
Shuttle mutagenesis and targeted disruption of a telomere-located essential gene of Leishmania
Upload
independentCategory
view
2download
0
BC10, a DUF266-containing and Golgi-located type IImembrane protein, is required for cell-wall biosynthesisin rice (Oryza sativa L.)
Yihua Zhou1,†, Shengben Li1,†, Qian Qian2,†, Dali Zeng1, Mu Zhang1, Longbiao Guo2, Xiangling Liu1, Baocai Zhang1,
Lingwei Deng1, Xinfang Liu1, Guanzheng Luo1, Xiujie Wang1 and Jiayang Li1,*
1State Key Laboratory of Plant Genomics and National Center for Plant Gene Research, Institute of Genetics and Developmental
Biology, Chinese Academy of Sciences, Beijing 100101, China, and2State Key Laboratory of Rice Biology, China National Rice Research Institute, Chinese Academy of Agricultural Sciences,
Hangzhou 310006, China
Received 7 June 2008; revised 6 September 2008; accepted 16 September 2008; published online 7 November 2008.
*For correspondence (fax +86 10 64873428; email [email protected]).†These authors contributed equally to this work.
Summary
Glycosyltransferases (GTs) are one of the largest enzyme groups required for the synthesis of complex wall
polysaccharides and glycoproteins in plants. However, due to the limited number of related mutants that have
observable phenotypes, the biological function(s) of most GTs in cell-wall biosynthesis and assembly have
remained elusive. We report here the isolation and in-depth characterization of a brittle rice mutant, brittle
culm 10 (bc10). bc10 plants show pleiotropic phenotypes, including brittleness of the plant body and retarded
growth. The BC10 gene was cloned through a map-based approach, and encodes a Golgi-located type II
membrane protein that contains a domain designated as ‘domain of unknown function 266’ (DUF266) and
represents a multiple gene family in rice. BC10 has low sequence similarity with the domain to a core 2 b-1,6-
N-acetylglucosaminyltransferase (C2GnT), and its in vitro enzymatic activity suggests that it functions as a
glycosyltransferase. Monosaccharide analysis of total and fractioned wall residues revealed that bc10 showed
impaired cellulose biosynthesis. Immunolocalization and isolation of arabinogalactan proteins (AGPs) in the
wild-type and bc10 showed that the level of AGPs in the mutant is significantly affected. BC10 is mainly
expressed in the developing sclerenchyma and vascular bundle cells, and its deficiency causes a reduction in
the levels of cellulose and AGPs, leading to inferior mechanical properties.
Keywords: mechanical strength, glycosyltransferase, cell wall, cellulose, arabinogalactan protein, rice.
Introduction
The plant cell wall is a rigid and dynamic network system
comprising polysaccharides, aromatic substances and gly-
coproteins. Walls are not only central to determining the
mechanical properties of all organs, but also play funda-
mental roles in regulating plant growth and development
and are critical for a wide range of cell functions including
cell adhesion, cell-to-cell communication, and defense
responses (Bacic et al., 1988; Carpita and Gibeaut, 1993; Ellis
et al., 2002; Vogel et al., 2004). The Arabidopsis genome
sequencing project is now complete, and it has been
estimated that, of approximately 27 000 Arabidopsis genes,
approximately 15% are dedicated to the processes of cell-
wall synthesis, modification, assembly and degradation,
and correct regulation of these processes during growth and
development (Carpita et al., 2001). Therefore, the cell wall is
one of the most complicated and the least-understood plant
cell structures. A powerful approach to probe the functions
of individual components of cell walls is through the iden-
tification and characterization of mutants (Reiter et al.,
1997). Mutants defective in mechanical strength, a promi-
nent physical property of the cell wall that depends upon the
structure of various wall polymers and how they interact,
446 ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd
The Plant Journal (2009) 57, 446–462 doi: 10.1111/j.1365-313X.2008.03703.x

have been proven to be valuable for identifying genes
involved in the biogenesis and modification of cell walls (Li
et al., 2003b; Tanaka et al., 2003; Taylor et al., 2003; Zhong
et al., 2002). Although much progress has been made in this
area, the molecular mechanisms of how the wall polysac-
charides and glycoproteins are synthesized, deposited and
regulated remain to be elucidated (Cosgrove, 2005; Farrokhi
et al., 2006; Lerouxel et al., 2006; Lytovchenko et al., 2007;
Somerville, 2006; Zhong and Ye, 2007).
Wall polysaccharides and glycoproteins have tremendous
structural complexity, and therefore plants require large
families of glycosyltransferases (GTs) to facilitate their
biosynthesis. Based on sequence similarities and the exis-
tence of certain motifs, hydrophobic clusters and their
catalytic specificity (Rosen et al., 2004; Ross et al., 2001),
GTs are divided into 91 families (http://www.cazy.org/CAZY/).
Among these, both cellulose synthase active subunits
(CESA) (Somerville, 2006) and CESA-like proteins (CSL) are
classified into the GT2 family; the former are responsible for
synthesizing cellulose and the latter are believed to be
responsible for the formation of glycan backbones in the
endoplasmic reticulum (ER) or Golgi apparatus (Burton
et al., 2006; Dhugga et al., 2004). The remaining GTs
distributed in various GT families are usually Golgi-localized
type II integral membrane proteins (Geshi et al., 2004),
which synthesize the backbones of pectin (Bacic, 2006) and
xylan (Faik et al., 2000; Madson et al., 2003; Pena et al.,
2007), or add side chains to polysaccharides (Persson et al.,
2007; Sarria et al., 2001). In addition to polysaccharides,
most wall proteins are also highly glycosylated, and some
have been proposed to interact with the major carbohydrate
components of the wall to form a complex network. Of
these, arabinogalactan proteins (AGPs), located at the outer
leaflets of the plasma membrane or secreted into the cell
wall, are regarded as critical components in maintaining
both the physical and biological functions of walls (Takeda
and Fry, 2004). AGPs often contain >90% carbohydrate,
which is essential for their biological functions (van Hengel
et al., 2001; Wu et al., 2000). Therefore, a large number of
GTs are probably required to synthesize the highly complex
sugar chains of AGPs. However, no enzymes responsible for
the glycosylation of AGPs have yet been identified or
characterized (Gaspar et al., 2001).
In Arabidopsis, more than 400 putative GT genes have
been identified based on their sequences (http://afmb.
cnrs-mrs.fr/CAZY/) (Scheible and Pauly, 2004). However, a
recent survey of the Arabidopsis and rice genomes has
increased that number, and indicates that the proportion of
these enzymes in plant genomes is greater than that in
human or other fully sequenced genomes (Coutinho et al.,
2003). It is likely that each linkage in polysaccharides and
glycoproteins requires the action of a distinct glycosyl-
transferase, leading to the prediction that multi-cellular
organisms contain hundreds of GTs (Henrissat and Davies,
2000). However, little is known of their molecular basis of
reaction specificity, including donor selectivity and acceptor
preference, or their biological functions (Pagny et al., 2003;
Wenderoth and von Schaewen, 2000). Glycosylation in
animal systems most frequently occurs as a post-transla-
tional modification of proteins rather than in polysaccharide
biosynthesis, the exception being hyaluronic acid. Unlike
plants, only about 200 human genes encode proteins related
to the biosynthesis, function and degradation of sugar
chains. For example, core 2 b-1,6-N-acetylglucosaminyl-
transferase (C2GnT), which forms the core 2 branched
O-glycan, is a glycosyltransferase that is involved in bio-
synthesis of mucin-type glycoproteins (O-glycans) (Fukuda,
1996). Generally, GTs identified in animals show low
sequence identity to those in plants, indicating significant
variation between plants and animals.
The enormous number of sequences available in
databases makes accurate sequence annotation a great
challenge, especially when there are large numbers of
functionally uncharacterized families. Many proteins that
contain domains of unknown function (DUF) have been
predicted in plants (rice and Arabidopsis) and other eukary-
otic genomes (Bateman et al., 2004), but their biological
functions remain to be elucidated. Here, we report the
isolation of a brittle culm mutant (bc10) from rice, cloning of
the gene responsible for the mutant phenotype, and molec-
ular characterization of BC10, which is a DUF266-containing
Golgi-localized glycosyltransferase that perturbs the levels
of AGPs and cellulose, thereby affecting the mechanical
properties of rice plants.
Results
bc10 plants are defective in mechanical strength
Culms and leaves are two major aerial organs of the rice
plant. Sclerenchyma cells and vascular bundles within
these two organs provide mechanical support for body
of the rice plant. To understand the mechanism that regu-
lates the mechanical properties of cereal plants, we sys-
tematically isolated and characterized rice mutants that
show the brittle culm (bc) phenotype (Li et al., 2003b). bc10
was isolated from a japonica cultivar Huang Jin Qin
(Figure 1). The prominent phenotype of bc10 plants is their
decreased mechanical strength in aerial organs, resulting in
brittle culms and leaves that can be broken easily by
bending. To accurately describe the phenotype, we quan-
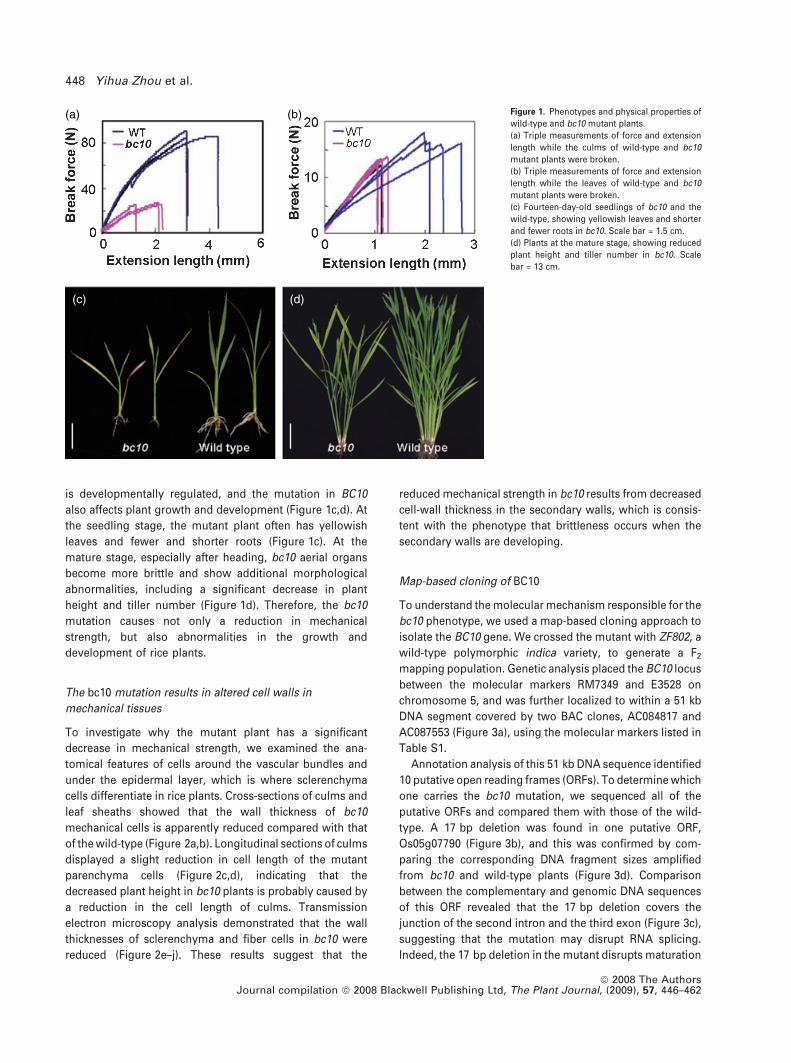
titatively compared the breaking forces of 140-day-old bc10
and wild-type plants. Figure 1(a,b) shows that the forces
needed to break the bc10 culms and leaves were reduced to
25 and 80%, respectively, compared to the wild-type. The
extension lengths, which reflect the elasticity of plant
tissues, were also significantly decreased in mutant culms
and leaves (Figure 1a,b). In addition, the brittle phenotype
BC10 regulates mechanical strength in rice 447
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462
is developmentally regulated, and the mutation in BC10
also affects plant growth and development (Figure 1c,d). At
the seedling stage, the mutant plant often has yellowish
leaves and fewer and shorter roots (Figure 1c). At the
mature stage, especially after heading, bc10 aerial organs
become more brittle and show additional morphological
abnormalities, including a significant decrease in plant
height and tiller number (Figure 1d). Therefore, the bc10
mutation causes not only a reduction in mechanical
strength, but also abnormalities in the growth and
development of rice plants.
The bc10 mutation results in altered cell walls in
mechanical tissues
To investigate why the mutant plant has a significant
decrease in mechanical strength, we examined the ana-
tomical features of cells around the vascular bundles and
under the epidermal layer, which is where sclerenchyma
cells differentiate in rice plants. Cross-sections of culms and
leaf sheaths showed that the wall thickness of bc10
mechanical cells is apparently reduced compared with that
of the wild-type (Figure 2a,b). Longitudinal sections of culms
displayed a slight reduction in cell length of the mutant
parenchyma cells (Figure 2c,d), indicating that the
decreased plant height in bc10 plants is probably caused by
a reduction in the cell length of culms. Transmission
electron microscopy analysis demonstrated that the wall
thicknesses of sclerenchyma and fiber cells in bc10 were
reduced (Figure 2e–j). These results suggest that the
reduced mechanical strength in bc10 results from decreased
cell-wall thickness in the secondary walls, which is consis-
tent with the phenotype that brittleness occurs when the
secondary walls are developing.
Map-based cloning of BC10
To understand the molecular mechanism responsible for the
bc10 phenotype, we used a map-based cloning approach to
isolate the BC10 gene. We crossed the mutant with ZF802, a
wild-type polymorphic indica variety, to generate a F2
mapping population. Genetic analysis placed the BC10 locus
between the molecular markers RM7349 and E3528 on
chromosome 5, and was further localized to within a 51 kb
DNA segment covered by two BAC clones, AC084817 and
AC087553 (Figure 3a), using the molecular markers listed in
Table S1.
Annotation analysis of this 51 kb DNA sequence identified
10 putative open reading frames (ORFs). To determine which
one carries the bc10 mutation, we sequenced all of the
putative ORFs and compared them with those of the wild-
type. A 17 bp deletion was found in one putative ORF,
Os05g07790 (Figure 3b), and this was confirmed by com-
paring the corresponding DNA fragment sizes amplified
from bc10 and wild-type plants (Figure 3d). Comparison
between the complementary and genomic DNA sequences
of this ORF revealed that the 17 bp deletion covers the
junction of the second intron and the third exon (Figure 3c),
suggesting that the mutation may disrupt RNA splicing.
Indeed, the 17 bp deletion in the mutant disrupts maturation
(a)
(c) (d)
(b) Figure 1. Phenotypes and physical properties of
wild-type and bc10 mutant plants.
(a) Triple measurements of force and extension
length while the culms of wild-type and bc10
mutant plants were broken.
(b) Triple measurements of force and extension
length while the leaves of wild-type and bc10
mutant plants were broken.
(c) Fourteen-day-old seedlings of bc10 and the
wild-type, showing yellowish leaves and shorter
and fewer roots in bc10. Scale bar = 1.5 cm.
(d) Plants at the mature stage, showing reduced
plant height and tiller number in bc10. Scale
bar = 13 cm.
448 Yihua Zhou et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462
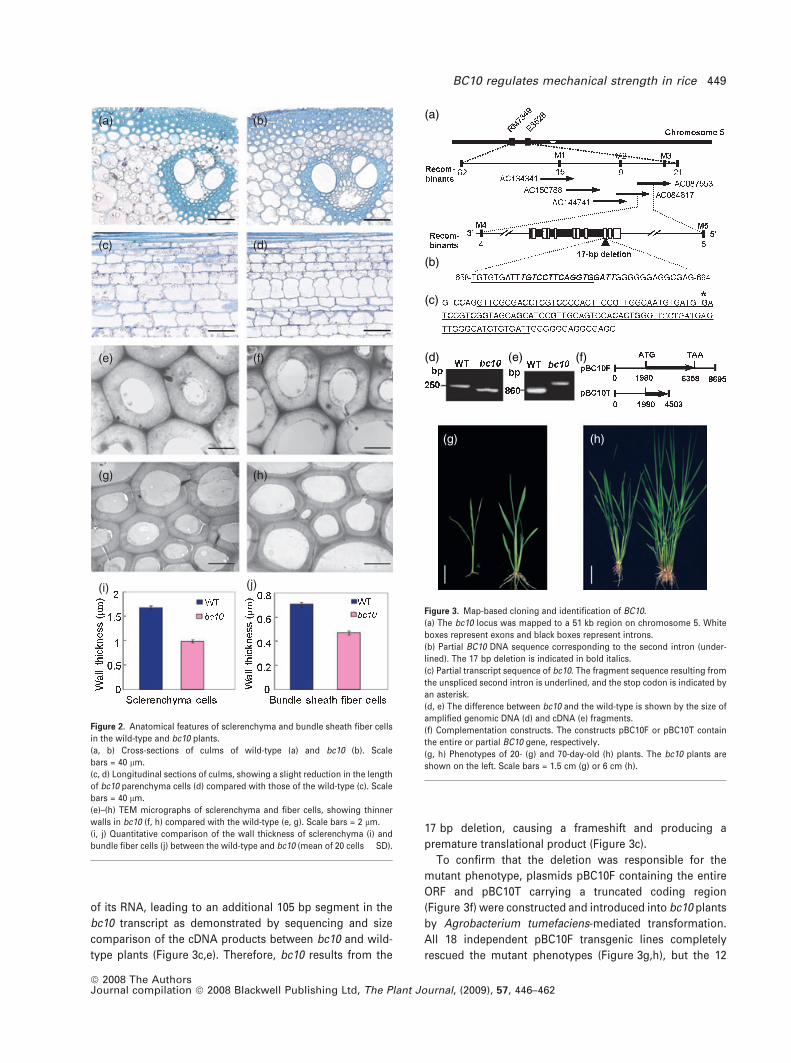
of its RNA, leading to an additional 105 bp segment in the
bc10 transcript as demonstrated by sequencing and size
comparison of the cDNA products between bc10 and wild-
type plants (Figure 3c,e). Therefore, bc10 results from the
17 bp deletion, causing a frameshift and producing a
premature translational product (Figure 3c).
To confirm that the deletion was responsible for the
mutant phenotype, plasmids pBC10F containing the entire
ORF and pBC10T carrying a truncated coding region
(Figure 3f) were constructed and introduced into bc10 plants
by Agrobacterium tumefaciens-mediated transformation.
All 18 independent pBC10F transgenic lines completely
rescued the mutant phenotypes (Figure 3g,h), but the 12
(a) (b)
(c) (d)
(e) (f)
(g) (h)
(i) (j)
Figure 2. Anatomical features of sclerenchyma and bundle sheath fiber cells
in the wild-type and bc10 plants.
(a, b) Cross-sections of culms of wild-type (a) and bc10 (b). Scale
bars = 40 lm.
(c, d) Longitudinal sections of culms, showing a slight reduction in the length
of bc10 parenchyma cells (d) compared with those of the wild-type (c). Scale
bars = 40 lm.
(e)–(h) TEM micrographs of sclerenchyma and fiber cells, showing thinner
walls in bc10 (f, h) compared with the wild-type (e, g). Scale bars = 2 lm.
(i, j) Quantitative comparison of the wall thickness of sclerenchyma (i) and
bundle fiber cells (j) between the wild-type and bc10 (mean of 20 cells � SD).
(a)
(b)
(c)
(d)
(g) (h)
(e) (f)
Figure 3. Map-based cloning and identification of BC10.
(a) The bc10 locus was mapped to a 51 kb region on chromosome 5. White
boxes represent exons and black boxes represent introns.
(b) Partial BC10 DNA sequence corresponding to the second intron (under-
lined). The 17 bp deletion is indicated in bold italics.
(c) Partial transcript sequence of bc10. The fragment sequence resulting from
the unspliced second intron is underlined, and the stop codon is indicated by
an asterisk.
(d, e) The difference between bc10 and the wild-type is shown by the size of
amplified genomic DNA (d) and cDNA (e) fragments.
(f) Complementation constructs. The constructs pBC10F or pBC10T contain
the entire or partial BC10 gene, respectively.
(g, h) Phenotypes of 20- (g) and 70-day-old (h) plants. The bc10 plants are
shown on the left. Scale bars = 1.5 cm (g) or 6 cm (h).
BC10 regulates mechanical strength in rice 449
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462
pBC10T lines failed to complement the bc10 mutant, and we
therefore conclude that the Os05g07790 is the BC10 gene.
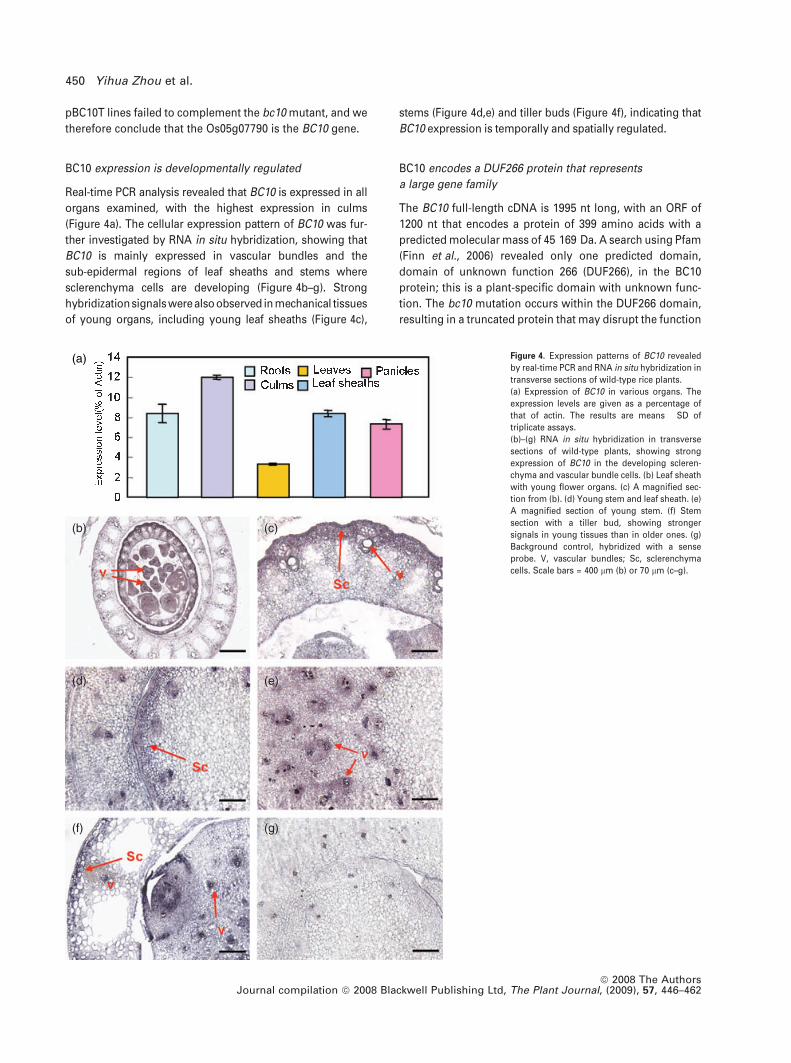
BC10 expression is developmentally regulated
Real-time PCR analysis revealed that BC10 is expressed in all
organs examined, with the highest expression in culms
(Figure 4a). The cellular expression pattern of BC10 was fur-
ther investigated by RNA in situ hybridization, showing that
BC10 is mainly expressed in vascular bundles and the
sub-epidermal regions of leaf sheaths and stems where
sclerenchyma cells are developing (Figure 4b–g). Strong
hybridizationsignalswerealsoobserved in mechanical tissues
of young organs, including young leaf sheaths (Figure 4c),
stems (Figure 4d,e) and tiller buds (Figure 4f), indicating that
BC10 expression is temporally and spatially regulated.
BC10 encodes a DUF266 protein that represents
a large gene family
The BC10 full-length cDNA is 1995 nt long, with an ORF of
1200 nt that encodes a protein of 399 amino acids with a
predicted molecular mass of 45 169 Da. A search using Pfam
(Finn et al., 2006) revealed only one predicted domain,
domain of unknown function 266 (DUF266), in the BC10
protein; this is a plant-specific domain with unknown func-
tion. The bc10 mutation occurs within the DUF266 domain,
resulting in a truncated protein that may disrupt the function
(a)
(b) (c)
(d) (e)
(f) (g)
Figure 4. Expression patterns of BC10 revealed
by real-time PCR and RNA in situ hybridization in
transverse sections of wild-type rice plants.
(a) Expression of BC10 in various organs. The
expression levels are given as a percentage of
that of actin. The results are means � SD of
triplicate assays.
(b)–(g) RNA in situ hybridization in transverse
sections of wild-type plants, showing strong
expression of BC10 in the developing scleren-
chyma and vascular bundle cells. (b) Leaf sheath
with young flower organs. (c) A magnified sec-
tion from (b). (d) Young stem and leaf sheath. (e)
A magnified section of young stem. (f) Stem
section with a tiller bud, showing stronger
signals in young tissues than in older ones. (g)
Background control, hybridized with a sense
probe. V, vascular bundles; Sc, sclerenchyma
cells. Scale bars = 400 lm (b) or 70 lm (c–g).
450 Yihua Zhou et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462

of DUF266 (Figure 5a). A BLASTX search for BC10 homologs
identified 18 and 23 DUF266-containing proteins in the Oryza
sativa (japonica) and Arabidopsis thaliana genomes,
respectively. Phylogenetic analysis revealed that these BC10
homologs are clustered into three major clades, implying
that they may have related but distinct functions (Figure 5b).
The closest homolog of BC10 is an Arabidopsis protein,
At5g14550, with 82% sequence identity. RT-PCR amplifica-
tion of total RNAs isolated from various organs of rice
revealed that the BC10-homologous genes tested are
transcriptionally active, with distinct expression patterns
(Figure 5c), suggesting that these DUF266-containing genes
are transcriptionally regulated and may play similar or
partially redundant roles in plant growth and development.
To further confirm whether the DUF266 domain is only
present in plants, we performed profile search for all
proteins in the NCBI nr database (ftp://ftp.ncbi.nih.gov/
blast/db/FASTA/nr.gz). The complete DUF266 domain was
only found in predicted proteins from plant species, such as
Arabidopsis, rice (Oryza sativa), spruce (Picea sitchensis),
poplar (Populus trichocarpa), clover (Medicago truncatula),
grape (Vitis vinifera) and moss (Physcomitrella patens),
indicating that DUF266 is very likely a plant-specific domain.
BC10 is a Golgi-localized type II integral membrane protein
Analysis using TMHMM2.0 (http://www.cbs.dtu.dk/services/
TMHMM-2.0/) showed that BC10 is a type II membrane
(a)
(c)
(b)
Figure 5. Prediction of the primary sequence, phylogenetic tree, and RT-PCR analyses of BC10 homologs.
(a) Deduced amino acid sequence of BC10. Numbers on the left refer to the positions of amino acid residues. The N-terminal cytosolic tail is indicated in bold italics,
the transmembrane domain is shown in white letters on a grey background, and the DUF266 domain is underlined.
(b) Phylogenetic analysis of BC10 homologs from rice and Arabidopsis. The numbers at each node represent the bootstrap support (percentage), and the scale bar is
an indicator of genetic distance based on branch length.
(c) RT-PCR analyses of BC10 homologs, indicating that the BC10 homologs tested are transcribed with various organ patterns.
BC10 regulates mechanical strength in rice 451
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462

protein that consists of a short cytoplasmic N-terminus
followed by a single transmembrane helix and a long
non-cytoplasmic C-terminus containing the DUF266 domain
(Figure 6a). To determine the membrane topology of BC10,
we generated transgenic rice plants that over-express a
BC10–GFP fusion protein in the bc10 background. The mu-
tant phenotypes were fully rescued in the transgenic plants
(data not shown), indicating that the BC10–GFP fusion pro-
tein functions as a native BC10. Unlike dynamin, a peripheral
membrane protein that may be dissolved by high salt (1 M
NaCl) and alkaline pH (pH 11), the BC10–GFP fusion protein
was partially extracted from the microsomal fraction of
transgenic plants using detergent (Triton X-100) (Figure 6b),
indicating that BC10 is an integral membrane protein. Fur-
thermore, immunoblotting analysis revealed that the BC10–
GFP fusion protein was sensitive to proteinase K treatment
only when microsomes were pre-treated with detergent
(Figure 6c), suggesting that the C-terminus of BC10 is lumi-
nal and that BC10 has type II membrane topology.
Most type II membrane proteins are associated with the
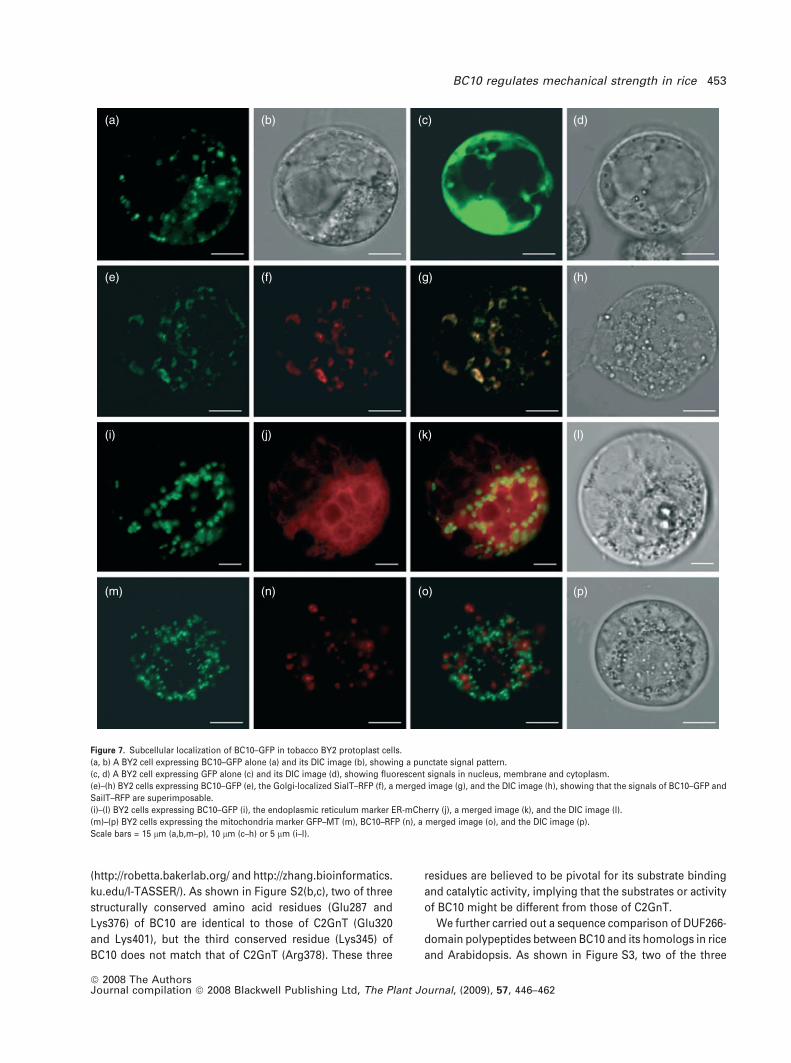
ER and/or Golgi apparatus. To ascertain the actual subcel-
lular location of BC10, we expressed the BC10–GFP fusion in
tobacco Bright Yellow 2 (BY2) protoplasts, and found that
the fluorescent signal of BC10–GFP displayed a punctate
pattern (Figure 7a,b), suggesting that BC10 may be targeted
to certain organelles. Co-localization of BC10 with various
organelle-specific markers in BY2 protoplasts demonstrated
that the punctate pattern of BC10–GFP was identical to that
of the red fluorescent protein (RFP)-tagged Golgi marker
sialyltransferase (SialT) (Figure 7e–h); no such match was
found with any other organelle-specific markers, including
the endoplasmic reticulum marker (Figure 7i–l) and the
mitochondrial marker (Figure 7m–p). Therefore, BC10 is a
Golgi-localized type II membrane protein. In addition, we
also showed that a polypeptide consisting of the first 44
amino acid residues at the N-terminus of BC10 is essential
for its subcellular localization and biological function
(Figure S1).
BC10 may function as a glycosyltransferase
As DUF266 is the only domain identified in BC10, it may be
critical for its function. A BLASTP search of BC10 protein
against the NCBI nr database failed to reveal any homolog
with known functions. However, a second iteration of a PSI-
BLAST search identified b-1,6-N-acetylglucosaminyltrans-
ferase (C2GnT), a glycosyltransferase of animals (Van den
Steen et al., 2000), as a homolog, with an E-value less than
1e-25. Moreover, analyses using the Conserved Domain
Architecture Retrieval Tool (CDART) (Geer et al., 2002) and
the FUGUE server showed that the structure similarity
between BC10 and C2GnT is with high certainty
(ZSCORE = 11.72; a ZSCORE > 6 indicates 99% prediction
confidence). The prediction was further supported by a
secondary structure analysis using the PSIPRED Protein
Structure Prediction Program, showing that the top hits (P
value = 1e-09) were also C2GnT (Figure S2a). We therefore
measured the C2GnT enzyme activity of BC10 by expressing
BC10 in Chinese hamster ovary (CHO) cells, which have an
extremely low background of enzyme activity. The enzyme
activity assay with human C2GnT substrates demonstrated
that the expressed BC10 showed a low but significant GT
activity (Table 1), suggesting that BC10 is a glycosyltrans-
ferase. Compared with the positive control, which showed
much higher enzyme activity, the low enzyme activity of
BC10 is probably due to the lack of authentic acceptor or
donor substrates in the assay system. This result was further
strengthened by an in-depth structural comparison between
the published crystal structure of C2GnT (Pak et al., 2006)
and the predicted three-dimensional structure of BC10
(a)
(b)
(c)
Figure 6. BC10 is a type II membrane protein.
(a) BC10 is a type II membrane protein as predicted by the TMHMM2.0
program, showing a transmembrane helix domain between amino acid
residues 20 and 42, a short N-terminal region located on the cytoplasmic side
of the membrane (inside), and a long C-terminal region containing the
DUF266 domain on the non-cytoplasmic side of the membrane (outside).
(b) Supernatant (S) and pellet (P) fractions of the ultracentrifuged microsomal
fractions from BC10–GFP transgenic plants resuspended in the control buffer,
1 M NaCl, alkaline buffer (pH 11) or 1% Triton X-100. Each fraction was probed
with anti-GFP and dynamin antibodies.
(c) Microsomal fractions from BC10–GFP transgenic plants were incubated in
the presence (+) or absence ()) of Triton X-100 and proteinase K and then
subjected to immunoblot analysis with anti-GFP antibodies. LC, loading
control comprising a Coomassie blue-stained gel of the immunoblot.
452 Yihua Zhou et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462
(http://robetta.bakerlab.org/ and http://zhang.bioinformatics.
ku.edu/I-TASSER/). As shown in Figure S2(b,c), two of three
structurally conserved amino acid residues (Glu287 and
Lys376) of BC10 are identical to those of C2GnT (Glu320
and Lys401), but the third conserved residue (Lys345) of
BC10 does not match that of C2GnT (Arg378). These three
residues are believed to be pivotal for its substrate binding
and catalytic activity, implying that the substrates or activity
of BC10 might be different from those of C2GnT.
We further carried out a sequence comparison of DUF266-
domain polypeptides between BC10 and its homologs in rice
and Arabidopsis. As shown in Figure S3, two of the three
(a) (b) (c) (d)
(e) (f) (g) (h)
(i) (j) (k) (l)
(m) (n) (o) (p)
Figure 7. Subcellular localization of BC10–GFP in tobacco BY2 protoplast cells.
(a, b) A BY2 cell expressing BC10–GFP alone (a) and its DIC image (b), showing a punctate signal pattern.
(c, d) A BY2 cell expressing GFP alone (c) and its DIC image (d), showing fluorescent signals in nucleus, membrane and cytoplasm.
(e)–(h) BY2 cells expressing BC10–GFP (e), the Golgi-localized SialT–RFP (f), a merged image (g), and the DIC image (h), showing that the signals of BC10–GFP and
SailT–RFP are superimposable.
(i)–(l) BY2 cells expressing BC10–GFP (i), the endoplasmic reticulum marker ER-mCherry (j), a merged image (k), and the DIC image (l).
(m)–(p) BY2 cells expressing the mitochondria marker GFP–MT (m), BC10–RFP (n), a merged image (o), and the DIC image (p).
Scale bars = 15 lm (a,b,m–p), 10 lm (c–h) or 5 lm (i–l).
BC10 regulates mechanical strength in rice 453
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462
conserved amino acid residues essential for substrate
binding and enzyme activity of C2GnT are unchanged in
all the members. However, the third conserved residue
showed variation in more than half of the members. These
results suggest that this group of proteins may function as
GTs with different donor or acceptor substrates from those
of human C2GnT.
Cellulose contents of total and fractionated cell walls are
decreased in bc10
The cellular and morphological phenotypes suggested that
the cell-wall composition in the mutant plant may also be
altered. We therefore compared the cellulose and lignin
contents between bc10 and wild-type culms. Compared
with the wild-type, the cellulose content of bc10 was
decreased by approximately 30%, but the lignin content
was increased by approximately 20% in bc10 (Table 2).
These results were further confirmed by quantitative
analysis of the neutral monosaccharide and uronic acid
composition in alcohol-insoluble residues of bc10 and
wild-type culms. As shown in Table 2, glucose content was
reduced by approximately 20% in bc10 culms, but the
contents of xylose, arabinose and galactose were slightly
increased, suggesting that the BC10 mutation has an effect
on cell-wall biosynthesis.
To understand the detailed changes of polysaccharides in
bc10, we sequentially extracted alcohol-insoluble residues
from walls under conditions that allowed individual frac-
tions enriched in a specific type of wall polymer to be
obtained (see Experimental procedures). The sugar compo-
sition of the five fractions, including cellulose, was then
analyzed by GC–MS. As shown in Table 3, individual
monosaccharides in fractions of pectin extracted using
EGTA-Na2CO3 or digested with endopolygalaturonase plus
pectin methylesterase were not significantly altered in bc10.
Similar results were also obtained for the hemicellulose
fraction, although the contents of some sugars, such as
xylose, showed a slightly increase in bc10 extracts obtained
using 1.0 and 4.0 N KOH. We further investigated the
oligosaccharides of xylan and xyloglucan using MALDI-
TOF by digesting them with xylanase and xyloglucanase,
but found no significant differences between them (Fig-
ure S4). However, the glucose content in the cellulose
fraction was significantly reduced in bc10 (Table 3), consis-
tent with the measured cellulose contents (Table 2). Because
BC10 shows similarity to C2GnT, which adds GlcNAc to
glycan, we determined the N-acetyl-glucosamine content in
the above fractions and found no significant reduction,
suggesting that the substrates of BC10 might be different
from those of C2GnT. Therefore, the mutation in BC10
causes a significant decrease in cellulose content. The
increase in lignin (Table 2) and minor alterations in non-
cellulosic sugars (Tables 2 and 3) probably result from
unidentified compensatory regulations responding to cellu-
lose reduction (Li et al., 2003a; Manfield et al., 2004).
Table 1 Assay for C2GnT enzyme activity of BC10 in CHO cells
Transfectants Enzyme activitya
C2GnT cDNA 9625 � 243BC10 cDNA 101 � 14Denatured crude extracts <10Empty vector <10No transfection <10
aThe enzyme activity (pmol mg)1 protein h)1) is the mean of triplicatemeasurements � SE.
Table 2 Comparison of cell-wall components between wild-type and bc10 culms
Sample Glc Xyl Man Gal Ara Rha Uronic acids Cellulose Lignin
Wild-type 395.0 � 6.5 49.2 � 0.7 7.49 � 0.1 14.1 � 1.0 10.3 � 0.2 4.74 � 0.1 1.38 � 0.09 236.2 � 2.5 144.9 � 0.8bc10 316.0 � 6.3 62.8 � 1.0 7.72 � 0.2 16.6 � 0.6 16.7 � 0.3 5.58 � 0.2 1.51 � 0.07 160.6 � 6.7 177.2 � 5.1
The results are means � SE of five independent assays. Each wall component was calculated as mg g)1 of alcohol-insoluble cell-wall residue.
Table 3 Glycosyl composition of fractionated cell walls fromwild-type and bc10 culms
Cell wallfraction Rha Fuc Ara Xyl Man Glc Gal N-GlcA Total
PectinWild-type 0.8 0.0 4.6 7.7 4.2 110.0 7.1 0.8 135.1bc10 0.5 0.0 4.8 7.9 3.9 111.5 7.3 0.7 136.5
EGTA/Na2CO3
Wild-type 0.1 0.0 1.5 0.8 0.7 1.2 2.2 0.2 6.6bc10 0.1 0.0 1.4 0.7 0.7 0.9 2.3 0.3 6.3
1 N KOHWild-type 0.2 0.2 6.2 24.4 0.3 3.7 1.4 0.1 36.5bc10 0.3 0.1 9.6 36.8 0.3 4.0 2.0 0.2 53.2
4 N KOHWild-type 0.1 0.1 1.0 4.1 0.2 4.1 0.4 0.0 9.9bc10 0.1 0.1 1.8 5.8 0.3 3.4 0.8 0.1 12.4
CelluloseWild-type 318.2 318.2bc10 202.5 202.5
TotalWild-type 1.2 0.2 13.3 37.1 5.4 437.1 11.1 1.1 506.3bc10 1.0 0.2 17.5 51.2 5.1 322.2 12.3 1.3 410.8
The results are means of three independent assays (mg g)1 ofalcohol-insoluble residues). The variance is not shown but is <0.2.
454 Yihua Zhou et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462
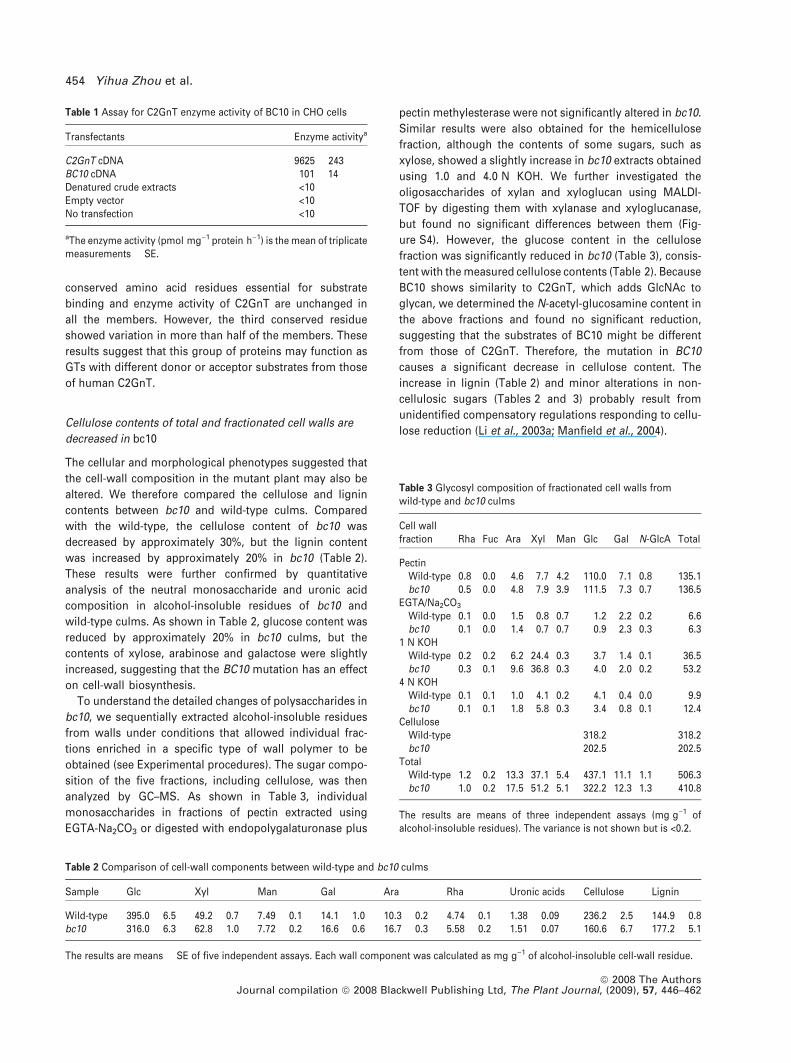
Formation of AGPs is impaired in bc10
Given that the cellulose content is significantly decreased in
bc10 and BC10 is predicted to function as a type II membrane
glycosyltransferase, we therefore analyzed glycoprotein and
polysaccharide contents using seven monoclonal antibodies
(Appendix S1), which enabled us to compared the carbo-
hydrate epitopes of polysaccharides and glycoproteins
between wild-type and mutant plants. As shown in Figure 8,
immunolabeling of culm sections with these antibodies
indicated an apparent decrease in AGP epitopes recognized
by LM2, LM6 and JIM13 in the bc10 plants. AGPs recognized
by LM2 (Figure 8a–d) and JIM13 (Figure 8e–h) were found in
all cells including vascular bundles, sclerenchyma and
parenchyma cells in the wild-type, but were significantly
decreased in mutant plants (Figure 8a–h and Table S2). A
difference in signal intensity when using LM6 was only de-
tected in the vascular bundles and sclerenchyma cells (Fig-
ure 8i–l and Table S2). In contrast, JIM7 equally labeled the
pectin in wild-type and mutant plants (Figure 8m,n and Ta-
ble S2). Staining with b-glucosyl Yariv reagent (bGlcY), a
chemical that is widely used to localize and quantify AGPs,
also revealed a reduction in AGPs in hand-cut stem sections
of the bc10 mutant plant, as indicated by a substantial
diminution in the reddish-brown color (Figure 8o, p), but no
difference was found using b-mancosyl Yariv reagent
(bManY), an inactive analog of bGlcY (data not shown). To
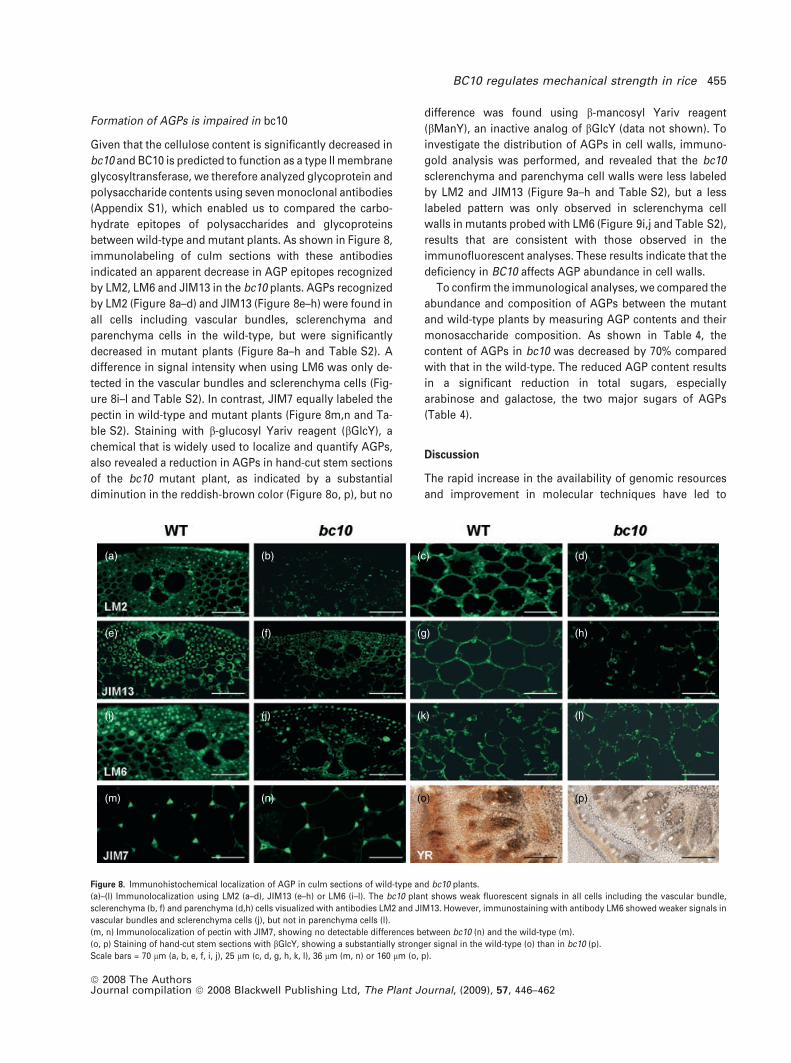
investigate the distribution of AGPs in cell walls, immuno-
gold analysis was performed, and revealed that the bc10
sclerenchyma and parenchyma cell walls were less labeled
by LM2 and JIM13 (Figure 9a–h and Table S2), but a less
labeled pattern was only observed in sclerenchyma cell
walls in mutants probed with LM6 (Figure 9i,j and Table S2),
results that are consistent with those observed in the
immunofluorescent analyses. These results indicate that the
deficiency in BC10 affects AGP abundance in cell walls.
To confirm the immunological analyses, we compared the
abundance and composition of AGPs between the mutant
and wild-type plants by measuring AGP contents and their
monosaccharide composition. As shown in Table 4, the
content of AGPs in bc10 was decreased by 70% compared
with that in the wild-type. The reduced AGP content results
in a significant reduction in total sugars, especially
arabinose and galactose, the two major sugars of AGPs
(Table 4).
Discussion
The rapid increase in the availability of genomic resources
and improvement in molecular techniques have led to
(a) (b) (c) (d)
(e) (f) (g) (h)
(i) (j) (k) (l)
(m) (n) (o) (p)
Figure 8. Immunohistochemical localization of AGP in culm sections of wild-type and bc10 plants.
(a)–(l) Immunolocalization using LM2 (a–d), JIM13 (e–h) or LM6 (i–l). The bc10 plant shows weak fluorescent signals in all cells including the vascular bundle,
sclerenchyma (b, f) and parenchyma (d,h) cells visualized with antibodies LM2 and JIM13. However, immunostaining with antibody LM6 showed weaker signals in
vascular bundles and sclerenchyma cells (j), but not in parenchyma cells (l).
(m, n) Immunolocalization of pectin with JIM7, showing no detectable differences between bc10 (n) and the wild-type (m).
(o, p) Staining of hand-cut stem sections with bGlcY, showing a substantially stronger signal in the wild-type (o) than in bc10 (p).
Scale bars = 70 lm (a, b, e, f, i, j), 25 lm (c, d, g, h, k, l), 36 lm (m, n) or 160 lm (o, p).
BC10 regulates mechanical strength in rice 455
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462
numerous GTs being identified through bioinformatics and
an increasing number being predicted to play roles in cell-
wall metabolism. However, only a small number have been
functionally characterized (Somerville et al., 2004). In this
study, we demonstrate that a loss-of-function mutation of
BC10, a DUF266-containing Golgi-localized protein that
represents a large family of proteins in higher plants and
may function as a glycosyltransferase, disrupts the levels of
cellulose and arabinogalactan proteins in cell walls. These
findings are important for understanding cell-wall biosyn-
thesis in monocotyledonous plants.
BC10 encodes a Golgi-localized DUF266-containing protein
belonging to a superfamily
The plant Golgi apparatus is a critical cytoplasmic organelle
involved in cell-wall polysaccharide and glycoprotein bio-
synthesis and modification (Andreeva et al., 1998). Based
upon bioinformatic analysis, microscopic co-localization
with organ-specific markers and biochemical experiment,
we conclude that BC10 encodes a 399 amino acid long Golgi-
localized type II membrane protein. Identification of the first
44 amino acid residues of BC10 comprising the N-terminal
cytosolic tail (CT) and transmembrane (TM) domains as the
Golgi targeting signal (Figure S1), and characterization of
the mutant’s inferior mechanical properties strongly suggest
that BC10 functions in cell-wall biosynthesis.
Proteins associated with cell-wall biosynthesis and mod-
ification are generally believed to be clustered into families
(a) (b) (c) (d)
(e) (f) (g) (h)
(i) (j) (k) (l)
Figure 9. Immunogold labeling of sclerenchyma and parenchyma cell walls of wild-type and bc10 plants.
(a)–(d) Labeling with the LM2 monoclonal antibody, showing that the bc10 plant has fewer particles in sclerenchyma (b) and parenchyma (d) cell walls.
(e)–(h) Labeling with the JIM13 monoclonal antibody, showing that the bc10 plant has fewer particles in sclerenchyma (f) and parenchyma (h) cell walls.
(i, j) Labeling with the LM6 monoclonal antibody, showing that the bc10 plant has fewer particles only in sclerenchyma (j) cell walls.
(k, l) Labeling with the JIM7 monoclonal antibody, showing equal labeling in wild-type (k) and mutant (l) cell walls.
Scale bars = 500 nm (a–j) or 1 lm (k, l).
Table 4 Comparison of the abundance of AGPs and sugar compo-sition in bc10 and wild-type plants
Samples Wild-type bc10
AGPs 69.8 � 2.6 20.1 � 1.1Total sugar 40.3 � 2.3 17.9 � 1.0Gal 24.0 � 2.3 5.7 � 0.3Ara 14.5 � 1.1 5.5 � 0.3
The AGP content was measured and calculated as lg g)1 fresh weightof tissues. The results are means � SE of three independent assays.
456 Yihua Zhou et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462

(Sarria et al., 2001; Scheible and Pauly, 2004). For example,
the Arabidopsis genome contains a total of nine sequences
with high homology to AtFUT1 (Sarria et al., 2001) and eight
genes with significant similarity to GalT (Edwards et al.,
1999, 2004; Keegstra and Raikhel, 2001). BC10 contains a
DUF266 domain that does not display homology with any
domain of known function. Intensive genomic surveys in the
NCBI and pFam databases indicated that this family of
proteins is plant-specific and ubiquitous in higher plants,
with 18 homologs in rice, 23 in Arabidopsis, and some in
poplar, spruce, grape, etc.
BC10 functions as a glycosyltransferase
As the first identified mutant in the family, bc10 provides an
intriguing opportunity to unravel functions of the BC10
family genetically and biologically. Bioinformatic studies
involving CDART analysis and the prediction of protein
secondary and crystal structures revealed that BC10 shows
some similarities to the animal glycosyltransferase C2GnT,
consistent with a previous prediction that At5g11730 and
At3g21310, two BC10 homologs in Arabidopsis, may func-
tion as GTs related to PF02485 (C2GnT) based on pFam
bioinformatic analysis (Egelund et al., 2004). BC10 shares
two of the three essential conserved amino acid residues
and three of nine conserved Cys residues that are important
for binding substrates and maintaining an active confor-
mation of C2GnT (Pak et al., 2006; Yang et al., 2003). On the
other hand, sequence variations between BC10 and C2GnT
may also reflect their activity differences. This speculation is
substantiated by the low but significant GT enzyme activity
of BC10 in the in vitro assay system. Further studies are
required to understand the biochemical nature of BC10,
especially donor selectivity and acceptor preference, which
has proven to be a real challenge for most GTs in higher
plants (Farrokhi et al., 2006; Keegstra and Raikhel, 2001;
Zhang, 2003).
GTs involved in cell-wall biosynthesis have been grouped
into more than 70 families (http://www.cazy.org) based on
their sequence similarities or sometimes on the existence of
certain motifs, hydrophobic clusters and catalytic specifici-
ties (Rosen et al., 2004). All the BC10 homologs in the current
databases, which share common features including a length
of 340–420 amino acid residues, the presence of a DUF266
domain and prediction as type II membrane proteins, and
should thus be grouped into a separate sub-family or family
of GTs. However, which GT family does BC10 and its
homologs belong to? Based on several lines of experimental
and bioinformatic evidence, we categorize BC10 and its
homologs into family GT14 (http://www.cazy.org). First,
bioinformatic studies, especially the structural comparison
between BC10 and C2GnT using the CDART and PSIPRED
programs, have shown that BC10 is homologous to C2GnT,
which is a GT14 member. Second, the BC10 protein has low
GT enzymatic activity using a C2GnT assay system. Third,
some BC10 homologs, such as At5g11730 and At3g21310,
show a close association with GT14 (Egelund et al., 2004).
However, when BC10, C2GnT and some randomly selected
members from individual GT families were analyzed for their
phylogenetic relationship, BC10 did not cluster in the same
clade as C2GnT and other GT14 proteins (data not shown),
suggesting the possibility that BC10 and its homologs might
form a new GT family. Elucidation of its biochemical nature
will enable this question to be answered.
BC10 regulates mechanical properties by perturbing the
levels of cellulose and AGPs
Based on its subcellular localization and biochemical anal-
ysis, BC10 very probably functions in a manner that leads to
perturbation of the biosynthesis of glycoprotein and/or non-
cellulosic polysaccharides. Therefore, the defect in cellulose
biosynthesis in bc10 revealed by the quantitative analysis of
cell-wall fractions may not be the primary effect that results
in the mutant phenotypes because cellulose is formed at the
plasma membrane involving CESAs in the catalytic subunits
(Delmer, 1999). However, deficiency in formation of glyco-
proteins or the polysaccharide matrix often causes aberrant
cellulose synthesis. For example, an Arabidopsis cyt1
mutant that is defective in mannose-1-phosphate guanylyl-
transferase involved in N-linked glycosylation was found to
have a reduced level of cellulose (Lukowitz et al., 2001).
Xylan synthesis mutants also exhibit reduced secondary
wall deposition and hence reduced cellulose (Brown et al.,
2005, 2007). A likely explanation might be that proteins such
as BC10 play a glycosylation role, modifying CESA subunits
and/or other component(s) of the cellulose synthase
machinery, because CESAs have been predicted to have
several putative N-linked glycosylation sites and mutants of
Arabidopsis with defects in the processing of N-linked gly-
cans are deficient in cellulose synthesis (Gillmor et al., 2002).
Another possibility is that BC10 might play a role in modi-
fying plasma membrane-associated proteins such as
KORRIGAN, COBRA or KOBITO, which have been demon-
strated to be important for cellulose biosynthesis in the
cellulose-deficient mutants kor (His et al., 2001; Molhoj
et al., 2001), cobra (Schindelman et al. 2001; Roudier et al.,
2005) and bc1 (Li et al., 2003b). N-glycosylation of KOR and
COB has been demonstrated to be required for the enzy-
matic activity, which in turn affect cellulose synthesis.
Therefore, the perturbation of cellulose levels in bc10
probably results from malfunction in glycosylation of CESAs
or related proteins.
Monosaccharide analysis of total and fractioned wall
residues revealed no significant alteration in pectin and
hemicellulose in bc10. Therefore, the decrease in the cellu-
lose content in bc10 may result from deficient biosynthesis
of some wall glycoproteins, for example AGPs. It has been
BC10 regulates mechanical strength in rice 457
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462

predicted that AGPs may be secreted to the cell surface in
parallel with cellulose synthesis, and that their carbohydrate
chains bind to cellulose, directly or indirectly regulating the
cellulose synthesis and mechanical properties of cell walls
(Casero et al., 1998; Ito et al., 2005; Loopstra et al., 2000).
Indeed, AGP-like glycoproteins show a tight physical asso-
ciation with flax fiber cellulose (Girault et al., 2000). The
presence of AGP carbohydrate epitopes (Dolan and Roberts,
1995; Stacey et al., 1995) and backbone peptides (Gao and
Showalter, 1999, 2000) in the secondary wall suggest their
crucial roles in the secondary cell-wall thickening. Because
AGPs are often composed of a short protein backbone and
branched carbohydrate chains (Nothnagel, 1997; Seifert and
Roberts, 2007), glycosylation of AGPs is a complicated
process. Therefore, it is plausible that BC10 is responsible
for adding a key moiety to carbohydrate portion of AGPs that
is required for complete synthesis of AGPs. The dramatic
decrease in AGP carbohydrate epitopes in the secondary
walls of bc10 may explain its brittle phenotype, as the lower
level of AGPs in bc10 might directly or indirectly result in a
significant reduction in the cellulose content, which in turn
affects the formation of the secondary cell walls and thus
leads to the brittle phenotype.
Experimental procedures
Plant materials
The bc10 mutant was isolated from a natural population of thejaponica cultivar Huang Jin Qin. A F2 mapping population wasgenerated from a cross between bc10 and Zhefu802 (ZF802), apolymorphic indica cultivar.
Measurements of physical properties
The breaking force and extensibility (measured as extension length)of rice culms and leaves were measured using a digital force/lengthtester (5848 microtester, Istron, http://www.instron.com).
Microscopy
Culms and leaves were fixed in 2.5% v/v glutaraldehyde. Sampleswere post-fixed in 2% w/v OsO4 for 2 h, dehydrated through anethanol gradient, and infiltrated and embedded in butyl methylmethacrylate. Sections 3 lm thick were cut, stained with toluidineblue, and viewed under a microscope. For transmission electronmicroscopy, the samples were embedded using a Suprr kit forelectron microscope (Sigma, http://www.sigmaaldrich.com/), sec-tioned (80 nm ultra-thin) using an Ultracut E ultramicrotome (Leica,http://www.leica.com), and viewed under a Hitachi (http://www.hitachi-hta.com) H7500 transmission electron microscope.
Mapping of BC10
BC10 was mapped and cloned using 870 F2 mutant plants and themolecular markers listed in Table S1. For complementation anal-ysis, an 8.69 kb genomic DNA fragment containing the entire
BC10 coding region, a 1.98 kb upstream sequence and a 2.3 kbdownstream sequence were inserted into the binary vectorpCAMBIA 1300 to generate the transformation plasmid pBC10F. Acontrol plasmid, pBC10T, containing a 3¢ truncated BC10 gene(4.5 kb) was also constructed. The binary plasmids were intro-duced into Agrobacterium tumefaciens strain EHA105 by electro-poration and transformed into bc10 as described previously (Hieiet al., 1994).
Bioinformatics analysis of the BC10 gene
Domain prediction for BC10 was performed using Pfam onlineprediction software (http://pfam.sanger.ac.uk/). Systematic identifi-cation of DUF266-containing proteins in the NCBI nr databases wasperformed using a TimeLogic implemented profile search enginewith default parameters through a CodeQuest work station (http://www.timelogic.com). Sequences with the identified DUF266domain were selected as DUF266-containing proteins. A phyloge-netic tree of Arabidopsis and rice DUF266-containing proteins wasgenerated using MAGE4 (Tamura et al., 2007), with 1000 bootstrapreplications. A PSI-BLAST search for BC10 homologs was per-formed using the NCBI BLAST server (http://blast.ncbi.nlm.nih.gov/Blast.cgi). A structure homology search for BC10 was carried outusing the FUGUE server (http://www-cryst.bioc.cam.ac.uk/fugue/).
Gene expression analysis
Total RNA was extracted from rice plants as described previously(Wadsworth et al., 1988). Complementary DNA (cDNA) was syn-thesized from total RNA using a reverse transcriptional kit fromPromega (http://www.promega.com/). The RT-PCR primers foramplification of BC10, its homologs and actin are given in Table S3,and the reactions were performed on quantitative PCR system7900HT from Applied Biosystems (http://www.appliedbiosystems.com/).
RNA in situ hybridization was performed as described byKao et al. (2002). The 3¢ end of BC10 was sub-cloned into thepGEM-T vector (Promega) and used as a template to generateRNA probes. Hybridization was performed on wax-embeddedtransverse sections (10 lm) using probe labeled with digoxin(Roche, http://www.roche.com).
Subcellular localization
To determine its exact subcellular location, BC10 cDNA was fusedin-frame with either EGFP or mRFP and ligated into the PUC19vector. The expression constructs were co-transfected into tobaccoBY2 protoplasts with mRFP/mCherry- or EGFP-tagged markers forvarious organelles. These markers include the Golgi-localized sial-yltransferase (Lee et al., 2002), and ER and mitochondria localiza-tion constructs containing the ER and mitochondria retentionsequences, respectively (Nelson et al., 2007). The transformedprotoplasts were examined using a confocal microscope (LSM 510META, Zeiss, http://www.zeiss.com/).
C2GnT enzyme activity assay
Human C2GnT and BC10 cDNA clones were inserted into the pEGFP-N1 vector (Clontech, http://www.clontech.com/). The plasmid DNAswere transiently transfected into Chinese hamster ovary (CHO) cellsusing a modified lipofection procedure. The cells were harvested,washed with PBS buffer, and suspended in lysis buffer (10 mM TrisCl,
458 Yihua Zhou et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462

pH 8.0, 0.1 mM EDTA, 5 mM DTT, 0.9% NaCl, 1% Triton X-100). Aftergently shaking at 4�C for at least 2 h, the cell lysate was centrifuged at800 g at 4�C for 5 min. The supernatant was transferred, divided intoaliquots (250 ll each), and stored at )80�C until use. Protein con-centration was determined using a Bio-Rad protein assay kit (http://www.bio-rad.com/) with bovine serum albumin (BSA) as the stan-dard. C2GnT activities were determined as described previously(Bierhuizen and Fukuda, 1992). Briefly, a total of 50 ll reaction mix-ture containing 50 mM HEPES-NaOH, pH 7.0, 1 mM UDP-[glucosa-mine-U-14C] GlcNAc (3.7 kBq, Amersham Pharmacia Biotech, http://www5.amershambiosciences.com/), 1 mM Galb1 fi 3GalNAca1 fip-nitrophenyl (Sigma), 0.1 M GlcNAc, 5 mM DTT and 250 lg lysatewas incubated at 37�C for 2–3 h and the reaction was stopped byadding 5 ml deionized water. The reaction products were purifiedusing a C18 Sep-Pak column (Waters, http://www.waters.com),washed with 20 ml of water, and eluted with methanol. Finally, theproducts were measured using a liquid scintillation counter (1450MicroBeta TriLux; Perkin-Elmer, http://www.perkinelmer.com).
Compositional analysis of neutral glycosyl residues
of whole cell walls
Alcohol-insoluble residues of culms were prepared as described byLi et al. (2003b), and the cellulose content was determined as pre-viously described (Tanaka et al., 2003). A phenol/sulfuric acid assaywas used to quantify hexoses attributed to cellulose content.Sugars, alditol acetates and lignin were analyzed as describedpreviously (Hoebler et al., 1989; Kirk and Obst, 1988).
Fractionation of walls for glycosyl composition analysis and
measurement of uronic acids
Ground material from the second culm internode was fractioned asdescribed by Persson et al. (2007) and Zhong et al. (2005). Uronicacid content was determined as described by Filisetti-Cozzi andCarpita (1991). For details, see Appendix S1.
Chemical and immunological analyses of AGPs
For immunolocalization analyses, see Appendix S1. For histo-chemical staining, fresh hand-cut sections of stems of wild-type andbc10 plants were incubated in bGlcY (Yariv et al., 1962) [2 mg ml)1
(b-D-glucosyl)3 in 0.1 M NaCl] or the control reagent bManY[2 mg ml)1 (b-D-mannosyl)3 in 0.1 M NaCl] for 1 h. The sectionswere then visualized using a fluorescent microscope (Leica).
AGPs were isolated from the aerial organs of wild-type and bc10plants as described previously (Kreuger and van Holst, 1993). Theconcentration of AGPs was determined by the single radial diffusionmethod developed by van Holst and Clarke (1985). Monosaccharidecomposition was determined by GC–MS analysis of alditol acetatesderived from sugars in the hydrolysates of AGPs (van Hengel andRoberts, 2002).
Acknowledgements
We thank Antony Bacic (The School of Botany, University of Mel-bourne, Melbourne) and Vincent L. Chiang (Department of Forestry,North Carolina State University, Raleigh) for critical comments onthe manuscript, Taihua Zhang for measurement of breaking force ofrice plants, Dongqiao Shi for confocal microscope examination,Minoru Fukuda (Cancer Research Center, Burnham Institute, LaJolla, CA) for providing the C2GnT cDNA clone, Inhwan Hwang(Division of Molecular and Life Sciences, Pohang University of
Science and Technology, Pohang, Korea) for providing the Golgimarker gene SailT–RFP, and Xianming Pan (Department of Biolo-gical Sciences and Biotechnology, Tsinghua University) and Yun He(Institute of Biophysics, Chinese Academy of Sciences, Beijing) forprotein structure prediction. Development and distribution ofantibodies JIM7 and JIM13 were supported in part by NationalScience Foundation grants DBI-0421683 and RCN-0090281. Thiswork was supported by grants from the National Natural ScienceFoundation of China (30221002, 30470112 and 30330040).
Supporting Information
Additional Supporting Information may be found in the onlineversion of this article:Figure S1. Subcellular localization of GFP-tagged BC10 andBC10DN44 in onion epidermal and rice root cells.Figure S2. Secondary and three-dimensional structures of the BC10protein.Figure S3. Alignment of BC10 homologs in rice and Arabidopsis.Figure S4. Comparison of MALDI-TOF mass spectra of b-endoxy-lanase- and xyloglucanase-digested products from 1 and 4 N KOHfractions of wild-type and bc10 culms.Table S1. Molecular markers developed for map-based cloning ofBC10.Table S2. Quantification of fluorescent intensity and gold particlesdetected in immunostaining.Table S3. Primers for RT-PCR analysis of BC10 homolog in rice.Appendix S1. Additional experimental procedures and referencesPlease note: Wiley-Blackwell are not responsible for the content orfunctionality of any supporting materials supplied by the authors.Any queries (other than missing material) should be directed to thecorresponding author for the article.
References
Andreeva, A., Kutuzov, M., Evans, D. and Hawes, C. (1998) Thestructure and function of the Golgi apparatus: a hundred years ofquestions. J. Exp. Bot. 49, 1281–1291.
Bacic, A. (2006) Breaking an impasse in pectin biosynthesis. Proc.Natl Acad. Sci. USA, 103, 5639–5640.
Bacic, A., Harris, P.J. and Stone, B.A. (1988) Structure and functionof plant cell walls. In The Biochemistry of Plants (Priess, J., ed.).New York: Academic Press, pp. 297–371.
Bateman, A., Coin, L., Durbin, R. et al. (2004) The Pfam proteinfamilies database. Nucleic Acids Res. 32, 138–141.
Bierhuizen, M.F. and Fukuda, M. (1992) Expression cloning of acDNA encoding UDP-GlcNAc:Galb 1-3-GalNAc-R (GlcNAc toGalNAc) b1-6GlcNAc transferase by gene transfer into CHO cellsexpressing polyoma large tumor antigen. Proc. Natl Acad. Sci.USA, 89, 9326–9330.
Brown, D.M., Zeef, L.A., Ellis, J., Goodacre, R. and Turner, S.R.
(2005) Identification of novel genes in Arabidopsis involved insecondary cell wall formation using expression profiling andreverse genetics. Plant Cell, 17, 2281–2295.
Brown, D.M., Goubet, F., Wong, V.W., Goodacre, R., Stephens, E.,
Dupree, P. and Turner, S.R. (2007) Comparison of five xylansynthesis mutants reveals new insight into the mechanisms ofxylan synthesis. Plant J. 52, 1154–1168.
Burton, R.A., Wilson, S.M., Hrmova, M., Harvey, A.J., Shirley, N.J.,
Medhurst, A., Stone, B.A., Newbigin, E.J., Bacic, A. and Fincher,
G.B. (2006) Cellulose synthase-like CslF genes mediate thesynthesis of cell wall (1,3;1,4)-b-D-glucans. Science, 311, 1940–1942.
BC10 regulates mechanical strength in rice 459
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462

Carpita, N.C. and Gibeaut, D.M. (1993) Structural models of primarycell walls in flowering plants: consistency of molecular structurewith the physical properties of the walls during growth. Plant J. 3,1–30.
Carpita, N., Tierney, M. and Campbell, M. (2001) Molecularbiology of the plant cell wall: searching for the genes thatdefine structure, architecture and dynamics. Plant Mol. Biol. 47,1–5.
Casero, P.J., Casimiro, I. and Knox, J.P. (1998) Occurrence of cellsurface arabinogalactan-protein and extensin epitopes in relationto pericycle and vascular tissue development in the root apex offour species. Planta, 204, 252–259.
Cosgrove, D.J. (2005) Growth of the plant cell wall. Nature Rev. 6,850–861.
Coutinho, P.M., Stam, M., Blanc, E. and Henrissat, B. (2003) Why arethere so many carbohydrate-active enzyme-related genes inplants? Trends Plant Sci. 8, 563–565.
Delmer, D.P. (1999) Cellulose biosynthesis: exciting times for a dif-ficult field of study. Annu. Rev. Plant Physiol. Plant Mol. Biol. 50,245–276.
Dhugga, K.S., Barreiro, R., Whitten, B. et al. (2004) Guar seedb-mannan synthase is a member of the cellulose synthase supergene family. Science, 303, 363–366.
Dolan, L. and Roberts, K. (1995) Secondary thickening in roots ofArabidopsis thaliana – anatomy and cell-surface changes. NewPhytol. 131, 121–128.
Edwards, M.E., Dickson, C.A., Chengappa, S., Sidebottom, C.,
Gidley, M.J. and Reid, J.S. (1999) Molecular characterisation of amembrane-bound galactosyltransferase of plant cell wall matrixpolysaccharide biosynthesis. Plant J. 19, 691–697.
Edwards, M.E., Choo, T.S., Dickson, C.A., Scott, C., Gidley, M.J. and
Reid, J.S. (2004) The seeds of Lotus japonicus lines transformedwith sense, antisense, and sense/antisense galactomannangalactosyltransferase constructs have structurally altered galac-tomannans in their endosperm cell walls. Plant Physiol. 134,1153–1162.
Egelund, J., Skjot, M., Geshi, N., Ulvskov, P. and Petersen, B.L.
(2004) A complementary bioinformatics approach to identify po-tential plant cell wall glycosyltransferase-encoding genes. PlantPhysiol. 136, 2609–2620.
Ellis, C., Karafyllidis, I., Wasternack, C. and Turner, J.G. (2002) TheArabidopsis mutant cev1 links cell wall signaling to jasmonateand ethylene responses. Plant Cell, 14, 1557–1566.
Faik, A., Bar-Peled, M., DeRocher, A.E., Zeng, W., Perrin, R.M.,
Wilkerson, C., Raikhel, N.V. and Keegstra, K. (2000) Biochemicalcharacterization and molecular cloning of an a-1,2-fucosyltrans-ferase that catalyzes the last step of cell wall xyloglucan biosyn-thesis in pea. J. Biol. Chem. 275, 15082–15089.
Farrokhi, N., Burton, R.A., Brownfield, L., Hrmova, M., Wilson,
S.M., Bacic, A. and Fincher, G.B. (2006) Plant cell wall bio-synthesis: genetic, biochemical and functional genomicsapproaches to the identification of key genes. Plant Biotechnol.J. 4, 145–167.
Filisetti-Cozzi, T.M. and Carpita, N.C. (1991) Measurement of uronicacids without interference from neutral sugars. Anal. Biochem.197, 157–162.
Finn, R.D., Mistry, J., Schuster-Bockler, B. et al. (2006) Pfam: clans,web tools and services. Nucleic Acids Res. 34, 247–251.
Fukuda, M. (1996) Possible roles of tumor-associated carbohydrateantigens. Cancer Res. 56, 2237–2244.
Gao, M. and Showalter, A.M. (1999) Yariv reagent treatmentinduces programmed cell death in Arabidopsis cell cultures andimplicates arabinogalactan protein involvement. Plant J. 19, 321–331.
Gao, M. and Showalter, A.M. (2000) Immunolocalization ofLeAGP-1, a modular arabinogalactan-protein, reveals its devel-opmentally regulated expression in tomato. Planta, 210, 865–874.
Gaspar, Y., Johnson, K.L., McKenna, J.A., Bacic, A. and Schultz, C.J.
(2001) The complex structures of arabinogalactan-proteins andthe journey towards understanding function. Plant Mol. Biol. 47,161–176.
Geer, L.Y., Domrachev, M., Lipman, D.J. and Bryant, S.H. (2002)CDART: protein homology by domain architecture. Genome Res.12, 1619–1623.
Geshi, N., Jorgensen, B. and Ulvskov, P. (2004) Subcellular locali-zation and topology of b(1 fi 4)galactosyltransferase that elon-gates b(1 fi 4)galactan side chains in rhamnogalacturonan I inpotato. Planta, 218, 862–868.
Gillmor, C.S., Poindexter, P., Lorieau, J., Palcic, M.M. and Somer-
ville, C. (2002) a-Glucosidase I is required for cellulose biosyn-thesis and morphogenesis in Arabidopsis. J. Cell Biol. 156, 1003–1013.
Girault, R., His, I., Andeme-Onzighi, C., Driouich, A. and Morvan, C.
(2000) Identification and partial characterization of proteins andproteoglycans encrusting the secondary cell walls of flax fibres.Planta, 211, 256–264.
van Hengel, A.J. and Roberts, K. (2002) Fucosylated arabinogalac-tan-proteins are required for full root cell elongation in Arabid-opsis. Plant J. 32, 105–113.
van Hengel, A.J., Tadesse, Z., Immerzeel, P., Schols, H., van
Kammen, A. and de Vries, S.C. (2001) N-acetylglucosamine andglucosamine-containing arabinogalactan proteins control soma-tic embryogenesis. Plant Physiol. 125, 1880–1890.
Henrissat, B. and Davies, G.J. (2000) Glycoside hydrolases andglycosyltransferases: families, modules and implications forgenomics. Plant Physiol. 124, 1515–1519.
Hiei, Y., Ohta, S., Komari, T. and Kumashiro, T. (1994) Efficienttransformation of rice (Oryza sativa L.) mediated by Agrobacte-rium and sequence analysis of the boundaries of the T-DNA. PlantJ. 6, 271–282.
His, I., Driouich, A., Nicol, F., Jauneau, A. and Hofte, H. (2001)Altered pectin composition in primary cell walls of korrigan, adwarf mutant of Arabidopsis deficient in a membrane-boundendo-1,4-b-glucanase. Planta, 212, 348–358.
Hoebler, C., Barry, L.D. and Delort-Laval, J. (1989) Rapid hydrolysisof plant cell wall polysaccharides by gas–liquid chromatography.J. Agric. Food. Chem. 37, 360–367.
van Holst, G.J. and Clarke, A.E. (1985) Quantification of arabino-galactan-protein in plant extracts by single radial gel diffusion.Anal. Biochem. 148, 446–450.
Ito, S., Suzuki, Y., Miyamoto, K., Ueda, J. and Yamaguchi, I. (2005)AtFLA11, a fasciclin-like arabinogalactan-protein, specificallylocalized in sclerenchyma cells. Biosci. Biotechnol. Biochem. 69,1963–1969.
Kao, Y.Y., Harding, S.A. and Tsai, C.J. (2002) Differential expressionof two distinct phenylalanine ammonia-lyase genes in condensedtannin-accumulating and lignifying cells of quaking aspen. PlantPhysiol. 130, 796–807.
Keegstra, K. and Raikhel, N. (2001) Plant glycosyltransferases. Curr.Opin. Plant Biol. 4, 219–224.
Kirk, T.K. and Obst, J.R. (1988) Lignin determination. MethodsEnzymol. 161, 87–101.
Kreuger, M. and van Holst, G.J. (1993) Arabinogalactan proteins areessential in somatic embryogenesis of Daucus carota L. Planta,189, 243–248.
Lee, M.H., Min, M.K., Lee, Y.J., Jin, J.B., Shin, D.H., Kim, D.H., Lee,
K.H. and Hwang, I. (2002) ADP-ribosylation factor 1 of Arabidopsisplays a critical role in intracellular trafficking and maintenance of
460 Yihua Zhou et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462

endoplasmic reticulum morphology in Arabidopsis. Plant Phys-iol. 129, 1507–1520.
Lerouxel, O., Cavalier, D.M., Liepman, A.H. and Keegstra, K. (2006)Biosynthesis of plant cell wall polysaccharides – a complex pro-cess. Curr. Opin. Plant Biol. 9, 621–630.
Li, L., Zhou, Y., Cheng, X., Sun, J., Marita, J.M., Ralph, J. and
Chiang, V.L. (2003a) Combinatorial modification of multiple lignintraits in trees through multigene cotransformation. Proc. NatlAcad. Sci. USA, 100, 4939–4944.
Li, Y., Qian, Q., Zhou, Y. et al. (2003b) BRITTLE CULM1, whichencodes a COBRA-like protein, affects the mechanical propertiesof rice plants. Plant Cell, 15, 2020–2031.
Loopstra, C.A., Puryear, J.D. and No, E.G. (2000) Purification andcloning of an arabinogalactan-protein from xylem of loblollypine. Planta, 210, 686–689.
Lukowitz, W., Nickle, T.C., Meinke, D.W., Last, R.L., Conklin, P.L.
and Somerville, C.R. (2001) Arabidopsis cyt1 mutants are defi-cient in a mannose-1-phosphate guanylyltransferase and point toa requirement of N-linked glycosylation for cellulose biosynthe-sis. Proc. Natl Acad. Sci. USA, 98, 2262–2267.
Lytovchenko, A., Sonnewald, U. and Fernie, A.R. (2007) The com-plex network of non-cellulosic carbohydrate metabolism. Curr.Opin. Plant Biol. 10, 227–235.
Madson, M., Dunand, C., Li, X., Verma, R., Vanzin, G.F., Caplan, J.,
Shoue, D.A., Carpita, N.C. and Reiter, W.D. (2003) The MUR3 geneof Arabidopsis encodes a xyloglucan galactosyltransferase that isevolutionarily related to animal exostosins. Plant Cell, 15, 1662–1670.
Manfield, I.W., Orfila, C., McCartney, L., Harholt, J., Bernal, A.J.,
Scheller, H.V., Gilmartin, P.M., Mikkelsen, J.D., Paul Knox, J.
and Willats, W.G. (2004) Novel cell wall architecture ofisoxaben-habituated Arabidopsis suspension-cultured cells:global transcript profiling and cellular analysis. Plant J. 40, 260–275.
Molhoj, M., Ulvskov, P. and Dal Degan, F. (2001) Characterization ofa functional soluble form of a Brassica napus membrane-anchored endo-1,4-b-glucanase heterologously expressed inPichia pastoris. Plant Physiol. 127, 674–684.
Nelson, B.K., Cai, X. and Nebenfuhr, A. (2007) A multicolored set ofin vivo organelle markers for co-localization studies in Arabid-opsis and other plants. Plant J. 51, 1126–1136.
Nothnagel, E.A. (1997) Proteoglycans and related components inplant cells. Int. Rev. Cytol. 174, 195–291.
Pagny, S., Bouissonnie, F., Sarkar, M., Follet-Gueye, M.L., Driouich,
A., Schachter, H., Faye, L. and Gomord, V. (2003) Structuralrequirements for Arabidopsis b1,2-xylosyltransferase activity andtargeting to the Golgi. Plant J. 33, 189–203.
Pak, J.E., Arnoux, P., Zhou, S., Sivarajah, P., Satkunarajah, M., Xing,
X. and Rini, J.M. (2006) X-ray crystal structure of leukocyte typecore 2 b1,6-N-acetylglucosaminyltransferase. Evidence for aconvergence of metal ion-independent glycosyltransferasemechanism. J. Biol. Chem. 281, 26693–26701.
Pena, M.J., Zhong, R., Zhou, G.K., Richardson, E.A., O’Neill, M.A.,
Darvill, A.G., York, W.S. and Ye, Z.H. (2007) Arabidopsis irregularxylem8 and irregular xylem9: implications for the complexity ofglucuronoxylan biosynthesis. Plant Cell, 19, 549–563.
Persson, S., Caffall, K.H., Freshour, G., Hilley, M.T., Bauer, S.,
Poindexter, P., Hahn, M.G., Mohnen, D. and Somerville, C. (2007)The Arabidopsis irregular xylem8 mutant is deficient in glucu-ronoxylan and homogalacturonan, which are essential for sec-ondary cell wall integrity. Plant Cell, 19, 237–255.
Reiter, W.D., Chapple, C. and Somerville, C.R. (1997) Mutants ofArabidopsis thaliana with altered cell wall polysaccharide com-position. Plant J. 12, 335–345.
Rosen, M.L., Edman, M., Sjostrom, M. and Wieslander, A. (2004)Recognition of fold and sugar linkage for glycosyltransferasesby multivariate sequence analysis. J. Biol. Chem. 279, 38683–38692.
Ross, J., Li, Y., Lim, E. and Bowles, D.J. (2001) Higher plant glyco-syltransferases. Genome Biol. 2, reviews, 3004.1–3004.6.
Roudier, F., Fernandez, A.G., Fujita, M. et al. (2005) COBRA,an Arabidopsis extracellular glycosyl-phosphatidyl inositol-anchored protein, specifically controls highly anisotropic expan-sion through its involvement in cellulose microfibril orientation.Plant Cell, 17, 1749–1763.
Sarria, R., Wagner, T.A., O’Neill, M.A., Faik, A., Wilkerson, C.G.,
Keegstra, K. and Raikhel, N.V. (2001) Characterization of a familyof Arabidopsis genes related to xyloglucan fucosyltransferase I.Plant Physiol. 127, 1595–1606.
Scheible, W.R. and Pauly, M. (2004) Glycosyltransferases and cellwall biosynthesis: novel players and insights. Curr. Opin. PlantBiol. 7, 285–295.
Schindelman, G., Morikami, A., Jung, J., Baskin, T.I., Carpita, N.C.,
Derbyshire, P., McCann, M.C. and Benfry, P.N. (2001) COBRAencodes a putative GPI-anchored protein, which is polarly loca-lized and necessary for oriented cell expansion in Arabidopsis.Genes Dev. 15, 1115–1127.
Seifert, G.J. and Roberts, K. (2007) The biology of arabinogalactanproteins. Annu. Rev. Plant Biol. 58, 137–161.
Somerville, C. (2006) Cellulose synthesis in higher plants. Annu.Rev. Cell Dev. Biol. 22, 53–78.
Somerville, C., Bauer, S., Brininstool, G. et al. (2004) Toward asystems approach to understanding plant cell walls. Science, 306,2206–2211.
Stacey, N.J., Roberts, K., Carpita, N.C., Wells, B. and McCann, M.C.
(1995) Dynamic changes in cell surface molecules are very earlyevents in the differentiation of mesophyll cells from Zinnia ele-gans into tracheary elements. Plant J. 8, 891–906.
Takeda, T. and Fry, S.C. (2004) Control of xyloglucan endotrans-glucosylase activity by salts and anionic polymers. Planta, 219,722–732.
Tamura, K., Dudley, J., Nei, M. and Kumar, S. (2007) MEGA4:Molecular Evolutionary Genetics Analysis (MEGA) softwareversion 4.0. Mol. Biol. Evol. 24, 1598–1599.
Tanaka, K., Murata, K., Yamazaki, M., Onosato, K., Miyao, A. and
Hirochika, H. (2003) Three distinct rice cellulose synthase catalyticsubunit genes required for cellulose synthesis in the secondarywall. Plant Physiol. 133, 73–83.
Taylor, N.G., Howells, R.M., Huttly, A.K., Vickers, K. and Turner, S.R.
(2003) Interactions among three distinct CesA proteins essentialfor cellulose synthesis. Proc. Natl Acad. Sci. USA, 100, 1450–1455.
Van den Steen, P.E., Rudd, P.M., Wormald, M.R., Dwek, R.A. and
Opdenakker, G. (2000) O-linked glycosylation in focus. TrendsGlycosci. Glycotechnol. 12, 35–49.
Vogel, J.P., Raab, T.K., Somerville, C.R. and Somerville, S.C. (2004)Mutations in PMR5 result in powdery mildew resistance andaltered cell wall composition. Plant J. 40, 968–978.
Wadsworth, G.J., Redinbaugh, M.G. and Scandalios, J.G. (1988) Aprocedure for the small-scale isolation of plant RNA suitable forRNA blot analysis. Anal. Biochem. 172, 279–283.
Wenderoth, I. and von Schaewen, A. (2000) Isolation and charac-terization of plant N-acetyl glucosaminyltransferase I (GntI) cDNAsequences. Functional analyses in the Arabidopsis cgl mutantand in antisense plants. Plant Physiol. 123, 1097–1108.
Wu, H.M., Wong, E., Ogdahl, J. and Cheung, A.Y. (2000) Apollen tube growth-promoting arabinogalactan protein fromNicotiana alata is similar to the tobacco TTS protein. Plant J. 22,165–176.
BC10 regulates mechanical strength in rice 461
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462

Yang, X., Qin, W., Lehotay, M., Toki, D., Dennis, P., Schutzbach, J.S.
and Brockhausen, I. (2003) Soluble human core 2 b6-Nacetyl-glucosaminyltransferase C2GnT1 requires its conservedcysteine residues for full activity. Biochem. Biophys. Acta, 1648,62–74.
Yariv, J., Rapport, M.M. and Graf, L. (1962) The interaction ofglycosides and saccharides with antibody to the correspondingphenylazo glycosides. Biochem. J. 85, 383–388.
Zhang, J.Z. (2003) Overexpression analysis of plant transcriptionfactors. Curr. Opin. Plant Biol. 6, 430–440.
Zhong, R. and Ye, Z.H. (2007) Regulation of cell wall biosynthesis.Curr. Opin. Plant Biol. 10, 564–572.
Zhong, R., Burk, D.H., Morrison, W.H. III and Ye, Z.H. (2002) A kinesin-like protein is essential for oriented deposition of cellulosemicrofibrils and cell wall strength. Plant Cell, 14, 3101–3117.
Zhong, R., Pena, M.J., Zhou, G.K., Nairn, C.J., Wood-Jones, A.,
Richardson, E.A., Morrison, W.H. III, Darvill, A.G., York, W.S. and
Ye, Z.H. (2005) Arabidopsis fragile fiber8, which encodes a puta-tive glucuronyltransferase, is essential for normal secondary wallsynthesis. Plant Cell, 17, 3390–3408.
The GenBank/EMBL accession number for the BC10 sequence is EF140884.
462 Yihua Zhou et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2009), 57, 446–462
Copyright © 2022 FDOKUMEN




















