isolation and Identification of Coprophilous Fungi
50
A number of the genera are recorded in the literature as being found on koala faeces;
Sordaria, Aspergillus and Penicillium species were identified by morphology only
(Bell, 2005; Cribb, 1997), and Aspergillus, Penicillium, Sordaria, Fusarium,
Neurospora and Trichoderma were identified by DNA sequencing in the formative
research by Peterson et al. (2009). Some of the Ascomycetes isolated in this work
have not been definitively recorded in the literature as having been observed on koala
faeces. However, it would be rash to assume that this is the first occasion of their
discovery on koala faeces. The majority of morphological identifications that have
been made during dung surveys in Australia have not been submitted to peer-
reviewed journals. Rather, the samples were often collected by keen naturalists and
identifications, if published, were limited to club newsletters and the like.
One survey that was more systematic in nature is the illustrated guide compiled by
Ann Bell (2005), in which she identified 176 species of Ascomycetes from
approximately 300 dung samples (including koala) from Australia that were
collected mostly by volunteers. Bell also makes reference to two other mycologists
who studied coprophilous fungi in Australia, Dade and Cribb, the former identifying
both Sordaria and Fimetariella species on dung collected in Victoria, Australia. Of
the remaining genera isolated and identified in this research, the Ascomycetes
Thielavia and Anthostomella and the Basidiomycete Bjerkandera are known to occur
as coprophilous fungi (Baker et al., 2013; Misra et al., 2014; Reina et al., 2013). The
genus Neurospora is closely related to Sordaria; however, while the latter contains
predominantly coprophilous species, the majority of species within the sister genus
Neurospora are found primarily on other substrates such as soil (Krug et al., 2004;
Raju, 2002). Finally, several Plectosphaerella species are pathogens of plant species
in Australia, causing root rot (Jahromi, 2007; Tesoriero, 2014), but to the best of my
knowledge, this genus has not been reported to occur on faeces in Australia.

Chapter 3: Isolation and Identification of Coprophilous Fungi
51
3.3.5 Comparison of the ITS regions of the fungal isolates
A multiple sequence alignment of the ITS regions of the fungal isolates was created
using ClustalW to explore the number and location of differences in nucleotide bases
between selected isolates. Jalview software produced a visualisation of the analysis
including the construction of the consensus graph shown in Figure 3.9.
The ITS regions, as expected, demonstrated a higher degree of difference between
the isolates compared with the highly conserved sequences in the adjacent 18S, 5.8S
and 28S rDNA regions. In terms of the ITS regions, similarities were most common
between isolates of the same genus (e.g., Neurospora intermedia, F4P1, KT071718
and Neurospora pannonica, S3P1, KT071728; Table 3.3 and Figure 3.9), and then
between isolates belonging to the same phylum (N. intermedia, F4P1, KT071718;
N. pannonica, S3P1, KT071728; Trichoderma pseudokoningii, S4P4, KT071741; all
Ascomycetes; Figure 3.9). In comparison, further differences can be seen between
the three representative Ascomycete isolates and the isolate Bjerkandera adusta
(F3W1, KT071734; Figure 3.9) from the phylum Basidiomycota. Insertions in the
ITS regions of the T. pseudokoningii isolate accounted for its sequence length being
larger than that of the other Ascomycetes (Table 3.3, final column). B. adusta also
contained insertions, notably within the mostly conserved 5.8S rDNA region,
accounting for its second largest sequence length.
The multiple sequence alignment of the ITS regions of the fungi was used to
construct a phylogenetic tree with MEGA software version 5. The neighbour-joining
method with bootstrap re-sampling performed over 1,000 replicates was used to infer
evolutionary history of the fungal isolates. As shown in Figure 3.10, this method
placed the thirty Ascomycete isolates into five different orders (Sordariales,
Hypocreales, Incertae sedis, Xylariales and Eurotiales), all belonging to the
Sordariomycetes class; the single Basidiomycete isolate, B. adusta, is shown on a
separate branch.

Chapter 3: Isolation and Identification of Coprophilous Fungi
52
Figu
re 3
.9 Ja
lvie
w re
pres
enta
tion
of C
lusta
lW m
ultip
le a
lignm
ent o
f ITS
1, 5
.8S
rDN
A, I
TS2
and
adja
cent
18S
rDN
A a
nd 2
8S rD
NA
of
four
fung
i iso
late
d fr
om k
oala
faec
es.
Neur
ospo
ra i
nter
med
ia (
F4P1
) K
T071
718,
Neu
rosp
ora
pann
onic
a (S
3P1)
KT0
7172
8 an
d Tr
icho
derm
a ps
eudo
koni
ngii
(S4P
4)
KT0
7174
1 fr
om th
e ph
ylum
Asc
omyc
ota;
Bje
rkan
dera
adu
sta (F
3W1)
KT0
7173
4, a
repr
esen
tativ
e of
the
Basid
iom
ycet
e ph
ylum
. The
bl
ack
bar g
raph
repr
esen
ts th
e pe
rcen
tage
of b
ases
mat
chin
g th
e co
nsen
sus s
eque
nce
at e
ach
resid
ue.

Chapter 3: Isolation and Identification of Coprophilous Fungi
53
Figure 3.10 Phylogenetic position of fungi isolated from koala faeces. Note: The evolutionary history was inferred using neighbour-joining method (Saitou and Nei, 1987) and the percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (≥50%, 1,000 replicates) is shown next to the branches (Felsenstein, 1985). Five orders within the Sordariomycetes class are annotated and the one Basidiomycete isolate is positioned at the lowest branch.

Chapter 3: Isolation and Identification of Coprophilous Fungi
54
3.4 Summary
Coprophilous fungi are an important part of the Australian ecosystem, but there is
little published data describing these communities compared with their Northern
hemisphere counterparts. In this chapter, a collection of coprophilous fungi were
isolated from koala faeces and identified via ITS sequencing. All but one of the 31
isolates were members of the phylum Ascomycota, a weighted diversity that is
common in numerous Ascomyceteous dominated coprophilous collections in the
literature. This work did not reveal any previously unknown species, but the findings
are of interest because they add to the very small amount of research into Australian
coprophilous fungi. The isolates were assigned accession numbers KT071713–
KT071744 and lodged with the NCBI database, where they represent only the second
set of DNA sequences of coprophilous fungi isolated from koala faeces.
Coprophilous fungi have shown potential as a source of antimicrobial compounds in
the literature, and this bioactivity will be explored in the following chapter. Chapter
Four describes the preparation of extracts from liquid fermentations of isolated
coprophilous fungi and the screening of these extracts against selected bacteria and
fungi.

Coprophilous Fungi from Koala Faeces: A Novel Source of Antimicrobial Compounds
55
CHAPTER 4
Screening of Coprophilous Fungi for Antimicrobial Activity

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
56
4.1 Introduction
Natural products and their derivatives have long been exploited for their
pharmaceutical potential, and it is estimated that more than 70% of the antimicrobial
drugs currently in clinical use are derived from natural products (Qadri et al., 2014).
However, new drug approvals are declining and it has been suggested that a
discovery void exists (Li and Vederas, 2009). Given the increased resistance of
pathogens to current drugs and the rising incidence of opportunistic infections in
immune-compromised individuals, it is essential that previously untapped biological
niches are explored for novel microbes and natural products. An alternative system
of bacterial control is through the disruption of QS signals. An ideal quenching
compound would reduce the virulence of, rather than kill, the target bacteria, and in
turn, may reduce the development of resistant strains (Hentzer and Givskov, 2003).
Phytopathogenic fungi are becoming increasingly problematic for important food
crops, particularly in developing countries (Vurro et al., 2010). These plant diseases
cause a reduced harvest yield (or post-harvest rot), which has serious implications in
terms of food security and a sustainable economy (Fears et al., 2014). Resistance to
current agrochemicals is an ongoing challenge, and concerns relating to the safety
and environmental impact of agrochemicals have led to a more stringent regulatory
process (Lamberth et al., 2013). Thus, there is now a demand for new safer and more
selective compounds, and it is important that natural products continue to be studied
for their potential application in crop protection (Dayan et al., 2009; Olufolaji, 2010).
Fungi have provided numerous chemically and biologically diverse secondary
metabolites that have proved immensely useful in medical, agricultural and industrial
settings. Coprophilous fungi remain a relatively untapped source compared with
fungi from other substrata and biological niches. However, coprophilous fungi are
prime candidates for discovery of antimicrobials due to their ubiquity and their
dominance in a highly competitive environment. The limited research to date has

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
57
revealed a number of bioactive secondary metabolites and further reinforces their
potential for antibiotic discovery (Bills et al., 2013).
Australian coprophilous fungi have yet to be explored in terms of their potential to
produce antimicrobial secondary metabolites. This chapter presents, for the first time,
the screening of coprophilous fungi from koala faeces for antibacterial, antifungal
and anti-QS activity.
4.1.1 Chapter aims
The experiments described in this chapter were designed to evaluate the bioactivity
of coprophilous fungi from koala faeces. More specifically, this work aimed to assess
the antimicrobial potential of their liquid cultivated extracts against:
i. the Gram-positive bacterium S. aureus
ii. three Gram-negative bacteria: E. coli, P. aeruginosa and K. pneumoniae
iii. the filamentous fungus D. brizae
iv. the yeast C. albicans.
In addition, the research sought to establish whether the extracts were capable of
anti-QS activity using the indicator bacterium C. violaceum.
4.2 Materials and methods
4.2.1 Bacterial cultures
Antibacterial testing was performed against S. aureus (ATCC 25923), E. coli (ATCC
25922), P. aeruginosa (ATCC 9721) and K. pneumoniae (Australian Collection of
Microorganisms: accession no. 90), which were part of the culture collection at
Swinburne University of Technology, Victoria, Australia. Anti-QS activity was
tested using the Gram-negative indicator strain C. violaceum (University of New
South Wales collection: accession no. 040100).

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
58
Agar slants (Section 3.2.5) were prepared for medium-term storage of bacterial
cultures. Overnight cultures were subcultured onto Mueller–Hinton agar (MHA;
Oxoid, Basingstoke, UK) slants and incubated at 37 °C for 24 hours prior to storage
at 4 °C for up to six months. Recovery of bacteria was achieved by subculture to
Mueller–Hinton broth (MHB) and incubation for 18 hours at 37 °C. Storage and
maintenance of C. violaceum used Luria–Bertani agar (Merck, Germany) and Luria–
Bertani broth. For long-term storage of cultures, the Protect Bead storage system
(Technical Service Consultants, Lancashire, UK) was used to enable frozen storage
at −80 °C for several years.
The preparation of a standardised bacterial inoculation was required for the
antibacterial and anti-QS assays. One colony was removed from a plate culture and
then suspended in 2 mL of the appropriate liquid medium and incubated at 37 °C
(30 °C for C. violaceum) for approximately 24 hours with shaking (at 220 rpm).
Following incubation, the turbidity of the solution was adjusted with sterile saline to
visually match a 0.5 McFarland standard, thus producing an inoculum with an
approximate cell density of 1.5 × 108 colony forming units (cfu)/mL that could be
further diluted if required.
4.2.2 Fungal cultures
Antifungal testing was performed against one filamentous phytopathogenic fungi,
D. brizae (Phillips et al., 2011), and one yeast strain, C. albicans (FRR5580), which
were both part of the culture collection at Swinburne University. D. brizae and
C. albicans were both maintained on PDA and PDB with incubation at 25 °C for 7
days and 30 °C for 18 hours, respectively. Long-term storage of C. albicans used the
Protect Bead system, whereas glycerol stocks were prepared for D. brizae (Section
3.2.5).
A standardised fungal inoculum of C. albicans was prepared following the protocol
used for bacteria (Section 4.2.1) with the use of PDA and PDB media and incubation

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
59
at 30 °C. A 0.5 McFarland standard equates to 1–5 × 106 yeast cells/mL (Espinel-
Ingroff and Canton, 2007d). Standardised suspensions of D. brizae were created
from cultures that were grown in PDB for 7 days at 25 °C with shaking (at 180 rpm).
Mycelia were vortexed vigorously and repeatedly drawn and aspirated through a 25
gauge needle (Terumo Corp., Macquarie Park, Australia) to separate cells. The
resulting solution was adjusted to a concentration of 0.4–5 × 106 mycelia
fragments/mL (OD600 = 0.15–0.17) (Espinel-Ingroff and Canton, 2007a).
4.2.3 Fungal fermentation for preparation of crude extracts
Extracts were produced from liquid fermentations of the isolated coprophilous fungi
described in Chapter Three. For each isolate, a 1 cm2 agar plug of fungi was placed
in an Erlenmeyer flask containing 100 mL of PDB. The flask was stoppered with
cotton wool and incubated at 25 °C in the dark with shaking at 150 rpm for 22 ± 2
days. Following incubation, the contents were filtered through Whatman (No. 1)
filter paper under vacuum to separate the bulk of the mycelium from the fermentation
liquor. The remaining fungal cells were removed from the liquid by membrane
filtration through a 0.22 μm filter (Merck Millipore, MA, USA) under vacuum. This
resulted in approximately 100 mL of sterile liquor and the separated mycelial
biomass for each sample.
4.2.3.1 Mycelia extraction
Mycelial biomass that had been separated from the fermentation broth was
transferred to weighed sterile round-bottom flasks and frozen at −80 °C in
preparation for lyophilisation. Following freeze-drying in a Cryodos-50 (Telstar,
Spain), the mycelia were weighed and resuspended in 100% ethanol to a
concentration of 100 mg/mL. Extracts were stored at 4 °C until use. When the
mycelial extracts were tested for antimicrobial activity, the control consisted of an
equivalent volume of ethanol.

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
60
4.2.3.2 Liquor extraction
The filtered liquor was extracted twice with equal volumes of ethyl acetate (EtOAc,
Merck) in a separating flask for a total of 22 hours. The organic phases were
combined and dried with anhydrous sodium sulphate Sigma), evaporated to
approximately 2 mL on a rotary evaporator at 45 °C (Buchi, Switzerland) and then
transferred to a microcentrifuge tube and evaporated to dryness in a micro-rotary
evaporator (John Morris Scientific, Chatswood, Australia) with centrifugation at
40 °C. The dried extracts were then resuspended in methanol (Merck) to a
concentration of 100 mg/mL and stored at 4 °C until use (Corrado and Rodrigues,
2004). When the EtOAc extracts were tested for antimicrobial activity, the control
consisted of equivalent volumes of methanol.
The aqueous phase of the liquid–liquid separation was rotary evaporated to remove
any residual EtOAc; it was then frozen and lyophilized to dryness. The aqueous
extracts were resuspended in MilliQ water to a concentration of 100 mg/mL and
stored at 4 °C until use.
4.2.4 Antibacterial testing
Coprophilous fungal extracts were tested against one Gram-positive bacterium,
(S. aureus) and three Gram-negative bacteria (E. coli, P. aeruginosa and
K. pneumoniae).
4.2.4.1 Plate-hole diffusion
Screening of all extracts was performed using a modification of the plate-hole
diffusion assay described by Rahman et al. (2001). Molten MHA (15 mL, 45 °C) was
inoculated with 200 µL of a bacterial suspension (1.5 × 108 cfu/mL), mixed and then
poured into a sterile petri dish. Once set, a sterile stainless steel cork borer (6 mm
diameter) was used to make wells in the agar and 20 µL of extract was added to each

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
61
well. Control wells contained ethanol for mycelial extracts and methanol for organic
liquor extracts. A chlorhexidine (CHX) gluconate solution (0.5% w/v in 70% v/v
ethanol) was included for comparison (J & J Medical, USA). The plates were kept at
room temperature for 30 minutes prior to incubation to allow for absorption of the
extracts and controls. After incubation for 18 hours at 37 °C, the zones of inhibited
growth were measured to the nearest millimetre (inclusive of well diameter) and the
average of triplicate results was recorded.
4.2.4.2 Microdilution assay
Minimum inhibitory concentrations (MIC) were determined for the fungal extracts
that produced a zone of inhibited bacterial growth that was greater than 8 mm in the
plate-hole diffusion assays. A microdilution method was employed in accordance
with a modification of the Clinical Laboratory Standards Institute document, M7-A7
(CLSI, 2006). In sterile round-bottom microtitre plates (Corning, USA), 150 µL of
MHB was added to each of the test wells and 50 µL of each extract (5 mg) was
added to the wells in the first column. The contents were mixed via gentle aspiration
with a micropipette and a two-fold serial dilution was prepared by sequentially
transferring 100 µL aliquots (with mixing) across each row. Bacterial inocula were
prepared by diluting a 0.5 McFarland standardised suspension 1:200 in MHB to a
final concentration of 7.5 × 105 cfu/mL. One hundred microlitres of the suspension
was added to each of the test wells, producing an extract concentration range of
12,500–98 µg/mL. Each extract was tested in triplicate against each bacterium. A
medium sterility control well contained only MHB, a growth control well contained
bacteria in the presence of MHB only, and a positive control comprised two-fold
dilutions of 0.5% CHX in addition to the bacterial inoculum and MHB. The plates
were incubated overnight at 37 °C. After incubation, the wells were inspected for
turbidity and the MIC for each extract was determined as the lowest concentration
that inhibited visible growth.

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
62
Minimum bactericidal concentrations (MBC) were also deduced via a growth test
(Qaiyumi, 2007) by sub-culturing a 100 µL aliquot from the microtitre wells that had
no visible growth onto MHA. Following incubation, the MBC was defined as the
lowest concentration of the extract that resulted in zero growth on the MHA plate.
4.2.4.3 Time-course growth assay
To visualise the antibacterial activity of the coprophilous fungal extracts over 24
hours, a modification of the time-course assay described by Verma (2007) was used.
Two extracts were selected for the assay based on their performance in the plate-hole
diffusion and MIC assays. Inocula of E. coli and S. aureus were prepared by diluting
a 0.5 McFarland standardised suspension 1:100 in MHB to a final concentration of
1.5 × 106 cfu/mL. For each bacterium, 1 mL of inoculum was added to six sterile
tubes containing 4 mL of MHB. Fungal extract was then added to two tubes
(duplicate) to a final concentration of 1 × MIC and to two tubes (duplicate) at a final
concentration of 2 × MIC. The final two tubes served as duplicate growth controls.
Two aliquots of 100 µL were removed from each tube and spread onto the surface of
two MHA plates. A further two 100 µL aliquots were removed from each tube and
duplicate ten-fold serial dilutions were performed from 10−1 to 10−6; 100 µL aliquots
from each dilution were spread onto MHA plates. These plates represented the
time=0 viable count. The time-kill tubes and all plates were incubated at 37 °C. This
process was repeated at 0.5, 1.5, 1, 3, 6, 12 and 24 hours. Following overnight
incubation of all plates, the viable cell numbers were counted for dilutions containing
30–300 cfu and the number of viable bacteria at each time point was calculated.
Each assay was performed in triplicate and a two-way analysis of variance with the
Bonferroni’s post-hoc correction was performed using GraphPad Prism v.6
(GraphPad, La Jolla, CA) to assess statistical differences across conditions at each
time interval. Values of p < 0.05 were considered statistically significant. Data are
presented as the means ± standard deviation.

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
63
4.2.5 Antifungal testing
Coprophilous fungal extracts were tested against the pathogenic yeast strain C.
albicans and the phytopathogenic fungus D. brizae.
4.2.5.1 C. albicans inhibition
Antifungal activity against C. albicans was assessed using modifications of the two
methods that were used for the antibacterial testing. Plate-hole diffusion assays were
carried out in PDA with the addition of 200 µL of a 1–5 × 106 cells/mL (McFarland
standard 0.5) inoculum, and the plates were incubated at 30 °C (see Section 4.2.4.1).
Extracts that exhibited anti-candidal activity were tested in a microdilution assay to
determine their MIC and growth tested to assign a minimum fungicidal concentration
(MFC). Modifications of this method included the use of PDB, addition of 100 µL of
a 1–5 × 103 cells/mL inoculum to each test well and incubation of the 96-well
microtitre plate at 30 °C (see Section 4.2.4.2).
4.2.5.2 D. brizae inhibition
Four methods were used to evaluate the inhibitory potential of the extracts against
the filamentous fungus D. brizae.
Spore germination assay
To establish if the fungal extracts were able to inhibit spore germination by
D. brizae, bioautographic assays were performed using the method of Quiroga et al.
(2009) with some adjustments. Silica gel aluminium plates (0.2 mm thick, Sigma)
were cut to size and then placed into empty sterile Petri plates and spotted with
15 µL of each extract and controls. After the plates had dried, 12 mL of semi-solid
PDA (0.6% agar) inoculated with 0.3 mL of a D. brizae spore suspension (0.4–
5 × 106 spores/mL) was poured over the surface of the plates. Following incubation

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
64
at 25 °C for 3 days, 100 µL of a 0.25% (w/v) thiazolyl blue tetrazolium bromide
(MTT, Sigma) solution was spread over the surface of the plates and they were
incubated for 1 hour. Plates were observed for the presence of a colourless zone
surrounding an extract, which signified inhibition of spore germination. Each extract
was tested in triplicate and results are presented as the mean clear zone diameter ±
standard deviation.
Radial growth assay
In this antifungal assay, extracts and controls were incorporated into PDA prior to
inoculation with the test fungus D. brizae to assess the effect of the extracts on radial
growth (Boyraz and zcan, 2005; Quiroga et al., 2009; Zhao et al., 2010).
Extracts were mixed evenly with 10 mL of molten (45 °C) PDA to a final
concentration of 5 mg/mL and poured into Petri plates. The agar was allowed to set
and a fungal plug (8 mm diameter) of D. brizae was placed in the centre of the plate.
All plates were incubated at 25 °C until the growth of the control approached the
plate edge. Following incubation, the diameter of growth was measured in two
directions and growth reduction was calculated using the following formula:
퐺푟표푤푡ℎ 푟푒푑푢푐푡푖표푛 (%) =푔푟표푤푡ℎ 푖푛 푐표푛푡푟표푙 − 푔푟표푤푡ℎ 푖푛 푒푥푡푟푎푐푡
푔푟표푤푡ℎ 푖푛 푐표푛푡푟표푙 × 100
Plates containing appropriate solvent (without extracts) were included as controls.
Data are presented as the average growth reduction (%) from triplicate assays ±
standard deviation.
Hyphal extension assay
The ability of the extracts to inhibit hyphal extension of D. brizae was assessed by
disk diffusion assays using the method of Quiroga et al. (2009) with some

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
65
modifications. A fungal plug (8 mm diameter) of D. brizae was placed in the centre
of a PDA plate. Sterile paper disks (Oxoid, 6 mm diameter) were impregnated with
20 µL of each extract. The discs were allowed to dry in a sterile dish and were then
placed on the PDA surface, approximately 2 cm from the fungal plug with a
maximum of 4 disks per plate. Controls included disks impregnated with 20 µL of an
appropriate solvent, including water for the aqueous liquor extracts. Plates were
inverted and incubated in the dark for 5 days at 25 °C. The assay was performed in
triplicate, and any inhibition of fungal growth was visualised as a crescent-shaped
inhibitory zone at the mycelial front adjacent to the disk.
Microdilution assay
Extracts that exhibited positive results in two or more of the antifungal tests were
selected for inclusion in a microdilution assay to determine their MIC against
D. brizae (Espinel-Ingroff and Canton, 2007a).
The assay was performed in 96-well sterile polystyrene plates with low evaporation
lids (Corning). Wells were filled with 200 µL of PDB and then 100 µL of each
extract was added to the first well of each row and a 2-fold serial dilution was carried
out across the row. A mycelial suspension of D. brizae was prepared (see Section
4.2.2) and diluted 1:50 in PDB to a final concentration of 0.8–10 × 104 mycelial
fragments/mL and each well was inoculated with 50 µL of the suspension. The final
concentration of the extracts was within the range of 25–0.78 mg/mL and all extracts
were tested in triplicate. Growth control wells did not contain extract, and wells
containing 0.5% CHX were included as an antifungal control. The wells at the plate
perimeter were filled with 200 µL of MilliQ water to reduce evaporation of the test
wells and the plates were incubated in the dark for 7 days at 25 °C. Following
incubation, the plates were placed on a light box and the wells were inspected for
fungal growth. The MIC was defined as the lowest concentration of extract to
completely inhibit fungal hyphal growth (Espinel-Ingroff and Canton, 2007a).

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
66
4.2.6 Anti-QS testing
For each extract, QS inhibition was tested via the standard antibacterial disk
diffusion assay (Bauer and Kirby, 1966) using the indicator strain C. violaceum
following the method described by Adonizio et al. (2006) with some modifications.
Each extract (20 L) was loaded onto a sterile paper disk (6 mm diameter), allowed
to dry and then placed on the surface of Luria–Bertani agar plates spread with
100 μL (approximately 1.5 × 108 cfu/mL) of C. violaceum culture. Gentamicin disks
(10 µg, Oxoid) were used to compare QS inhibition with an antibiotic effect and
appropriate solvents were loaded onto disks for a negative control. Plates were
incubated at 30 °C for 40 hours. QS inhibition was detected by a halo of colourless
but viable cells around the disk, whereas antibacterial activity, when present,
produced a zone devoid of cells.
Additionally, the ability of cells to recover from the anti-QS activity and regain
production of violacein was established. A small sample of colourless colonies was
removed from around an active extract and then subcultured onto Luria–Bertani agar
and incubated for 40 hours at 30 °C. Following incubation, the plates were inspected
for the presence of purple pigmented growth, which signals QS ability. Assays were
performed in triplicate and measurements (to the nearest half millimetre) were taken
from the edge of the disk to the edge of the halo.
4.3 Results and discussion
4.3.1 Production of mycelial extracts
Following incubation of the coprophilous fungi and separation of the mycelia from
the fermentation liquor, the mycelia were freeze dried and dissolved in ethanol to
produce 100 mg/mL mycelial extracts.

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
67
Mycelial extracts were produced in this manner from four fungi (F2P1, F13P1,
S6W2 and S4P1). For the remaining isolates, only a liquor extract was created. The
decision to omit the mycelium extract protocol for the remaining isolates was due, in
part, to the difficult and time-consuming nature of working with the separated
mycelia (compared with working with the sterile liquid filtrate). This decision was
supported by numerous published studies describing the preparation liquor extracts
for the screening of bioactive compounds (Bhagobaty and Joshi, 2012; Ganesh
Kumar et al., 2010; Hoffman et al., 2008; Lin et al., 2005; Nath et al., 2012; Swathi
et al., 2013; Xing et al., 2011).
4.3.2 Production of EtOAc and aqueous extracts from liquor
The fermentation liquor from each coprophilous fungus was separated from the
mycelia and then extracted with EtOAc, thus producing an aqueous extract and an
EtOAc extract. This protocol was followed with four fungi (F2P1, F13P1, S6W2 and
S4P1), but it was decided to omit the liquid–liquid separation step for the remaining
fungi and simply lyophilise the fermentation liquor to produce the liquor extract. The
reason for this change in the protocol was that preliminary studies indicated that
there was little difference in the bioactivity if the liquor was extracted with EtOAc,
and this was further supported by the results of the screening assays (see Sections
4.3.3, 4.3.4 and 4.3.5. Given that the liquid–liquid partitioning, drying and screening
of the extracts was a very time-intensive process, omitting this step meant that more
of the fungi could be screened. Although extraction of the fermentation liquor with
EtOAc is a common method in the literature (Gamble et al., 1995; Swathi et al.,
2013; Weber and Gloer, 1991; Whyte et al., 1997), there are many published
examples where the liquor is used in screening for bioactivity, either as the only
approach or as a preliminary screening tool that precedes further extraction and
separation (Ganesh Kumar et al., 2010; Liu et al., 2012; Zhang et al., 2012b).
In one such study, Phongpaichit et al. (2006) isolated endophytic fungi from
Garcinia plants and screened them for antimicrobial activity. The fermentation liquor

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
68
was used for preliminary screening via plate-hole diffusion assays, and EtOAc
extracts were produced from the liquor (and mycelia) for determining the MIC. The
study found that of the 18.6% of fermentation liquors that exhibited antimicrobial
activity, only 0.9–1.9% of the corresponding EtOAc extracts inhibited the same test
organism. These results suggest that testing the fermentation liquor is a useful
approach for evaluating antimicrobial activity, at least as the preliminary phase of
screening. Therefore, for the remaining coprophilous fungi, EtOAc extraction was
not performed and the fermentation liquor was lyophilised to produce extracts for
screening assays. Liquor extracts were produced from the four aforementioned fungi,
so that a comparison could be made in relation to the bioactivity potential of the
different extraction methods.
4.3.3 Antibacterial activity of coprophilous fungi extracts
4.3.3.1 Plate-hole diffusion screening of extracts
The coprophilous fungal extracts were screened for antibacterial activity via the
plate-hole diffusion assay against S. aureus, E. coli, K. pneumoniae and
P. aeruginosa. The final amount of extract added to each well of bacteria-seeded
agar was 2 mg. Following incubation, the appearance of a clear zone surrounding a
well, as shown in Figure 4.1, indicated antibacterial activity for the extract. The zone
diameter (inclusive of the well diameter, 6 mm) was measured and the mean values
from triplicate assays are presented in Table 4.1.

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
69
Figure 4.1 S. aureus plate-hole diffusion assay, following incubation. Clear zones indicate inhibition of S. aureus by liquor extracts from the fungi S4P4 and F14P1 and by the positive control CHX. The extracts from fungi F13P1 and F14W1 did not inhibit growth of the bacterium.
Seventeen (54.8%) of the 31 coprophilous fungi produced extracts that were capable
of inhibiting the growth of at least one test bacterium. Six fungi demonstrated broad-
spectrum antibacterial activity, and three fungi (F16EP1, F14P1 and S5P3) were able
to inhibit the growth of every bacterium to some degree. The Gram-positive
bacterium S. aureus was more sensitive to the fungal extracts than the three Gram-
negative bacteria. S. aureus was affected by the extracts from 14 fungi, whereas
E. coli, the second most susceptible bacterium, was inhibited by nine. Only three of
the extracts were capable of inhibiting the growth of P. aeruginosa. Similar
observations have been reported from the screening of extracts from other fungal
species, whereby S. aureus was shown to be the most sensitive of the test bacteria
(Buatong et al., 2011; Ganesh Kumar et al., 2010; Hoffman et al., 2008;
Phongpaichit et al., 2006). The differential sensitivity to the extracts by Gram-
negative and Gram-positive bacteria are likely to be due to differences in their cell
walls; Gram-negative bacteria have an outer polysaccharide membrane that acts as a
barrier to environmental substances, including antibiotics (Madigan et al., 2003).
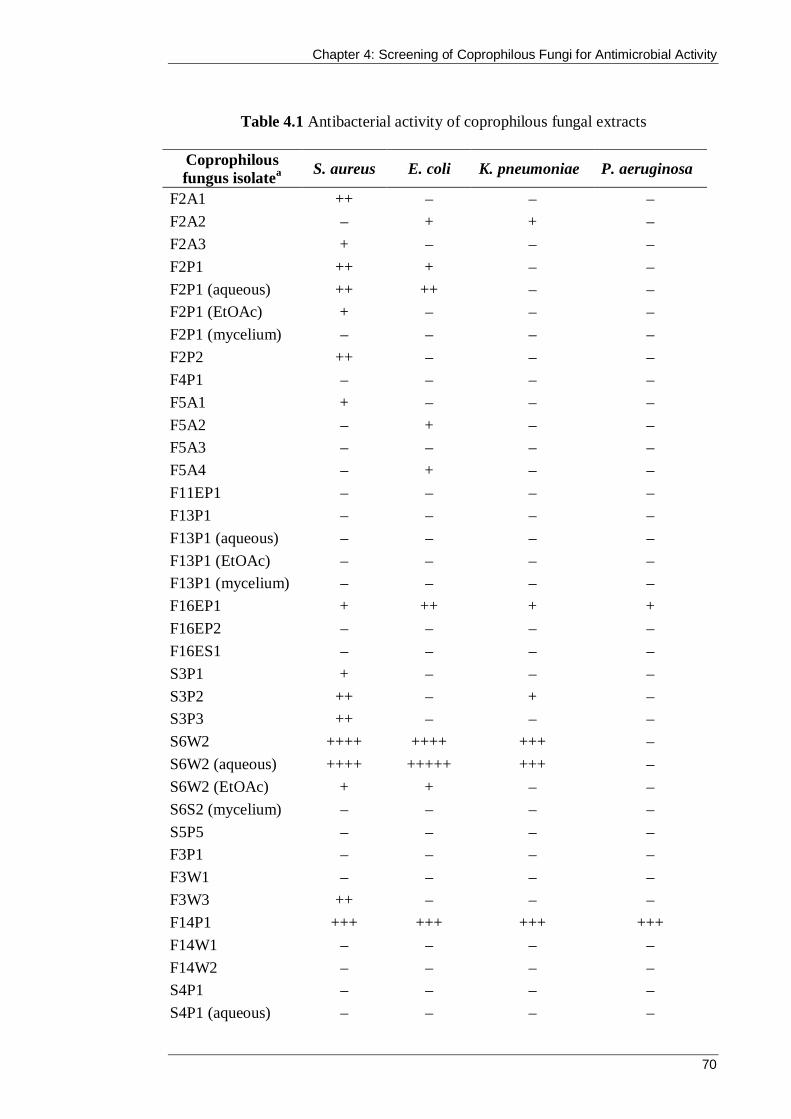
Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
70
Table 4.1 Antibacterial activity of coprophilous fungal extracts
Coprophilous fungus isolatea S. aureus E. coli K. pneumoniae P. aeruginosa
F2A1 ++ – – – F2A2 – + + – F2A3 + – – – F2P1 ++ + – – F2P1 (aqueous) ++ ++ – – F2P1 (EtOAc) + – – – F2P1 (mycelium) – – – – F2P2 ++ – – – F4P1 – – – – F5A1 + – – – F5A2 – + – – F5A3 – – – – F5A4 – + – – F11EP1 – – – – F13P1 – – – – F13P1 (aqueous) – – – – F13P1 (EtOAc) – – – – F13P1 (mycelium) – – – – F16EP1 + ++ + + F16EP2 – – – – F16ES1 – – – – S3P1 + – – – S3P2 ++ – + – S3P3 ++ – – – S6W2 ++++ ++++ +++ – S6W2 (aqueous) ++++ +++++ +++ – S6W2 (EtOAc) + + – – S6S2 (mycelium) – – – – S5P5 – – – – F3P1 – – – – F3W1 – – – – F3W3 ++ – – – F14P1 +++ +++ +++ +++ F14W1 – – – – F14W2 – – – – S4P1 – – – – S4P1 (aqueous) – – – –
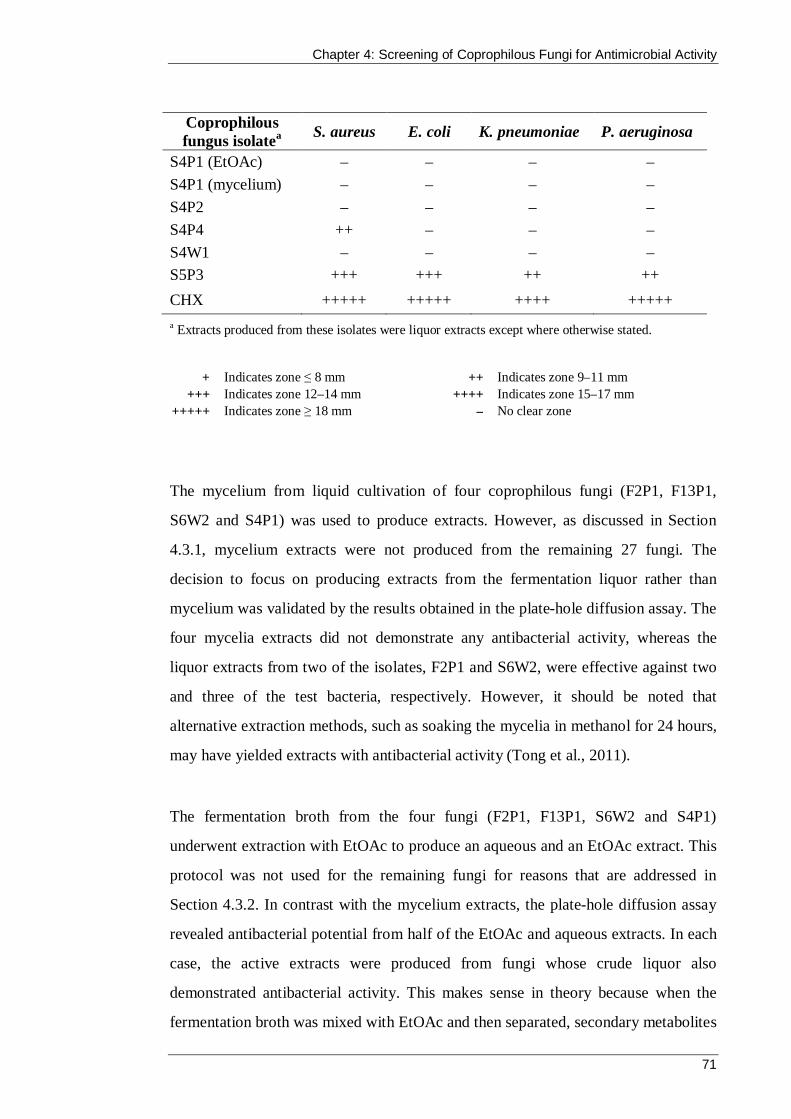
Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
71
Coprophilous fungus isolatea S. aureus E. coli K. pneumoniae P. aeruginosa
S4P1 (EtOAc) – – – – S4P1 (mycelium) – – – – S4P2 – – – – S4P4 ++ – – – S4W1 – – – – S5P3 +++ +++ ++ ++ CHX +++++ +++++ ++++ +++++ a Extracts produced from these isolates were liquor extracts except where otherwise stated.
+ Indicates zone ≤ 8 mm ++ Indicates zone 9–11 mm +++ Indicates zone 12–14 mm ++++ Indicates zone 15–17 mm
+++++ Indicates zone ≥ 18 mm – No clear zone
The mycelium from liquid cultivation of four coprophilous fungi (F2P1, F13P1,
S6W2 and S4P1) was used to produce extracts. However, as discussed in Section
4.3.1, mycelium extracts were not produced from the remaining 27 fungi. The
decision to focus on producing extracts from the fermentation liquor rather than
mycelium was validated by the results obtained in the plate-hole diffusion assay. The
four mycelia extracts did not demonstrate any antibacterial activity, whereas the
liquor extracts from two of the isolates, F2P1 and S6W2, were effective against two
and three of the test bacteria, respectively. However, it should be noted that
alternative extraction methods, such as soaking the mycelia in methanol for 24 hours,
may have yielded extracts with antibacterial activity (Tong et al., 2011).
The fermentation broth from the four fungi (F2P1, F13P1, S6W2 and S4P1)
underwent extraction with EtOAc to produce an aqueous and an EtOAc extract. This
protocol was not used for the remaining fungi for reasons that are addressed in
Section 4.3.2. In contrast with the mycelium extracts, the plate-hole diffusion assay
revealed antibacterial potential from half of the EtOAc and aqueous extracts. In each
case, the active extracts were produced from fungi whose crude liquor also
demonstrated antibacterial activity. This makes sense in theory because when the
fermentation broth was mixed with EtOAc and then separated, secondary metabolites

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
72
were either extracted in the organic phase or retained in the aqueous phase.
Therefore, the EtOAc and aqueous extracts should encompass all the components of
the liquor extract, but their distribution between the two phases will differ. Both the
EtOAc and aqueous extracts from F2P1 and S6W2 produced zones of inhibition
against one or more test bacteria, but the aqueous extracts demonstrated greater
bacterial inhibition compared with the EtOAc extracts. When fungi produced
antibacterial aqueous extracts, the resulting clear zone measurement was the same
diameter as the corresponding liquor extract, or, as in the case of the E. coli assays, it
was larger than the crude liquor extract. A possible explanation for this is that the
compounds within the fermentation liquor that were responsible for the inhibition
were retained in the aqueous phase during partitioning with EtOAc. Extraction of
non-active material into the EtOAc phase would therefore increase the concentration
of active metabolites in the aqueous phase. All extracts were tested at the same
concentration (2 mg in 20 µL) and, therefore, this would result in a larger inhibitory
effect.
The plate-hole diffusion assay is ideal for screening extracts for antibacterial activity
because it is easy to perform and relatively inexpensive. However, the size of a zone
of inhibition depends on a number of variables relating to the inoculum, media and
extract. Therefore, the assay is generally considered to be an indirect qualitative
method that provides categorical results rather than quantitative values (Wanger,
2007). Essentially, the size of a clear zone is determined by the competing
interactions of two dynamic processes: first, the growth of the test bacteria
throughout the agar, and second, the diffusion of the fungal extract compounds from
the well into the surrounding agar. Diffusion of the extract from the well produces a
descending concentration gradient radiating outwards from the well. The growth of
the test bacteria (and consequently the zone size) is influenced by variables including
the initial inoculum density, the nutritional composition of the agar, and the
temperature and length of incubation for the assay (Jorgensen et al., 1999). For
reproducible zone sizes, all of the assay parameters must be kept constant.

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
73
To address the above variables, a standardised inoculum density (1.5 × 108 cfu/mL)
was used and all plates were incubated for 18 hours at 37 °C. MHA was used for the
assay because it is a commonly used media for agar diffusion methods and is suitable
for the cultivation of a range of non-fastidious aerobic Gram-negative and Gram-
positive bacteria (Reade, 1994). The diffusion of the fungal extract compounds is
also affected by the agar composition, the incubation conditions and the depth of the
agar in the plate (Wanger, 2007). Therefore, a measured volume (15 mL) of MHA
was added to every plate. Furthermore, every extract was added to a well as a 20 µL
aliquot (100 mg/mL) and the wells were consistently 6 mm diameter. It is important
to remember that the diffusion of an antibacterial compound through the agar is also
influenced by its molecular properties such as weight, size, ionic charge and aqueous
solubility (Wanger, 2007). Given that the diffusion rate of the antibacterial
compound directly affects the size of the zone of inhibition, it cannot always be
assumed that a larger zone signifies greater susceptibility of the bacteria to the
extract.
More than half of the isolated fungi produced an extract that elicited a visible zone of
inhibition against one or more species of pathogenic bacteria. These results support
the hypothesis that coprophilous fungi from koala faeces are a possible source of
antibacterial compounds. Further studies of the fungus collection could include
testing the extracts against additional clinically relevant bacteria such as methicillin-
resistant strains of S. aureus.
4.3.3.2 Determination of MIC and MBC for most active extracts
The MIC is the lowest concentration of extract that is required to inhibit the growth
of the test bacterium. Compared with the plate-hole diffusion assay, this provides a
quantitative measurement of the efficacy of an antibiotic against a particular
bacterium. The MIC and MBC were established for the fungal extracts that were
capable of eliciting a zone of inhibition greater than 8 mm for one or more test
bacteria in the plate-hole diffusion assay (see Table 4.1). The extracts were tested

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
74
using a 96-well microdilution assay at a concentration range of 12,500–98 µg/mL.
Following incubation, the test wells were inspected for turbidity, with comparison to
the growth control and sterility control wells, and the MIC was determined for each
extract. MBC was assigned to the lowest concentration of extract that prevented
regrowth of the bacteria following subculture to MHA plates. Table 4.2 presents the
observed MIC and MBC values for the 11 liquor extracts and the two aqueous
extracts that were tested.

75
Table 4.2 MIC and MBC of coprophilous fungal extracts against Gram-positive and Gram-negative bacteria*
S. aureus E. coli K. pneumoniae P. aeruginosa
Coprophilous fungus isolatea Genusb MIC MBC MIC MBC MIC MBC MIC MBC F2A1 Aspergillus 6.25 >12.5 – – – – – – F2P1 Fimetariella 6.25 >12.5 – – – – – – F2P1 (aqueous) 6.25 12.5 3.125 6.25 – – – – F2P2 Fimetariella 6.25 12.5 – – – – – – F16EP1 Anthostomella – – 3.125 3.125 – – – – S3P2 Neurospora 3.125 6.25 – – – – – – S3P3 Neurospora 3.125 6.25 – – – – – – S6W2 Fusarium 3.125 >12.5 1.56 6.25 1.56 3.125 – – S6W2 (aqueous) 1.563 >12.5 0.78 6.25 1.56 3.125 – – F3W3 Thielavia 3.125 6.25 – – – – – – F14P1 Sordaria 3.125 6.25 3.125 12.5 6.25 12.5 3.125 6.25 S4P4 Trichoderma 6.25 >12.5 – – – – – – S5P3 Aspergillus 3.125 6.25 6.25 >12.55 6.25 12.5 6.25 >12.5
* Values represent the average concentration (mg/mL) of triplicate experiments and sd = (±0) a Extracts produced from these isolates are liquor extracts, except where otherwise stated b Identification of fungi was achieved by comparison of ITS regions of fungal rDNA with the GenBank database (see Section 3.3.4) – Not tested.

Chapter 5: Screening of Coprophilous Fungi for Antimicrobial Activity
76
The MICs and MBCs of the tested extracts were in the range of 6.25–0.78 mg/mL
and 12.5–3.125 mg/mL, respectively. The positive control, CHX, had an MIC and
MBC below the lowest concentration that was tested (4.88 µg/mL) for each bacteria.
In general, the relative antibacterial activity that was observed between the extracts
in the plate-hole diffusion assay was mostly consistent with the MIC values. For
example, the extracts that produced the largest clear zones (S6W2 liquor and aqueous
extracts, F14P1 and S5P3; see Table 4.1) also exhibited the lowest MIC values.
Although there was an overall relationship between zone size and MIC, there were
variations between the results from the two methods for each zone diameter range.
For example, in the twelve instances that an extract produced a zone of inhibition
that was 9–11 mm (++) in diameter, seven of the corresponding MICs were
6.25 mg/mL and five were 3.125 mg/mL. These differences could have been
influenced by the diffusion rate of the active compounds through the agar in the
plate-hole diffusion assay, a factor that is not a consideration in the microdilution
MIC assay. Two aqueous extracts were tested, and similar to the plate-hole diffusion
results, they performed equally or better than their liquor extract counterparts. In
particular, the aqueous extract from S6W2 was capable of inhibiting the growth of
both S. aureus and E. coli at half the MIC of the S6W2 liquor extract for each
bacterium.
The MIC of an extract is capable of limiting bacterial growth without necessarily
killing it. Therefore, it is possible that if the extract is removed, the bacteria will
recover and begin to grow again (Qaiyumi, 2007). To ascertain which concentration
of extract was bactericidal, 100 µL was taken from the wells showing no growth and
subcultured onto MHA plates. The lowest concentration of extract that led to the
inability of the bacteria to recover and produce visible growth was reported as the
MBC. One study into the antibacterial effects of natural compounds found that MBC
values for natural product extracts and compounds are commonly twice the MIC, and
sometimes this is used rather than an experimentally determined value (Furiga et al.,
2008). This was true for more than half of the experiments in this study; of the 24

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
77
extracts and bacteria combinations tested, 13 displayed MBCs that were twice the
corresponding MIC.
Although the MIC is a quantitative assessment of antibacterial activity compared
with the plate-hole diffusion assay, it is not a constant. It is affected by parameters
such as the inoculum size, the composition of the medium and the incubation
conditions (Madigan et al., 2003). To address this, these variables were kept
consistent throughout the study, which meant that it was possible to compare the
extracts and determine which was the most effective against each organism (see
Table 4.3).
Table 4.3 Summary of the most effective extracts against each test bacteria
S. aureus E. coli K. pneumoniae P. aeruginosa
MIC (mg/mL) 1.56 0.78 1.56 1.56 3.13
Extract S6W2 aqueous
S6W2 aqueous
S6W2 liquor
S6W2 aqueous
F14P1 liquor
Comparisons of MICs of extracts, fractions and compounds between different
research studies are only meaningful if conditions are rigorously standardised within
each study, and if conditions are identical between studies. Nonetheless, reporting of,
and comparisons between, fungal extract MIC values is considered standard practice.
The research into bioactive coprophilous fungi is scarce despite their proven
potential for antibiotic discovery. This is thought to be because they have not yet
been incorporated in screening programs at levels competitive with fungi from other
substrata and niche groups (Bills et al., 2013). The vast majority of studies detail the
structural elucidation and antibacterial activity of isolated compounds, and very few
report the early stages of research and state the activity of fermentation broths or
crude extracts like those that were tested in this chapter.

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
78
In contrast, there are numerous studies of endophytic fungi that report the
antibacterial activity of broths and crude extracts, many of which have not yet
ascertained the MIC and instead rely on agar diffusion methods (Gond et al., 2012;
Hazalin et al., 2009; Tolulope et al., 2015; Zhang et al., 2012a; Zhang et al., 2015).
MIC values were ascertained by Phongpaichit et al. (2006), who studied the
antibacterial activity of fermentation broths of endophytic fungi isolated from
Garcinia plants. The EtOAc extracts of 10 fungal broths demonstrated inhibition of
S. aureus, with MICs in the range of 32–512 µg/mL. Similar MIC values (31.25–
250 µg/mL) were obtained by Arivudainambi et al. (2011) for the extracts of the
endophyte fungus Colletotrichum gloeosporioides against 10 clinical strains of
S. aureus. In comparison, the lowest MIC against S. aureus observed in this study
was 1.563 mg/mL, which is between three and 50 times larger.
The results presented in this chapter represent preliminary screening efforts and must
not be disregarded. First, the extracts prepared in this chapter are crude in nature.
Therefore, it is likely that any bioactive compounds that are present make up only a
small percentage of the extract by weight, which lessens the observed antibacterial
affect. Fractionation of the extracts, guided by antibacterial assays, would isolate
active constituents for identification and provide an accurate assessment of their
antibacterial potential. Second, there has been no attempt to optimise the
fermentation conditions to improve antibacterial activity. Changes to the liquid
medium (e.g., variations to carbon and nitrogen sources and pH) and the
fermentation temperature could affect the secondary metabolite profile of an isolate
and potentially upregulate the production of an antibacterial compound and decrease
the observed MIC (Thammajaruk et al., 2010; Xu et al., 2010).
4.3.3.3 Time-course growth analysis of liquor extracts F14P1 and
S6W2
While the plate-hole diffusion and MIC methods provided an end-point assessment
of an extract’s ability to inhibit bacterial growth, the time-course assay allowed the
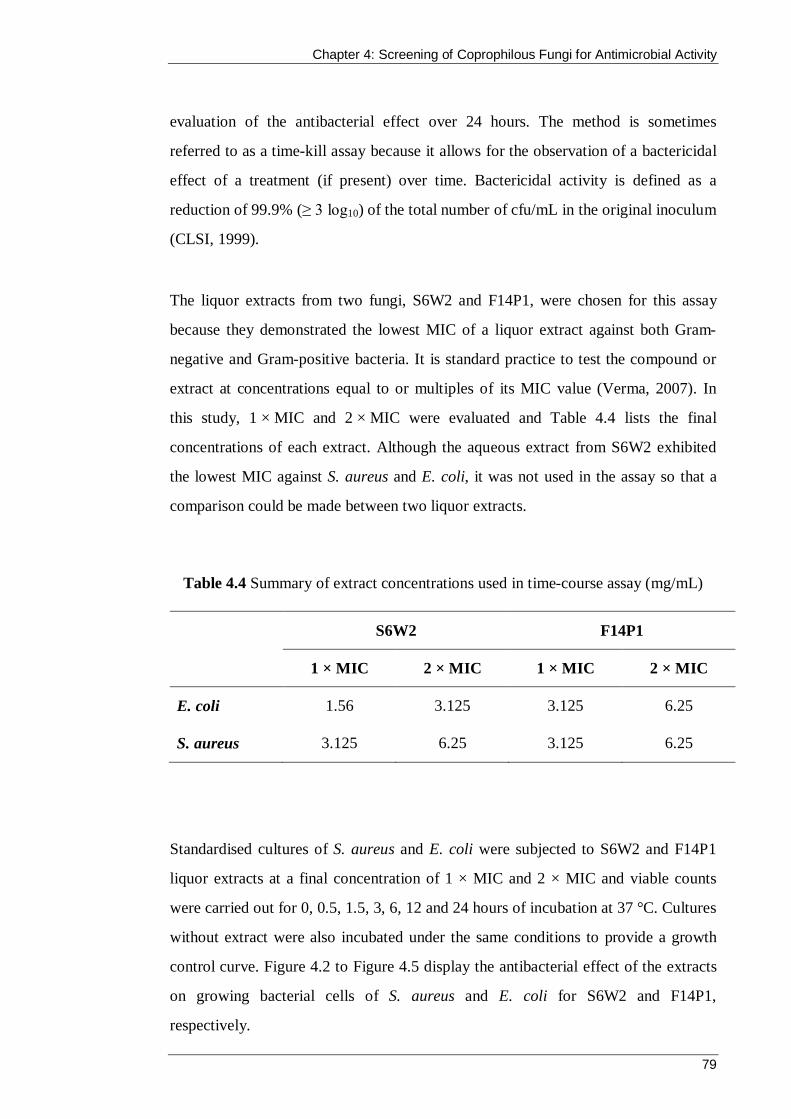
Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
79
evaluation of the antibacterial effect over 24 hours. The method is sometimes
referred to as a time-kill assay because it allows for the observation of a bactericidal
effect of a treatment (if present) over time. Bactericidal activity is defined as a
reduction of 99.9% (≥ 3 log10) of the total number of cfu/mL in the original inoculum
(CLSI, 1999).
The liquor extracts from two fungi, S6W2 and F14P1, were chosen for this assay
because they demonstrated the lowest MIC of a liquor extract against both Gram-
negative and Gram-positive bacteria. It is standard practice to test the compound or
extract at concentrations equal to or multiples of its MIC value (Verma, 2007). In
this study, 1 × MIC and 2 × MIC were evaluated and Table 4.4 lists the final
concentrations of each extract. Although the aqueous extract from S6W2 exhibited
the lowest MIC against S. aureus and E. coli, it was not used in the assay so that a
comparison could be made between two liquor extracts.
Table 4.4 Summary of extract concentrations used in time-course assay (mg/mL)
S6W2 F14P1
1 × MIC 2 × MIC 1 × MIC 2 × MIC
E. coli 1.56 3.125 3.125 6.25
S. aureus 3.125 6.25 3.125 6.25
Standardised cultures of S. aureus and E. coli were subjected to S6W2 and F14P1
liquor extracts at a final concentration of 1 × MIC and 2 × MIC and viable counts
were carried out for 0, 0.5, 1.5, 3, 6, 12 and 24 hours of incubation at 37 °C. Cultures
without extract were also incubated under the same conditions to provide a growth
control curve. Figure 4.2 to Figure 4.5 display the antibacterial effect of the extracts
on growing bacterial cells of S. aureus and E. coli for S6W2 and F14P1,
respectively.
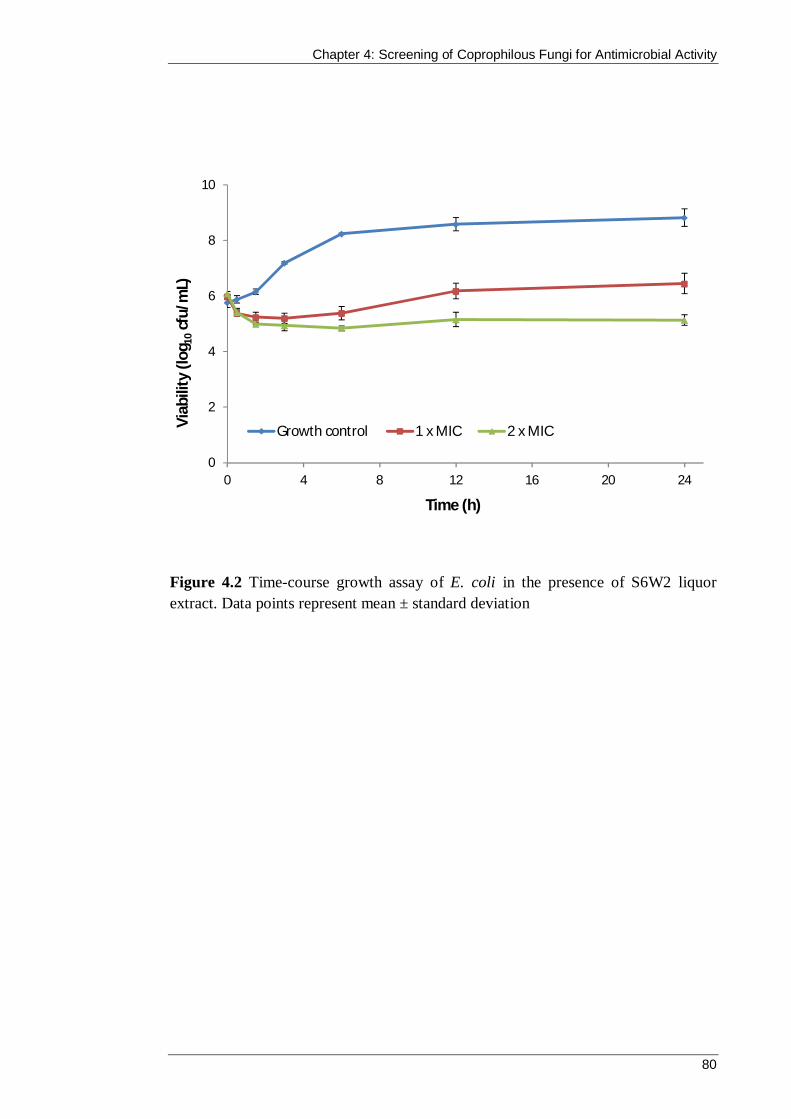
Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
80
Figure 4.2 Time-course growth assay of E. coli in the presence of S6W2 liquor extract. Data points represent mean ± standard deviation
0
2
4
6
8
10
0 4 8 12 16 20 24
Viab
ility
(log
10cf
u/m
L)
Time (h)
Growth control 1 x MIC 2 x MIC

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
81
Figure 4.3 Time-course growth assay of S. aureus in the presence of S6W2 liquor extract. Data points represent mean ± standard deviation.
0
2
4
6
8
10
0 4 8 12 16 20 24
Viab
ility
(log
10cf
u/m
L)
Time (h)
Growth control 1 x MIC 2 x MIC

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
82
Figure 4.4 Time-course growth assay of E. coli in the presence of F14P1 liquor
extract. Data points represent mean ± standard deviation
0
2
4
6
8
10
0 4 8 12 16 20 24
Viab
ility
(log
10cf
u/m
L)
Time (h)
Growth control 1 x MIC 2 x MIC

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
83
Figure 4.5 Time-course growth assay of S. aureus in the presence of F14P1 liquor extract. Data points represent mean ± standard deviation.
0
2
4
6
8
10
0 4 8 12 16 20 24
Viab
ility
(log
10cf
u/m
L)
Time (h)
Growth control 1 x MIC 2 x MIC

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
84
The S6W2 extract exhibited a significant reduction in viability of E.coli from 1.5
hours onwards compared with the growth control (Figure 4.2). However, the
difference between the original inoculum (at time = 0) and the 1 × MIC and 2 × MIC
treated cells after 24 hours was only approximately +0.5 log10 and −1.0 log10 units,
respectively. Because this represents less than a 3 log10 reduction, the S6W2 extract
cannot be described as bactericidal against E. coli. The 1 × MIC treatment did cause
an initial decline in viable cells during the first three hours of incubation, but the
bacteria were able to recover and grow to a density greater than the original
inoculum, although not significantly. In the presence of the two-fold concentrated
S6W2 extract, E. coli was also able to recover growth following an initial steep
decline, but not to a level beyond the original inoculum. In conclusion, the S6W2
extract can be described as bacteriostatic against E. coli at both 1.56 mg/mL and
3.125 mg/mL. Bactericidal activity may have been observed for the extract using the
MBC (6.25 mg/mL) in the assay, but this was not performed.
The S6W2 extract demonstrated significant inhibitory activity against S. aureus from
3 hours onwards compared with the growth control (see Figure 4.3). At 24 hours, the
number of surviving cells represented approximately a 0.7 log10 and 1.0 log10
reduction from the original inoculum upon treatment with 1 × MIC and 2 × MIC,
respectively. According to the definition, extract S6W2 did not exhibit bactericidal
activity against S. aureus within 24 hours. The antibacterial effect of the 2 × MIC
extract (6.25 mg/mL) was not significantly different to the inhibition caused by the
1 × MIC extract (3.125 mg/mL) at every time point. This is not surprising given that,
in the microdilution assay, an MBC was not assigned because the highest
concentration of the extract (12.5 mg/mL) was not able to prevent regrowth of
S. aureus in the subsequent growth test (see Table 4.2).
The F14P1 extract exhibited a significant reduction in viability of E. coli from 1.5
hours onwards compared with the growth control (see Figure 4.4). Similar to the
S6W2 extract, the 1 × MIC treatment resulted in an initial decline in viable E. coli
cells, but the bacteria were able to recover and grow to a density approximately

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
85
0.7 log10 units greater than the original inoculum, which represents a significant
increase. The two-fold concentrated F14P1 extract caused an initial steep incline in
viability followed by a plateau of growth inhibition at approximately 1.2 log10 units
lower than the original inoculum. In conclusion, the F14P1 extract can be described
as bacteriostatic against E. coli at both 3.125 mg/mL and 6.25 mg/mL. Bactericidal
activity may have been observed for the extract using the MBC (12.5 mg/mL) in the
assay, but this test was not performed.
Figure 4.5 shows the significant reduction in viable S. aureus cells treated with
1 × MIC and 2 × MIC F14P1 extracts from 1.5 hours onwards compared with the
growth control. The 1 × MIC extract exhibited a bacteriostatic effect that inhibited
growth of the organism, but this did not reduce the viability by more than 3 log10
units. In contrast, at 12 hours the 2 × MIC extract had produced an approximate
3.8 log10 reduction in viable cells, thus demonstrating bactericidal activity against
S. aureus. This result was expected because the 2 × MIC extract equated to
6.25 mg/mL, which was the MBC value ascertained in the microdilution and growth
test.
Overall, the results of the time-course experiments correlated with the MIC and
MBC values that were determined for both extracts in the microdilution method (see
Table 4.2). The S6W2 extract exhibited bacteriostatic inhibitory activity against both
test organisms at both concentrations. The F14P1 extract also demonstrated
bacteriostatic activity against E. coli at both concentrations. For these tests, the
2 × MIC represented a concentration that was half the experimentally determined
MBC, or as was the case with S6W2 against S. aureus, an MBC could not be
determined. The F14P1 extract exhibited bacteriostatic activity against S. aureus at
1 × MIC and bactericidal activity at 2 × MIC that was equal to the MBC. It would be
interesting to conduct the time-course assay using the MBC for all extracts. This
work could be carried out in the future, but it would be more useful to perform these
additional experiments with an extract that has undergone activity-guided separation
rather than with the crude extracts.

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
86
4.3.4 Antifungal activity of coprophilous fungi extracts
The coprophilous fungal extracts were screened for antifungal activity against the
yeast C. albicans and the filamentous fungus D. brizae.
4.3.4.1 C. albicans inhibition
Plate-hole diffusion assays were performed with all extracts to determine which were
capable of inhibiting the growth of C. albicans. The total amount of extract within
each well was 2 mg and the presence of a clear zone surrounding a well indicated
inhibitory activity by the respective extract, such as in Figure 4.6.
Eight (18.6%) of the 31 coprophilous fungi produced extracts that were capable of
inhibiting the growth of C. albicans. Extracts that produced a clear zone greater than
8 mm were tested in a microdilution assay (with a concentration range of 12.5–
0.098 mg/mL) to ascertain their MIC and MFC. Table 4.5 summarises the results of
these assays from the extracts that demonstrated anti-candida activity.
In the plate-hole diffusion assay, C. albicans exhibited a similar sensitivity to E. coli
against the collection of fungal extracts. For both microorganisms, eight fungi
produced extracts that produced a zone of inhibition that was greater than 6 mm. In
general, the MICs of the active extracts against C. albicans were approximately
twice the MICs against E. coli. However, the aqueous extract of the S6W2 liquor was
capable of preventing visible growth at a concentration of 0.52 mg/mL, which was
lower than any MIC obtained against the test bacteria.

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
87
Figure 4.6 C. albicans plate-hole diffusion assay following incubation. Clear zones indicate inhibition of the yeast by the liquor extract from the fungus S5P3 (left) and by the positive control CHX (right).
Table 4.5 Inhibition of C. albicans by coprophilous fungi extracts
Coprophilous fungal extractsa Genusb
Plate-hole diffusion
assayc
Microdilution assay (mg/mL)
MIC MFC
F11EP1 Penicillium ++ 12.5 ± 0.0 >12.5 ± 0.0 S3P1 Neurospora + – – S6W2 Fusarium +++ 0.78 ± 0.0 1.56 ± 0.0 S6W2 (aqueous) ++++ 0.52 ± 0.23 1.04 ± 0.45 S5P5 Plectosphaerella + – – F14P1 Sordaria ++ 6.25 ± 0.0 12.5 ± 0.0 S4P2 Sordaria ++ 12.5 ± 0.0 >12.5 ± 0.0 S4P4 Trichoderma ++ 6.25 ± 0.0 12.5 ± 0.0 S5P3 Aspergillus ++ 6.25 ± 0.0 >12.5 ± 0.0 a Extracts produced from these isolates were liquor extracts except where otherwise stated. b Identification of fungi was achieved by comparison of ITS regions of fungal rDNA with the GenBank database (see Section 3.3.4) - Not tested. c Zone of inhibition diameter includes 6 mm disk:
+ Indicates zone ≤ 8 mm ++ Indicates zone 9–11 mm +++ Indicates zone 12–14 mm ++++ Indicates zone 15–17 mm

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
88
Four of the seven extracts demonstrated an MFC that was twice the corresponding
MIC, which suggests that they may be good candidates for further separation and the
identification of a fungicidal constituent. A time-course growth assay would provide
a better indication as to whether these extracts were fungicidal or fungistatic, but this
test was not performed. Treatment with a fungicidal (as opposed to a fungistatic)
drug is often important for the treatment of immunosuppressed individuals suffering
from invasive infections caused by C. albicans (Lewis and Graybill, 2008).
Fungistatic drugs are still immensely useful, as demonstrated by one of the most
commonly administered drugs for Candida infections, fluconazole. This azole
compound is classified as a fungistatic agent and is listed on the WHO’s current
Model List of Essential Medicines (WHO, 2015). Unfortunately, Candida species are
rapidly acquiring resistance to fluconazole, and therefore the identification of novel
compounds that can attenuate Candida infections would be beneficial. Extracts
produced from the fungus S6W2 represent ideal candidates for further exploration of
this activity and were chosen to undergo bioassay-directed separation (see Chapter
Five).
In addition to discovering new stand-alone anti-candidal compounds, current
research efforts are working to potentiate the efficacy of fluconazole via drug
synergism, with the aim of affecting previously resistant strains and improving
fluconazole’s activity from fungistatic to fungicidal (Fiori and Van Dijck, 2012; Liu
et al., 2014; Spitzer et al., 2011; Zhai et al., 2010). This was achieved by
Phaopongthai et al. (2013) who combined fluconazole with a secondary metabolite
that they isolated from an endophytic fungus Alternaria alternata that was obtained
from a Thai medicinal plant. Screening of the coprophilous fungal extracts for
synergistic activity is an alternative approach to exploring their therapeutic potential,
although it would be more beneficial to use isolated compounds rather than crude
extracts.

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
89
4.3.4.2 D. brizae inhibition
The ability of the extracts to inhibit the growth of D. brizae was evaluated using
three agar-based assays and one broth microdilution assay. This methodology was
employed because it assessed the effect of the extract on the fungus at different
stages of growth. The spore germination assay detected antifungal activity in the
earliest stages of growth, the radial growth assay assessed inhibition during the first
phases of filamentous growth and the hyphal extension assay analysed the effect on
active growth (Quiroga et al., 2009). The extracts that inhibited D. brizae during two
or more stages of growth were subsequently included in the microdilution assay to
determine their MIC. Table 4.6 presents the results of the three agar assays for each
extract.
Twelve (38.7%) of the coprophilous fungi produced extracts that exhibited antifungal
activity in one or more of the agar assays. Three extracts (S6W2, S6W2 aqueous and
S4P2) were able to inhibit the test fungus in all three assays. The hyphal extension
assay recorded the highest number of extracts demonstrating inhibition (15),
followed by the radial growth (10) and spore germination (five) assays. The variation
in inhibition between the methods may reflect the effect that the D. brizae growth
stage had on the antifungal capabilities of the extracts. For example, of the 15
extracts that inhibited hyphal extension, only four were also able to inhibit spore
germination, which suggests that this early stage of growth may be more resistant to
interference from the secondary metabolites present in the extracts.

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
90
Table 4.6 Inhibition of D. brizae
Coprophilous fungal extracta
Agar assay
Hyphal extensionb
Radial growth (% growth
inhibition ± sd)c
Spore germination (mm ± sd)d
F2A1 + – – F2A2 + – – F2A3 – – – F2P1 – – – F2P1 (aqueous) + – – F2P1 (EtOAc) – – – F2P1 (mycelium) – – – F2P2 + 14.60 ± 2.53 – F4P1 + – 11.00 ± 0.50 F5A1 – – – F5A2 – – – F5A3 – – – F5A4 – – – F11EP1 + 11.16 ± 2.11 – F13P1 – – – F13P1 (aqueous) – – – F13P1 (EtOAc) – – – F13P1 (mycelium) – – – F16EP1 + 15.82 ± 2.46 – F16EP2 + – – F16ES1 – – – S3P1 – – 14.17 ± 0.76 S3P2 – – – S3P3 – – – S6W2 + 17.85 ± 3.80 17.67 ± 0.76 S6W2 (aqueous) + 31.03 ± 2.88 20.50 ± 1.80 S6W2 (EtOAc) – – – S6W2 (mycelium) – – – S5P5 – – – F3P1 – – – F3W1 + 14.00 ± 3.06 – F3W3 – – – F14P1 + 18.86 ± 2.74 – F14W1 – – – F14W2 – – –
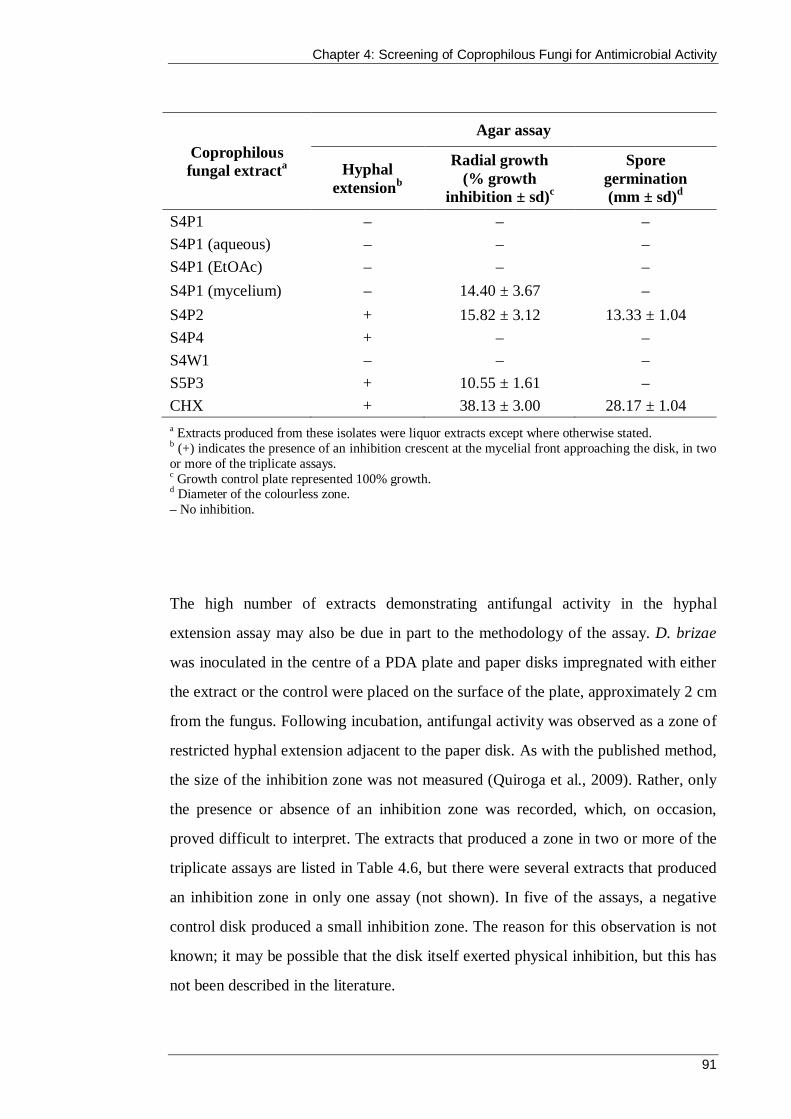
Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
91
Coprophilous fungal extracta
Agar assay
Hyphal extensionb
Radial growth (% growth
inhibition ± sd)c
Spore germination (mm ± sd)d
S4P1 – – – S4P1 (aqueous) – – – S4P1 (EtOAc) – – – S4P1 (mycelium) – 14.40 ± 3.67 – S4P2 + 15.82 ± 3.12 13.33 ± 1.04 S4P4 + – – S4W1 – – – S5P3 + 10.55 ± 1.61 – CHX + 38.13 ± 3.00 28.17 ± 1.04 a Extracts produced from these isolates were liquor extracts except where otherwise stated. b (+) indicates the presence of an inhibition crescent at the mycelial front approaching the disk, in two or more of the triplicate assays. c Growth control plate represented 100% growth. d Diameter of the colourless zone. – No inhibition.
The high number of extracts demonstrating antifungal activity in the hyphal
extension assay may also be due in part to the methodology of the assay. D. brizae
was inoculated in the centre of a PDA plate and paper disks impregnated with either
the extract or the control were placed on the surface of the plate, approximately 2 cm
from the fungus. Following incubation, antifungal activity was observed as a zone of
restricted hyphal extension adjacent to the paper disk. As with the published method,
the size of the inhibition zone was not measured (Quiroga et al., 2009). Rather, only
the presence or absence of an inhibition zone was recorded, which, on occasion,
proved difficult to interpret. The extracts that produced a zone in two or more of the
triplicate assays are listed in Table 4.6, but there were several extracts that produced
an inhibition zone in only one assay (not shown). In five of the assays, a negative
control disk produced a small inhibition zone. The reason for this observation is not
known; it may be possible that the disk itself exerted physical inhibition, but this has
not been described in the literature.

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
92
Unlike the method of Quiroga et al. (2009) this study included a positive control that
consisted of disks impregnated with 20 µL of CHX. However, this positive control
could not generate a reproducible inhibition zone and in some assays it did not
exhibit any inhibition at all (see Figure 4.7). CHX is a commonly used biocidal agent
for testing against bacteria and yeast, and one study has confirmed its antifungal
activity against some filamentous fungi (Sandle et al., 2014). CHX was capable of
inhibiting D. brizae in the radial growth, spore germination and microdilution assays,
but an alternative positive control (such as Derosal Plus®; Bayer, Germany) may
better serve the hyphal extension assay. For future studies, further modifications
could be made to this assay to improve the quality of the data. For example, a similar
but quantitative method was employed by Flores et al. (2013) in their evaluation of
the antifungal activity of extracts prepared from the fermentation liquor of the
endophytic fungus Phomopsis longicolla. Disks impregnated with extract were
placed 4 cm from the fungal pathogen plug (one extract per PDA plate), and on day
seven, the distance between the disk and the mycelial front was measured with
comparison to the negative and positive controls. Because the inhibitory zones were
measured, the mean diameter could be compared with those produced by the controls
and a statistical test (the Tukey method) was used to determine if statistically
significant inhibition had occurred.

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
93
Figure 4.7 Example of a hyphal extension assay where the positive control (CHX) disk demonstrated only very minimal inhibition of D. brizae.
In the radial growth assays, 10 extracts were capable of reducing the growth of
D. brizae compared with the growth control. The S6W2 aqueous extract
demonstrated the greatest inhibition, producing a 31.03 ± 2.88% reduction in radial
growth. The mycelium extract of S4P1 exhibited a 14.40 ± 3.67% reduction in radial
growth and was the first and only occurrence of antimicrobial activity from a
mycelium extract.
This assay required the incorporation of the extracts into the agar at a final
concentration of 5 mg/mL. For each plate, 10 mL of PDA was poured and 500 µL of
each extract stock solution (100 mg/mL) had to be added to each replicate assay.
This agar-dilution method is a common approach to antifungal testing; however, it
became apparent during this research that it is more appropriate for testing extracts or
compounds when supply is in abundance. Although the supply of the extracts was
not strictly limited, their production was a time-intensive process. Thus, it was
CHX
EtOAc Broth
Aqueous

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
94
important to consider the efficient use of the extracts and it was decided that any
subsequent antifungal testing of fractions or compounds that were separated from the
extracts would not be carried out via the radial growth test.
In the spore germination assay, MTT was spread onto the surface of the agar to
increase the clarity of the zones of inhibition surrounding the antifungal extracts.
Viable fungi cells are able to reduce the MTT, which leads to the intracellular
accumulation of a purple formazan compound that can be seen macroscopically
(Meletiadis et al., 2000). Non-viable cells cannot reduce the MTT and therefore
remain colourless. Hence, an extract that is capable of inhibiting spore germination
will produce a colourless zone in the surrounding agar, such as in Figure 4.8 Five
extracts inhibited spore germination in the bioautographic assay, and the S6W2
aqueous extract was again the most effective, producing a colourless zone that
measured 20.50 ± 1.80 mm.
Ten extracts demonstrated inhibition of D. brizae in two or more of the agar assays
and these were subsequently included in the microdilution assay to determine their
MIC. The MIC of the extracts ranged from 8.33 ± 3.6 to 25.00 ± 0.0 mg/mL (see
Table 4.7).

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
95
Figure 4.8 Spore germination plate following incubation and treatment with MTT. A clear zone surrounds the S6W2 aqueous extract.
Table 4.7 MIC of coprophilous fungal extracts against D. brizae
Coprophilous fungal a
Genusb MIC (mg/mL ± sd) F2P2 Fimetariella 12.5 ± 0.0 F4P1 Neurospora >25.00 ± 0.0 F11EP1 Penicillium 25.00 ± 0.0 F16EP1 Anthostomella 20.83 ± 7.22 S6W2 Fusarium 16.67 ± 7.22 S6W2 (aqueous) 8.33 ± 3.61 F3W1 Bjerkandera 12.50 ± 0.0 F14P1 Sordaria 10.42 ± 3.61 S4P2 Sordaria 25.00 ± 0.0 S5P3 Aspergillus 6.25 ± 0.0
a Extracts produced from these isolates were liquor extracts except where otherwise stated. b Identification of fungi was achieved by comparison of ITS regions of fungal rDNA with the GenBank database (see Section 3.3.4)
F2P1 aqueous
S6W2 aqueous
F13P1 aqueous
S4P1 aqueous

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
96
The F4P1 liquor extract did not inhibit D. brizae at 25 mg/mL despite demonstrating
inhibitory activity in the hyphal extension and spore germination assays. However, as
discussed previously, the hyphal extension results may have been unreliable given
the presence of some inhibitory zones surrounding the negative controls. The clear
zone that this extract produced in the spore germination assay (11.00 ± 0.50 mm)
was the smallest of those recorded.
The lowest extract MIC against D. brizae (8.33 ± 3.6 mg/mL) was attributed to the
aqueous extract from coprophilous isolate S6W2. CHX was able to prevent visible
growth of the fungus at the lowest concentration tested, 39.06 µg/mL, which is
approximately 200-fold lower than the MIC of the S6W2 aqueous extract. Although
the MICs of the extracts appear modest when compared with CHX, there are two
important factors that need to be considered. These factors have been
comprehensively addressed in the discussion of the antibacterial MIC assays (see
Section 4.3.3.2). Briefly, the extracts tested in this chapter are crude and the research
represents a preliminary exploration of their antifungal potential prior to activity-
guided separation. Furthermore, optimisation of fermentation conditions was not
carried out, and it is likely that the bioactivity of the crude extracts can be enhanced.
Given that there has been very little research into the exploitation of coprophilous
fungi against plant pathogens (Sarrocco et al., 2015a), the extracts warrant further
evaluation as possible control agents.
4.3.5 Anti-QS activity of coprophilous fungi extracts
The coprophilous fungal extracts were tested for anti-QS activity using a disk
diffusion assay against the screening bacterium C. violaceum (Figure 4.9). In this
wild type strain, the production of the purple pigment violacein is under N-
acylhomoserine lactone QS control (McClean et al., 1997). When QS signals are
quenched, non-pigmented bacteria grow and can be observed as a turbid halo of
viable but colourless cells surrounding the extract impregnated disk. Growth
inhibition by an extract results in a zone of inhibition similar to those observed in the

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
97
antibacterial plate-hole diffusion assay (see Section 4.3.3.1, Figure 4.1). Thus, C.
violaceum is an excellent screening tool to detect the presence of anti-QS activity by
an extract.
Figure 4.9 Anti-QS activity using C. violaceum in a disk diffusion assay. Disks were impregnated with 2 mg of liquor extracts from fungi S4P4, F13P1, F14W1 and F14P1. Note: A 20µg gentamicin disk was included (centre) for comparison to an antibacterial zone of inhibition. When the bacteria grow to a threshold density, N-acylhomoserine lactone molecules are produced and detected, resulting in the production of purple violacein pigment.
Following incubation of the disk diffusion assay, the extracts were inspected for the
presence of anti-QS or antibacterial activity, and zones were measured to the nearest
millimetre. For the purpose of this thesis, photos were taken with the plate on top of
printed text to demonstrate the difference in appearance between the turbid anti-QS
zone and the clear antibacterial zone. In Figure 4.10a, the aqueous extract of fungus
S6W2 produced an anti-QS halo. Colourless C. violaceum colonies were observed
surrounding the disk and caused the underlying text to appear blurred. In
comparison, the gentamicin disk completely inhibited the growth of the bacterium,
S4P4 F13P1
F14P1 F14W1
Gentamicin

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
98
resulting in a clear halo and well-defined text (see Figure 4.10b). The liquor extract
of S4P4 did not demonstrate anti-QS or antibacterial activity and therefore was
surrounded by confluent purple growth (see Figure 4.10c).
Figure 4.10 Example of results obtained in anti-QS disk diffusion assays. In each photo the disk is 6 mm: a) colourless, turbid anti-QS halo, b) clear antibacterial halo and c) no halo.
Table 4.8 Anti-QS activity of coprophilous fungi extracts against C. violaceum
Coprophilous fungi extract a Genus b QS inhibition
(mm ± sd) c Growth inhibition
(mm ± sd) d S6W2 Fusarium 1.8 ± 0.29 0.5 ± 0 S6W2 (aqueous) 3.5 ± 0 0.67 ± 0.29 F3W3 Thielavia 2.8 ± 0.29 0 ± 0 F14P1 Sordaria 3.0 ± 0 0.5 ± 0 S5P3 Aspergillus 2.0 ± 0 0.17 ± 0.29 Gentamicin 0 ± 0 5.17 ± 0.29 a Extracts produced from these isolates were liquor extracts except where otherwise stated. b Identification of fungi was achieved by comparison of ITS regions of fungal rDNA with the GenBank database (see Section 3.3.4) c Measurements of turbid, colourless zones were taken from the edge of the purple growth to the perimeter of the disk or growth inhibition (if present). d Measurements of clear zones were taken from the edge of the disk to the boundary of clear and turbid zones.
Four (12.9%) of the coprophilous fungi produced extracts that exhibited anti-QS
activity in the disk diffusion assays. The largest zone of colourless C. violaceum
growth was produced by the aqueous extract of S6W2. Table 4.8 presents the widths

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
99
of the turbid colourless anti-QS zones and clear antibacterial zones produced by the
active extracts.
QS systems regulate diverse physiological processes in bacterial populations,
including several functions that are associated with increased virulence. Disruption
of QS, rather than killing bacteria, represents an alternative approach to bacterial
control that may also reduce the development of resistant strains (Hentzer and
Givskov, 2003). The potential of an extract to be an effective anti-QS agent is also
predicted by its absence of antibacterial activity in the assay. All but one of the active
extracts (F3W3) produced very small antibacterial clear zones immediately adjacent
to the disk, preceding turbid anti-QS halos. As described in Section 4.3.3.1, assays
that rely on diffusion of an extract through agar result in a descending concentration
gradient radiating outwards from the disk or well. Hence, the area closest to the disk
contained the highest concentration of extract. This begs the question: was the
observed anti-QS halo simply the result of incomplete inhibition by the extract (at
decreasing concentrations) that prevented C. violaceum from reaching the density
threshold required to initiate QS and produce violacein? Although this conclusion
cannot be entirely disregarded, it was not observed in the disk diffusion results for
gentamicin. A halo completely devoid of bacteria surrounded the antibiotic disk and
there were no colourless cells preceding the purple growth, despite a similarly
decreasing concentration gradient (see Figure 4.11). The same observation has been
reported with another common antibiotic, tetracycline (Adonizio et al., 2006).

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
100
Figure 4.11 Magnified view of the clear zone produced by the gentamicin antibiotic disk.
Additionally, the ability of cells to recover from the anti-QS activity and regain
production of violacein was established. A sample of colourless colonies that were
surrounding an active extract was subcultured onto Luria–Bertani agar alongside a
sample of pigmented cells from the same plate for comparison. Following
incubation, the previously colourless bacteria were purple, indicating that their
viability had not been affected by the extract and that there had been recovery of QS
communication among the cells (Figure 4.12).

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
101
Figure 4.12 Colourless cells were able to grow upon subculture and incubation and regained QS directed production of violacein (right). A sample of purple cells from the same plate was also subcultured for comparison (left).
The results of the anti-QS screening in this chapter are promising, particularly as
there is currently no published research describing anti-QS activity by coprophilous
fungi or their extracts. In particular, the liquor extract of isolate F3W3 warrants
further exploration because it did not exhibit any growth inhibition. The fungi S6W2
and F14P1 were chosen for bioactivity-guided separation (Chapter Five) with the aim
of identifying the compound responsible for the observed QS inhibition.
4.4 Summary
The premise behind this work was that the fitness of coprophilous fungi is challenged
by competing and invading organisms within koala faeces and, therefore, to survive
they must produce chemicals to mediate these interactions. In accordance with this
premise, many of the isolates were, indeed, found to exhibit antimicrobial activity.
This was the first work, to my knowledge, in which the antibacterial, antifungal and
anti-QS potential of Australian coprophilous fungi have been explored.
Violacein-producing bacteria
Bacteria recovered from region adjacent
to active extract

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
102
Lyophilised fermentation liquors of 31 fungi were produced in addition to 12 extracts
that were made from the mycelial biomass and liquid separation of the fermentation
broth from four isolates. All 43 coprophilous fungi extracts were screened for
antibacterial, antifungal and anti-QS activity.
Antibacterial activity was assessed against S. aureus, E. coli, K. pneumoniae and
P. aeruginosa. In plate-hole diffusion assays, 54.8% of the coprophilous fungi
produced extracts that were capable of inhibiting at least one test bacterium. Six
fungi demonstrated broad-spectrum antibacterial activity, and three fungi (F16EP1,
F14P1 and S5P3) were able to inhibit the growth of all four bacteria. The MIC and
MBC were determined for the 13 extracts that produced inhibition zones greater than
8 mm in the plate-hole diffusion assay. The most effective extracts were produced by
isolates S6W2 and F14P1. Time-course growth assays suggested that the
antibacterial activity of S6W2 against S. aureus and E. coli at 2 × MIC was
bacteriostatic. Under the same conditions, F14P1 demonstrated bacteriostatic
inhibition against E. coli and bactericidal activity against S. aureus.
Antifungal activity was tested for against C. albicans and D. brizae and 8.6% and
38.7%, respectively, of the coprophilous fungi produced extracts that inhibited the
test fungi in agar assays. The MIC was ascertained for extracts against both fungi,
and there were six (19.4%) that were capable of inhibiting visible growth in both the
yeast and filamentous fungi.
The extracts were screened for their ability to inhibit the production of violacein in
C. violaceum, which is under QS control mediated by N-acylhomoserine lactone. In
disk diffusion assays, four (12.9%) of the coprophilous fungi produced extracts that
inhibited pigment production, thus exhibiting anti-QS activity. This is the first time,
to my knowledge, that coprophilous fungi have been screened for anti-QS activity.
The antimicrobial activity exhibited by the extracts in this chapter, while modest, are
encouraging given the high percentage of bioactive isolates from one fermentation

Chapter 4: Screening of Coprophilous Fungi for Antimicrobial Activity
103
method and the production of only crude extracts. Optimisation of the fermentation
conditions and the extraction methodology would likely improve the efficacy of the
extracts and could be considered for further research. This work has suggested that
coprophilous fungi from koala faeces may represent a source of novel antimicrobials
and they warrant further exploration, especially given the paucity of research on this
particular source.
Chapter Five describes the selection of an extract and the scaled-up fermentation
process and bioactivity-guided separation to identify its active compound.

Coprophilous Fungi from Koala Faeces: A Novel Source of Antimicrobial Compounds
104
CHAPTER 5
Bioactivity-Guided Separation of Extract S6W2

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
105
5.1 Introduction
Bioactivity-guided separation is a technique commonly used in natural product drug
discovery. The aim of the process is to isolate compounds with a particular biological
activity from crude extracts. To achieve this, the abundant and often diverse
compounds in the extract must be separated into fractions that are then screened for
the biological activity of interest. Fractions without bioactivity are disregarded and
fractions that demonstrate bioactivity are further separated, often using a variety of
techniques, until the bioactive constituent has been isolated and can be identified.
It is important to note that a standardised nomenclature does not exist when
describing bioactivity-guided separation (Weller, 2012). The separation component
of the phrase may also be referred to as fractionation, which is then guided or
directed by the bioactivity, activity or bioassay (Giltrap et al., 2009; Rodríguez-
Algaba et al., 2015; Seida et al., 2015). These interchangeable terms must all be used
when performing literature searches to ensure that all relevant research is identified.
In this chapter, the scaled-up fermentation liquor of S6W2 was separated using
adsorption to Amberlite® XAD-16 resin, to isolate and enrich the active substance in
solution so that it is more suitable for further purification by preparative reversed-
phase HPLC. The fractions obtained from these isolation and purification procedures
were screened for antibacterial, anti-candidal, antifungal and anti-QS activity using a
combination of the plate-hole diffusion, microdilution and disk diffusion assays.
Identification of the active compound was achieved using ultraviolet (UV)
spectroscopy, mass spectrometry (MS) and NMR spectroscopy analyses.
5.1.1 Chapter aim
The aim of this chapter was to use bioactivity-guided separation to isolate and
identify the active compound from the S6W2 extract.

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
106
5.2 Materials and methods
5.2.1 Liquid fermentation of fungi
Two Erlenmeyer flasks containing 400 mL of PDB were each inoculated with four
1 cm2 agar plugs of the fungal isolate. The flasks were then stoppered with cotton
wool and incubated at 25 °C in the dark with shaking at 150 rpm for 27 days.
Following incubation, the bulk of the mycelium was removed by filtration through
Whatman (No.1) filter paper under vacuum. The fermentation liquor then underwent
membrane filtration using a 0.22 µm filter under vacuum to remove any remaining
fungal cells. The filtrate was dried by lyophilisation to determine its mass, and
bioassays were performed to confirm the presence of the bioactivity that was
originally observed in the smaller scale fermentation liquid.
5.2.2 Amberlite XAD-16 resin
Amberlite® XAD-16 (Rohm and Haas, USA) is a non-ionic polymeric macroporous
adsorbent resin that is supplied as insoluble off-white beads. This is regularly used in
natural product research for the removal or recovery of compounds from an aqueous
solution, such as microbial fermentation liquor, manufacturing waste water or plant
extracts (Li et al., 2008; Pu et al., 2013; Scoma et al., 2012).
Prior to use, one volume of XAD-16 resin was soaked in two volumes of methanol in
a five-volume beaker with occasional stirring for 30 minutes. Distilled water was
then slowly added with stirring to the XAD-16 in methanol. When the beaker had
filled the resin was allowed to settle. Two-thirds of the aqueous methanol supernatant
was then decanted and the addition of distilled water continued until the beaker was
again filled. Two-thirds of the aqueous methanol supernatant was again decanted,
and the process of adding distilled water and decanting was repeated three more
times, so that the aqueous solution was mostly distilled water. The XAD-16 resin
should settle to the bottom of the beaker when the stirring stops. If this procedure of

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
107
wetting the XAD-16 resin is not followed, the resin floats and would not adsorb
hydrophobic material from the aqueous extract very efficiently. The XAD-16 resin
was transferred to a glass chromatography column and further washed with another
10 bed volumes of distilled water, to ensure that all methanol had been removed from
the resin. It was vital that all methanol had been removed from the XAD-16 resin to
ensure the efficient adsorption of hydrophobic compounds from the aqueous extract.
The extract was loaded slowly onto the XAD-16 resin in a glass chromatography
column at a flow rate of approximately one bed volume per hour. The flow-rate was
not allowed to exceed two bed volumes per hour to ensure efficient adsorption of
weakly hydrophobic compounds from the solution. The adsorption of weakly
hydrophobic compounds of interest to the resin can be improved by increasing the
ionic strength of the extract solution through addition of sodium chloride or
ammonium formate to produce a 3–5% salt solution (N. Hart, personal
communication). Following collection of the column loading effluent, the XAD-16
resin was rinsed with distilled water to wash the remaining non-adsorbed material
from the resin. The adsorbed hydrophobic compounds were then eluted from the
XAD-16 resin using 3 to 4 bed volumes of methanol at a flow rate of approximately
5 bed volumes per hour. A final methanol rinse ensured that all adsorbed material
had been eluted from the column. The fractions were collected in glass screw-top
vials, capped and kept at 2–8 °C until they had been analysed by HPLC and
concentrated for bioassays.
The small-scale separation of S6W2 liquor included a preliminary capture step with
the XAD-16 resin in a beaker before pouring the resin into the glass column.About
10 mL of XAD-16 resin was added with gentle stirring to a beaker with 10 mL of
aqueous solution containing 100 mg of the extract. The suspension was gently stirred
at room temperature for one hour to ensure complete adsorption to the XAD-16 resin
before being poured into a glass chromatography column. Elution of the XAD-16
resin was then performed using the procedure for the scaled-up experiment, with the
first distilled water rinse of the beaker being used to transfer any remaining XAD-16

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
108
resin into the column. Rinsing the XAD-16 resin in the column with distilled water
washed the remaining non-adsorbed liquor extract from the resin column effluent.
The adsorbed material was eluted with methanol.
5.2.3 Analytical and preparative reversed-phase HPLC
Fractions were analysed by reversed-phase HPLC using a Waters 600 gradient pump
and the Waters Millennium v4.0 Chromatography Manager Software with an
Alltech Alltima C18 column (150 × 4.6 mm, 5 µm). Isocratic and gradient elution
with acetonitrile/water (ACN/H2O) mobile phase at a flow rate of 1 mL/min was
used. Peak detection was made with a Waters 2996 Photodiode Array UV detector.
Extracts were separated by preparative reversed-phase HPLC using a Waters 600E
gradient pumping system with an Alltech Alltima C18 column (150 × 4.6 mm, 5
µm). Peaks were detected with a Waters 2487 UV detector at λ 254 nm and recorded
on a Curken 250-2 model chart recorder.
5.2.4 Evaporation and drying of fractions
The XAD-16 fractions and the bioactive fractions collected from the preparative
reversed-phase HPLC were evaporated to dryness to determine their mass. Organic
solvents were removed from the fractions using a Heidolph Laborota 4000 rotary
evaporator (Germany) equipped with a dry ice cold trap connected to a Vacuubrand
CVC 2000 II vacuum pump (Germany). The water bath was kept below 40 °C to
avoid the possible degradation of compounds, and the vacuum pressure was
regulated to avoid bumping and loss of product. While water could be removed from
the fractions using the dry ice cold trap attached to the rotary evaporator, large
volumes of water were removed from the aqueous solutions in round-bottom flasks
by lyophilisation on a Telstar Cryodos-50 freeze dryer.

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
109
Following evaporation of fractions to smaller volumes in a round-bottom flask on the
rotary evaporator, the concentrated solutions were transferred to pre-weighed 10 mL
glass screw-top vials. A custom-made adaptor allowed for attachment of the vials
directly to the rotary evaporator vapour duct. This meant that the fractions could be
evaporated to dryness in the vials and immediately capped and weighed when
removed from the rotary evaporator.
5.2.5 Bioassays
Bioassays were carried out to guide the separation of the extract. Fractions that
exhibited bioactivity were further chromatographed to purify the active material.
Antibacterial and anti-candidal activity was assessed using plate-hole diffusion
assays (Sections 4.2.4.1 and 4.2.5.1), anti-QS was determined by disk diffusion
(Section 4.2.6) and antifungal activity was assessed using microdilution assays
Section 4.2.5.2). When the mass of a fraction was known (after drying in pre-
weighed glass vials; Section 5.2.4), it was reconstituted in MilliQ water or aqueous
methanol to a concentration of 12.5 mg/mL for the bioassays. Aqueous methanol was
used as the control. The final concentration range in the antifungal microdilution
assays was 12.5–0.39 mg/mL. All bioassays were performed in duplicate.
The preparative reversed-phase HPLC fractions were tested after they had been
concentrated to approximately 2 mL. In these bioassays, negative controls consisted
of 5% ACN/H2O and 50% ACN/H2O that had undergone the same drying process as
the fractions. Antifungal activity of the fractions against D. brizae was assessed
using the microdilution assay described in 4.2.5.2, with the following adjustment. To
duplicate wells, 100 µL of PDB and 50 µL of each fraction were combined with
50 µL of the D. brizae suspension. Following incubation, the growth of fungi in the
presence of each fraction was compared with growth control wells that did not
contain any fraction.

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
110
The MIC was determined for the final isolated compound against the three
susceptible test bacteria and the yeast using the antibacterial and anti-candidal
microdilution assays described in Sections 4.2.4.2 and 4.2.5.1, respectively. To
reduce the amount of isolated material that was needed in these assays, the method
was modified as follows. Instead of adding 50 µL of a 100 mg/mL solution of the
extract (5 mg) to the first microtitre well, 50 µL of a 12.5 mg/mL solution of the
fraction (625 µg) was added. This produced a final concentration range of 1,560–
12 µg/mL. The MIC of the isolated compound against D. brizae was determined
using the microdilution method described in Section 4.2.5.2.
5.2.6 Identification of active compound
5.2.6.1 Mass spectrometry
Gas chromatography mass spectrometry (GC-MS) was performed with a
ThermoQuest TRACE DSQ GC mass spectrometer in the positive ion mode with an
ionisation energy of 70 eV. GC was performed with a SGE BPX5 (15 m × 0.1 mm
ID, 0.1 µm film thickness) using a temperature program of 40 °C for 2 minutes, then
increasing the temperature by 25 °C per minute to 300 °C after which the
temperature was held for 17.6 minutes with a split ratio of 10, an injector
temperature of 280 °C and the transfer line was set to 280 °C. High-purity helium
was used as carrier gas with a flow rate of 1 mL/min.
Liquid chromatography mass spectrometry (LC-MS) electrospray ionisation (ESI)
analyses were conducted on a Shimadzu liquid chromatography mass spectrometer–
2010EV quadrupole mass spectrometer equipped with a solvent pump and Mercury
MS C18 column (3 µm, 20 × 2.0 mm) using a gradient solvent of 5% to 100% ACN
in water (0.1% formic acid) with a flow rate of 0.3 mL/min. The analyses were
conducted in both positive and negative ionisation modes with a cone voltage of
50 V and the source maintained at 80 °C.

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
111
5.2.6.2 NMR spectroscopy
The structure of the isolated compound was determined using NMR spectroscopy.
NMR spectra were obtained on a Bruker Av400 NMR spectrometer using the
Topspin software package and deuterium oxide (D2O) as the solvent. Gradient
versions of the 2-dimensional 1H -1H correlated spectroscopy (COSY) and 1H -13C
heteronuclear single quantum coherence (HSQC) experiments were performed using
the standard pulse sequences as supplied by Bruker. The 1H peaks were referenced to
residual solvent resonances (4.81 ppm for D2O) and the 13C NMR chemical shifts
were externally referenced using trimethylsilyl propionic salt in D2O prior to the
sample analysis.
The sodium salt of fusaric acid was prepared as a reference compound for NMR
studies. Fusaric acid (Sigma; 10 mg) was dissolved in D2O (0.5 mL) containing
sodium deuteroxide (NaOD) (Sigma; 2.23 mg).
5.3 Results and discussion
5.3.1 Selection of extract for bioactivity-guided separation
The preparation of extracts from 31 coprophilous fungi and the antimicrobial
screening of the extracts are described in Chapter Four. The extracts produced from
isolates S6W2 and F14P1 demonstrated the greatest inhibitory activity. However, the
yield of lyophilized liquor from F14P1 was small and S6W2 was therefore chosen
for bioactivity-guided separation to isolate the active constituent. Table 5.1 shows a
summary of the bioactivity of the liquor extract from S6W2. The extract exhibited
broad spectrum antibacterial activity and was capable of inhibiting the test yeast and
filamentous fungi. In addition, the extract inhibited violacein production in
C. violaceum, which signifies QS quenching.
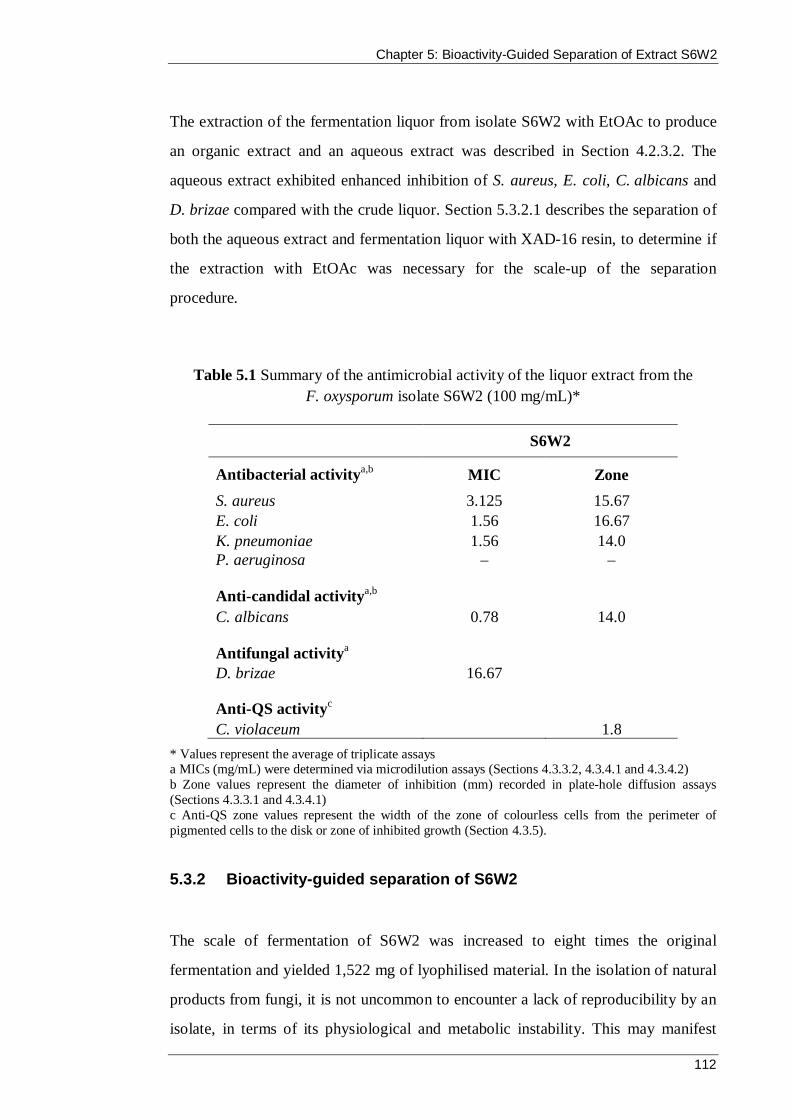
Chapter 5: Bioactivity-Guided Separation of Extract S6W2
112
The extraction of the fermentation liquor from isolate S6W2 with EtOAc to produce
an organic extract and an aqueous extract was described in Section 4.2.3.2. The
aqueous extract exhibited enhanced inhibition of S. aureus, E. coli, C. albicans and
D. brizae compared with the crude liquor. Section 5.3.2.1 describes the separation of
both the aqueous extract and fermentation liquor with XAD-16 resin, to determine if
the extraction with EtOAc was necessary for the scale-up of the separation
procedure.
Table 5.1 Summary of the antimicrobial activity of the liquor extract from the F. oxysporum isolate S6W2 (100 mg/mL)*
S6W2
Antibacterial activitya,b MIC Zone S. aureus 3.125 15.67 E. coli 1.56 16.67 K. pneumoniae 1.56 14.0 P. aeruginosa – –
Anti-candidal activitya,b C. albicans 0.78 14.0
Antifungal activitya D. brizae 16.67
Anti-QS activityc C. violaceum 1.8
* Values represent the average of triplicate assays a MICs (mg/mL) were determined via microdilution assays (Sections 4.3.3.2, 4.3.4.1 and 4.3.4.2) b Zone values represent the diameter of inhibition (mm) recorded in plate-hole diffusion assays (Sections 4.3.3.1 and 4.3.4.1) c Anti-QS zone values represent the width of the zone of colourless cells from the perimeter of pigmented cells to the disk or zone of inhibited growth (Section 4.3.5).
5.3.2 Bioactivity-guided separation of S6W2
The scale of fermentation of S6W2 was increased to eight times the original
fermentation and yielded 1,522 mg of lyophilised material. In the isolation of natural
products from fungi, it is not uncommon to encounter a lack of reproducibility by an
isolate, in terms of its physiological and metabolic instability. This may manifest

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
113
itself in a change in bioactivity after a scaled-up fermentation or even result in the
complete loss of activity (Martin et al., 2006). Therefore, before any separation
efforts, the bioactivity of the scaled-up fermentation liquor was tested to confirm that
it had reproduced a similar level of inhibitory activity that was observed by the
original smaller scale fermentation (Table 5.2). Both fermentation liquors were
assayed at 12.5 mg/mL because this this was the concentration of the fractions in the
bioassays that guided their separation.
Isolation of the active compound from the scaled-up S6W2 liquor was carried out
using adsorption to XAD-16 resin and preparative reversed-phase HPLC. A
summary of the separation is shown in Figure 5.1.
Table 5.2 Confirmation of bioactivity in scaled-up S6W2 liquor (12.5 mg/mL)*
Organism Original S6W2 liquor Scaled-up S6W2 liquor
S. aureusa 9.33 9.0 E. colia 11.0 11.33 K. pneumoniaea 8.0 8.0 P. aeruginosaa – – C. albicansa 7.67 7.67 D. brizaeb 16.67 16.67 C. violaceumc 1.0 1.0
* Results are presented as the average of triplicate assays a Values represent the zone of inhibition (mm) in plate-hole diffusion assays bAntifungal activity against D. brizae was determined by microdilution assays and the values represent the MIC (mg/mL); The MIC of the original liquor was presented in Chapter Four (Table 4.7) c Anti-QS activity against C. violaceum measured the width (mm) of the zone of colourless cells from the perimeter of pigmented cells to the disk or zone of inhibited growth.

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
114
Figure 5.1 Bioactivity-guided separation and chromatography of S6W2 liquor from the scaled-up fermentation.
5.3.2.1 Preliminary investigation of separation methodology
Before the scaled-up fermentation of isolate S6W2, smaller amounts of the crude
fermentation liquor and the aqueous extract following extraction with EtOAc were
applied to XAD-16 resin to assess the adsorption of their active constituents. XAD-
16 resin was used to separate each extract into two fractions: the non-adsorbed
inorganic salts and highly polar compounds in the effluent, and the hydrophobic
compounds that were adsorbed by the XAD-16 resin and subsequently eluted with
methanol.
The fermentation liquor and aqueous extract (100 mg in 10 mL of water) were both
chromatographed on XAD-16 resin in a glass chromatography column (70 × 15 mm,
12 mL bed volume). Fraction collection was guided by observing the colour of the
S6W2 fermentation liquor
Step 1: Separation XAD-16 Resin
1350 mg
Effluent 1036 mg
Eluate 204 mg
Step 2: Chromatography Preparative reversed-phase HPLC
185 mg
Fraction 12-14 42.2 mg

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
115
effluent coming from the column; there appeared to be no difference in this respect
between the liquor and the aqueous extract. The initial dark pink effluent (8 mL) was
denoted the column effluent, followed by elution with distilled water (15 mL) that
rinsed the remaining non-adsorbed material from the XAD-16 resin. Elution of the
column with methanol produced a yellow eluate (13 mL) that contained most of the
adsorbed material; further elution with methanol (23 mL) ensured all the adsorbed
material was stripped from the XAD-16 resin. The difference in colour between the
effluent and eluate fractions can be seen in Figure 5.2.
Figure 5.2 Fractions collected from the separation of the aqueous extract of isolate S6W2 on XAD-16 resin: a) Initial column effluent that passed through the XAD-16 resin without interaction; b) first water wash; c) second water wash; d) methanol eluate; e) final methanol eluate.
The effluent fractions were combined and dried by lyophilisation. The methanol
eluate fractions were combined and evaporated to dryness in a pre-weighed screw-
top vial using the rotary evaporator. The dried methanol eluate residues were
dissolved in 5% (v/v) methanol/water and the lyophilised effluent residues were
reconstituted in MilliQ water. Bioassays of the column effluent and eluate of both the
liquor and the aqueous extracts showed that the active compounds had been adsorbed
by the XAD-16 resin and were eluted by the methanol in the eluate fractions (Table
5.3). The original extracts (prior to separation with XAD-16) were also assayed at
a. b. c. d. e.
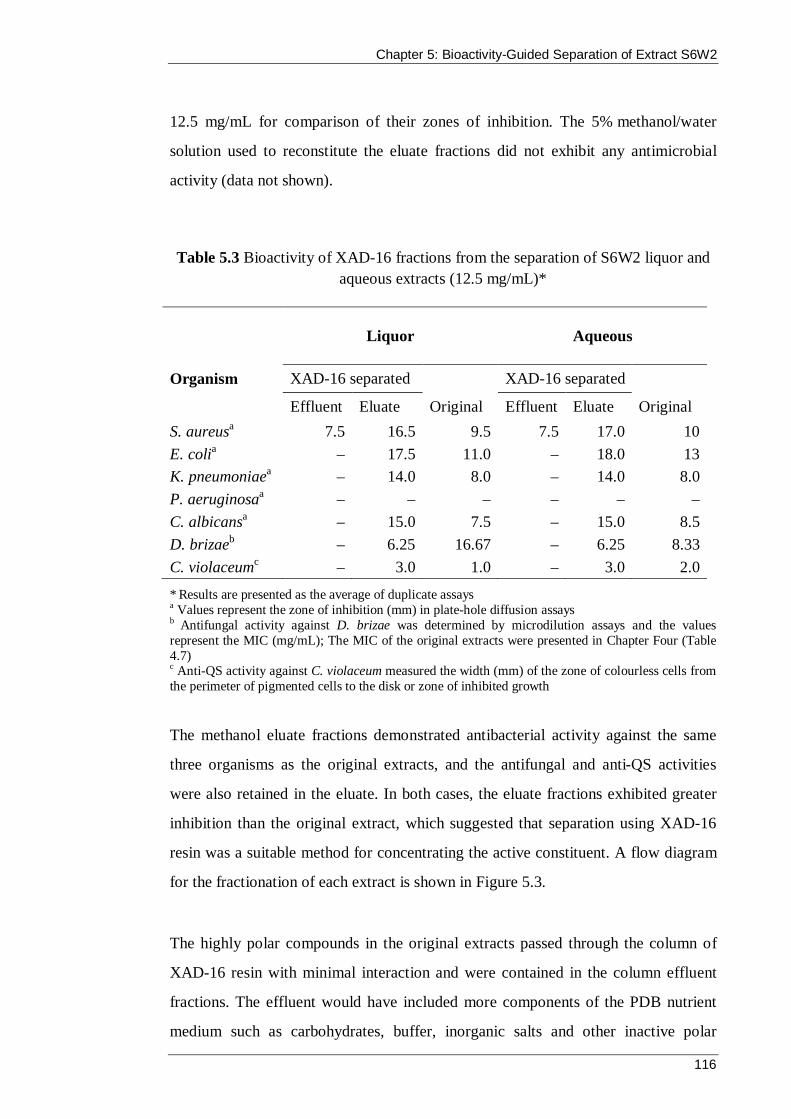
Chapter 5: Bioactivity-Guided Separation of Extract S6W2
116
12.5 mg/mL for comparison of their zones of inhibition. The 5% methanol/water
solution used to reconstitute the eluate fractions did not exhibit any antimicrobial
activity (data not shown).
Table 5.3 Bioactivity of XAD-16 fractions from the separation of S6W2 liquor and aqueous extracts (12.5 mg/mL)*
Liquor Aqueous
Organism XAD-16 separated XAD-16 separated
Effluent Eluate Original Effluent Eluate Original S. aureusa 7.5 16.5 9.5 7.5 17.0 10 E. colia – 17.5 11.0 – 18.0 13 K. pneumoniaea – 14.0 8.0 – 14.0 8.0 P. aeruginosaa – – – – – – C. albicansa – 15.0 7.5 – 15.0 8.5 D. brizaeb – 6.25 16.67 – 6.25 8.33 C. violaceumc – 3.0 1.0 – 3.0 2.0
* Results are presented as the average of duplicate assays a Values represent the zone of inhibition (mm) in plate-hole diffusion assays b Antifungal activity against D. brizae was determined by microdilution assays and the values represent the MIC (mg/mL); The MIC of the original extracts were presented in Chapter Four (Table 4.7) c Anti-QS activity against C. violaceum measured the width (mm) of the zone of colourless cells from the perimeter of pigmented cells to the disk or zone of inhibited growth
The methanol eluate fractions demonstrated antibacterial activity against the same
three organisms as the original extracts, and the antifungal and anti-QS activities
were also retained in the eluate. In both cases, the eluate fractions exhibited greater
inhibition than the original extract, which suggested that separation using XAD-16
resin was a suitable method for concentrating the active constituent. A flow diagram
for the fractionation of each extract is shown in Figure 5.3.
The highly polar compounds in the original extracts passed through the column of
XAD-16 resin with minimal interaction and were contained in the column effluent
fractions. The effluent would have included more components of the PDB nutrient
medium such as carbohydrates, buffer, inorganic salts and other inactive polar
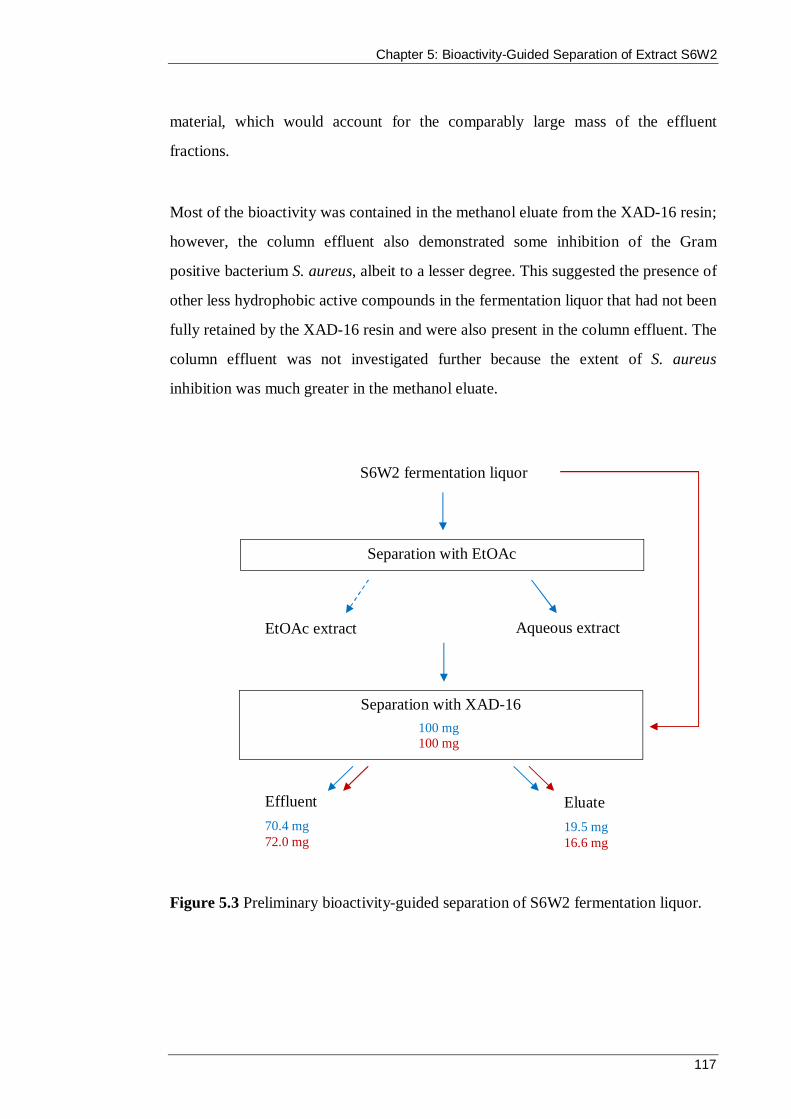
Chapter 5: Bioactivity-Guided Separation of Extract S6W2
117
material, which would account for the comparably large mass of the effluent
fractions.
Most of the bioactivity was contained in the methanol eluate from the XAD-16 resin;
however, the column effluent also demonstrated some inhibition of the Gram
positive bacterium S. aureus, albeit to a lesser degree. This suggested the presence of
other less hydrophobic active compounds in the fermentation liquor that had not been
fully retained by the XAD-16 resin and were also present in the column effluent. The
column effluent was not investigated further because the extent of S. aureus
inhibition was much greater in the methanol eluate.
Figure 5.3 Preliminary bioactivity-guided separation of S6W2 fermentation liquor.
S6W2 fermentation liquor
Separation with EtOAc
Separation with XAD-16 100 mg 100 mg
Aqueous extract EtOAc extract
Effluent 70.4 mg 72.0 mg
Eluate 19.5 mg 16.6 mg
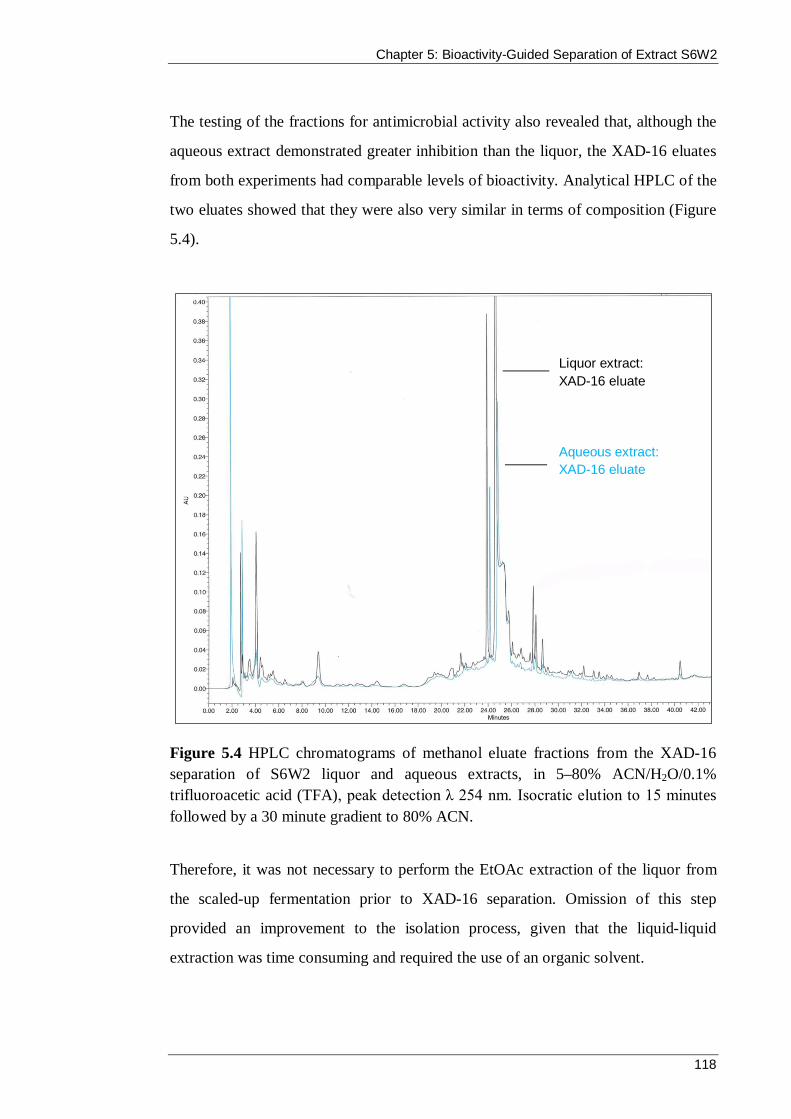
Chapter 5: Bioactivity-Guided Separation of Extract S6W2
118
The testing of the fractions for antimicrobial activity also revealed that, although the
aqueous extract demonstrated greater inhibition than the liquor, the XAD-16 eluates
from both experiments had comparable levels of bioactivity. Analytical HPLC of the
two eluates showed that they were also very similar in terms of composition (Figure
5.4).
Figure 5.4 HPLC chromatograms of methanol eluate fractions from the XAD-16 separation of S6W2 liquor and aqueous extracts, in 5–80% ACN/H2O/0.1% trifluoroacetic acid (TFA), peak detection λ 254 nm. Isocratic elution to 15 minutes followed by a 30 minute gradient to 80% ACN.
Therefore, it was not necessary to perform the EtOAc extraction of the liquor from
the scaled-up fermentation prior to XAD-16 separation. Omission of this step
provided an improvement to the isolation process, given that the liquid-liquid
extraction was time consuming and required the use of an organic solvent.
Aqueous extract: XAD-16 eluate
Liquor extract: XAD-16 eluate

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
119
5.3.2.2 Stage 1 Separation – Amberlite XAD-16 resin
Having established that extraction of the S6W2 liquor with EtOAc was not necessary
to concentrate the bioactivity, the first step of the separation was solid phase
extraction using XAD-16 resin.
XAD-16 resin in a glass chromatography column (70 × 30 mm, bed volume 49 mL)
was equilibrated with 10 bed volumes of distilled water before loading the liquor
extract (1,350 mg in 100 mL). The resulting effluent (80 mL) displayed the same
pink colour that was observed in the smaller scale separation, and this was combined
with the effluent from a water rinse (200 mL). The adsorbed material was eluted with
methanol (400 mL), yielding 204 mg of crude product on evaporating the eluate to
dryness using a rotary evaporator.
Table 5.4 Bioactivity of the effluent and eluate fractions from the separation of S6W2 liquor using XAD-16 resin (12.5 mg/mL)*
Organism Effluent Eluate
S. aureusa 7.0 16.0 E. colia – 17.5 K. pneumoniaea – 14.0 P. aeruginosaa – – C. albicansa – 15.5 D. brizaeb – 6.25 C. violaceumc – 3.0
* Results are presented as the average of duplicate assays a Values represent the zone of inhibition (mm) in plate-hole diffusion assays b Antifungal activity against D. brizae was determined by microdilution assays and the values represent the MIC (mg/mL) c Anti-QS activity against C. violaceum measured the width (mm) of the zone of colourless cells from the perimeter of pigmented cells to the disk or zone of inhibited growth.

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
120
Bioassays of the methanol eluate confirmed that the activity was contained in this
adsorbed fraction and the strength of inhibition was equivalent to the XAD-16 eluate
from the smaller scale separation of the S6W2 liquor. A sample of the effluent was
also tested to verify that it contained only inactive material, with the exception of
minor antibacterial activity against S. aureus (Table 5.4).
5.3.2.3 Stage 2 chromatography of S6W2 – preparative reversed-
phase HPLC
The XAD-16 eluate of S6W2 was further separated by reversed-phase HPLC using a
preparative Alltima C18 column that had been pre-equilibrated with 5% ACN/H2O.
The sample (185 mg) was dissolved in 5% ACN/H2O (2 mL) and loaded onto the
column at a flow rate of 8 mL/min. The column was eluted 5% ACN/H2O at a flow-
rate of 8 mL/min for 5 minutes, after which the flow-rate was increased to
10 mL/min for a further 15 minutes. The column eluate was monitored using a UV
detector at λ 254 nm and was collected in 25 mL screw top vials. The fraction
collections were marked on the chart paper recording the detector response. After 20
minutes, a linear solvent gradient to 100% ACN over 60 minutes at flow rate
10 mL/min was applied, which gradually eluted all the material injected onto the
column. A total of 65 fractions were collected. Each fraction was concentrated to
approximately 2 mL by rotary evaporation and tested for antimicrobial activity in
duplicate. To conserve material, antibacterial assays were performed against only
two bacteria, S. aureus and E. coli. In addition, an adjusted antifungal assay (see
Section 5.2.5) was carried out against D. brizae that required a smaller volume of
fraction compared with the serial dilution method. Fractions 12 and 13 were the only
fractions that exhibited bioactivity, and both demonstrated antibacterial, anti-
candidal, antifungal and anti-QS activity (Table 5.5).

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
121
Table 5.5 Bioactivity of fractions 12 and 13 from the Stage 2 Separation of S6W2 liquor using preparative reversed-phase HPLC*
Organism Fraction 12 Fraction 13
S. aureusa 26.0 8.0 E. colia 33.0 8.5 C. albicansa 38.5 9.0 D. brizaeb No growth No growth C. violaceumc 6.5 3.0
* Unknown concentration; results are presented as the average of duplicate assays a Values represent the zone of inhibition (mm) in plate-hole diffusion assays b The fractions were tested without serial dilution by an adjusted antifungal assay c Anti-QS activity against C. violaceum measured the width (mm) of the zone of colourless cells from the perimeter of pigmented cells to the disk or zone of inhibited growth.
The collection of fraction 12 corresponded with the detection of a broad peak at
retention time 35 minutes, which was initially thought to be a group of poorly
resolved peaks. Fractions 13 and 14 were collected at the tail end of the broad peak.
Analytical HPLC of fractions 12, 13 and 14 showed that they all had a major peak at
retention time 24.5 minutes, which is the same as that of the XAD eluate peak in the
analytical HPLC chromatogram of the methanol eluate from the XAD-16 separation
shown in Figure 5.4. The overlay of the HPLC chromatograms of fractions 12, 13
and 14 presented in Figure 5.5, shows that fraction 12 contained the same
compounds as present in fractions 13 and 14, and that most of the material had been
eluted in fraction 12. Fraction 13 demonstrated the same pattern of bioactivity as
fraction 12, albeit on a lower scale of activity. It is likely that fraction 14 would have
produced similar results upon further concentration. HPLC analysis of the other main
fractions did not reveal a peak with retention time 24.5 minutes. Thus, fractions 12,
13 and 14 were pooled, evaporated to dryness in a pre-weighed glass screw-top vial
on the rotary evaporator and weighed (42.2 mg). The combined fractions were
designated F12–14.

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
122
Figure 5.5 An overlay of analytical HPLC chromatograms of fractions 12, 13 and 14 from the preparative reversed-phase HPLC of S6W2 liquor, at λ 215 nm. Isocratic elution with 5%ACN/H2O/0.1% TFA for 15 minutes followed by a 30 minute gradient to 80% ACN/H2O/0.1% TFA. The three fractions were combined and designated F12–14.
The bioactivity of F12–14 was assayed at a concentration of 12.5 mg/mL against all
microorganisms (Table 5.6), for comparison of bioactivity with the XAD-16 eluate
fraction and original liquor activity (Table 5.3). In addition, the MIC was also
determined.
The inhibition zones in the antibacterial and anti-candidal assays were slightly
reduced compared with those produced by the concentrated preparative HPLC
fraction 12, which suggested that the concentration of the active compound was
higher in the 2 mL fraction than in the 12.5 mg/mL solution. This was consistent
with the recording of a combined weight of fractions 12, 13 and 14 of 42.2 mg, and
the vast majority of material being eluted into fraction 12. Bioactivity-guided
Fraction 12
Fraction 13
Fraction 14

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
123
separation of the S6W2 liquor increased the antibacterial inhibition zones by an
average of 62.4% (standard deviation 2.5). The anti-candidal inhibition zone was
increased by 40%, and the anti-QS zone, which was measured on only one side of the
paper disk, was increased by 36%. The MICs against the test bacteria and yeast were
reduced more than fifteen-fold and the fraction was capable of inhibiting the growth
of D. brizae at a concentration of only 2% of the original liquor MIC against the
fungus.
Table 5.6 Bioactivity of F12–14 from the preparative chromatography of S6W2*
Organism F12–14
Zone (mm) c MIC (µg/mL) S. aureusa 24.5 195 E. colia 28.0 98 K. pneumoniaea 22.0 98 P. aeruginosa – – C. albicansa 35.0 49 D. brizae n/a 390 C. violaceumb 5.0 n/a
* Results are presented as the average of duplicate assays a Antibacterial and anti-candidal activity were determined by plate-hole diffusion assays b Anti-QS activity against C. violaceum measured the width (mm) of the zone of colourless cells from the perimeter of pigmented cells to the disk or zone of inhibited growth c The fraction was tested at 12.5 mg/mL.
HPLC analysis of F12–14 under isocratic conditions showed a strong sharp peak at
20.15 minutes followed by two weaker broad shoulders of decreasing absorbance at
21.9 minutes and 23 minutes (Figure 5.6). The spectral data of signals from the
photodiode array detector were collected, and the UV profile of the main peak was
compared with the profile of the broad shoulder at 21.9 minutes (Figure 5.7). Both
peaks had absorption maxima at λ 200.1, 227.1 and 272.0 nm, which strongly
suggested that they were the same compound.

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
124
Figure 5.6 Analytical reversed-phase HPLC chromatogram of F12–14 from the preparative reversed-phase HPLC of S6W2 liquor, using 10% ACN/H2O/0.1% TFA at λ 254 nm.
Figure 5.7 Diode array UV profile (λ 190–400 nm) of the main peak of F12–14 at 20.15 minutes and of the first shoulder at 21.9 minutes.
Main peak (20.15 minutes)
Shoulder (21.90 minutes)

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
125
Ultraviolet–visible spectroscopy can be helpful in the analysis of natural products if
they contain functional groups with absorption frequencies in the easily accessible
region of the UV spectrum (190–400 nm). Absorption in this region usually signifies
the presence of polyunsaturated chromophores in the compound, such as carbonyl,
carboxyl, carboxylic acid, amide functional groups and aromatic ring systems
(Larsen and Hansen, 2008). The strong absorbance at λ 272 nm in the UV spectrum
of F12–14 indicated the presence of a carboxyl group in conjugation with a double
bond or attached to an aromatic ring.
The reproducible broad shoulders on the tail of the main peak seen in the HPLC
analysis of F12–14 (Figure 5.6) are typical of a compound such as a weak acid that is
in a state of partial protonation or ionisation in the acid and salt forms in the acidic
mobile phase being used (N. Hart, personal communication). A smaller injection
volume (5 µL instead of 20 µL) of F12–14 produced only the two broad peaks and
was missing the sharp, faster eluting peak (Figure 5.8), which indicated that the
acidic moiety in the smaller injected amount had been fully protonated and the faster
eluting salt form was absent.
HPLC analysis of fraction F12–14 using an alkaline mobile phase (10% ACN/0.02M
disodium phosphate (Na2HPO4), pH 9) resulted in the compound eluting much faster
as a single peak at 12 minutes (Figure 5.8). Because the salt forms of ionisable
compounds elute faster than the protonated forms, this observation confirmed the
presence of an acidic group in F12–14.
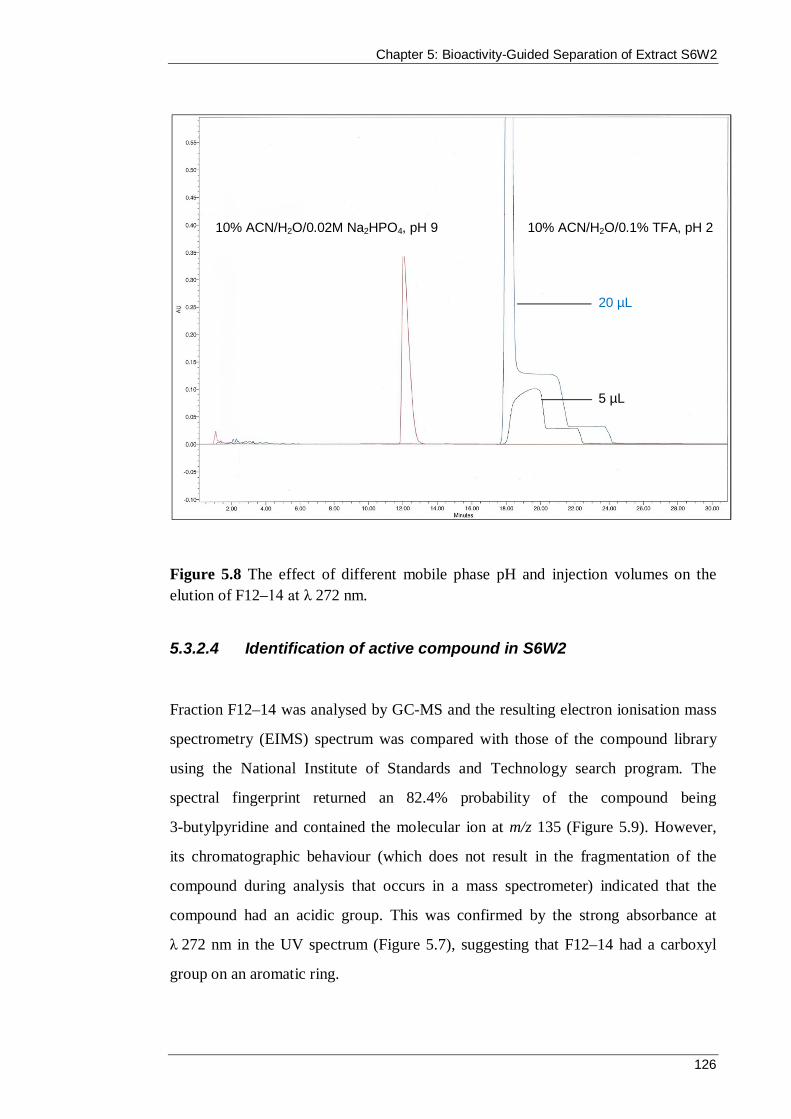
Chapter 5: Bioactivity-Guided Separation of Extract S6W2
126
Figure 5.8 The effect of different mobile phase pH and injection volumes on the elution of F12–14 at λ 272 nm.
5.3.2.4 Identification of active compound in S6W2
Fraction F12–14 was analysed by GC-MS and the resulting electron ionisation mass
spectrometry (EIMS) spectrum was compared with those of the compound library
using the National Institute of Standards and Technology search program. The
spectral fingerprint returned an 82.4% probability of the compound being
3-butylpyridine and contained the molecular ion at m/z 135 (Figure 5.9). However,
its chromatographic behaviour (which does not result in the fragmentation of the
compound during analysis that occurs in a mass spectrometer) indicated that the
compound had an acidic group. This was confirmed by the strong absorbance at
λ 272 nm in the UV spectrum (Figure 5.7), suggesting that F12–14 had a carboxyl
group on an aromatic ring.
10% ACN/H2O/0.1% TFA, pH 2 10% ACN/H2O/0.02M Na2HPO4, pH 9
20 µL
5 µL

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
127
Figure 5.9 The EIMS of F12–14 and of the top match chemical structure 3-butylpyridine in the National Institute of Standards and Technology spectral database.
LC-MS (ESI) analysis of F12–14 produced a different molecular ion and
fragmentation pattern (Figure 5.10) to that of the EIMS (Figure 5.9). It was
determined that the peak at m/z 179.9 [M]+ represented the aforementioned 3-
butylpyridine with an attached carboxylic acid functional group.
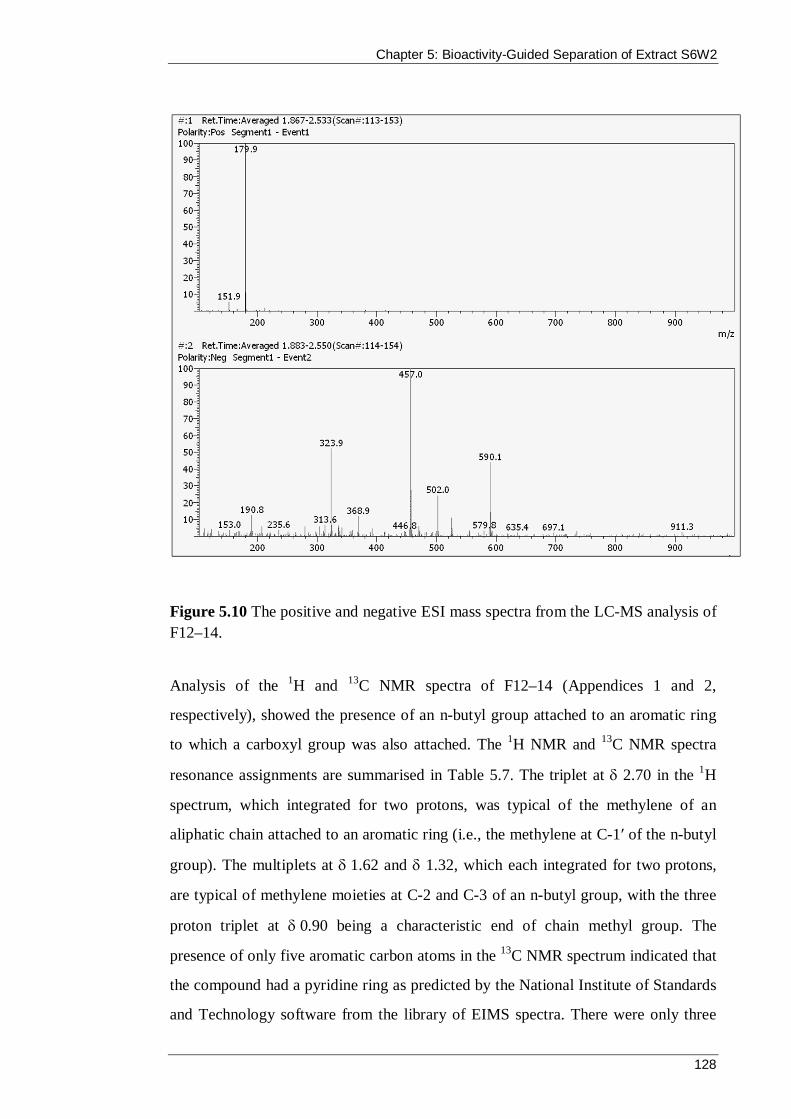
Chapter 5: Bioactivity-Guided Separation of Extract S6W2
128
Figure 5.10 The positive and negative ESI mass spectra from the LC-MS analysis of F12–14.
Analysis of the 1H and 13C NMR spectra of F12–14 (Appendices 1 and 2,
respectively), showed the presence of an n-butyl group attached to an aromatic ring
to which a carboxyl group was also attached. The 1H NMR and 13C NMR spectra
resonance assignments are summarised in Table 5.7. The triplet at 2.70 in the 1H
spectrum, which integrated for two protons, was typical of the methylene of an
aliphatic chain attached to an aromatic ring (i.e., the methylene at C-1ʹ of the n-butyl
group). The multiplets at 1.62 and 1.32, which each integrated for two protons,
are typical of methylene moieties at C-2 and C-3 of an n-butyl group, with the three
proton triplet at 0.90 being a characteristic end of chain methyl group. The
presence of only five aromatic carbon atoms in the 13C NMR spectrum indicated that
the compound had a pyridine ring as predicted by the National Institute of Standards
and Technology software from the library of EIMS spectra. There were only three

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
129
aromatic proton signals in the 1H spectrum: The doublets at 7.79 and 7.90 were
coupled and therefore adjacent to each other, and the singlet at 8.45 is indicative of
a proton on a carbon atom adjacent to a pyridine nitrogen and a quaternary carbon
(i.e., it is the methine proton at C-6). This being the only signal in the 1H spectrum of
a proton on a carbon next to the pyridine N atom, it indicated that the carboxyl group
must therefore be attached to the pyridine ring at C-2.
The COSY spectrum of F12–14 (Appendix 3) confirmed the proton–proton
connectivity in the n-butyl chain. There was connectivity between the signals at
7.90 and 7.79 of the protons on C-3 and C-4 of the pyridine ring, although it was
difficult to see because of their close proximity to one another and the poor peak
shape of the C-3 proton signal. The connectivity between the proton on C-4 of the
pyridine ring at 7.79 to a proton on C-1ʹ of the n-butyl group at 2.70, was
evidence of long range allylic coupling between these two protons, which showed
that the n-butyl group was attached to the pyridine ring at C-5. The HSQC spectrum
(Appendix 4) confirmed the connectivity between the assigned carbon and proton
atoms in the n-butyl chain and the connectivity between the carbon and proton atoms
at C-4 of the pyridine ring. These assignments led to the presumptive identification
of the active compound F12–14 as 5-butylpyridine-2-carboxylic acid, which is better
known as fusaric acid (Figure 5.11). Fusaric acid is not very soluble in water, but its
solubility can be improved under acidic or alkaline conditions.
Figure 5.11 Structure of fusaric acid with assigned atom numbering.

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
130
The 1H and 13C NMR spectra of the reference fusaric acid in NaOD/D2O are shown
in Appendices 5 and 6, respectively. Comparison of the 1H and 13C NMR spectra of
F12–14 with those of fusaric acid (Figure 5.12 and Figure 5.13, respectively)
confirmed the identification of F12–14 as fusaric acid. A summary of the 1H and 13C
NMR spectral data of the reference fusaric acid are included in Table 5.7. The small
differences in the chemical shifts and peak multiplicity are due to the differences in
concentration of the NMR samples and to the reference fusaric acid spectra being
acquired in D2O containing NaOD. Although F12–14 appears to be present in a salt
form in the NMR analysis, the bioactive constituent of the S6W2 liquor extract was
identified as fusaric acid. Fusaric acid is an amphoteric compound that is capable of
chelating metals and can also form a salt with monovalent ions, such as a sodium ion
(Na+), available in the fermentation media or in solution during the isolation
process (Bochner et al., 1980; Stack Jr et al., 2004).
Table 5.7 Summary of the 1H (400 MHz) and 13C (100 MHz) NMR spectral data of F12–14 and FA–Na+ in D2O
F12–14 FA–Na+
Position δ 13C in ppm
δ 1H in ppm, Mult.*
(J in Hz)
δ 13C in ppm
δ 1H in ppm, Mult.* (J in Hz)
2 148.26, qC – 148.51, qC – 3 123.70, CH 7.90, d (8.0) 123.53, CH 7.86, d (8.0) 4 137.80, CH 7.79, d (8.0) 137.60, CH 7.76, dd (8.0, 1.8) 5 141.20, qC – 141.04, qC – 6 150.30, CH 8.45, bs 150.55, CH 8.41, bs 7 173.10, qC – 173.18, qC – 1’ 31.73, CH2 2.70, t (7.5) 31.67, CH2 2.68, t (7.6) 2’ 32.37, CH2 1.62, m 32.35, CH2 1.60, m 3’ 21.45, CH2 1.32, m 21.44, CH2 1.31, m 4’ 13.06, CH3 0.90, t (7.4) 13.07, CH3 0.90, t (7.4)
* Mult. means multiplicity; bs = broad singlet; d = doublet; dd = doublet of doublets; m = multiplet; t = triplet; qC = quaternary carbon; CH = methane; CH2 = methylene; CH3 = methyl.
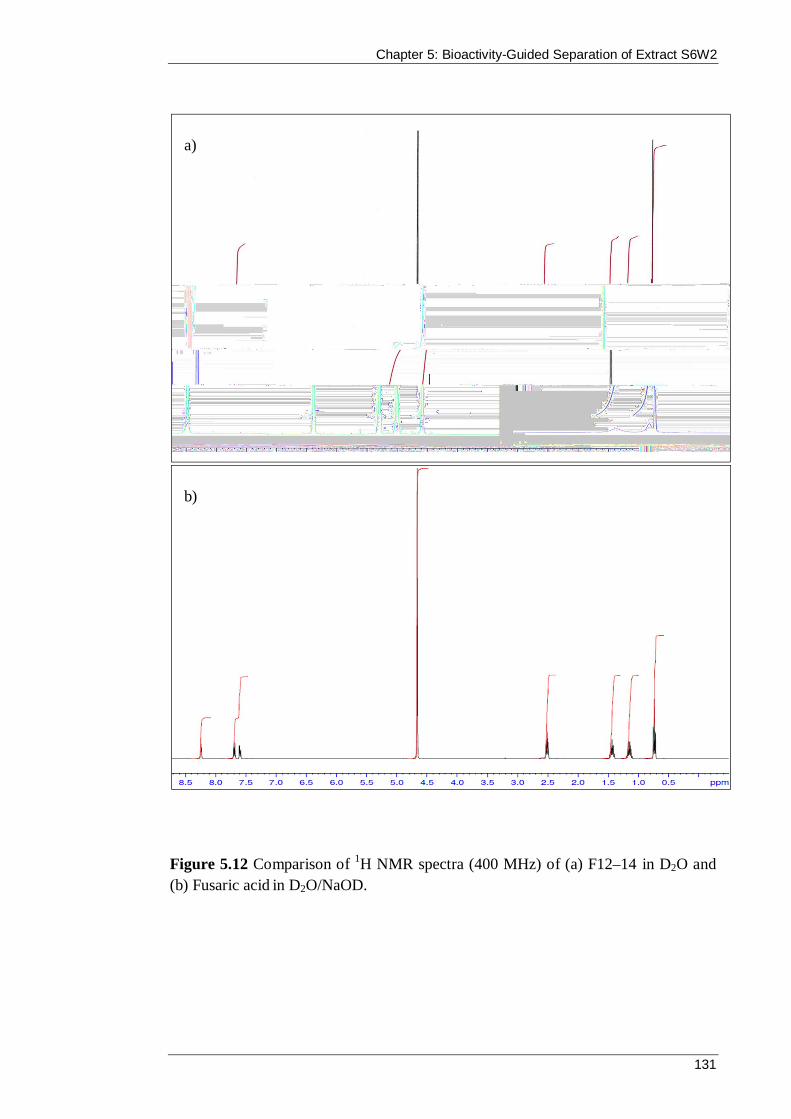
Chapter 5: Bioactivity-Guided Separation of Extract S6W2
131
Figure 5.12 Comparison of 1H NMR spectra (400 MHz) of (a) F12–14 in D2O and (b) Fusaric acid in D2O/NaOD.
a)
b)

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
132
Figure 5.13 Comparison of 13C NMR spectra (100 MHz) of (a) F12–14 in D2O and (b) Fusaric acid in D2O/NaOD.
Fusaric acid is a well-documented fungal toxin that was first isolated by Yabuta et al.
(1934) from Fusarium heterosporum as a compound inhibiting the growth of rice
seedlings. Fusaric acid has subsequently been implicated in a number of plant
diseases and has been isolated from many Fusarium species including
F. moniliforme, F. crookswellense, F. subglutinans, F. sambucinum, F. napiforme,
F. oxysporum, F. solani, F. proliferatum, F. verticillioides, F. sacchari and
a)
b)

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
133
F. fujikuroi (Bacon et al., 1996; Brown et al., 2012; Ghannam et al., 2014; Niehaus et
al., 2014). The production of fusaric acid by pathogenic Fusarium strains plays a role
in the pathogenesis of various wilt, rot and blight diseases in commercially important
plants such as barley, corn, wheat, rice, banana and cottonseed. These lead to
reduced crop yields and contamination of the harvested product, particularly those
used for animal feed (Arias, 1985; Barna et al., 2011; Dong et al., 2012; Rofiat et al.,
2015; Saremi and Okhovvat, 2006; Shimshoni et al., 2013; Stipanovic et al., 2011).
In addition to phytotoxic activity, fusaric acid exhibits diverse biological activities
including anti-oomycete and antifungal (Son et al., 2008), antibacterial (May et al.,
2000; Son et al., 2008), anti-mycobacterial (in metal complexes) (Pan et al., 2011),
acanthamoebicidal (Boonman et al., 2012), nematicidal (Kwon et al., 2007), anti-
hypotensive (Hidaka et al., 1969), dopamine β-hydroxylase inhibition (Goodwin and
Sack, 1974) and anti-tumour activity (Jaglowski and Stack Jr, 2006). Anti-QS
activity has been attributed to fusaric acid, in the repression of the production of QS
signal N-hexanoyl-L-homoserine lactone in Pseudomonas chlororaphis (Van Rij et
al., 2005). Recently, Martín-Rodríguez et al. (2014) identified a marine endophytic
isolate of F. oxysporum that demonstrated QS inhibition against C. violaceum, and it
is likely the active compound was fusaric acid. The compound has also been used to
screen for tetracycline-resistant mutants of E. coli and Salmonella typhimurium
because they are more susceptible to fusaric acid than wild type strains (Herrin Jr et
al., 1982). In this thesis, P. aeruginosa was not susceptible to the extracts from the
F. oxysporum isolate S6W2 or to the isolated fusaric acid. This may be explained by
the presence of a fusaric acid resistance gene in the bacterium, similar to that
reported by Utsumi et al. (1991) in Pseudomonas cepacia and by Ruiz et al. (2015)
in Pseudomonas protegens.
F. oxysporum are commonly isolated from soil, although they have previously been
isolated from faeces (Krug et al., 2004; Peterson et al., 2009). They are considered a
facultative coprophile that can take advantage of the variable nutrients provided by
dung from different herbivores. As discussed in Chapter Three, koalas feed

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
134
exclusively on eucalyptus leaves and their faeces consist of undigested cellulose,
highly lignified fibre and phenolic compounds. Fusarium species are known
producers of the enzymes cellulase, hemicellulase, ligninase and tannase, and are
therefore prime candidates to use the recalcitrant substrate for nutrition (Demain et
al., 2005; Kikot et al., 2009; Murugan et al., 2007). The production of fusaric acid by
the fungus may also confer a competitive advantage to the species by inhibiting the
growth of other fungi in the dung as the compound exhibited antifungal activity in
the bioassays performed in this thesis. Interestingly, antifungal activity by fusaric
acid has only been reported in the literature once. Son et al. (2008) demonstrated
growth inhibition of two plant pathogenic fungi by fusaric acid in an antifungal assay
similar to the radial growth assay described in Section 4.2.5.2.
The isolation of fusaric acid from extract S6W2 supports the hypothesis that
coprophilous fungi are present in the koala faeces and that these fungi are capable of
producing bioactive compounds. Bérdy (2005) summarises the difficulties of
bioactivity-guided separation as follows: “The identification of active compounds
from fermentation samples (extracts, whole broths) is one of the most complicated,
labour intensive, time consuming steps of the screening protocols” (p.23). Since
fusaric acid is a known compound, any efforts to identify active compounds within
the remaining active extracts should be focused on the lesser studied fungi genera,
such as Fimetariella and Anthostomella, to increase the chances of identifying a
novel compound. Furthermore, if resources permit, dereplication strategies that
provide fast and unambiguous identification of previously described compounds
without reference standards should be used (Cordell, 1999). For example, this could
include ultra-HPLC coupled to photodiode array and high-resolution MS in
combination with searching a microbial compound database such as AntiBase
(Wiley-VCH, Germany) with additional in-house reference standards from previous
screening efforts, if available (El-Elimat et al., 2013; Klitgaard et al., 2014; Nielsen
and Larsen, 2015). Recently, a new method was described by Sica et al. (2015) that
eliminates the need for any extract preparation from a fungus, by achieving
identification of secondary metabolites directly on the surface, or surrounding, a
fungal culture in a Petri dish. Continued development in technology will contribute

Chapter 5: Bioactivity-Guided Separation of Extract S6W2
135
to the improvement of natural product drug discovery and aid researchers in finding
novel compounds.
5.4 Summary
Australian coprophilous fungi have yet to be explored in terms of their potential to
produce antimicrobial secondary metabolites. In this chapter, the extract from the
fermentation liquor of F. oxysporum isolate S6W2 was separated to identify the
compound responsible for the observed antibacterial, antifungal and anti-QS
activities. Separation of the extract was achieved by bioactivity-guided fractionation
using adsorption to Amberlite® XAD-16 resin and preparative reversed-phase HPLC
in conjunction with plate-hole diffusion, microdilution and disk diffusion assays.
Following analysis with UV spectroscopy, mass spectrometry and NMR
spectroscopy, and after comparison with a reference standard, the bioactive
compound was identified as fusaric acid.
Fusaric acid is a well-documented mycotoxin of the Fusarium genus and is
implicated in various wilt, blight and rot diseases in commercially important plants.
The compound has also demonstrated a range of biological activities since its
isolation in 1934. In natural product drug discovery research, the main objective is to
find new compounds. The isolation of a known compound in this work highlights the
importance of employing dereplication protocols (if available) to reduce the costly
and time consuming efforts of bioactivity-guided separation. Nonetheless, the
isolation of fusaric acid from extract S6W2 supported the hypothesis that
coprophilous fungi capable of producing bioactive compounds were present in the
koala faeces.

Coprophilous Fungi from Koala Faeces: A Novel Source of Antimicrobial Compounds
136
CHAPTER 6
Conclusion

Chapter 6: Conclusion
137
6.1 Introduction
Fungi have provided numerous chemically and biologically diverse secondary
metabolites that have proved useful in medical and agricultural settings. Given the
increasing resistance of human and plant pathogens to current treatments, it is
essential that previously untapped biological niches are explored for novel microbes
and natural products.
The preceding chapters described the studies undertaken to address the major aims of
the thesis, which were to:
i. isolate and identify fungi from koala faeces
ii. assess the antimicrobial activity of extracts prepared from the fungi
iii. identify the active compounds within the most active extract.
This chapter provides an overview of the major findings of this project as well as
identifying the scope for further research.
6.2 Summary of findings
6.2.1 Chapter Three
Very little research has been carried out on coprophilous fungi in Australia, despite
the important role they play in the ecosystem and their potential for producing
bioactive secondary metabolites. Chapter Three broadened our knowledge about the
community of coprophilous fungi that inhabit koala faeces. Thirty-one fungal isolates
from koala faeces were identified using ITS sequencing, representing only the
second time that fungi from koala faeces have been identified by molecular means
(Peterson et al., 2009). The ITS sequences now form a Popset (population sequence
data) in the NCBI database and serve as a good reference for studying coprophilous
fungi communities in the future. All but one of the isolates were members of the

Chapter 6: Conclusion
138
phylum Ascomycota, a weighted diversity that is common in numerous
Ascomyceteous dominated coprophilous collections in the literature (Bell, 2005;
Gupta, 2010; Melo et al., 2012; Pandey, 2009; Peterson et al., 2009; Richardson,
2001a, c; Watling and Richardson, 2010). Although this work did not reveal any
previously unknown species, the findings are important because they add to the
paucity of research into Australian coprophilous fungi.
6.2.2 Chapter Four
Studies of coprophilous fungi often focus on their taxonomy and their ecology rather
than investigating their secondary metabolites for bioactivity. The limited studies that
do exist suggest that coprophilous fungi are capable of producing bioactive
compounds, and that these are are potentially a source of bioactive compounds that is
on par with their more well-documented counterparts such as the marine and
endophytic fungi (Bills et al., 2013). The reason for this capability and the premise
behind this work is that the fitness of coprophilous fungi is challenged by competing
and invading organisms within koala faeces. To survive, the coprophilous fungi must
produce chemicals to mediate interactions with other organisms. In accordance with
this premise, many of the fungi isolated in this project were, indeed, found to exhibit
antimicrobial activity.
In plate-hole diffusion assays, 54.8% of the fungi produced extracts that were
capable of inhibiting at least one of the test bacteria: S. aureus, E. coli,
K. pneumoniae and P. aeruginosa. Extracts from the fermentation liquor of
A. leucospermi (F16EP1), S. alcina (F14P1) and A. niger (S5P3) isolates were able to
inhibit the growth of all four bacteria. The MICs of the most effective extracts were
determined via microdilution assays and the lowest MIC was exhibited by an extract
of F. oxysporum (S6W2, aqueous). This extract inhibited the growth of S. aureus,
E. coli and K. pneumoniae at a concentration of 1.56 mg/mL, 0.78 mg/mL and
1.56 mg/mL, respectively. Time-course growth assays provided an evaluation of the
antibacterial effect over 24 hours and suggested that the inhibition of S. aureus and

Chapter 6: Conclusion
139
E. coli by S6W2 was bacteriostatic at 2 × MIC. Antifungal activity was assessed
against C. albicans and D. brizae with 8.6% and 38.7%, respectively, of the
coprophilous fungi producing extracts that inhibited the test fungi in agar assays. Six
extracts were capable of inhibiting both fungi in microdilution assays, and the lowest
MICs against C. albicans and D. brizae were produced by F. oxysporum
(0.52 mg/mL) and A. niger (6.25 mg/mL), respectively. An alternative system of
bacterial control is through the disruption of QS signals. In this work, 12.9% of the
coprophilous fungi produced extracts that inhibited violacein production in the
screening bacterium C. violaceum, thus indicating anti-QS activity. This is the first
time that coprophilous fungi have been screened for anti-QS activity.
The antimicrobial activity exhibited by the extracts in this thesis, while modest, are
encouraging given the high percentage of bioactive isolates from one fermentation
method and the production of only crude extracts. This was the first work in which
the antibacterial, antifungal and anti-QS potential of Australian coprophilous fungi
have been explored.
6.2.3 Chapter Five
Isolate S6W2 (F. oxysporum) was chosen to undergo bioactivity-guided separation
based on its adequate extract yield and its moderate antibacterial, antifungal and anti-
QS activity. Separation of the scaled-up extract was achieved via adsorption to
XAD-16 resin and preparative RP-HPLC in conjunction with antimicrobial screening
assays. Following analysis with UV spectroscopy, MS, NMR spectroscopy and
comparison with a reference standard, the bioactive compound was identified as
fusaric acid.
Fusaric acid is produced by many Fusarium species and is most notable for its
implication in the pathogenesis of various wilt, blight and rot diseases in plants of
commercial importance. Additionally, fusaric acid has been reported to exhibit
diverse biological activities including antibacterial, antifungal and anti-QS activity.

Chapter 6: Conclusion
140
The isolation of fusaric acid from extract S6W2 supported the hypothesis that
coprophilous fungi that were capable of producing bioactive compounds were
present in the koala faeces.
6.3 Scope for further research
6.3.1 Immediate source of additional study
Several fungal isolates that warrant further investigation were identified from the
koala faeces. Isolates that exhibited antimicrobial activities but were not studied
further could be cultivated in a scaled-up fermentation and undergo bioactivity-
guided separation to identify the active compounds. The most likely candidates
include F2P2 (F. rabenhorstii), F16EP1 (A. leucospermi), F3W3 (T. hyrcaniae),
F14P1 (S. alcina) and S5P3 (A. niger).
6.3.2 Broadening of the fungal survey
Attempting to increase the diversity of fungi isolated from koala faeces would more
comprehensively address the research aims in two ways. First, a larger collection
would further broaden our understanding of the coprophilous community in koala
faeces, and second, an increased number of fungi for bioactivity screening would
improve the likelihood of uncovering novel bioactive secondary metabolites.
The fungi isolated in this research represent an unknown fraction of the species
present in a collection of faeces from 12 koalas living in two locations at one time of
the year. Given these considerations, an expanded survey of the coprophilous fungi
population could address experimental variables, including adjustment of the drying
time for faeces, changes to incubation temperature, the introduction of light and dark
cycles, and the use of a different medium, such as malt extract agar. For example,
some isolates had difficulty producing confluent growth and did not survive multiple
subculturing, which may have been resolved by changes to the growth medium or to

Chapter 6: Conclusion
141
the incubation temperature. Moist chamber incubation was not performed in this
study because the aim was to isolate fungi that are more likely to be cultivable with
PDB. However, because moist chamber incubation identifies isolates that are able to
rely solely on the faeces for nutrition (Krug, 2004), it may allow the isolation of
different species to those isolated in the agar methods and, therefore, may be worth
performing. In terms of variables relating to the collection of faeces, some studies
have suggested that season and location are likely to affect the diversity of the
coprophilous fungal community (Krug et al., 2004; Kruys and Ericson, 2008; Melo et
al., 2012; Richardson, 2001c). Therefore, although time intensive, a more
comprehensive survey would include many faecal pellets collected at different times
of the year from numerous koalas residing in various locations. These approaches
may identify fungal species that were not found in the current study and may
increase the opportunities for discovery of novel bioactive compounds produced as
secondary metabolites.
6.3.3 Optimisation of fermentation conditions
Modern genome sequencing has shown that many fungi are only capable of
producing a small fraction of their secondary metabolites under standard laboratory
conditions (Spraker and Keller, 2014; Takahashi et al., 2013). Therefore, in this
work, the true potential for natural product discovery in the isolated fungi may have
been underestimated. Activation of silent secondary metabolite gene clusters may
yield novel compounds with interesting chemistries. One method of triggering
production is to systematically alternate cultivation parameters such as the media
composition, temperature, aeration, and addition of enzyme inhibitors or biosynthetic
precursors (Kurtböke, 2012). This technique was termed the OSMAC (one strain –
many compounds) approach by Bode et al. (2002), and it has proven to be a simple
and effective tool to diversify metabolite production and to produce compounds from
silent metabolic pathways (Hewage et al., 2014; Wang et al., 2013; Wei et al., 2010).
Fungi are also capable of photosensing, and the presence (or absence) of light at
differing intensities and wavelengths has been shown to affect the production of

Chapter 6: Conclusion
142
secondary metabolites (Bayram et al., 2008; Castrillo et al., 2013; Schmidt-Heydt et
al., 2010). Another consideration is to take advantage of the role that secondary
metabolites play in interspecies communication, and co-cultivate the target fungus
with another fungus or bacterium also known to inhabit koala faeces. This would
mimic the physiological conditions of the dung related to communication and
competition, and could activate silent gene clusters, leading to the production of
different compounds (Brakhage, 2013; Netzker et al., 2015).
The cultivation parameters in this thesis were consistent for every fungus (PDB in a
flask stoppered with cotton wool and incubated at 25 °C in the dark with shaking at
150 rpm), and it would therefore be valuable to investigate if alternative conditions
could lead to the production of novel antimicrobial compounds.
6.4 Conclusion
The results from the current study provide compelling evidence that coprophilous
fungi from koala faeces produce antimicrobial compounds. Further study should be
carried out to identify the active constituents in the active extracts that did not
undergo bioactivity-guided separation. More broadly, this thesis encourages greater
investigation of fungi from minimally explored biological niches, such as animal
faeces.

References
143
References

References
144
References
Abraham, E.P., and Loder, P.B. (1972). Cephalosporin C. In Cephalosporins and
Penicilins: Chemistry and Biology (New York: Academic Press).
Adonizio, A.L., Downum, K., Bennett, B.C., and Mathee, K. (2006). Anti-quorum
sensing activity of medicinal plants in southern Florida. Journal of Ethnopharmacology
105, 427-435.
Alfatafta, A.A., Gloer, J.B., Scott, J.A., and Malloch, D. (1994). Apiosporamide, a new
antifungal agent from the coprophilous fungus Apiospora montagnei. Journal of Natural
Products 57, 1696-1702.
Anderson, I.C., and Cairney, J.W.G. (2004). Diversity and ecology of soil fungal
communities: Increased understanding through the application of molecular techniques.
Environmental Microbiology 6, 769-779.
Arias, J.A. (1985). Secretory organelle and mitochondrial alterations induced by fusaric
acid in root cells of Zea mays. Physiological Plant Pathology 27, 149-158.
Arivudainambi, U.E., Anand, T.D., Shanmugaiah, V., Karunakaran, C., and Rajendran,
A. (2011). Novel bioactive metabolites producing endophytic fungus Colletotrichum
gloeosporioides against multidrug-resistant Staphylococcus aureus. FEMS Immunology
and Medical Microbiology 61, 340-345.
Bacon, C.W., Porter, J.K., Norred, W.P., and Leslie, J.F. (1996). Production of fusaric
acid by Fusarium species. Applied and Environmental Microbiology 62, 4039-4043.
Baker, A.G., Bhagwat, S.A., and Willis, K.J. (2013). Do dung fungal spores make a
good proxy for past distribution of large herbivores? Quaternary Science Reviews 62,
21-31.

References
145
Barna, B., Jansen, C., and Kogel, K.H. (2011). Sensitivity of barley leaves and roots to
fusaric acid, but not to H2O2, is associated with susceptibility to Fusarium infections.
Journal of Phytopathology 159, 720-725.
Bastias, B.A., Zu, Z.H., and Cairney, J.W.G. (2006). Influence of long-term repeated
prescribed burning on mycelial communities of ectomycorrhizal fungi. New Phytologist
172, 149-158.
Bauer, A.W., and Kirby, W.M. (1966). Antibiotic susceptibility testing by a
standardized single disk method. American Journal of Clinical Pathology 45, 493-496.
Bayram, Ö., Krappmann, S., Ni, M., Jin, W.B., Helmstaedt, K., Valerius, O., Braus-
Stromeyer, S., Kwon, N.J., Keller, N.P., Yu, J.H., et al. (2008). VelB/VeA/LaeA
complex coordinates light signal with fungal development and secondary metabolism.
Science 320, 1504-1506.
Bell, A. (2005). Illustrated Guide to the Coprophilous Ascomycetes of Australia
(Utrecht, The Netherlands: CBS).
Bellemain, E., Carlsen, T., Brochmann, C., Coissac, E., Taberlet, P., and Kauserud, H.
(2010). ITS as an environmental DNA barcode for fungi: An in silico approach reveals
potential PCR bias. BMC Microbiology 10, 189-207.
Ben-Ami, R., Olshtain-Pops, K., Krieger, M., Oren, I., Bishara, J., Dan, M., Wiener-
Well, Y., Weinberger, M., Zimhony, O., Chowers, M., et al. (2012). Antibiotic
exposure as a risk factor for fluconazole-resistant Candida bloodstream infection.
Antimicrobial Agents and Chemotherapy 56, 2518-2523.
Bérdy, J. (2005). Bioactive microbial metabolites: A personal view. Journal of
Antibiotics 58, 1-26.

References
146
Bérdy, J. (2012). Thoughts and facts about antibiotics: Where we are now and where we
are heading. Journal of Antibiotics 65, 385-395.
Bergstrom, J.D., Dufresne, C., Bills, G.F., Nallin-Omstead, M., and Byrne, K. (1995).
Discovery, biosynthesis and of mechanism of action of the zaragozic acids: Potent
inhibitors of squalene synthase. Annual Review of Microbiology 49, 607-639.
Bhagobaty, R.K., and Joshi, S.R. (2012). Antimicrobial and antioxidant activity of
endophytic fungi isolated from ethnomedicinal plants of the "Sacred forests" of
Meghalaya, India. Mikologia Lekarska 19, 5-11.
Bills, G.F., Gloer, J.B., and An, Z. (2013). Coprophilous fungi: Antibiotic discovery and
functions in an underexplored arena of microbial defensive mutualism. Current Opinion
in Microbiology 16, 549-565.
Bills, G.F., Overy, D., Genilloud, O., and Pelaez, F. (2009). Contributions of
pharmaceutical antibiotic and secondary metabolite discovery to the understanding of
microbial defense and antagonism. In Defensive Mutualism in Microbial Symbiosis,
J.F. White, and M.S. Torres, eds. (Boca Raton, USA: CRC Press), pp. 258-285.
Bills, G.F., and Polishook, J.D. (1993). Selection isolation of fungi from dung of
Odocoileus hemionus (mule deer). Nova Hedwigia 57, 195 - 206.
Bloch, P., and Tamm, C. (1981). Isolation and structure of pseurotin A, a microbial
metabolite of Pseudeurotium ovalis Stolk with an unusual heterospirocyclic system.
Helv Chim Acta 64, 304-315.
Bochner, B.R., Huang, H.C., Schieven, G.L., and Ames, B.N. (1980). Positive selection
for loss of tetracycline resistance. Journal of Bacteriology 143, 926-933.
Bode, H.B., Bethe, B., Höfs, R., and Zeeck, A. (2002). Big effects from small changes:
Possible ways to explore nature's chemical diversity. ChemBioChem 3, 619-627.

References
147
Boonman, N., Prachya, S., Boonmee, A., Kittakoop, P., Wiyakrutta, S., Sriubolmas, N.,
Warit, S., and Dharmkrong-At Chusattayanond, A. (2012). In vitro acanthamoebicidal
activity of fusaric acid and dehydrofusaric acid from an endophytic fungus Fusarium sp.
Tlau3. Planta Medica 78, 1562-1567.
Boyraz, N., and zcan, M. (2005). Antifungal effect of some spice hydrols. Fitoterapia
76, 661-665.
Bradner, J.R., Sidhu, R.K., Yee, B., Skotnicki, M.L., Selkirk, P.M., and Nevalainen,
K.M.H. (2000). A new microfungal isolate, Embellisia sp., associated with the Antarctic
moss Bryum argenteum. Polar Biology 23, 730 - 732.
Brakhage, A.A. (2013). Regulation of fungal secondary metabolism. Nature Reviews
Microbiology 11, 21-32.
Breitenstein, W., Chexal, K.K., Mohr, P., and Tamm, C. (1981). Pseurotin B, C, D, and
E. Further new metabolites of Pseudeurotium ovalis Stolk. Helv Chim Acta 64, 379-
388.
Brkljača, R., and Urban, S. (2011). Recent advancements in HPLC-NMR and
applications for natural product profiling and identification. Journal of Liquid
Chromatography and Related Technologies 34, 1063-1076.
Brown, D.W., Butchko, R.A.E., Busman, M., and Proctor, R.H. (2012). Identification of
gene clusters associated with fusaric acid, fusarin, and perithecial pigment production in
Fusarium verticillioides. Fungal Genetics and Biology 49, 521-532.
Bruns, T.D., Arnold, A.E., and Hughes, K.W. (2007). Fungal networks made of
humans: UNITE, FESIN and frontiers in fungal ecology. New Phytologist 177, 586-
588.

References
148
Buatong, J., Phongpaichit, S., Rukachaisirikul, V., and Sakayaroj, J. (2011).
Antimicrobial activity of crude extracts from mangrove fungal endophytes. World
Journal of Microbiology and Biotechnology 27, 3005-3008.
Castrillo, M., García-Martínez, J., and Avalos, J. (2013). Light-dependent functions of
the Fusarium fujikuroi CryD DASH cryptochrome in development and secondary
metabolism. Applied and Environmental Microbiology 79, 2777-2788.
Chapman, A.D. (2009). Numbers of living species in Australia and the world, 2nd edn
(Canberra, A.C.T.: Australian Biological Resources Study).
Che, Y., Araujo, A.R., Gloer, J.B., Scott, J.A., and Malloch, D. (2005). Communiols E-
H: New polyketide metabolites from the coprophilous fungus Podospora communis.
Journal of Natural Products 68, 435-438.
Che, Y., Gloer, J.B., Koster, B., and Malloch, D. (2002). Decipinin A and
decipienolides A and B: New bioactive metabolites from the coprophilous fungus
Podospora decipiens. Journal of Natural Products 65, 916-919.
Che, Y., Gloer, J.B., Scott, J.A., and Malloch, D. (2004a). Communiols A-D: New
mono- and bis-tetrahydrofuran derivatives from the coprophilous fungus Podospora
communis. Tetrahedron Letters 45, 6891-6894.
Che, Y., Gloer, J.B., and Wicklow, D.T. (2004b). Curvicollides A-C: New polyketide-
derived lactones from a sclerotium-colonizing isolate of Podospora curvicolla (NRRL
25778). Organic Letters 6, 1249-1252.
Che, Y., Swenson, D.C., Gloer, J.B., Koster, B., and Malloch, D. (2001).
Pseudodestruxins A and B: New cyclic depsipeptides from the coprophilous fungus
Nigrosabulum globosum. Journal of Natural Products 64, 555-558.

References
149
Chen, D.M., and Cairney, J.W.G. (2002). Investigation of the influence of prescribed
burning on ITS profiles of ectomycorrhizal and other soil fungi at three Australian
sclerophyll forest sites. Mycological Research 106, 532-540.
CLSI (1999). Methods for determining bactericidal activity of antimicrobial agents;
Approved guideline M26-A (Wayne, PA: Clinical and Laboratory Standards Institute).
CLSI (2006). Methods for dilution antimicrobial susceptibility testing for bacteria that
grow aerobically; Approved standard M7-A7 (Wayne, PA.: Clinical and Laboratory
Standards Institute).
Colegate, S., M., and Molyneux, R., J. (2008). An introduction and overview. In
Bioactive Natural Products, S. Colegate, M., and R. Molyneux, J., eds. (Boca Raton,
FL: CRC Press), pp. 1-9.
Cordell, G.A. (1999). Finding the needle in the haystack. the dereplication of natural
product extracts. Pure and Applied Chemistry 71, 1089-1094.
Cork, S.J., and Foley, W.J. (1997). Digestive and metabolic adaptations of arboreal
marsupials for dealing with plant antinutrients and toxins. In Marsupial Biology: Recent
Research, New Perspectives, N. Saunders, and L. Hinds, eds. (Sydney, Australia:
UNSW Press), pp. 204-226.
Cork, S.J., and Hume, I.D. (1983). Microbial digestion in the koala (Phascolarctos
cinereus, Marsupialia), an arboreal folivore. Journal of Comparative Physiology 152,
131-135.
Corrado, M., and Rodrigues, K.F. (2004). Antimicrobial evaluation of fungal extracts
produced by endophytic strains of Phomopsis sp. Journal of Basic Microbiology 44,
157-160.

References
150
Cragg, G.M., and Boyd, M.R. (1996). Drug discovery and development at the National
Cancer Institute: The role of natural products of plant origin. In Medicinal Resources of
the Tropical Forest: Biodiversity and its Importance to Human Health, M.J. Balick, E.
Elisabetsky, and S.A. Laird, eds. (New York, USA: Columbia University Press), pp.
101-132.
Cragg, G.M., Grothaus, P.G., and Newman, D.J. (2014). New horizons for old drugs
and drug leads. Journal of Natural Products 77, 703-723.
Cragg, G.M., and Newman, D.J. (2005). International collaboration in drug discovery
and development from natural sources. Pure and Applied Chemistry 77, 1923-1942.
Cragg, G.M., and Newman, D.J. (2013). Natural products: A continuing source of novel
drug leads. Biochimica et Biophysica Acta - General Subjects 1830, 3670-3695.
Cribb, A.B. (1988). Fungi on kangaroo dung. In Lake Broadwater - The Natural History
of an Inland lake and its Environments, G. Scott, ed. (Toowoomba, Australia: Darling
Downs Institute Press).
Cribb, A.B. (1989). Fungi on wombat dung in Tasmania. Queensland Nature 29, 114-
115.
Cribb, A.B. (1997). Two coprophilous fungi on koala dung. Queensland Nature 35, 4-6.
CSL (2015). Our history: 1940-1959.
Dayan, F.E., Cantrell, C.L., and Duke, S.O. (2009). Natural products in crop protection.
Bioorganic and Medicinal Chemistry 17, 4022-4034.
Demain, A., L., Velasco, J., and Adrio, J., L. (2005). Industrial mycology. In Handbook
of Industrial Mycology, Z. An, ed. (New York: CRC Press), pp. 1-26.

References
151
Demain, A.L. (2009). Antibiotics: Natural products essential to human health.
Medicinal Research Reviews 29, 821-842.
Deslippe, J.R., Hartmann, M., Simard, S.W., and Mohn, W.W. (2012). Long-term
warming alters the composition of Arctic soil microbial communities. FEMS
Microbiology Ecology 82, 303-315.
Dong, X., Ling, N., Wang, M., Shen, Q., and Guo, S. (2012). Fusaric acid is a crucial
factor in the disturbance of leaf water imbalance in Fusarium-infected banana plants.
Plant Physiology and Biochemistry 60, 171-179.
Doveri, F., Pecchia, S., Vergara, M., Sarrocco, S., and Vannacci, G. (2012). A
comparative study of Neogymnomyces virgineus, a new keratinolytic species from
dung, and its relationships with the Onygenales. Fungal Diversity 52, 13-34.
Doveri, F., Sarrocco, S., Pecchia, S., Forti, M., and Vannacci, G. (2010). Coprinellus
mitrinodulisporus, a new species from chamois dung. Mycotaxon 114, 351-360.
Doveri, F., Sarrocco, S., and Vannacci, G. (2013). Studies on three rare coprophilous
plectomycetes from Italy. Mycotaxon 124, 279-300.
El-Elimat, T., Figueroa, M., Ehrmann, B., M., Cech, N., B., Pearce, C., J., and Oberlies,
N., H. (2013). High-Resolution MS, MS/MS, and UV database of fungal secondary
metabolites as a dereplication protocol for bioactive natural products. Journal of Natural
Products 76, 1709-1716.
Endo, A. (2010). A historical perspective on the discovery of statins. Proceedings of the
Japan Academy, Series B Physical and Biological Sciences 86, 484-493.
Espinel-Ingroff, A., and Canton, E. (2007a). Antifungal susceptibility testing of
filamentous fungi. In Antimicrobial Susceptibility Testing Protocols, R. Schwalbe, L.
Steele-Moore, and A. Goodwin, eds. (Boca Raton, USA: CRC Press), pp. 209-241.

References
152
Espinel-Ingroff, A., and Canton, E. (2007d). Antifungal susceptibility testing of yeasts.
In Antimicrobial Susceptibility Testing Protocols R. Schwalbe, L. Steele-Moore, and A.
Goodwin, eds. (Boca Raton, USA: CRC Press), pp. 174-207.
Essig, A., Hofmann, D., Münch, D., Gayathri, S., Künzler, M., Kallio, P.T., Sahl, H.G.,
Wider, G., Schneider, T., and Aebi, M. (2014). Copsin, a novel peptide-based fungal
antibiotic interfering with the peptidoglycan synthesis. Journal of Biological Chemistry
289, 34953-34964.
Fears, R., Aro, E.M., Pais, M.S., and Ter Meulen, V. (2014). How should we tackle the
global risks to plant health? Trends in Plant Science 19, 206-208.
Fehlhaber, H.W., Kogler, H., Mukhopadhyay, T., Vijayakumar, E.K.S., and Ganguli,
B.N. (1988). Structure of aranorosin, a new antibiotic of a novel skeletal type. Journal
of the American Chemical Society 110, 8242-8244.
Felsenstein, J. (1985). Confidence limits on phylogenies: An approach using the
bootstrap. Evolution 39, 783-791.
Fiori, A., and Van Dijck, P. (2012). Potent synergistic effect of doxycycline with
fluconazole against Candida albicans is mediated by interference with iron
homeostasis. Antimicrobial Agents and Chemotherapy 56, 3785-3796.
Fisch, K.M., Bakeer, W., Yakasai, A.A., Song, Z., Pedrick, J., Wasil, Z., Bailey, A.M.,
Lazarus, C.M., Simpson, T.J., and Cox, R.J. (2011). Rational domain swaps decipher
programming in fungal highly reducing polyketide synthases and resurrect an extinct
metabolite. Journal of the American Chemical Society 133, 16635-16641.
Fleming, A. (1929). On the antibacterial action of a Penicillium with special reference
to their use in the isolation of B. influenzae. British Journal of Experimental Pathology
10, 226-236.

References
153
Flores, A.C., Pamphile, J.A., Sarragiotto, M.H., and Clemente, E. (2013). Production of
3-nitropropionic acid by endophytic fungus Phomopsis longicolla isolated from
Trichilia elegans A. JUSS ssp. elegans and evaluation of biological activity. World
Journal of Microbiology and Biotechnology 29, 923-932.
Florey, H.W., Chain, E., Heatley, N.G., Jennings, M.A., Sanders, A.G., Abraham, A.P.,
and Florey, M.E. (1949). Antibiotics (London, UK: Oxford University Press).
Fukiharu, T., Shimizu, K., Nakajima, A., Miyamoto, T., Raut, J.K., and Kinjo, N.
(2015). Coprinopsis igarashii sp. nov., a coprophilous agaric fungus from Hokkaido,
northern Japan. Mycoscience 56, 413-418.
Furiga, L., Lonvaud-Funel, A., Dorignac, G., and Badet, C. (2008). In vitro antibacterial
and anti-adherance effects of natural polyphenolic compounds on oral bacteria. Journal
of Applied Microbiology 105, 1470-1476.
Galagan, J.E., Calvo, S.E., Borkovich, K.A., Selker, E.U., Read, N.O., Jaffe, D.,
FitzHugh, W., Ma, L.J., Smirnov, S., Purcell, S., et al. (2003). The genome sequence of
the filamentous fungus Neurospora crassa. Nature 422, 859-868.
Gamble, W.R., Gloer, J.B., Scott, J.A., and Malloch, D. (1995). Polytolypin, a new
antifungal triterpenoid from the coprophilous fungus Polytolypa hystricis. Journal of
Natural Products 58, 1983-1986.
Ganesh Kumar, C., Mongolla, P., Joseph, J., Nageswar, Y.V.D., and Kamal, A. (2010).
Antimicrobial activity from the extracts of fungal isolates of soil and dung samples from
Kaziranga National Park, Assam, India. Journal de Mycologie Medicale 20, 283-289.
Genilloud, O. (2014). The re-emerging role of microbial natural products in antibiotic
discovery. International Journal of General and Molecular Microbiology 106, 173-188.

References
154
Ghannam, I.A.Y., Roaiah, H.F., Hanna, M.M., El-Nakkady, S.S., and Cox, R.J. (2014).
Identification, Crystal structure and antitumor activity of fusaric acid from the
sugarcane fungal pathogen, Fusarium sacchari. International Journal of Pharmacy and
Technology 6, 6528-6535.
Giltrap, M., Macken, A., McHugh, B., Hernan, R., O' Rourke, K., McGovern, E., Foley,
B., and Davoren, M. (2009). Bioassay-directed fractionation of marine sediment solvent
extracts from the east coast of Ireland. Chemosphere 76, 357-364.
Gloer, J.B. (1995). The chemistry of fungal antagonism and defense. Canadian Journal
of Botany 73, S1265-S1274.
Goh, T.K., Lee, L.L., and Teo, K.C. (2013). A new coprophilous gilmaniella species
from Malaysia. Mycotaxon 125, 235-241.
Gomes, N.G.M., Lefranc, F., Kijjoa, A., and Kiss, R. (2015). Can some marine-derived
fungal metabolites become actual anticancer agents? Marine Drugs 13, 3950-3991.
Gond, S.K., Kharwar, R.N., and White, J.F. (2014). Will fungi be the new source of the
blockbuster drug taxol? Fungal Biology Reviews 28, 77-84.
Gond, S.K., Mishra, A., Sharma, V.K., Verma, S.K., Kumar, J., Kharwar, R.N., and
Kumar, A. (2012). Diversity and antimicrobial activity of endophytic fungi isolated
from Nyctanthes arbor-tristis, a well-known medicinal plant of India. Mycoscience 53,
113-121.
Goodfellow, M. (2010). Selective isolation of Actinobacteria. In Isolation and screening
of secondary metabolites and enzymes, A.T. Bull, and J.E. Davies, eds. (Washington:
ASM Press), pp. 12-27.

References
155
Goodwin, F.K., and Sack, R.L. (1974). Behavioral effects of a new dopamine-β-
hydroxylase inhibitor (fusaric acid) in man. Journal of Psychiatric Research 11, 211-
217.
Gupta, A.K. (2010). Studies on fungi from the dung of some herbivorous inmates of
Prince of Wales Zoological Garden, Lucknow; PhD thesis (Lucknow: University of
Lucknow), pp. 178.
Hauser, D., and Sigg, H.P. (1971a). Isolation and decomposition of sordarin. Helvetica
Chimica Acta 54, 1178-1190.
Hauser, D., and Sigg, H.P. (1971b). Isolierung und Abbau von Sordarin. 1. Mitteilung
über Sordarin. Helvetica Chimica Acta 54, 1178-1190.
Hazalin, N.A.M.N., Ramasamy, K., Lim, S.M., Wahab, I.A., Cole, A.L.J., and Abdul
Majeed, A.B. (2009). Cytotoxic and antibacterial activities of endophytic fungi isolated
from plants at the National Park, Pahang, Malaysia. BMC Complementary and
Alternative Medicine 9.
Hein, S.M., Gloer, J.B., Koster, B., and Malloch, D. (1998). Arugosin F: A new
antifungal metabolite from the coprophilous fungus Ascodesmis sphaerospora. Journal
of Natural Products 61, 1566-1567.
Hein, S.M., Gloer, J.B., Koster, B., and Malloch, D. (2001). Bombardolides: New
antifungal and antibacterial γ,-lactones from the coprophilous fungus Bombardioidea
anartia. Journal of Natural Products 64, 809-812.
Helman, Y., and Chernin, L. (2015). Silencing the mob: Disrupting quorum sensing as a
means to fight plant disease. Molecular Plant Pathology 16, 316-329.
Hensens, O.D., Helms, G.L., Jones, E.T.T., and Harris, G.H. (1995). Structure
elucidation of australifungin, a potent inhibitor of sphinganine N-acyltransferase in

References
156
sphingolipid biosynthesis from Sporormiella australis. Journal of Organic Chemistry
60, 1772-1776.
Hentzer, M., and Givskov, M. (2003). Pharmacological inhibition of quorum sensing for
the treatment of chronic bacterial infections. Journal of Clinical Investigation 112,
1300-1307.
Herrera, J., Poudel, R., and Khidir, H.H. (2011). Molecular characterization of
coprophilous fungal communities reveals sequences related to root-associated fungal
endophytes. Microbial Ecology 61, 239-244.
Herrin Jr, G.L., Russell, D.R., and Bennett, G.N. (1982). A stable derivative of pBR322
conferring increased tetracycline resistance and increased sensitivity to fusaric acid.
Plasmid 7, 290-293.
Hewage, R.T., Aree, T., Mahidol, C., Ruchirawat, S., and Kittakoop, P. (2014). One
strain-many compounds (OSMAC) method for production of polyketides, azaphilones,
and an isochromanone using the endophytic fungus Dothideomycete sp. Phytochemistry
108, 87-94.
Hidaka, H., Nagatsu, T., Takeya, K., Takeuchi, T., and Suda, H. (1969). Fusaric acid, a
hypotensive agent produced by fungi. Journal of Antibiotics 22, 228-230.
Hoffman, A.M., Mayer, S.G., Strobel, G.A., Hess, W.M., Sovocool, G.W., Grange,
A.H., Harper, J.K., Arif, A.M., Grant, D.M., and Kelley-Swift, E.G. (2008).
Purification, identification and activity of phomodione, a furandione from an
endophytic Phoma species. Phytochemistry 69, 1049-1056.
Hofle, G., and Reichenbach, H. (2012). Epothilone, a myxobacterial metabolite with
promising antitumor activity. In Anticancer Agents from Natural Products, G.M. Cragg,
D.G.I. Kingston, and D.J. Newman, eds. (Boca Raton, FL: Taylor and Francis), pp. 513-
573.

References
157
Hong, S.B., Yamada, O., and Samson, R.A. (2014). Taxonomic re-evaluation of black
koji moulds. Applied Microbiology and Biotechnology 98, 555-561.
Huang, W.Y., Cai, Y.Z., Surveswaran, S., Hyde, K.D., Corke, H., and Sun, M. (2009).
Molecular phylogenetic identification of endophytic fungi isolated from three Artemisia
species. Fungal Diversity 36, 69-88.
Hughes, K.W., Petersen, R.H., and Lickey, E.B. (2009). Using heterozygosity to
estimate a percentage DNA sequence similarity for environmental species' delimitation
across basidiomycete fungi. New Phytologist 182, 795-798.
Ikediugwa, F.E.O., and Webster, J. (1970). Hyphal interference in a range of
coprophilous fungi. Transactions of the British Mycological Society 54, 205-210.
Jaglowski, J.R., and Stack Jr, B.C. (2006). Enhanced growth inhibition of squamous cell
carcinoma of the head and neck by combination therapy of fusaric acid and paclitaxel or
carboplatin. Cancer Letters 243, 58-63.
Jahromi, F.G. (2007). Effect of environmental factors on disease development caused
by the fungal pathogen Plectosporium alismatis on the floating-leaf stage of starfruit
(Damasonium minus), a weed of rice. Biocontrol Science and Technology 17, 871-877.
Jayanetti, D.R., Yue, Q., Bills, G.F., and Gloer, J.B. (2015). Hypocoprins A-C: New
sesquiterpenoids from the coprophilous fungus hypocopra rostrata. Journal of Natural
Products 78, 396-401.
Jorgensen, J., Turnidge, J., and Washington, J. (1999). Antibacterial susceptibility tests:
dilution and disk diffusion methods. In Manual of Clinical Microbiology, P. Murray, R.,
ed. (Washington, DC.: ASM Press), pp. 1526-1542.

References
158
Kale-Pradhan, P.B., Wilhelm, S.M., and Johnson, L.B. (2012). Clinical relevance of in
vitro resistance of echinocandins: A focus on candida parapsilosis. Current Fungal
Infection Reports 6, 107-112.
Keller, N.P., Turner, G., and Bennett, J.W. (2005). Fungal secondary metabolism -
From biochemistry to genomics. Nature Reviews Microbiology 3, 937-947.
Kikot, G.E., Hours, R.A., and Alconada, T.M. (2009). Contribution of cell wall
degrading enzymes to pathogenesis of Fusarium graminearum: A review. Journal of
Basic Microbiology 49, 231-241.
Klitgaard, A., Iversen, A., Andersen, M.R., Larsen, T.O., Frisvad, J.C., and Nielsen,
K.F. (2014). Aggressive dereplication using UHPLC-DAD-QTOF: Screening extracts
for up to 3000 fungal secondary metabolites. Analytical and Bioanalytical Chemistry
406, 1933-1943.
Kociolek (2009). Quorum-Sensing Inhibitors and Biofilms Anti-Infective Agents in
Medicinal Chemistry 8, 315.
Krivoruchko, A., and Nielsen, J. (2015). Production of natural products through
metabolic engineering of Saccharomyces cerevisiae. Current Opinion in Biotechnology
35, 7-15.
Krug, J.C. (2004). Moist chambers for the development of fungi. In Biodiversity of
Fungi: Inventory and Monitoring, G.M. Mueller, G.F. Bills, and M.S. Foster, eds. (San
Diego, CA: Elsevier), pp. 589-593.
Krug, J.C., Benny, G.L., and Keller, H.W. (2004). Coprophilous fungi. In Biodiversity
of Fungi: Inventory and Monitoring, G.M. Mueller, G.F. Bills, and M.S. Foster, eds.
(San Diego, CA: Elsevier), pp. 589-593.

References
159
Kruys, Å. (2015). New species of Preussia with 8-celled ascospores (Sporormiaceae,
Pleosporales, Ascomycota). Phytotaxa 234, 143-150.
Kruys, A., and Ericson, L. (2008). Species richness of corpophilous asomycetes in
relation to variable food intake by herbivores. Fungal Diversity 30, 73-81.
Kumar, L., Khormi, H.M., Leis, K., and Taylor, S. (2015). Ecological research in
Australia: Identifying links versus gaps between hotspots of ecological research and
biodiversity. Austral Ecology 40, 581-590.
Kumaran, R.S., Muthumary, J., Kim, E.K., and Hur, B.K. (2009). Production of taxol
from Phyllosticta dioscoreae, a leaf spot fungus isolated from Hibiscus rosa-sinensis.
Biotechnology and Bioprocess Engineering 14, 76-83.
Kurtböke, D.I. (2012). Biodiscovery from rare actinomycetes: An eco-taxonomical
perspective. Applied Microbiology and Biotechnology 93, 1843-1852.
Kuznetsova, T.A., Smetanina, O.F., Afiyatullov, S.S., Pivkin, M.V., Denisenko, V.A.,
and Elyakov, G.B. (2001). The identification of fusidic acid, a steroidal antibiotic from
marine isolate of the fungus stilbella aciculosa. Biochemical Systematics and Ecology
29, 873-874.
Kwon, H.R., Son, S.W., Han, H.R., Choi, G.J., Jang, K.S., Choi, Y.H., Lee, S., Sung,
N.D., and Kim, J.C. (2007). Nematicidal activity of bikaverin and fusaric acid isolated
from Fusarium oxysporum against pine wood nematode, Bursaphelenchus xylophilus.
Plant Pathology Journal 23, 318-321.
Lam, K.S. (2007). New aspects of natural products in drug discovery. Trends in
Microbiology 15, 279-289.
Lamberth, C., Jeanmart, S., Luksch, T., and Plant, A. (2013). Current challenges and
trends in the discovery of agrochemicals. Science 341, 742-746.

References
160
Larsen, T.O., and Hansen, M.A.E. (2008). Dereplication and discovery of natural
products by UV spectroscopy In Bioactive Natural Products: Detection, Isolation, and
Structural Determination, S.M. Colegate, and R.J. Molyneux, eds. (Boca Raton, USA:
CRC Press).
Lee, K.K., Gloer, J.B., Scott, J.A., and Malloch, D. (1995). Petriellin A: A novel
antifungal depsipeptide from the coprophilous fungus Petriella sordida. The Journal of
Organic Chemistry 60, 5384-5385.
Lehr, N.A., Meffert, A., Antelo, L., Sterner, O., Anke, H., and Weber, R.W.S. (2006).
Antiamoebins, myrocin B and the basis of antifungal antibiosis in the coprophilous
fungus Stilbella erythrocephala (syn. S. fimetaria). FEMS Microbiology Ecology 55,
105-112.
Lewis, J.S., and Graybill, J.R. (2008). Fungicidal versus fungistatic: What's in a word?
Expert Opinion on Pharmacotherapy 9, 927-935.
Li, H., Lee, J.H., and Ha, J.M. (2008). Effective purification of ginsenosides from
cultured wild ginseng roots, red ginseng, and white ginseng with macroporous resins.
Journal of Microbiology and Biotechnology 18, 1789-1791.
Li, J.W.H., and Vederas, J.C. (2009). Drug discovery and natural products: End of an
era or an endless frontier? Science 325, 161-165.
Lin, X., Huang, Y., Fang, M., Wang, J., Zheng, Z., and Su, W. (2005). Cytotoxic and
antimicrobial metabolites from marine lignicolous fungi, Diaporthe sp. FEMS
Microbiology Letters 251, 53-58.
Liu, K., Ding, X., Deng, B., and Chen, W. (2009). Isolation and characterization of
endophytic taxol-producing fungi from Taxus chinensis. Journal of Industrial
Microbiology and Biotechnology 36, 1171-1177.

References
161
Liu, W., Li, L.P., Zhang, J.D., Li, Q., Shen, H., Chen, S.M., He, L.J., Yan, L., Xu, G.T.,
An, M.M., et al. (2014). Synergistic antifungal effect of glabridin and fluconazole.
PLoS ONE 9.
Liu, X.L., Huang, K.H., Zhou, J.Z., Meng, L., Wang, Y., and Zhang, L.X. (2012).
Identification and antibacterial characteristics of an endophytic fungus Fusarium
oxysporum from Lilium lancifolium. Letters in Applied Microbiology 55, 399-406.
Livermore, D.M. (2009). Has the era of untreatable infections arrived? Journal of
Antimicrobial Chemotherapy 64, 29-36.
Madigan, M., T., Martinko, J., M., and Parker, J. (2003). Brock Biology of
Microorganisms, 10th edn (Upper Saddle River, NJ: Pearson Education).
Martín-Rodríguez, A.J., Reyes, F., Martín, J., Pérez-Yépez, J., León-Barrios, M.,
Couttolenc, A., Espinoza, C., Trigos, A., Martín, V.S., Norte, M., et al. (2014).
Inhibition of bacterial quorum sensing by extracts from aquatic fungi: First report from
marine endophytes. Marine Drugs 12, 5503-5526.
Martin, S.M., Kau, D.A., and Wrigley, S.K. (2006). Scale-up of natural product
isolation. In Natural Products Isolation, S. Sarker, D., Z. Latif, and A. Gray, I., eds.
(New Jersey, USA: Humana Press).
May, H.D., Wu, Q., and Blake, C.K. (2000). Effects of the Fusarium spp. mycotoxins
fusaric acid and deoxynivalenol on the growth of Ruminococcus albus and
Methanobrevibacter ruminantium. Canadian Journal of Microbiology 46, 692-699.
McClean, K.H., Winson, M.K., Fish, L., Taylor, A., Chhabra, S.R., Camara, M.,
Daykin, M., Lamb, J.H., Swift, S., Bycroft, B.W., et al. (1997). Quorum sensing and
Chromobacterium violaceum: Exploitation of violacein production and inhibition for
the detection of N-acylhomoserine lactones. Microbiology 143, 3703-3711.

References
162
McMullan-Fisher, S.J.M., May, T.W., Robinson, R.M., Bell, T.L., Lebel, T.,
Catcheside, P., and York, A. (2011). Fungi and fire in Australian ecosystems: A review
of current knowledge, management implications and future directions. Australian
Journal of Botany 59, 70-90.
Meletiadis, J., Meis, J.F.G.M., Mouton, J.W., Donnelly, J.P., and Verweij, P.E. (2000).
Comparison of NCCLS and 3-(4,5-dimethyl-2-thiazyl)-2,5-diphenyl-2H-tetrazolium
bromide (MTT) methods of in vitro susceptibility testing of filamentous fungi and
development of a new simplified method. Journal of Clinical Microbiology 38, 2949-
2954.
Melo, R.F.R., Bezerra, J.L., and Cavalcanti, M.A.Q. (2012). Diversity of coprophilous
ascomycetes from captive wild animals in Dois Irmãos State Park, Brazil. Nova
Hedwigia 94, 153-162.
Milshteyn, A., Schneider, J.S., and Brady, S.F. (2014). Mining the metabiome:
Identifying novel natural products from microbial communities. Chemistry and Biology
21, 1211-1223.
Mishra, B.B., and Tiwari, V.K. (2011). Natural products: An evolving role in future
drug discovery. European Journal of Medicinal Chemistry 46, 4769-4807.
Misra, J.K., Pandey, S., Gupta, A.K., and Deshmukh, S.K. (2014). Coprophilous fungi:
A review and selected bibliography. In Fungi from Different Substrates, J.K. Misra, J.P.
Tewari, S.K. Deshmukh, and C. Vagvolgyi, eds. (Boca Raton, USA: CRC Press), pp.
170-200.
Morris, H.M., Smith, M.E., Rizzo, D.M., Rejmanek, M., and Bledsoe, C.S. (2008).
Contrasting ectomycorrhizal fungal communites on the roots of co-occurring oaks
(Quercus spp.) in a California woodland. New Phytologist 178, 167-176.
Moyal, A.M. (2008). Koala : A Historical Biography (Collingwood, Vic: CSIRO
Publishing).

References
163
Mudur, S.V., Gloer, J.B., and Wicklow, D.T. (2006). Sporminarins A and B: Antifungal
metabolites from a fungicolous isolate of Sporormiella minimoides. Journal of
Antibiotics 59, 500-506.
Mukhopadhyay, T., Bhat, R.G., Roy, K., Vijayakumar, E.K.S., and Ganguli, B.N.
(1998). Aranochlor A and aranochlor B, two new metabolites from Pseudoarachniotus
rosens: Production, isolation, structure elucidation and biological properties. Journal of
Antibiotics 51, 439-441.
Murugan, K., Saravanababu, S., and Arunachalam, M. (2007). Screening of tannin acyl
hydrolase (E.C.3.1.1.20) producing tannery effluent fungal isolates using simple agar
plate and SmF process. Bioresource Technology 98, 946-949.
Nath, A., Raghunatha, P., and Joshi, S.R. (2012). Diversity and biological activities of
endophytic fungi of Emblica officinalis, an ethnomedicinal plant of India. Mycobiology
40, 8-13.
Netzker, T., Fischer, J., Weber, J., Mattern, D.J., König, C.C., Valiante, V., Schroeckh,
V., and Brakhage, A.A. (2015). Microbial communication leading to the activation of
silent fungal secondary metabolite gene clusters. Frontiers in Microbiology 6.
Newman, D.J., and Cragg, G.M. (2012). Natural products as sources of new drugs over
the 30 years from 1981 to 2010. Journal of Natural Products 75, 311-335.
Ng, R. (2005). Drugs: From discovery to approval (New Jersey: John Wiley & Sons).
Niehaus, E.M., Von Bargen, K.W., Espino, J.J., Pfannmüller, A., Humpf, H.U., and
Tudzynski, B. (2014). Characterization of the fusaric acid gene cluster in Fusarium
fujikuroi. Applied Microbiology and Biotechnology 98, 1749-1762.
Nielsen, K.F., and Larsen, T.O. (2015). The importance of mass spectrometric
dereplication in fungal secondary metabolite analysis. Frontiers in Microbiology 6.

References
164
O'Brian, H.E., Parrent, J.L., Jackson, J.A., Moncalvo, J.-M., and Vilgalys, R. (2005).
Fungal community analysis by large scale sequencing of environmental samples.
Applied and Environmental Microbiology 71, 5544-5550.
O'Sullivan, J., Phillipson, D.W., Straub, H., and Ermann, P.H. (1993). 4-Methoxy-5-
methylpyran-3-ol natural products scopularin and lanomycin and derivatives thereof. In
US Patent (USA).
Odds, F.C. (2001). Sordarin antifungal agents. Expert Opinion on Therapeutic Patents
11, 283-294.
Olufolaji, D., B. (2010). Prospects of large-scale use of natural products as alternatives
to synthetic pesticides in developing countries. In Natural Products in Plant Pest
Management, N. Dubey, K., ed. (United Kingdom: CABI Publishing).
Osawa, R., Bird, P.S., Harbrow, D.J., Ogimoto, K., and Seymour, G.J. (1993).
Microbiological studies of the intestinal microflora of the koala, Phascolarctos cinereus.
I.Colonisation of the caecal wall by tannin-protein-complex-degrading enterobacteria.
Australian Journal of Zoology 41, 599-609.
Paech, K. (1950). Biochemie and physiologie der sekundaren pflanzenstoffe (Berlin:
Springer-Verlag).
Pan, J.H., Chen, Y., Huang, Y.H., Tao, Y.W., Wang, J., Li, Y., Peng, Y., Dong, T., Lai,
X.M., and Lin, Y.C. (2011). Antimycobacterial activity of fusaric acid from a mangrove
endophyte and its metal complexes. Archives of Pharmacal Research 34, 1177-1181.
Pandey, S. (2009). Studies on fungi growing on the dung of some herbivores; PhD
thesis (Lucknow: University of Lucknow), pp. 207.

References
165
Perrone, G., Haidukowski, M., Stea, G., Epifani, F., Bandyopadhyay, R., Leslie, J.F.,
and Logrieco, A. (2014). Population structure and Aflatoxin production by Aspergillus
Sect. Flavi from maize in Nigeria and Ghana. Food Microbiology 41, 52-59.
Peterson, R.A., Bradner, J.R., Roberts, T.H., and Nevalainen, K.M.H. (2009). Fungi
from koala (Phascolarctos cinereus) faeces exhibit a broad range of enzyme activities
against recalcitrant substrates. Letters in Applied Microbiology 48, 218-225.
Pfaller, M.A., and Diekema, D.J. (2007). Epidemiology of invasive candidiasis: A
persistent public health problem. Clinical Microbiology Reviews 20, 133-163.
Pfaller, M.A., Diekema, D.J., Castanheira, M., and Jones, R.N. (2011). Definitions and
epidemiology of Candida species not susceptible to echinocandins. Current Fungal
Infection Reports 5, 120-127.
Phaopongthai, J., Wiyakrutta, S., Meksuriyen, D., Sriubolmas, N., and Suwanborirux,
K. (2013). Azole-synergistic anti-candidal activity of altenusin, a biphenyl metabolite of
the endophytic fungus Alternaria alternata isolated from Terminalia chebula Retz.
Journal of Microbiology 51, 821-828.
Phillips, R.L., Palombo, E.A., Panozzo, J.F., and Bhave, M. (2011). Puroindolines, Pin
alleles, hordoindolines and grain softness proteins are sources of bactericidal and
fungicidal peptides. Journal of Cereal Science 53, 112-117.
Phongpaichit, S., Rungjindamai, N., Rukachaisirikul, V., and Sakayaroj, J. (2006).
Antimicrobial activity in cultures of endophytic fungi isolated from Garcinia species.
FEMS Immunology and Medical Microbiology 48, 367-372.
Prakash, V. (2015). Endophytic fungi as resource of bioactive compounds. International
Journal of Pharma and Bio Sciences 6, B887-B898.

References
166
Projan, S.J., and Youngman, P.J. (2002). Antimicrobials: New solutions badly needed.
Current Opinion in Microbiology 5, 463-465.
Pu, X., Qu, X., Chen, F., Bao, J., Zhang, G., and Luo, Y. (2013). Camptothecin-
producing endophytic fungus Trichoderma atroviride LY357: Isolation, identification,
and fermentation conditions optimization for camptothecin production. Applied
Microbiology and Biotechnology 97, 9365-9375.
Qadri, M., Rajput, R., Abdin, M.Z., Vishwakarma, R.A., and Riyaz-Ul-Hassan, S.
(2014). Diversity, molecular phylogeny, and bioactive potential of fungal endophytes
associated with the Himalayan blue pine (Pinus wallichiana). Microbial Ecology 67,
877-887.
Qaiyumi, S. (2007). Macro- and microdilution methods of antimicrobial susceptibility
testing. In Antimicrobial Susceptibility Testing Protocols, R. Schwalbe, L. Steele-
Moore, and A. Goodwin, eds. (Boca Raton, USA: CRC Press), pp. 75-79.
Quiroga, E.N., Sampietro, D.A., Sgariglia, M.A., Soberón, J.R., and Vattuone, M.A.
(2009). Antimycotic activity of 5′-prenylisoflavanones of the plant Geoffroea
decorticans, against Aspergillus species. International Journal of Food Microbiology
132, 42-46.
Rahman, A., Choudhary, M., and Thompson, W. (2001). Bioassay techniques for drug
development (EBL EBook Library, viewed 1 February 2012: Informa Healthcare).
Raju, N.B. (2002). Meiosis and ascospore development in nonlinear asci of Neurospora
pannonica. Mycologia 94, 99-104.
Rateb, M.E., and Ebel, R. (2011). Secondary metabolites of fungi from marine habitats.
Natural Product Reports 28, 290-344.

References
167
Reade, E. (1994). Techniques manual - Department of microbiology (Melbourne:
University of Melbourne Publishing).
Reina, R., Liers, C., Ocampo, J.A., Garcia-Romera, I., and Aranda, E. (2013). Solid
state fermentation of olive mill residues by wood- and dung-dwelling Agaricomycetes:
Effects on peroxidase production, biomass development and phenol phytotoxicity.
Chemosphere 93, 1406-1412.
Richardson, M.J. (2001a). Coprophilous fungi from Brazil. Brazilian Archives of
Biology and Technology 44, 283-289.
Richardson, M.J. (2001c). Diversity and occurrence of coprophilous fungi. Mycological
Research 105, 387-402.
Richardson, M.J. (2002). The coprophilous succession. Fungal Diversity 10, 101-111.
Ridderbusch, D.C., Weber, R.W.S., Anke, T., and Sterner, O. (2004). Tulasnein and
podospirone from the coprophilous xylariaceous fungus Podosordaria tulasnei.
Zeitschrift fur Naturforschung - Section C Journal of Biosciences 59, 379-383.
Rodríguez-Algaba, J., Sørensen, J.C., Sørensen, H., Khoa, N.Đ., Collinge, D.B., and
Jørgensen, H.J.L. (2015). Activity-guided separation of Chromolaena odorata leaf
extract reveals fractions with rice disease-reducing properties. European Journal of
Plant Pathology 143, 331-341.
Rofiat, A.S., Fanelli, F., Atanda, O., Sulyok, M., Cozzi, G., Bavaro, S., Krska, R.,
Logrieco, A.F., and Ezekiel, C.N. (2015). Fungal and bacterial metabolites associated
with natural contamination of locally processed rice (Oryza sativa L.) in Nigeria. Food
Additives and Contaminants - Part A Chemistry, Analysis, Control, Exposure and Risk
Assessment 32, 950-959.

References
168
Roy, K., Vijayakumar, E.K.S., Mukhopadhyay, T., Chatterjee, S., Bhat, R.G.,
Blumbach, J., and Ganguli, B.N. (1992). Aranorosinol A and aranorosinol B, two new
metabolites from Pseudoarachniotus roseus: Production, isolation, structure elucidation
and biological properties. Journal of Antibiotics 45, 1592-1598.
Ruiz, J.A., Bernar, E.M., and Jung, K. (2015). Production of siderophores increases
resistance to fusaric acid in Pseudomonas protegens Pf-5. PLoS ONE 10.
Ryberg, M., Nilsson, R.H., Kristiansson, E., Topel, M., Jacobsson, S., and Larsson, E.
(2008). Mining metadata from unidentified ITS sequences in GenBank: A case study in
Inocybe (Basidiomycota). BMC Evolutionary Biology 8, 50-64.
Saitou, N., and Nei, M. (1987). The neighbor-joining method: A new method for
reconstructing phylogenetic trees. Molecular biology and evolution 4, 406-425.
Sandle, T., Vijayakumar, R., Saleh Al Aboody, M., and Saravanakumar, S. (2014). In
vitro fungicidal activity of biocides against pharmaceutical environmental fungal
isolates. Journal of Applied Microbiology 117, 1267-1273.
Saremi, H., and Okhovvat, S.M. (2006). Mycotoxin producing Fusarium species
associated with plant disease on potato, wheat, corn and animal diseases in northwest
Iran. Communications in agricultural and applied biological sciences 71, 1175-1185.
Sarrocco, S., Diquattro, S., Avolio, F., Cimmino, A., Puntoni, G., Doveri, F., Evidente,
A., and Vannacci, G. (2015a). Bioactive metabolites from new or rare fimicolous fungi
with antifungal activity against plant pathogenic fungi. European Journal of Plant
Pathology 142, 61-71.
Sarrocco, S., Diquattro, S., Baroncelli, R., Cimmino, A., Evidente, A., Vannacci, G.,
and Doveri, F. (2015c). A polyphasic contribution to the knowledge of Auxarthron
(Onygenaceae). Mycological Progress 14.

References
169
Schmidt-Heydt, M., Bode, H., Raupp, F., and Geisen, R. (2010). Influence of light on
ochratoxin biosynthesis by Penicillium. Mycotoxin Research 26, 1-8.
Scoma, A., Pintucci, C., Bertin, L., Carlozzi, P., and Fava, F. (2012). Increasing the
large scale feasibility of a solid phase extraction procedure for the recovery of natural
antioxidants from olive mill wastewaters. Chemical Engineering Journal 198-199, 103-
109.
Segeth, M.P., Bonnefoy, A., Brönstrup, M., Knauf, M., Schummer, D., Toti, L.,
Vértesy, L., Wetzel-Raynal, M.C., Wink, J., and Seibert, G. (2003). Coniosetin, a novel
tetramic acid antibiotic from Coniochaeta ellipsoidea DSM 13856. Journal of
Antibiotics 56, 114-122.
Seida, A.A., El Tanbouly, N.D., Islam, W.T., Eid, H.H., El Maraghy, S.A., and El
Senousy, A.S. (2015). Bioassay-guided fractionation of a hepatoprotective and
antioxidant extract of pea by-product. Natural Product Research 29, 1578-1583.
Shimshoni, J.A., Cuneah, O., Sulyok, M., Krska, R., Galon, N., Sharir, B., and
Shlosberg, A. (2013). Mycotoxins in corn and wheat silage in Israel. Food Additives
and Contaminants - Part A Chemistry, Analysis, Control, Exposure and Risk
Assessment 30, 1614-1625.
Sica, V.P., Raja, H.A., El-Elimat, T., Kertesz, V., Van Berkel, G.J., Pearce, C.J., and
Oberlies, N.H. (2015). Dereplicating and spatial mapping of secondary metabolites
from fungal cultures in situ. Journal of Natural Products 78, 1926-1936.
Smith, M.E., Douhan, G.W., and Rizzo, D.M. (2007). Ectomycorrhizal community
structure in a xeric Quercus woodland based on rDNA sequence analysis of sporocarps
and pooled roots. New Phytologist 174, 847-863.
Sneader, W. (2005). Drug discovery: A history (Chichester, England: John Wiley &
Sons Ltd.).

References
170
Soman, A.G., Gloer, J.B., Koster, B., and Malloch, D. (1999). Sporovexins A-C and a
new preussomerin analog: Antibacterial and antifungal metabolites from the
coprophilous fungus Sporormiella vexans. Journal of Natural Products 62, 659-661.
Son, S., W., Kim, H., Y., Choi, G., J., Lim, H., K., Jang, K., S., Lee, S., O., Lee, S.,
Sung, N., D., and Kim, J.-C. (2008). Bikaverin and fusaric acid from Fusarium
oxysporum show antioomycete activity against Phytophthora infestans. Journal of
Applied Microbiology 104, 692-698.
Spitzer, M., Griffiths, E., Blakely, K.M., Wildenhain, J., Ejim, L., Rossi, L., De Pascale,
G., Curak, J., Brown, E., Tyers, M., et al. (2011). Cross-species discovery of syncretic
drug combinations that potentiate the antifungal fluconazole. Molecular Systems
Biology 7.
Spraker, J., and Keller, N. (2014). Waking Sleeping Pathways in Filamentous Fungi. In
Natural Products: Discourse, Diversity, and Design, pp. 277-292.
Stack Jr, B.C., Hansen, J.P., Ruda, J.M., Jaglowski, J., Shvidler, J., and Hollenbeak,
C.S. (2004). Fusaric acid: A novel agent and mechanism to treat HNSCC.
Otolaryngology - Head and Neck Surgery 131, 54-60.
Stierle, A., Strobel, G., and Stierle, D. (1993). Taxol and taxane production by
Taxomyces andreanae, an endophytic fungus of pacific yew. Science 260, 214-216.
Stipanovic, R.D., Wheeler, M.H., Puckhaber, L.S., Liu, J., Bell, A.A., and Williams,
H.J. (2011). Nuclear magnetic resonance (NMR) studies on the biosynthesis of fusaric
acid from fusarium oxysporum f. sp. vasinfectum. Journal of Agricultural and Food
Chemistry 59, 5351-5356.
Strobel, G., and Daisy, B. (2003). Bioprospecting for Microbial Endophytes and Their
Natural Products. Microbiology and Molecular Biology Reviews 67, 491-502.

References
171
Strobel, G., Yang, X., Sears, J., Kramer, R., Sidhu, R.S., and Hess, W.M. (1996). Taxol
from Pestalotiopsis microspora, an endophytic fungus of Taxus wallachiana.
Microbiology 142, 435-440.
Strobel, G.A. (2003). Endophytes as sources of bioactive products. Microbes and
Infection 5, 535-544.
Strobel, G.A. (2006). Harnessing endophytes for industrial microbiology. Current
Opinion in Microbiology 9, 240-244.
Swathi, J., Sowjanya, K.M., Narendra, K., and Krishna Satya, A. (2013). Bioactivity
assay of an isolated marine Fusarium sps. International Journal of Bio-Science and Bio-
Technology 5, 179-186.
Takahashi, J.A., Teles, A.P.C., De Almeida Pinto Bracarense, A., and Gomes, D.C.
(2013). Classical and epigenetic approaches to metabolite diversification in filamentous
fungi. Phytochemistry Reviews 12, 773-789.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. (2011).
MEGA5: Molecular evolutionary genetics analysis using maximum likelihood,
evolutionary distance, and maximum parsimony methods. Molecular Biology and
Evolution 28, 2731-2739.
Tesoriero, L. (2014). Emerging plant diseases for protected cropping. In Soilless
Australia (N.S.W, Australia: Protected Cropping Australia Ltd.), pp. 9.
Thammajaruk, N., Sriubolmas, N., Israngkul, D., Meevootisom, V., and Wiyakrutta, S.
(2010). Optimization of culture conditions for mycoepoxydiene production by
Phomopsis sp. Hant25. Journal of Industrial Microbiology and Biotechnology, 1-7.

References
172
Tolulope, R.A., Adeyemi, A.I., Erute, M.A., and Abiodun, T.S. (2015). Isolation and
screening of endophytic fungi from three plants used in traditional medicine in Nigeria
for antimicrobial activity. International Journal of Green Pharmacy 9, 58-62.
Tong, W.Y., Darah, I., and Latiffah, Z. (2011). Antimicrobial activities of endophytic
fungal isolates from medicinal herb Orthosiphon stamineus Benth. Journal of Medicinal
Plant Research 5, 831-836.
Tyndale-Biscoe, H. (2005). Life of Marsupials (Collingwood, Victoria: CSIRO
Publishing).
Utsumi, R., Yagi, T., Katayama, S., Katsuragi, K., Tachibana, K., Toyoda, H., Ouchi,
S., Obata, K., Shibano, Y., and Noda, M. (1991). Molecular cloning and
characterization of the fusaric acid-resistance gene from Pseudomonas cepacia.
Agricultural and Biological Chemistry 55, 1913-1918.
Van Dugteren, A. (1999). Shedding light on Lepidoptera. In Ecos, pp. 33-35.
Van Rij, E.T., Girard, G., Lugtenberg, B.J.J., and Bloemberg, G.V. (2005). Influence of
fusaric acid on phenazine-1-carboxamide synthesis and gene expression of
Pseudomonas chlororaphis strain PCL1391. Microbiology 151, 2805-2814.
Verma, P. (2007). Methods for determining bactericidal activity and antimicrobial
interactions: syngery testing, time-kill curves, and population analysis. In Antimicrobial
Susceptibility Testing Protocols, R. Schwalbe, L. Steele-Moore, and A. Goodwin, eds.
(Boca Raton, USA: CRC Press), pp. 275-298.
Vicente, M.F., Basilio, A., Cabello, A., and Peláez, F. (2003). Microbial natural
products as a source of antifungals. Clinical Microbiology and Infection 9, 15-32.

References
173
Vurro, M., Bonciani, B., and Vannacci, G. (2010). Emerging infectious diseases of crop
plants in developing countries: Impact on agriculture and socio-economic consequences.
Food Security 2, 113-132.
Waksman, S.A. (1954). My Life With Microbes (New York: Simon and Schuster).
Walker, J.F., Miller, O.K., and Horton, J.L. (2008). Seasonal dynamics of
ectoycorrhizal fungus assemblages on oak seedlings in the southeastern Appalachian
Mountains. Mycorrhizia 19, 123-132.
Wang, H.j., Gloer, J.B., Scott, J.A., and Malloch, D. (1995a). Coniochaetones A and B:
New antifungal benzopyranones from the coprophilous fungus Coniochaeta saccardoi.
Tetrahedron Letters 36, 5847-5850.
Wang, H.J., Gloer, K.B., Gloer, J.B., Scott, J.A., and Malloch, D. (1997). Anserinones
A and B: New antifungal and antibacterial benzoquinones from the coprophilous fungus
Podospora anserina. Journal of Natural Products 60, 629-631.
Wang, J., Zhang, H., Ying, L., Wang, C., Jiang, N., Zhou, Y., Wang, H., and Bai, H.
(2009). Five new epothilone metabolites from Sorangium cellulosum strain So0157-2.
The Journal of Antibiotics 62, 483-487.
Wang, Q.X., Bao, L., Yang, X.L., Guo, H., Ren, B., Guo, L.D., Song, F.H., Wang,
W.Z., Liu, H.W., and Zhang, L.X. (2013). Tricycloalternarenes F-H: Three new mixed
terpenoids produced by an endolichenic fungus Ulocladium sp. using OSMAC method.
Fitoterapia 85, 8-13.
Wang, Y., Gloer, J.B., Scott, J.A., and Malloch, D. (1993). Appenolides A-C: Three
new antifungal furanones from the coprophilous fungus Podospora appendiculata.
Journal of Natural Products (Lloydia) 56, 341-344.

References
174
Wang, Y., Gloer, J.B., Scott, J.A., and Malloch, D. (1995b). Terezines A-D: New amino
acid-derived bioactive metabolites from the coprophilous fungus Sporormiella
teretispora. Journal of Natural Products 58, 93-99.
Wanger, A. (2007). Disk diffusion test and gradient methodologies. In Antimicrobial
Susceptibility Testing Protocols, R. Schwalbe, L. Steele-Moore, and A. Goodwin, eds.
(Boca Raton, USA: CRC Press), pp. 53 - 73.
Warcup, J.H. (1950). The soil-plate method for isolation of fungi from soil. Nature 166,
117 - 118.
Waterhouse, A.M., Proctor, J.B., Martin, D.M.A., Clamp, M., and Barton, G.J. (2009).
Jalview Version 2 - a multiple sequence alignment editor and analysis workbench.
Watling, R., and Richardson, M.J. (2010). Coprophilous fungi of the Falkland Islands.
Edinburgh Journal of Botany 67, 399-423.
Watve, M.G., Tickoo, R., Jog, M.M., and Behole, B.D. (2001). How many antibiotics
are produced by the genus Streptomyces? Archives of Microbiology, 386-390.
Wawrzyn, G.T., Quin, M.B., Choudhary, S., Lopez-Gallego, F., and Schmidt-Dannert,
C. (2012). Draft genome of Omphalotus olearius provides a predictive framework for
sesquiterpenoid natural product biosynthesis in basidiomycota. Chemistry and Biology
19, 772-783.
Weber, H.A., Baenziger, N.C., and Gloer, J.B. (1990). Structure of preussomerin A: An
unusual new antifungal metabolite from the coprophilous fungus Preussia isomera.
Journal of the American Chemical Society 112, 6718-6719.
Weber, H.A., and Gloer, J.B. (1988). Interference competition among natural fungal
competitors: An antifungal metabolite from the coprophilous fungus Preussia
fleischhakii. Journal of Natural Products 51, 879-883.

References
175
Weber, H.A., and Gloer, J.B. (1991). The preussomerins: Novel antifungal metabolites
from the coprophilous fungus Preussia isomera Cain. Journal of Organic Chemistry 56,
4355-4360.
Weber, H.A., Swenson, D.C., Gloer, J.B., and Malloch, D. (1992). Similins A and B:
New antifungal metabolites from the coprophilous fungus Sporormiella similis.
Tetrahedron Letters 33, 1157-1160.
Wei, H., Lin, Z., Li, D., Gu, Q., and Zhu, T. (2010). OSMAC (one strain many
compounds) approach in the research of microbial metabolites--a review. Wei sheng wu
xue bao = Acta microbiologica Sinica 50, 701-709.
Weller, M.G. (2012). A unifying review of bioassay-guided fractionation, effect-
directed analysis and related techniques. Sensors (Switzerland) 12, 9181-9209.
White, T.J., Bruns, T.D., Lee, S.J., and Taylor, J. (1990). Amplification and direct
sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, M.
Innis, D. Gelfand, J. Sninsky, and T. White, eds. (NewYork, N. Y.: Academic Press
Inc.), pp. 315-322.
WHO (2014). Antimicrobial Resistance. Global Report on Surveillance : Global Report
on Surveillance (World Health Organization).
WHO (2015). WHO Model list of essential medicines (World Health Organisation).
Whyte, A.C., Gloer, J.B., Scott, J.A., and Malloch, D. (1996). Cercophorins A-C: Novel
antifungal and cytotoxic metabolites from the coprophilous fungus Cercophora
areolata. Journal of Natural Products 59, 765-769.
Whyte, A.C., Gloer, K.B., Gloer, J.B., Koster, B., and Malloch, D. (1997). New
antifungal metabolites from the coprophilous fungus Cercophora sordarioides.
Canadian Journal of Chemistry 75, 768-772.

References
176
Wicklow, D.T. (1981). Interference competition. In The Fungal Community: Its
Organization and the Role in the Ecosystem, D.T. Wicklow, and G.C. Carroll, eds.
(New York: Marcel Dekker), pp. 265-274.
Wicklow, D.T., Detroy, R.W., and Adams, S. (1980). Differential modification of the
lignin and cellulose components in wheat straw by fungal colonists of ruminant dung:
ecologicial implications. Mycologia 72, 1065-1076.
Wong, F.T., and Khosla, C. (2012). Combinatorial biosynthesis of polyketides-a
perspective. Current Opinion in Chemical Biology 16, 117-123.
Wu, M.C., Law, B., Wilkinson, B., and Micklefield, J. (2012). Bioengineering natural
product biosynthetic pathways for therapeutic applications. Current Opinion in
Biotechnology 23, 931-940.
Xing, Y.M., Chen, J., Cui, J.L., Chen, X.M., and Guo, S.X. (2011). Antimicrobial
activity and biodiversity of endophytic fungi in Dendrobium devonianum and
Dendrobium thyrsiflorum from Vietman. Current Microbiology 62, 1218-1224.
Xu, L.J., Liu, Y.S., Zhou, L.G., and Wu, J.Y. (2010). Optimization of a liquid medium
for beauvericin production in fusarium redolens dzf2 mycelial culture. Biotechnology
and Bioprocess Engineering 15, 460-466.
Yabuta, T., Kambe, K., and Hayashi, T. (1934). Fusaric acid, a new product of the
"bakanae" fungus. Journal of the Agricultural Chemical Society of Japan 10, 1059-
1068.
Yuhico, L.S.O., and Blair, D.C. (2011). Multi-Drug-Resistant Pseudomonas aeruginosa:
A Rapidly Growing Treatment Dilemma. Clinical Microbiology Newsletter 33, 159-
160.

References
177
Zaferanloo, B., Mahon, P.J., and Palombo, E.A. (2012). Endophytes from medicinal
plants as novel sources of bioactive compounds. In Medicinal Plants: Diversity and
Drugs, M. Rai, L. Rastrelli, M. Marinof, J.L. Martinez, and G.A. Cordell, eds. (USA:
Science Publisher), pp. 355-411.
Zenk, M., H. (1967). Biochemie und physiologie der sekundaren pflanzenstoffe.
Berichte der Deutschen Botanischen Gesellschaft 80, 573-591.
Zhai, B., Zhou, H., Yang, L., Zhang, J., Jung, K., Giam, C.Z., Xiang, X., and Lin, X.
(2010). Polymyxin B, in combination with fluconazole, exerts a potent fungicidal effect.
Journal of Antimicrobial Chemotherapy 65, 931-938.
Zhang, H., Bai, X., and Wu, B. (2012a). Evaluation of antimicrobial activities of
extracts of endophytic fungi from Artemisia annua. Bangladesh Journal of
Pharmacology 7, 120-123.
Zhang, H., Ruan, C., and Bai, X. (2015). Isolation and antimicrobial effects of
endophytic fungi from Edgeworthia chrysantha. Bangladesh Journal of Pharmacology
10, 529-532.
Zhang, H., Xiong, Y., Zhao, H., Yi, Y., Zhang, C., Yu, C., and Xu, C. (2012b). An
antimicrobial compound from the endophytic fungus Phoma sp. isolated from the
medicinal plant Taraxacum mongolicum. Journal of the Taiwan Institute of Chemical
Engineers 44, 177-181.
Zhao, J., Li, Y., Liu, Q., and Gao, K. (2010). Antimicrobial activities of some thymol
derivatives from the roots of Inula hupehensis. Food Chemistry 120, 510-516.
Zhou, X., Zhu, H., Liu, L., Lin, J., and Tang, K. (2010). A review: Recent advances and
future prospects of taxol-producing endophytic fungi. Applied Microbiology and
Biotechnology 86, 1707-1717.

References
178
Zotchev, S.B., Sekurova, O.N., and Katz, L. (2012). Genome-based bioprospecting of
microbes for new therapeutics. Current Opinion in Biotechnology 23, 941-947.

Appendices
179
Appendices
Appendices
180
Appendix 1: 1H NMR spectrum (400 MHz) of F12–14 in D2O

Appendices
181
Appendix 2: 13C NMR spectrum (100 MHz) of F12–14 in D2O

Appendices
182
Appendix 3: COSY Spectrum of F12–14 in D2O

Appendices
183
Appendix 4: HSQC Spectrum of F12–14 in D2O

Appendices
184
Appendix 5: 1H NMR spectrum (400 MHz) of fusaric acid in D2O/NaOD

Appendices
185
Appendix 6: 13C NMR spectrum (100 MHz) of fusaric acid in D2O/NaOD

Appendices
186
Appendix 7: Publications arising from this thesis
1. Book chapter
Hayhoe, E. and Palombo, E. A. (2013). Screening for antibacterial, antifungal, and anti
quorum sensing activity. In Metabolomics Tools for Natural Product Discovery:
Methods and Protocols, U. Roessner and A. Dias, eds. (New Jersey, U.S.A: Humana
Press) pp 219–225.
2. Conference poster presentation
Hayhoe, E., Hart, N., Yang, Q., Bhave, M. and Palombo, E. A. (2012). Fusaric acid
from a coprophilous Fusarium oxysporum isolated from koala faeces displays
antimicrobial and anti-quorum sensing activities. Poster presented at the Australian
Society for Microbiology Annual Scientific Meeting and Exhibition, Brisbane,
Australia.
Copyright © 2022 FDOKUMEN




















