
Effect of growth temperature on the biosynthesis of cell wall proteins from Bifidobacterium globosum
11
Effect of growth temperature on the biosynthesis of cell wall proteins from Bifidobacterium globosum Paola Mattarelli a *, Bruno Biavati a , Michela Pesenti a , Franca Crociani b a Institute of Agricultural Microbiology, University of Bologna, Bologna, Italy b Institute of Normal and Pathologic Cytomorphology, CNR, Bologna, Italy (Submitted 15 June 1998; accepted 2 December 1998) Abstract — Seventy strains of Bifidobacterium globosum isolated from gastrointestinal tracts of different animals were studied. Strains were grown at temperatures ranging from 25 to 46.5°C in order to examine changes both in the expression of bifidobacterial outer proteins (BIFOPs) and in their hydrophobic properties. It was observed that the expression of BIFOPs found on the cell-surface changes according to growth temperature, with quantitative and/or qualitative variations. Generally speaking, it was observed that BIFOP expression at low-growth temperature was considerably attenuated, while at medium- and high-growth temperature it increased. Furthermore, at high-growth temperatures, the presence of a new common protein was detected in all the strains studied. Cells from B. globosum strains grown under different temperature conditions were studied in terms of their cellular hydrophobicity properties. At medium-growth temperature, the cell hydrophobicity was strictly correlated with BIFOP expression, while at low and high-growth temperatures, the presence of BIFOP only partially influenced the hydrophobic features. © Elsevier, Paris Bifidobacterium / bacterial colonization / temperature-inducible proteins / cell wall proteins / hydropho- bicity 1. Introduction The Bifidobacterium genus is one of the most frequently found in the microflora of the human and animal gastrointestinal tracts [28]; under normal physiological conditions, the presence of bifidobacteria exerts beneficial effects on its host, such as the inhibition of microbial patho- gens [9], the alleviation of lactose intolerance, the enhancement of the immune system and anticarcinogenic activity [2, 24]. Furthermore, because of their probiotic properties, bifidobac- teria have been added to industrial foods and pharmaceutical preparations for human and animal consumption. A better understanding of the physiological mechanisms that enable bifi- dobacterial colonization of the host gastrointes- tinal tract can be of utmost importance when employing particular strains as probiotic or dietary complements. Colonization processes play a crucial role in the settlement of a microbial community in the host [6]. The outcome of the competition of microorganisms in particular environments can be influenced or determined by their adhesive properties. The initial adhesion of bacteria de- pends partially on the nature of the surfaces and on the bacterial surface properties [1, 5]. The most important cell wall components involved in surface colonization, such as fimbriae, polysaccharides, teichoic and lipoteichoic acids, and proteins, also called adhesins, have been extensively investigated [7, 15, 31]. For a wide variety of bacteria, it has been shown that * Correspondence and reprints [email protected] Abbreviations: BAS, buffered ammonium sulphate; BIFOP, bifidobacterial outer protein; HIC, hydrophobic interaction chromatography; HTB, high temperature BIFOP; PBS, phosphate-buffered saline; SAT, salting-out aggregation test; SDS-PAGE, SDS protein acrylamide gel electrophore- sis; SPB, sodium phosphate buffer Res. Microbiol. 150 (1999), 117-127 © Elsevier, Paris
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Effect of growth temperature on the biosynthesis of cell wall proteins from Bifidobacterium globosum

Effect of growth temperature on the biosynthesis of cell wall proteinsfrom Bifidobacterium globosum
Paola Mattarellia*, Bruno Biavatia, Michela Pesentia, Franca Crocianib
aInstitute of Agricultural Microbiology, University of Bologna, Bologna, ItalybInstitute of Normal and Pathologic Cytomorphology, CNR, Bologna, Italy
(Submitted 15 June 1998; accepted 2 December 1998)
Abstract — Seventy strains of Bifidobacterium globosum isolated from gastrointestinal tracts of different animalswere studied. Strains were grown at temperatures ranging from 25 to 46.5°C in order to examine changes bothin the expression of bifidobacterial outer proteins (BIFOPs) and in their hydrophobic properties. It wasobserved that the expression of BIFOPs found on the cell-surface changes according to growth temperature,with quantitative and/or qualitative variations. Generally speaking, it was observed that BIFOP expressionat low-growth temperature was considerably attenuated, while at medium- and high-growth temperature itincreased. Furthermore, at high-growth temperatures, the presence of a new common protein was detected inall the strains studied. Cells from B. globosum strains grown under different temperature conditions werestudied in terms of their cellular hydrophobicity properties. At medium-growth temperature, the cellhydrophobicity was strictly correlated with BIFOP expression, while at low and high-growth temperatures,the presence of BIFOP only partially influenced the hydrophobic features. © Elsevier, Paris
Bifidobacterium / bacterial colonization / temperature-inducible proteins / cell wall proteins / hydropho-bicity
1. Introduction
The Bifidobacterium genus is one of the mostfrequently found in the microflora of the humanand animal gastrointestinal tracts [28]; undernormal physiological conditions, the presenceof bifidobacteria exerts beneficial effects on itshost, such as the inhibition of microbial patho-gens [9], the alleviation of lactose intolerance,the enhancement of the immune system andanticarcinogenic activity [2, 24]. Furthermore,because of their probiotic properties, bifidobac-
teria have been added to industrial foods andpharmaceutical preparations for human andanimal consumption. A better understanding ofthe physiological mechanisms that enable bifi-dobacterial colonization of the host gastrointes-tinal tract can be of utmost importance whenemploying particular strains as probiotic ordietary complements.
Colonization processes play a crucial role inthe settlement of a microbial community in thehost [6]. The outcome of the competition ofmicroorganisms in particular environments canbe influenced or determined by their adhesiveproperties. The initial adhesion of bacteria de-pends partially on the nature of the surfaces andon the bacterial surface properties [1, 5]. Themost important cell wall components involvedin surface colonization, such as fimbriae,polysaccharides, teichoic and lipoteichoic acids,and proteins, also called adhesins, have beenextensively investigated [7, 15, 31]. For a widevariety of bacteria, it has been shown that
* Correspondence and [email protected]
Abbreviations: BAS, buffered ammonium sulphate; BIFOP,bifidobacterial outer protein; HIC, hydrophobic interactionchromatography; HTB, high temperature BIFOP; PBS,phosphate-buffered saline; SAT, salting-out aggregationtest; SDS-PAGE, SDS protein acrylamide gel electrophore-sis; SPB, sodium phosphate buffer
Res. Microbiol. 150 (1999), 117−127© Elsevier, Paris

proteins which play an essential role in host-bacterium interactions are similarly regulated atthe transcriptional level, and that their expres-sion is induced by environmental stimuli likelyto be encountered within the host [12]. Cell wallproteins can be variably expressed in relation totemperature growth with qualitative or quanti-tative variations. Cell surface hydrophobicity,which can be correlated with the presence of cellwall proteins [22, 23], has been associated, asshown in many studies, with the colonization ofvarious surfaces [5, 34]. Greater hydrophobicityof cells corresponds to greater attractive forcesand higher levels of adhesion [8, 20, 26]. In thepresent work, we studied the expression ofbifidobacterial outer proteins (BIFOPs) in theresponse to different temperature conditions inBifidobacterium globosum strains; the influence ofBIFOP presence on cell surface hydrophobicitywas also investigated.
2. Materials and methods
2.1. Bacterial strains and culture conditions
The strains studied are listed in table I.Strains were cultured anaerobically in Gas Packjars (BBL) in trypticase-phytone-yeast extract(TPY) broth [4] at 37 °C.
2.2. Growth cultures at different temperatures
All the strains studied were grown at 25, 37and 45 °C in large-volume cultures (1 or 2 L) inconical flasks almost completely filled with TPYbroth, containing 2.5 mL /L of a 10% NaHCO3solution sterilized by filtration. The broth wasinoculated with 100 mL of 24 h TPY anaerobicculture grown at the same temperature as thelarge-volume culture preparation. Flasks wereincubated statically. Bacterial growth was deter-mined by the optical density at 600 nm. Sixteen
Table I. Strains studied, their origin and the presence of BIFOP (Coomassie staining) when grown at 25, 37 and 45°C.
Strains Isolated from BIFOP at 25°C BIFOP at 37°C BIFOP at 45°C
AG 57, AG 73, AG 82 lamb faeces – + +AG 29, AG 46, AG 49B lamb faeces – – +AG53, AG 69, AG 70 lamb faeces – – +
B 607, B608, B 614 infant faeces – – +
F 468, F 593/2, F 512 sewage – + +F 461/1, F 591 sewage – – +
P 7/2, P 42 chicken faeces – + +P 24, P 28, P 30 chicken faeces – – +P 92, P112, P115 chicken faeces – – +
RA 35 rabbit faeces – + +RA 120, RA 122, RA 125 rabbit faeces – – +RA 143, RA 167 rabbit faeces – – +
RU 809/1, bovine rumen – + +RU 240A, RU 457 bovine rumen – – +RU 809/8, RU 844 bovine rumen – – +
SU 841, pig faeces – + +SU 844, SU 847, SU 861 pig faeces – – +SU 881, SU 884, SU 886 pig faeces – – +
VT 97, VT 378 calf faeces – + +
T19, T 26, T 121, T 125, T 126 rat faeces – + +T 140A, T 154 rat faeces – + +T21, T 55, T 55B, T 58, T 75, T 78 rat faeces – – +T 86, T 95, T 107, T 113, T 117, T119B rat faeces – – +T 130, T 132, T 140B rat faeces – – +T 142, T 143, T 175B rat faeces – – +
+, BIFOP positive; –, BIFOP negative
118 Mattarelli et al.
strains from different habitats (table II) wereselected to be grown also at 30, 40, 41.5, 43, and46.5 °C.
2.3. Cell wall protein preparations, SDS-PAGE,immunoblotting
Cell wall preparations were obtained fromsonicated bacteria by differential centrifugationaccording to Mattarelli et al. [16]. Cell wallproteins were examined by SDS-PAGE using
the method of Laemmli [11] with an 8–20%linear gradient; 60 mg of total protein per lanewere used. Following electrophoresis, proteinswere stained with Coomassie or were trans-ferred electrophoretically to cellulose nitrate/acetate filters (Millipore GS-0.22 µ, Millipore).Proteins on the blots were detected using the‘Biotin-Blot Protein Detection kit’ (Bio-Rad,Richmond, CA) to show total proteins or usingthe ‘Immunoblot GAR-AP Assay kit’ (Bio-Rad)
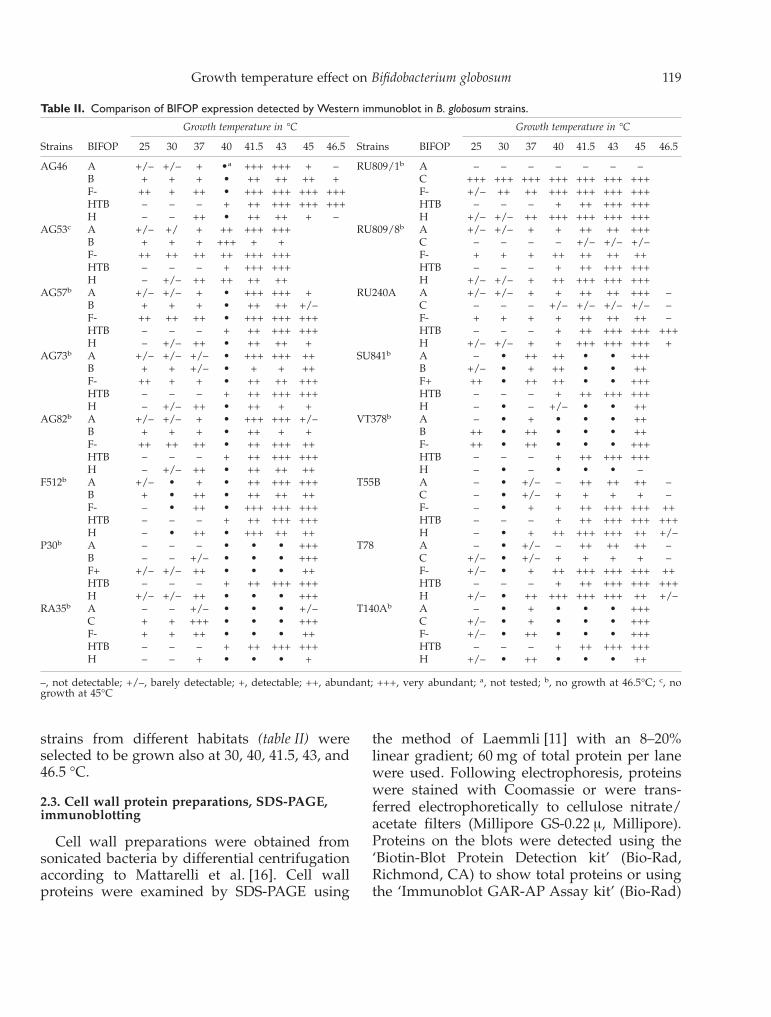
Table II. Comparison of BIFOP expression detected by Western immunoblot in B. globosum strains.
Strains BIFOP
Growth temperature in °C
Strains BIFOP
Growth temperature in °C
25 30 37 40 41.5 43 45 46.5 25 30 37 40 41.5 43 45 46.5
AG46 A +/– +/– + •a +++ +++ + – RU809/1b A – – – – – – –B + + + • ++ ++ ++ + C +++ +++ +++ +++ +++ +++ +++F- ++ + ++ • +++ +++ +++ +++ F- +/– ++ ++ +++ +++ +++ +++HTB – – – + ++ +++ +++ +++ HTB – – – + ++ +++ +++H – – ++ • ++ ++ + – H +/– +/– ++ +++ +++ +++ +++
AG53c A +/– +/ + ++ +++ +++ RU809/8b A +/– +/– + + ++ ++ +++B + + + +++ + + C – – – – +/– +/– +/–F- ++ ++ ++ ++ +++ +++ F- + + + ++ ++ ++ ++HTB – – – + +++ +++ HTB – – – + ++ +++ +++H – +/– ++ ++ ++ ++ H +/– +/– + ++ +++ +++ +++
AG57b A +/– +/– + • +++ +++ + RU240A A +/– +/– + + ++ ++ +++ –B + + + • ++ ++ +/– C – – – +/– +/– +/– +/– –F- ++ ++ ++ • +++ +++ +++ F- + + + + ++ ++ ++ –HTB – – – + ++ +++ +++ HTB – – – + ++ +++ +++ +++H – +/– ++ • ++ ++ + H +/– +/– + + +++ +++ +++ +
AG73b A +/– +/– +/– • +++ +++ ++ SU841b A – • ++ ++ • • +++B + + +/– • + + ++ B +/– • + ++ • • ++F- ++ + + • ++ ++ +++ F+ ++ • ++ ++ • • +++HTB – – – + ++ +++ +++ HTB – – – + ++ +++ +++H – +/– ++ • ++ + + H – • – +/– • • ++
AG82b A +/– +/– + • +++ +++ +/– VT378b A – • + • • • ++B + + + • ++ + + B ++ • ++ • • • ++F- ++ ++ ++ • ++ +++ ++ F- ++ • ++ • • • +++HTB – – – + ++ +++ +++ HTB – – – + ++ +++ +++H – +/– ++ • ++ ++ ++ H – • – • • • –
F512b A +/– • + • ++ +++ +++ T55B A – • +/– – ++ ++ ++ –B + • ++ • ++ ++ ++ C – • +/– + + + + –F- – • ++ • +++ +++ +++ F- – • + + ++ +++ +++ ++HTB – – – + ++ +++ +++ HTB – – – + ++ +++ +++ +++H – • ++ • +++ ++ ++ H – • + ++ +++ +++ ++ +/–
P30b A – – – • • • +++ T78 A – • +/– – ++ ++ ++ –B – – +/– • • • +++ C +/– • +/– + + + + –F+ +/– +/– ++ • • • ++ F- +/– • + ++ +++ +++ +++ ++HTB – – – + ++ +++ +++ HTB – – – + ++ +++ +++ +++H +/– +/– ++ • • • +++ H +/– • ++ +++ +++ +++ ++ +/–
RA35b A – – +/– • • • +/– T140Ab A – • + • • • +++C + + +++ • • • +++ C +/– • + • • • +++F- + + ++ • • • ++ F- +/– • ++ • • • +++HTB – – – + ++ +++ +++ HTB – – – + ++ +++ +++H – – + • • • + H +/– • ++ • • • ++
–, not detectable; +/–, barely detectable; +, detectable; ++, abundant; +++, very abundant; a, not tested; b, no growth at 46.5°C; c, nogrowth at 45°C
Growth temperature effect on Bifidobacterium globosum 119

to detect specific antigens. The detailed proce-dures for these methods are described in Mat-tarelli et al. [16].
2.4. Preparation of antisera
New Zealand white rabbits were given, onthe upper back, 8–10 intradermal injections(50–100 µL each) of antigen-Freund’s adjuvant,for a total of 100–150 µg of immunogen [3, 33].The animals were bled 8–10 weeks later and theantisera precipitated with ammonium sulphateand freeze-dried or stored at -20 °C in thepresence of 0.02% azide.
2.5.Hydrophobic interaction chromatography(HIC)
The general procedures of Smyth et al. [29]and Faris et al. [10] were used with slight modi-fications: 150 µL of octyl-Sepharose CL-4B(Pharmacia-LKB) (40 µmol mL–1 gel bed) werepacked in disposable 1 mL syringes (5 mm di-ameter) cut to 35 mm height and plugged witha disk of plastic sponge; after washing with3 mL of buffered ammonium sulphate (BAS)(10 mM sodium phosphate buffer, 1 M ammo-nium sulphate, pH 6.8), 100 µL of cell suspen-sion were allowed to drain into gel beds. Cells,previously washed with phosphate-buffered sa-line (PBS) (8 mM Na2HPO4, 2 mM KH2PO4,140 mM NaCl, 3 mM KCl, pH 7.0), were sus-pended in BAS at a concentration of 0.5 OD at550 nm, then 100 µL of this suspension wasdiluted into 3 µL of the same buffer. The drainedcells were eluted with 3 mL of BAS and eluateOD values were recorded. Values are given as apercentage of gel adsorption.
2.6. Salting-out aggregation test (SAT)
Salting out of bacterial cells was performed asdescribed by Lindahl et al. [13]. Briefly, 25 µLaliquots of cell suspension in 0.002 M sodiumphosphate buffer pH 6.8 (SPB) (ca 108 cellsmL–1) were mixed with equal volumes of SPBadded with ammonium sulphate over the rangeof 0.02 to 4.0 M usually in 0.025 decrements; themixture was made on glass depression slides
and gently rocked for 1–2 min; clumping wasvisually scored against a black background.Suspensions mixed with SPB without ammo-nium sulphate were used as negative control.The lowest concentration causing bacterial ag-gregation was the SAT value recorded.
2.7. Treatment with trypsin
Two 100 µL fractions of bacterial suspensionwere centrifuged and suspended in 200 µL of0.01 M Tris-HCl (pH 7.5) with 2 mg mL –1 (≈220total units) of trypsin (Boehringer cat.N°109–819); as control, two 100 µL fractions ofbacterial suspension were maintained in thesame buffer without adding trypsin: after 2 h at37 °C, the cells were washed twice with PBS andfinally suspended in 100 mL of BAS or SPB forthe evaluation of the HIC or SAT values, respec-tively.
3. Results
3.1. Coomassie detection
Previous studies of B. globosum strainsshowed that 53 strains out of 147 displayed thepresence of BIFOPs when grown at 37 °C [16].In the present study, 70 strains of B. globosumwere grown at temperatures of 25, 37 and 45 °C(table I). The data of BIFOP expression at 37 °Cpresented in table I are in agreement with theprevious work [16]. They show an increasingBIFOP expression from low to high growthtemperatures as revealed by SDS-PAGE; at25 °C there were no BIFOPs, at 37 °C BIFOPswere present in 20 out of 70 strains (as in theprevious study), and at 45 °C all the strainsdisplayed BIFOPs; one representative example(strain RA35) is displayed in figure 1A.
All strains showed the presence of one com-mon protein when grown at 45 °C (or at 43 °Cfor AG 53, its maximum growth temperature).For this particular protein, strains were alsogrown at 40 and 43 °C and, starting from 40 °C,the expression of this protein increased withincreasing temperature. AG 46, RU 240A, T 55Band T 78 strains, which can grow at a tempera-
120 Mattarelli et al.

ture of 46.5 °C, showed HTB (high temperatureBIFOPs) to be the only cell wall protein at thistemperature (data not shown). This protein wascalled HTB. Apart from this protein, other pro-teins were detected in different strains, up tofive to six proteins per strain (figure 1B). Theseproteins, which were also present when thestrains were grown at 37 °C, corresponded toBIFOPs A (94.5 kDa), B (87.5 kDa), C (85.5 kDa),F+ (48 kDa), F- (50 kDa), and H (37.2 kDa), asrevealed by immunoblotting reactions usingspecific antisera obtained previously [16] (datanot shown). In one strain, P 30, one additionalBIFOP of approximately 40 molr was present at45 °C (figure 2): this protein, probably typical ofthe higher temperature, was not further inves-tigated in this work. The SDS-PAGE BIFOPprofiles at 45 °C did not correlate with thehabitat from which the strains were obtained, incontrast to the results obtained at 37 °C [16].
3.2. HTB characterization
HTB migrated very close to BIFOP E(49 kDa), F+ and F-, which were detected insome BIFOP-positive strains at 37 °C [16]. More-over, it was very difficult to distinguish betweenthem because HTB was expressed in very largeamounts at 45 °C. In order to be sure that HTBwas a new BIFOP, different tests were per-formed. First, cell wall preparations and puri-fied HTB from strains grown at 45 °C andpurified BIFOP E and F+ from strains grown at37 °C were compared by SDS-PAGE; the appar-ent molr of HTB was lower than that of BIFOP Eand higher than that of BIFOP F+ (figure 2).Furthermore, the immunoblotted cell wall pro-tein profiles obtained from cells grown at 45 °Cand stained either with anti-BIFOP F+ and F-sera or for total proteins confirmed that HTBmigrated in an intermediate position compared
Figure 1. SDS-PAGE of cell-wall-related proteins from selected strains of B.globosum (Coomassie staining). A. (lanes 1–3): RA35grown at 25, 37 and 45°C, respectively. B. (lanes 1–8): T55B, T78, T86, RA143, SU861, P92, T142, T119B (grown at 45°C). Capitalletters on the right of each section refer to the BIFOP designations and the numbers in brackets are the molecular masses (in kDa).
Growth temperature effect on Bifidobacterium globosum 121

to BIFOP F+ and F- (figure 3). For better detec-tion of HTB, a specific polyclonal antiserum wasgenerated in rabbit [16] against this BIFOP pu-rified from SU 861 grown at 45 °C. The compari-son of F+, F-, E BIFOP, and HTB immunostainedusing specific antisera confirmed their differingmigrations (data not shown).
3.3. Immunological detection
In order to confirm and extend the resultsobtained with Coomassie detection, which re-vealed only abundant cell wall proteins, weused the more sensitive approach of immunob-lotting on 16 strains out of 71. With the immu-noblot technique we were able to display moreBIFOPs (table II) compared with Coomassiestaining (table III).
These strains, isolated from different animalhabitats, were selected in order to study BIFOPand HTB expression at different temperatures
(table II). All these strains showed the presenceof HTBs at temperatures of 40 °C or higher inagreement with the results obtained with Coo-massie staining. The amount of HTB expressedin these strains increased with increasing tem-perature.
The variations in A, B, F and H BIFOP con-centrations (in addition to HTB) at the differentgrowth temperatures were examined by immu-noblotting, as shown in table II. Immunologicaltests showed that the majority of BIFOP bandsincreased in intensity when cells were grown athigh temperatures.
The unusual feature of the strains isolatedfrom lamb was that BIFOP A was expressed ingreater quantity at 41.5–43 °C than at othergrowth temperatures. In the RU 809/1 plasmid-positive clone and in its ‘cured’ derivativeplasmid-negative RU 809/8, [17], BIFOP A dis-played a distinct pattern. Unexpectedly, the
Figure 2. SDS-PAGE of protein preparations (Coomassie staining). Lanes 1–3: P 30 grown at 45°C; total extract, cytoplasmic contentand cell wall preparation, respectively. Lanes 4 and 6: BIFOP E purified from T 126 and T 121 grown at 37°C. Lanes 5 and 9: cell wallpreparations of T 126 and P42 grown at 45°C. Lane 7: HTB purified from T 121. Lane 8: BIFOP F+ purified from P 42 grown at 37°C.Letters on the right refer to the BIFOP designations and the molecular masses in kDa are indicated in brackets. Arrow in lane 3 indicatesthe additional BIFOP (of approx. 40 kDa).
122 Mattarelli et al.

plasmid-positive clone grown at different tem-peratures did not express BIFOP A, which onthe contrary was present in RU 809/8 grownunder the same conditions. This suggests acorrelation between the absence of BIFOP A andthe plasmid in the RU 809/1 clone. Other fea-tures characteristic of this clone related to thepresence of the plasmid included auxotrophyfor L-leucine and higher sensitivity to lysozymeand poly-D-lysine [1, 19]. As shown in figure 4,RU809/1 grown at 25, 30, 37, and 40 °C did notshow a remarkable difference in the intensity ofBIFOP C in the cell wall preparation. An in-creasing amount of BIFOP H was observed inthe cell wall preparation with the increase intemperature; this pattern of expression does notappear in the total extract and BIFOP H wasalmost completely absent at 40 °C in the super-natant preparation. This finding suggests thatgrowth temperature could influence the trans-
location of BIFOP H to the cell wall but not itssynthesis.
3.4. Surface hydrophobicity of RU 809/1and RU 809/8 strains
As several proteins are cell wall-associated inB. globosum, their individual contribution togeneral cell hydrophobicity can be investigatedby growing cells under different temperatureconditions, in case these conditions should leadto a different modulation of the individualprotein. RU 809/1 and RU 809/8 clones, whichshow different cell wall protein patterns atdifferent temperatures, were tested for differ-ences in hydrophobic characteristics whengrown at different temperatures. Cell hydro-phobicity was measured by SAT and HICmethods; comparison of values obtained usingthe two methods showed a good correlation.
Figure 3. Cell wall preparations of cells grown at 45°C. Lanes1–2, SU861 and RA143 (SDS-PAGE/Coomassie staining); lanes3–4, RA143 and SU861 (Western blotting): 3a and 4a, immunos-taining with anti-BIFOP F; 3b and 4b, staining for total proteins.
Figure 4. Western blot reactivities of anti-BIFOP C and anti-BIFOP H polyclonal sera. Lanes 1–4, total extract, lanes 5–8,cytoplasmic content, lanes 9–12, cell wall preparation of RU809/1 cells grown at 25, 30, 37 and 40°C, respectively.
Growth temperature effect on Bifidobacterium globosum 123

The two clones examined showed high valuesof hydrophobicity at low-growth temperatures(25–27 °C), with RU 809/8 being more hydro-
phobic than RU 809/1. At medium-high growthtemperatures (35–45 °C), RU 809/1 was morehydrophobic than RU 809/8, and both clones
Table III. Comparison of BIFOP expression detected by Coomassie in B. globosum strains.
Growth temperature in °C Growth temperature in °C
Strains BIFOP 25 37 45 Strains BIFOP 25 37 45
AG 46 A – – – RU809/1 A – – –
B – – – C – +++ +++F- – – – F- – – –HTB – – +++ HTB – – +++H – – – H – +++ +++
AG 53 A – – +++a RU 809/8 A – – +
B – – –a C – – –F- – – –a F- – – –HTB – – +++a HTB – – +++H – – –a H – – –
AG 57 A – +++ +++ RU 240 A A – – –
B – – – C – – –F- – – – F- – – –HTB – – +++ HTB – – +++H – +++ ++ H – – –
AG 73 A – +++ – SU 841 A – ++ ++
B – – – B – – –F- – – – F+ – – –HTB – – +++ HTB – – +++H – + – H – ++ +++
AG 82 A – +++ – VT 378 A – +++ +++
B – – – B – ++ +++F- – – – F- – ++ +++HTB – – +++ HTB – – +++H – +++ +++ H – ++ +++
F 512 A – +++ +++ T 55 B A – – –
B – +++ – C – – –F- – ++ +++ F- – – –HTB – – +++ HTB – – +++H – +++ – H – – +++
P 30 A – – +++ T 78 A – – –
B – – – C – – –F+ – – – F- – – –HTB – – +++ HTB – – +++H – – H – – ++
RA 35 A – – – T 140A A – +++ +++
C – +++ +++ C – – –F- – – – F- – +++ +++HTB – – +++ HTB – – +++H – – – H – ++ –
–, not detectable; +, detectable; ++, abundant; +++, very abundant; a, BIFOP at 43 °C, the maximum growth temperaturefor this strain
124 Mattarelli et al.
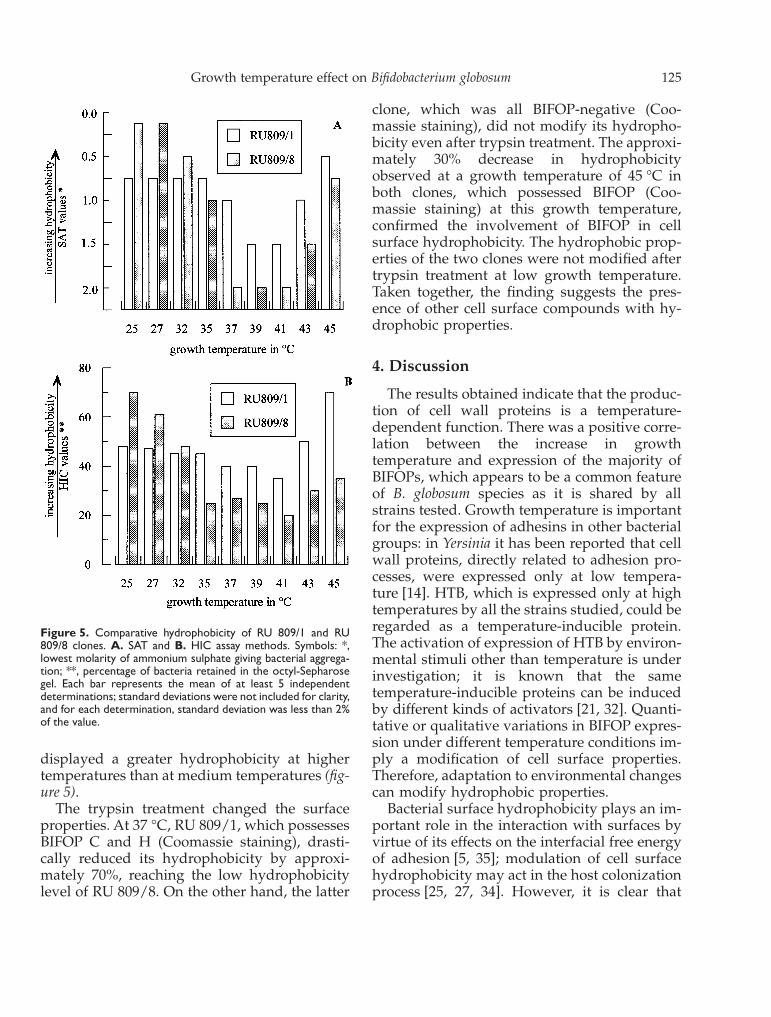
displayed a greater hydrophobicity at highertemperatures than at medium temperatures (fig-ure 5).
The trypsin treatment changed the surfaceproperties. At 37 °C, RU 809/1, which possessesBIFOP C and H (Coomassie staining), drasti-cally reduced its hydrophobicity by approxi-mately 70%, reaching the low hydrophobicitylevel of RU 809/8. On the other hand, the latter
clone, which was all BIFOP-negative (Coo-massie staining), did not modify its hydropho-bicity even after trypsin treatment. The approxi-mately 30% decrease in hydrophobicityobserved at a growth temperature of 45 °C inboth clones, which possessed BIFOP (Coo-massie staining) at this growth temperature,confirmed the involvement of BIFOP in cellsurface hydrophobicity. The hydrophobic prop-erties of the two clones were not modified aftertrypsin treatment at low growth temperature.Taken together, the finding suggests the pres-ence of other cell surface compounds with hy-drophobic properties.
4. Discussion
The results obtained indicate that the produc-tion of cell wall proteins is a temperature-dependent function. There was a positive corre-lation between the increase in growthtemperature and expression of the majority ofBIFOPs, which appears to be a common featureof B. globosum species as it is shared by allstrains tested. Growth temperature is importantfor the expression of adhesins in other bacterialgroups: in Yersinia it has been reported that cellwall proteins, directly related to adhesion pro-cesses, were expressed only at low tempera-ture [14]. HTB, which is expressed only at hightemperatures by all the strains studied, could beregarded as a temperature-inducible protein.The activation of expression of HTB by environ-mental stimuli other than temperature is underinvestigation; it is known that the sametemperature-inducible proteins can be inducedby different kinds of activators [21, 32]. Quanti-tative or qualitative variations in BIFOP expres-sion under different temperature conditions im-ply a modification of cell surface properties.Therefore, adaptation to environmental changescan modify hydrophobic properties.
Bacterial surface hydrophobicity plays an im-portant role in the interaction with surfaces byvirtue of its effects on the interfacial free energyof adhesion [5, 35]; modulation of cell surfacehydrophobicity may act in the host colonizationprocess [25, 27, 34]. However, it is clear that
Figure 5. Comparative hydrophobicity of RU 809/1 and RU809/8 clones. A. SAT and B. HIC assay methods. Symbols: *,lowest molarity of ammonium sulphate giving bacterial aggrega-tion; **, percentage of bacteria retained in the octyl-Sepharosegel. Each bar represents the mean of at least 5 independentdeterminations; standard deviations were not included for clarity,and for each determination, standard deviation was less than 2%of the value.
Growth temperature effect on Bifidobacterium globosum 125

caution is necessary in attempting to correlatesingle surface molecules with complex ‘bulk’properties of the whole bacterium such as hy-drophobicity or adherence, without detailedanalysis of the molecules that are present.
In a previous study on Bifidobacterium bifidum,Op den Camp et al. [23] concluded that bothproteins and lipoteichoic acids, independentlyor in dynamic complexes, are responsible forthe hydrophobic feature of bifidobacterial cellsurfaces. Mukai et al. [22] suggested that the cellsurface proteinaceous components are involvedin the adhesion of Bifidobacterium adolescentis.
In the present study, the influence of growthtemperature on hydrophobic properties wasstudied in RU 809/1 (plasmid-positive) and RU809/8 (plasmid-negative) clones. Our data sug-gest that low and high growth temperaturescorrespond to high values of hydrophobicitycompared to a medium-growth temperature of37 °C. There is a positive, though not absolute,correlation between the presence of BIFOP, asrevealed by Coomassie staining, and hydropho-bicity, even if no evidence is given that one ormore of these cell wall proteins is directlyresponsible for the hydrophobicity properties.At 25 °C, no clones express BIFOP and treat-ment with trypsin does not influence cell hydro-phobicity. At 37 °C, RU 809/1 expresses BIFOPand trypsin treatment reduces hydrophobicityonly in this clone. At 45 °C, both clones expressBIFOP and, after trypsin treatment, a decreasein hydrophobicity of approximately 30% wasobserved in both clones. Comparable effects,showing that hydrophobicity properties wereprotease-sensitive, were observed in other mi-croorganisms such as streptococci [30]. Further-more, the high hydrophobicity of cells in bothclones grown at 25 °C compared to 37 °C sug-gests the presence of other nonproteinaceousstructures, i.e. lipoteichoic acids, or othermechanisms involved in conferring surfaceproperties. Future studies will investigate themolecular events involved in Bifidobacterium ad-herence and lead to a better characterization ofadhesion molecules.
Résumé — Effet de la température de croissancesur la biosynthèse de la paroi de Bifidobacteriumglobosum. Soixante-dix souches de Bifidobacteriumglobosum isolées du tube digestif de plusieurs ani-maux ont été l’objet de cette étude. L’effet d’unegamme de températures de culture de 25 à 46,5 °C aété observé sur la composition des protéines, BIFOP(bifidobacterial outer protein), spécifiques de la paroibactérienne des bifidobactéries. et sur l’hydrophobi-cité des bactéries. Nous avons montré que la biosyn-thèse des BIFOP est dépendante de la températurede croissance. En effet, elle est très faible à bassetempérature (25–30°C) et augmente avec la tempéra-ture de croissance. En outre, nous avons mis enévidence chez toutes les souches étudiées, une nou-velle protéine de 48.5 kDa, appelée HTB (high tempe-rature BIFOP). L’hydrophobicité de quelques sou-ches a aussi été étudiée après croissance des bactériesà différentes températures. Il semble que cette hy-drophobicité soit principalement due aux BIFOPlorsque les bactéries se développent à 37 °C; si lesbactéries croissent à des températures inférieures ousupérieures à 37 °C, les BIFOP semblent contribuerdans une proportion bien plus faible à l’hydropho-bicité cellulaire. © Elsevier, Paris
Bifidobacterium / colonisation bactérienne /hydrophobicité / protéines température-induc-tibles / protéines de la paroi bactérienne
References
[1] Absolom D.R., Lamberti F.V., Policova Z., Zingg W., VanOss C.J.,Neumann A.W., Surface thermodynamics of bacterial adhesion,Appl. Environ. Microbiol. 46 (1983) 90–97.
[2] Ballongue J., Bifidobacteria and probiotic action, in: SalminenS.,Avonwright A., (Eds.), Lactic Acid Bacteria, Marcel Dekker Inc.,New York, N.Y., 1993, pp. 357–428.
[3] Bailey G.S., The production of antisera, in: Walker J. M., (Ed.),Methods in molecular biology, Vol. I, Proteins, Humana Press,Clifton, N.J., 1984.
[4] Biavati B., Sgorbati B., Scardovi V., The genus Bifidobacterium, in:Balows A., Trüper H. G., Dworkin M., Harder W., Schleifer K. H.,(Eds.), The Prokaryotes a handbook on the biology of bacteriaecophysiology, isolation, identification, applications, Second Edi-tion, Springer Verlag, Heidelberg, New York, N.Y., 1991, pp.816–837.
[5] Busscher H.J., Weerkamp A.H., Van Der Mei H.C., VanPelt A.W.J.,Jong H.P., Arends J., Measurements of the surface free energy ofbacterial cell surfaces and its relevance for adhesion, Appl. Environ.Microbiol. 48 (1984) 980–983.
[6] Christensen G.D., Simpson W.A., Beachey E.H., Adhesion of bac-teria to animal tissues complex mechanisms, in: Savage D. C.,Fletcher M., (Eds.), Bacterial adhesion, Plenum Press, New York,N.Y., 1985, pp. 279–305.
[7] Conway P.L., Kjelleberg S., Protein-mediated adhesion of Lactoba-cillus fermentum strain 737 to mouse stomach squamous epithelium,J. Gen. Microbiol. 135 (1989) 1175–1186.
126 Mattarelli et al.

[8] Doyle R., Rosenberg M., Microbial cell surface hydrophobici-ty.(Eds.), American Society for Microbiology, Washington, D.C.,1990.
[9] Elmer G.W., Surawicz C.M., McFarlane L.V., Biotherapeutic agents,JAMA 275 (1996) 870–876.
[10] Faris A., Wadström T., Freer J.H., Hydrophobic adsorptive andhemagglutinating properties of Escherichia coli possessing colonisa-tion factor antigens (CFA/I or CFA/II), type 1 pili, or other pili, Curr.Microbiol. 5 (1981) 67–72.
[11] Laemmli, U.K., Cleavage of structural proteins during the assemblyof the head of bacteriophage T4, Nature 227 (1970) 680–685.
[12] Lathigra R.B., Butcher P.D., Garbe T.R., Young D.B., Heat shockproteins as virulence factors of pathogens, Curr. Top. Microbiol.Immunol. 167 (1991) 125–143.
[13] Lindahl M., Faris A., Wadström T., Hjertén S., A new based on‘salting out’ to measure relative surface hydrophobicity of bacterialcells, Biochim. Biophys. Acta 677 (1981) 471–476.
[14] Maclagan R.M., Old D.C., Haemagglutinins and fimbriae in differentserotypes and biotypes of Yersinia enterocolitica, J. Appl. Bacteriol. 49(1980) 353–360.
[15] Mafu A.A., Roy D., Goulet J., Savoie L., Characterization of physi-cochemical forces involved in adhesion of Listeria monocytogenesto surfaces, Appl. Environ. Microbiol. 57 (1991) 1969–1973.
[16] Mattarelli P., Biavati B., Crociani F., Scardovi V., Prati G., Bifidobac-terial cell-wall proteins (BIFOP) in Bifidobacterium globosum, Res.Microbiol. 144 (1993) 581–590.
[17] Mattarelli, P., Biavati, B., Alessandrini, A., Crociani, F., Scardovi, V.,Characterization of the plasmid pVS809 from Bifidobacterium globo-sum, Microbiologica, 17 (1994) 327–331.
[18] Mattarelli P., Crociani F.,, Biavati B., Bactericidal activity of poly-D-lysine and lysozyme in Bifidobacterium globosum strains, Ann. Micro-biol. Enzymol. 47 (1997) 185–191.
[19] Mattarelli P., Biavati B., L-leucine auxotrophy in Bifidobacteriumglobosum strains, Microbiologica (1998) in press.
[20] Millsap K.W., Reid G., Van DerMei H.C., Busscher H.J., Adhesion ofLactobacillus species in urine and phosphate buffer to siliconerubber and glass under flow, Biomaterials 18 (1997) 87–91.
[21] Morgan R.W., Christman M.F., Jacobson F.S., Storz G., Ames B.N.,Hydrogen-peroxide-inducible proteins in Salmonella typhimuriumoverlap heat shock and other stress proteins, Proc. Natl. Acad. Sci.USA 83 (1988) 8059–8063.
[22] Mukai T., Taba T., Ohori H., Collagen binding of Bifidobacteriumadolescentis, Curr. Microbiol. 34 (1997) 326–331.
[23] Op DenCamp H.J.M., Oosterhof A., Veerkamp J.H., Cell surfacehydrophobicity of Bifidobacterium bifidum subsp. Pennsylvanicum,Antonie van Leeuwenhoek, 51 (1985) 303–312.
[24] O’ Sullivan M.G., Thornton G.C., O’Sullivan G.C., Collins J.K.,Probiotic bacteria myth or reality ? Trends Food Sci. Technol. 31(1992) 309–314.
[25] Pérez P.F., Minnaard Y., Disalvo E.A., DeAntoni G.L., Surface prop-erties of bifidobacterial strains of human origin, Appl. Environ.Microbiol. 64 (1998) 21–26.
[26] Rijnaarts H.H.M., Norde W., Bouwer E.J., Lyklema J., ZehnderA.J.B., Bacterial adhesion under static and dynamic conditions,Appl. Environ. Microbiol. 59 (1993) 3255–3265.
[27] Savoia D., Malcangi A., Martinetto P., The effect of subinhibitoryconcentrations of some antibiotics on the hydrophobicity of gram-negative bacteria, J. Chemoth. 2 (1990) 20–25.
[28] Scardovi, V., Genus Bifidobacterium Orla-Jense, in: Sneath P. H. A.,Mair N. S., Sharpe M. E., Holt J. G., (Eds.), Bergey’s Manual ofSystematic Bacteriology, Second Volume, Williams and Wilkins,Baltimore, B.A., 1986, pp. 1418–1434.
[29] Smyth C.J., Jonsson P., Olsson E., Söderlind O., Rosengren J.,Hjertén S., Wadström T., Differences in hydrophobic surface char-acteristics of porcine enteropathogenic Escherichia coli with orwithout K88 antigen as revealed by hydrophobic interaction chro-matography, Inf. Immun. 22 (1978) 462–472.
[30] Tamura G.S., Kuypers J.M., Smith S., Raff H., Rubens C.E., Adher-ence of group B streptococci to cultured epithelial cells roles ofenvironmental factors and bacterial surface components, Inf. Im-mun. 62 (1994) 2450–2458.
[31] Teti G.F., Tomasello F., Chiofalo M.S., Orefici G., Mastroeni P.,Adherence of group B streptococci to adult and neonatal epithelialcells mediated by lipoteichoic acids, Inf. Immun. 55 (1987)3057–3064.
[32] Young R.A., Stress proteins and immunology, Ann. Rev. Immunol. 8(1990) 401–420.
[33] Vaitukaitis J.L., Production of antisera with small doses of immuno-gen mutiple intradermal injections, in: Langone J. J., Van Vunaakis H.,Methods in enzymolgy, vol.73, Academic Press, New York, N.Y.,1981, pp. 46–57.
[34] VanLoosdrecht M.C.M., Lyklema J., Norde W., Schraa G., ZehnderA.J.B., The role of bacterial cell wall hydrophobicity in adhesion,App. Environm. Microbiol. 53 (1987) 1893–1897.
[35] Weerkamp A.H., Van DerMei H.C., Busscher H.J., The surface freeenergy of oral streptococci after being coated with saliva and itsrelation to adhesion in the mouth, J. Dent. Res. 64 (1985)1204–1210.
Growth temperature effect on Bifidobacterium globosum 127




















