
Expression of glycosylated haloalkane dehalogenase LinB in Pichia pastoris

Arabidopsis Reversibly Glycosylated Polypeptides1 and 2 Are Essential for Pollen Development1[W]
Georgia Drakakaki, Olga Zabotina, Ivan Delgado2, Stephanie Robert, Kenneth Keegstra,and Natasha Raikhel*
The Center for Plant Cell Biology and Department of Botany and Plant Sciences, University of California,Riverside, California 92521 (G.D., O.Z., I.D., S.R., N.R.); and Department of Energy Plant Research Laboratory,Michigan State University, East Lansing, Michigan 48824 (K.K.)
Reversibly glycosylated polypeptides (RGPs) have been implicated in polysaccharide biosynthesis. To date, to our knowledge,no direct evidence exists for the involvement of RGPs in a particular biochemical process. The Arabidopsis (Arabidopsisthaliana) genome contains five RGP genes out of which RGP1 and RGP2 share the highest sequence identity. We characterizedthe native expression pattern of Arabidopsis RGP1 and RGP2 and used reverse genetics to investigate their respective functions.Although both genes are ubiquitously expressed, the highest levels are observed in actively growing tissues and in maturepollen, in particular. RGPs showed cytoplasmic and transient association with Golgi. In addition, both proteins colocalized in thesame compartments and coimmunoprecipitated from plant cell extracts. Single-gene disruptions did not show any obviousmorphological defects under greenhouse conditions, whereas the double-insertion mutant could not be recovered. We presentevidence that the double mutant is lethal and demonstrate the critical role of RGPs, particularly in pollen development. Detailedanalysis demonstrated that mutant pollen development is associated with abnormally enlarged vacuoles and a poorly definedinner cell wall layer, which consequently results in disintegration of the pollen structure during pollen mitosis I. Taken together,our results indicate that RGP1 and RGP2 are required during microspore development and pollen mitosis, either affecting celldivision and/or vacuolar integrity.
Reversibly glycosylated polypeptides (RGPs) havebeen implicated in polysaccharide biosynthesis (Dhuggaet al., 1991, 1997) because they are localized to theGolgi apparatus (Dhugga et al., 1997) and are ableto react with UDP-Glc, UDP-Xyl, and UDP-Gal. TheRGPs of a number of species have been studied: Arabi-dopsis (Arabidopsis thaliana; Delgado et al., 1998), maize(Zea mays; Rothschild and Tandecarz, 1994; Sagi et al.,2005), potato (Solanum tuberosum; Bocca et al., 1999),wheat (Triticum aestivum) and rice (Orzya sativa;Langeveld et al., 2002), cotton (Gossypium hirsutum;Zhao et al., 2001), and tomato (Lycopersicon esculentum;Selth et al., 2006). The RGPs of wheat, rice, and potatohave been implicated in the initiation of starch bio-synthesis (Bocca et al., 1997; Langeveld et al., 2002),although there is no direct evidence that confirms thishypothesis. However, the fact that none of the RGP
sequences contain a predicted plastid-targeting signalrenders this hypothesis unlikely. The existence of RGPsin both monocots and dicots and their absence in non-plant species indicates a plant-specific function.
RGPs have been detected in both the membrane andsoluble fractions of most of the above-mentionedspecies. Furthermore, experiments conducted in maizeand transgenic tobacco (Nicotiana tabacum) overexpress-ing Arabidopsis RGPs have shown them to be associ-ated with the plasmodesmata in a Golgi-dependentmanner (Sagi et al., 2005).
Although RGP was initially purified from pea(Pisum sativum) hypocotyls (Dhugga et al., 1991) andmaize upper mesocotyls (Sagi et al., 2005), RGP1 wasalso found to be highly expressed in roots and in cellcultures in Arabidopsis (Delgado et al., 1998). Stablytransformed tobacco plants containing a cotton RGPpromoterTb-glucuronidase fusion construct showedhigh expression in the roots, styles, and stigmas (Wuet al., 2006).
Data from transcriptome and proteomic analyseshave shown that genes related to cell wall biosynthesisand regulation are highly expressed in Arabidopsispollen (Becker et al., 2003; Honys and Twell, 2003;Holmes-Davis et al., 2005; Noir et al., 2005; Pina et al.,2005). This finding underscores the importance ofthe cell wall in the growth process of the pollen tube.Recent genetic evidence emphasizes the role of callosesynthases and pectin methyl esterases (PMEs) for pol-len plant development and fertility (Bosch et al., 2005;Bosch and Hepler, 2005; Enns et al., 2005; Jiang et al.,
1 This work was supported by the National Science Foundation(plant genome grant no. DBI–0211797).
2 Present address: Identigene, Inc., 5615 Kirby Dr., Ste. 800,Houston, TX 77005–2451.
* Corresponding author; e-mail [email protected]; fax 951–827–2155.
The author responsible for distribution of materials integral to thefindings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Natasha Raikhel ([email protected]).
[W] The online version of this article contains Web-only data.www.plantphysiol.org/cgi/doi/10.1104/pp.106.086363
1480 Plant Physiology, December 2006, Vol. 142, pp. 1480–1492, www.plantphysiol.org � 2006 American Society of Plant Biologists www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

2005; Nishikawa et al., 2005). Other cell wall-relatedenzymes that have been shown to be important forpollen development include AtCSLA7 and UDP-sugarpyrophosphorylase (Goubet et al., 2003; Schnurret al., 2006). Several monosaccharide transporters ex-hibiting high expression during various stages of pol-len development have been identified (Truernit et al.,1996, 1999; Schneidereit et al., 2003, 2005; Scholz-Starkeet al., 2003). Interestingly, RGP1 and RGP2 were alsodetected in Arabidopsis and rice pollen proteomes(Holmes-Davis et al., 2005; Noir et al., 2005; Dai et al.,2006).
The Arabidopsis genome contains five RGP genes(Girke et al., 2004). RGP1 (At3g02230) and RGP2(At5g15650) have high sequence identity and similargene structures and thus may have arisen through agene duplication event (Blanc et al., 2000). To begin tounderstand the biological roles of RGPs, we createdtranslational fusions of Arabidopsis RGP1 and RGP2and determined their expression patterns and subcel-lular localization. We also undertook a genetic approachto determine the respective functions of AtRGP1 andAtRGP2. We demonstrated that a double-mutant crossbetween rgp1-1 and rgp2-1 resulted in lethality. Furtheranalysis demonstrated the critical role of RGPs inpollen development.
RESULTS
The Arabidopsis RGP Gene Family
The Arabidopsis genome encodes five RGPs (Fig. 1).AtRGP1 and AtRGP2 share 93% identity at the amino
acid level. RGP1 is 80% identical to RGP3 and 74%identical to RGP4. All RGPs are at least 43% identicalto each other. Arabidopsis RGP1 was previously char-acterized and shown to reversibly autoglycosylatewith UDP-Glc, UDP-Xyl, or UDP-Gal as substrates(Delgado et al., 1998). Because we have previouslystudied RGP1 (Delgado et al., 1998), and because amongthe Arabidopsis RGP genes it has the highest similarityto RGP2 (Girke et al., 2004), we decided to continueour studies on these two genes.
RGP1 and RGP2 Are Strongly Expressed in ActivelyGrowing Tissues as Well as Pollen
To examine the potential tissue specificity of RGP1and RGP2 expression, we utilized the fluorescent tag-ging of full-length proteins technique (Tian et al., 2004).In this approach, which has been shown to faithfullyreproduce the native levels and expression patterns, aswell as subcellular localizations of the fluorescentlytagged gene products (Tian et al., 2004), RGP1 andRGP2 fused to yellow fluorescent protein (YFP) wereexpressed from their native promoters in both thepresence and absence of 35S enhancers. The 35S en-hancers have been shown to increase expression levelsof YFP fusion proteins, but not to alter their subcellularlocalizations (Tian et al., 2004). Characterization ofspatial expression patterns detected RGP1-YFP andRGP2-YFP in all tissues, but both were most stronglyexpressed in actively growing tissues. Figure 2, A andD, shows that RGP1-YFP was expressed predomi-nantly in the root tip (Fig. 2A) and the apical meristem(Fig. 2D) of young seedlings. Strong expression ofRGP1-YFP was also observed in lateral root primordia
Figure 1. Phylogenetic analysis of the RGP familyin Arabidopsis. The alignments are used to calcu-late phylogenetic trees with the PHYLIP package(Felsenstein, 2005), as described in the Cell WallNavigator database (Girke et al., 2004). Scalerepresents 10 substitutions per 100 amino acids.Bootstrap values for 100 replications are given atthe branch points.
Reversibly Glycosylated Poplypeptides in Pollen Development
Plant Physiol. Vol. 142, 2006 1481 www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

(Fig. 2C). RGP2-YFP, with expression augmented bythe 35S enhancer, was expressed in the same tissues(Fig. 2B). More interestingly, during later developmen-tal stages, both RGP fusions were highly expressed inpollen (Figs. 2E and 3, A and D). These data are inagreement with in silico microarray analyses that showessentially the same expression patterns, with strongexpression of AtRGP1 and AtRGP2 in roots, vegetativeshoots, flowers, and mature pollen (http://csbdb.mpimp-golm.mpg.de). Microarray data from both thepublic databases and RGP1-YFP show that the highestexpression levels are in tricellular pollen (Supplemen-tal Fig. S1). The RGP1 transcript and protein havepreviously been detected at high levels in suspensioncultures and roots (Delgado et al., 1998).
Confocal laser-scanning microscopy (CLSM) wasused to examine the subcellular localization of theRGP-YFP fusions in various Arabidopsis organs.These studies revealed cytoplasmic (especially inroots) and punctate labeling for both the RGP1 andRGP2 fusion proteins (Fig. 3). The RGP2 fusion protein(with 35S enhancer) was observed in leaf epidermalcells as small fluorescent foci throughout the cyto-plasm (Supplemental Video S1), indicating a charac-teristic pattern of association with Golgi. The punctatepatterns were very pronounced and dynamic in grow-ing pollen for both RGPs (Fig. 3, A and D; Supple-mental Video S2). Occasionally, we observed largerfluorescing foci in RGP2-YFP leaves and hypocotyls,which was not indicative of a known pattern for aspecific compartment (data not shown).
Arabidopsis RGPs Exhibit Cytoplasmic and
Golgi-Associated Localization
Previous reports have shown that a significantamount of RGPs were present in soluble cellularfractions (Dhugga et al., 1991, 1997; Bocca et al., 1997,1999), indicating that RGPs can be localized to thecytoplasm. However, it was assumed that most ofthe RGP present in the cytoplasm was released fromthe Golgi membranes or from plasmodesmata, respec-tively (Dhugga et al., 1997; Sagi et al., 2005). Consistentwith the biochemical evidence (Dhugga et al., 1991;Delgado et al., 1998), our results clearly demonstratethat the fusion protein was present in the cytoplasm.
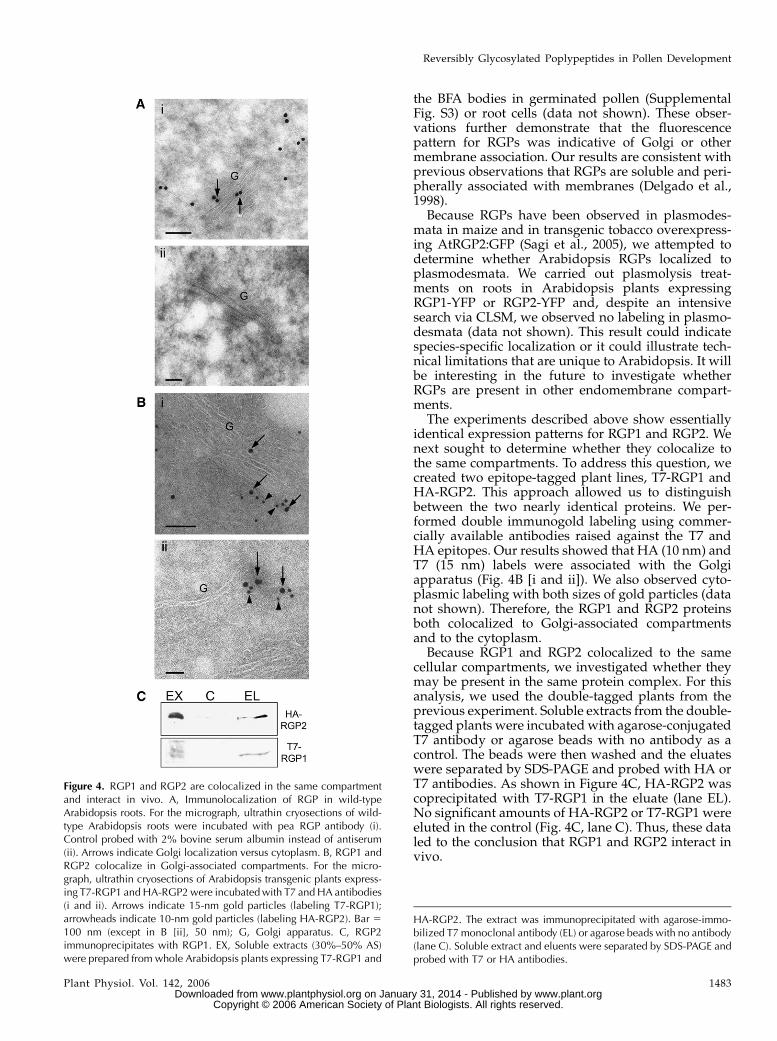
To validate the subcellular localization of Arabidop-sis RGP1 and RGP2 in both the cytoplasm and Golgi,we performed electron microscopy immunolocaliza-tion on wild-type Arabidopsis roots. The pea RGPantibody was used for its high specificity and lowbackground signal on western blots (Dhugga et al.,1997). Our analysis showed that Arabidopsis RGPslocalized to both Golgi and cytoplasm (Fig. 4A). Thesedata are in agreement with the fluorescence localiza-tion of RGP1-YFP and RGP2-YFP.
Subcellular fractionation of fluorescence-taggedplants was used to verify the Golgi association of theYFP fusion proteins. Both fusion proteins were de-tected in the Golgi/microsomal-enriched fraction (Sup-plemental Fig. S2).
Brefeldin A (BFA), a trafficking inhibitor that dis-rupts the Golgi apparatus and results in aggregates ofGolgi membranes (for review, see Nebenfuhr et al.,2002), was used to further examine the association ofthe RGP fusions with this compartment. Germinatedpollen (Supplemental Fig. S3) or root cells from theelongation zone (data not shown) that were treatedwith BFA displayed the characteristic fluorescent ag-gregates referred to as BFA bodies. The endocyticmarker FM4-64 (Bolte et al., 2004) also colocalized with
Figure 2. Expression patterns of RGP1 and RGP2. Expression pattern ofRGP1-YFP (A, C, D, and E) and RGP2-YFP (B) are primarily observed inthe apical meristem and root tip in 6-d-old seedlings. C, High expres-sion of RGP1-YFP occurred in lateral root primordia (C, inset). E,Expression pattern of RGP1-YFP in flowers is strongly localized inanthers (in mature pollen). The same pattern was observed for RGP2-YFP (data not shown). Bright field (A, B, and E, inset). YFP filter (A, B, C,and E). D, Overlay of single confocal images of YFP filter (yellow) andred autofluoresence (red). Bar 5 400 mm (A, B, and D); 80 mm (Cand E).
Figure 3. Subcellular localization of RGP1-YFP and RGP2-YFP. A,RGP1-YFP in pollen tube, apical meristem (B), and root tip (C).Subcellular localization of RGP2-YFP (with 35S enhancers) in pollentube (D), leaf (E), and root tip (F). All images are single confocalsections. Bar 5 20 mm.
Drakakaki et al.
1482 Plant Physiol. Vol. 142, 2006 www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.
the BFA bodies in germinated pollen (SupplementalFig. S3) or root cells (data not shown). These obser-vations further demonstrate that the fluorescencepattern for RGPs was indicative of Golgi or othermembrane association. Our results are consistent withprevious observations that RGPs are soluble and peri-pherally associated with membranes (Delgado et al.,1998).
Because RGPs have been observed in plasmodes-mata in maize and in transgenic tobacco overexpress-ing AtRGP2:GFP (Sagi et al., 2005), we attempted todetermine whether Arabidopsis RGPs localized toplasmodesmata. We carried out plasmolysis treat-ments on roots in Arabidopsis plants expressingRGP1-YFP or RGP2-YFP and, despite an intensivesearch via CLSM, we observed no labeling in plasmo-desmata (data not shown). This result could indicatespecies-specific localization or it could illustrate tech-nical limitations that are unique to Arabidopsis. It willbe interesting in the future to investigate whetherRGPs are present in other endomembrane compart-ments.
The experiments described above show essentiallyidentical expression patterns for RGP1 and RGP2. Wenext sought to determine whether they colocalize tothe same compartments. To address this question, wecreated two epitope-tagged plant lines, T7-RGP1 andHA-RGP2. This approach allowed us to distinguishbetween the two nearly identical proteins. We per-formed double immunogold labeling using commer-cially available antibodies raised against the T7 andHA epitopes. Our results showed that HA (10 nm) andT7 (15 nm) labels were associated with the Golgiapparatus (Fig. 4B [i and ii]). We also observed cyto-plasmic labeling with both sizes of gold particles (datanot shown). Therefore, the RGP1 and RGP2 proteinsboth colocalized to Golgi-associated compartmentsand to the cytoplasm.
Because RGP1 and RGP2 colocalized to the samecellular compartments, we investigated whether theymay be present in the same protein complex. For thisanalysis, we used the double-tagged plants from theprevious experiment. Soluble extracts from the double-tagged plants were incubated with agarose-conjugatedT7 antibody or agarose beads with no antibody as acontrol. The beads were then washed and the eluateswere separated by SDS-PAGE and probed with HA orT7 antibodies. As shown in Figure 4C, HA-RGP2 wascoprecipitated with T7-RGP1 in the eluate (lane EL).No significant amounts of HA-RGP2 or T7-RGP1 wereeluted in the control (Fig. 4C, lane C). Thus, these dataled to the conclusion that RGP1 and RGP2 interact invivo.
Figure 4. RGP1 and RGP2 are colocalized in the same compartmentand interact in vivo. A, Immunolocalization of RGP in wild-typeArabidopsis roots. For the micrograph, ultrathin cryosections of wild-type Arabidopsis roots were incubated with pea RGP antibody (i).Control probed with 2% bovine serum albumin instead of antiserum(ii). Arrows indicate Golgi localization versus cytoplasm. B, RGP1 andRGP2 colocalize in Golgi-associated compartments. For the micro-graph, ultrathin cryosections of Arabidopsis transgenic plants express-ing T7-RGP1 and HA-RGP2 were incubated with T7 and HA antibodies(i and ii). Arrows indicate 15-nm gold particles (labeling T7-RGP1);arrowheads indicate 10-nm gold particles (labeling HA-RGP2). Bar 5
100 nm (except in B [ii], 50 nm); G, Golgi apparatus. C, RGP2immunoprecipitates with RGP1. EX, Soluble extracts (30%–50% AS)were prepared from whole Arabidopsis plants expressing T7-RGP1 and
HA-RGP2. The extract was immunoprecipitated with agarose-immo-bilized T7 monoclonal antibody (EL) or agarose beads with no antibody(lane C). Soluble extract and eluents were separated by SDS-PAGE andprobed with T7 or HA antibodies.
Reversibly Glycosylated Poplypeptides in Pollen Development
Plant Physiol. Vol. 142, 2006 1483 www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

Genetic Analysis of rgp1 and rgp2 and
Double-Mutant Studies
We took a reverse-genetics approach to understandthe functions of the RGP1 and RGP2 proteins. Mutantplant lines with T-DNA insertions in each of the geneswere obtained from different collections. The rgp1-1line (Gabi_652F12) contained an insert in the firstintron of the RGP1 gene, approximately 763 bp down-stream from the start codon (Fig. 5A [i]). An rgp2-1mutant was obtained from the Salk collection (SALK_132152); it contained a T-DNA insertion in the lastexon of RGP2, approximately 150 nucleotides up-stream of the stop codon (Fig. 5A [ii]). As shown inFigure 5B, reverse transcription (RT)-PCR of tran-scripts isolated from homozygous rgp1 or rgp2 plantsshowed no mRNAs for RGP1 or RGP2, respectively.We carried out RT-PCR of the Arabidopsis UBIQUI-TIN10 gene as a control (Fig. 5B). Under normalgrowth conditions, we did not observe any obviousphenotypes associated with the RGP1 or RGP2 T-DNAinsertions (Fig. 5C).
To address the possibility of genetic redundancy, wecrossed the two single mutants to create a doublemutant. We determined the zygosity of the RGP1 andRGP2 genes at their respective loci by genotyping 100F2 progeny using PCR-based markers. The results ofthe genotyping experiments are reported in Table I. Wefound no homozygous double mutants among the 100F2 progeny. Furthermore, we detected fewer rgp1/rgp1RGP2/rgp2, and no RGP1/rgp1 rgp2/rgp2 plants. Inan independent experiment using 303 F2 progeny, wecame to the same conclusion, namely, no homozygousdouble mutant was identified (Supplemental TableS1). In this case, plants that contained only one func-tional copy of RGP1 (RGP1/rgp1 rgp2/rgp2) were iden-tified; however, they did not survive after transfer tosoil. We also examined the seed set from siliquesremoved from the F2 plants shown in Table I. Seedsfrom the selfing of wild type (data not shown) or thetwo single-mutant plants rarely aborted (Fig. 5D).However, siliques isolated from selfing of doubleheterozygous plants (RGP1/rgp1 RGP2/rgp2) or plantswith only one functional copy of RGP2 (rgp1/rgp1RGP2/rgp2) had vacant spaces (Fig. 5D). We did notobserve aborted embryos or shriveled seeds. There-fore, analysis of the F2 population strongly supportsthe conclusion that the rgp1 rgp2 double mutant islethal.
The Double-Mutant Defect Occurs in Pollen and ImpactsPollen Morphology
To examine more closely the nature of the lethalphenotype, we created reciprocal crosses of rgp1/rgp1RGP2/rgp2 and the wild-type Columbia parent. Whenwild-type pollen was used to fertilize rgp1/rgp1RGP2/rgp2 plants, we observed a reduction in thenumber of progeny that carried both mutant alleles,suggesting a reduction of rgp1 rgp2 transmission
through the female gametophytes (Table II). Moreinterestingly, when pollen isolated from plants thatcarried one copy of RGP2 (rgp1/rgp1 RGP2/rgp2) wasused to fertilize wild-type plants, we were able todetect only progeny with a T-DNA insert in RGP1. The
Figure 5. rgp1-1 and rgp2-1 mutants and their genetic cross. A, T-DNAinsertion in RGP1 and RGP2. T-DNA insertion is located in the firstintron in rgp1-1 (i). T-DNA insertion is located in the last exon in rgp2-1(ii). Arrows indicate the position of the primers used for RT-PCR. B, RT-PCR of RGP1 and RGP2. Total RNAs from seedlings of UBIQUITIN 10using the same templates with gene-specific primers are shown at thebottom. C, None of the mutant plants of RGP1 or RGP2 exhibited anyobvious differences under normal growth conditions as compared towild type. D, Effect of rgp1-1 and/or rgp2-1 mutation on seed develop-ment. Whole-mount images of the mature silique cleared by NaOHsolution. i, rgp1/rgp1; ii, rgp2/rgp2; iii, RGP1/rgp1 RGP2/rgp2; iv, rgp1/rgp1 RGP2/rgp2. Arrowheads indicate unnatural intervals. Bar 5 1 mm.
Drakakaki et al.
1484 Plant Physiol. Vol. 142, 2006 www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

RGP1/rgp1 RGP2/rgp2 genotype was not detected(Table II). These data indicate that pollen carryingboth insertions (rgp1 rgp2) is not viable. Thus, failureduring gametogenesis in rgp1 rgp2 pollen is the basisfor the lethality of the double mutant.
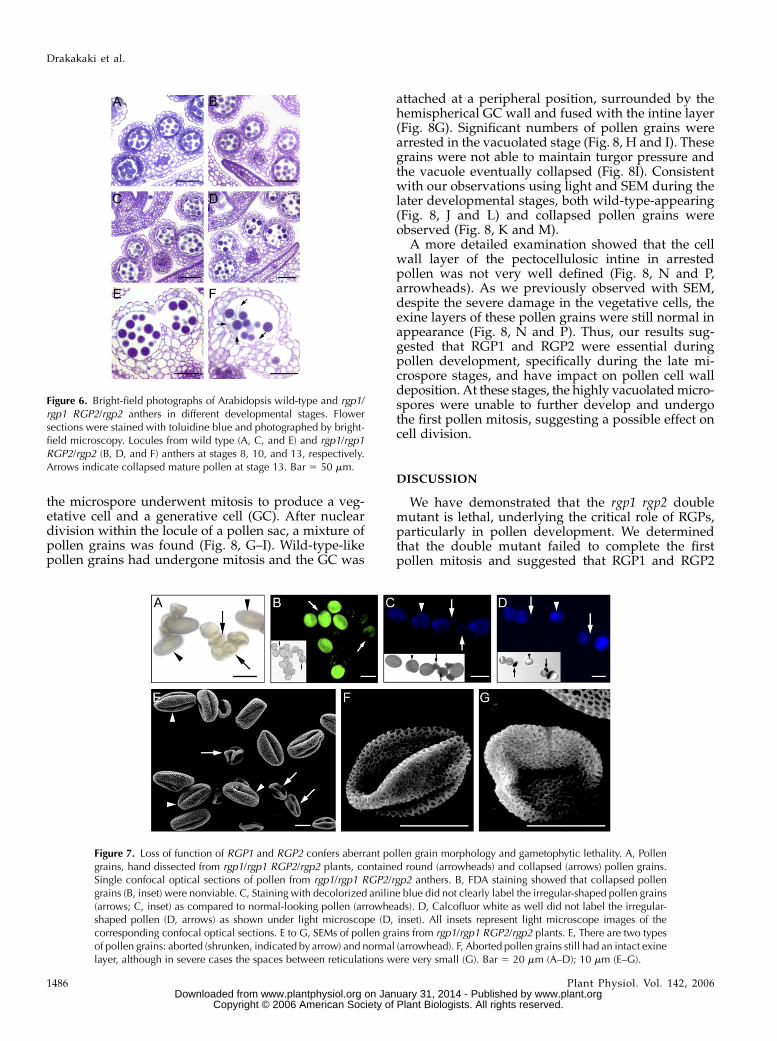
Because reciprocal crosses indicate that the malegametophyte is not viable in the homozygous rgp1 rgp2double mutant, we closely examined the developmentof anthers in wild-type and rgp1/rgp1 RGP2/rgp2mutants. We examined toluidine blue-stained sectionsof anthers at various stages of development using lightmicroscopy. Anther development and microspore for-mation follow a well-established pattern in dicotyle-donous plants (Sanders et al., 1999) and are separatedinto 14 stages. No morphological differences wereobserved between wild type and rgp1/rgp1 RGP2/rgp2 anthers at stages 8 to 10 (Fig. 6, A, B, C, and D,respectively). At this stage, meiosis was already com-plete, the callose walls surrounding the tetrads haddegenerated, and microspores had been released. Fol-lowing this stage, individual microspores further de-veloped into pollen grains. During the later stages(stages 11–14), numerous pollen grains from the rgp1/rgp1 RGP2/rgp2 anthers were not completely devel-oped (Figs. 6F and 7A). Compared to mature wild-typepollen grains, many of those in mutant anthers werecollapsed in appearance (Fig. 6, E and F, respectively).Taking into account the data from the reciprocal
crosses, we concluded that the collapsed pollen grains(Figs. 6F and 7A) harbored mutations in both RGP1 andRGP2.
Mature pollen was released at various stages ofdevelopment by hand dissection and further analyzed.We determined pollen viability by using fluorescencemicroscopy analysis after fluorescein diacetate (FDA)staining. Malformed pollen grains did not stain withFDA (Fig. 7B, arrows) and were incapable of germi-nation in vitro, suggesting that they were not viable.We further investigated the small and flattened pollengrains for their nuclear constitution by staining with4#,6#-diamidino-2-phenylindole. In normal-looking pol-len, the vegetative nucleus and the two generativenuclei were clearly distinguishable (Supplemental Fig.S4, arrowheads). In the malformed pollen, only diffusestaining was observed (Supplemental Fig. S4, arrows).Staining with calcofluor white (for cellulose) or anilineblue (for callose) failed to label the collapsed pollengrains (Fig. 7, C and D, arrows). Pollen released fromsingle-mutant anthers did not show any differences inmorphology or staining compared to that of wild-typeones (data not shown).
Using scanning electron microscopy (SEM), we ex-amined more closely pollen from wild type or plantswith only one functional copy of RGP2 (rgp1/rgp1RGP2/rgp2). Although some pollen grains appearednormal (Fig. 7E, arrowheads), significant numbers ofabnormal and collapsed pollen grains were detected inthe rgp1/rgp1 RGP2/rgp2 line (Fig. 7, E [arrows]–G),which is consistent with light microscopic observa-tions (Figs. 6F and 7A). Although mutant pollen grainswere smaller in size and less robust, the basic structureof the exine layer was still present (Fig. 7, F and G).
Pollen Grains with One Functional Copy of RGP2Are Arrested during Pollen Mitosis I
Cross sections of rgp1/rgp1 RGP2/rgp2 were fur-ther analyzed by transmission electron microscopy(TEM). Microspores did not show any structural ab-normalities (Fig. 8, A–C). Later in microspore devel-opment, the vacuoles fused into a single large vacuoleduring microspore expansion and nuclear migration(Fig. 8, D–F). During this stage, some of the spores hadunusually large vacuoles (Fig. 8, E and F). Thereafter,
Table I. Summary of the lethality of rgp1-1 and rgp2-1 andmutant progeny
Percentage of genotypes (by PCR) observed in the F2 populationfollowing the genetic cross of rgp1-1 3 rgp2-1. The percentage of theexpected genotypes following the genetic cross of rgp1 3 rgp2 in F2 islisted.
Genotype No. Expected Observed
RGP1/rgp1 RGP2/rgp2 35 25% 35%rgp1/rgp1 RGP2/rgp2 2 12.5% 2%RGP1/rgp1 rgp2/rgp2 0 12.5% 0%rgp1/rgp1 rgp2/rgp2 0 6.25% 0%rgp1/rgp1 RGP2/RGP2 17 6.25% 17%RGP1/rgp1 RGP2/RGP2 12 12.5% 12%RGP1/RGP1 rgp2/rgp2 18 6.25% 18%RGP1/RGP1 RGP2/rgp2 10 12.5% 10%RGP1/RGP1 RGP2/RGP2 6 6.25% 6%Total 100
Table II. Genetic analysis of rgp1/rgp1 RGP2/rgp2 gametophytes
Individuals inheriting the rgp1 or the rgp2 mutation were scored by PCR and then subjected to a X2 test. P, Statistical significance; n, sample size.
Classes Observed (o) Expected (e) Deviation d (o – e) d 2 X2(d 2/e)
Back cross of wild type (female) 3 rgp1/1rgp1 RGP2/rgp2 (male)a
RGP1/rgp1 RGP2/RGP2 100% (42) 50% (21) 21 441 21RGP1/rgp1 RGP2/rgp2 0% (0) 50% (21) 221 441 21
Back cross of rgp1/rgp1 RGP2/rgp2 (female) 3 wild type (male)b
RGP1/rgp1 RGP2/RGP2 62.10% (59) 50% (47.5) 11.5 132.25 2.784RGP1/rgp1 RGP2/rgp2 37.89% (36) 50% (47.5) 211.5 132.25 2.784
an 542, X2 5 42, P , 0.001. bn 5 95, X 2 5 5.568, 0.01,P , 0.02.
Reversibly Glycosylated Poplypeptides in Pollen Development
Plant Physiol. Vol. 142, 2006 1485 www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.
the microspore underwent mitosis to produce a veg-etative cell and a generative cell (GC). After nucleardivision within the locule of a pollen sac, a mixture ofpollen grains was found (Fig. 8, G–I). Wild-type-likepollen grains had undergone mitosis and the GC was
attached at a peripheral position, surrounded by thehemispherical GC wall and fused with the intine layer(Fig. 8G). Significant numbers of pollen grains werearrested in the vacuolated stage (Fig. 8, H and I). Thesegrains were not able to maintain turgor pressure andthe vacuole eventually collapsed (Fig. 8I). Consistentwith our observations using light and SEM during thelater developmental stages, both wild-type-appearing(Fig. 8, J and L) and collapsed pollen grains wereobserved (Fig. 8, K and M).
A more detailed examination showed that the cellwall layer of the pectocellulosic intine in arrestedpollen was not very well defined (Fig. 8, N and P,arrowheads). As we previously observed with SEM,despite the severe damage in the vegetative cells, theexine layers of these pollen grains were still normal inappearance (Fig. 8, N and P). Thus, our results sug-gested that RGP1 and RGP2 were essential duringpollen development, specifically during the late mi-crospore stages, and have impact on pollen cell walldeposition. At these stages, the highly vacuolated micro-spores were unable to further develop and undergothe first pollen mitosis, suggesting a possible effect oncell division.
DISCUSSION
We have demonstrated that the rgp1 rgp2 doublemutant is lethal, underlying the critical role of RGPs,particularly in pollen development. We determinedthat the double mutant failed to complete the firstpollen mitosis and suggested that RGP1 and RGP2
Figure 6. Bright-field photographs of Arabidopsis wild-type and rgp1/rgp1 RGP2/rgp2 anthers in different developmental stages. Flowersections were stained with toluidine blue and photographed by bright-field microscopy. Locules from wild type (A, C, and E) and rgp1/rgp1RGP2/rgp2 (B, D, and F) anthers at stages 8, 10, and 13, respectively.Arrows indicate collapsed mature pollen at stage 13. Bar 5 50 mm.
Figure 7. Loss of function of RGP1 and RGP2 confers aberrant pollen grain morphology and gametophytic lethality. A, Pollengrains, hand dissected from rgp1/rgp1 RGP2/rgp2 plants, contained round (arrowheads) and collapsed (arrows) pollen grains.Single confocal optical sections of pollen from rgp1/rgp1 RGP2/rgp2 anthers. B, FDA staining showed that collapsed pollengrains (B, inset) were nonviable. C, Staining with decolorized aniline blue did not clearly label the irregular-shaped pollen grains(arrows; C, inset) as compared to normal-looking pollen (arrowheads). D, Calcofluor white as well did not label the irregular-shaped pollen (D, arrows) as shown under light microscope (D, inset). All insets represent light microscope images of thecorresponding confocal optical sections. E to G, SEMs of pollen grains from rgp1/rgp1 RGP2/rgp2 plants. E, There are two typesof pollen grains: aborted (shrunken, indicated by arrow) and normal (arrowhead). F, Aborted pollen grains still had an intact exinelayer, although in severe cases the spaces between reticulations were very small (G). Bar 5 20 mm (A–D); 10 mm (E–G).
Drakakaki et al.
1486 Plant Physiol. Vol. 142, 2006 www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.
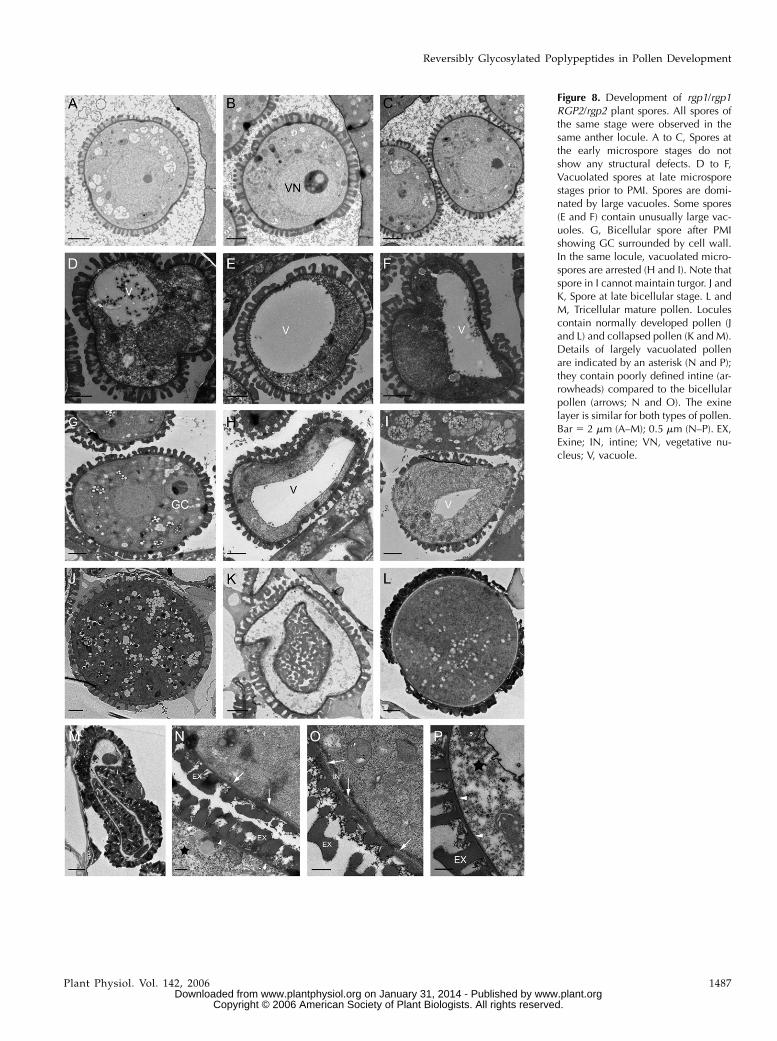
Figure 8. Development of rgp1/rgp1RGP2/rgp2 plant spores. All spores ofthe same stage were observed in thesame anther locule. A to C, Spores atthe early microspore stages do notshow any structural defects. D to F,Vacuolated spores at late microsporestages prior to PMI. Spores are domi-nated by large vacuoles. Some spores(E and F) contain unusually large vac-uoles. G, Bicellular spore after PMIshowing GC surrounded by cell wall.In the same locule, vacuolated micro-spores are arrested (H and I). Note thatspore in I cannot maintain turgor. J andK, Spore at late bicellular stage. L andM, Tricellular mature pollen. Loculescontain normally developed pollen (Jand L) and collapsed pollen (K and M).Details of largely vacuolated pollenare indicated by an asterisk (N and P);they contain poorly defined intine (ar-rowheads) compared to the bicellularpollen (arrows; N and O). The exinelayer is similar for both types of pollen.Bar 5 2 mm (A–M); 0.5 mm (N–P). EX,Exine; IN, intine; VN, vegetative nu-cleus; V, vacuole.
Reversibly Glycosylated Poplypeptides in Pollen Development
Plant Physiol. Vol. 142, 2006 1487 www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

affect cell division and/or vacuolar integrity. We dem-onstrated in detail that RGPs are essential for pollendevelopment and possibly for other stages in the lifecycle of plants.
RGP1 and RGP2 Are Functionally Redundant
The Arabidopsis genome encodes five RGPs (Girkeet al., 2004), one of which, RGP1, was previously char-acterized (Delgado et al., 1998). Plant RGPs are dividedinto two distinct phylogenetic classes (Langeveld et al.,2002); class 1 contains AtRGP1 to AtRGP4, as well asseveral other monocot and dicot RGPs, and class 2 islimited to RGPs from wheat and rice, as well as theremaining Arabidopsis paralog, AtRGP5. The highlyconserved sequences of AtRGP1 and AtRGP2 suggestpartial functional redundancy. Indeed, our geneticanalysis confirms this hypothesis; single knockoutsare viable and the double knockout is lethal.
Given that RGP1 to RGP4 are in the same class, itcould still suggest that there may be some functionalredundancy among the four family members. How-ever, because the rgp1 rgp2 double mutant is lethal, weposit that the RGP3 and RGP4 proteins cannot com-pensate for their loss, at least during pollen develop-ment. If true, this deficiency may reflect true functionaldivergence within the family or it may be caused bydifferent expression patterns or levels of expressionof the other RGP genes. Ectopic expression of eitherRGP3 or RGP4 in the rgp1 rgp2 mutant backgroundand determining whether they are sufficient to rescuethe mutant phenotype may help distinguish betweenthese possibilities. We propose that, in both singlemutants (rgp1 or rgp2), sufficient RGP levels remain sothat active RGP forms may function during pollendevelopment.
Previous studies have shown that wheat RGP1 andRGP2 can assemble to form either homooligomers orheterooligomers with a predicted molecular mass of230 kD (Langeveld et al., 2002). Potato RGP proteinsare also predicted to form a complex, although, in thiscase, the architecture is thought to be pentameric orhexameric (Bocca et al., 1997). In Arabidopsis, thecolocalization of RGP1 and RGP2 and coimmunopre-cipitation provide evidence for their interaction. Be-cause RGP proteins are thought to be more active asmembers of complexes (Langeveld et al., 2002), weanticipate that the Arabidopsis RGPs in a complexwould show greater autoglycosylation efficiency thanthose present as monomers. It is also possible that theArabidopsis RGP complex contains other RGP familymembers, a hypothesis that we are currently investi-gating.
RGP1 and RGP2 Expression Is Associated withActive Growth
Our results showed that RGP1 and RGP2 are highlyexpressed in actively growing tissues, which requiresynthesis of large amounts of cell wall components.
This finding supports the hypothesis that RGPs areinvolved in cell wall biosynthesis and therefore in cellgrowth and division. In agreement with this result,RGPs have previously been found in growing tissues,such as the mesocotyl of pea and maize (Dhugga et al.,1991, 1997; Sagi et al., 2005), Arabidopsis (Delgadoet al., 1998), and cotton (Zhao et al., 2001).
The high expression levels of RGP1 and RGP2 inpollen suggest an important function during pollendevelopment. In agreement with the expression pat-terns, our genetic analysis showed that rgp1 rpg2double-mutant pollen, derived from plants withone functional copy of RGP2, has severe defects indevelopment.
The cotton RGP promoterTb-glucuronidase fusionwas expressed strongly in the roots, styles, and stigmaof transgenic tobacco plants, but less strongly in pol-len, indicating species-specific expression patterns(Wu et al., 2006). RGPs are also strongly expressed inpollinated melon (Cucumis melo; Nagasawa et al., 2005)and in Medicago seeds (stage 18 d after pollination;Gallardo et al., 2003), providing further evidence for acrucial role after pollination.
RGPs are present both in the cytoplasm (wheremany aspects of sugar nucleotide synthesis and metab-olism occur) and at the surface of the Golgi compart-ments (where matrix polysaccharides are synthesized).As previously stated, phylogenetic analysis dividesthe RGPs into two classes. Langeveld et al. (2002) havesuggested that the class 1 RGPs (AtRGPs 1–4) mayplay a role in cell wall synthesis and class 2 RGPs(AtRGP5) may be required for starch biosynthesis.Indeed, our localization results are in agreement withthe possible role of RGP1 and RGP2 in cell wall poly-saccharide biosynthesis. Sagi et al. (2005) proposedthat the final destination of RGPs could be the cell wallor plasmodesmata, but we were unable to confirm thishypothesis for Arabidopsis. Continuing research onRGP3, RGP4, and RGP5 is necessary to test whetherthey might function in starch synthesis.
RGP1 and RGP2 Are Crucial for Normal MaleGametophyte Development
Our genetic analysis clearly demonstrates the im-portance of RGPs by illustrating that the rgp1 rgp2double mutant is lethal and suggests that RGP1 andRGP2 are essential, at least for pollen development.Application of cell wall stains to wild type and pollengrains of double mutants with one copy of RGPshowed that the stains failed to label the collapsedrpg1 rgp2 double-mutant pollen. Ultrastructural anal-ysis showed that the double-mutant pollen had ir-regular organization with poorly defined intine(pectocellulosic cell wall layer). Taken together, theseresults suggested that the loss of RGP function affectscell wall formation in the pollen grains of the doublemutant. As previously stated, RGPs have been impli-cated in noncellulosic polysaccharide biosynthesis(Dhugga et al., 1997; Delgado et al., 1998). Because
Drakakaki et al.
1488 Plant Physiol. Vol. 142, 2006 www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

the double mutant is male gametophytically lethal, it isexpected that, in the double mutant pollen, cell wallformation could be affected. It would therefore beinteresting in the future to find out which polysac-charides are affected in the double mutant.
There are numerous examples of male gametophyticmutants that have defects in cell wall formation,particularly in callose biosynthesis and pectin esteri-fication. Mutants defective in callose biosynthesis ex-hibit several gametophytic defects. For example, theArabidopsis callose synthase 5 (calS5) mutant has severedefects in the exine wall, although weaker alleles al-low fertile pollen to be produced (Dong et al., 2005;Nishikawa et al., 2005). The CalS11 (gsl1) and CalS12(gsl5) genes were shown to have partially overlappingroles in the formation of the callose wall that separatesthe microspores within the tetrad and, later, duringpollen grain maturation (Enns et al., 2005). The rgp1rpg2 double mutant formed microspores that arenormal in appearance, but arrested at a postmeioticstage, suggesting that tetrad formation and microspo-rogenesis were not affected in this background. Be-cause there were no obvious defects during tetradformation, or in the exine layer, and because RGP1-YFP and RGP2-YFP were not confined to the calloseplug of the germinated pollen tubes (data not shown),we concluded that RGPs do not play a major or in-dispensable role in callose biosynthesis.
Pectin is another major component of pollen graintube walls and is the major component of the outerfibril cell wall layer (Li et al., 1995). Recent evidencesuggests that PMEs play an important role in pollentube growth. Although there was no apparent pheno-type in pollen grains, mutations in the ArabidopsisVanguard1 gene, which encodes a pollen-specific PME,caused profound retardation of in vivo growth andstructurally unstable pollen tubes when grown invitro (Jiang et al., 2005). YFP tagging of VANGUARD1demonstrated that the fusion protein was distributedthroughout the pollen tube and was localized to thecell wall (Jiang et al., 2005). Similarly, tobacco plants inwhich the tobacco pollen-specific PME (NtPPME1)gene was silenced using an RNAi construct showedsignificantly retarded pollen tube growth in vivo(Bosch and Hepler, 2005). Mutations in the Arabidop-sis pollen-specific PME1 (AtPPME1) gene did notaffect the pollen grains themselves, but compromisedthe morphology and growth rate of pollen tubes (Tianet al., 2006). Although our double-mutant phenotypedoes not resemble those described above, it is possiblethat RGPs have an indirect effect on pectin biosynthe-sis in the pollen cell wall. Because pectin is the majorcomponent of the pollen grain, cell wall defects inpectin composition could have a severe impact onpollen development similar to that observed in thedouble mutant. Specific antibodies (for polysaccha-rides) will be used in the future to determine whichpolysaccharide, or cell wall component, is affected.Additionally, biochemical analysis of mutant pollencould determine the sugar or cell wall composition.
Detailed analysis of rgp1 rgp2 double-mutant pollenat various stages showed that pollen development wasnot affected until microspore release. However, aber-rant pollen grains with enlarged vacuoles were ob-served in midmicrospore stage and they appearedto be collapsed at the bicellular stage, suggesting thatthe mutant was arrested before or during the firstmitotic division. Several mutants in male gameto-phytic development, such as gem-1 (Park et al., 1998),stud (Hulskamp et al., 1997), and tio (Oh et al., 2005),have previously been described to have defects incytokinesis. The gem-1 mutant produces a substantialproportion of microspores that either fail to establish acell plate at pollen mitosis I (PMI) or produce partialand/or irregularly branching cell walls that alter di-vision symmetry (Park et al., 1998; Park and Twell,2001). TIO encodes a FUSED (FU) Ser-Thr proteinkinase ortholog in Arabidopsis and tio mutants pro-duce binucleate pollen grains that arise from the fail-ure of cytokinesis at PMI (Oh et al., 2005). One of thecharacteristics of tio microspores is the incompleteformation of callose walls. In addition, ArabidopsisSTUD/TETRASPORE/NACK2 encodes a kinesin-likeprotein; the corresponding mutants fail to undergocytokinesis after meiosis and large multinucleatepollen grains are formed with aberrant cell walls(Hulskamp et al., 1997; Yang et al., 2003; Tanaka et al.,2004). The phenotypes of the tio and stud mutantsunderscore the importance of phosphorylation duringcytokinesis in male gametophytes. Regulation of selfglycosylation by RGPs may also be mediated by phos-phorylation (Testasecca et al., 2004; G. Drakakaki andN. Raikhel, unpublished data). Because our doublemutant is arrested during PMI, it is tempting tospeculate that RGPs are involved in cell division andperhaps cell plate formation. Considering that RGPsare implicated in polysaccharide biosynthesis, cellplate formation during the first mitotic division couldbe affected. A more detailed analysis of rpg1 rpg2 mu-tant pollen in the early bicellular stage will enable us toexplore this hypothesis.
Recent studies have shown that proteins involved insugar nucleotide metabolism are important duringmale gametophyte development. The presence of amutation in a UDP-sugar pyrophosphorylase (usp-2)resulted in lack of a pectocellulosic intine and degen-eration of cytoplasm in mutant pollen (Schnurr et al.,2006). Like the usp mutant, our rgp1 rgp2 double mu-tant produced pollen that did not remain turgid; theabnormally enlarged vacuoles in this pollen collapsedduring the early bicellular stage. The common featureof the two mutants may be defects in cell wall biosyn-thesis. In the usp mutant, the phenotype may arisebecause of defects in the sugar nucleotide supply. Inthe case of the rgp1 rgp2 double mutant, pollen defectsmay have arisen because of an inability to convertsugar nucleotides into either wall polysaccharides orother carbohydrates. If correct, this interpretation sup-ports the hypothesis that RGPs are involved in cellwall polysaccharide biosynthesis and could explain
Reversibly Glycosylated Poplypeptides in Pollen Development
Plant Physiol. Vol. 142, 2006 1489 www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

the loss of turgor and the defects in the intine cell walllayer in mutant pollen.
Several monosaccharide transporters are also ex-pressed at various stages of pollen maturation (Truernitet al., 1996, 1999; Schneidereit et al., 2003, 2005; Scholz-Starke et al., 2003). In particular, AtSTP2 transporterexpression is confined to the early stages of pollendevelopment. Both AtSTP2 mRNA and protein arefirst observed at the beginning of callose degradationand microspore release from the tetrads and are nolonger detected after the first mitotic division andthe formation of the trinuclear gametophyte (Truernitet al., 1999). Our rgp1 rgp2 double mutant arrestedbefore or during the first mitotic division. It is there-fore tempting to speculate that the loss of RGP1 andRGP2 prevented the downstream utilization of sugarsderived from callose degradation. RGP1 and RGP2may be involved in specific product formation (e.g.polysaccharide) that is necessary for cell/pollen via-bility. Alternatively, RGP1 and RGP2 may be respon-sible for maintaining and regulating a supply of sugarsfor wall biosynthesis that are important during thisessential stage in plant reproduction.
MATERIALS AND METHODS
Plant Material
All Arabidopsis (Arabidopsis thaliana) plants used in this study were in the
Columbia background. Plants were grown under standard long-day condi-
tions at 22�C under a 16-h light/8-h dark cycle.
In Vitro Pollen Germination
Pollen was germinated on agar as previously described by Hicks et al.
(2004). Germination medium was applied to the surface of a microscope slide
and allowed to solidify. Pollen from individual flowers was then dabbed onto
the agar and further incubated 3 to 8 h at 100% humidity.
Preparation of Constructs
Transgenic Arabidopsis plants expressing YFP-tagged RGP1 and RGP2
from its native regulatory sequences were generated employing the fluores-
cence tagging of full-length proteins technique (Tian et al., 2004). First,
AtRGP1 and AtRGP2 were amplified from genomic DNA as two fragments
using two sets of primers: RGP1-P1, 5#-gctcgatccacctaggctttgccttgttgggtt-
tatcctt-3#, RGP1-P2, 5#-cacagctccacctccacctccaggccggccccaagcttcaatccaagtgac-3#,
RGP1-P3, 5#-tgctggtgctgctgcggccgctggggccgatgagcttaacccacccacta-3#, RGP1-P4,
5#-cgtagcgagaccacaggatttgatttcggctcatttctca-3#, RGP2-P1, 5#-gctcgatccacctagg-
ctgccaaagctaacctctggtcat-3#, RGP2-P2, 5#-cacagctccacctccacctccaggccggccctca-
tcccaagcttcaatccat-3#, RGP2-P3, 5#-tgctggtgctgctgcggccgctggggcccttaacccacca-
gcagccagt-3#, and RGP2-P4, 5#-cgtagcgagaccacaggacggactattttcccccatacct-3#.
For the second PCR reaction, a pair of gene nonspecific Gateway primers
was used and triple template PCR was performed to produce the full-length
RGP1 and RGP2 with YFP coding sequence inserted into their last exon
approximately 30 bp upstream of the stop codon (Tian et al., 2004). The
resulting DNA fragments were recombined into pDONR207 (Invitrogen),
verified by DNA sequencing, and transferred by Gateway recombination into
the binary destination vectors pBIN-GW and pMN-GW (Tian et al., 2004).
For the His-T7-AtRGP1, the EcoRI-NotI fragment of AtRGP1 cDNA (ac-
cession no. AF013627) was cloned into the EcoRI-NotI sites of the N-terminal
His-T7-tag vector pET28c (Novagen). The resulting His-T7-AtRGP1 fusion
was excised with XbaI-XhoI digestion and religated into the XbaI-SalI sites of a
pCAMBIA 1300 modified vector.
Similarly, the NaeI-SalI fragment of AtRGP2 cDNA (accession no.
AF013628) was cloned into the SmaI-XhoI sites of the N-terminal HA-tag
vector pACT2 (Novagen). The resulting HA-AtRGP2 fusion was excised with
BglII digestion and religated to a pCAMBIA 3300 modified vector.
Plant Transformation
Wild-type seedlings were transformed by an infiltration method using
Agrobacterium tumefaciens strain GV3101 carrying the appropriate constructs
(Clough and Bent, 1998). Five to ten independent lines were analyzed. The
lines were amplified and the T3 plants were used for further experiments. The
double-tagged T7-RGP1 3 HARGP2 were obtained by genetic crosses of
single-tagged homozygous lines.
Genomic DNA Extraction and Isolation of T-DNAInsertion lines
Seedlings of T-DNA insertion lines Salk _132152 (Alonso et al., 2003) and
Gabi Kat_ 652F12 (Li et al., 2003) were grown individually. One fully
expanded leaf of a 4-week-old plant was used to extract genomic DNA. We
used two RGP1 gene-specific primers: forward (RGP1F), 5#-tcacttcctcaccataa-
caagg-3# and reverse (RGP1R), 5#-ttcacggtatgggtcgtaca-3#, and the T-DNA
primer, Gabi T-DNA, 5#-cccatttggacgtgaatgtagacac-3#. For RGP2, two gene-
specific primers—forward (RGP2F), 5#-agcaacccttttgttaacctga-3# and reverse
(RGP2R), 5#-aatatcgaccaaatgtgaaaagc-3# and the LBb, 5#-gcgtggaccgcttgctg-
caact-3#—were used on genomic DNA.
RT-PCR Analysis
Total RNA was extracted from rosette leaves or Arabidopsis seedlings
using the RNeasy plant mini kit (Qiagen) and treated with DNAse (Roche).
RT-PCR was carried out using 500 ng total RNA with the One-Step RT-PCR kit
(Qiagen) following the manufacturer’s protocol. Primers used for RGP1 were
forward (RGP1 RT-F), 5#-ctctccaaattctcttctctct-3# and reverse (RGP1 RT-R),
5#-caaaacggagaagatagatgatat-3#, to generate a 1,176-bp product; for RGP2,
forward (RGP2 RT-F), 5#-ataccctttcagtctccgtg-3# and reverse (RGP2 RT-R),
5#-gctcatactgctctcaagcttttgccac-3# to generate a 620-bp product. Ubiquitin
UBQ-10 forward, 5#-gatctttgccggaaaacaattggaggatggt-3# and reverse 5#-cgact-
tgtcattagaaagaaagagataacagg-3# primers were used as control.
Immunoprecipitation
Whole 4-week-old plants were homogenized in extraction buffer (40 mM
HEPES-KOH, pH 8; 0.45 M Suc; 1 mM EDTA; 1 mM MgCl2; 1 mM dithiothreitol)
and protease inhibitors as complete mixture (Roche). Soluble extracts were
separated by centrifugation at 100,000g for 40 min on a Ti70 rotor (Beckman)
and further enriched by ammonium sulfate precipitation (30%–50%). Proteins
were desalted using a PD10 (Amersham-Pharmacia Biotech) desalting column
and immunoprecipitated using the column mode of a T7 tag affinity purifi-
cation kit (Novagen) according to manufacturer’s instructions. Control col-
umns were prepared with unlinked agarose beads. Eluates from different
fractions were separated on SDS-PAGE, followed by immunoblotting using
various antibodies.
Genetic Crosses
To determine gametophytic transmission of the rgp1 and rgp2 insertions,
reciprocal crosses were performed between wild type and plants with only
one functional copy of RGP2 (rgp1/rgp1 RGP2/rgp2). Approximately 30
independent reciprocal crosses were done. Seeds were harvested and pooled
for (rgp1/rgp1 RGP2/rgp2) male or female. After sterilization and germination
on plates, DNA was extracted at the appearance of the fourth true leaf.
Progeny were genotyped for the presence of rgp1 and rgp2 T-DNA insertions
by PCR. X2 and other values are indicated. P values were calculated using
GraphPad Quickcals (available online at http://www.GraphPad.com).
Light Microscopy
Plant tissues were fixed in formaldehyde acetic acid containing 37% (w/v)
formaldehyde, glacial acetic acid, and 70% (v/v) ethanol (5:5:90; [v/v/v]),
overnight at 4�C. Samples were then dehydrated through an ethanol series of
up to 100% ethanol. The ethanol was replaced gradually by Technovit 7100
embedding solution (EMS). After polymerization, 3-mm sections were cut,
Drakakaki et al.
1490 Plant Physiol. Vol. 142, 2006 www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

mounted on glass slides, and stained with 0.5% toluidine blue for visualiza-
tion on a Nikon Microphot FXA microscope. Anthers from stage 13 were
squashed to release pollen and mounted in 50% (v/v) glycerol and viewed on
a Nikon microscope.
TEM and SEM
Stamens for TEM were taken from different flower stages and fixed with
2% glutaraldehyde in 50 mM phosphate buffer, pH 7.4, overnight at 4�C.
Samples were postfixed in 1% osmium tetroxide for 6 h on ice, dehydrated in
graduated ethanol series, embedded in Spurr’s resin, and polymerized at 50�C
for approximately 72 h. Ultrathin sections (60–70 nm) were cut with a
diamond knife and mounted on Formvar-coated copper grids. The sections
were stained with lead citrate and uranyl acetate (Reynolds, 1963) and viewed
using a FEI Tecnai 12-electron microscope (FEI).
Cryosections of Arabidopsis roots were used for all immunogold labeling
experiments as described in Sanderfoot et al. (1998). Controls were performed
with the use of 2% bovine serum albumin substituted for the antisera. Many
sections from independent plants were observed for each combination of
antibodies. In all cases, the antisera demonstrated high specificity of the
labeling.
For SEM, released pollen grains were mounted on stubs over double-sided
tape and sputter coated with gold particles (Cressington 108 Auto). Specimens
were examined with a scanning electron microscope (XL30 FEG SEM 30
ESEM; Philips) at an accelerating voltage of 10 kV.
CLSM
Live seedlings and plant tissue samples were mounted in water between
1.5 cover glasses, using silicon vacuum grease to create spaces between glass
surfaces. Images were collected using Leica TCS SP2/UV. A 514-nm laser line
from an argon ion was used to excite YFP. Released pollen grains were stained
directly on slides with 0.1% aniline blue for callose, 0.01% calcofluor white for
cellulose (as described by Nishikawa et al., 2005), and observed under UV
light excitation. Similarly, FDA (Heslop-Harrison and Heslop-Harrison, 1970)
was used to stain released pollen directly on slides viewed under standard
fluorescein isothiocyanate conditions under CLSM (Leica TCS SP2/UV). In all
cases, water immersion objectives 203 or 633 were employed. The optimal
pinhole diameter was set at 1 Airy unit in all cases.
Material and methods used for supplementary data are described in the
supplemental data.
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Materials and Methods S1.
Supplemental Figure S1. RGP1 expression in pollen grains.
Supplemental Figure S2. Subcellular fractionation of plants expressing
RGP1-YFP and RGP2-YFP.
Supplemental Figure S3. Pollen expressing RGP1-YFP and RGP2-YFP are
BFA sensitive.
Supplemental Figure S4. Fluorescent image of DAPI-stained tricellular
pollen.
Supplemental Table S1. Supplemental summary of the lethality of the
rgp1-1 and rgp2-1 and mutant progeny.
Supplemental Video S1. Z series of Arabidopsis leaf cells expressing
RGP2-YFP.
Supplemental Video S2. Time series of germinating pollen expressing
RGP1-YFP.
ACKNOWLEDGMENTS
We thank Kanwarpal Dhugga for kindly providing the anti-pea RGP
antibody. We also wish to thank Valentina Kovaleva for skillful immunolo-
calization; Marci Surpin for critical reading of this manuscript; Glenn Hicks
for advice and useful comments on pollen confocal microscopy; Thomas
Girke for advice on phylogenetic analysis; Elizabeth Lord and Juan Dong for
useful suggestions; Jocelyn Brimo for assistance with graphic arts during
submission of this manuscript; and Karen Bird for editorial assistance.
Received July 5, 2006; accepted October 17, 2006; published October 27, 2006.
LITERATURE CITED
Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen HM, Shinn P,
Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al (2003) Genome-
wide insertional mutagenesis of Arabidopsis thaliana. Science 301:
653–657
Becker JD, Boavida LC, Carneiro J, Haury M, Feijo JA (2003) Transcrip-
tional profiling of Arabidopsis tissues reveals the unique characteristics
of the pollen transcriptome. Plant Physiol 133: 713–725
Blanc G, Barakat A, Guyot R, Cooke R, Delseny I (2000) Extensive
duplication and reshuffling in the Arabidopsis genome. Plant Cell 12:
1093–1101
Bocca SN, Kissen R, Rojas-Beltran JA, Noel F, Gebhardt C, Moreno S, du
Jardin P, Tandecarz JS (1999) Molecular cloning and characterization of
the enzyme UDP-glucose: protein transglucosylase from potato. Plant
Physiol Biochem 37: 809–819
Bocca SN, Rothschild A, Tandecarz JS (1997) Initiation of starch biosyn-
thesis: purification and characterization of UDP-glucose:protein trans-
glucosylase from potato tubers. Plant Physiol Biochem 35: 205–212
Bolte S, Talbot C, Boutte Y, Catrice O, Read ND, Satiat-Jeunemaitre B
(2004) FM-dyes as experimental probes for dissecting vesicle trafficking
in living plant cells. J Microsc (Oxf) 214: 159–173
Bosch M, Cheung AY, Hepler PK (2005) Pectin methylesterase, a regulator
of pollen tube growth. Plant Physiol 138: 1334–1346
Bosch M, Hepler PK (2005) Pectin methylesterases and pectin dynamics in
pollen tubes. Plant Cell 17: 3219–3226
Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agro-
bacterium-mediated transformation of Arabidopsis thaliana. Plant J 16:
735–743
Dai S, Li L, Chen T, Chong K, Xue Y, Wang T (2006) Proteomic analyses of
Oryza sativa mature pollen reveal novel proteins associated with pollen
germination and tube growth. Proteomics 6: 2504–2529
Delgado IJ, Wang ZH, de Rocher A, Keegstra K, Raikhel NV (1998)
Cloning and characterization of AtRGP1—a reversibly autoglycosylated
Arabidopsis protein implicated in cell wall biosynthesis. Plant Physiol
116: 1339–1349
Dhugga KS, Tiwari SC, Ray PM (1997) A reversibly glycosylated poly-
peptide (RGP1) possibly involved in plant cell wall synthesis: purifica-
tion, gene cloning, and trans-Golgi localization. Proc Natl Acad Sci USA
94: 7679–7684
Dhugga KS, Ulvskov P, Gallagher SR, Ray PM (1991) Plant polypeptides
reversibly glycosylated by UDP-glucose—possible components of Golgi
beta-glucan synthase in pea cells. J Biol Chem 266: 21977–21984
Dong XY, Hong ZL, Sivaramakrishnan M, Mahfouz M, Verma DPS
(2005) Callose synthase (CalS5) is required for exine formation during
microgametogenesis and for pollen viability in Arabidopsis. Plant J 42:
315–328
Enns LC, Kanaoka MM, Torii KU, Comai L, Okada K, Cleland RE (2005)
Two callose synthases, GSL1 and GSL5, play an essential and redun-
dant role in plant and pollen development and in fertility. Plant Mol Biol
58: 333–349
Felsenstein J (2005) Using the quantitative genetic threshold model for
inferences between and within species. Philos Trans R Soc Lond B Biol
Sci 360: 1427–1434
Gallardo K, Le Signor C, Vandekerckhove J, Thompson RD, Burstin J
(2003) Proteomics of Medicago truncatula seed development establishes
the time frame of diverse metabolic processes related to reserve accu-
mulation. Plant Physiol 133: 664–682
Girke T, Lauricha J, Tran H, Keegstra K, Raikhel N (2004) The cell wall
navigator database: a systems-based approach to organism-unrestricted
mining of protein families involved in cell wall metabolism. Plant
Physiol 136: 3003–3008
Goubet F, Misrahi A, Park SK, Zhang ZN, Twell D, Dupree P (2003)
AtCSLA7, a cellulose synthase-like putative glycosyltransferase, is
Reversibly Glycosylated Poplypeptides in Pollen Development
Plant Physiol. Vol. 142, 2006 1491 www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

important for pollen tube growth and embryogenesis in Arabidopsis.
Plant Physiol 131: 547–557
Heslop-Harrison J, Heslop-Harrison Y (1970) Evaluation of pollen viabil-
ity by enzymatically induced fluorescence; intracellular hydrolysis of
fluorescein diacetate. Stain Technol 45: 115–120
Hicks GR, Rojo E, Hong SH, Carter DG, Raikhel NV (2004) Geminating
pollen has tubular vacuoles, displays highly dynamic vacuole biogen-
esis, and requires VACUOLESS1 for proper function. Plant Physiol 134:
1227–1239
Holmes-Davis R, Tanaka CK, Vensel WH, Hurkman WJ, McCormick S
(2005) Proteome mapping of mature pollen of Arabidopsis thaliana.
Proteomics 5: 4864–4884
Honys D, Twell D (2003) Comparative analysis of the Arabidopsis pollen
transcriptome. Plant Physiol 132: 640–652
Hulskamp M, Parekh NS, Grini P, Schneitz K, Zimmermann I, Lolle SJ,
Pruitt RE (1997) The STUD gene is required for male-specific cytokinesis
after telophase II of meiosis in Arabidopsis thaliana. Dev Biol 187:
114–124
Jiang LX, Yang SL, Xie LF, Puah CS, Zhang XQ, Yang WC, Sundaresan V,
Ye D (2005) VANGUARD1 encodes a pectin methylesterase that en-
hances pollen tube growth in the Arabidopsis style and transmitting
tract. Plant Cell 17: 584–596
Langeveld SMJ, Vennik M, Kottenhagen M, van Wijk R, Buijk A, Kijne
JW, de Pater S (2002) Glucosylation activity and complex formation of
two classes of reversibly glycosylated polypeptides. Plant Physiol 129:
278–289
Li Y, Rosso MG, Strizhov N, Viehoever P, Weisshaar B (2003) GABI-Kat
SimpleSearch: a flanking sequence tag (FST) database for the identifica-
tion of T-DNA insertion mutants in Arabidopsis thaliana. Bioinformatics
19: 1441–1442
Li YQ, Faleri C, Geitmann A, Zhang HQ, Cresti M (1995) Immunogold
localization of arabinogalactan proteins, unesterified and esterified
pectins in pollen grains and pollen tubes of Nicotiana-tabacum-L.
Protoplasma 189: 26–36
Nagasawa M, Mori H, Shiratake K, Yamaki S (2005) Isolation of cDNAs for
genes expressed after/during fertilization and fruit set of melon
(Cucumis melo L.). J Jpn Soc Hortic Sci 74: 23–30
Nebenfuhr A, Ritzenthaler C, Robinson DG (2002) Brefeldin A: decipher-
ing an enigmatic inhibitor of secretion. Plant Physiol 130: 1102–1108
Nishikawa S, Zinkl GM, Swanson RJ, Maruyama D, Preuss D (2005)
Callose (beta-1,3 glucan) is essential for Arabidopsis pollen wall pat-
terning, but not tube growth. BMC Plant Biol 5: 22
Noir S, Brautigam A, Colby T, Schmidt J, Panstruga R (2005) A reference
map of the Arabidopsis thaliana mature pollen proteome. Biochem
Biophys Res Commun 337: 1257–1266
Oh SA, Johnson A, Smertenko A, Rahman D, Park SK, Hussey PJ, Twell
D (2005) A divergent cellular role for the FUSED kinase family in the
plant-specific cytokinetic phragmoplast. Curr Biol 15: 2107–2111
Park SK, Howden R, Twell D (1998) The Arabidopsis thaliana gameto-
phytic mutation gemini pollen1 disrupts microspore polarity, division
asymmetry and pollen cell fate. Development 125: 3789–3799
Park SK, Twell D (2001) Novel patterns of ectopic cell plate growth and
lipid body distribution in the Arabidopsis gemini pollen1 mutant. Plant
Physiol 126: 899–909
Pina C, Pinto F, Feijo JA, Becker JD (2005) Gene family analysis of the
Arabidopsis pollen transcriptome reveals biological implications for cell
growth, division control, and gene expression regulation. Plant Physiol
138: 744–756
Reynolds ES (1963) The use of lead citrate at high pH as an electron-opaque
stain in electron microscopy. J Cell Biol 17: 208–212
Rothschild A, Tandecarz JS (1994) Udp-glucose-protein transglucosylase
in developing maize endosperm. Plant Sci 97: 119–127
Sagi G, Katz A, Guenoune-Gelbart D, Epel BL (2005) Class 1 reversibly
glycosylated polypeptides are plasmodesmal-associated proteins deliv-
ered to plasmodesmata via the Golgi apparatus. Plant Cell 17: 1788–1800
Sanderfoot AA, Ahmed SU, Marty-Mazars D, Rapoport I, Kirchhausen T,
Marty F, Raikhel NV (1998) A putative vacuolar cargo receptor partially
colocalizes with AtPEP12p on a prevacuolar compartment in Arabidop-
sis roots. Proc Natl Acad Sci USA 95: 9920–9925
Sanders PM, Bui AQ, Weterings K, McIntire KN, Hsu YC, Lee PY, Truong
MT, Beals TP, Goldberg RB (1999) Anther developmental defects in
Arabidopsis thaliana male-sterile mutants. Sex Plant Reprod 11: 297–322
Schneidereit A, Scholz-Starke J, Buttner M (2003) Functional characteriza-
tion and expression analyses of the glucose-specific AtSTP9 monosac-
charide transporter in pollen of Arabidopsis. Plant Physiol 133: 182–190
Schneidereit A, Scholz-Starke J, Sauer N, Buttner M (2005) AtSTP11, a
pollen tube-specific monosaccharide transporter in Arabidopsis. Planta
221: 48–55
Schnurr JA, Storey KK, Jung HJ, Somers DA, Gronwald JW (2006) UDP-
sugar pyrophosphorylase is essential for pollen development in Arabi-
dopsis. Planta 224: 520–532
Scholz-Starke J, Buttner M, Sauer N (2003) AtSTP6, a new pollen-specific H1-
monosaccharide symporter from Arabidopsis. Plant Physiol 131: 70–77
Selth LA, Dogra SC, Rasheed MS, Randles JW, Rezaian MA (2006)
Identification and characterization of a host reversibly glycosylated
peptide that interacts with the tomato leaf curl virus V1 protein. Plant
Mol Biol 61: 297–310
Tanaka H, Ishikawa M, Kitamura S, Takahashi Y, Soyano T, Machida C,
Machida Y (2004) The AtNACK1/HINKEL and STUD/TETRASPORE/
AtNACK2 genes, which encode functionally redundant kinesins, are
essential for cytokinesis in Arabidopsis. Genes Cells 9: 1199–1211
Testasecca P, Wald FA, Cozzarin ME, Moreno S (2004) Regulation of self-
glycosylation of reversibly glycosylated polypeptides from Solanum
tuberosum. Physiol Plant 121: 27–34
Tian GW, Chen MH, Zaltsman A, Citovsky V (2006) Pollen-specific pectin
methylesterase involved in pollen tube growth. Dev Biol 294: 83–91
Tian GW, Mohanty A, Chary SN, Li SJ, Paap B, Drakakaki G, Kopec CD,
Li JX, Ehrhardt D, Jackson D, et al (2004) High-throughput fluorescent
tagging of full-length Arabidopsis gene products in planta. Plant
Physiol 135: 25–38
Truernit E, Schmid J, Epple P, Illig J, Sauer N (1996) The sink-specific and
stress-regulated Arabidopsis STP4 gene: enhanced expression of a gene
encoding a monosaccharide transporter by wounding, elicitors, and
pathogen challenge. Plant Cell 8: 2169–2182
Truernit E, Stadler R, Baier K, Sauer N (1999) A male gametophyte-specific
monosaccharide transporter in Arabidopsis. Plant J 17: 191–201
Wu AM, Ling C, Liu JY (2006) Isolation of a cotton reversibly glycosylated
polypeptide (GhRGP1) promoter and its expression activity in trans-
genic tobacco. J Plant Physiol 163: 426–435
Yang CY, Spielman M, Coles JP, Li Y, Ghelani S, Bourdon V, Brown RC,
Lemmon BE, Scott RJ, Dickinson HG (2003) TETRASPORE encodes a
kinesin required for male meiotic cytokinesis in Arabidopsis. Plant J 34:
229–240
Zhao GR, Liu JY, Du XM (2001) Molecular cloning and characterization of
cotton cDNAs expressed in developing fiber cells. Biosci Biotechnol
Biochem 65: 2789–2793
Drakakaki et al.
1492 Plant Physiol. Vol. 142, 2006 www.plant.org on January 31, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.
Copyright © 2022 FDOKUMEN




















