
Exposure of Intestinal Epithelial Cells to UV-Killed Lactobacillus GG but Not Bifidobacterium breve...
10
Fax +41 61 306 12 34 E-Mail [email protected] www.karger.com Original Paper Int Arch Allergy Immunol 2010;152:159–168 DOI: 10.1159/000265537 Exposure of Intestinal Epithelial Cells to UV-Killed Lactobacillus GG but Not Bifidobacterium breve Enhances the Effector Immune Response in vitro Els van Hoffen a Nicoline M. Korthagen b Sander de Kivit b Bastiaan Schouten b Bart Bardoel a Arjan Duivelshof c Jan Knol c Johan Garssen b, c Linette E.M. Willemsen b a Dermatology/Allergology, University Medical Center Utrecht, b Pharmacology and Pathophysiology, Utrecht Institute Pharmaceutical Sciences, Utrecht University, Utrecht, and c Danone Research Centre for Specialised Nutrition, Wageningen, The Netherlands TLR2 and NOD2 ligands, but not Bb, enhanced IFN- , IL-12 and/or TNF- secretion. Bacteria did not induce cytokine se- cretion when added to HT-29/unstimulated PBMC co-cul- tures, whereas direct incubations with PBMC did. Conclu- sion: UV-killed LGG as well as Bb supported a T helper 1 and/ or regulatory phenotype when added directly to activated PBMC, similar to live bacteria. In contrast, LGG, TLR2 or NOD2 ligands – but not Bb – enhanced T helper 1 type cytokine secretion when added to IEC, while IL-10 secretion remained suppressed. Co-cultures combining IEC and PBMC may re- veal differences between bacterial strains relevant for the in vivo situation. Copyright © 2009 S. Karger AG, Basel Introduction In Western societies, every 1 out of 5 people suffers from allergies. Many of these allergies are food related, affecting 6–8% of young children and 1–4% of adults, and involve the gastrointestinal tract as well as the gut im- mune system [1]. In food-allergic patients, allergens en- tering the gut fail to elicit oral tolerance and activate the adaptive immune system resulting in a T helper (Th) 2 polarized inflammation [1] . It has been suggested that Key Words Bifidobacterium breve Co-culture, intestinal epithelial cells HT-29 Immune modulation Lactobacillus GG Nucleotide-binding oligomerization domain Probiotics Toll-like receptor Abstract Background: Intestinal bacteria and intestinal epithelial cells (IEC) may modulate the mucosal immune response. In this study, immune modulation by Lactobacillus GG (LGG) and Bifidobacterium breve (Bb1, Bb2) in the presence or ab- sence of IEC was addressed in an in vitro transwell co-culture model. Methods: UV-killed LGG, Bb1, Bb2 or Toll-like recep- tor (TLR) 2 or nucleotide oligomerization domain (NOD) 2 ligands were added directly to unstimulated or anti-CD3/ CD28-stimulated PBMC, or applied apically to human IEC (HT-29) co-cultured with PBMC. A mixture of live bacteria was used as reference. The effect on T helper 1 (IFN- , IL-12), T helper 2 (IL-13), inflammatory (TNF- ) and regulatory (IL-10) cytokine secretion was determined. Results: Both UV-killed LGG and Bb enhanced IL-12, IFN- , TNF- and IL-10, and re- duced IL-13 secretion when added directly to stimulated PBMC, similar to live bacteria. IEC reduced IL-13, IFN- and IL-10 secretion by stimulated PBMC. Apically added LGG, Received: February 9, 2009 Accepted after revision: August 26, 2009 Published online: December 16, 2009 Correspondence to: Dr. Linette E.M. Willemsen Division of Pharmacology and Pathophysiology Utrecht Institute for Pharmaceutical Sciences, Utrecht University Sorbonnelaan 16, PO Box 80082, NL–3508 TB Utrecht (The Netherlands) Tel. +31 30 253 6950, Fax +31 30 253 7420, E-Mail l.e.m.willemsen @ uu.nl © 2009 S. Karger AG, Basel 1018–2438/10/1522–0159$26.00/0 Accessible online at: www.karger.com/iaa
Transcript of Exposure of Intestinal Epithelial Cells to UV-Killed Lactobacillus GG but Not Bifidobacterium breve...

Fax +41 61 306 12 34E-Mail [email protected]
Original Paper
Int Arch Allergy Immunol 2010;152:159–168 DOI: 10.1159/000265537
Exposure of Intestinal Epithelial Cells to UV-Killed Lactobacillus GG but Not Bifidobacterium breve Enhances the Effector Immune Response in vitro
Els van Hoffen a Nicoline M. Korthagen b Sander de Kivit b Bastiaan Schouten b
Bart Bardoel a Arjan Duivelshof c Jan Knol c Johan Garssen b, c
Linette E.M. Willemsen b
a Dermatology/Allergology, University Medical Center Utrecht, b Pharmacology and Pathophysiology,Utrecht Institute Pharmaceutical Sciences, Utrecht University, Utrecht , and c Danone Research Centre for Specialised Nutrition, Wageningen , The Netherlands
TLR2 and NOD2 ligands, but not Bb , enhanced IFN- � , IL-12 and/or TNF- � secretion. Bacteria did not induce cytokine se-cretion when added to HT-29/unstimulated PBMC co-cul-tures, whereas direct incubations with PBMC did. Conclu-
sion: UV-killed LGG as well as Bb supported a T helper 1 and/or regulatory phenotype when added directly to activated PBMC, similar to live bacteria. In contrast, LGG , TLR2 or NOD2 ligands – but not Bb – enhanced T helper 1 type cytokine secretion when added to IEC, while IL-10 secretion remained suppressed. Co-cultures combining IEC and PBMC may re-veal differences between bacterial strains relevant for the in vivo situation.
Copyright © 2009 S. Karger AG, Basel
Introduction
In Western societies, every 1 out of 5 people suffers from allergies. Many of these allergies are food related, affecting 6–8% of young children and 1–4% of adults, and involve the gastrointestinal tract as well as the gut im-mune system [1] . In food-allergic patients, allergens en-tering the gut fail to elicit oral tolerance and activate the adaptive immune system resulting in a T helper (Th) 2 polarized inflammation [1] . It has been suggested that
Key Words
Bifidobacterium breve � Co-culture, intestinal epithelial cells � HT-29 � Immune modulation � Lactobacillus GG � Nucleotide-binding oligomerization domain � Probiotics � Toll-like receptor
Abstract
Background: Intestinal bacteria and intestinal epithelial cells (IEC) may modulate the mucosal immune response. In this study, immune modulation by Lactobacillus GG (LGG) and Bifidobacterium breve (Bb1, Bb2) in the presence or ab-sence of IEC was addressed in an in vitro transwell co-culture model. Methods: UV-killed LGG, Bb1, Bb2 or Toll-like recep-tor (TLR) 2 or nucleotide oligomerization domain (NOD) 2 ligands were added directly to unstimulated or anti-CD3/CD28-stimulated PBMC, or applied apically to human IEC (HT-29) co-cultured with PBMC. A mixture of live bacteria was used as reference. The effect on T helper 1 (IFN- � , IL-12), T helper 2 (IL-13), inflammatory (TNF- � ) and regulatory (IL-10) cytokine secretion was determined. Results: Both UV-killed LGG and Bb enhanced IL-12, IFN- � , TNF- � and IL-10, and re-duced IL-13 secretion when added directly to stimulated PBMC, similar to live bacteria. IEC reduced IL-13, IFN- � and IL-10 secretion by stimulated PBMC. Apically added LGG,
Received: February 9, 2009 Accepted after revision: August 26, 2009 Published online: December 16, 2009
Correspondence to: Dr. Linette E.M. Willemsen Division of Pharmacology and Pathophysiology Utrecht Institute for Pharmaceutical Sciences, Utrecht University Sorbonnelaan 16, PO Box 80082, NL–3508 TB Utrecht (The Netherlands) Tel. +31 30 253 6950, Fax +31 30 253 7420, E-Mail l.e.m.willemsen @ uu.nl
© 2009 S. Karger AG, Basel1018–2438/10/1522–0159$26.00/0
Accessible online at:www.karger.com/iaa

van Hoffen et al.
Int Arch Allergy Immunol 2010;152:159–168160
differences in the composition of the gut microbiota might be related to the aberrant development of the im-mune system, leading to allergic diseases [2] .
According to the currently adopted definition by the Food and Agriculture Organization/World Health Orga-nization (2001), probiotics are: ‘ Live microorganisms which when administered in adequate amounts confer a health benefit on the host ’ . It has been shown that these bacteria can strengthen the intestinal barrier and en-hance T helper (Th) 1 and regulatory T cell (Treg) func-tion [3, 4] , which may be beneficial in allergic disease. Several placebo-controlled clinical trials have shown that Lactobacillus rhamnosus GG in particular, but also mixed preparations containing Lactobacillus and Bifidobacte-rium species reduce the development or severity of atop-ic eczema in infants at risk for atopic disease [4–6] . Hence, use of probiotic bacteria is increasingly explored for pre-vention or treatment of allergic diseases. In this regard, the selection of beneficial bacterial strains to be used in clinical studies is of key importance.
In the gut, bacteria are sampled by dendritic cells (DC) or M-cells. In this way they will enter primary immune organs like the Peyer’s patches and mesenteric lymph nodes [7] . It is known that probiotics may affect immune function via these routes. However, the majority of pro-biotics remain within the intestinal lumen. The immune cells of the gut-associated lymphoid tissue and lamina propria in particular are located directly underneath a monolayer of intestinal epithelial cells (IEC) [7] . IEC maintain homeostasis in the gut, and probiotic bacteria are essential for the development of oral tolerance and the support of epithelial function [8] . When studying effects of probiotics in vitro, mostly the potential immune mod-ulation by IEC is neglected. Furthermore, numerous studies using in vitro co-culture models have shown that this microbial-epithelial ‘cross-talk’ may dampen or stimulate the immune response [9–13] . Both IEC and mucosal immune cells contain Toll-like receptors (TLR) and nucleotide-binding oligomerization domain (NOD) that may initiate cell signaling after binding of bacterial cell wall components [14] . Cell membranes of Gram-pos-itive Lactobacilli and Bifidobacteria contain lipoproteins, peptidoglycans and lipoteichoic acids that might be li-gands for TLR2 and co-receptors TLR1 and TLR6 and NOD2 expressed by IEC [14, 15] . NOD is expressed by IEC and APC; NOD1 senses peptidoglycans of Gram-negative bacteria, while NOD2 senses muramyl dipeptide (MDP) also present on Gram-positive bacteria [14] .
In this study, an in vitro co-culture model was used to evaluate the differential potential of UV-killed Lactoba-
cillus GG and Bifidobacterium breve bacteria to modulate the immune response in the presence or absence of IEC. A mixture of live bacteria, that was previously shown to skew the response of stimulated PBMC in vitro towards a Th1/Treg phenotype [16] , was used as a reference. Hu-man intestinal IEC (HT-29) were grown on filters, and unstimulated or anti-CD3/CD28-activated PBMC were added basolaterally in a separate compartment. The bac-teria were added apically to the IEC or applied directly to the PBMC in absence of IEC.
Materials and Methods
Probiotic Preparations and TLR2 or NOD2 Ligand A mixture of live probiotics as well as UV-killed individual
strains were used. The mixture of bacteria contained the Lactoba-cilli L. acidophilus , L. casei and L. salivarius , as well as Lactococcus lactis , Bifidobacterium infantis , B. lactis and B. longum (Orthiflor Junior; Orthica, Almere, The Netherlands). This mixture of live bacteria, that was previously shown to skew the response of stimu-lated PBMC in vitro towards a Th1/Treg phenotype [16] , was used as a reference. The individual strains were L. rhamnosus strain GG (LGG) and B. breve strain NumRes 200 (Bb1) and B. breve strain NumRes 204 ( Bb2 ; Danone Research BV, Wageningen, The Neth-erlands). These individual strains were UV-killed for 20 min at 70% of the maximal intensity using a Gibco BRL UV Transillumi-nator (Invitrogen, Carlsbad, Calif., USA), and cell death was con-firmed by plating on MRS agar. The strains were aliquoted and stored at –80 ° C in 10% glycerol. Before use the glycerol-stored strains were quickly thawed and centrifuged for 15 min at 10,000 rpm at 4 ° C. The supernatant was removed and the pellet was washed in cold PBS. After spinning 5 min at 10,000 rpm, the su-pernatant was removed and the pellet was resuspended in culture medium. Bacterial solutions of 5 ! 10 7 CFU/ml were prepared in culture medium. TLR2 ligand Pam 3 Cys-SKKKK (1 �g/ml; EMC Microcollections, Tübingen, Germany) and NOD2 ligand mur-amyl dipeptide (MDP, active L-D isomer) and a negative control for MDP (inactive D-D isomer) (5 �g/ml) were obtained from Invivo-gen (San Diego, Calif., USA).
PBMC Cultures Buffy coats (Sanquin, Amsterdam, The Netherlands) or ve-
nous heparinized blood was obtained from healthy adult volun-teers. PBMC were isolated using Ficoll-Isopaque centrifugation (Amersham Biosciences, Uppsala, Sweden). After isolation and washing, remaining erythrocytes were removed by adding 5 ml 4 ° C lysis buffer (4.14 g NH 4 Cl, 0.5 g KHCO 3 , 18.6 mg Na 2 EDTA in 500 ml demi-water, pH adjusted to 7.4 and filter sterilized) and incubation for 5 min on ice. PBMC were cultured in Iscove’s Mod-ified Dulbecco’s Medium (IMDM; Invitrogen) supplemented with 5% heat-inactivated pooled human serum (Lonza, Verviers, Belgium), penicillin (100 U/ml)/streptomycin (100 � g/ml; Sigma, UK) and glutamine (1 m M ; Invitrogen), or in RPMI 1640 (Lonza) supplemented with 2.5% heat-inactivated fetal calf serum (Invi-trogen), penicillin/streptomycin (Sigma) and pyruvate (1 m M; Sigma).

Immune Modulation in a HT-29/PBMC Transwell Co-Culture Model
Int Arch Allergy Immunol 2010;152:159–168 161
IEC Culture Conditions The human colon adenocarcinoma cell line HT-29 (HTB-38;
American Type Culture Collection, Manassas, Va., USA; passag-es 136–150) was used as IEC. HT-29 cells were cultured in Mc-Coy’s 5A medium (Invitrogen) supplemented with 10% heat-in-activated fetal calf serum, penicillin (100 U/ml)/streptomycin (100 � g/ml; Sigma) and pyruvate (1 m M ; Sigma). Cells were grown in 25-cm 2 culture flasks (Greiner, Frickenhausen, Germany) or cultured on transwell plates (Costar; Corning Inc., Corning, N.Y., USA; 0.4 � m pore size to prevent crossing of whole bacteria) and medium was refreshed every 2–3 days. HT-29 cells were grown to confluence in 1 week, confluency of the cell layer was confirmed using light microscopy. During transwell experiments, PBMC medium was used, containing 2.5% fetal calf serum in order to allow determination of endogenous TGF- � 1 secretion.
Transwell Co-Cultures and pH Measurements IEC were cultured on transwell filters (1 cm 2 surface, 0.4- � m
filter, 12-well plate). HT-29 were used just after reaching con-fluency. PBMC were added to the basolateral compartment (1.5 ml) at a concentration of 2 ! 10 6 cells/ml in presence or absence of anti-CD3 (clone T3/4.E) and anti-CD28 (clone CD28/1) stimu-lation (Sanquin, both 1: 10,000). In the apical compartment0.5 ml of 5 ! 10 7 CFU/ml bacteria were added on top of the IEC (IEC:bacteria approx. 1: 10; fig. 1 ).
Additionally in 12-well plates in absence of IEC, 0.5 ml bacte-rial suspension (1.25 ! 10 6 CFU/ml) was added to 1 ml PBMC(3 ! 10 6 cells/ml) in presence or absence of anti-CD3/CD28 stim-ulation (bacteria:PBMC 1: 5). After 24 h, 500 � l of basolateral su-pernatant was refreshed. Plates were incubated for 48 h in an in-
cubator at 37 ° C and 5% CO 2 . Basolateral supernatants at 48 h were collected and stored at –20 ° C for cytokine measurements. The pH was tested in order to determine whether the pH re-mained in physiological ranges (SevenGo pH meter; Mettler To-ledo, Hengelo, The Netherlands).
Proliferation Assays Lymphocyte stimulation tests were performed in triplicate in
96-well flat-bottom plates (Greiner). Each well contained 1 ! 10 5 PBMCs, and cells were either left unstimulated, or were stimu-lated with anti-CD3/CD28 (Sanquin), in the presence or absence of probiotics in a 1: 5 or 1: 1 ratio (0.2 ! 10 5 or 1 ! 10 5 CFU/well). Supernatants were harvested at 24 and 72 h and stored at –20 ° C for cytokine measurement. Proliferation was measured by [ 3 H]-TdR incorporation (1 � Ci/well; Amersham, Aylesbury, UK), which was added at 72 h, harvested on glass-fiber filters after18 h, and counted using a 1205 TM betaplate counter (Wallac, Turku, Finland). Proliferative responses are expressed as the mean [ 3 H]-TdR incorporation (cpm) of triplicates.
Cytokine ELISA The production of cytokines was measured in the supernatant
of PBMC cultures. ELISAs for IL-10, TNF- � , IL-13, IFN- � , IL-12 p40/p70 (Sanquin and/or Biosource; Invitrogen) and TGF- � 1 and TSLP (R&D, Abingdon, UK) were used according to the manufac-turer’s instructions. The detection limits were 2.3 pg/ml for IL-10, 3.1 pg/ml for TNF- � , 1.2 pg/ml for IL-13, 3.1 pg/ml for IFN- � , 10 pg/ml for IL-12, 4.6 pg/ml for TGF- � 1 and 31 pg/ml for TSLP.
Statistical Analyses Data were analyzed by means of paired t test, or by repeated
measures 1-way ANOVA with post hoc Dunnett correction for multiple comparisons (SPSS v.13). To obtain a normal distribu-tion, the cytokine data were 10 log transformed to permit paramet-ric statistical analyses.
Results
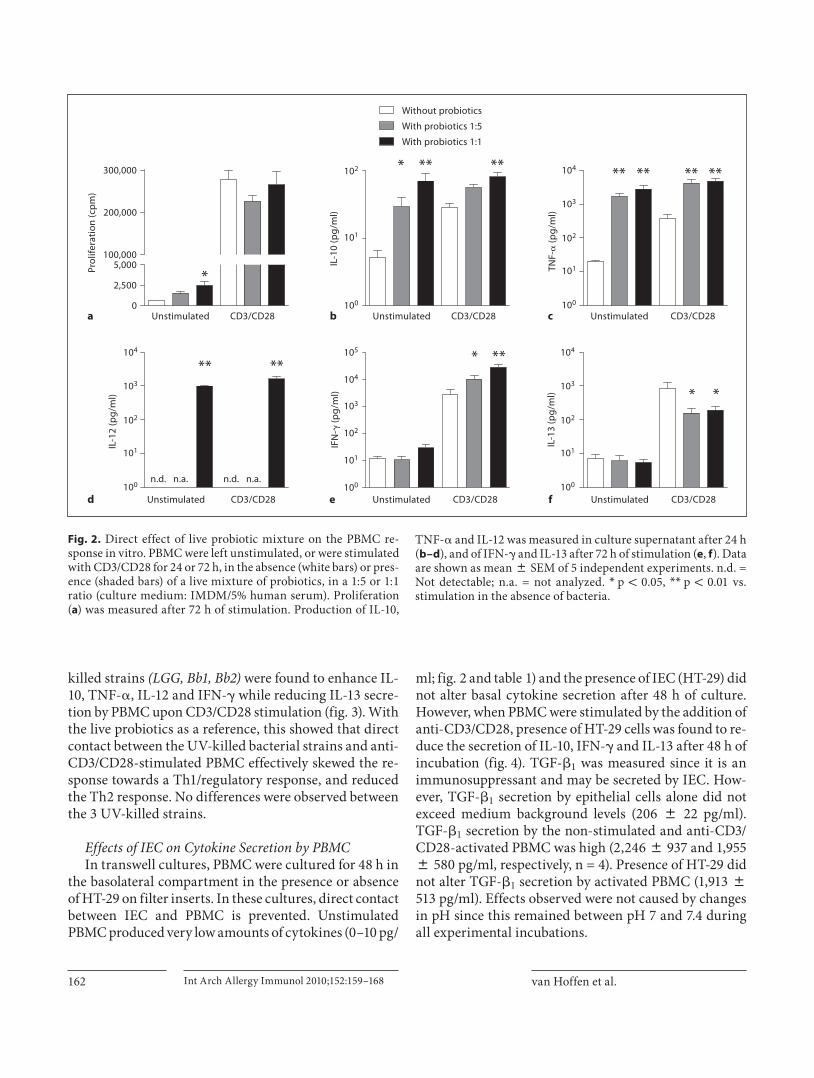
Direct Effects of Probiotics on PBMC Responses in vitro To study the direct effect of the bacteria on the re-
sponse of PBMC the live bacteria were added in a 1: 1 or 1: 5 ratio, this live probiotic mixture was used as reference. T cell proliferation was significantly enhanced by the probiotics in the unstimulated, but not in the anti-CD3/CD28-stimulated cultures ( fig. 2 a). This effect was inde-pendent of the medium used (IMDM/5% human serum culture medium vs. RPMI/2.5% fetal calf serum; data not shown). Addition of live probiotics enhanced the produc-tion of IL-10, TNF- � , IL-12 in unstimulated and stimu-lated cultures both at 24 and 72 h (shown for 24 h in fig. 2 b–d). Production of IFN- � and IL-13 was observed in anti-CD3/CD28-stimulated cultures at 72 h. IFN- � was enhanced and IL-13 was reduced upon addition of bacteria ( fig. 2 e, f). Similar to the live bacteria, the 3 UV-
Bacteriaor
TLR/NOD agonists
± anti-CD3/CD28
Fig. 1. Co-culture model to study immune modulation by IEC. IEC (HT-29, a human intestinal adenocarcinoma cell line) were cultured on transwell inserts and PBMC were added to the baso-lateral compartment. PBMC were stimulated with anti-CD3/CD28 to bring about an effector immune response. Bacteria or TLR or NOD agonists were added to the apical compartment.
van Hoffen et al.
Int Arch Allergy Immunol 2010;152:159–168162
killed strains (LGG, Bb1, Bb2) were found to enhance IL-10, TNF- � , IL-12 and IFN- � while reducing IL-13 secre-tion by PBMC upon CD3/CD28 stimulation ( fig. 3 ). With the live probiotics as a reference, this showed that direct contact between the UV-killed bacterial strains and anti-CD3/CD28-stimulated PBMC effectively skewed the re-sponse towards a Th1/regulatory response, and reduced the Th2 response. No differences were observed between the 3 UV-killed strains.
Effects of IEC on Cytokine Secretion by PBMC In transwell cultures, PBMC were cultured for 48 h in
the basolateral compartment in the presence or absence of HT-29 on filter inserts. In these cultures, direct contact between IEC and PBMC is prevented. Unstimulated PBMC produced very low amounts of cytokines (0–10 pg/
ml; fig. 2 and table 1 ) and the presence of IEC (HT-29) did not alter basal cytokine secretion after 48 h of culture. However, when PBMC were stimulated by the addition of anti-CD3/CD28, presence of HT-29 cells was found to re-duce the secretion of IL-10, IFN- � and IL-13 after 48 h of incubation ( fig. 4 ). TGF- � 1 was measured since it is an immunosuppressant and may be secreted by IEC. How-ever, TGF- � 1 secretion by epithelial cells alone did not exceed medium background levels (206 8 22 pg/ml). TGF- � 1 secretion by the non-stimulated and anti-CD3/CD28-activated PBMC was high (2,246 8 937 and 1,955 8 580 pg/ml, respectively, n = 4). Presence of HT-29 did not alter TGF- � 1 secretion by activated PBMC (1,913 8 513 pg/ml). Effects observed were not caused by changes in pH since this remained between pH 7 and 7.4 during all experimental incubations.
0
2,500
5,000100,000
200,000
300,000
Without probiotics
With probiotics 1:5
With probiotics 1:1
*Prol
ifera
tion
(cp
m)
102 104
103
102
101
100
104 105
104
103
102
101
100
103
102
101
100
104
103
102
101
100
101
100
** ***
IL-1
0 (p
g/m
l)
** ** ** **
TNF-
� (p
g/m
l)
Unstimulated
***
IFN
-� (p
g/m
l) **
IL-1
3 (p
g/m
l)
n.d. n.a. n.d. n.a.
** **
IL-1
2 (p
g/m
l)
CD3/CD28
Unstimulated CD3/CD28 Unstimulated CD3/CD28 Unstimulated CD3/CD28
Unstimulated CD3/CD28 Unstimulated CD3/CD28a
d e f
b c
Fig. 2. Direct effect of live probiotic mixture on the PBMC re-sponse in vitro. PBMC were left unstimulated, or were stimulated with CD3/CD28 for 24 or 72 h, in the absence (white bars) or pres-ence (shaded bars) of a live mixture of probiotics, in a 1: 5 or 1: 1 ratio (culture medium: IMDM/5% human serum). Proliferation ( a ) was measured after 72 h of stimulation. Production of IL-10,
TNF- � and IL-12 was measured in culture supernatant after 24 h ( b–d ), and of IFN- � and IL-13 after 72 h of stimulation ( e , f ). Data are shown as mean 8 SEM of 5 independent experiments. n.d. = Not detectable; n.a. = not analyzed. * p ! 0.05, * * p ! 0.01 vs. stimulation in the absence of bacteria.

Immune Modulation in a HT-29/PBMC Transwell Co-Culture Model
Int Arch Allergy Immunol 2010;152:159–168 163
103 104
103
102
101
100
104 105
104
103
102
101
100
103
102
101
100
104
103
102
101
100
102
101
100
** ** ** **
IL-1
0 (p
g/m
l)
** ** ** **
TNF-
� (p
g/m
l)
n.d.
** ** ** **
IL-1
2 (p
g/m
l)
** ** ** **
IFN
-� (p
g/m
l)
** ** ** **
IL-1
3 (p
g/m
l)
Control Mix LGG Bb1 Bb2
Control Mix LGG Bb1 Bb2
Control Mix LGG Bb1 Bb2
Control Mix LGG Bb1 Bb2
Control Mix LGG Bb1 Bb2c
e
d
a b
Fig. 3. Direct effect of live mix of probiot-ics or UV-killed LGG or Bb on the PBMC response in vitro. PBMC were stimulated with CD3/CD28 for 24 or 72 h, in the ab-sence (control) or presence of a live mix-ture of bacteria (Mix), or of UV-killed strains LGG, Bb1 (NumRes 200) or Bb2 (NumRes204) in a 1: 1 ratio. Production of IL-10, TNF- � and IL-12 was measured in culture supernatant after 24 h ( a–c ), and of IFN- � and IL-13 after 72 h of stimulation ( d , e ). Data are shown as mean 8 SEM of 3 independent experiments. n.d. = Not de-tectable. * * p ! 0.01 vs. control stimulation in the absence of bacteria.
Table 1. Effect of UV-killed bacteria on cytokine secretion by non-stimulated PBMC in presence or absence of HT-29
IL-13 IFN-� IL-10 IL-12 TNF-�
PBMCNo bacteria 0.080.0 0.080.0 3.683.6 0.080.0 45.9831.9LGG 0.080.0 0.080.0 96.2828.5** 670.08246.9** 2,075.98784.9**Bb1 (NumRes 200) 0.080.0 0.080.0 57.8826.4** 503.28160.9** 1,048.18471.7**Bb2 (NumRes 204) 3.083.0 0.080.0 90.5828.9** 539.98184.9** 1,549.98574.8**
HT-29/PBMCNo bacteria 0.080.0 0.080.0 0.980.9 0.080.0 0.080.0LGG 0.080.0 0.080.0 1.181.1 0.080.0 0.080.0Bb1 (NumRes 200) 0.080.0 0.080.0 1.081.0 0.080.0 0.080.0Bb2 (NumRes 204) 4.184.1 0.080.0 1.581.5 13.1813.1 20.384.9
Cytokine concentrations are shown as pg/ml, mean 8 SEM of 3 independent experiments. ** p < 0.01 vs. cultures without bacte-ria.

van Hoffen et al.
Int Arch Allergy Immunol 2010;152:159–168164
Effects of Bacteria on Cytokine Secretion byIEC/PBMC Co-Cultures Direct addition of UV-killed LGG as well as Bb to un-
stimulated PBMC in a 1: 5 ratio was found to dramati-cally enhance IL-10, IL-12 and TNF- � secretion ( table 1 ). However, when these strains were added apically to IEC (HT-29) in a 10: 1 ratio, no cytokines were produced by unstimulated PBMC in the basolateral compartment af-ter 48 h of culture ( table 1 ), indicating that the bacteria did not leak to the basolateral compartment.
The presence of HT-29 reduced the production ofIL-10, IFN- � and IL-13 by anti-CD3/CD28-stimulated PBMC. Addition of LGG and Bb to the IEC was not able to reverse the reduction in IL-10 caused by the presence of HT-29. Hence, unlike direct incubations with the PBMC, the LGG and Bb did not enhance IL-10 when add-ed to IEC/PBMC co-cultures ( fig. 4 a). Similar to direct incubations with activated PBMC, UV-killed LGG was found to enhance TNF- � , IL-12 and IFN- � secretion by activated PBMC when added to HT-29. In contrast, the presence of HT-29 prevented Bb1 and Bb2 to exert this ef-
fect ( fig. 4 b–d). None of the probiotic strains further re-duced the IL-13 secretion as compared to HT-29 alone ( fig. 3 e). TGF- � 1 secretion by the activated PBMC was al-ready high (see above) and the probiotic bacteria did not affect those levels (data not shown). Concentrations of the epithelial-derived cytokine TSLP were below detection level.
Effects of TLR2 or NOD2 Ligands on Cytokine Secretion by IEC/PBMC Co-Cultures TLR2 and NOD2 are potential receptors for LGG on
IEC. Ligands of these receptors were added apically, in absence of bacteria, to simulate the effects of LGG and address the potential involvement of these receptors in immune modulation by HT-29 cells. Similar to LGG, Pam 3 Cys-SKKKK and MDP did not affect IL-10 and IL-13 secretion of activated PBMC in presence of HT-29 ( fig. 5 a, e). Pam 3 Cys-SKKKK was found to enhance IL-12 secretion of activated PBMC when added to HT-29, whereas MDP, but not its negative control peptide, en-hanced IFN- � and TNF- � secretion ( fig. 5 b–d).
HT/Bb2
**
103 105
104
103
102
101
100
102
101
100
104 104
103
102
101
100
103
102
101
100
103
102
101
100
IL-1
0 (p
g/m
l) #
TNF-
� (p
g/m
l)
#
IL-1
2 (p
g/m
l)
*
#
IFN
-� (p
g/m
l)
*
IL-1
3 (p
g/m
l)
Control HT HT/LGG HT/Bb1
HT/Bb2Control HT HT/LGG HT/Bb1
HT/Bb2Control HT HT/LGG HT/Bb1
HT/Bb2Control HT HT/LGG HT/Bb1
HT/Bb2Control HT HT/LGG HT/Bb1
a
c
e
d
b
Fig. 4. Effects of IEC and UV-killed LGG and Bb on cytokine secretion by IEC/PBMC co-cultures. In a transwell system, HT-29 cells were co-cultured with anti-CD3/CD28 stimulated PBMC for 48 h. Production of IL-10, TNF- � , IL-12, IFN- � and IL-13 in the basolateral compartment was measured after 48 h of co-culture. The data of 6 independent experiments are shown as mean 8 SEM. Control = CD3/CD28 stimulation of PBMC in the absence of HT-29; HT = CD3/CD28 stimulation of PBMC in the presence of HT-29; HT/ LGG , HT/ Bb1 , HT/ Bb2 = CD3/CD28 stimula-tion of PBMC in the presence of HT-29, with apical addition of the respective bac-terial strains. * p ! 0.05, * * p ! 0.01 vs. cul-tures without HT-29; # p ! 0.05 vs. cultures with HT-29.

Immune Modulation in a HT-29/PBMC Transwell Co-Culture Model
Int Arch Allergy Immunol 2010;152:159–168 165
Discussion
Emerging evidence suggests an immunomodulatory role for IEC that separate the external from the internal milieu, thereby contributing to mucosal homeostasis. Di-rect stimulation of PBMC with bacteria has often been used as a way to screen the immunomodulatory candi-date probiotic strains. Our data fit with previous studies showing that probiotics skew the T- cell response towards a Th1 and/or regulatory phenotype [17, 18] . Both the UV-killed LGG, Bb1 and Bb2 were effective in skewing the response of stimulated PBMC and resembled effects ob-served using the live mixture of probiotics, used as a ref-erence for its known capacities to polarize the immune response [16] . Indeed, it has been shown that Lactobacilli and Bifidobacteria , either live or heat-killed, can polarize the immune response in vitro [18–20] . However, in hu-mans it was shown that direct in vitro effects of probiot-ics on PBMC responses are discrepant from ex vivo or in vivo effects [16, 21] . This may be due to the fact that pro-biotics in vivo act via the intestinal mucosa, which differs from the direct contact between probiotics and PBMC in an in vitro system. Therefore, in this study immune mod-ulation by UV-killed single-strain LGG or Bb was com-pared when applied directly to PBMC or when added to intestinal epithelial cells co-cultured with non-stimulat-ed or activated PBMC.
Although LGG, Bb1 and Bb2 dramatically enhanced secretion of IL-10, IL-12 and TNF- � when added directly to non-stimulated PBMC, these bacteria did not induce any response when added apically to the IEC/PBMC co-culture. Hence, the bacteria remained within the apical compartment being unable to elicit an immune response by unstimulated PBMC via activation of IEC. In a similar model, the responsiveness of IEC was shown to be depen-dent of individual strain properties, since L. johnsonii and L. gasseri were not found to elicit an immune re-sponse in unstimulated PBMC while L. sakei was able to induce TNF- � and IL-10 secretion by PBMC via activa-tion of HT-29 and Caco-2 epithelial cells [9, 22] .
HT-29 cells did not affect cytokine secretion by un-stimulated PBMC. However, they dampened Th1 (IFN- � ), Th2 (IL-13) but also regulatory-type IL-10 secretion of anti-CD3/CD28-stimulated PBMC, used to simulate an effector immune response. It has been shown previously that IEC-conditioned medium is able to reduce lympho-cyte proliferation upon CD3 stimulation [23] . IL-10 is known to be involved in maintenance of homeostasis within the lamina propria and IEC or conditioned super-natant has been shown to enhance IL-10 secretion of
monocyte-derived DC [7, 24] . Although IEC can instruct DC to induce tolerogenic responses, under inflammatory conditions a dramatic reduction in IL-10 secretion was found by Salmonella -activated colonic DC as well as IEC-conditioned moDC, while TNF- � secretion was not af-fected [25, 26] . IEC can become activated when IL-10 se-cretion is neutralized [22] . Hence, HT-29 cells may se-crete mediators that selectively reduce IL-10 secretion by monocytes, enabling the IEC to actively participate in the effector immune response.
The frequently used human IEC lines HT-29 (model for colonic crypt cells) and Caco-2 (model for fully dif-ferentiated villus enterocytes) are known to possess re-ceptors for pro-inflammatory cytokines and secrete me-diators upon exposure to inflammatory mediators or mi-crobes [9, 10, 24, 27, 28] . However, HT-29 were shown to be more responsive than Caco-2 cells when co-cultured with unstimulated PBMC and apically exposed to Esch-erichia coli or Lactobacillus sakei [9] . Both cell lines have been shown to express TLRs 1–4, although expression of TLR-2 co-receptors TLR-1 and -6 was very low or absent in Caco-2 compared to HT-29 [29, 30] . In the current study, bacteria-exposed HT-29 cells were not found to af-fect the immune response, whereas upon activation of the PBMC, simulating a mucosal effector response, differen-tial effects were observed between UV-killed LGG and Bb . Intestinal inflammation results in epithelial regen-eration coinciding with loss of epithelial cell polarity [31] . Hence, HT-29 cells may be of particular relevance in this HT-29/activated PBMC coculture model.
Although no differences were found between LGG and Bb when added directly to PBMC, in the co-culture mod-el only LGG potentiated the effector response. Lactoba-cilli were shown to have a differential potential compared to Bifidobacteria to activate epithelial cells. Live and UV-killed Lactobacilli, in particular LGG, were found to en-hance TSLP secretion by IEC while Bifidobacteria did not [24] . Hence Bifidobacteria may have lesser capacities than Lactobacilli in provoking IEC/immune cell cross-talk. IEC are known to express a whole panel of chemokines and cytokines [9, 10, 24, 27, 28] and apical LGG exposure may have contributed to the release of mediators that po-tentiated Th1, B cell and/or monocyte-derived cytokine responses. The enhanced TNF- � may partially have been derived from HT-29 since IEC are known to express TNF- � when activated [24, 28] . In particular, TSLP and TGF- � have been shown to modulate the IEC-immune cell cross-talk [24, 26] . However, in our model, TSLP concentrations were below detectable levels and TGF- � 1 did not differ between the treatments.

van Hoffen et al.
Int Arch Allergy Immunol 2010;152:159–168166
Although LGG is able to enhance IL-10 secretion dur-ing direct incubation with activated PBMC, in the co-cul-ture model LGG did not reverse the suppression of IL-10 that occurred in presence of IEC. Also, in the lamina propria of virus-infected mice several Lactobacilli were found to effectively enhanced TNF- � secretion; however, only L. casei tended to enhance the number of IL-10-pos-itive cells [32] . Especially in preventive study designs, in vivo probiotic supplementation with LGG and L. reuteri have been shown to reduce the incidence of atopic derma-titis in high-risk infants [4–6] . LGG was effective in re-ducing atopic eczema, but did not enhance plasma IL-10 levels [33, 34] . It has been proposed that induction of low-grade inflammation contributes to the effect exerted by LGG [34–36] .
Both live as well as UV-killed bacteria and bacterial fragments can induce changes in cytokine production by PBMC [20] . IEC show low homeostatic TLR expression [29] . However, under inflammatory conditions, apical TLR and intracellular NOD2 expression, as well as pep-tide transporter 1 (hPepT1) membrane expression re-
quired for cellular uptake of NOD2 ligand MDP, are in-duced in the intestinal epithelium [15, 26, 37, 38] . In the co-culture model, TLR2 (Pam 3 Cys-SKKKK) and NOD2 ligand were found to potentiate the effector immune re-sponse when added apically to IEC. The TLR2 ligand was found to enhance IL-12 in the presence of HT-29 cells, while the NOD2 ligand enhanced TNF- � and IFN- � . Al-though only LGG significantly enhanced IL-12 secretion upon apical exposure of IEC, Bb1 and Bb2 showed the same tendency. However, in contrast to LGG and NOD2 ligand, Bb1 and Bb2 did not have any effect on TNF- � and IFN- � secretion. This may imply that Lactobacilli have the capacity to signal via NOD2 whereas Bb are not effec-tive in signaling via this route in the co-culture model. Although differential NOD2 signaling between Lactoba-cilli and Bifidobacteria is not known from literature, in-deed NOD2 signaling and uptake of MDP containing peptidoglycans by hPepT1 have been shown to be size and structure dependent [39, 40] .
In conclusion, the present data show that UV-killed LGG or Bb1 and Bb2 , like the reference mixture of live
HT/MDP–
103
102
101
100
104
103
102
101
100
105
104
103
102
101
100
104
103
102
101
100
103
102
101
100
IL-1
0 (p
g/m
l)
*
TNF-
� (p
g/m
l)
*
IL-1
2 (p
g/m
l) **
IFN
-� (p
g/m
l)
IL-1
3 (p
g/m
l)
HT HT/Pam3Cys HT/MDP+ HT/MDP–HT HT/Pam3Cys HT/MDP+
HT/MDP–HT HT/Pam3Cys HT/MDP+ HT/MDP–HT HT/Pam3Cys HT/MDP+
HT/MDP–HT HT/Pam3Cys HT/MDP+
a
c
b
d
e
Fig. 5. TLR2 and NOD2 are potential re-ceptors for LGG on IEC. The effect of TLR2 and NOD2 ligands Pam 3 Cys-SKKKK and MDP on cytokine secretion in the co-culture of HT-29 with CD3/CD28-stimulated PBMC was analyzed. Pam 3 Cys-SKKKK, MDP (MDP+) and the MDP negative control (MDP–) were added to the apical compartment during the co-culture. Cytokines in the basolateral com-partment of the co-culture of HT-29 cells with activated PBMC were measured after 48 h of incubation. The data of 5 indepen-dent experiments are shown as mean 8 SEM. Control (HT) = CD3/CD28-stimu-lated PBMC in the presence of HT-29 in the absence of TLR2/NOD2 ligands. * p ! 0.05, * * p ! 0.01 vs. control.

Immune Modulation in a HT-29/PBMC Transwell Co-Culture Model
Int Arch Allergy Immunol 2010;152:159–168 167
bacteria, provide the typical regulatory type of Th1 po-larization when incubated directly with PBMC, while differential effects were found in the HT-29/activated PBMC co-culture. IEC themselves were found to dampen the effector response. Although in the presence of HT-29 cells, UV-killed LGG and Bb strains lost the capacity to enhance regulatory IL-10 secretion, UV-killed LGG but
not the Bb strains, were still able to enhance the Th1 type effector response when added to IEC. The co-culture model provides additional information on immune ef-fects of different candidate probiotic strains since the modulatory role of IEC is included. Therefore, it may pro-vide a useful tool to predict the in vivo effectiveness of probiotic strains.
References
1 Bischoff S, Crowe SE: Gastrointestinal food allergy: new insights into pathophysiology and clinical perspectives. Gastroenterology 2005; 128: 1089–1113.
2 Laiho K, Hoppu U, Ouwehand AC, Salminen S, Isolauri E: Probiotics: on-going research on atopic individuals. Br J Nutr 2002; 88 (suppl 1):S19–S27.
3 Isolauri E, Majamaa H, Arvola T, Rantala I, Virtanen E, Arvilommi H: Lactobacillus ca-sei strain GG reverses increased intestinal permeability induced by cow milk in suck-ling rats. Gastroenterology 1993; 105: 1643–1650.
4 Savilahti E, Kuitunen M, Vaarala O: Pre-and probiotics in the prevention and treat-ment of food allergy. Curr Opin Allergy Clin Immunol 2008; 8: 243–248.
5 Abrahamsson TR, Jakobsson T, Bottcher MF, Fredrikson M, Jenmalm MC, Bjorksten B, Oldaeus G: Probiotics in prevention of IgE-associated eczema: a double-blind, ran-domized, placebo-controlled trial. J Allergy Clin Immunol 2007; 119: 1174–1180.
6 Prescott SL, Bjorksten B: Probiotics for the prevention or treatment of allergic diseases. J Allergy Clin Immunol 2007; 120: 255–262.
7 Mowat AM: Anatomical basis of tolerance and immunity to intestinal antigens. Nat Rev Immunol 2003; 3: 331–341.
8 Kagnoff MF, Eckmann L: Epithelial cells as sensors for microbial infection. J Clin Invest 1997; 100: 6–10.
9 Haller D, Bode C, Hammes WP, Pfeifer AM, Schiffrin EJ, Blum S: Non-pathogenic bacte-ria elicit a differential cytokine response by intestinal epithelial cell/leucocyte co-cul-tures. Gut 2000; 47: 79–87.
10 Hormannsperger G, Clavel T, Hoffmann M, Reiff C, Kelly D, Loh G, Blaut M, Holzlwim-mer G, Laschinger M, Haller D: Post-trans-lational inhibition of IP-10 secretion in IEC by probiotic bacteria: impact on chronic in-flammation. PLoS One 2009; 4:e4365.
11 Iliev ID, Spadoni I, Mileti E, Matteoli G, Son-zogni A, Sampietro GM, Foschi D, Caprioli F, Viale G, Rescigno M: Human intestinal epithelial cells promote the differentiation of tolerogenic dendritic cells. Gut 2009, E-pub ahead of print.
12 Vinderola G, Matar C, Perdigon G: Role of intestinal epithelial cells in immune effects mediated by gram-positive probiotic bacte-ria: involvement of toll-like receptors. Clin Diagn Lab Immunol 2005; 12: 1075–1084.
13 Zoumpopoulou G, Tsakalidou E, Dewulf J, Pot B, Grangette C: Differential crosstalk between epithelial cells, dendritic cells and bacteria in a co-culture model. Int J Food Microbiol 2009; 131: 40–51.
14 Strober W, Murray PJ, Kitani A, Watanabe T: Signalling pathways and molecular interac-tions of NOD1 and NOD2. Nat Rev Immunol 2006; 6: 9–20.
15 Forchielli ML, Walker WA: The role of gut-associated lymphoid tissues and mucosal de-fence. Br J Nutr 2005; 93(suppl 1):S41–S48.
16 Flinterman AE, Knol EF, van Ieperen-van Dijk AG, Timmerman HM, Knulst AC, Bruijnzeel-Koomen CA, Pasmans SG, van Hoffen E: Probiotics have a different immu-nomodulatory potential in vitro versus ex vivo upon oral administration in children with food allergy. Int Arch Allergy Immunol 2007; 143: 237–244.
17 Mohamadzadeh M, Olson S, Kalina WV, Ruthel G, Demmin GL, Warfield KL, Bavari S, Klaenhammer TR: Lactobacilli activate human dendritic cells that skew T cells to-ward T helper 1 polarization. Proc Natl Acad Sci USA 2005; 102: 2880–2885.
18 Niers LE, Timmerman HM, Rijkers GT, van Bleek GM, van Uden NO, Knol EF, Kapsen-berg ML, Kimpen JL, Hoekstra MO: Identi-fication of strong interleukin-10 inducing lactic acid bacteria which down-regulate T helper type 2 cytokines. Clin Exp Allergy 2005; 35: 1481–1489.
19 Iwabuchi N, Takahashi N, Xiao JZ, Miyaji K, Iwatsuki K: In vitro Th1 cytokine-indepen-dent Th2 suppressive effects of bifidobacte-ria. Microbiol Immunol 2007; 51: 649–660.
20 Medina M, Izquierdo E, Ennahar S, Sanz Y: Differential immunomodulatory properties of Bifidobacterium logum strains: relevance to probiotic selection and clinical applica-tions. Clin Exp Immunol 2007; 150: 531–538.
21 Kopp MV, Goldstein M, Dietschek A, Sofke J, Heinzmann A, Urbanek R: Lactobacillus GG has in vitro effects on enhanced interleu-kin-10 and interferon-gamma release of mononuclear cells but no in vivo effects in supplemented mothers and their neonates. Clin Exp Allergy 2008; 38: 602–610.
22 Haller D, Serrant P, Peruisseau G, Bode C, Hammes WP, Schiffrin E, Blum S: IL-10 producing CD14 low monocytes inhibit lym-phocyte-dependent activation of intestinal epithelial cells by commensal bacteria. Mi-crobiol Immunol 2002; 46: 195–205.
23 Christ AD, Colgan SP, Balk SP, Blumberg RS: Human intestinal epithelial cell lines pro-duce factor(s) that inhibit CD3-mediated T-lymphocyte proliferation. Immunol Lett 1997; 58: 159–165.
24 Zeuthen LH, Fink LN, Frokiaer H: Epithelial cells prime the immune response to an array of gut-derived commensals towards a tolero-genic phenotype through distinct actions of thymic stromal lymphopoietin and trans-forming growth factor-beta. Immunology 2008; 123: 197–208.
25 Baba N, Samson S, Bourdet-Sicard R, Rubio M, Sarfati M: Commensal bacteria trigger a full dendritic cell maturation program that promotes the expansion of non-Tr1 suppres-sor T cells. J Leukoc Biol 2008; 84: 468–476.
26 Rimoldi M, Chieppa M, Salucci V, Avogadri F, Sonzogni A, Sampietro GM, Nespoli A, Viale G, Allavena P, Rescigno M: Intestinal immune homeostasis is regulated by the crosstalk between epithelial cells and den-dritic cells. Nat Immunol 2005; 6: 507–514.
27 Berin MC, Dwinell MB, Eckmann L, Kag-noff MF: Production of MDC/CCL22 by hu-man intestinal epithelial cells. Am J Physiol Gastrointest Liver Physiol 2001; 280:G1217–G1226.
28 O’Hara AM, O’Regan P, Fanning A, O’Mahony C, Macsharry J, Lyons A, Bienen-stock J, O’Mahony L, Shanahan F: Function-al modulation of human intestinal epithelial cell responses by Bifidobacterium infantis and Lactobacillus salivarius . Immunology 2006; 118: 202–215.

van Hoffen et al.
Int Arch Allergy Immunol 2010;152:159–168168
29 Furrie E, Macfarlane S, Thomson G, Macfar-lane GT: Toll-like receptors-2, -3 and -4 ex-pression patterns on human colon and their regulation by mucosal-associated bacteria. Immunology 2005; 115: 565–574.
30 Melmed G, Thomas LS, Lee N, Tesfay SY, Lu-kasek K, Michelsen KS, Zhou Y, Hu B, Ar-diti M, Abreu MT: Human intestinal epithe-lial cells are broadly unresponsive to Toll-like receptor 2-dependent bacterial ligands: im-plications for host-microbial interactions in the gut. J Immunol 2003; 170: 1406–1415.
31 Hanby AM, Chinery R, Poulsom R, Playford RJ, Pignatelli M: Downregulation of E-cad-herin in the reparative epithelium of the hu-man gastrointestinal tract. Am J Pathol 1996; 148: 723–729.
32 Maassen CB, van Holten-Neelen C, Balk F, den Bak-Glashouwer MJ, Leer RJ, Laman JD, Boersma WJ, Claassen E: Strain-dependent induction of cytokine profiles in the gut by orally administered Lactobacillus strains. Vaccine 2000; 18: 2613–2623.
33 Viljanen M, Kuitunen M, Haahtela T, Jun-tunen-Backman K, Korpela R, Savilahti E: Probiotic effects on faecal inflammatory markers and on faecal IgA in food allergic atopic eczema/dermatitis syndrome infants. Pediatr Allergy Immunol 2005; 16: 65–71.
34 Viljanen M, Pohjavuori E, Haahtela T, Kor-pela R, Kuitunen M, Sarnesto A, Vaarala O, Savilahti E: Induction of inflammation as a possible mechanism of probiotic effect in atopic eczema-dermatitis syndrome. J Aller-gy Clin Immunol 2005; 115: 1254–1259.
35 Marschan E, Kuitunen M, Kukkonen K, Poussa T, Sarnesto A, Haahtela T, Korpela R, Savilahti E, Vaarala O: Probiotics in infancy induce protective immune profiles that are characteristic for chronic low-grade inflam-mation. Clin Exp Allergy 2008; 38: 611–618.
36 Pohjavuori E, Viljanen M, Korpela R, Kui-tunen M, Tiittanen M, Vaarala O, Savilahti E: Lactobacillus GG effect in increasing IFN-gamma production in infants with cow’s milk allergy. J Allergy Clin Immunol 2004; 114: 131–136.
37 Begue B, Dumant C, Bambou JC, Beaulieu JF, Chamaillard M, Hugot JP, Goulet O, Schmitz J, Philpott DJ, et al: Microbial in-duction of CARD15 expression in intestinal epithelial cells via toll-like receptor 5 trig-gers an antibacterial response loop. J Cell Physiol 2006; 209: 241–252.
38 Charrier L, Merlin D: The oligopeptide transporter hPepT1:gateway to the innate immune response. Lab Invest 2006; 86: 538–546.
39 Girardin SE, Travassos LH, Herve M, Blanot D, Boneca IG, Philpott DJ, Sansonetti PJ, Mengin-Lecreulx D: Peptidoglycan molecu-lar requirements allowing detection by Nod1 and Nod2. J Biol Chem 2003; 278: 41702–41708.
40 Ismair MG, Vavricka SR, Kullak-Ublick GA, Fried M, Mengin-Lecreulx D, Girardin SE: hPepT1 selectively transports muramyl di-peptide but not Nod1-activating muramyl peptides. Can J Physiol Pharmacol 2006; 84: 1313–1319.




















