
Recombinant expression of Pleurotus ostreatus laccases in Kluyveromyces lactis and Saccharomyces...
12
Appl Microbiol Biotechnol (2005) 69: 428–439 DOI 10.1007/s00253-005-0004-z APPLIED GENETICS AND MOLECULAR BIOTECHNOLOGY Alessandra Piscitelli . Paola Giardina . Cristina Mazzoni . Giovanni Sannia Recombinant expression of Pleurotus ostreatus laccases in Kluyveromyces lactis and Saccharomyces cerevisiae Received: 4 February 2005 / Revised: 14 April 2005 / Accepted: 15 April 2005 / Published online: 14 July 2005 # Springer-Verlag 2005 Abstract Heterologous expression of Pleurotus ostreatus POXC and POXA1b laccases in two yeasts, Kluyveromyces lactis and Saccharomyces cerevisiae, was performed. Both transformed hosts secreted recombinant active laccases, although K. lactis was much more effective than S. cere- visiae. rPOXA1b transformants always had higher secreted activity than rPOXC transformants did. The lower tendency of K. lactis with respect to S. cerevisiae to hyperglycosy- late recombinant proteins was confirmed. Recombinant laccases from K. lactis were purified and characterised. Specific activities of native and recombinant POXA1b are similar. On the other hand, rPOXC specific activity is much lower than that of the native protein, perhaps due to in- complete or incorrect folding. Both recombinant laccase signal peptides were correctly cleaved, with rPOXA1b protein having two C-terminal amino acids removed. The availability of the established recombinant expression sys- tem provides better understanding of laccase structure– function relationships and allows the development of new oxidative catalysts through molecular evolution techniques. Introduction Laccases (E.C.1.10.3.2, p-benzenediol:oxygen oxidore- ductase) belong to the blue oxidase family of proteins, along with ascorbic oxidases and ceruloplasmins. These copper-containing enzymes (multicopper oxidases) couple the oxidation of various substrates with the simultaneous reduction of molecular oxygen to water (Yaropolov et al. 1994). Laccases are widely distributed in higher plants and fungi, and laccase-like activity has been found in some insects (Dittmer et al. 2004; Kramer et al. 2001) and bacte- ria (Claus 2003). Roles attributed to these enzymes include pigment production in fungi and bacteria (Aramayo and Timberlake 1993; Williamson et al. 1998), lignification and delignification of plant cell walls (O’ Malley et al. 1993), virulence factors (Williamson et al. 1998), and copper and iron homeostasis (Stoj and Kosman 2003). Laccases con- tain four copper atoms central to the enzyme catalytic mechanisms (Yaropolov et al. 1994), distributed between a mononuclear site (one type 1 copper ion) and a trinu- clear site (one type 2 and two type 3 copper ions). Laccases have a very broad substrate range and can ox- idize polyphenols, aminophenols, methoxyphenols and aryl amines and are being increasingly evaluated for use in a variety of applications including the pulp and paper in- dustry, bioremediation, and for analytical purposes (Mayer et al. 2002). The white-rot fungus Pleurotus ostreatus expresses mul- tiple laccase genes encoding isoenzymes with different physico-chemical and catalytic characteristics. The amount of the various enzymes produced depends on culture con- ditions. So far, five isoenzymes secreted by this fungus have been purified and characterised: POXC (Palmieri et al. 1993), POXA1w (Palmieri et al. 1997), POXA1b (Giardina et al. 1999) and, more recently, the two closely related iso- enzymes POXA3a and POXA3b (Palmieri et al. 2003). POXC is the most abundantly produced in all growth con- ditions tested so far; POXA1w has unusual metal ion content; POXA1b is the most stable at alkaline pH; and POXA3a and POXA3b are heterodimers. Four genes en- coding laccases are known, poxc (previously named pox2) (Giardina et al. 1996), pox1 (which codes for a laccase isoenzyme not identified yet) (Giardina et al. 1995), poxa1b and poxa3 (Giardina et al. 1999; Palmieri et al. 2003). The achievement of tailored enzymes for different in- dustrial needs (i.e. high stability, wide variety of substrates, A. Piscitelli . P. Giardina . G. Sannia (*) Dipartimento di Chimica Organica e Biochimica, Università di Napoli “Federico II”, Complesso Universitario Monte S. Angelo, via Cinthia, 80126 Naples, Italy e-mail: [email protected] Tel.: +39-081-674310 Fax: +39-081-674313 C. Mazzoni Dipartimento di Biologia Cellulare e dello Sviluppo, Università di Roma “La Sapienza”, Piazzale Aldo Moro, 5, 00185 Rome, Italy
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Recombinant expression of Pleurotus ostreatus laccases in Kluyveromyces lactis and Saccharomyces...

Appl Microbiol Biotechnol (2005) 69: 428–439DOI 10.1007/s00253-005-0004-z
APPLIED GENETICS AND MOLECULAR BIOTECHNOLOGY
Alessandra Piscitelli . Paola Giardina .Cristina Mazzoni . Giovanni Sannia
Recombinant expression of Pleurotus ostreatus laccasesin Kluyveromyces lactis and Saccharomyces cerevisiae
Received: 4 February 2005 / Revised: 14 April 2005 / Accepted: 15 April 2005 / Published online: 14 July 2005# Springer-Verlag 2005
Abstract Heterologous expression of Pleurotus ostreatusPOXC and POXA1b laccases in two yeasts,Kluyveromyceslactis and Saccharomyces cerevisiae, was performed. Bothtransformed hosts secreted recombinant active laccases,although K. lactis was much more effective than S. cere-visiae. rPOXA1b transformants always had higher secretedactivity than rPOXC transformants did. The lower tendencyof K. lactis with respect to S. cerevisiae to hyperglycosy-late recombinant proteins was confirmed. Recombinantlaccases from K. lactis were purified and characterised.Specific activities of native and recombinant POXA1b aresimilar. On the other hand, rPOXC specific activity is muchlower than that of the native protein, perhaps due to in-complete or incorrect folding. Both recombinant laccasesignal peptides were correctly cleaved, with rPOXA1bprotein having two C-terminal amino acids removed. Theavailability of the established recombinant expression sys-tem provides better understanding of laccase structure–function relationships and allows the development of newoxidative catalysts through molecular evolution techniques.
Introduction
Laccases (E.C.1.10.3.2, p-benzenediol:oxygen oxidore-ductase) belong to the blue oxidase family of proteins,along with ascorbic oxidases and ceruloplasmins. These
copper-containing enzymes (multicopper oxidases) couplethe oxidation of various substrates with the simultaneousreduction of molecular oxygen to water (Yaropolov et al.1994). Laccases are widely distributed in higher plants andfungi, and laccase-like activity has been found in someinsects (Dittmer et al. 2004; Kramer et al. 2001) and bacte-ria (Claus 2003). Roles attributed to these enzymes includepigment production in fungi and bacteria (Aramayo andTimberlake 1993; Williamson et al. 1998), lignification anddelignification of plant cell walls (O’ Malley et al. 1993),virulence factors (Williamson et al. 1998), and copper andiron homeostasis (Stoj and Kosman 2003). Laccases con-tain four copper atoms central to the enzyme catalyticmechanisms (Yaropolov et al. 1994), distributed betweena mononuclear site (one type 1 copper ion) and a trinu-clear site (one type 2 and two type 3 copper ions).
Laccases have a very broad substrate range and can ox-idize polyphenols, aminophenols, methoxyphenols and arylamines and are being increasingly evaluated for use in avariety of applications including the pulp and paper in-dustry, bioremediation, and for analytical purposes (Mayeret al. 2002).
The white-rot fungus Pleurotus ostreatus expresses mul-tiple laccase genes encoding isoenzymes with differentphysico-chemical and catalytic characteristics. The amountof the various enzymes produced depends on culture con-ditions. So far, five isoenzymes secreted by this fungushave been purified and characterised: POXC (Palmieri et al.1993), POXA1w (Palmieri et al. 1997), POXA1b (Giardinaet al. 1999) and, more recently, the two closely related iso-enzymes POXA3a and POXA3b (Palmieri et al. 2003).POXC is the most abundantly produced in all growth con-ditions tested so far; POXA1w has unusual metal ioncontent; POXA1b is the most stable at alkaline pH; andPOXA3a and POXA3b are heterodimers. Four genes en-coding laccases are known, poxc (previously named pox2)(Giardina et al. 1996), pox1 (which codes for a laccaseisoenzyme not identified yet) (Giardina et al. 1995), poxa1band poxa3 (Giardina et al. 1999; Palmieri et al. 2003).
The achievement of tailored enzymes for different in-dustrial needs (i.e. high stability, wide variety of substrates,
A. Piscitelli . P. Giardina . G. Sannia (*)Dipartimento di Chimica Organica e Biochimica,Università di Napoli “Federico II”,Complesso Universitario Monte S. Angelo,via Cinthia,80126 Naples, Italye-mail: [email protected].: +39-081-674310Fax: +39-081-674313
C. MazzoniDipartimento di Biologia Cellulare e dello Sviluppo,Università di Roma “La Sapienza”,Piazzale Aldo Moro, 5,00185 Rome, Italy

high efficiency in catalysis, etc.) through recombinant ex-pression and directed evolution would be of special interest.So far, heterologous expression of laccases has been at-tained in Saccharomyces cerevisiae (Cassland and Jonsson1999; Kiiskinen and Saloheimo 2004; Klonowska et al.2005; Kojima et al. 1990), Pichia pastoris (Gelo-Pujic etal. 1999; Jonsson et al. 1997; Liu et al. 2003; Otterbein etal. 2000; Soden et al. 2002), Yarrowia lipolytica (Jolivaltet al. 2005), Trichoderma reesei (Kiiskinen et al. 2004),Aspergillus oryzae (Berka et al. 1997; Sigoillot et al. 2004),Aspergillus sojae (Hatamoto et al. 1999), Aspergillus niger(Larrondo et al. 2003; Record et al. 2002) and Aspergillusnidulans (Larrondo et al. 2003). Filamentous fungi are thebest producers of recombinant laccases, with protein yieldsranging from 70 (Record et al. 2002) to 230 mg l−1
(Kiiskinen et al. 2004). However, yeasts are the organismsof choice to create new enzymes with desirable physico-chemical characters, these microorganisms being easilymanipulable and more cost-effective hosts; for example,the directed evolution of a Myceliophthora thermophilalaccase expressed in S. cerevisiae has been reported (Bulteret al. 2003). Kluyveromyces lactis is also being used asan expression system for the secretion of a number ofeukaryotic proteins, proving to be more efficient than S.cerevisiae (Hsieh and Da Silva 1998), with reported yieldsranging from 20 mg l−1 to a few grams per liter (Bergquistet al. 2002); other advantages are faster growth and lack ofhyperglycosylation of proteins (Hsieh and Da Silva 1998).
In this paper we describe for the first time the recom-binant expression of two P. ostreatus laccases, POXC andPOXA1b, in both K. lactis and S. cerevisiae, and compar-ison of the two systems in laccase production.
Materials and methods
Strains, media and plasmids
The Escherichia coli strain Top 10 (F-mcrA D (mrr-hsdRMS-mcrBC) f80lacZDM15 DlacX74 deoR recA1araD139 D (ara-leu) 7697 galU galK rpsL (StrR) endA1nupG) was used in all DNA manipulations. E. coli wasgrown in Luria–Bertani (LB) medium (in g l−1: 10 bacto
tryptone, 10 NaCl, 5 yeast extract), supplemented, whenrequired, with 100 μg ml−1 of ampicillin.
The K. lactis strain used for heterologous expression wasCMK5 (a thr lys pgi1 adh3 adh1URA3 adh2URA3) (Saliolaet al. 1999). Plasmid pYG132 (Saliola et al. 1999) wasengineered from pKD1, a plasmid originally isolated fromKluyveromyces drosophilarum (Falcone et al. 1986) thatcan replicate stably in K. lactis. Both K. lactis strain andplasmid were kindly given by Prof. C. Falcone (University“La Sapienza”, Rome, Italy). Insert expression is controlledby the ethanol-inducible KlADH4 promoter (Mazzoni et al.1992) and the S. cerevisiae phosphoglycerate kinase (PGK)terminator. K. lactis was grown in different media, sup-plemented, when required, with 100 μg ml−1 of geneticinG418: YPPD (in g l−1: 10 yeast extract, 40 bacto tryptone,20 glucose); YPD (in g l−1: 10 yeast extract, 20 bacto tryp-tone, 20 glucose); YPPG (in g l−1: 10 yeast extract, 40 bactotryptone, 20 galactose); and YPPDG (in g l−1: 10 yeastextract, 40 bacto tryptone, 20 glucose, 20 galactose).
The S. cerevisiae strain used for heterologous expressionwasW303-1A (MATade2-1, his3-11, 15, leu2-3, 112, trp1-1, ura3-1, can1-100). The plasmids used for S. cerevisiaeexpression were pAC1 (ADH1 constitutive promoter),pSAL4 (copper-inducible CUP1 promoter) and YEp352(PGK1 constitutive promoter) (Hill et al. 1986). All of themcarry URA3 gene for auxotrophic selection. S. cerevisiaestrain and plasmids were kindly given by Dr. T. Tron(CNRS UMR 6517, Marseille, France). S. cerevisiae wasgrown on a selective medium [6.7 g l−1 yeast nitrogen basewithout amino acids and ammonium sulfate, 5 g l−1 casa-mino acids, 30mg l−1 adenine, 40mg l−1 tryptophan, 50mMsuccinate buffer (pH 5.3)] containing glucose or galactose:SD (selective medium containing 20 g l−1 glucose), Sd (se-lective medium containing 10 g l−1 glucose), SG (selectivemedium containing 20 g l−1 galactose) and Sg (selectivemedium containing 10 g l−1 galactose).
Vector construction
The GenBank accession numbers of the sequences of theP. ostreatus laccase cDNAs poxc (Giardina et al. 1996)and poxa1b (Giardina et al. 1999) reported in this paperare Z34848 and AJ005018, respectively.
Table 1 List of the primers usedin the amplification experiments
Nucleotides in bold are comple-mentary to the laccase cDNAsequences. Nucleotides in italicsare recognized by restrictionenzymes
Primer Nucleotide sequences Annealingtemperature (°C)
Fwpoxc TTTGAATTCAAGCTTATGTTTCCAGGCGCACGG 60Revpoxc GGCCACGATATGGTACCAATCC 60Fwpoxa1b ATAAGAATTCAAGCTTATGGCGGTTGCATTCG 54Revpoxa1b GTGTGCCCGACAAGATCTGCAACAAC 54A1bfw AAAAGAATTCATGCATCTAGCATTGGGCCCCGCGG 54A1brev TAAGGATCCGAATTCTTATAATCATGCTTTCAATGGC 54FwBglII TGTTGCAGATCTTGTCGG 54Cfw AAAAGAATTCATGCATCTGCCATTGGGCCCGCTGGC 60FwEcorV AGCCGATATCAACATCAATCTCG 60Crev AAAATCTAGAGAATTCTTAAGAAGTAGGAATAAGAAG 60
429

The 5′ regions of both cDNAs were amplified (Pfu poly-merase, Stratagene) with Fwpoxc/Revpoxc and Fwpoxa1b/Revpoxa1b primers (Table 1), using pUC13 containingPOXC cDNA (Cpox13) and pUC18 containing POXA1bcDNA (A1bpox18), respectively, as templates in order toinsert a HindIII site (Fig. 1 and Fig. 2, step 1). The clonedPCR products were checked by sequencing. The HindIIIDNA fragments containing the laccase encoding sequences,including their signal peptides, were ligated into pYG132(Fig. 1 and Fig. 2, step 2), giving pYCps and pYA1bpsvectors. To insert the laccase cDNAs in the expressionvector pSAL4, the 3′ regions of both cDNAswere amplifiedwith FwEcoRV/Crev and FwBglII/A1brev primers thatallowed insertion of an EcoRI sequence (Fig. 1 and Fig. 2,step 3). The EcoRI DNA fragments containing the laccase-encoding sequences, including their signal peptides, wereligated into the expression vectors (Fig. 1 and Fig. 2, step 4),giving C-pSAL4 andB-pSAL4 vectors. The same strategies
were exploited to clone laccase cDNAs into pAC1 plasmids(step 4), obtaining C-pAC1 and B-pAC1 vectors.
Vectors in which the endogenous signal peptide codingsequences were replaced with those of K. lactis killer toxin(α subunit) (Stark and Boyd 1986) or S. cerevisiae invertase(SUC2) were constructed. A synthetic DNA fragment cor-responding to the presequence (16 amino acids) of theK. lactis killer toxin (α subunit) was ligated to the DNAfragment carrying the poxc mature cDNA. A HindIII frag-ment containing the K. lactis killer toxin sequence in framewith the poxc cDNA was subcloned into pYG132, givingpYCtk vector.
The 5′ regions of both mature cDNAs were amplifiedwith Cfw/Revpoxc and A1bfw/Revpoxa1b primers, usingCpox13 and A1bpox18, respectively, as templates. Thefragments containing the invertase signal peptide encodingsequence in frame with the cDNAs were subcloned intoYEp 352, giving C-YEp and B-YEp vectors.
Fig. 1 Construction of poxc expression vectors. pYCps, pYG132vector containing poxc cDNA with its signal peptide; C-pSAL4,pSAL4 vector containing poxc cDNA with its signal peptide. Se-lection markers, promoter regions and replication origins are indi-
cated. The hatched region encodes the native signal peptide. Fwpoxc/Revpoxc and FwEcoRV/Crev are the primers used to amplify poxccDNA. H HindIII, E EcoRI, K KpnI, X XbaI, EV EcoRV
430

Yeast transformation, cultivation and laccaseproduction
K. lactis transformation was performed by electroporationwith a Bio-Rad Micro-Pulser apparatus, as specified by themanufacturer. The cells were spread on YPPD mediumcontaining 100 μg ml−1 of geneticin G418, after an over-night incubation at 28°C in YPPD. Agar plate assays onYPPD supplemented with 100 μg ml−1 of geneticin G418,2% ethanol, 0.1 mMCuSO4 and 0.2mMABTS [2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)] were used toselect transformants. Plates were incubated upside down for4 days at 28°C and checked for the development of greencolour. One hundred microliters of ethanol was added eachday to the lid of the plate to compensate for its evaporation.Thirty transformed clones for each expression vector werescreened for laccase production by growing them in 10 mlof selective medium (YPPD containing geneticin) supple-mented with 0.1 mM CuSO4 and 2% ethanol, at 28°C on arotary shaker (150 rpm.). Two percent ethanol was added
daily and samples were taken at intervals to measure opticaldensity and laccase activity. The best producing cloneswere chosen to study laccase production. Inocula grown onselective YPPD medium were used to seed flasks (250 ml)containing 50 ml of selective medium, starting from 0.06OD600. Cultures were grown at 28°C on a rotary shaker for4 days. Preparative culture was done in 1-l flasks containing200 ml of medium.
S. cerevisiae transformation was done by using a lithiumacetate protocol (Gietz et al. 1992). The cells were spreadon the selective medium and the plates incubated upsidedown for 4 days at 28°C. Laccase production by trans-formants was first assayed on SD plates supplemented with0.6 mM CuSO4 and 0.2 mM ABTS. Inocula grown onselective SD medium were used to seed flasks (250 ml)containing 50 ml of selective medium, starting from 0.5OD600. Cultures were grown at 28°C on a rotary shakerat 150 rpm. Preparative culture was done in 1-l flasks con-taining 200 ml of medium.
Fig. 2 Construction of poxa1b expression vectors. pYA1bps, pYG132vector containing poxa1b cDNA with its signal peptide; B-pSAL4,pSAL4 vector containing poxa1b cDNA with its signal peptide. Se-lection markers, promoter regions and replication origins are indi-
cated. The hatched region encodes the native signal peptide. Fwpoxa1b/Revpoxa1b, FwBglII/A1brev, are the primers used to amplify poxa1bcDNA. H HindIII, E EcoRI, B BglII, Bh BamHI
431

Assay of laccase activity
Culture aliquots (1 ml) were collected daily and cells werepelleted by centrifugation (12,000×g for 2 min at 4°C).Laccase activity in the culture supernatant was assayed at25°C by monitoring the oxidation of ABTS at 420 nm(ɛ420=3.6×10
4 M−1 cm−1). The assay mixture contained2 mMABTS, 0.1M sodium citrate buffer (pH 3.0). Laccaseactivity towards 2,6-dimethoxyphenol (DMP) was assayedin a mixture containing 1 mM DMP and McIlvaine’scitrate–phosphate buffer adjusted to pH 5.3. Oxidation ofDMP was followed by an increase in A477 (ɛ477=1.48×10
4
M−1 cm−1). Activity was measured using ABTS as sub-strate unless otherwise stated.
Electrophoresis and isoelectric focusing
Polyacrylamide (9%) gel slab electrophoresis in 0.1% SDSwas carried out as described by Laemmli (1970). Celllysates were obtained by resuspending cellular pellets(corresponding to 0.7 OD) in SDS loading buffer, incubat-ing for 15 min at 90°C, and occasionally vortexing them.Culture supernatants were tenfold concentrated by ultra-filtration and 25 μl of ultrafiltered broths were loaded onSDS-PAGE.
Analytical isoelectric focusing (IEF) in the pH range 3.5–9.5 was performed on 4.6% acrylamide gel slab with aMultiphor electrophoresis system (Amersham Biosciences)according to the manufacturer’s instructions.
Western blotting
Proteins were separated on SDS-PAGE and electroblottedonto polyvinylidene difluoride membranes (Immobilion P,Millipore). Electroblotting was performed in 10 mM 3-(cy-clohexylamino)-1-propanesulfonic acid (CAPS), pH 11.0,10% (v/v) methanol at 50 V for 180 min at room tem-perature. The washing solution was 5% (w/v) dried milkin phosphate-buffered saline solution supplemented with0.2% (v/v) Triton X-100 (washing buffer). The membranewas washed and incubated with rabbit anti-POXC anti-bodies diluted 1:5,000 or rabbit anti-POXA1b antibodiesdiluted 1:2,000 in washing buffer, at room temperature for1 h under continuous shaking. Subsequently, the membranewas washed and incubated as previously described withanti-rabbit IgG, peroxidase conjugate (Sigma), diluted1:2,000 in washing buffer. Blots were visualised with100 mM Tris–HCl (pH 7.5), 0.5 mg ml−1 3,3′-diaminoben-zidine, 0.03% (w/v) NiCl2, and 0.006% (v/v) H2O2.
Laccase purification
K. lactis culture media were harvested 3 days after inoc-ulation, and cells were sedimented by centrifugation at12,000×g at 4°C for 15 min. Phenylmethylsulphonyl fluo-ride (1 mM) was added to the culture supernatant, and
secreted proteins were precipitated by the addition of(NH4)2SO4 up to 80% saturation at 4°C and centrifugedat 10,000×g for 40 min. After this step, different purifi-cation strategies were exploited for rPOXA1b and rPOXC.
The rPOXA1b ammonium sulphate precipitate was re-suspended in 50 mM sodium phosphate (pH 6.0) and afterextensive dialysis against the same buffer was loaded ontoan SP-Sepharose Fast Flow (Amersham Biosciences) col-umn equilibrated with the phosphate buffer. After washing,the enzyme was eluted at a flow rate of 1 ml min−1 with alinear gradient from 0 to 0.4 M NaCl. The active fractionswere pooled, concentrated and desalted on an Amicon PM-30 membrane.
The rPOXC ammonium sulphate precipitate was resus-pended in 50 mM sodium phosphate (pH 7.0) containing1 M (NH4)2SO4, and loaded onto a Phenyl Sepharosehigh-performance 35/100 column (Amersham Biosciences)equilibrated with the same buffer. After an isocratic step at1 M (NH4)2SO4, a linear gradient from 1 to 0 M (NH4)2SO4
was applied. The active fractions were pooled, concentratedand desalted on an Amicon PM-30 membrane. Then, thesample was loaded onto a Q-Sepharose Fast Flow (Amers-ham Biosciences) column equilibrated with the phosphatebuffer and eluted from ionic exchange chromatography at aflow rate of 1 ml min−1 with a linear gradient from 0 to0.4MNaCl. The active fractions were pooled, concentratedand desalted as above.
Protein determination
Protein concentration was determined using the Bio-Radprotein assay with bovine serum albumin as standard.
Reduction and alkylation of cysteine residues
Mass spectrometry analyses were performed on the solublerecombinant proteins (1 nmol) reduced in 300 mMTris (pH8.0) and 20 mM EDTA (containing 6 M guanidiniumchloride) by incubation with a 10:1 molar excess of di-thiothreitol over the protein SH groups at 37°C for 2 h. Thefree cysteine residues were alkylated by using a 5:1 excessof iodoacetamide over the total SH groups at room tem-perature for 1 h in the dark under a nitrogen atmosphere.Protein samples were freed from salt and excess reagent bypassing the reaction mixture through a PD10 prepackedcolumn (Pharmacia Biotech), equilibrated and eluted in0.4% ammonium bicarbonate (pH 8.5).
Mass spectrometry analyses were also performed on theCoomassie blue-stained proteins (0.4 nmol) excised from apreparative SDS electrophoresis on a 9% polyacrylamidegel. Excised bands were washed with acetonitrile and thenwith 0.1 M ammonium bicarbonate. Protein samples werereduced by incubation in 10 mM dithiothreitol for 45 minat 56°C and carboxamidomethylated by using 55 mMiodoacetamide in 0.1 M NH4HCO3 for 30 min, in the dark,under nitrogen atmosphere at room temperature.
432

Enzymatic hydrolysis
The gel particles were washed with ammonium bicarbonateand acetonitrile. Enzymatic digestions were carried out withtrypsin (10 μg ml−1) in 50 mM ammonium bicarbonate (pH8.5) at 4°C for 2 h. The buffer solution was then removedand a new aliquot of the enzyme/buffer solution was addedfor 18 h at 37°C. Minimum reaction volume, sufficient forcomplete rehydration of the gel, was used. Peptides werethen extracted by washing the gel particles with 20 mMammonium bicarbonate and 0.1% trifluoroacetic acid in50% acetonitrile at room temperature and then lyophilised.Aliquots of the digests were directly analysed by matrixassisted laser desorption ionisation mass spectrometry(MALDI-MS). The tryptic peptide mixture was deglyco-sylated with peptide N-glycosidase F (PNGaseF) by in-cubation overnight in 0.4% ammonium bicarbonate (pH8.5) at 37°C. Aliquots of the digest were analysed directlyby MALDI-MS. V8, trypsin and AspN digestions in so-lution were performed on carboxamidomethylated laccasesamples in 0.4% ammonium bicarbonate (pH 8.5) at 37°Cfor 18 h using an enzyme/substrate ratio of 1:50.
Protein deglycosylation was done by incubating laccasesamples (1 U) with 1 mU of endoglycosidase H in 20 mMsodium acetate (pH 5.2), 0.01% SDS and 5 mM DTT over-night at 37°C.
Mass spectrometry analysis
MALDI mass spectra were recorded using a Voyager DEand Voyager DE Pro MALDI-TOF mass spectrometer (Ap-plied Biosystems); a mixture of analyte solution, α-cyano-4-hydroxy-cinnamic acid or di-hydroxy-benzoic as matrices,with bovine insulin and horse heart myoglobin as standards,were applied to the sample plate and air-dried. Mass cali-bration was obtained using the quasi-molecular ions (MH+)from horse myoglobin (m/z 16,952.50), bovine insulin(m/z 5734.59), and α-cyano-4-hydroxy-cinnamic acid (m/z379.06) as internal standards. Raw data were analysed byusing computer software provided by the manufacturer andare reported as average or monoisotopic masses.
Results
Two laccase isoenzymes secreted by P. ostreatus, POXC(Palmieri et al. 1993) and POXA1b (Giardina et al. 1999)were expressed in K. lactis and S. cerevisiae. The sequenceof the poxc cDNA used was the same as the GenBanksequence, whereas that of the poxa1b cDNA differed byone nucleotide from the GenBank sequence at position 687.This mutation results in a single amino acid substitution,involving residue 201 of the mature protein (codon GACinstead of AAC, coding Asp instead of Asn).
K. lactis transformants
The production of recombinant laccases was assayed onplates containing ABTS as substrate. Several transformants(85%) carrying pYA1bps vector caused a colour changeafter 3 days, whereas no colour change was observed eitherfor transformants carrying pYCps or the pYCtk vectors,both of which contained the poxc cDNA.
Thirty recombinant clones from each transformationwere grown in liquid cultures and evaluated for laccaseproduction. The amount of extracellular laccase activityproduced after 3 days by the pYA1bps transformants rangedfrom (0 to 9)×10−2 U ml−1, whereas activities ranging from(0 to 6)×10−4 U ml−1 were measured for pYCps trans-formants. No laccase activity and no signal onWestern blotswere detected for cellular extracts or culture supernatantsfrom either the pYCtk transformants or the control cultures.The best producing clones (named HpYC for r-POXC andHpYA1b for r-POXA1b) were used to study the time courseof laccase production and to characterise the recombinantenzymes. Western blot analyses showed that the secretedrecombinant POXA1b had a slightly higher apparentmolecular mass (Mr) than the intracellular recombinantand the native proteins, whereas the protein recognized bythe anti-POXC antibodies had aMr very close to that of thenative laccase (Fig. 3a-1,b-1).
Cultures of HpYC and HpYA1b clones were carried outto improve laccase production varying the growth con-ditions (inducer concentration, medium composition andcopper concentration):
1. Both HpYC and HpYA1b produced the highest amountof laccase activity (0.007 and 2.03 U ml−1, correspond-ing to 0.25 and 95 mU/OD, respectively) in the pres-ence of 0.5% ethanol (Fig. 4a). Cell growth decreased(50% after 4 days) in the presence of 2% ethanol.
a
Celllysate
CulturesupernatantPOXC
1
Celllysate
CulturesupernatantPOXC
2
b
CelllysatePOXA1b
Culturesupernatant
1
CelllysatePOXA1b
Culturesupernatant
2K. lactis S. cerevisiae P. ostreatus
c
deglycosylated-POXA1b
Fig. 3 Western blot analyses of cellular extracts and culture super-natants. a rPOXC-expressing clones: 1 K. lactis clone HpYC; 2 S.cerevisiae clone C-pSAL4. b rPOXA1b-expressing clones: 1 K. lactisclone HpYA1b; 2 S. cerevisiae clone B-pSAL4. c Deglycosylatednative and recombinant POXA1b
433
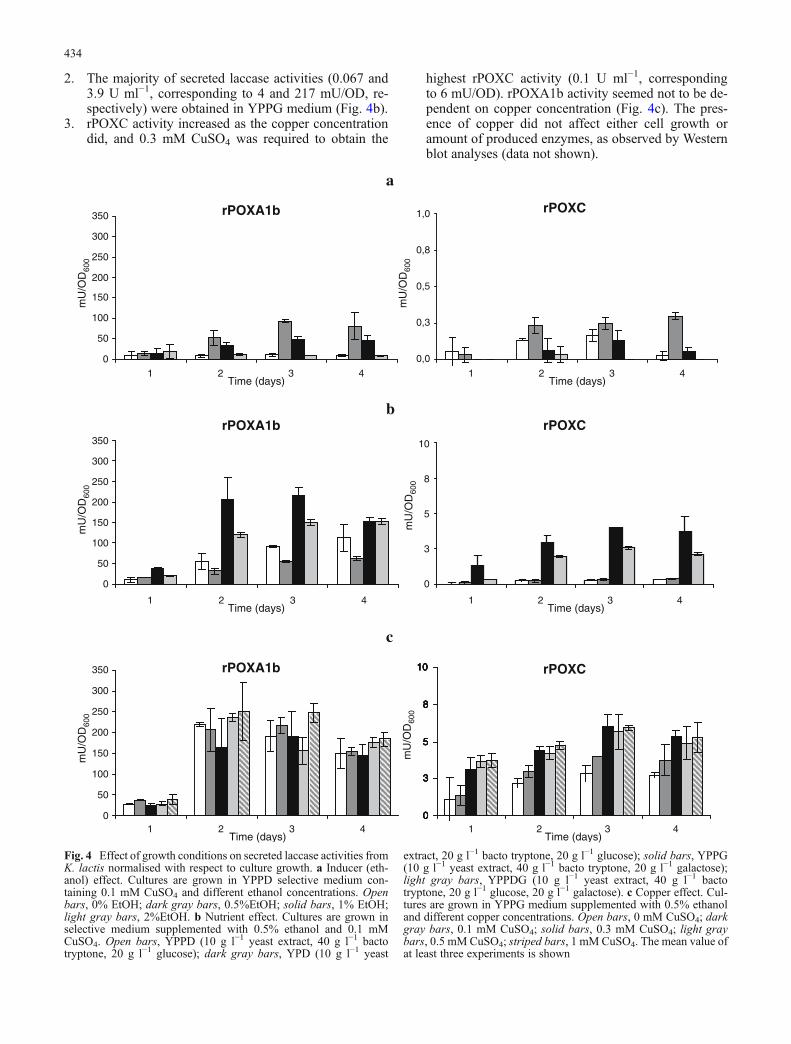
2. The majority of secreted laccase activities (0.067 and3.9 U ml−1, corresponding to 4 and 217 mU/OD, re-spectively) were obtained in YPPG medium (Fig. 4b).
3. rPOXC activity increased as the copper concentrationdid, and 0.3 mM CuSO4 was required to obtain the
highest rPOXC activity (0.1 U ml−1, correspondingto 6 mU/OD). rPOXA1b activity seemed not to be de-pendent on copper concentration (Fig. 4c). The pres-ence of copper did not affect either cell growth oramount of produced enzymes, as observed by Westernblot analyses (data not shown).
Fig. 4 Effect of growth conditions on secreted laccase activities fromK. lactis normalised with respect to culture growth. a Inducer (eth-anol) effect. Cultures are grown in YPPD selective medium con-taining 0.1 mM CuSO4 and different ethanol concentrations. Openbars, 0% EtOH; dark gray bars, 0.5%EtOH; solid bars, 1% EtOH;light gray bars, 2%EtOH. b Nutrient effect. Cultures are grown inselective medium supplemented with 0.5% ethanol and 0.1 mMCuSO4. Open bars, YPPD (10 g l−1 yeast extract, 40 g l−1 bactotryptone, 20 g l−1 glucose); dark gray bars, YPD (10 g l−1 yeast
extract, 20 g l−1 bacto tryptone, 20 g l−1 glucose); solid bars, YPPG(10 g l−1 yeast extract, 40 g l−1 bacto tryptone, 20 g l−1 galactose);light gray bars, YPPDG (10 g l−1 yeast extract, 40 g l−1 bactotryptone, 20 g l−1 glucose, 20 g l−1 galactose). c Copper effect. Cul-tures are grown in YPPG medium supplemented with 0.5% ethanoland different copper concentrations. Open bars, 0 mM CuSO4; darkgray bars, 0.1 mM CuSO4; solid bars, 0.3 mM CuSO4; light graybars, 0.5 mMCuSO4; striped bars, 1 mM CuSO4. The mean value ofat least three experiments is shown
0
3
5
8
10
rPOXA1b
0
3
5
8
10
rPOXCb
0
3
5
8
10
0
3
5
8
10rPOXA1b
0
3
5
8
10
0
3
5
8
10 rPOXC
c
0
50
100
150
200
250
300
350
1 2 3 4Time (days)
1 2 3 4Time (days)
1 2 3 4Time (days)
1 2 3 4Time (days)
1 2 3 4Time (days)
1 2 3 4Time (days)
rPOXA1b
mU
/OD
600
mU
/OD
600
mU
/OD
600
mU
/OD
600
mU
/OD
600
mU
/OD
600
0
50
100
150
200
250
300
350
0
50
100
150
200
250
300
350
a
0,0
0,3
0,5
0,8
1,0 rPOXC
434
S. cerevisiae transformants
Two of the three S. cerevisiae transformants containing thepoxc cDNA (C-YEp and C-pSAL4), and all poxa1b trans-formants were found positive by plate assay. Both thecolour intensity and the halo diameter suggested a markeddifference in the levels of laccase expression either amongthe different constructs used or between the two laccases.pSAL4 transformants were the best laccase producers forboth P. ostreatus laccases, all being poxa1b transformantsmore efficient than poxc transformants.
Western blot analyses of cellular extracts and culturesupernatants of C-pSAL4 and B-pSAL4 transformants(Fig. 3a-2,b-2) showed that the secreted recombinantPOXA1b had a higher apparent Mr than those of the intra-cellular recombinant and the native proteins, whereas intra-and extracellular rPOXC had a Mr very close to that ofthe native laccase.Moreover, enzymatically deglycosylatednative and recombinant S. cerevisiae andK. lactis POXA1bmigrated similarly in SDS-PAGE (Fig. 3c), thus demon-strating that the higher apparent molecular masses are due todifferent glycosylation.
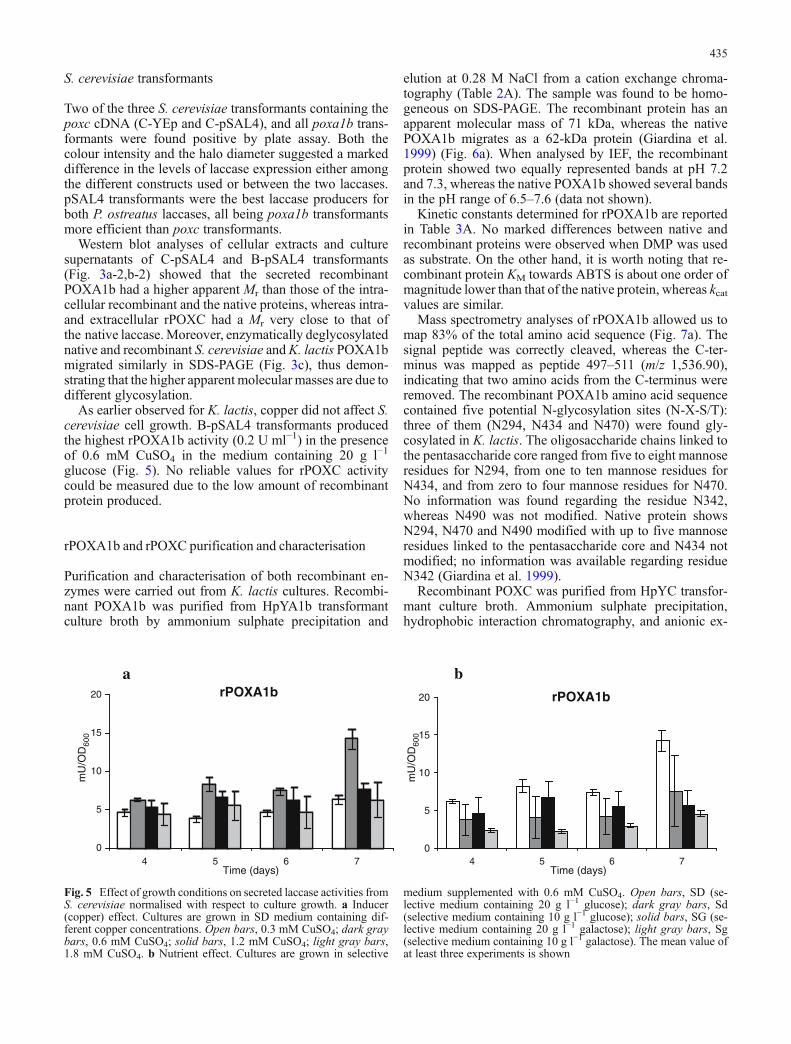
As earlier observed for K. lactis, copper did not affect S.cerevisiae cell growth. B-pSAL4 transformants producedthe highest rPOXA1b activity (0.2 U ml−1) in the presenceof 0.6 mM CuSO4 in the medium containing 20 g l−1
glucose (Fig. 5). No reliable values for rPOXC activitycould be measured due to the low amount of recombinantprotein produced.
rPOXA1b and rPOXC purification and characterisation
Purification and characterisation of both recombinant en-zymes were carried out from K. lactis cultures. Recombi-nant POXA1b was purified from HpYA1b transformantculture broth by ammonium sulphate precipitation and
elution at 0.28 M NaCl from a cation exchange chroma-tography (Table 2A). The sample was found to be homo-geneous on SDS-PAGE. The recombinant protein has anapparent molecular mass of 71 kDa, whereas the nativePOXA1b migrates as a 62-kDa protein (Giardina et al.1999) (Fig. 6a). When analysed by IEF, the recombinantprotein showed two equally represented bands at pH 7.2and 7.3, whereas the native POXA1b showed several bandsin the pH range of 6.5–7.6 (data not shown).
Kinetic constants determined for rPOXA1b are reportedin Table 3A. No marked differences between native andrecombinant proteins were observed when DMP was usedas substrate. On the other hand, it is worth noting that re-combinant protein KM towards ABTS is about one order ofmagnitude lower than that of the native protein, whereas kcatvalues are similar.
Mass spectrometry analyses of rPOXA1b allowed us tomap 83% of the total amino acid sequence (Fig. 7a). Thesignal peptide was correctly cleaved, whereas the C-ter-minus was mapped as peptide 497–511 (m/z 1,536.90),indicating that two amino acids from the C-terminus wereremoved. The recombinant POXA1b amino acid sequencecontained five potential N-glycosylation sites (N-X-S/T):three of them (N294, N434 and N470) were found gly-cosylated in K. lactis. The oligosaccharide chains linked tothe pentasaccharide core ranged from five to eight mannoseresidues for N294, from one to ten mannose residues forN434, and from zero to four mannose residues for N470.No information was found regarding the residue N342,whereas N490 was not modified. Native protein showsN294, N470 and N490 modified with up to five mannoseresidues linked to the pentasaccharide core and N434 notmodified; no information was available regarding residueN342 (Giardina et al. 1999).
Recombinant POXC was purified from HpYC transfor-mant culture broth. Ammonium sulphate precipitation,hydrophobic interaction chromatography, and anionic ex-
0
5
10
15
20 rPOXA1b
b
0
5
10
15
20
4 5 76
rPOXA1b
Time (days)4 5 76
Time (days)
mU
/OD
600
mU
/OD
600
a
Fig. 5 Effect of growth conditions on secreted laccase activities fromS. cerevisiae normalised with respect to culture growth. a Inducer(copper) effect. Cultures are grown in SD medium containing dif-ferent copper concentrations. Open bars, 0.3 mM CuSO4; dark graybars, 0.6 mM CuSO4; solid bars, 1.2 mM CuSO4; light gray bars,1.8 mM CuSO4. b Nutrient effect. Cultures are grown in selective
medium supplemented with 0.6 mM CuSO4. Open bars, SD (se-lective medium containing 20 g l−1 glucose); dark gray bars, Sd(selective medium containing 10 g l−1 glucose); solid bars, SG (se-lective medium containing 20 g l−1 galactose); light gray bars, Sg(selective medium containing 10 g l−1 galactose). The mean value ofat least three experiments is shown
435

change chromatography were sequentially used (Table 2B).The active fraction eluted at 0.28 M NaCl from Q-Se-pharose chromatography appeared to be homogeneous onSDS-PAGE and the recombinant protein migration wassimilar to that of the native POXC (Fig. 6b). Recombinantand native proteins also displayed the same mobility whenanalysed by native PAGE and IEF (data not shown).
Kinetic constants determined for rPOXC are reported inTable 3B. As earlier observed for rPOXA1b, rPOXC KM
towards ABTS is about one order of magnitude lower thanthat of the native protein, whereas KM of both proteinstowards DMP are similar. On the other hand, kcat values forboth substrates are significantly lower than those of thenative enzyme. No activity increase was observed afterroom temperature incubation of rPOXC with Cu(II) (5 mMCuSO4) or Cu(I) (5 mM CuSO4 in the presence of 0.1, 0.5and 1 mM sodium ascorbate) (Larrondo et al. 2003).
Mass spectrometry analyses of rPOXC allowed us tomap 70% of the amino acid sequence (Fig. 7b). The signalpeptide was correctly cleaved, whereas no information re-garding the C-terminal sequence could be deduced. POXCamino acid sequence contains two potential N-glycosyla-tion sites, one of them (N444) was found glycosylated inK. lactis. The oligosaccharide chains linked to the penta-saccharide core ranged from three to seven mannoseresidues. In the native protein, the same site was foundglycosylated with up to six mannose residues linked to thepentasaccharide core (Palmieri et al. 1993).
bPOXA1b
31 2POXC
a
31 2
116 kDa
66 kDa
35 kDa
45 kDa
25 kDa
Fig. 6 SDS-PAGE analyses ofrPOXA1b (a) and rPOXC (b).Lane 1, recombinant protein;lane 2, native protein; lane 3,molecular mass standards
Table 2 rPOXA1b and rPOXCpurification from K. lactiscultures
Purification step Total activity(U)
Total protein(mg)
Specific activity(U/mg)
Recovery(%)
Purificationfold
A. rPOXA1bBroth 1,392 94.7 14.7 100 1.0(NH4)2SO4 precipitate 910 32.0 28.4 65.4 1.9SP-Sepharose 745 0.6 1,307.0 53.5 88.9B. rPOXCBroth 54.6 106.0 0.51 – 1.0(NH4)2SO4 precipitate 73.2 100.8 0.73 100 1.4Phenyl sepharose 59.3 3.8 15.6 81.1 30.6Q-Sepharose 59.1 1.1 51.8 80.7 101.6
Table 3 rPOXA1b and rPOXC kinetic constants compared to thoseof the native proteins (POXA1b and POXC)
Substrate KM (mM) kcat (min−1)
A. rPOXA1bPOXA1b rPOXA1b POXA1b rPOXA1b
ABTS 0.47±0.06 0.04±0.01 (9.0±1.7)×104 (9.0±1.2)×104
DMP 0.26±0.09 0.29±0.04 (1.5±0.1)×104 (5.5±0.7)×104
B. rPOXCPOXC rPOXC POXC rPOXC
ABTS 0.28±0.04 0.03±0.01 (1.4±0.2)×105 (3.2±0.4)×103
DMP 0.07±0.01 0.13±0.02 (4.8±0.2)×105 (1.0±0.1)×103
436

Discussion
Two yeast expression systems were developed and com-paratively analysed for P. ostreatus laccase heterologousproduction.
Laccase expression in yeasts had already been reportedusing both native laccase or yeast signal peptides (S. cere-visiae α-factor, invertase) and different results were ob-tained for various expressed enzymes. In P. pastoris, betterresults were found using the laccase signal peptide insteadof the S. cerevisiae α-factor fusion (Jonsson et al. 1997; Liuet al. 2003; Soden et al. 2002), whereas other authors re-ported comparable extracellular activities via the two signalpeptides (Otterbein et al. 2000). In S. cerevisiae, a highersecreted activity was found using the yeast α-factor signalpeptide (Kiiskinen and Saloheimo 2004) or the yeastinvertase signal peptide (Klonowska et al. 2005). There-fore, the best performing signal peptide to drive the se-cretion of recombinant laccases in yeasts seems not to be apriori predictable.
Three expression vectors were constructed to expressboth POXC and POXA1b in S. cerevisiae, differing for
replication origins, promoter regions and signal peptidesequences. The plasmid with 2μ as replication origin, copperinducible CUP1 promoter region, and homologous laccasesignal peptides gave the best results for both laccases. How-ever, the native signal peptide always proved to be moreeffective in extracellular laccase production.
Recombinant expression of POXC in K. lactis was per-formed using both laccase and killer toxin signal peptide,and intra- and extracellular laccase production were an-alysed. No recombinant enzyme was detected by immuno-blot analysis in both compartments when killer toxin signalpeptide was used. On the other hand, the laccase signalpeptide allowed production and secretion of active rPOXCin the medium. Therefore, the signal peptide not only af-fects secretion but also production of recombinant enzymesin yeasts.
The apparent molecular mass of both recombinant pro-teins expressed in K. lactis is similar to that of the nativeones, whereas S. cerevisiae extracellular rPOXA1b showshigher molecular mass. These data confirm the tendency ofK. lactis with respect to S. cerevisiae to not hyperglyco-sylate recombinant proteins.
SIGPRGTLNI ANKVIQPDGF YRSTVLAGGS YPGPLI
POXC
AIGPAGNMYI VNEDVSPDGF ARSAVVARSV PATDPTPATA SIPGVLVQGN KGDNFQLNVV 60
NQLSDTTMLK TTSIHWHGFF QAGSSWADGP AFVTQCPVAS GDSFLYNFNV PDQAGTFWYH 120
SHLSTQYCDG LRGPFVVYDP SDPHLSLYDI DNADTVITLE DWYHIVAPQN AAIPTPDSTL 180
INGKGRYAGG PTSPLAIINV ESNKRYRFRL VSMSCDPNFT FSIDGHSLLV IEADAVNIVP 240
ITVDSIQIFA GQRYSFVLTA NQAVDNYWIR ANPNLGSTGF VGGINSAILR YAGATEDDPT 300
TTSSTSTPLL ETNLVPLENP GAPGPPVPGG ADININLAMA FDFTTFELTI NGVPFLPPTA 360
PVLLQILSGA STAASLLPSG SIYELEANKV VEISMPALAV GGPHPFHLHG HTFDVIRSAG 420
*STTYNFDTPA RRDVVNTGTG ANDNVTIRFV TDNPGPWFLH CHIDWHLEIG LAVVFAEDVT 480
SISAPPAAWD DLCPIYNALS DNDKGGIVPS 510
POXA1b
KGKT GDRFQINVVN KLADTSMPVD 60
TSIHWHGLFV KGHNWADGPA MVTQCPIVPG HSFLYDFEVP DQAGTFWYHS HLGTQYCDGL 120
RGPLVVYSKN DPHKRLYDVD DESTVLTVGD WYHAPSLSLT GVPHPDSTLF NGLGRSLNGP 180
ASPLYVMNVV KGKRYRIRLI DTSCDSNYQF SIDGHTFTVI EADGENTQPL QVDQVQIFAG 240
*QRYSLVLNAN QAVGNYWIRA NPNSGDPGFE NQMNSAILRY KGARSIDPTT PEQNATNPLH 300
EYNLRPLIKK PAPGKPFPGG ADHNINLNFA FDPATALFTA NNHTFVPPTV PVLLQILSGT 360
RDAHDLAPAG SIYDIKLGDV VEITMPALVF AGPHPIHLHG HTFAVVRSAG SSTYNYENPV 420
* *RRDVVSIGDD PTDNVTIRFV ADNAGPWFLH CHIDWHLDLG FAVVFAEGVN QTAAANPVPE 480
AWNNLCPIYN SSNPSKLLMG TNAIGRLPAP LKA 513
a
b
Fig. 7 Amino acid sequence ofPOXA1b (a) and POXC (b). Theunderlined stretches have beenverified by MALDI-MS for therecombinant proteins. PutativeN-glycosylation sites are in bold;sites found glycosylated are in-dicated by an asterisk
437

It is worth noting that rPOXA1b transformants alwaysgave higher activity than rPOXC transformants, and pro-duction of both laccases in K. lactis was always signif-icantly higher than that in S. cerevisiae.
Both K. lactis recombinant laccases were purified tohomogeneity, characterised, and their properties comparedwith those of the corresponding native proteins. Amountsof rPOXA1b and rPOXC purified from 1-l culture brothwere similar (1.1 and 1.4 mg, respectively, taking into ac-count purification yield). Mass spectrometry analyses al-lowed mapping of the primary structures of K. lactisrPOXA1b and rPOXC and determining their glycosydicmoieties. POXC contains two potential N-glycosylationconsensus sites in its sequence, one of them being modifiedboth in the recombinant and in the native protein. POXA1bcontains five potential N-glycosylation consensus sites, andthree of them were found modified in the recombinantprotein as well as in the native one. Taking into account thatthe average number of mannose residues per site in re-combinant proteins is higher than that of native ones andthat the molecular mass differences are 11.3% for POXA1band 1.6% for POXC (by SDS-PAGE), the extent of glyco-sylation per site affects recombinant enzyme molecularmasses more than the number of N-glycosylated sites.
rPOXC kcat is about two orders of magnitude lower thanthat of the native one, whereas no significant differencewas observed for rPOXA1b kcat value. When the effect ofcopper concentration on K. lactis laccase production wasevaluated, rPOXC showed a positive correlation betweencopper availability and laccase activity up to 0.3 mMCuSO4, higher CuSO4 concentration having no further ef-fect. On the other hand, no activity increase was observedwhen purified rPOXC was incubated with either Cu(II) orCu(I) (Larrondo et al. 2003). These results lead us to ex-clude that the low rPOXC kcat depends on the presence ofapoproteins lacking some copper ions or on its glycosyla-tion status and to hypothesize that rPOXC undergoes anincomplete or incorrect protein folding.
It is worth noting that rPOXA1b undergoes a C-terminalprocessing, and a C-terminus heterogeneity was observedfor the native POXA1b (Giardina et al. 1999). Unfortu-nately, no information on native and recombinant POXCC-termini are available. C-terminal processing at a con-served processing site has been reported for ascomycetelaccases, and a role of the C-terminus in modulating enzy-matic acivity has been hypothesized (Hakulinen et al. 2002;Kiiskinen and Saloheimo 2004). If C-terminus can reallyaffect laccase activity, it could be responsible for the dif-ferent affinity between native and recombinant POXA1btowards ABTS. The same difference was found betweennative and recombinant POXC, but the lack of informationregarding their C-termini does not allow inferring generalconclusions. Further investigations are needed to evaluatethe influence of C-terminus on laccase catalytic parameters.
The availability of established recombinant expressionsystems can represent the starting point for projecting newoxidative catalysts suitable for specific industrial needs.
Acknowledgements This work was supported by grants from theMinistero dell’Università e della Ricerca Scientifica (Progetti diRilevante Interesse Nazionale, PRIN 2002 and 2004), INTAS (In-ternational Association for the promotion of cooperation with thescientists from the New Independent States of the former SovietUnion, ref. no. 03-51-5889), and Centro Regionale di CompetenzaBioTekNet. A. Piscitelli was the recipient of a French/Italian Uni-versity fellowship. The authors thank Prof. Claudio Falcone forkindly making availableK. lactis strain and plasmid, Prof. Piero Puccifor helpful discussions, and Dr. Thierry Tron for the critical reading ofthe manuscript and for kindly making available S. cerevisiae strainand plasmids.
References
Aramayo R, Timberlake WE (1993) The Aspergillus nidulans yAgene is regulated by abaA. EMBO J 12:2039–2048
Bergquist P, Te’o V, Gibbs M, Cziferszhy A, De Faria FP, AzevedoM, Nevalainen H (2002) Expression of xylanase enzymes fromthermophilic microorganisms in fungal hosts. Extremophiles6:177–184
Berka RM, Schneider P, Golightly EJ, Brown SH, MaddenM, BrownKM, Halkier T, Mondorf K, Xu F (1997) Characterization of thegene encoding an extracellular laccase of Myceliophtora ther-mophila and analysis of the recombinant enzyme expressed inAspergillus oryzae. Appl Environ Microbiol 63:3151–3157
Bulter T, Alcalde M, Sieber V, Meinhold P, Schlachtbauer C, ArnoldFH (2003) Functional expression of a fungal laccase in Saccha-romyces cerevisiae by directed evolution. Appl Environ Micro-biol 69:987–995
Cassland P, Jonsson LJ (1999) Characterization of a gene encodingTrametes versicolor laccase A and improved heterologous ex-pression in Saccharomyces cerevisiae by decreased cultivationtemperature. Appl Microbiol Biotechnol 52:393–400
Claus H (2003) Laccases and their occurrence in prokaryotes. ArchMicrobiol 179:145–150
Dittmer NT, Suderman RJ, Jiang H, Zhu YC, Gorman MJ, KramerKJ, Kanost MR (2004) Characterization of cDNAs encodingputative laccase-like multicopper oxidases and developmentalexpression in the tobacco hornworm, Manduca sexta, and themalaria mosquito, Anopheles gambiae. Insect BiochemMol Biol34:29–41
Falcone C, Saliola M, Chen XJ, Frontali L, Fukuhara H (1986)Analysis of a 1.6-micron circular plasmid from the yeastKluyveromyces drosophilarum: structure and molecular dimor-phism. Plasmid 15:248–252
Gelo-Pujic M, Kim HH, Butlin NG, Palmore GT (1999) Electro-chemical studies of a truncated laccase produced in Pichiapastoris. Appl Environ Microbiol 65:5515–5521
Giardina P, Cannio R, Martirani L, Marzullo L, Palmieri G, Sannia G(1995) Cloning and sequencing of a laccase gene from thelignin-degrading basidiomycete Pleurotus ostreatus. Appl En-viron Microbiol 61:2408–2413
Giardina P, Aurilia V, Cannio R, Marzullo L, Amoresano A, SicilianoR, Pucci P, Sannia G (1996) The gene, protein and glycan struc-tures of laccase from Pleurotus ostreatus. Eur J Biochem 235:508–515
Giardina P, Palmieri, G, Scaloni A, Fontanella B, Faraco V, CennamoG, Sannia G (1999) Protein and gene structure of a blue laccasefrom Pleurotus ostreatus. Biochem J 341:655–663
Gietz D, St. Jean A, Woods RA, Schiestl RH (1992) Improvedmethod for high efficiency transformation of intact yeast cells.Nucleic Acids Res 2:1425
Hakulinen N, Kiiskinen LL, Kruus K, Saloheimo M, Paananen A,Koivula A, Rouvinen J (2002) Crystal structure of a laccase fromMelanocarpus albomyces with an intact trinuclear copper site.Nat Struct Biol 9:601–605
438

Hatamoto O, Sekine H, Nakano E, Abe K (1999) Cloning and ex-pression of a cDNA encoding the laccase from Schizophyllumcommune. Biosci Biotechnol Biochem 63:58–64
Hill JE, Myers AM, Koerner TJ, Tzagoloff A (1986) Yeast/E. colishuttle vectors with multiple unique restriction sites. Yeast 2:163–167
Hsieh HP, Da Silva NA (1998) Partial-pKD1 plasmids provideenhanced structural stability for heterologous protein productionin Kluyveromyces lactis. Appl Microbiol Biotechnol 49:411–416
Jolivalt C, Madzak C, Brault A, Caminade E, Malosse C, Mougin C(2005) Expression of laccase IIIb from the white-rot fungusTrametes versicolor in the yeast Yarrowia lipolytica for envi-ronmental applications. Appl Microbiol Biotechnol 66:450–456
Jonsson LJ, SaloheimoM, Penttila M (1997) Laccase from the white-rot fungus Trametes versicolor: cDNA cloning of lcc1 and ex-pression in Pichia pastoris. Curr Genet 32:425–430
Kiiskinen LL, Saloheimo M (2004) Molecular cloning and expres-sion in Saccharomyces cerevisiae of a laccase gene from theascomycete Melanocarpus albomyces. Appl Environ Microbiol70:137–144
Kiiskinen LL, Kruus K, Bailey M, Ylosmaki E, Siika-Aho M,Saloheimo M (2004) Expression of Melanocarpus albomyceslaccase in Trichoderma reesei and characterisation of the pu-rified. Microbiology 150:3065–3074
Klonowska A, Gaudin C, Asso M, Fournel A, Reglier M, Tron T(2005) LAC3, a new low redox potential laccase from Trametessp strain C30 obtained as recombinant protein in yeasts. EnzymeMicrob Technol 36:34–41
Kojima Y, Tsukuda Y, Kawai Y, Tsukamoto A, Sugiura J, Sakaino M,Kita Y (1990) Cloning, sequence analysis, and expression ofligninolytic phenoloxidase genes of the white-rot basidiomyceteCoriolus hirsutus. J Biol Chem 265:15224–15230
Kramer KJ, Kanost MR, Hopkins TL, Jing H, Zhu YC, Xhu R,Kerwin JL, Turecek F (2001) Oxidative conjugation of catecholswith proteins in insect skeletal systems. Tetrahedron 57:385–392
Laemmli UK (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227:680–685
Larrondo LF, Avila M, Salas L, Cullen D, Vicuna R (2003) Het-erologous expression of laccase cDNA from Ceriporiopsis sub-vermispora yields copper-activated apoprotein and complexisoform patterns. Microbiology 149:1177–1182
Liu W, Chao Y, Liu S, Bao H, Qian S (2003) Molecular cloning andcharacterization of a laccase gene from the basidiomycete Fomelignosus and expression in Pichia pastoris. Appl MicrobiolBiotechnol 63:174–181
Mayer AM, Staples RC (2002) Laccase: new functions for an oldenzyme. Phytochemistry 60:551–565
Mazzoni C, Saliola M, Falcone C (1992) Ethanol-induced andglucose-insensitive alcohol dehydrogenase activity in the yeastKluyveromyces lactis. Mol Microbiol 6:2279–2286
O’Malley DM,Whetten R, BaoW, Chen CL, Seedorf RR (1993) Therole of laccase in lignification. Plant J 4:751–757
Otterbein L, Record E, Longhi S, Asther M, Moukha S (2000)Molecular cloning of the cDNA encoding laccase from Pycno-porus cinnabarinus I-937 and expression in Pichia pastoris. EurJ Biochem 267:1619–1625
Palmieri G, Giardina P, Marzullo L, Desiderio B, Nitti G, Cannio R,Sannia G (1993) Stability and activity of a phenol oxidase fromthe ligninolytic fungus Pleurotus ostreatus. Appl MicrobiolBiotechnol 39:632–636
Palmieri G, Giardina P, Bianco C, Scaloni A, Capasso A, Sannia G(1997) A novel white laccase from Pleurotus ostreatus. J BiolChem 272:31301–31307
Palmieri G, Cennamo G, Faraco V, Amoresano A, Sannia G, GiardinaP (2003) Atypical laccase isoenzymes from copper supple-mented Pleurotus ostreatus cultures. Enzyme Microb Technol33:220–230
Record E, Punt PJ, Chamkha M, Labat M, van Den Hondel CAMJJ,Aster M (2002) Expression of the Pycnoporus cinnabarinuslaccase gene in Aspergillus niger and characterization of therecombinant enzyme. Eur J Biochem 269:602–609
Saliola M, Mazzoni C, Solimando N, Crisà A, Falcone C, Jung G,Fleer R (1999) Use of the KlADH4 promoter for ethanol-dependent production of recombinant human serum albumin inKluyveromyces lactis. Appl Environ Microbiol 65:53–60
Sigoillot C, Record E, Belle V, Robert JL, Levasseur A, Punt PJ, VanDen Hondel CA, Fournel A, Sigoillot JC, Asther M (2004)Natural and recombinant fungal laccases for paper pulp bleach-ing. Appl Microbiol Biotechnol 64:346–352
Soden DM, O’Callaghan J, Dobson AD (2002) Molecular cloning ofa laccase isozyme gene from Pleurotus sajor-caju and expres-sion in the heterologous Pichia pastoris host. Microbiology 148:4003–4014
Stark MJ, Boyd A (1986) The killer toxin of Kluyveromyces lactis:characterization of the toxin subunits and identification of thegenes which encode them. EMBO J 5:1995–2002
Stoj C, Kosman DJ (2003) Cuprous oxidase activity of yeast Fet3pand human ceruloplasmin: implication for function. FEBS Lett554:422–426
Williamson PR, Wakamatsu K, Ito S (1998) Melanin biosynthesis inCryptococcus neoformans. J Bacteriol 180:1570–1572
Yaropolov AI, Skorobogat’ko OV, Vartanov SS, Varfolomeyev SD(1994) Laccase: properties, catalytic mechanism, and applica-bility. Appl Biochem Biotechnol 49:257–280
439




















