
A Reversibly Autoglycosylated Arabidopsis Protein Implicated in Cell Wall Biosynthesis
11
Cloning and Characterization of AtRGP1 1 A Reversibly Autoglycosylated Arabidopsis Protein Implicated in Cell Wall Biosynthesis Ivan J. Delgado, Zhaohong Wang, Amy de Rocher, Kenneth Keegstra, and Natasha V. Raikhel* Michigan State University, Department of Energy Plant Research Laboratory, Michigan State University, East Lansing, Michigan 48824–1312 A reversibly glycosylated polypeptide from pea (Pisum sativum) is thought to have a role in the biosynthesis of hemicellulosic polysac- charides. We have investigated this hypothesis by isolating a cDNA clone encoding a homolog of Arabidopsis thaliana, Reversibly Gly- cosylated Polypeptide-1 (AtRGP1), and preparing antibodies against the protein encoded by this gene. Polyclonal antibodies detect homologs in both dicot and monocot species. The patterns of ex- pression and intracellular localization of the protein were exam- ined. AtRGP1 protein and RNA concentration are highest in roots and suspension-cultured cells. Localization of the protein shows it to be mostly soluble but also peripherally associated with mem- branes. We confirmed that AtRGP1 produced in Escherichia coli could be reversibly glycosylated using UDP-glucose and UDP- galactose as substrates. Possible sites for UDP-sugar binding and glycosylation are discussed. Our results are consistent with a role for this reversibly glycosylated polypeptide in cell wall biosynthesis, although its precise role is still unknown. The primary cell wall of dicot plants is laid down by young cells prior to the cessation of elongation and second- ary wall deposition. Making up to 90% of the cell’s dry weight, the extracellular matrix is important for many pro- cesses, including morphogenesis, growth, disease resis- tance, recognition, signaling, digestibility, nutrition, and decay. The composition of the cell wall has been exten- sively described (Bacic et al., 1988; Levy and Staehelin, 1992; Zablackis et al., 1995), and yet many questions remain unanswered regarding the synthesis and interaction of these components to provide cells with a functional wall (Carpita and Gibeaut, 1993; Carpita et al., 1996). Heteropolysaccharide biosynthesis can be divided into four steps: (a) chain or backbone initiation, (b) elongation, (c) side-chain addition, and (d) termination and extracellu- lar deposition (Waldron and Brett, 1985). The similarity between various polysaccharide backbones leads to the prediction that the synthesizing machinery would be con- served between them. For example, the backbone of xylo- glucan polymers, b-1,4 glucan, can be synthesized inde- pendently of or concurrently with side-chain addition (Campbell et al., 1988; White et al., 1993), and this polymer and the chains that make up cellulose are identical. The later addition of side chains to xyloglucan are catalyzed by specific transferases (Kleene and Berger, 1993) such as xy- losyltransferase (Campbell et al., 1988), galactosyltrans- ferase, and fucosyltransferase (Faı ¨k et al., 1997), all of which are localized to the Golgi compartment (Brummell et al., 1990; Driouich et al., 1993; Staehelin and Moore, 1995). The enzymes involved in wall biosynthesis have been recalcitrant to isolation (Carpita et al., 1996; Albersheim et al., 1997). Only recently has the first gene encoding puta- tive cellulose biosynthetic enzymes, celA, been isolated from cotton (Gossypium hirsutum) and rice (Oryza sativa; Pear et al., 1996). During studies of polysaccharide synthesis in pea (Pisum sativum) Golgi membranes, Dhugga et al. (1991) identified a 41-kD protein doublet that they suggested was involved in polysaccharide synthesis. The authors showed that this protein could be glycosylated by radiolabeled UDP-Glc but that this labeling could be reversibly competed with by unlabeled UDP-Glc, UDP-Xyl, and UDP-Gal, the sugars that make up xyloglucan (Hayashi, 1989). The 41-kD pro- tein was named PsRGP1 (P. sativum Reversibly Glycosy- lated Polypeptide-1; Dhugga et al., 1997). Furthermore, the conditions that stimulate or inhibit Golgi-localized b-glucan synthase activity are the same conditions that stimulate or inhibit the glycosylation of PsRGP1 (Dhugga et al., 1991). To address the role of this protein in polysac- charide synthesis, the authors purified the polypeptides and obtained the sequences from tryptic peptides (Dhugga and Ray, 1994). Antibodies raised against PsRGP1 showed that it is soluble and localized to the plasma membrane (Dhugga et al., 1991) and Golgi compartment (Dhugga et al., 1997). In addition to its Golgi localization, the steady- state glycosylation of PsRGP1 is approximately 10:7:3 (UDP-Glc:-Xyl:-Gal), which is similar to the typical sugar composition of xyloglucan (1.0:0.75:0.25; Dhugga et al., 1997). We were interested in studying various aspects of cell wall metabolism, including the synthesis of polysacchar- ides and their delivery to the cell wall. Studies in pea have shown that a 41-kD protein may be involved in cell wall polysaccharide synthesis, possibly that of xyloglucan (Dhugga et al., 1997). Here we report the characterization 1 This research was supported by a grant from the U.S. Depart- ment of Energy (no. De-FG02-91ER20021). * Corresponding author; e-mail [email protected]; fax 1–517–353–9168. Abbreviations: EST, expressed sequence tag; GST, glutathione S-transferase; RGP, reversibly glycosylated polypeptide. Plant Physiol. (1998) 116: 1339–1349 1339 www.plant.org on January 31, 2014 - Published by www.plantphysiol.org Downloaded from Copyright © 1998 American Society of Plant Biologists. All rights reserved.
Transcript of A Reversibly Autoglycosylated Arabidopsis Protein Implicated in Cell Wall Biosynthesis

Cloning and Characterization of AtRGP11
A Reversibly Autoglycosylated Arabidopsis Protein Implicated in Cell Wall Biosynthesis
Ivan J. Delgado, Zhaohong Wang, Amy de Rocher, Kenneth Keegstra, and Natasha V. Raikhel*
Michigan State University, Department of Energy Plant Research Laboratory, Michigan State University,East Lansing, Michigan 48824–1312
A reversibly glycosylated polypeptide from pea (Pisum sativum) isthought to have a role in the biosynthesis of hemicellulosic polysac-charides. We have investigated this hypothesis by isolating a cDNAclone encoding a homolog of Arabidopsis thaliana, Reversibly Gly-cosylated Polypeptide-1 (AtRGP1), and preparing antibodies againstthe protein encoded by this gene. Polyclonal antibodies detecthomologs in both dicot and monocot species. The patterns of ex-pression and intracellular localization of the protein were exam-ined. AtRGP1 protein and RNA concentration are highest in rootsand suspension-cultured cells. Localization of the protein shows itto be mostly soluble but also peripherally associated with mem-branes. We confirmed that AtRGP1 produced in Escherichia colicould be reversibly glycosylated using UDP-glucose and UDP-galactose as substrates. Possible sites for UDP-sugar binding andglycosylation are discussed. Our results are consistent with a rolefor this reversibly glycosylated polypeptide in cell wall biosynthesis,although its precise role is still unknown.
The primary cell wall of dicot plants is laid down byyoung cells prior to the cessation of elongation and second-ary wall deposition. Making up to 90% of the cell’s dryweight, the extracellular matrix is important for many pro-cesses, including morphogenesis, growth, disease resis-tance, recognition, signaling, digestibility, nutrition, anddecay. The composition of the cell wall has been exten-sively described (Bacic et al., 1988; Levy and Staehelin,1992; Zablackis et al., 1995), and yet many questions remainunanswered regarding the synthesis and interaction ofthese components to provide cells with a functional wall(Carpita and Gibeaut, 1993; Carpita et al., 1996).
Heteropolysaccharide biosynthesis can be divided intofour steps: (a) chain or backbone initiation, (b) elongation,(c) side-chain addition, and (d) termination and extracellu-lar deposition (Waldron and Brett, 1985). The similaritybetween various polysaccharide backbones leads to theprediction that the synthesizing machinery would be con-served between them. For example, the backbone of xylo-glucan polymers, b-1,4 glucan, can be synthesized inde-pendently of or concurrently with side-chain addition(Campbell et al., 1988; White et al., 1993), and this polymer
and the chains that make up cellulose are identical. Thelater addition of side chains to xyloglucan are catalyzed byspecific transferases (Kleene and Berger, 1993) such as xy-losyltransferase (Campbell et al., 1988), galactosyltrans-ferase, and fucosyltransferase (Faık et al., 1997), all ofwhich are localized to the Golgi compartment (Brummell etal., 1990; Driouich et al., 1993; Staehelin and Moore, 1995).
The enzymes involved in wall biosynthesis have beenrecalcitrant to isolation (Carpita et al., 1996; Albersheim etal., 1997). Only recently has the first gene encoding puta-tive cellulose biosynthetic enzymes, celA, been isolatedfrom cotton (Gossypium hirsutum) and rice (Oryza sativa;Pear et al., 1996).
During studies of polysaccharide synthesis in pea (Pisumsativum) Golgi membranes, Dhugga et al. (1991) identifieda 41-kD protein doublet that they suggested was involvedin polysaccharide synthesis. The authors showed that thisprotein could be glycosylated by radiolabeled UDP-Glc butthat this labeling could be reversibly competed with byunlabeled UDP-Glc, UDP-Xyl, and UDP-Gal, the sugarsthat make up xyloglucan (Hayashi, 1989). The 41-kD pro-tein was named PsRGP1 (P. sativum Reversibly Glycosy-lated Polypeptide-1; Dhugga et al., 1997). Furthermore,the conditions that stimulate or inhibit Golgi-localizedb-glucan synthase activity are the same conditions thatstimulate or inhibit the glycosylation of PsRGP1 (Dhuggaet al., 1991). To address the role of this protein in polysac-charide synthesis, the authors purified the polypeptidesand obtained the sequences from tryptic peptides (Dhuggaand Ray, 1994). Antibodies raised against PsRGP1 showedthat it is soluble and localized to the plasma membrane(Dhugga et al., 1991) and Golgi compartment (Dhugga etal., 1997). In addition to its Golgi localization, the steady-state glycosylation of PsRGP1 is approximately 10:7:3(UDP-Glc:-Xyl:-Gal), which is similar to the typical sugarcomposition of xyloglucan (1.0:0.75:0.25; Dhugga et al.,1997).
We were interested in studying various aspects of cellwall metabolism, including the synthesis of polysacchar-ides and their delivery to the cell wall. Studies in pea haveshown that a 41-kD protein may be involved in cell wallpolysaccharide synthesis, possibly that of xyloglucan(Dhugga et al., 1997). Here we report the characterization1 This research was supported by a grant from the U.S. Depart-
ment of Energy (no. De-FG02-91ER20021).* Corresponding author; e-mail [email protected]; fax
1–517–353–9168.Abbreviations: EST, expressed sequence tag; GST, glutathione
S-transferase; RGP, reversibly glycosylated polypeptide.
Plant Physiol. (1998) 116: 1339–1349
1339 www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 1998 American Society of Plant Biologists. All rights reserved.

of AtRGP1 (Arabidopsis thaliana Reversibly GlycosylatedPolypeptide-1), a soluble protein that can also be foundweakly associated with membrane fractions, most likelythe Golgi fraction. The reversible nature of the glycosyla-tion of this Arabidopsis homolog by the substrates used tomake polysaccharides (nucleotide sugars) suggests a pos-sible role for AtRGP1 in polysaccharide biosynthesis.
MATERIALS AND METHODS
Plant and Cell Growth
Arabidopsis thaliana wild-type plants (Columbia ecotype)were grown in soil at 22°C and 80% humidity and with 16 hof light. To obtain large amounts of soil-free root tissue,seeds were germinated in liquid culture medium (1 seed100 mL21, 10 g L21 Suc, 4.3 g L21 Murashige-Skoog salts[GIBCO-BRL], 0.5 g L21 Mes, 0.1 g L21 myo-inositol, 1 mgL21 thiamine-HCl, and 0.5 mg L21 nicotinic acid, pH 5.7).Plants grown in liquid culture and maintained under con-stant agitation (50–60 rpm) and light grew mostly roots.
A. thaliana cell-suspension cultures were obtained fromAxelos et al. (1992) and were maintained by diluting a2-week-old culture 1:5 with fresh suspension-culture me-dium (3.2 g L21 Gamborg’s B5 medium [Sigma], 20 g L21
Suc, 2.5 mm 2,4-D, and 0.5 g L21 Mes, pH 5.7).
Cloning, Sequencing, and Sequence Analysis
Peptide sequences from purified pea (Pisum sativum)proteins (Dhugga and Ray, 1994; Fig. 1) were used tosearch the EST database (dBEST) comprising the GenBank,EMBL, and DDBJ databases. We used a database searchprogram (Basic BLAST) with the “tblastn” option, whichcompares a protein query sequence against a nucleotidesequence database dynamically translated in all readingframes (Altschul et al., 1990, http://www.ncbi.nlm.nih-.gov/BLAST/). Twenty-eight ESTs have been obtainedso far from the original search and subsequent searches(Altschul et al., 1997) using the ESTs themselves to findoverlapping ESTs. These ESTs can be grouped based onsequence similarity using a software package (Meg-Align module, DNAStar, Genetics Computer Group, Mad-ison, WI).
Based on sequence similarity, the following ESTs havebeen assigned: eight to group I (AtRGP1; T20512, T46245,T42672, T22507, T22943, H37657, N37306, and T44971),five to group II (AtRGP2; T23020, T46745, AA597661,AA650721, and AA650701), and three to group III (AtRGP3;ATTS4214, R30021, and R90614). The rest (ATTS3942,T44394, T45672, ATTS0381, H76915, N65402, N65528,N65622, AA042694, AA650802, AA651536, and AA597661)were too short to be unambiguously assigned to a specificgroup. Furthermore, similar genomic sequences to AtRGP1have thus far been identified in only two Arabidopsischromosomes. A 129-nucleotide stretch at the end of abacteria artificial chromosome clone (T24B2-Sp6) of chro-mosome I (http://cbil.humgen.upenn.edu/approximatelyatgc/SPP.html) is 96% identical to AtRGP1, and a 113-nucleotide stretch at the end of a bacteria artificial chromo-
some clone (T20M8-Sp6) of chromosome II (http://www.tigr.org/tdb/at/atgenome/atgenome.html) is 96% iden-tical to AtRGP1. Full-length cDNA clones (T20512 andT46745) were obtained from the PRL2 Arabidopsis cDNALibrary (Newman et al., 1994) kept by the ArabidopsisBiological Research Center at Ohio State University (Co-lumbus). Automated sequencing of the cDNAs was per-formed by the Plant Biochemistry Facility at Michigan StateUniversity (East Lansing).
BLAST searches of the GenBank database using thefull-length AtRGP1 cDNA also detected 19 rice ESTs(RICC1486A, RICC2546A, RICR0440A, RICR1510A, RICR-2980A, RICS1545A, RICS1750A, RICS1848A, RICS2305A,RICS5091A, C28170, C73320, C27450, C26584, C72510,C26475, C25974, and C71661), four of which (C72510,C26584, RICS5091A, and C27450) can be fused to give afull-length clone (Oryza sativa, OsRGP). Other RGPs iden-tified in this search include those from P. sativum (PsRGP1;U31565), maize (Zea mays, ZmRGP; U89897), and cowpea(Vigna unguiculata, VuRGP; AF005279).
Hydropathy analysis of AtRGP1 was performed usingthe DNAStar package, which uses the method by Kyte andDoolittle (1982). Protein-sequence analysis of AtRGP1 wasalso performed with this software package. In addition, thepSORT program (http://psort.nibb.ac.jp/) was used,which compares a given protein sequence against a data-base of known sequences that mediate protein sorting tovarious membranes and organelles.
Isolation of RNA and Northern Analysis
Total RNA from Arabidopsis flowers, leaves, roots, andstems was extracted as described previously (Puissant andHoudebine, 1990). Total Arabidopsis RNA (30 mg/lane)was separated on a gel containing 1% agarose and 2%formaldehyde. Electrophoresis was stopped when the dyefront had migrated three-quarters of the way down the gel.Sizes were estimated using the sizes of rRNAs as markers.Gels were washed for 20 min in 103 SSC and analyzed asdescribed by Sambrook et al. (1992). Transfer was per-formed by capillary action onto a nylon membrane (Hy-bond N, Amersham) overnight. Membranes were washedin 23 SSC for 5 min, cross-linked in a UV-cross-linker, andstored at room temperature between filter papers. Hybrid-ization was overnight at 65°C in northern hybridizationbuffer (53 SSC, 103 Denhardt’s solution [2 mg mL21 Fi-coll, 2 mg mL21 PVP, 2 mg mL21 BSA], 0.1 m KPO4, pH 6.8,100 mg/mL salmon-sperm DNA [freshly boiled], 10% dex-tran, and 30% deionized formamide) using a random-primed 32P-labeled 1.0-kb EcoRI-BbvI fragment of theAtRGP1 cDNA at 1,000,000 dpm mL21. Membranes werewashed once with 23 SSC and 0.5% SDS, twice with 23SSC and 0.1% SDS, and once with 0.23 SSC and 0.1% SDS(13 SSC is 0.15 m NaCl and 15 mm sodium nitrate, pH 7.0),each for 30 min at 65°C prior to autoradiographic exposure.
Protoplast Preparation
Twenty grams of cells from a 5- to 8-d-old cell-suspensionculture collected on an 80-mm filter was incubated with 50
1340 Delgado et al. Plant Physiol. Vol. 116, 1998
www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 1998 American Society of Plant Biologists. All rights reserved.

mL of freshly made protoplasting solution (15.4% [w/v] Suc,0.32% [w/v] Gamborg’s B5 minimal organic, 5 mg mL21
cellulase [Onozuka R10, Yakutt Honsha, Tokyo], and 1.2 mgmL21 Macerozyme R10 [Yakutt Honsha]) for 2 h in a rotaryshaker at 80 rpm. All manipulations were done at roomtemperature. The cells were then poured into Babcock cen-trifuge bottles and centrifuged for 10 min at 1100 rpm in aclinical centrifuge swinging bucket rotor (model HNSII, IEC,Needham Heights, MA). Floated protoplasts were collected,mixed with 20 mL of 0.4 m betaine, 3 mm Mes, and 10 mmCaCl2, pH 5.7, and then pelleted for 5 min at 50g.
Protein Extraction
Four milliliters of cold lysis buffer (20 mm Hepes-KOH,pH 7.0, 13.5% [w/v] Suc, 10 mm potassium acetate, 1 mmDDT, 0.5 mm PMSF, and 1 mm EDTA) was added permilliliter of packed protoplasts and passed through a 25–5/8-gauge needle at 4°C until no unbroken protoplastscould be detected under the microscope. Cell debris werepelleted by centrifugation at 500g for 5 min and this ho-mogenate was called the total protoplast protein. Totalplant protein from different tissues (flowers, leaves, roots,stems, and roots grown in liquid culture) was extracted bygrinding 1 to 5 g of the given tissue in liquid N2 and mixingwith 5 mL of lysis buffer at 4°C. For all plant tissue sam-ples, cell debris were pelleted by centrifugation at 500g for5 min. This homogenate was called the total tissue protein,where “tissue” specifies the tissue used.
Overexpression of a Fusion Protein in Escherichia coli andAntibody Preparation
The 1.4-kb EcoRI-NotI fragment of AtRGP1 (GenBankaccession no. AF013627), encoding all but the first nineamino acids from the 59 end of the cDNA, was cloned intothe EcoRI-NotI sites of pGEX-5X-2 (Pharmacia) to generatean N-terminal in-frame fusion with the 26-kD domain ofGST. This fusion was overexpressed in E. coli (strain DH5a)by growing cells in Luria-Bertani medium at 37°C to anA600 of 0.7 to 0.8, then adding isopropylthio-b-d-gal-actosidase to a final concentration of 0.2 mm, and incubat-ing the culture at 28°C for 4 h. The soluble GST fusionprotein was purified as described previously (Bar-Peledand Raikhel, 1996). Five hundred microliters (0.4–0.5 mg)of eluted GST-AtRGP1 fusion protein was emulsified bysonication with an equal amount of TiterMax adjuvant(CytRx Corp., Norcross, GA) and injected into New Zeal-and White rabbits. An equal amount of GST-AtRGP1 pro-tein was injected into the rabbits two more times to boostthe immune response over a 2-month period. Blood wastaken from the rabbits 1 month after the first injection andevery other week thereafter and treated as described pre-viously (Harlow and Lane, 1988).
Figure 1. Predicted protein sequence of AtRGP1 (GenBank acces-sion no. AF013627) and comparison with other RGP proteins. A,Amino acid sequence alignment of AtRGP1, PsRGP1 (U31565),VuRGP (AF005279), ZmRGP (U89897), and OsRGP using theMegAlign module of DNAStar. The black regions represent aminoacid differences between AtRGP1 and the other RGPs. The overlinehighlights a putative N-myristoylation site. The shaded box repre-sents the predicted UDP-Glc-binding domain U1 from cotton CelA.The open box is the site of glycosylation as determined in sweet corn.Underlined are the two pea tryptic peptides used to search the EST
database. B, Alignment of UDP-Glc-binding domains from variousorganisms, as determined by Delmer and Amor (1995), with thepredicted binding domain of RGP.
Reversibly Autoglycosylated Arabidopsis Protein 1341
www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 1998 American Society of Plant Biologists. All rights reserved.

Timed Studies of RNA and Protein Levels
A 0.5-L Arabidopsis cell-suspension culture was startedby adding 50 mL of a 7-d-old culture to 450 mL of freshsuspension-culture medium. Fifteen-milliliter aliquotswere taken at 24-h intervals over a 12-d span. Each fractionwas centrifuged at 50g for 5 min, the supernatant wasremoved, and the weight of the packed cells was measured.Equal amounts of protein (50 mg/lane) were separated bySDS-PAGE (Laemmli et al., 1970), and the presence ofAtRGP1 was determined by immunoblotting. Equalamounts of RNA (30 mg/lane) were separated in a 1%agarose and 2% formaldehyde gel, and the level of AtRGP1RNA was determined by northern analysis.
Reaction with UDP-Sugars
One hundred to 250 mg of total Arabidopsis, pea, to-bacco, or maize total protein from roots was labeled asdescribed previously (Dhugga et al., 1991). First, 0.1 mCi ofUDP-d-[U-14C]Glc (specific activity 263 mCi mmol21 [ICN])or 0.5 mCi of UDP-d-[6-3H]Gal (specific activity 10.6 mCimmol21 [ICN]), was added to the protein sample in avolume of 50 to 150 mL of lysis buffer containing 3 mmMgCl2. The reactions were stopped after a 10-min incuba-tion at room temperature by adding 10 to 30 mL of SDS-PAGE loading buffer (120 mm Tris, pH 6.8, 200 mm DTT,4% SDS, 0.02% bromphenol blue, and 20% glycerol). Theprotein samples were separated by SDS-PAGE and treatedas described below. Since the UDP-Glc and UDP-Gal reac-tions gave the same results, only the UDP-Glc reactions arepresented.
Reactions with purified GST-AtRGP1 protein were car-ried out using 2 mg of protein in the absence of Mg21. Thereactions were mixed by inversion and incubated staticallyat room temperature for 10 min. For displacement reac-tions, after a 10-min incubation with 20 pmol (0.2 mCi) ofUDP-[3H]Glc (specific activity 10.9 Ci mmol21; Sigma),UDP, Glc, UDP-Glc, UDP-Xyl, UDP-Gal, or UDP-Man wasadded to a final concentration of 3 mm and the reactionscontinued for 10 more min. To stop the reactions, SDS-PAGE loading buffer (120 mm Tris, pH 6.8, 200 mm DTT,4% SDS, 0.02% bromphenol blue, and 20% glycerol) wasadded. The protein samples were separated by SDS-PAGEand treated as described below.
Immunochemical Studies
Immunoprecipitation
Immunoprecipitations were as described previously(Harlow and Lane, 1988). Anti-AtRGP1 polyclonal antibod-ies were cross-linked to protein A-Sepharose 6MB (Phar-macia) and mixed with total protein from roots grown inliquid culture. After the samples were washed three timeswith PBS and once with 10 mm potassium phosphatebuffer, pH 7.0, proteins bound to the antibodies were
eluted with 100 mm Gly-HCl, pH 2.5. Protoplasts labeledwith [35S]Met were incubated overnight in a rotary shakerat 80 rpm, and total protein was prepared as describedabove, and immunoprecipitated.
SDS-PAGE and Immunoblotting
Protein concentration was determined as described pre-viously (Bradford, 1976) using BSA as the standard. Proteinmolecular weight standards were purchased from Bio-Rad(low- and broad-range markers, catalog nos. 161-0304 and161-0317, respectively). All protein samples were separatedon 12.5% SDS-PAGE gels using 13 running buffer (250 mmTris, 1 m Gly, and 0.5% SDS).
After separation, gels containing UDP-d-[U-14C]Glc- andUDP-d-[3H]Gal-labeled reactions were stained with 0.1%Coomassie blue overnight, destained with 5 volumes of30% methanol and 10% acetic acid, washed twice withdistilled water, incubated with Fluoro-Hance (ResearchProducts International, Prospect, IL) for 15 min, and driedfor 2 h in a gel drier (model 583, Bio-Rad). The dried gelswere exposed to film for 7 to 10 d at 280°C prior to filmdevelopment.
For immunoblotting, gels containing various proteinsamples (30–50 mg/lane) were washed with 5 volumes of13 transfer buffer (63 mm Trizma base, 250 mm Gly, 0.13%SDS, and 20% methanol) and transferred in the same bufferto a 0.45-mm nitrocellulose membrane (Hybond N, Amer-sham) using a semidry apparatus at 2 mA cm22 for 2 h.Filters were stained with 13 Ponceau-S (2% Ponceau-S,30% TCA, and 30% sulfosalicylic acid) and blocked over-night in 10% (w/v) dry milk powder in PBST (13 PBS and0.1% Tween 20). AtRGP1 polyclonal antibodies were usedat a 1:1000 dilution. Anti-RD28 plasma membrane markersera was used at 1:500 dilution and anti-binding protein ERlumen protein at 1:1000 dilution. They were a gift fromMaarten Chrispeels (University of California, San Diego).Anti-AtELP (partially Golgi-localized protein, Ahmed etal., 1997) and anti-AtPEP12p (post-Golgi compartment pro-tein, Conceicao et al., 1997) polyclonal antibodies wereused at a 1:500 dilution. ARA4 monoclonal antibody (aGolgi membrane-localized and -soluble protein, Ueda etal., 1996) was used at 1:500 dilution.
After primary antibody incubation for 1 to 2 h in PBST,membranes were washed three times with PBST and thenreacted with a 1:2500 dilution of alkaline phosphatase-conjugated goat anti-rabbit secondary antibody (Kirke-gaard and Perry Laboratories, Gaithersburg, MD) for 1 to2 h, except the ARA4 antibody, which required alkalinephosphatase-conjugated goat anti-mouse secondary anti-body. Finally, filters were washed twice with PBST, twicewith PBS, and once with alkaline buffer (100 mm Tris, 100mm NaCl, and 5 mm MgCl2 , pH 9.5) prior to detection ofthe immune complexes using the substrates 5-bromo-4-chloro-3-indolyl phosphate (150 mg mL21 final concentra-tion) and nitroblue tetrazolium (300 mg mL21 final concen-tration) in alkaline buffer.
1342 Delgado et al. Plant Physiol. Vol. 116, 1998
www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 1998 American Society of Plant Biologists. All rights reserved.

Localization
Differential Centrifugation
Total protoplast protein was centrifuged at 1,000g for 20min at 4°C. The pellet (p1) was washed with lysis bufferand centrifuged again. The supernatant (s1) was centri-fuged at 5,000g for 30 min. The pellet was washed withlysis buffer and centrifuged again (now called p5). Thesupernatant (now s5) was treated in the same way forsequential differential centrifugations at 15,000, 25,000,50,000, and 100,000g, with the last two centrifugations car-ried out for 40 min. All pellets (p1, p5, p15, p25, p50, andp100) were resuspended in lysis buffer and the protein wasquantified.
Membrane Association
Total microsomes were prepared by centrifugation oftotal protoplast protein at 150,000g for 1 h and washing thepellet with lysis buffer. Pellets were resuspended with lysisbuffer (total protein) or lysis buffer containing 0.1, 0.5, or1.0% Triton X-100, 0.5 or 2 m urea, 0.1 m sodium carbonate,0.1, 0.5, or 1.0 m NaCl, or 0.1 m potassium phosphate, pH7.0, for 1 h on ice, and centrifuged again at 150,000g for 1 h.The resulting pellets were washed with lysis buffer, resus-pended in loading buffer, and separated by SDS-PAGEprior to analysis by immunoblot using anti-AtRGP1 anti-bodies.
Suc-Density-Gradient Centrifugation
Total protoplast protein was centrifuged at 1000g for 10min at 4°C, and 6 mL of the supernatant (S1) was loaded ontop of a 16 to 55% equilibrium-density Suc gradient mod-ified from Gibeaut and Carpita (1994). To prepare thegradient, stock solutions of 55, 40, 33.5, 26.5, and 16%(w/v) Suc buffered in 10 mm Hepes-KOH, pH 7.0, 10 mmpotassium acetate, and 2 mm EDTA were prepared. Gra-dients were prepared by the sequential addition of 2.7 mLof 55% Suc solution, 8.8 mL of 40% Suc solution, 7.0 mL of33.5% Suc solution, 6.0 mL of 26.5% Suc solution, and 4.5mL of 16% Suc solution to 35-mL polyallomer tubes (cata-log no. 326823, Beckman). After these solutions were care-fully layered, they were centrifuged at 150,000g for 18 to20 h at 4°C in a rotor (model SW28, Beckman). This resultedin an equilibrium-density gradient similar to one obtainedusing a gradient maker. Equal-volume fractions were col-lected from the bottom of the gradients, and the Suc con-centration was estimated by measuring the refractive indexof the individual fractions using a refractometer. Aliquotsof the fractions (50–100 mL) were separated by SDS-PAGE,and the separation of AtRGP1 and membrane marker pro-teins were analyzed by immunoblotting.
RESULTS
Identification of an Arabidopsis cDNA Encoding anAtRGP1 Protein
Sequences from three purified tryptic peptides from thepea RGP protein doublet were presented by Dhugga and
Ray (1994) and used to search the dBEST database. A totalof 28 Arabidopsis ESTs have been obtained so far from theoriginal and subsequent searches. Based on sequence sim-ilarity, these ESTs can be assigned to at least three differentgroups, denoted AtRGP1 (AF013627), AtRGP2 (AF013628),and AtRGP3 (AF034255). These cDNAs share between 93and 99% sequence identity at the amino acid level. Becausethe similarity is spread through the whole sequence,only one cDNA, AtRGP1, was used for further analysis(Fig. 1A).
We used the full-length cDNA of AtRGP1 to search thedBEST databases and obtained, in addition to the 28 Ara-bidopsis cDNAs, 19 rice cDNAs, a full-length maize cDNA,and a full-length cowpea cDNA, all of which shared sig-nificant sequence similarity with AtRGP1. None of thesesequences belonged to a gene with an assigned function atthe time of the BLAST search. AtRGP1 is 84, 87, 83, and 85%identical to the pea, cowpea, maize, and rice proteins,PsRGP1, VuRGP, ZmRGP, and OsRGP, respectively. Afull-length cDNA clone encoding the rice RGP has not yetbeen isolated, but available ESTs can be combined to makeup a full-length clone (OsRGP). Searches of nonplant data-bases gave mixed results. Although similar sequences werefound in both the yeast genome database (http://speedy.mips.biochem.mpg.de/mips/yeast/index.htmlx)and the Synechocystis sp. strain PCC6803 database (http://www.kazusa.or.jp/cyano/cyano.html), none shared sig-nificant similarity to AtRGP1 (P . 0.01), all of the sequencealignments were very short (10–20 amino acids), and noneof the predicted open reading frames encoded a 41-kDprotein. These findings are further supported by the im-munoscreening of various species described below.
Hydropathy profile analysis (Kyte and Doolittle, 1982)predicts that AtRGP1 is a hydrophilic protein (data notshown). Visual and computer-aided (pSORT, http://psort.nibb.ac.jp/) analyses of the protein sequence predict thatthe protein lacks a signal sequence that might directAtRGP1 to a membrane compartment within the cell. Thesame computer program predicts the existence of anN-myristoylation site in AtRGP1 (Fig. 1A). N-myristoyla-tion has been shown to aid in the association of otherwisesoluble proteins to the cytosolic surface of membranes.
The sequence of AtRGP1 was compared with the re-ported predicted binding domains for UDP-Glc. The deter-mination of these UDP-Glc-binding domains is based onthe observation that there is a substantial sequence conser-vation between the bacterial catalytic subunit and otherenzymes that catalyze the polymerization of b-glycosylresidues from either UDP-Glc or UDP-GlcNAc (Saxena etal., 1995), as well as UDP-Glc-binding proteins (Delmerand Amor, 1995; Pear et al., 1996). AtRGP1 is 40% identicaland 53% similar to the predicted UDP-Glc-binding domain(U1) of cotton CelA (Delmer and Amor, 1995; Pear et al.,1996; Saxena and Brown, 1997). Furthermore, AtRGP1 con-tained the critical Asp residue (Fig. 1B). In contrast, se-quence identity between the deduced amino acid sequenceof cotton CelA and AtRGP1 is very low, approximately 15%.
Singh et al. (1995) demonstrated that the Glc from UDP-Glc was attached to an Arg residue of a maize protein withhigh sequence similarity to AtRGP1. The tryptic peptide
Reversibly Autoglycosylated Arabidopsis Protein 1343
www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 1998 American Society of Plant Biologists. All rights reserved.
containing the glucosylated Arg is 93% identical to a regionof AtRGP1 (Fig. 1A). Dhugga et al. (1997) noted this samesimilarity to PsRGP1 and postulated that Arg-158 is thelocation of a sugar addition in the pea protein. Similarly,we postulate that Arg-158 is the location of Glc addition inthe Arabidopsis protein.
Distribution of RGP1 Transcript and Protein
The isolation of RGP proteins in both monocots anddicots suggests a general function for this protein. To con-firm the presence of proteins similar to RGP, as evidencedby database searches, we used AtRGP1 antibodies in im-munoblots against protein from various organisms. Immu-noreactive polypeptides of 41 kD were detected in Arabi-dopsis, pea, tobacco, and maize (Fig. 2). Although peaRGP1 has been referred to as a polypeptide of about 40 kD(Dhugga et al., 1991, 1997), it is clear from the cloned cDNAthat PsRGP1 has a predicted size of 41 kD. Since all RGPsso far identified encode proteins of approximately the samesize, we will refer to them as 41-kD proteins.
A 64-kD protein was also detected by AtRGP1 antibodiesin immunoblots of Arabidopsis, pea, tobacco, maize, andSynechocystis sp. strain PCC6803 (Fig. 2). The relationshipof this protein to the 41-kD protein is not yet clear. Al-though four hypothetical proteins were found in the Syn-echocystis sp. strain PCC6803 database (sll0524, sll0501,sll0602, and sll1080) that shared limited sequence similaritywithin separate regions in AtRGP1, none was of the sizedetected by immunoblotting. AtRGP1 antibodies did notdetect any proteins in either yeast or mammalian extracts(Fig. 2), as expected from database searches.
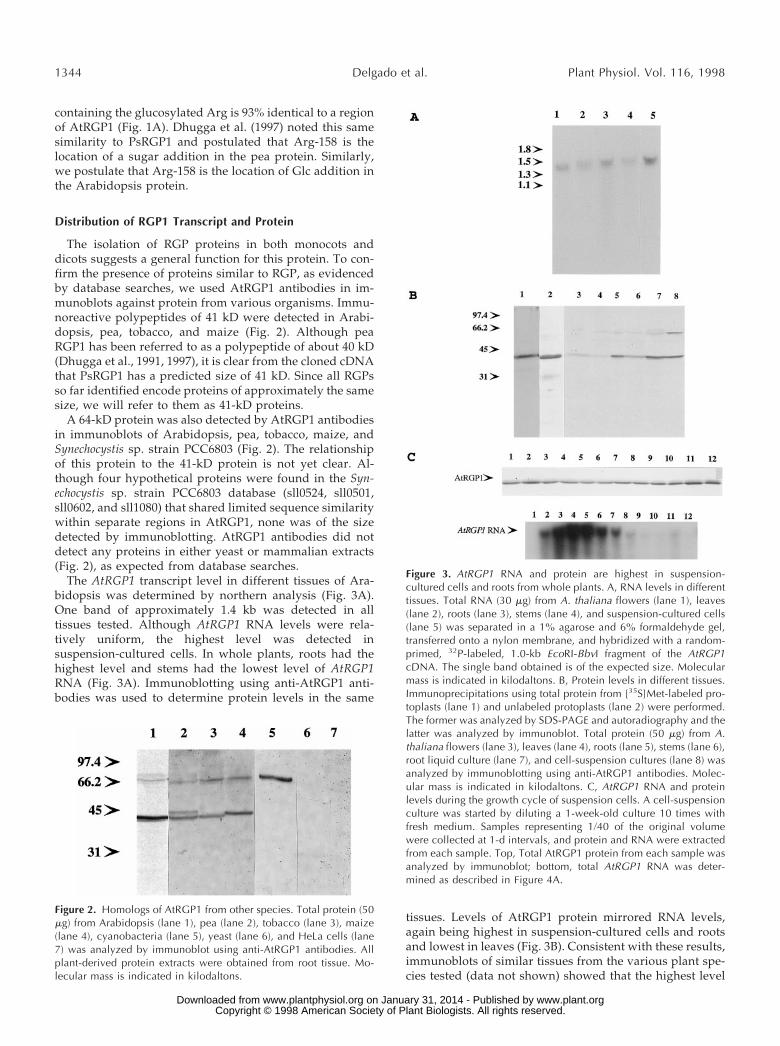
The AtRGP1 transcript level in different tissues of Ara-bidopsis was determined by northern analysis (Fig. 3A).One band of approximately 1.4 kb was detected in alltissues tested. Although AtRGP1 RNA levels were rela-tively uniform, the highest level was detected insuspension-cultured cells. In whole plants, roots had thehighest level and stems had the lowest level of AtRGP1RNA (Fig. 3A). Immunoblotting using anti-AtRGP1 anti-bodies was used to determine protein levels in the same
tissues. Levels of AtRGP1 protein mirrored RNA levels,again being highest in suspension-cultured cells and rootsand lowest in leaves (Fig. 3B). Consistent with these results,immunoblots of similar tissues from the various plant spe-cies tested (data not shown) showed that the highest level
Figure 2. Homologs of AtRGP1 from other species. Total protein (50mg) from Arabidopsis (lane 1), pea (lane 2), tobacco (lane 3), maize(lane 4), cyanobacteria (lane 5), yeast (lane 6), and HeLa cells (lane7) was analyzed by immunoblot using anti-AtRGP1 antibodies. Allplant-derived protein extracts were obtained from root tissue. Mo-lecular mass is indicated in kilodaltons.
Figure 3. AtRGP1 RNA and protein are highest in suspension-cultured cells and roots from whole plants. A, RNA levels in differenttissues. Total RNA (30 mg) from A. thaliana flowers (lane 1), leaves(lane 2), roots (lane 3), stems (lane 4), and suspension-cultured cells(lane 5) was separated in a 1% agarose and 6% formaldehyde gel,transferred onto a nylon membrane, and hybridized with a random-primed, 32P-labeled, 1.0-kb EcoRI-BbvI fragment of the AtRGP1cDNA. The single band obtained is of the expected size. Molecularmass is indicated in kilodaltons. B, Protein levels in different tissues.Immunoprecipitations using total protein from [35S]Met-labeled pro-toplasts (lane 1) and unlabeled protoplasts (lane 2) were performed.The former was analyzed by SDS-PAGE and autoradiography and thelatter was analyzed by immunoblot. Total protein (50 mg) from A.thaliana flowers (lane 3), leaves (lane 4), roots (lane 5), stems (lane 6),root liquid culture (lane 7), and cell-suspension cultures (lane 8) wasanalyzed by immunoblotting using anti-AtRGP1 antibodies. Molec-ular mass is indicated in kilodaltons. C, AtRGP1 RNA and proteinlevels during the growth cycle of suspension cells. A cell-suspensionculture was started by diluting a 1-week-old culture 10 times withfresh medium. Samples representing 1/40 of the original volumewere collected at 1-d intervals, and protein and RNA were extractedfrom each sample. Top, Total AtRGP1 protein from each sample wasanalyzed by immunoblot; bottom, total AtRGP1 RNA was deter-mined as described in Figure 4A.
1344 Delgado et al. Plant Physiol. Vol. 116, 1998
www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 1998 American Society of Plant Biologists. All rights reserved.

of RGP1 protein was found in roots (Fig. 2). Although RNAlevels are similar in leaves and stems, more AtRGP1 pro-tein can be found in stems than in leaves.
AtRGP1 was detected only as a doublet in tissues inwhich it was highly expressed (Figs. 2 and 3B), but it wasclearly not the same kind of doublet as that seen for pea(Fig. 2). In addition, a 64-kD protein was detected byimmunoblot analysis of total protein in nearly all tissuestested (Fig. 3B), as well as in all of the other plant speciestested (Fig. 2). The nature of this protein is not understood,and it does not seem to be well recognized by AtRGP1antibodies, since immunoprecipitation studies recognizedAtRGP1 almost exclusively (Fig. 3B).
Since AtRGP1 RNA and the corresponding protein werehighest in suspension-cultured cells, its behavior was stud-ied by starting cultures and then following them through atime course. As seen in Figure 3C, cells were grown for aperiod of 12 d and aliquots were taken at 24-h intervals todetermine the amount of AtRGP1 RNA and its correspond-ing protein. Although there was a clear increase in RNAaccumulation during the first 5 d, peaking at d 4 andalmost undetectable by d 8, the corresponding protein levelwas constant throughout the experiment.
AtRGP1 Is Soluble and Membrane Associated
One strategy for understanding the function of AtRGP1is to localize this protein. Hydropathy plots predict thatAtRGP1 is a soluble protein (not shown) although analysisof the sequence shows that it contains a putativeN-myristoylation site that may be involved in its associa-tion with membranes (Fig. 1A). Fractionation studies werecarried out to determine whether AtRGP1 is soluble ormembrane associated. Protoplasts derived from cell-suspension cultures were lysed and subjected to sequentialdifferential centrifugation. The resulting pellets were ana-lyzed by immunoblot using AtRGP1 antibodies (Fig. 4A).Several intrinsic membrane proteins (AtPEP12p, RD28, andAtELP), membrane-associated and soluble proteins(ARA4), and soluble proteins (ER lumen and BiP) wereanalyzed in the same fractions and used as a control.AtRGP1 was enriched in fractions composed of solubleproteins (s100) but was also detected in membrane-containing fractions (p5-p100, Fig. 4A). Our results indicatethat AtRGP1 is both soluble (s100) and membrane associ-ated.
The presence of AtRGP1 in both soluble and membrane-bound fractions is interesting and may provide hints re-garding its role in the cell. Since AtRGP1 is predicted to bea soluble protein, we were specifically interested in study-ing the nature of its association with membranes. Equalamounts of total microsomes were prepared (p150) andpellets were resuspended under various conditions. Aftertreated membranes were centrifuged at 150,000g, pelletswere resuspended in lysis buffer to analyze by immunoblotany protein left that was associated with membranes. Totalprotein (Fig. 4B) and microsomes resuspended in lysisbuffer containing Triton X-100, urea, sodium carbonate,NaCl, or potassium phosphate buffer alone were pelletedand resuspended in lysis buffer for immunoblot analysis
(Fig. 4B). Nonionic detergents, high levels of urea (2 m),and alkaline treatment eluted AtRGP1 from the membrane(Fig. 4B), whereas low levels of urea (0.5 m), NaCl treat-ment, and a hypotonic wash with a neutral buffer left someAtRGP1 associated with the membrane (Fig. 4B). This sug-gests that AtRGP1 is a peripheral membrane protein.
As a further attempt to determine the localization ofAtRGP1, a postnuclear supernatant (s1) fraction of proto-plast lysates derived from cell-suspension cultures wasfractionated on Suc gradients and the individual fractionswere analyzed by immunoblot (Fig. 5). AtRGP1 peaked infraction 10, corresponding to approximately 30% Suc. BiP,
Figure 4. AtRGP1 is soluble and membrane associated. A, Totalprotoplast protein from suspension-cultured cell protoplasts was cen-trifuged at 1,000, 5,000, 10,000, 15,000, 25,000, 50,000, and100,000g. The resultant pellets were resuspended in lysis buffer andmake up fractions p1, p5, p10, p15, p25, p50, and p100, respec-tively. s100 denotes the supernatant after the 100,000g centrifuga-tion and represents total soluble proteins. Equal volumes of proteinwere separated by SDS-PAGE, transferred to nitrocellulose, and an-alyzed by immunoblot. Various soluble proteins (BiP), integral mem-brane proteins (AtPEP12p, RD28, and AtELP), and peripheral mem-brane proteins (ARA4) were compared with the membraneassociation of AtRGP1. The fraction of total protein (T) present ineach pellet is shown. B, AtRGP1 is a peripheral membrane protein.Total microsomes were prepared by centrifuging total protein at150,000g for 1 h and washing the pellet with lysis buffer. Pellets wereresuspended with various buffers for 1 h on ice and centrifuged againat 150,000g for 1 h, and the pellets were analyzed by immunoblotusing anti-AtRGP1 antibodies. Microsomes were resuspended withlysis buffer (lane 1, total protein) or lysis buffer containing 0.1, 0, or1.0% Triton X-100 (lanes 2, 3, and 4, respectively), 0.5 or 2 M urea(lanes 5 and 6, respectively), 0.1 M sodium carbonate (lane 7), 0.1,0.5, or 1.0 M NaCl (lanes 8, 9, and 10, respectively), or 0.1 M
potassium phosphate buffer, pH 7.0, alone (lane 11).
Reversibly Autoglycosylated Arabidopsis Protein 1345
www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 1998 American Society of Plant Biologists. All rights reserved.
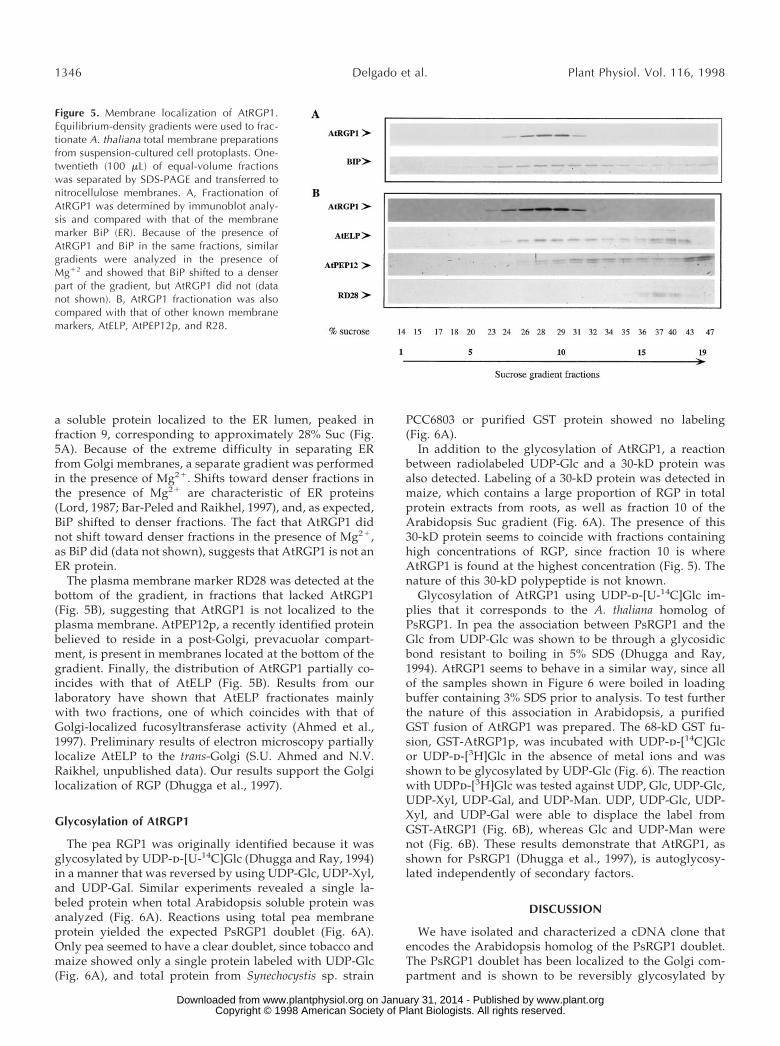
a soluble protein localized to the ER lumen, peaked infraction 9, corresponding to approximately 28% Suc (Fig.5A). Because of the extreme difficulty in separating ERfrom Golgi membranes, a separate gradient was performedin the presence of Mg21. Shifts toward denser fractions inthe presence of Mg21 are characteristic of ER proteins(Lord, 1987; Bar-Peled and Raikhel, 1997), and, as expected,BiP shifted to denser fractions. The fact that AtRGP1 didnot shift toward denser fractions in the presence of Mg21,as BiP did (data not shown), suggests that AtRGP1 is not anER protein.
The plasma membrane marker RD28 was detected at thebottom of the gradient, in fractions that lacked AtRGP1(Fig. 5B), suggesting that AtRGP1 is not localized to theplasma membrane. AtPEP12p, a recently identified proteinbelieved to reside in a post-Golgi, prevacuolar compart-ment, is present in membranes located at the bottom of thegradient. Finally, the distribution of AtRGP1 partially co-incides with that of AtELP (Fig. 5B). Results from ourlaboratory have shown that AtELP fractionates mainlywith two fractions, one of which coincides with that ofGolgi-localized fucosyltransferase activity (Ahmed et al.,1997). Preliminary results of electron microscopy partiallylocalize AtELP to the trans-Golgi (S.U. Ahmed and N.V.Raikhel, unpublished data). Our results support the Golgilocalization of RGP (Dhugga et al., 1997).
Glycosylation of AtRGP1
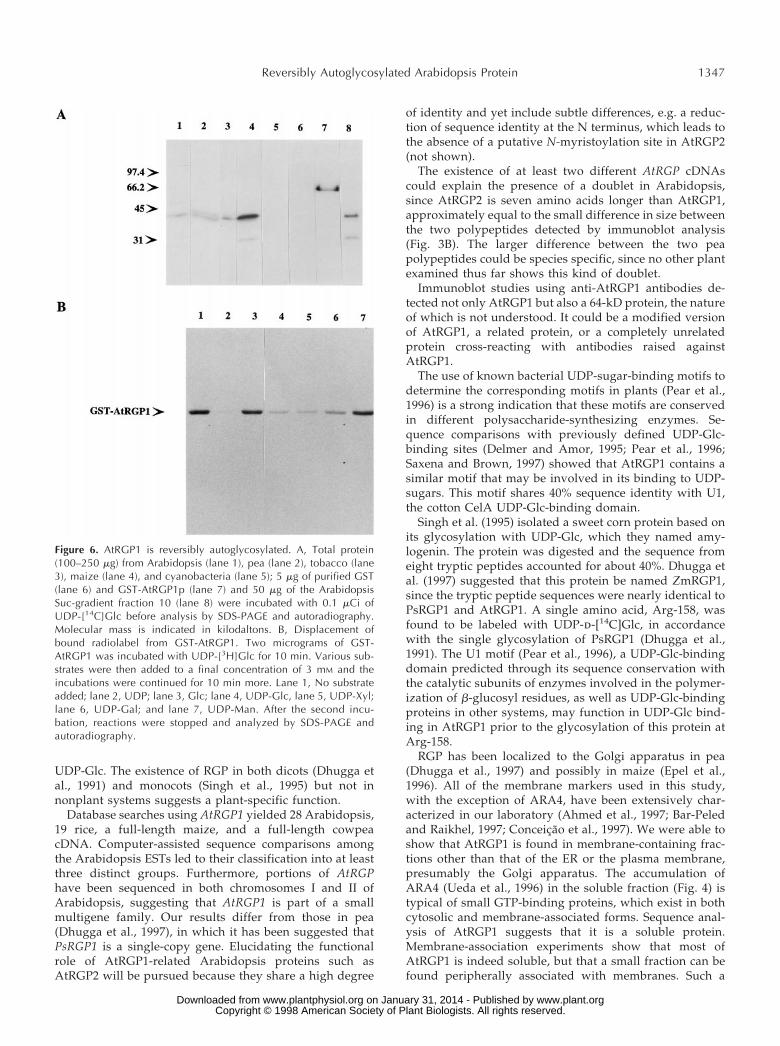
The pea RGP1 was originally identified because it wasglycosylated by UDP-d-[U-14C]Glc (Dhugga and Ray, 1994)in a manner that was reversed by using UDP-Glc, UDP-Xyl,and UDP-Gal. Similar experiments revealed a single la-beled protein when total Arabidopsis soluble protein wasanalyzed (Fig. 6A). Reactions using total pea membraneprotein yielded the expected PsRGP1 doublet (Fig. 6A).Only pea seemed to have a clear doublet, since tobacco andmaize showed only a single protein labeled with UDP-Glc(Fig. 6A), and total protein from Synechocystis sp. strain
PCC6803 or purified GST protein showed no labeling(Fig. 6A).
In addition to the glycosylation of AtRGP1, a reactionbetween radiolabeled UDP-Glc and a 30-kD protein wasalso detected. Labeling of a 30-kD protein was detected inmaize, which contains a large proportion of RGP in totalprotein extracts from roots, as well as fraction 10 of theArabidopsis Suc gradient (Fig. 6A). The presence of this30-kD protein seems to coincide with fractions containinghigh concentrations of RGP, since fraction 10 is whereAtRGP1 is found at the highest concentration (Fig. 5). Thenature of this 30-kD polypeptide is not known.
Glycosylation of AtRGP1 using UDP-d-[U-14C]Glc im-plies that it corresponds to the A. thaliana homolog ofPsRGP1. In pea the association between PsRGP1 and theGlc from UDP-Glc was shown to be through a glycosidicbond resistant to boiling in 5% SDS (Dhugga and Ray,1994). AtRGP1 seems to behave in a similar way, since allof the samples shown in Figure 6 were boiled in loadingbuffer containing 3% SDS prior to analysis. To test furtherthe nature of this association in Arabidopsis, a purifiedGST fusion of AtRGP1 was prepared. The 68-kD GST fu-sion, GST-AtRGP1p, was incubated with UDP-d-[14C]Glcor UDP-d-[3H]Glc in the absence of metal ions and wasshown to be glycosylated by UDP-Glc (Fig. 6). The reactionwith UDPd-[3H]Glc was tested against UDP, Glc, UDP-Glc,UDP-Xyl, UDP-Gal, and UDP-Man. UDP, UDP-Glc, UDP-Xyl, and UDP-Gal were able to displace the label fromGST-AtRGP1 (Fig. 6B), whereas Glc and UDP-Man werenot (Fig. 6B). These results demonstrate that AtRGP1, asshown for PsRGP1 (Dhugga et al., 1997), is autoglycosy-lated independently of secondary factors.
DISCUSSION
We have isolated and characterized a cDNA clone thatencodes the Arabidopsis homolog of the PsRGP1 doublet.The PsRGP1 doublet has been localized to the Golgi com-partment and is shown to be reversibly glycosylated by
Figure 5. Membrane localization of AtRGP1.Equilibrium-density gradients were used to frac-tionate A. thaliana total membrane preparationsfrom suspension-cultured cell protoplasts. One-twentieth (100 mL) of equal-volume fractionswas separated by SDS-PAGE and transferred tonitrocellulose membranes. A, Fractionation ofAtRGP1 was determined by immunoblot analy-sis and compared with that of the membranemarker BiP (ER). Because of the presence ofAtRGP1 and BiP in the same fractions, similargradients were analyzed in the presence ofMg12 and showed that BiP shifted to a denserpart of the gradient, but AtRGP1 did not (datanot shown). B, AtRGP1 fractionation was alsocompared with that of other known membranemarkers, AtELP, AtPEP12p, and R28.
1346 Delgado et al. Plant Physiol. Vol. 116, 1998
www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 1998 American Society of Plant Biologists. All rights reserved.
UDP-Glc. The existence of RGP in both dicots (Dhugga etal., 1991) and monocots (Singh et al., 1995) but not innonplant systems suggests a plant-specific function.
Database searches using AtRGP1 yielded 28 Arabidopsis,19 rice, a full-length maize, and a full-length cowpeacDNA. Computer-assisted sequence comparisons amongthe Arabidopsis ESTs led to their classification into at leastthree distinct groups. Furthermore, portions of AtRGPhave been sequenced in both chromosomes I and II ofArabidopsis, suggesting that AtRGP1 is part of a smallmultigene family. Our results differ from those in pea(Dhugga et al., 1997), in which it has been suggested thatPsRGP1 is a single-copy gene. Elucidating the functionalrole of AtRGP1-related Arabidopsis proteins such asAtRGP2 will be pursued because they share a high degree
of identity and yet include subtle differences, e.g. a reduc-tion of sequence identity at the N terminus, which leads tothe absence of a putative N-myristoylation site in AtRGP2(not shown).
The existence of at least two different AtRGP cDNAscould explain the presence of a doublet in Arabidopsis,since AtRGP2 is seven amino acids longer than AtRGP1,approximately equal to the small difference in size betweenthe two polypeptides detected by immunoblot analysis(Fig. 3B). The larger difference between the two peapolypeptides could be species specific, since no other plantexamined thus far shows this kind of doublet.
Immunoblot studies using anti-AtRGP1 antibodies de-tected not only AtRGP1 but also a 64-kD protein, the natureof which is not understood. It could be a modified versionof AtRGP1, a related protein, or a completely unrelatedprotein cross-reacting with antibodies raised againstAtRGP1.
The use of known bacterial UDP-sugar-binding motifs todetermine the corresponding motifs in plants (Pear et al.,1996) is a strong indication that these motifs are conservedin different polysaccharide-synthesizing enzymes. Se-quence comparisons with previously defined UDP-Glc-binding sites (Delmer and Amor, 1995; Pear et al., 1996;Saxena and Brown, 1997) showed that AtRGP1 contains asimilar motif that may be involved in its binding to UDP-sugars. This motif shares 40% sequence identity with U1,the cotton CelA UDP-Glc-binding domain.
Singh et al. (1995) isolated a sweet corn protein based onits glycosylation with UDP-Glc, which they named amy-logenin. The protein was digested and the sequence fromeight tryptic peptides accounted for about 40%. Dhugga etal. (1997) suggested that this protein be named ZmRGP1,since the tryptic peptide sequences were nearly identical toPsRGP1 and AtRGP1. A single amino acid, Arg-158, wasfound to be labeled with UDP-d-[14C]Glc, in accordancewith the single glycosylation of PsRGP1 (Dhugga et al.,1991). The U1 motif (Pear et al., 1996), a UDP-Glc-bindingdomain predicted through its sequence conservation withthe catalytic subunits of enzymes involved in the polymer-ization of b-glucosyl residues, as well as UDP-Glc-bindingproteins in other systems, may function in UDP-Glc bind-ing in AtRGP1 prior to the glycosylation of this protein atArg-158.
RGP has been localized to the Golgi apparatus in pea(Dhugga et al., 1997) and possibly in maize (Epel et al.,1996). All of the membrane markers used in this study,with the exception of ARA4, have been extensively char-acterized in our laboratory (Ahmed et al., 1997; Bar-Peledand Raikhel, 1997; Conceicao et al., 1997). We were able toshow that AtRGP1 is found in membrane-containing frac-tions other than that of the ER or the plasma membrane,presumably the Golgi apparatus. The accumulation ofARA4 (Ueda et al., 1996) in the soluble fraction (Fig. 4) istypical of small GTP-binding proteins, which exist in bothcytosolic and membrane-associated forms. Sequence anal-ysis of AtRGP1 suggests that it is a soluble protein.Membrane-association experiments show that most ofAtRGP1 is indeed soluble, but that a small fraction can befound peripherally associated with membranes. Such a
Figure 6. AtRGP1 is reversibly autoglycosylated. A, Total protein(100–250 mg) from Arabidopsis (lane 1), pea (lane 2), tobacco (lane3), maize (lane 4), and cyanobacteria (lane 5); 5 mg of purified GST(lane 6) and GST-AtRGP1p (lane 7) and 50 mg of the ArabidopsisSuc-gradient fraction 10 (lane 8) were incubated with 0.1 mCi ofUDP-[14C]Glc before analysis by SDS-PAGE and autoradiography.Molecular mass is indicated in kilodaltons. B, Displacement ofbound radiolabel from GST-AtRGP1. Two micrograms of GST-AtRGP1 was incubated with UDP-[3H]Glc for 10 min. Various sub-strates were then added to a final concentration of 3 mM and theincubations were continued for 10 min more. Lane 1, No substrateadded; lane 2, UDP; lane 3, Glc; lane 4, UDP-Glc, lane 5, UDP-Xyl;lane 6, UDP-Gal; and lane 7, UDP-Man. After the second incu-bation, reactions were stopped and analyzed by SDS-PAGE andautoradiography.
Reversibly Autoglycosylated Arabidopsis Protein 1347
www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 1998 American Society of Plant Biologists. All rights reserved.

distribution within the cell suggests that the reaction ofAtRGP1 with UDP-sugars takes place in the cytoplasm.
Our working hypothesis is that AtRGP1 may be in someway involved in polysaccharide synthesis, possibly as aprotein primer or as some form of intermediate. There is noevidence for a primer function other than an analogy toprotein-primed starch and glycogen synthesis (Moreno etal., 1986). AtRGP1’s reversible glycosylation, its prominentresidence in the cytoplasm, where nucleotide sugars arefound, and its transient association with membranes sug-gests that it functions as a carrier of UDP-sugars from thecytoplasm to membranes such as the Golgi apparatus.
Our understanding of the enzymes involved in cell wallbiosynthesis is scant at best, and so far, little progress hasbeen made in isolating these enzymes, which is a require-ment for studying their function. With this in mind, the useof cell-suspension cultures for the study of cell wall bio-synthesis may prove to be highly beneficial. Our prelimi-nary observations show that at the transcriptional levelAtRGP1 RNA can accumulate severalfold within 4 d aftersubculture and disappear completely after d 8. The fact thatAtRGP1 protein persisted throughout this time course isnot surprising, since cell-suspension systems are known tobe very active. For example, processes such as the secretionof cell wall polysaccharides continue unhindered becausethere is less spatial limit to cell wall deposition due tocell-to-cell contact.
Further investigation is necessary to determine the func-tion of AtRGP1. Our studies support the notion thatAtRGP1 is capable of autoglycosylation using UDP-Glcand UDP-Gal, most likely from the cytoplasmic pool ofsugar nucleotides, and carry these sugars to membranes,most likely the Golgi apparatus. Its function when associ-ated with membranes is not clear but may involve thepriming of polysaccharide synthesis. These observations,along with those made for the PsRGP1 doublet (Dhugga etal., 1991), suggest that AtRGP1 may play a role in cell wallbiosynthesis.
ACKNOWLEDGMENTS
We thank all of the members of the Raikhel group, especiallyAnton Sanderfoot and Diane Bassham, for their help with technicaland conceptual matters; the members of the cell wall group, in-cluding Hans Kende and Jonathan Walton, for helpful discussions;Maarten Chrispeels for the gifts of BiP and RD 28 antibodies; andTakashi Ueda for ARA-4 antibodies.
Received September 5, 1997; accepted December 1, 1997.Copyright Clearance Center: 0032–0889/98/116/1339/12.
LITERATURE CITED
Ahmed SU, Bar-Peled M, Raikhel NV (1997) Cloning and subcel-lular localization of an Arabidopsis receptor-like protein thatshares common features with protein-sorting receptors of eu-karyotic cells Plant Physiol 114: 325–336
Albersheim P, Darvill A, Roberts K, Staehelin AL, Varner JE(1997) Do the structures of cell wall polysaccharides define theirmode of synthesis? Plant Physiol 113: 1–3
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990)Basic local alignment search tool. J Mol Biol 215: 403–410
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, MillerW, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a newgeneration of protein database search programs. Nucleic AcidsRes 25: 3389–3402
Axelos M, Curie C, Mazzolini L, Bardet C, Lescure B (1992) Aprotocol for transient expression in Arabidopsis thaliana proto-plasts isolated from cell suspension cultures. Plant Physiol Bio-chem 30: 123–128
Bacic A, Harris PJ, Stone BA (1988) Structure and function of plantcell walls. In J Preiss, ed, The Biochemistry of Plants, Vol 14.Academic Press, San Diego, CA, pp 297–371
Bar-Peled M, Raikhel NV (1996) A method for isolation andpurification of specific antibodies to a protein fused to GST.Anal Biochem 241: 140–142
Bar-Peled M, Raikhel NV (1997) Characterization of AtSEC12 andAtSAR1 proteins likely involved in ER and Golgi traffic. PlantPhysiol 114: 315–324
Bradford MM (1976) A rapid and sensitive method for the quan-tification of nanogram quantities of proteins utilizing the prin-ciple of dye-binding. Anal Biochem 72: 248–254
Brummell DA, Camirand A, Maclachlan GA (1990) Differentialdistribution of xyloglucan glycosyl transferases in pea Golgidictyosomes and secretory vesicles. J Cell Sci 96: 705–710
Campbell RE, Brett CT, Hillman JR (1988) A xylosyltransferaseinvolved in the synthesis of a protein-associated xyloglucan insuspension-cultured dwarf-French-bean (Phaseolus vulgaris) cellsand its interaction with a glycosyltransferase. Biochem J 253:795–800
Carpita NC, Gibeaut DM (1993) Structural models of the primarycell walls of flowering plants: consistency of molecular structurewith the physical properties of the walls during growth. Plant J3: 1–30
Carpita NC, McCann MC, Griffing LR (1996) The plant extracel-lular matrix: news from the cell’s frontier. Plant Cell 8: 1451–1463
Conceicao AS, Marty-Mazars D, Bassham DC, Sanderfoot A,Marty F, Raikhel NV (1997) The syntaxin homologue AtPEP12presides on a late post-Golgi compartment in plants. Plant Cell 9:571–581
Delmer DP, Amor Y (1995) Cellulose biosynthesis. Plant Cell 7:987–1000
Dhugga KS, Ray PM (1994) Purification of the reversibly glyco-sylated polypeptides from pea. Purified polypeptides exhibit thesame properties as the Golgi-bound form (abstract no. 684).Plant Physiol 105: S-126
Dhugga KS, Tiwari SC, Ray PM (1997) A reversibly glycosylatedpolypeptide (RGP1) possibly involved in plant cell wall synthe-sis: purification, gene cloning and trans Golgi localization. ProcNatl Acad Sci USA 94: 7679–7684
Dhugga KS, Ulvskov P, Gallagher SR, Ray PM (1991) Plantpolypeptides reversibly glycosylated by UDP-glucose: possiblecomponents of Golgi b-glucan synthase in pea cells. J Biol Chem266: 21977–21984
Driouich A, Faye L, Staehelin LA (1993) The plant Golgi appara-tus: a factory for complex polysaccharides and glycoproteins.Trends Biochem Sci 18: 210–214
Epel BL, Van Lent JWM, Cohen L, Kotlizky G, Katz A, YahalomA (1996) A 41 kDa protein isolated form maize mesocotyl cellwalls immunolocalizes to plasmodesmata. Protoplasma 191:70–78
Faık A, Chileshe C, Sterling J, Maclachan G (1997) Xyloglucangalactosyl- and fucosyltransferase activities from pea epicotylmicrosomes. Plant Physiol 114: 245–254
Gibeaut DM, Carpita NC (1994) Biosynthesis of plant cell wallpolysaccharides. FASEB J 8: 904–915
Harlow E, Lane D (1988) Antibodies: A Laboratory Manual. ColdSpring Harbor Laboratory Press, Cold Spring Harbor, NY
Hayashi T (1989) Xyloglucans in the primary cell wall. Annu RevPlant Physiol Plant Mol Biol 40: 139–168
Kleene R, Berger EG (1993) The molecular and cell biology ofglucosyltransferases. Biochim Biophys Acta 1154: 283–325
Kyte J, Doolittle RF (1982) A simple method for displaying thehydropathic character of a protein. J Mol Biol 157: 105–132
1348 Delgado et al. Plant Physiol. Vol. 116, 1998
www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 1998 American Society of Plant Biologists. All rights reserved.

Laemmli, UK (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227: 680–685
Levy S, Staehelin LA (1992) Synthesis, assembly and function ofplant cell wall macromolecules. Curr Opin Cell Biol 4: 856–862
Lord JM (1987) Isolation of endoplasmic reticulum: general prin-ciples, enzymatic markers, and endoplasmic reticulum-boundpolysomes. Methods Enzymol 148: 576–584
Moreno S, Cardini CE, Tandecarz JS (1986) a-Glucan synthesis ona protein primer, uridine diphosphoglucose:protein transglyco-sylase. I. Separation from starch synthetase and phosphorylaseand a study of its properties. Eur J Biochem 157: 539–545
Newman T, de Brujn FJ, Green P, Keegstra K, Kende H, McIn-tosh L, Ohlrogge J, Raikhel NV, Somerville S, Tomashow M,and others (1994) Genes galore. A summary of methods foraccessing results from a large-scale partial sequencing of anon-ymous Arabidopsis cDNA clones. Plant Physiol 106: 1241–1255
Pear JR, Kawagoe Y, Schreckengost WE, Delmer DP, Stalker DM(1996) Higher plants contain homologs of the bacterial celAgenes encoding the catalytic subunit of cellulose synthase. ProcNatl Acad Sci USA 93: 12637–12642
Puissant C, Houdebine L-M (1990) An improvement of the single-step method of RNA isolation by acid guanidium thiocyanate-phenol-chloroform extraction. Biotechniques 8: 148–149
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular Cloning: ALaboratory Manual, Ed 2. Cold Spring Harbor Laboratory Press,Cold Spring Harbor, NY
Saxena IM, Brown RM Jr (1997) Identification of cellulose syn-thase(s) in higher plants: sequence analysis of processive b-gly-
cosyltransferases with the common motif “D,D,D35Q(R,Q)XRW”.Cellulose 4: 33–49
Saxena IM, Brown RM Jr, Fevre M, Geremia RA, Henrissat B(1995) Multidomain architecture of b-glycosyl transferases: im-plications for a mechanism of action. J Bacteriol 177: 1419–1424
Singh DG, Lomako J, Lomako WM, Whelan WJ, Meyer HE,Serwe M, Metzger JW (1995) b-Glycosylarginine: a newglucose-protein bond in a self-glucosylating protein from sweetcorn. FEBS Lett 376: 61–64
Staehelin LA, Moore I (1995) The plant Golgi apparatus: structure,functional organization and trafficking mechanisms. Annu RevPlant Physiol Plant Mol Biol 46: 261–288
Ueda T, Anai T, Tsukaya H, Hirata A, Uchimiya H (1996) Char-acterization and subcellular localization of a small GTP-bindingprotein (ARA-4) from Arabidopsis: conditional expression underthe control of the promoter of the gene for heat-shock proteinHSP81–1. Mol Gen Genet 250: 533–539
Waldron KW, Brett CT (1985) Interaction of enzymes involved incell wall heteropolysaccharide biosynthesis. In CT Brett, JR Hill-man, eds, Biochemistry of Plant Cell Walls, SEB Seminar Series.Cambridge University Press, Cambridge, UK, pp 79–97
White AR, Xin Y, Pezeshik V (1993) Xyloglucan glucosyltrans-ferase in Golgi membranes from Pisum sativum (pea). Biochem J294: 231–238
Zablackis E, Huang J, Muller B, Darvill AG, Albersheim P (1995)Characterization of the cell wall polysaccharides of Arabidopsisthaliana leaves. Plant Physiol 107: 1129–1138
Reversibly Autoglycosylated Arabidopsis Protein 1349
www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 1998 American Society of Plant Biologists. All rights reserved.




















