
*" 3 53 >, . ,5 *&3>* > 5 2 > 5*.4>5 5> ) "6 ' > &5 5;> 8 #+,$ &5> &
Upload
independentCategory
view
1download
0
Int. J. Med. Arom. Plants, ISSN 2249 – 4340RESEARCH ARTICLE
Vol. 2, No. 4, pp. 579-588, December 2012
*Corresponding author: (E-mail) dangreanand <@> gmail.com http://www.openaccessscience.com© 2012 Copyright by the Authors, licensee Open Access Science Research Publisher. [email protected]
This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported (CC BY-NC-ND 3.0) License (http://creativecommons.org/licenses/by-nc-nd/3.0)
Effect of Pleurotus nebrodensis extract on melanin synthesis: A natural
alternative for cosmetics
D. M. DANGRE1*, L. P. DAFNE1, R. P BHAGAT2, C. J. CHANDEKAR3
1Department of AYUSH, Maharashtra University of Health Sciences, Vani-Dindori Road, Mhasrul, Nashik,Maharashtra, India.2Department of Biotechnology, Rajarshi Shahu College, Latur, Maharshtra, India.3Department of Biotechnology, Shri Shivaji Science College, Congress Nagar, Nagpur, India.
*Corresponding Author, Tel: +91- 253-2539125, Fax: +91-253-2539125
Article History: Received 6th October 2012, Revised 27th December 2012, Accepted 28th December 2012.
Abstract: The mushroom Pleurotus nebrodensis is an edible mushroom widely used for nutritional and medicinal pur-poses. It was observed that the skin of mushroom farmers is turn out to be fairer as they were regularly involved in mush-room handling and process. In order to examine this effect and as a preliminary step to apply for cosmetic ingredients,the extracts as well as its fractionates with certain organic solvents have been prepared to characterize the effect bytyrosinase inhibition assay and melanogenesis inhibition capability in cultured human melanocytes as an in vitro model.The investigation revealed that certain extracts from P. nebrodensis are more effective in skin whitening than that ofArbutin, a widely used whitening ingredient in functional cosmetic products.
Keywords: Cosmetic ingredients; edible mushroom; melanogenesis inhibition; tyrosinase inhibition assay.
Introduction
The mushroom Pleurotus nebrodensis is anedible mushroom widely used for nutritionaland medicinal purposes. The extract of thismushroom with hot water have been reported tohave medicinal effects such as the reduction ofcholesterol level in blood and the activation ofmacrophages and lymphocytes in human beings(Alam et al. 2011). Recently, it was observedthat among the woman workers directly in-volved in handling of mushrooms at mushroomfarm were having whitening effect on their skin.In order to examine this effect and as a prelimi-nary step to apply for cosmetic ingredients, theextracts as well as its fractionates with certainorganic solvents have been prepared to charac-terize the effect by tyrosinase inhibition assayand melanogenesis inhibition capability in cul-tured Melanocytess or in human melanocytes asan in vitro model.
Melanin is one of the most widely distribut-ed pigments and is found in bacteria, fungi,plants as well as animals. It is a heterogeneous
polyphenol-like biopolymer with a complexstructure and color varying from yellow to black(Prota 1988). The color of mammalian skin andhair is determined by a number of factors, themost important of which is the degree and dis-tribution of melanin pigmentation. Melanin issecreted by melanocytes distributed in the basallayer of the dermis (Spritz and Hearing 1994).The role of melanin is to protect the skin fromultraviolet (UV) damage by absorbing UV sun-light and removing reactive oxygen species(ROS).
Great interest has been shown in the in-volvement of melanins in malignant Melanoma,the most life-threatening skin tumors. Melaninis formed through a series of oxidative reactionsinvolving the amino acid tyrosine in the pres-ence of tyrosinase. Tyrosinase (EC 1.14.18.1) isa copper-containing enzyme that catalyzes twodistinct reactions of melanin synthesis: the hy-droxylation of tyrosine by monophenolase ac-tion and the oxidation of 3,4-dihydroxyplhenylalanine (L-DOPA) to o-dopaquinone by diphenolase action. However, if

580Int. J. Med. Arom. Plants Mycoanalysis of Pleurotus nebrodensis against melanin synthesis
Dangre et al. http://[email protected]
L-DOPA is an active cofactor, its formation asan intermediate during o-dopaquinone produc-tion is still controversial. o-Dopaquinone is un-stable in aqueous solution and rapidly suffers anon-enzymatic cyclization to leukodopachrome,which is further oxidize enzymatically by an-other molecule of o-dopaquinone to yielddopachrome and one molecule of regenerated L-DOPA (Garcia-Canovas et al. 1982; Rodriguez-Lopez et al.1991; Cooksey et al. 1997).
Overview of melanin biosynthesis in mammals
Mammalian melanocytes can produce twotypes of melanin; eumelanin is black or brownand pheomelanin is red or yellow in color (Prota1992; Ito 1993). Switching between these twotypes of melanins in follicular melanocytes elic-its a temporary shift from eumelanogenesis topheomelanogenesis, which is responsible for thewild-type agouti pigment of murine hair color(Hirobe 1991). For many decades,melanosomal proteins that regulate melanin bio-synthesis have been studied and characterized,especially those required for eumelanogenesis(Hearing and King 1993). The pathway ofeumelanogenesis may be divided into two phas-es, one proximal and the other distal (Alam etal. 2011; Kobayashi et al. 1995). The proximalphase consists of the enzymatic oxidation of ty-rosine or L-DOPA to its corresponding o-dopaquinone catalyzed by tyrosinase. This nas-cent o-dopaquinone can undergo two differenttypes of reactions; intramolecular 1,4-additionto the benzene ring which causes its cyclizationinto leukodopachrome. This intermediate isquickly oxidized to dopachrome by another o-dopaquione, which is reduced back to L-DOPA(Brun and Rosset 1974; Pawelek 1991; Aroca etal. 1990). The second reaction occurs withcyclizable and noncyclizable quinines and con-sists of a water addition to the benzene ring,which leads to the formation of a three-hyddroxylated phenol, 2,4,5-trihydroxyphenylalanine (TOPA), which ischemically oxidized to p-topaquinone by anoth-er o-dopaquinone (Garcia-Canovas et al. 1982;Brun and Rosset 1974). This p-topaquinoneevolves through a series of slow reactions todopachrome, which is the final product of theproximal phase. The distal phase is represented
by chemical and enzymatic reactions which oc-cur after dopachrome formation and lead to thesynthesis of eumelanins. This phase starts withthe slow chemical decarboxylation ofdopachrome to 5,6-dihydroxyindole (DHI) andits subsequent oxidation to indole-5,6-quinone.As an alternative to this chemical evolution inthe distal phase, dopachrome may be enzymati-cally transformed into 5,6-dihydroxyindole-2-carboxylic acid (DHICA) by dopachrometautomerase (Pawelek 1991 and Aroca etal.1990). DHICA is further oxidized by a redoxreaction with o-dopaquinone to form indole-5,6-quinone carboxylic acid, which can exist inthree tautomeric forms, including the quinone-imine and the corresponding highly reactivequinone-methide (Lambert et al. 1989;Sugumaran and Semesi 1991). Properties ofDHI-derived and DHICA-derived melanins dif-fer in each other; the former are black and floc-culent, while the latter are yellowish-brown andfinely dispersed (Aroca et al. 1990). Duringpheomelanogenesis, the thiol group of Sulfhy-dryl compounds such as glutathione and Cyste-ine nucleophililcally attacks o-dopaquinonemade enzymatically by tyrosinase to produceCysteinyldopa or Glutathionyldopa. This thiolgroup can be added to different ring positions,although the 5-position is the favored position.Subsequent cyclization and polymerization ofCysteinyldopa or Glutathionyldopa in an un-characterized series of reaction result in the pro-duction of pheomelanins and trichochromes(Hearing and King 1993; Deshpande et al.1984). The interaction between the eumelaninand pheomelanin compounds gives rise to a het-erogeneous pool of mixed-type melanins.
Tyrosinase inhibitors
Melanin plays a crucial protective roleagainst skin photocarcinogenesis. However, theproduction of abnormal melanin pigmentation isa serious esthetic problem is human beings(Preiestley 1993). The cytotoxicity of L-DOPAhas been attributed to its selective uptake bymelanocytic cells and to the formation of reac-tive quinines and semiquinones formed in situduring metabolic activation of L-DOPA bytyrosinase (Wick et al. 1977; Graham et al.1978; Korytowski et al. 1987). The truphenolic

581Int. J. Med. Arom. Plants Mycoanalysis of Pleurotus nebrodensis against melanin synthesis
Dangre et al. http://[email protected]
amino acid TOPA also shows cytotoxic proper-ties which seem to be due to the susceptibility ofthis substance to oxidation (Wick et al. 1979;Sachs and Jonsson 1975). The oxidation prod-ucts of L-DOPA and TOPA, quinines, arechemically reactive compounds that are poten-tially harmful, but in melanocytes the normalprocess of melanogenesis is not usually associ-ated with significant toxicity due to thecompartmentation of the reaction within mem-brane-limited organelles (melanosomes) andbecause of the rapid cyclization of the quinoneintermediate. The discovery that certain substi-tuted phenols have a depigmenting action due totheir ability to act as substrates for tyrosinase,resulting in the generation of quinines, has ledto the examination of this system as a possibletargeted anti-Melanoma therapeutic strategy inthe case of disseminated Melanoma (Riely et al.1975; Morgan et al. 1981). Moreover, thisprodrug has to avoid alternative cellular metabo-lism and enter melanosomes, leading to oxida-tion by tyrosinase to generate significantamounts of the quinone product, which requiresthe ability to initiate reactions damaging to theMelanoma cell. Melanin biosynthesis can beinhibited by avoiding UV exposure, the inhibi-tion of tyrosinase, the inhibition of melanocytemetabolism and proliferation, or the removal ofmelanin with corneal ablation (Seiberg et al.2000 and Seiberg et al. 2000). Standard topicaltreatments for hyperpigmentation disorders suchas melasma and postinflammatory hyperpig-mentation include bleaching withhydroquinones, anti-inflammatory therapy byretinoids and use of tyrosinase inhibitors. Anumber of tyrosinase inhibitors from both natu-ral and synthetic sources that inhibitedmonophenolase, diphnolase of both of these ac-tivities have been identified to date.
Tyrosinase inhibitors application in cosmetics
Use of tyrosinase inhibitors is becoming in-creasingly important in the cosmetic and medic-inal industries due to their preventive effect onpigmentation disorders. Tyrosinase inhibitorsmay result in a reduction in melanin biosynthe-sis and are used in cosmetic products for hyper-pigmentation- related concerns, including the
formation of freckles. Tyrosinase and its inhibi-tors may also be targets for developing medi-cines to treat hypopigmentation-realated prob-lems, such as albinism and piebaldism. A num-ber of tyrosinase inhibitors have been reportedfrom both natural and synthetic sources, but on-ly a few of them are used as skin-whiteningagents, primarily due to various safety concerns.
Currently Arbutin and Aloesin are used inthe cosmetic industry as skin-whitening agentsbecause they show strong inhibition towardtyrosinase, which is responsible for pigmenta-tion in human beings. On the other hand the co-treatment of Arbutin and Aloesin exhibited theinhibitory effect on tyrosinase in a synergisticmanner by acting through different mechanisms:arbutin inhibited competitively, whereas aloesininhibited noncompetitively (Jin et al. 1999).Taken together they inhibit melanin productionsynergistically by a combined mechanism ofcompetitive and noncompetitive inhibition.This result indicates that it is beneficial to useArbutin and Aloesin as a mixture in depigmen-tation because the co-treatment reduces the dos-es of these agents for the same inhibitory effecton tyrosinase activity and can diminish adverseside effects.
In this study, we investigated tyrosinase in-hibition activity of the samples extracted fromPleurotus nebrodensis directly by using enzymeassays and indirectly through melanin synthesisin Melanocytes.
Materials and methods
The mushroom Pleurotus nebrodensis wassliced and completely removed the moisture byfreeze dry at -70 oC for 3 days. The 80% or100% pure Methanol (MeOH) were used forextraction under cold and hot temperature con-dition. Every sample extracted in MeOH for 3hours and repeated 3 times the extraction witheach sample.
The organic solvents (n-Hexane, Chloro-form and Butanol) were used for fractionation.From these solvents, we obtained n-Hexanefractionate sample, Chloroform fractionate sam-ple and Butanol fractionate sample for furtherstudy. Samples were dissolved in DMSO.

582Int. J. Med. Arom. Plants Mycoanalysis of Pleurotus nebrodensis against melanin synthesis
Dangre et al. http://[email protected]
Figure 1: Methanol and organic solvent extraction of P. nebrodensis.
Enzyme assay
The tyrosinase activity, using L-Tyrosine asa substrate was measured according to themethods of Kubu and Kinst-Hori, 2000 withslight modifications. The mushroom tyrosinase(EC 232-653-4) used for bioassay was pur-chased from Sigma-Aldrich Co. 40 µl of 1.5mML-tyrosine, 190 µl of 0.1M phosphate buffer(pH 6.8) and sample dissolved in DMSO weremixed, and 20 µl of mushroom tyrosinase(1,500 U/ml) was added. The enzyme-reactedsolution was incubated for 15 min at 37 oC andthe solution was immediately monitored for theformation of dopachrome by measuring the lin-ear increase in absorbance at 490 nm. Arbutinwas taken as a positive control to get compara-tive output.
The percentage of inhibition of enzyme activitywas expressed as
(1-(C-D)/(A-B))x100
(A = absorbance at 490 nm with enzyme andsubstrate but without test sample; B = absorb-
ance at 490 nm without test sample and enzyme;C = absorbance at 490 nm with test sample, en-zyme and substrate; D = absorbance at 490 nmwith test sample and subsrate, without enzyme).
Means of triplicates were determined andthe 50% inhibition (IC50) of tyrosinase activitywas calculated as the concentrations of testsamples that inhibited tyrosinase activity by50% under the experimental conditions.
The samples were gone through Thin LayerChromatography (TLC) to separate the com-pounds in various conditions. For each sampleswe choose a suitable liquid phase such as Hex-ane: Ethyl Acetate with ratio was 2:1 and 4:1,Methanol: Chloroform with ratio 1:20.
Cell viability and proliferation test
The MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay was used todetermine cellular viability and proliferationafter the treatment of cells by the samples for 24hrs as described. The culture plate was further

583Int. J. Med. Arom. Plants Mycoanalysis of Pleurotus nebrodensis against melanin synthesis
Dangre et al. http://[email protected]
incubated in a humidified atmosphere contain-ing 5% CO2 at 37 oC for 24 hrs, after 24hrs theold media was replaced by fresh media and in-cubated for 24hrs at similar condition mentionedabove. MTT was dissolved in H2O at a concen-tration of 5 mg/ml, and added 20µl to 200µl cul-ture medium. After removing the medium,150µl DMSO was added, and the absorbancewas measured at 570 nm.
Melanogenesis inhibition analysis with culturedcells
Melanocytes were grown in plastic tissueculture flasks and plates in DMEM supplement-ed with 10% FBS and 1% penicillin and strep-tomycin. Cells were incubated at 37 oC in anatmosphere of 5% CO2. The cells were placedin 24-well plastic culture plates at a density of 1x 105cells/well and incubated for 24 hrs in me-dium prior to treatment with the sample. After24 hrs, the medium was replaced with 990µl offresh medium. To this was added 30µl of sam-ple dissolved in DMSO or DMSO only (con-trol). The cells were exposed to the test com-pounds for 4 days at 37 oC and 5% CO2. Themelanin content of cells after treatment was de-termined as follows. After removing the medi-um, cells were washed with PBS and lysed byadding 0.3 ml of 1N NaOH. The crude cell ex-tracts were assayed and analyzed using amicroplate reader (Bio-Rad, USA) at 490 nm todetermine the melanin content. Results fromsamples were analyzed with percent of the con-trol.
Results and discussion
Concentration of sample to inhibit 50% oftyrosinase activity (IC50)
To investigate the tyrosinase activity inhibi-tion, enzyme assay technique was carried outwith L-tyrosine as a substrate. The enzyme-reacted solution was incubated for 15 min at 37oC and the solution was immediately monitoredfor the formation of dopachrome by measuringthe linear increase in absorbance at 490nm. Theresults bring out that the inhibition ability ofsamples extracted under cold condition werelower than that of hot extracts (table 1 and fig-
ure 2). Among the 3 fractionates of the extract,Chloroform fractionate have lower IC50 value(0.42mg) which indicates its highest inhibitionperformance towards tyrosinase activity (Table1). The inhibition by methanol extract waspoorer, while its fractionates had shown the bet-ter inhibitory effect (Figure 2). Arbutin, a posi-tive control had IC50 of 0.55mg.
Table 1: Sample description and IC50 value.
S.No. Samples IC50
1 Arbutin 0.552 80% MeOH extract 0.533 n-Hexane fractionate 0.474 Chloroform fractionate 0.425 Butanol fractionate 0.46
Figure 2: Comparison of IC50 values amongvarious extracts and fractionate samples
Separation of compounds by Thin Layer Chro-matography
To know our extraction sample has single ormulti compound that inhibited tyrosinase activi-ty, we were applied Thin Layer Chromatog-raphy (TLC) to identified.
We detected 2 different compounds in 80%MeOH under cold extraction, 3 different com-pounds in 80% MeOH under hot extraction,100% MeOH under cold and hot extractionwere detected 3 different compounds of each. Ineach organic solvent fraction, sample were alsodetected for finding the compounds by TLCtechnique, the results were revealed and de-scribed as shown in figure 3.
The n-Hexane fractionate of 80% coldMeOH extract shows 4 spots assumed to be ofseparated compounds, whereas n-Hexane frac-

584Int. J. Med. Arom. Plants Mycoanalysis of Pleurotus nebrodensis against melanin synthesis
Dangre et al. http://[email protected]
tionate of 80% hot MeOH extract shows 5 com-pounds spots; Chloroform fractionate of 80%cold MeOH extract and 80% hot MeOH extractshowed 5 and 4 compound spots respectively,while Butanol fractionate of 80% cold and hotMeOH extracts revealed 3 and 1 spots for sepa-rated compounds respectively. In n-Hexanefractionate of absolute cold and hot MeOH ex-tracts 4 and 2 spots of compounds were ob-served respectively, however, in Chloroformfractionate of absolute cold and hot MeOH ex-tracts, 4 and 2 spots observed respectively,while 3 and 2 compounds spots were observedin Butanol fractionate of absolute cold and hotMeOH extract.
a1 a2 b1 b2 c1 c2 d1 d2 e1 e2 f1 f2Figure 3: Thin Layer chromatography of testfractionates.a1 = n-Hexane fractionate of 80% cold MeOHextract, a2 = n-Hexane fractionate of 80% hotMeOH extract; b1 = Chloroform fractionate of80% cold MeOH extract, Chloroform fraction-ate of 80% hot MeOH extract; c1 = Butanolfractionate of 80% cold MeOH extract, c2 =Butanol fractionate of 80% hot MeOH extract;d1 = n-Hexane fractionate of absolute coldMeOH extract, d2 = n-Hexane fractionate ofabsolute hot MeOH extract; e1 = Chloroformfractionate of absolute cold MeOH extract, e2 =Chloroform fractionate of absolute hot MeOH
extract; f1 = Butanol fractionate of absolutecold MeOH extract, f2 = Butanol fractionate ofabsolute hot MeOH extract.
Sample stabilization against high temperatureand UV irradiation
a) Against high temperature: High temperatureand UV radiation are the agents who can af-fect on some compounds which result inchemical structure which leads to lose infunction. To find the compounds which caninhibit tyrosinase activity but also stablewith high temperature and UV light are im-portant to apply sample for life such aswhitening agent or food industry. There-fore, we were exposed sample with hightemperature (60 oC) and UV light, the sam-ple was exposed for 1,2,3,4,5,10 days. Theeffect on sample function was determined bymeasuring IC50 value.
High temperature revealed no effect onsample when exposed for 5 days, the IC50
value was similar as compare with controlsample (not exposed with high temperature),but the sample exposed in 60 oC up to 10days decreased tyrosinase inhibition abilityindicated by increase in the IC50 value (Ta-ble 2a; Figure 4).
b) Against UV irradiation: The UV radiation isalso important agent causes break to somecompound. The sample was exposed on UVlight for 1, 2, 3, 4, 5 and 10 days. The stabi-lization of sample was observed until day 3exposed, but up to 5 and further 10 dayssample was showing decreased tyrosinaseinhibition activity indicated by increase inIC50 value (Table 2b; Figure 4).
Table 2a: Effect of high temperature (60 oC) on sample.
S.No. DaysIC50 of samples in mM
Arbutin 80% MeOHextract
n-Hexane frac-tionate
Chloroformfractionate
Butanol frac-tionate
1 0 0.55 0.53 0.47 0.42 0.442 1 0.56 0.55 0.48 0.43 0.463 2 0.57 0.55 0.48 0.45 0.464 3 0.57 0.57 0.50 0.45 0.495 5 0.59 0.57 0.51 0.47 0.516 10 0.70 0.72 0.69 0.62 0.63

585Int. J. Med. Arom. Plants Mycoanalysis of Pleurotus nebrodensis against melanin synthesis
Dangre et al. http://[email protected]
Table 2b: Effect of UV irradiation on sample.
S.No. DaysIC50 of samples in mM
Arbutin80% MeOH
extractn-Hexane frac-
tionateChloroformfractionate
Butanol frac-tionate
1 0 0.52 0.46 0.42 0.40 0.492 1 0.53 0.48 0.44 0.42 0.493 2 0.54 0.5 0.44 0.41 0.504 3 0.59 0.55 0.50 0.49 0.575 5 0.66 0.66 0.60 0.63 0.646 10 0.78 0.74 0.73 0.69 0.70
Figure 4: Relative samples stabilization and activity check on exposure of (A) Temperature at 60 oCand (B) UV.
Cell viability and proliferation test on melano-cytes
The MTT assay was used to measure viabil-ity and proliferation of cells. The culture cellswere treated with the extracts for 24 hours asdescribed in methods. The degree of cellgrowth was determined by means of the MTTassay. We applied our samples to investigate thecytotoxicity of the sample on melanin cells.Dimethyl sulfoxide (DMSO), arbutin, n-Hexanefractionate, Chloroform fractionate and Butanolfractionate were used for MTT test as toxicagents, with different concentrations as 0.5µl/ml, 1.0µl/ml, 2.0µl/ml, 4.0µl/ml and 8µl/mlfor each. The results were obtained by measur-ing the OD at 570 nm with microplate reader(Bio-Rad).
a) DMSO: Dimethyl Sulfoxide (DMSO), acolorless liquid, is an important polar aproticsolvent that dissolves both polar and nonpo-lar compounds and is miscible in a widerange of organic solvents as well as water.All the samples were dissolved in DMSOand to know the toxicity, DMSO was ap-plied for MTT test on melanocytes as a con-trol. The results were obtained by measur-
ing the OD at 570 nm, the results indicatedno effect on cell viability (figure 5).
b) Arbutin: Arbutin is the inhibitor oftyrosinase and thus prevent formation ofmelanin. Arbutin is therefore used as a skin-lightening agent. Arbutin was added in cul-ture media at different concentration as pre-viously mentioned. In these concentrationsrange, the results revealed that Arbutin doesnot affect the melanocytes viability (figure5), up to 2µg/ml of concentration but thereare a decline in viability which indicates thecytotoxicity with increase in concentrationof Arbutin.
c) n-Hexane fractionate of P. nebrodensis: n-Hexane is a non-polar solvent was used forfractionation. The Hexane fractionate sam-ples dissolved in DMSO was added in cul-ture media at different concentration as men-tioned above. The effects on melanocyte vi-ability were observed by measuring the re-duction of 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) to pur-ple Formazan in the mitochondria of livingcells at OD570nm. We added n-Hexane sam-ple to culture media with concentration
586Int. J. Med. Arom. Plants Mycoanalysis of Pleurotus nebrodensis against melanin synthesis
Dangre et al. http://[email protected]
range (0.5µl/ml, 1µl/ml, 2µl/ml, 4µl/ml,8µl/ml). From 0.5µl/ml to 2µl/ml did not af-fect the Melanocytes, but 4µl/ml sample wasaffecting on viability of cells by 41% and93% when up to 8µl/ml sample.
d) Chloroform fractionate of P. nebrodensis:The Chloroform fractionate was also appliedto investigate cell viability. The testing pro-cess was similar with n-Hexane fractionate.The concentration up to 4µl/ml shows bettertolerance with no significant effect. Howev-er, the culture media having 8µl/ml of sam-ple showed a sharp decline by 90% of cellquantity as compare to the control (figures5).
e) The Butanol fractionate of P. nebrodensis:Sample was applied for cell viability test atsimilar concentrations mentioned above.Sample concentrations up to 2µl/ml werenot cytotoxic for melanocytes, but with4µl/ml concentration, the sample showed asuccessive decline with increase in concen-tration for toxicity (Figure 5).
Figure 5: Comparative analysis of melanocyteviability against different concentrations ofsamples.
Analysis of melanogenesis inhibition in humanmelanocytes with various extracts
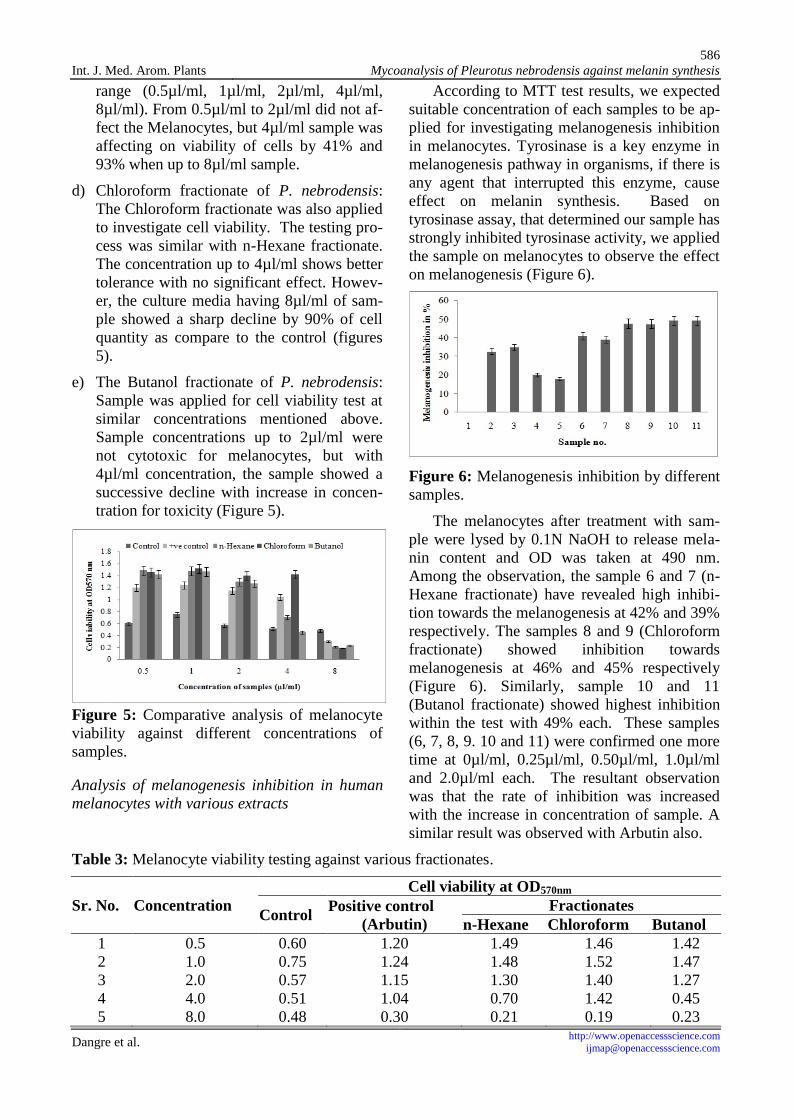
According to MTT test results, we expectedsuitable concentration of each samples to be ap-plied for investigating melanogenesis inhibitionin melanocytes. Tyrosinase is a key enzyme inmelanogenesis pathway in organisms, if there isany agent that interrupted this enzyme, causeeffect on melanin synthesis. Based ontyrosinase assay, that determined our sample hasstrongly inhibited tyrosinase activity, we appliedthe sample on melanocytes to observe the effecton melanogenesis (Figure 6).
Figure 6: Melanogenesis inhibition by differentsamples.
The melanocytes after treatment with sam-ple were lysed by 0.1N NaOH to release mela-nin content and OD was taken at 490 nm.Among the observation, the sample 6 and 7 (n-Hexane fractionate) have revealed high inhibi-tion towards the melanogenesis at 42% and 39%respectively. The samples 8 and 9 (Chloroformfractionate) showed inhibition towardsmelanogenesis at 46% and 45% respectively(Figure 6). Similarly, sample 10 and 11(Butanol fractionate) showed highest inhibitionwithin the test with 49% each. These samples(6, 7, 8, 9. 10 and 11) were confirmed one moretime at 0µl/ml, 0.25µl/ml, 0.50µl/ml, 1.0µl/mland 2.0µl/ml each. The resultant observationwas that the rate of inhibition was increasedwith the increase in concentration of sample. Asimilar result was observed with Arbutin also.
Table 3: Melanocyte viability testing against various fractionates.
Sr. No. ConcentrationCell viability at OD570nm
Control Positive control(Arbutin)
Fractionatesn-Hexane Chloroform Butanol
1 0.5 0.60 1.20 1.49 1.46 1.422 1.0 0.75 1.24 1.48 1.52 1.473 2.0 0.57 1.15 1.30 1.40 1.274 4.0 0.51 1.04 0.70 1.42 0.455 8.0 0.48 0.30 0.21 0.19 0.23

587Int. J. Med. Arom. Plants Mycoanalysis of Pleurotus nebrodensis against melanin synthesis
Dangre et al. http://[email protected]
ConclusionThe mushroom Pleurotus nebrodensis was
used as main source to extract the compoundswhich were expected to inhibit tyrosinase activi-ty involved in melanosgenesis during pigmenta-tion. In order to obtain the effective compounds,the fruit body of the mushroom was extractedwith Methanol followed by fractionation of theextract with various organic solvents. The ex-tracted compounds were analyzed by enzymeassay with mushroom tyrosinase and L-tyrosineas substrate to screen the effective ones beforeinvestigating their effectiveness with culturedcells. The enzyme assay revealed that amongfractionates with organic solvents using n-Hexane, Chloroform and Butanol, the Butanolfractionate was more effective on tyrosinase in-hibition.
To identify the relative amounts of eachcompound in samples, Thin Layer Chromatog-raphy was carried out. Relative to the inhibitionassay, it is necessary to characterize spots ob-served in Butanol fractionates and Chloroformfractionates extensively as we assumes that the-se spots contains the responsible compound formelanogenesis inhibition. All the samples werechosen to test for stabilization against high tem-perature and UV exposed. Samples were ex-posed to 60 oC for 10 days. They showed 90%of initial activity remained up to 5 days and thenthe activity was decreased up to 50%. Similar-ly, 80% of activity was observed by 3 days ofUV exposure on the samples, but dropped to50% after 5 day exposure.
The ultimate goal of this study is to applythe extracts to cosmetic for whitening humanskin with safety on human health and environ-ment. MTT test was carried out to analyze thecytotoxicity of samples with melanocytess andwith human melanocytes as well. The resultsrevealed that the n-Hexane, Chloroform andButanol fractionates have no cytotoxicity up to2µl/ml in average. Melanogenesis inhibitionwas investigated by measuring the melanin con-tent in melanocytes. The fractionates with n-Hexane, Chloroform and Butanol had shownhigh melanogenesis inhibition in melanocytes as41%, 46% and 49% respectively. The Butanolfractionate has revealed a high inhibition in themelanin sythesis within the cells. The fraction-
ate with Chloroform and Butanol showed thehighest melanogenensis inhibition effect amongthe tests, and more efficient than arbutin inmelanogenesis inhibition even in cell cultures ofmelanocytes.
In this study, though potent melanogenesisinhibiting materials were successfully fraction-ated with n-Hexane, Chloroform and Butanolfrom mushroom, Pleurotus nebrodensis, still theresearch is under development for determinationof responsible compound(s) from this mush-room for whitening the skin.
Acknowledgement: The authors are grateful toDr. Arun Jamkar, Hon’ble Vice-Chancellor,MUHS, Nashik, for his continuous encourage-ment towards the research work we have done.
ReferencesAlam, N, Yoon, K.N., Lee, J.S., Lee, M.W.,
Lee, T.S. 2011. Pleurotusnebrodensis ameliorates, atherogenic lipidand histological function inhypercholesterolemic rats. InternationalJournal of Pharmacology, 7: 455-462.
Aroca, P., Garcia-Borron, J.C., Solano, F.,Lozano, J.A. 1990. Regulation of mamma-lian melanogenesis I: partial purificationand characterization of a dopachrome con-verting factor: dopachrome tautomerase.Biochemistry and Biphysics Acta, 1035:266-275.
Brun, A., Rosset, R. 1974. Etudeelectrochimique de 1’oxydation de ladihydroxy-3, 4 phenylalanine (Dopa):mecanisme d’oxydation des catechols enmilieu acide. Journal of ElectroanalyticalChemistry, 49: 287-300.
Cooksey, C.J., Garratt, P.J., Land, E.J., Pavel,S., Ramsden, C.A., Riley, P.A. 1997. Evi-dence of the indirect formation of thecatecholic intermediate substrate responsi-ble for the auto activation kinetics oftyrosinase. Journal of Biological Chemis-try, 272: 26226-26235.
Deshpande, S.S., Sathe, S.K., Salunkhe, D.K.1984. Chemistry and safety of plant poly-phenols. In: Friedman M. (eds), Nutritionaland Toxicological Aspects of Food Safety,Plenum, New York, pp. 457-495,

588Int. J. Med. Arom. Plants Mycoanalysis of Pleurotus nebrodensis against melanin synthesis
Dangre et al. http://[email protected]
Garcia-Canovas, F., Garcia-Carmona, F., Vera-Sanchez, J., Iborra-Pastor, J.L., Lozano-Teruel, J.A. 1982. The role of pH in themelanin biosynthesis pathway. Journal ofBiological Chemistry, 257: 8738-8744.
Graham, D.G., Tiffany, S.M., Vogel, S.F. 1978.The toxixity of melanin precursors. Journalof Investigative Dermatology, 70: 113-116.
Hearing, V.J., King, R.A. 1993. Determinants ofskin color: melanocytes and melanization.In: Levine N. (eds), Pigmentation andPigmentary Abnormalities, CRC Press,New York, pp. 3-32.
Hirobe, T. 1991. Control of melanocyte prolif-eration and dirrerentiation in mouse epi-dermis. Journal of Investigative Dermatol-ogy, 5: 1-11.
Ito, S. 1993. Biochemistry and physiology ofmelanin. In: Levine N. (eds), Pigmentationand Pigmentary Disorders, CRC Press, Bo-ca Raton, FL, pp. 33-59.
Jin, Y.H., Lee, S.J., Chung, M.H.. Park, J.H.,Park, Y.I., Cho, T.H. 1999. Aloesin andarbutin inhibit tyrosinase activity in a syn-ergistic manner via a different action mech-anism. Archives of Pharmacological Re-search 22: 232-236.
Kobayashi, T., Vieira, W.D., Protterf, B., Sakai,C., Imokawa, G. 1995. Modulation ofmelanogenic protein expression during theswitch from eu- to pheomelanogenesis.Journal of Cell Science 108: 2301-2309.
Korytowski, W., Sarna, T., Kalyanaraman, B.,Sealy, R.C. 1987. Tyrosinase-catalyzed ox-idation of dopa and related cate-chol(amine)s: a kinetic electron spin reso-nance investigation using spin-stabilizationand spin label oximetry. Biochemistry andBiophysics Acta, 924: 383-392.
Lambert, C., Chacon, J.N., Chedekel, M.R.,Land, E.J., Riley, P.A., Thompson, A.1989. A pulse radiolysis investigation ofthe oxidation of indolic melanin precursors:evidence for indolequinones and subse-quent intermediates. Biochemistry and Bio-physics Acta, 993: 12-20.
Morgan, B.D.G., O’Neill, T., Dewey, D.L.,Galpine, A.R., Riely, P.A. 1981. Treatmentof malignant Melanoma by intravascular 4-
hydroxyanisole. Clinical Oncology, 7: 227-234.
Pawelek, J.M. 1991. After dopachrome? Pig-ment Cell Research, 4: 53-62.
Priestley, G.C. 1993, Molecular Aspects ofDermatology, Second ed. Wiley Publica-tions, Chichester, UK.
Prota, G. 1988. Progress in the chemistry ofmelanin and related metabolites. MedicalResearch Review, 8: 525-556.
Prota, G. 1992. Melanins and Melanigenesis,Academic Press, New York, pp. 1-290.
Riely, P.A., Sawyer, B., Wolff, M.A. 1975. Themelanocytotoxic effect of 4-hydroxyanisole. Journal of InvestigativeDermatology, 64: 86-89.
Rodriguez-Lopez, J.N., Tudela, J., Varon, R.,Garcia-Canovas, F. 1991. Kinetic study onthe effect of pH on the melanin biosynthe-sis pathway. Biochemistry and BiophysicsActa, 1076: 379-386.
Sachs, C., Jonsson, G. 1975. Mechanisms ofaction of 6-hydroxydopamine. Biochemis-try and Pharmacoogy, 24: 1-8.
Seiberg, M., Paine, C., Sharlow, E., Andrade-Gordon, P., Constanzo, M., Eisinger, M.2000. The protease-activated receptor 2regulates pigmentation via keratinocyte-melanocyte interactions. Experimental CellResearch 254: 25-32.
Seiberg, M., Paine, C., Sharlow, E., Andrade-Gordon, P., Costanzo, M., Eisinger, M.2000. Inhibition of melanosome transfer re-sults in skin lightening. Journal of Investi-gative Dermatology, 115: 162-167.
Spritz, R.A., Hearing, V.J.Jr. 1994. Geneticdisorders of pigmentation. Advances inHuman Genetics, 22:1-45.
Sugumaran, M., Semesi, V. 1991. Quinonemethide as a new intermediate in eumelaninbiosynthesis. Journal of Biological Chemis-try, 266: 6073-6078.
Wick, M.M., Byers, L., Frei, F. 1977. L-DOPA:selective toxicity for Melanoma cells invitro. Science, 197: 468-469.
Wick, M.M., Byers, L., Ratliff, J. 1979. Selec-tive toxicity of 6-hydroxydopa for Mela-noma cells. Journal of Investigative Der-matology, 72: 67-69.
Copyright © 2022 FDOKUMEN




















