
Conservation of key elements of natural competence in Lactococcus lactis ssp
11
Conservation of key elements of natural competence in Lactococcus lactis ssp. Sandra Wydau 1 , Rozenn Dervyn 1 , Jamila Anba 2 , S. Dusko Ehrlich 1 & Emmanuelle Maguin 1 1 G´ en´ etique Microbienne and 2 Unit ´ e d’Ecologie et de Physiologie du Tube Digestif, INRA, Domaine de Vilvert, Jouy-en-Josas Cedex, France Correspondence: Emmanuelle Maguin, Unit ´ e de G ´ en´ etique Microbienne, INRA, Domaine de Vilvert, 78352 Jouy-en-Josas Cedex, France. Tel.: 133 1 34 65 25 18; fax: 133 1 34 65 25 21; e-mail: [email protected] Received 23 September 2005; revised 22 December 2005; accepted 2 January 2006. First published online 16 February 2006. doi:10.1111/j.1574-6968.2006.00141.x Editor: Andre Klier Keywords sigma factor; regulation; late competence genes; genetic diversity; comX, dprA. Abstract Natural competence is active in very diverse species of the bacterial kingdom and probably participates in horizontal gene transfer. Recently, the genome sequence of various species, including Lactococcus lactis, revealed the presence of homologues of competence genes in bacteria, which were not previously identified as naturally transformable. We investigated the conservation among lactococcal strains of key components of the natural competence process in streptococci: (i) comX which encodes a sigma factor, allowing the expression of the late competence genes involved in DNA uptake, (ii) its recognition site, the cin-box and (iii) dprA which encodes a protein shown to determine the fate of incoming DNA. The comX and dprA genes and the cin-box appeared conserved among strains, although some L. lactis ssp. lactis strains presented an inactivated dprA gene. We established that ComX controls the expression of the late competence genes in L. lactis. In conclusion, our work strongly suggests that ComX has the same role in streptococci and L. lactis, i.e. the regulation of late competence genes. It also allowed the identification of a set of L. lactis strains and the construction of a comX overexpression system, which should facilitate the investigation of the natural competence activity in lactococci. Introduction Natural competence is a cellular state allowing the uptake of exogenous DNA by bacteria and its integration in the chromosome. This process is active in diverse species of the bacterial kingdom and participates in intra- and probably also inter-species gene transfer (Dreiseikelmann, 1994; Lor- enz & Wackernagel, 1994; Cvitkovitch, 2001; Claverys & Martin, 2003). Natural competence has been reported in several streptococci, e.g. in the Streptococcus mitis and Streptococcus mutans groups, and studied in detail in Strep- tococcus pneumoniae (Havarstein et al., 1997; Cvitkovitch, 2001). In streptococci, natural competence is controlled by a quorum-sensing system, which involves the production, the export-maturation and the detection by a two-component system (ComD, ComE) of a competence stimulating peptide (CSP) (Pestova et al., 1996). The phosphorylated response regulator, ComEP, stimulates the transcription of the genes responsible for CSP synthesis, export and detection, and also of the comX1 and comX2 genes that encode two homologous sigma factors (Lee & Morrison, 1999). Each of these sigma factors can initiate the next step of the competence process by switching on the expression of the late competence genes and stimulating the expression of competence-associated genes (Lee & Morrison, 1999). In order to stimulate gene expression, ComX interacts with the RNA polymerase core enzyme and the resulting holoenzyme recognizes a specific sequence, the cin-box located upstream of late competence genes (Lee & Morrison, 1999; Opdyke et al., 2001). In this system, ComX has a key role, linking the regulatory module constituted by the early competence genes to the DNA uptake module encoded by the late competence genes. The ComX-regulated genes are directly involved in the uptake of exogenous DNA (Claverys & Havarstein, 2002), in its protection from intracellular nu- cleases by the DprA protein (Berge et al., 2003), and in its incorporation in the chromosome by homologous recombi- nation (Mortier-Barriere et al., 1998). Lactococcus lactis is a gram-positive lactic acid bacterium (LAB), which is closely related to the streptococci (Stiles & Holzapfel, 1997). The two major L. lactis subspecies, L. lactis ssp. lactis and L. lactis ssp. cremoris, are industrially impor- tant as they are used as starters for the production of fermented dairy food (Stiles & Holzapfel, 1997). Lactococcus lactis was never identified as a species possessing a natural FEMS Microbiol Lett 257 (2006) 32–42 c 2006 Federation of European Microbiological Societies No claim to original French government works
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Conservation of key elements of natural competence in Lactococcus lactis ssp

Conservationof keyelementsof natural competence inLactococcus lactis ssp.Sandra Wydau1, Rozenn Dervyn1, Jamila Anba2, S. Dusko Ehrlich1 & Emmanuelle Maguin1
1Genetique Microbienne and 2Unite d’Ecologie et de Physiologie du Tube Digestif, INRA, Domaine de Vilvert, Jouy-en-Josas Cedex, France
Correspondence: Emmanuelle Maguin,
Unite de Genetique Microbienne, INRA,
Domaine de Vilvert, 78352 Jouy-en-Josas
Cedex, France. Tel.: 133 1 34 65 25 18;
fax: 133 1 34 65 25 21; e-mail:
Received 23 September 2005; revised 22
December 2005; accepted 2 January 2006.
First published online 16 February 2006.
doi:10.1111/j.1574-6968.2006.00141.x
Editor: Andre Klier
Keywords
sigma factor; regulation; late competence
genes; genetic diversity; comX, dprA.
Abstract
Natural competence is active in very diverse species of the bacterial kingdom and
probably participates in horizontal gene transfer. Recently, the genome sequence of
various species, including Lactococcus lactis, revealed the presence of homologues
of competence genes in bacteria, which were not previously identified as naturally
transformable. We investigated the conservation among lactococcal strains of key
components of the natural competence process in streptococci: (i) comX which
encodes a sigma factor, allowing the expression of the late competence genes
involved in DNA uptake, (ii) its recognition site, the cin-box and (iii) dprA which
encodes a protein shown to determine the fate of incoming DNA. The comX
and dprA genes and the cin-box appeared conserved among strains, although
some L. lactis ssp. lactis strains presented an inactivated dprA gene. We
established that ComX controls the expression of the late competence genes in
L. lactis. In conclusion, our work strongly suggests that ComX has the same role in
streptococci and L. lactis, i.e. the regulation of late competence genes. It also
allowed the identification of a set of L. lactis strains and the construction of a comX
overexpression system, which should facilitate the investigation of the natural
competence activity in lactococci.
Introduction
Natural competence is a cellular state allowing the uptake of
exogenous DNA by bacteria and its integration in the
chromosome. This process is active in diverse species of the
bacterial kingdom and participates in intra- and probably
also inter-species gene transfer (Dreiseikelmann, 1994; Lor-
enz & Wackernagel, 1994; Cvitkovitch, 2001; Claverys &
Martin, 2003). Natural competence has been reported in
several streptococci, e.g. in the Streptococcus mitis and
Streptococcus mutans groups, and studied in detail in Strep-
tococcus pneumoniae (Havarstein et al., 1997; Cvitkovitch,
2001).
In streptococci, natural competence is controlled by a
quorum-sensing system, which involves the production, the
export-maturation and the detection by a two-component
system (ComD, ComE) of a competence stimulating peptide
(CSP) (Pestova et al., 1996). The phosphorylated response
regulator, ComE�P, stimulates the transcription of the
genes responsible for CSP synthesis, export and detection,
and also of the comX1 and comX2 genes that encode
two homologous sigma factors (Lee & Morrison, 1999).
Each of these sigma factors can initiate the next step of the
competence process by switching on the expression of the
late competence genes and stimulating the expression of
competence-associated genes (Lee & Morrison, 1999). In
order to stimulate gene expression, ComX interacts with the
RNA polymerase core enzyme and the resulting holoenzyme
recognizes a specific sequence, the cin-box located upstream
of late competence genes (Lee & Morrison, 1999; Opdyke
et al., 2001). In this system, ComX has a key role, linking the
regulatory module constituted by the early competence
genes to the DNA uptake module encoded by the late
competence genes. The ComX-regulated genes are directly
involved in the uptake of exogenous DNA (Claverys &
Havarstein, 2002), in its protection from intracellular nu-
cleases by the DprA protein (Berge et al., 2003), and in its
incorporation in the chromosome by homologous recombi-
nation (Mortier-Barriere et al., 1998).
Lactococcus lactis is a gram-positive lactic acid bacterium
(LAB), which is closely related to the streptococci (Stiles &
Holzapfel, 1997). The two major L. lactis subspecies, L. lactis
ssp. lactis and L. lactis ssp. cremoris, are industrially impor-
tant as they are used as starters for the production of
fermented dairy food (Stiles & Holzapfel, 1997). Lactococcus
lactis was never identified as a species possessing a natural
FEMS Microbiol Lett 257 (2006) 32–42c� 2006 Federation of European Microbiological SocietiesNo claim to original French government works

competence pathway and to the best of our knowledge,
natural transformation was never observed in L. lactis
strains. However, the analysis of the complete genome
sequence of IL1403, an L. lactis ssp. lactis strain, revealed
the presence of orthologs of several genes involved in the
process of natural competence in various bacteria (Bolotin
et al., 1999, 2001; Claverys & Martin, 2003). Based on
sequence homology, a comX gene and all the late compe-
tence genes required for the formation of the DNA entry
pore [comC (also referred to as CCL in S. pneumoniae),
comEA, comEC, comFA, comFC and comGA to comGD] were
identified (Bolotin et al., 1999, 2001). Comparison of the
regions located upstream of these late competence genes
allowed the identification of a conserved sequence which
may constitute a recognition site of the ComX-RNA poly-
merase holoenzyme (Bolotin et al., 1999, 2001). These
genetic features of IL1403 raise the question whether L. lactis
can activate a natural competence system which may con-
tribute to genetic transfer in dairy food ecosystems (Gasson,
2000; Jonas et al., 2001; Kharazmi et al., 2002; van den Eede
et al., 2004).
Two observations questioned the functionality of natural
competence in Lactococcus lactis. The first observation is that
orthologs of the early competence genes (the ABC transpor-
ter encoding comA and comB genes, the two-component
system encoding genes comD and comE, and the CSP
encoding gene) responsible for the regulation by the CSP,
were not clearly identified in IL1403 (Bolotin et al., 2001).
The strain does, however, possess multigenic families
encoding ABC transporters (at least 60 ABC transporters
in IL1403) and two-component systems (seven complete
systems and an additional response regulator) (Bolotin
et al., 2001). In IL1403, Redon et al. (2005) observed a
slight progressive overexpression of comX during glucose
starvation and proposed that specific starvation condi-
tions may lead to the induction of natural competence in
L. lactis.
The second observation is that the IL1403 dprA gene (also
referred to as smf in several microorganisms as Bacillus
subtilis) is inactivated by a frameshift resulting in a pre-
mature stop at nt 100 (protein of 33 amino acids). In several
bacteria (Karudapuram et al., 1995; Ando et al., 1999;
Smeets et al., 2000; Berka et al., 2002; Friedrich et al., 2002;
Ogura et al., 2002), a determining role for DprA in the fate
of the exogenous DNA taken up during competence was
demonstrated. In an S. pneumoniae strain mutated for dprA,
the incoming DNA is degraded and the nucleotides are
randomly incorporated in the chromosome, while in a wild-
type background the incoming DNA fragments are inte-
grated in the chromosome by homologous recombination
(Berge et al., 2003). Since DprA activity is crucial for the
outcome of the natural transformation process and its
contribution to horizontal gene transfer, we investigated
whether L. lactis dprA1 strains could be identified in a
collection of strains.
In order to evaluate if natural competence may allow the
development of a general genetic tool for the modification
of lactococcal strains, we investigated the conservation of
two other key elements of the natural competence system,
comX and the cin-box, in lactococci. Finally, as a prelude to a
study on the functionality of natural competence in L. lactis,
we overexpressed two alleles of the comX gene in IL1403 to
test whether they regulate the expression of late competence
genes.
Materials and methods
Bacterial strains and culture conditions
For this study, 31 lactococcal strains (Table 1) were chosen
in the INRA national collection (INRA collection, URLGA,
Jouy-en-Josas, France). These strains were classified in the
different lactococcal subspecies (ssp. lactis, ssp. lactis biovar.
diacetylactis and ssp. cremoris) according to their pheno-
types (growth at 40 1C or in the presence of 4% NaCl and
arginine or citrate metabolism, J. C. Ogier, pers. comm.).
The strain VI 7101 is a derivative of IL1403, which
contains the nisR and nisK genes integrated in the chromo-
somal histidine biosynthesis operon (S. Calero and P.
Renault, pers. comm.). Strain VI 7238 is a comX-deleted
mutant of VI 7101 which resulted from a double crossing-
over (DCO) using plasmid pVI6237 (carrying a 2 kb DNA
insert constituted of the upstream (1 kb) and downstream
(1 kb) regions of comX fused together). The DCO was
performed as previously described for L. lactis (Biswas
et al., 1993) and the mutated strain was identified among
the ery-sensitive clones by PCR with the S99 and S102
primers (Table 2). The chromosomal structure of VI 7238
was then confirmed by Southern hybridization of digested
chromosomal DNA (HindIII, XmnI and EcoRI) using a PCR
fragment generated with primers S124 and S185 (Table 2) as
a probe. The VI 7293, VI 7247 and VI 7298 strains resulted
from the transformation of VI 7238 with plasmids pVI6253
(pnisA), pVI6213 (pnisA:comXIL) and pVI6255 (pnisA::-
comXMG), respectively. The pnisA promoter was induced by
the addition of nisin (0.8 ng mL�1, Sigma, Lyon, France) in
exponential phase cultures (OD600 � 0.1) growing in SA
medium (Jensen & Hammer, 1993) at 30 1C. Otherwise,
strains were propagated at 30 1C in M17 (Terzaghi &
Sandine, 1975) supplemented with 0.5% glucose. Erythro-
mycin (ery) was added at 5mg mL�1.
DNA extraction and PCR amplification
Chromosomal DNA was extracted as previously described
(Biswas et al., 1993). PCR DNA amplifications were per-
formed with the ExTaq DNA polymerase (Takara, St Germain
FEMS Microbiol Lett 257 (2006) 32–42 c� 2006 Federation of European Microbiological SocietiesNo claim to original French government works
33Natural competence in L. lactis

en laye, France) and the appropriate primers (Table 2) as
recommended by the supplier. For the comX and dprA
genes, primers homologous to the intergenic regions flank-
ing the gene of interest in the IL1403 genome sequence were
designed. These primers allowed to amplify fragments from
all the strains of the lactis subspecies (as determined by the
16S analysis, Table 1) but not from the strains belonging to
the cremoris ssp. For the cremoris ssp. strains (as determined
with the 16S analysis), an internal fragment of the gene of
interest was first amplified and sequenced (comX, a�300 bp
fragment obtained with the primer pairs S141–S143 or
S142–S143 and dprA, a 703 bp fragment obtained with
S183–S184). Then, the upstream and downstream regions
were amplified using primers homologous to this internal
fragment (comX, S143 or S141 and dprA, S183 or S184) and
the flanking genes (comX, S125, S126 or S127 and dprA, S98
or S186). The promoter region of the comG operon was
amplified using primers homologous to the upstream (S149,
S152 or S150) and downstream (S150, S154 or S155)
sequences.
16S analysis
In order to characterize the 16S sequence of the 31 lacto-
coccal strains selected from the INRA collection, an internal
fragment (343 bp) of the 16S ribosomal RNA genes was
amplified using the Y1 and Y2 primers described by Ward
et al. (1998) and chromosomal DNA as templates. The PCR
products were then digested by CfoI and MboII to determine
to which subspecies the strain belonged (Ward et al., 1998).
DNA sequencing and analysis
PCR generated fragments were sequenced using the Big Dye
Terminator v3.1 (Applied Biosystems, Foster City, CA).
Reaction products were analysed using a capillary sequencer
(3700 DNA Analyser, ABI PRISM). Sequences were analysed
using the GCG package (University of Wisconsin) and were
aligned using CLUSTALW (Infobiogen, http://www.infobio-
gen.fr/) (Thompson et al., 1994), the percentage of diver-
gence between the alleles was calculated by MEGA 2.1
(Kumar et al., 2001) and the data of the phylogenetic
analysis were represented using TREEVIEW (Page, 1996).
All sequences were deposited in the EMBL database under
the following accession numbers: comX genes AJ890847,
AJ890848, AJ890849, AJ890850, AJ890851, AJ890852,
AJ890853, AJ890854, AJ890855, AJ890856, AJ890857,
AJ890858, AJ890859, AJ890860, AJ890861, AJ890862,
AJ890863, AJ890864, AJ890865, AJ890866, AJ890867,
AJ890868, AJ890869, AJ890873, AJ890874, AJ890875,
AJ890876, AJ890878, AJ890879, AJ890881, dprA genes
AJ890918, AJ890919, AJ890920, AJ890921, AJ890922,
AJ890923, AJ890924, AJ890925, AJ890926, AJ890927,
AJ890928, AJ890929, AJ890930, AJ890932, AJ890933,
AJ890934, AJ890935, AJ890936, AJ890937, AJ890938,
AJ890944, AJ890945, AJ890946, AJ890947, AJ890948,
AJ890949, AJ890950, AJ890951, AJ890952, AJ890953.
Construction of plasmids
pVI6237 is a derivative of the thermosensitive pG1host9
plasmid (Maguin et al., 1996) which carries a �2 kb
SpeI–XhoI insert (a fusion of two 1 kb fragments
Table 1. Strains of the INRA collection used in this study
Strains
Subspecies according to the
Phenotype 16S analysis�
RAPD Group 1
IL1403 lactis lactis
IL581w lactis cremoris
IL584 lactis lactis
IL1321 lactis lactis
A13 lactis lactis
NCDO2146 lactis lactis
IL1306 lactis bv. diacetylactis lactis
DRC1 lactis bv. diacetylactis lactis
A15w cremoris lactis
A11 lactis lactis
A17 lactis lactis
A26 lactis lactis
NCDO604 lactis lactis
A7 lactis lactis
A8 lactis bv. diacetylactis lactis
A27 lactis bv. diacetylactis lactis
RAPD Group 2
CNRZ359 cremoris cremoris
C7 cremoris cremoris
AM1w cremoris lactis
AM2w cremoris lactis
RAPD Group 3
CO2 cremoris cremoris
CO4 cremoris cremoris
NCDO276w lactis bv. diacetylactis cremoris
SK1w lactis cremoris
MG1363 cremoris cremoris
A140w lactis cremoris
IL578w lactis cremoris
IL582w lactis cremoris
RAPD individual patterns
NCDO2091 lactis lactis
NCDO2118 lactis lactis
NCDO2633 lactis lactis
Groups 1, 2 and 3 correspond to the RAPD patterns the most frequently
identified in the CNRZ collection of lactococci (Tailliez et al., 1998).
Strains exhibiting specific RAPD patterns are referred to as RAPD
individual patterns.�The 16S analysis did not discriminate between Lactococcus lactis ssp.
lactis and L. lactis ssp. lactis biovar diacetylactis strains.wStrains indicated with were misclassified on the basis of their phenotypes.
Bv., biovariant.
FEMS Microbiol Lett 257 (2006) 32–42c� 2006 Federation of European Microbiological SocietiesNo claim to original French government works
34 S. Wydau et al.

Table 2. Oligonucleotides used in this study
Primer Localization Orientation Sequence 50 to 30
comX sequence
S57 Downstream of comX Reverse TCTTCTCTTATCAAAAAACTCCC
S125 16S Reverse AGAGCCGCTTTCGCCAC
S126 ezrA Forward TTAGCTGAACAATTGATTCAATATG
S127 16S Reverse CCTGAGCCAGGATCAAACTCTC
S141 comX Forward TAGTTATGAAATTAATGAAACAAATTCG
S142 comX Forward CATTAGAACATGGAAAATAGAGGATTAT
S143 comX Reverse GTAGTAAGTCTTGATAATGTGCGCTC
dprA sequence
S58 Upstream of dprA Forward AAATTGCTGACAAAGCTGTCAG
S59 Downstream of dprA Reverse TAGCGAAGTGGTTTTTCTCCG
S97 dprA Forward ATCTTGCTAAAAACCAACTCATAC
S98 topA Reverse TTGTTTTACTTTTAGTTGAAGTTGG
S183 dprA Forward TGGCTCAAGTCAAATCAATCCC
S184 dprA Reverse TAAAATATCTTGAGCTTGATAGAC
S186 aldR Forward GTCCTTATTGAAATAGAAGTCATTG
cin-box sequence
S149 polC Forward GAATGCCTGATGATAATCAATTAG
S150 comGA Reverse TTTCCTCTATTTCATACCAACAAG
S152 polC Forward TAGAGCGTGGTTTCACTTTTGG
S153 polC Reverse CTTAATTGATTATCATCAGGCATTC
S154 comGA Reverse CGTTGACTTGTAACTGAATAAAATC
S155 comGA Forward TTGTTGGTATGAAATAGAGGAAAG
Gene expression
S50 comX Forward CATGGAAAATAGAGGATTATCTTC
S51 comX Reverse GCGATACCGTTGCATGCGAG
S192 comX Forward GGTGGTCATATGAAACCAATCATCAGAAAAT
S193 comX Reverse GGTGGTCCGCGGTTAATCATCATCTCGAGAAAAT
S19 comGA Reverse GGGTTCTTCTATATTGATAACTTG
S21 comGA Forward TTAGAAGTTGGCTTAGCACTAC
S22 comGB Forward TTTGGTAGAAGTGCATGGTAAC
S23 comGB Reverse TGAGCAATTAGATTTCCCCATTC
S24 comGC Forward GCCGTCAGAAAGAGCTAAAAG
S25 comGC Reverse TTTTTGAGTAATCATCCCTGCAC
S26 comGD Forward CATTTACTTTACTAGAGTCTCTTC
S27 comGD Reverse TCTTTAACTGCTACCTCCTTTGG
S28 comEA Forward GTGCGGTAACAAAGCCTAATG
S29 comEA Reverse CCGAAAATCGATGATGTCTTGAG
S30 comEC Forward GAATGGCTAATTCATGAGGTTG
S31 comEC Reverse AGCTGGTCGATTTGACTTACTC
S32 comFA Forward TGTTGGTCTAGCTAGTCCAAG
S33 comFA Reverse TGAAAACGACGCGGGAGAAAC
S34 comFC Forward CGTGGTTTCAATCAAGTGACTG
S35 comFC Reverse ATGCGTGATATAAGGTGGTACC
S36 comC Forward TGGCTCACATTGCCGTTATTG
S37 comC Reverse TTGAATTTTTCAGCTAAGACGGC
S42 dprA Forward GAGAGAAGAGTATAAAATATACCC
S43 dprA Reverse TAATGGCAGAGAGATGACTTGC
S44 coiA Forward TCTCAAAGCTTGCGACTTATGG
S20 coiA Reverse AGGGATTTTAATCTCATCTTCTGAGC
S38 radA Forward TTGGTCGAAGTGACTAATCCC
S39 radA Reverse CAACCGCAACCGCGAGGTC
S266 radC Forward TTCTTCAACATTTTGAAACTTTGG
S267 radC Reverse CAGTTGCCTGATTAACCGCC
S72 recA Forward ATGCAAAAGCGCTCGGTGTC
S73 recA Reverse TCTTGTTATCTCCAGAACCTTC
S40 recQ Forward CAGGACGGGACGGGCTTG
FEMS Microbiol Lett 257 (2006) 32–42 c� 2006 Federation of European Microbiological SocietiesNo claim to original French government works
35Natural competence in L. lactis

corresponding to the upstream and downstream regions of
comX and amplified with primers S99–S135 and S136–S102,
respectively). pVI6213 is a derivative of the pJIM2246 vector
(Renault et al., 1996) which contains between its XhoI and
SacI sites a DNA insert containing 3 successive transcription
terminators (T3) followed by a fusion between the inducible
promoter pnisA and the comX gene of IL1403 (comXIL). This
XhoI-SacI insert was generated as described below. (i) The
pnisA::comXIL fusion was amplified by PCR using the S106
and S107 primers and as template the chromosomal DNA of
VI 7145, which contains the pnisA::comXIL fusion. This PCR
product was cloned in the pGEM-T vector (Promega,
Charbonnieres, France) leading to pVI6205. (ii) The
EcoRV-SacI fragment of pVI6205 (containing the fusion)
was cloned in pVI6207 (pBluescriptSKII containing T3)
leading to pVI6212 which contains the T3::pnisA::comXIL
fusion on an XhoI–SacI fragment. In order to generate
pVI6253 (pJIM2246::T3::pnisA), the comXIL gene was deleted
from pVI6213 by SacI and NcoI digestions followed by a
Klenow-T4 DNA polymerase treatment and self-ligation.
Finally, plasmid pVI6255 (pJIM2246::T3::pnisA::comXMG)
was obtained after SacI and NcoI digestions of pVI6253 and
ligation with a SacI–NcoI PCR fragment containing comXMG
(amplified from the MG1363 chromosomal DNA with
primers S260 and S261).
Extraction of total RNAs
A volume of 100 mL of cultures at OD600 � 0.1 were
centrifuged (5 min, 9800 g at 4 1C) and the cell pellet was
washed with cold TE. The pellet was resuspended in 500mL
of cold water and transferred in a 2 mL screw-cap micro-
centrifuge tube containing 0.6 g of glass beads (Sigma),
200 mL of Macaloid (2%) (Sambrook et al., 1989), 500 mL of
phenol–chloroform pH 4.7 and 25mL of SDS (20%). Cells
were disrupted by shaking in a Fastprep machine (BIO101)
for 40 s at speed 5.5. After centrifugation at 12 000 g for
15 min (4 1C), the aqueous supernatant, which contains the
RNA, was treated with phenol-chloroform pH 4.7, precipi-
tated with ethanol and resuspended in 30 mL of water.
Dnase treatment and RT-PCR
In all, 5mg of total RNAs diluted in water (15mL final) were
incubated 3 min at 95 1C and 10 min on ice. 3mL of Dnase
(2 unitsmL�1, Ambion, Huntington, UK) and 2mL of buffer
were added, and the mixture was incubated 1.5 h at 37 1C. To
synthesize cDNAs 1mg of total RNA was incubated 10 min at
25 1C and 5 min on ice with 1mL of dNTPs (10 mM,
Promega) and 1mL of random primer (500 ngmL�1, NEB,
Beverly, MA) in a final volume of 13mL. A volume of 1mL
of reverse transcriptase (M-MLV Reverse transcriptase kit,
Invitrogen Cergy-Pontoise, France), 4mL of 5� buffer, 2mL
of DTT (0.1 mM) were then added and the mixture was
incubated 1 h at 37 1C and 15 min at 75 1C. The PCR
amplification (final volume 50mL) was performed from
cDNA (1mL) using appropriate primers (Table 2), Extaq
polymerase (Takara), dNTPs and the reaction buffer as
recommended by the supplier (Takara). The amplification
was carried out as follows: 5 min at 94 1C, (30 s at 94 1C, 30 s
Primer Localization Orientation Sequence 50 to 30
S41 recQ Reverse TCGGACGACACAAGATAGGAC
S264 ssbA Forward GTCCGTGTGACCTTGGCAG
S265 ssbA Reverse CTCGTAAAGCGATTGTTGCCC
S268 yqfG Forward TGCCTCCTATGCTGGCTTAG
S269 yqfG Reverse TTGGAAATTCTATACTACTTATGG
S272 ywgA Forward TTAAAACGACTTTACCGTGTGG
S273 ywgA Reverse GTTTTTTGTAAGAATATTTATGACCTC
S52 hu Forward CTAACAAACAAGATCTTATCGCTG
S53 hu Reverse GAACAACTGTAGCAGCGATTTTG
Plasmid construction
S99 Upstream of comX Forward GATATACTAGTGTTAGAGGTTATCAAGAAC
S102 Downstream of comX Reverse CTCAACTCGAGGTCTACCAGTTTCCAATG
S106 Upstream of pnisa Forward CATCAGATATCAACCAATCACGTCCGAG
S107 Downstream of comX Reverse TCACTGAGCTCAGTGATTGGGAATTCCTC
S124 Upstream of comX Forward ACGTTCATCAGTTCTCAAAG
S135 Upstream of comX Reverse AAACTCCCTAGAATTCCTATAGGTTTCATTTCTGAAAAAAG
S136 Downstream of comX Forward AAACCTATAGGAATTCTAGGGAGTTTTTTGATAAGAGAAG
S185 Upstream of comX Reverse GCTGCACCCGGTTCTAC
S260 comXMG Forward GGTGGTCCATGGAAAAGAGGAATATCAATGACAT
S261 comXMG Reverse ACCACCGAGCTCAGGGAGCTTTTTTTAATCATC
All the oligonucleotides were designed on the sequence of IL1403, except S183 and S184 which were designed on the sequence of MG1363.
Table 2. Continued
FEMS Microbiol Lett 257 (2006) 32–42c� 2006 Federation of European Microbiological SocietiesNo claim to original French government works
36 S. Wydau et al.

at 57 1C, 1 min at 72 1C) 30 cycles. Note that the expression of
ydbC was not monitored since the 2 pairs of oligonucleotides
that we tested did not allow PCR amplification from a DNA
template.
Results
Characterization of the lactococcal strains
We selected 31 lactococcal strains including the two best
characterized Lactococcus lactis strains, IL1403 (ssp. lactis)
and MG1363 (ssp. cremoris) in the INRA national collection.
These strains belong to the three major RAPD groups
defined for the L. lactis strains (Tailliez et al., 1998) and
according to their phenotypes, they belong to the two major
L. lactis subspecies used in dairy industry (ssp. lactis and ssp.
cremoris, Table 1).
In order to better characterize the chosen strains, their
16S sequences were analysed in a region which allowed to
distinguish between the lactis and cremoris ssp. (Ward et al.,
1998). Our data revealed that nine strains out of 31 (more
than 29%) were misclassified on the basis of their pheno-
types (Table 1). According to the 16S data, our set of strains
comprises 20 strains of the lactis ssp. and 11 of the cremoris
ssp. It is noteworthy that according to this new strain
classification, our sample of the RAPD group 3 contained
exclusively strains of cremoris spp.
Sequence analysis of the dprA gene
Sequence analysis revealed that in IL1403, the dprA gene
contained a premature stop codon at position 34; the
resulting truncated protein of 33 residues (instead of 282)
is likely to be inactive. As DprA is expected to play a crucial
role in natural transformation, its sequence was determined
in all of the other 30 strains.
In all the strains belonging to the lactis ssp. according
to the 16S analysis, two fragments (632 and 592 bp in
IL1403), which together contained the dprA gene were
amplified. For the remaining 11 strains of the cremoris ssp.,
unsuccessful amplification assays prompted us to change
our strategy. An internal fragment (703 bp) of dprA was
amplified and sequenced. The upstream and downstream
regions were then amplified using oligonucleotides located
in the internal sequenced region and in the dprA neighbour-
ing genes.
The maximal dprA divergence was 21.7% and the sequence
analyses revealed two allelic types, one detected in all the
L. lactis ssp. lactis strains and the other in the ssp. cremoris
strains; they are referred to as the lactis- and the cremoris-
allelic types, respectively (Fig. 1a). The lactis- and cremoris-
type exhibited an intratype variation of 3.1% and 3.7%,
respectively. In the lactis-type, 10 genes (50%) presented a
premature stop codon that would result in truncated proteins
of 23 aa (A8), 33 aa (IL584, IL1403, DRC1), 167 aa (A7) or
193 aa (IL1306, A17, A15, AM2, IL1321). In all these cases,
the resulting protein is expected to be inactive. For the
cremoris-allelic type, all the predicted proteins had the full
size of 282 aa.
Complete dprA genes were identified in �68% of the
tested strains, and in both the lactis and cremoris ssp. Since
some strains contain an intact dprA gene, we were interested
to see if the upstream regulatory elements were conserved;
we therefore investigated the conservation of two other key
elements for natural competence, comX and the cin-box,
which regulate the expression of the late competence genes
in S. pneumoniae.
Genetic variability of the comX gene amonglactococci
As in the case of the dprA gene, the primers homologous to
the intergenic regions flanking the comX gene in IL1403 led
to a productive PCR only for the strains belonging to the
lactis ssp. Other primers were designed for amplification
from the cremoris strains (material and methods). The comX
comX
max div. = 27.5%
0.1
IL1403IL584DRC1AM1A11A13A26A8NCDO2118NCDO2633NCDO2146NCDO604A27A7A17IL1321IL1306A15AM2NCDO2091
CO4CO2C7CNRZ359IL578NCDO276MG1363SK1IL582IL581A140
(b)(a) dprA
max div. = 21.7%
0.1
A15A17AM2IL1321IL1306DRC1IL584AM1A11A26A13A8IL1403NCDO2146NCDO604NCDO2633NCDO2118A27A7NCDO2091
CO4CO2IL581C7CNRZ359IL578MG1363SK1A140NCDO276IL582
*******
**
*
Fig. 1. Neighbour-joining unrooted phylogenetic trees inferred from the
sequences of Lactococcus lactis genes. (a) dprA sequences. (b) comX
sequences. The multiple nucleotide sequences were compared by
CLUSTALW (Thompson et al., 1994). The tree representation was
obtained with TREEVIEW (Page, 1996). The maximal divergence (max.
div.) was calculated with MEGA 2.1 (Kumar et al., 2001). Asterisks (�)
indicate the strains in which the gene is inactivated.
FEMS Microbiol Lett 257 (2006) 32–42 c� 2006 Federation of European Microbiological SocietiesNo claim to original French government works
37Natural competence in L. lactis

gene was detected in the sequence of all of the 31 fragments.
The maximal divergence of the comX nucleotide sequences
was 27.5% and all genes were intact. Further sequence
comparison using CLUSTALW revealed two allelic types
(Fig. 1b).
One allelic type was detected in all the lactis ssp. lactis
strains (20 strains including IL1403); It exhibits a maximal
intratype divergence of 4.5%. In total the 20 predicted
proteins corresponding to this lactis-allelic type presented
eight positions with substitution and one with an insertion
(Fig. 2). However compared to the ComX of IL1403, the
most divergent proteins (strains NCDO2091 and A7) only
differed by four amino-acids: three substitutions (N38S,
D114E, Q129E) and one insertion (D40) for NCDO2091 and
four substitutions for A7 (N38S, L108S, Q129E, D145E). In the
other strains presenting this comX allelic type, the deduced
proteins only differed by one (six strains), two (four strains)
or three (two strains) amino acids from the IL1403 ComX.
The second allelic type was identified in the eleven L.
lactis ssp. cremoris strains (Fig. 1b); it has a maximal
intratype divergence of 1.4%. Consequently, the 11 pre-
dicted proteins corresponding to this allelic type are very
well conserved: only three positions with amino-acid sub-
stitutions (H54R, R67W and R130C) are observed (Fig. 2).
Comparison of the predicted proteins from the two comX
allelic types (represented by the sequences from IL1403 and
MG1363), revealed two clusters of amino-acid substitutions
(Fig. 2 grey boxes) in regions of the sigma factor which are
embedded in the functional domains 2.3 and 3.1 (Lonetto
et al., 1992; Wosten, 1998; Gruber & Gross, 2003). Since
both domains are proposed to be involved in the interaction
with DNA (opening of the DNA duplex and DNA binding,
respectively), we wondered whether this variability may be
correlated to any divergence of the sigma recognition site in
the promoters. We therefore determined the sequence of the
putative cin-box in these strains.
Sequence of the cin-box in various lactococcalstrains
In agreement with Bolotin et al. (2001), MEME (Bailey &
Elkan, 1994) generated a consensus sequence (GTTACAA-
TN9TTTTCGTATA, Fig. 3) from the upstream region of the
late competence operons comE, comF, comC, comG, several
competence associated genes dprA, coiA, radA, recQ, ssbA
and a gene of unknown function yqfG in IL1403. This
sequence is proposed as the cin-box required for ComX
recognition. Using PATSCAN (Dsouza et al., 1997) to search
the IL1403 genome, a similar sequence was also identified
upstream of the radC, recA, ydbC (encoding a conserved
hypothetical protein, COG 4443) and ywgA (encoding a
IL1403 MKPIVMKLMKQIRIRTWKIEDYLQEGMIILHLLLEEQNDGQKLHTKFKVKYHQRLIDELRRSYAKKRSHDHFIG
lactis-T -----------------E-------------------S--H--------------------------N------1 13 1 1
MG1363 ----IR--------KA-D----Y--------H----NHPSTNIY---------H------H------L----V-
cremoris-T -----------------------------------------------------R------------W-------4 1
IL1403 LDVYECSDWINSGDTSPDNEVVFNHLLAEVYEGLSAHYQDLLLRQMRGEELTRMQRYRLREKIKAILFSEDEE
lactis-T ---------------------------------S----E/N-------------E---------------E--1 1 1 7 1
MG1363 --I-------DA-GST-ES-L------------------E--V-----------E---------N----R-DD
cremoris-T -------------------------------------------------------C-----------------2
2.1 2.2 2.3 2.4
4.24.13.1
D
Fig. 2. Amino-acid sequences of ComX. The black lines shown above the protein sequences indicate the putative functional domains (2.1, 2.2, 2.3,
2.4, 3.1, 4.1 and 4.2) as defined for other sigma factors (Lonetto et al., 1992). The first protein sequence corresponds to the ComX protein of the IL1403
strain. The lactis-T line shows all the amino-acid differences observed in the lactis-allelic type. The third sequence indicates the differences in the
MG1363 ComX protein in comparison with the IL1403 protein; most of the substitutions indicated were found in all of the ssp. cremoris strains. The
cremoris-T line shows amino-acid differences (other than that indicated for the MG1363 ComX protein) observed in the cremoris-allelic type. The
numbers below the substitutions correspond to the number of strains in which the difference was observed. NCDO2091 is the strain exhibiting the aa
(D) insertion in position 40 of ComX.
FEMS Microbiol Lett 257 (2006) 32–42c� 2006 Federation of European Microbiological SocietiesNo claim to original French government works
38 S. Wydau et al.
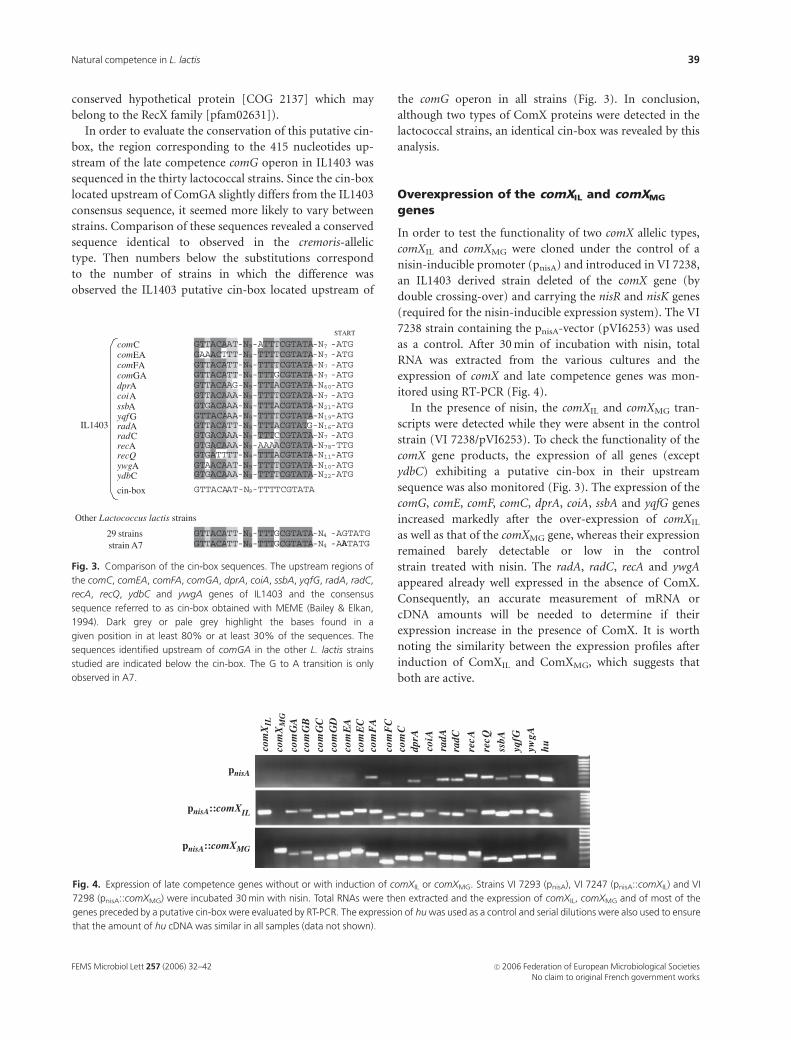
conserved hypothetical protein [COG 2137] which may
belong to the RecX family [pfam02631]).
In order to evaluate the conservation of this putative cin-
box, the region corresponding to the 415 nucleotides up-
stream of the late competence comG operon in IL1403 was
sequenced in the thirty lactococcal strains. Since the cin-box
located upstream of ComGA slightly differs from the IL1403
consensus sequence, it seemed more likely to vary between
strains. Comparison of these sequences revealed a conserved
sequence identical to observed in the cremoris-allelic
type. Then numbers below the substitutions correspond
to the number of strains in which the difference was
observed the IL1403 putative cin-box located upstream of
the comG operon in all strains (Fig. 3). In conclusion,
although two types of ComX proteins were detected in the
lactococcal strains, an identical cin-box was revealed by this
analysis.
Overexpression of the comXIL and comXMG
genes
In order to test the functionality of two comX allelic types,
comXIL and comXMG were cloned under the control of a
nisin-inducible promoter (pnisA) and introduced in VI 7238,
an IL1403 derived strain deleted of the comX gene (by
double crossing-over) and carrying the nisR and nisK genes
(required for the nisin-inducible expression system). The VI
7238 strain containing the pnisA-vector (pVI6253) was used
as a control. After 30 min of incubation with nisin, total
RNA was extracted from the various cultures and the
expression of comX and late competence genes was mon-
itored using RT-PCR (Fig. 4).
In the presence of nisin, the comXIL and comXMG tran-
scripts were detected while they were absent in the control
strain (VI 7238/pVI6253). To check the functionality of the
comX gene products, the expression of all genes (except
ydbC) exhibiting a putative cin-box in their upstream
sequence was also monitored (Fig. 3). The expression of the
comG, comE, comF, comC, dprA, coiA, ssbA and yqfG genes
increased markedly after the over-expression of comXIL
as well as that of the comXMG gene, whereas their expression
remained barely detectable or low in the control
strain treated with nisin. The radA, radC, recA and ywgA
appeared already well expressed in the absence of ComX.
Consequently, an accurate measurement of mRNA or
cDNA amounts will be needed to determine if their
expression increase in the presence of ComX. It is worth
noting the similarity between the expression profiles after
induction of ComXIL and ComXMG, which suggests that
both are active.
pnisA
pnisA::comXIL
pnisA::comXMG
com
XM
G
com
XIL
com
GA
com
GB
com
GC
com
GD
com
EA
com
EC
com
FA
com
FC
com
Cdp
rAco
iAra
dAra
dCre
cAre
cQss
bAyq
fGyw
gAhu
Fig. 4. Expression of late competence genes without or with induction of comXIL or comXMG. Strains VI 7293 (pnisA), VI 7247 (pnisA::comXIL) and VI
7298 (pnisA::comXMG) were incubated 30 min with nisin. Total RNAs were then extracted and the expression of comXIL, comXMG and of most of the
genes preceded by a putative cin-box were evaluated by RT-PCR. The expression of hu was used as a control and serial dilutions were also used to ensure
that the amount of hu cDNA was similar in all samples (data not shown).
-NGTTACAAT-N -ATTTCGTATA -ATGGAAACTTT-N -TTTTCGTATA-N -ATGGTTACATT-N -TTTTCGTATA-N -ATGGTTACATT-N -TTTGCGTATA-N -ATG
GTTACAAA-N -TTTTCGTATA-NGTTACAAG-N -TTTACGTATA-N -ATG
comCcomEAcomFAcomGA
coiAdprA
GTGACAAA-N -TTTACGTATA-N -ATGssbA
GTGACAAA-N -TTTCCGTATA-N -ATGradCGTGACAAA-N -AAAACGTATA-N -TTGrecA
GTTACATT-N -TTTGCGTATA-N -AGTATGGTTACATT-N -TTTGCGTATA-N -AATATGstrain A7
Other Lactococcus lactis strains
29 strains
ydb GTGACAAA-N -TTTTCGTATA-N -ATGCGTAACAAT-N -TTTTCGTATA-N -ATGywgA
GTTACAAT-N -TTTTCGTATAcin-box
IL1403
-
GTGATTTT-N -TTTACGTATA-NrecQ -ATG
GTTACATT-N -TTTACGTATG-NradA -ATGGTTACAAA-N -TTTTCGTATA-N -ATGyqfG
ATG
Fig. 3. Comparison of the cin-box sequences. The upstream regions of
the comC, comEA, comFA, comGA, dprA, coiA, ssbA, yqfG, radA, radC,
recA, recQ, ydbC and ywgA genes of IL1403 and the consensus
sequence referred to as cin-box obtained with MEME (Bailey & Elkan,
1994). Dark grey or pale grey highlight the bases found in a
given position in at least 80% or at least 30% of the sequences. The
sequences identified upstream of comGA in the other L. lactis strains
studied are indicated below the cin-box. The G to A transition is only
observed in A7.
FEMS Microbiol Lett 257 (2006) 32–42 c� 2006 Federation of European Microbiological SocietiesNo claim to original French government works
39Natural competence in L. lactis

Discussion
Comparison of 16S analysis, dprA and comXsequences
The classification of the lactis strains used in this study on
the basis of their 16S rRNA revealed that more than 29% of
the selected strains had been misclassified on the basis of the
phenotypic tests classically used to distinguish the L. lactis
ssp. The comX and dprA sequences each revealed two allelic
types, which perfectly matched the 16S classification of
strains while only partially fitting the phenotypic or RAPD
groups. The mean nucleic divergence observed between the
lactis and cremoris alleles was 26.9% for comX and 20.9% for
dprA. This observation strongly suggests that the natural
competence genes were present in lactococci before the
divergence between the lactis and cremoris ssp., i.e. around
17 million years ago (Bolotin et al., 2004) since the rate
of divergence was estimated at �0.9% per million years
(Ochman et al., 1999).
Conservation of key elements of the naturalcompetence system
In all strains, intact comX genes were found and identical
cin-boxes were identified upstream of the comG operon.
Two types of ComX were distinguished, each being well
conserved among the strains of one ssp. The observation
that the same cin-box is conserved among all tested strains
suggests that these two types of ComX recognize the same
DNA sequence. We showed that in IL1403, both ComX
variants were active and allowed the expression of all the late
competence genes required for DNA uptake in other micro-
organisms. It establishes that comX is not degenerated in L.
lactis, suggesting that having an active ComX might be an
advantage for the strain for an as yet undiscovered reason.
Contrary to comX, dprA was inactivated in about 32% of the
strains all belonging to the ssp. lactis. Although the exact role
of DprA remains to be established in L. lactis, this high
occurrence of a mutated dprA gene may indicate that the
natural competence process is used for another purpose
than genetic diversity (i.e. natural transformation) such as a
competitive advantage as previously shown in Escherichia
coli (Finkel & Kolter, 2001). In a dprA background, the
natural competence could provide DNA, which will be
degraded leading to an increase of the intracellular nucleo-
tide pool. As free bases are present in limited amounts in
milk, one can propose that a dprA mutation might be
advantageous. In addition to DNA uptake, the natural
induction of competence may also allow the predation of
non-competent cells or of other species via the expression of
bacteriocin as being recently described for Streptococcus
pneumoniae (Guiral et al., 2005) and S. mutans (Kreth
et al., 2005).
For both the lactis and cremoris ssp., strains presenting a
complete dprA gene were also found. The L. lactis strains
presenting the full DprA (lactis-, cremoris-types) or a
truncated form would be useful to test whether natural
competence is functional in L. lactis and to investigate the
role of this protein in lactococci. We demonstrated in this
work that ComXMG (present in the cremoris ssp.) can
stimulate the expression of the late competence genes in a
ssp. lactis strain. Consequently a comX overexpression
system could be introduced in the various strains exhibiting
a complete or a truncated dprA gene (identified in this
work) to test whether the upregulation of comX could lead
to natural transformation. This system constitutes an inter-
esting alternative to the laborious approach that is to seek
conditions allowing the induction of competence in a given
species by testing different media, starvation conditions,
growth phases etc. This latter approach may be time
consuming since the conditions of induction of natural
competence vary between species (Dubnau, 1991).
In conclusion, this study revealed that the homologues of
three key elements of the natural competence process are
conserved among lactococci and it demonstrates for the first
time in lactococci the activity of the ComX–cin-box regula-
tory circuit. Strains with different allelic types of the comX
and dprA genes and the comX overexpression system con-
stitute promising tools to test the functionality of natural
transformation in this species.
Acknowledgements
We are grateful to D. Vailhen, A. Bolotin, M. van de Guchte,
P. Serror for helpful discussion and suggestions. S. Wydau is
the recipient of a fellowship from the Ministere de la
Recherche.
References
Ando T, Israel DA, Kusugami K & Blaser MJ (1999) HP0333, a
member of the dprA family, is involved in natural
transformation in Helicobacter pylori. J Bacteriol 181:
5572–5580.
Bailey TL & Elkan C (1994) Fitting a mixture model by
expectation maximization to discover motifs in biopolymers.
Proceedings of the Second International Conference on Intelligent
Systems for Molecular Biology 2: 28–36. AAAI Press, Menlo
Park, CA.
Berge M, Mortier-Barriere I, Martin B & Claverys JP (2003)
Transformation of Streptococcus pneumoniae relies on
DprA- and RecA-dependent protection of incoming DNA
single strands. Mol Microbiol 50: 527–536.
Berka RM, Hahn J, Albano M, Draskovic I, Persuh M, Cui X,
Sloma A, Widner W & Dubnau D (2002) Microarray analysis
FEMS Microbiol Lett 257 (2006) 32–42c� 2006 Federation of European Microbiological SocietiesNo claim to original French government works
40 S. Wydau et al.

of the Bacillus subtilis K-state: genome-wide expression
changes dependent on ComK. Mol Microbiol 43: 1331–1345.
Biswas I, Gruss A, Ehrlich SD & Maguin E (1993) High-efficiency
gene inactivation and replacement system for gram-positive
bacteria. J Bacteriol 175: 3628–3635.
Bolotin A, Mauger S, Malarme K, Ehrlich SD & Sorokin A (1999)
Low-redundancy sequencing of the entire Lactococcus lactis
IL1403 genome. Antonie Van Leeuwenhoek 76: 27–76.
Bolotin A, Wincker P, Mauger S, Jaillon O, Malarme K,
Weissenbach J, Ehrlich SD & Sorokin A (2001) The complete
genome sequence of the lactic acid bacterium Lactococcus lactis
ssp. lactis IL1403. Genome Res 11: 731–753.
Bolotin A, Quinquis B, Sorokin A & Ehrlich DS (2004) Recent
genetic transfer between Lactococcus lactis and enterobacteria.
J Bacteriol 186: 6671–6677.
Claverys JP & Havarstein LS (2002) Extracellular-peptide control
of competence for genetic transformation in Streptococcus
pneumoniae. Front Biosci 7: d1798–d1814.
Claverys JP & Martin B (2003) Bacterial ‘‘competence’’ genes:
signatures of active transformation, or only remnants? Trends
Microbiol 11: 161–165.
Cvitkovitch DG (2001) Genetic competence and transformation
in oral streptococci. Crit Rev Oral Biol Med 12: 217–243.
Dreiseikelmann B (1994) Translocation of DNA across bacterial
membranes. Microbiol Rev 58: 293–316.
Dsouza M, Larsen N & Overbeek R (1997) Searching for patterns
in genomic data. Trends Genet 13: 497–498.
Dubnau D (1991) The regulation of genetic competence in
Bacillus subtilis. Mol Microbiol 5: 11–18.
van den Eede G, Aarts H, Buhk HJ, et al. (2004) The relevance of
gene transfer to the safety of food and feed derived from
genetically modified (GM) plants. Food Chem Toxicol 42:
1127–1156.
Finkel SE & Kolter R (2001) DNA as a nutrient: novel role for
bacterial competence gene homologs. J Bacteriol 183:
6288–6293.
Friedrich A, Prust C, Hartsch T, Henne A & Averhoff B (2002)
Molecular analyses of the natural transformation machinery
and identification of pilus structures in the extremely
thermophilic bacterium Thermus thermophilus strain HB27.
Appl Environ Microbiol 68: 745–755.
Gasson MJ (2000) Gene transfer from genetically modified food.
Curr Opin Biotechnol 11: 505–508.
Gruber TM & Gross CA (2003) Multiple sigma subunits and the
partitioning of bacterial transcription space. Annu Rev
Microbiol 57: 441–466.
Guiral S, Mitchell TJ, Martin B & Claverys JP (2005)
Competence-programmed predation of noncompetent cells in
the human pathogen Streptococcus pneumoniae: genetic
requirements. Proc Natl Acad Sci USA 102: 8710–8715.
Havarstein LS, Hakenbeck R & Gaustad P (1997) Natural
competence in the genus Streptococcus: evidence that
streptococci can change pherotype by interspecies
recombinational exchanges. J Bacteriol 179: 6589–6594.
Jensen PR & Hammer K (1993) Minimal requirements for
exponential growth of Lactococcus lactis. Appl Environ
Microbiol 59: 4363–4366.
Jonas DA, Elmadfa I, Engel KH, Heller KJ, Kozianowski G, Konig
A, Muller D, Narbonne JF, Wackernagel W & Kleiner J (2001)
Safety considerations of DNA in food. Ann Nutr Metab 45:
235–254.
Karudapuram S, Zhao X & Barcak GJ (1995) DNA sequence and
characterization of Haemophilus influenzae dprA1, a gene
required for chromosomal but not plasmid DNA
transformation. J Bacteriol 177: 3235–3240.
Kharazmi M, Hammes WP & Hertel C (2002) Construction of a
marker rescue system in Bacillus subtilis for detection of
horizontal gene transfer in food. Syst Appl Microbiol 25:
471–477.
Kreth J, Merritt J, Shi W & Qi F (2005) Co-ordinated bacteriocin
production and competence development: a possible
mechanism for taking up DNA from neighbouring species.
Mol Microbiol 57: 392–404.
Kumar S, Tamura K, Jakobsen IB & Nei M (2001) MEGA2:
molecular evolutionary genetics analysis software.
Bioinformatics 17: 1244–1245.
Lee MS & Morrison DA (1999) Identification of a new regulator
in Streptococcus pneumoniae linking quorum sensing to
competence for genetic transformation. J Bacteriol 181:
5004–5016.
Lonetto M, Gribskov M & Gross CA (1992) The sigma 70 family:
sequence conservation and evolutionary relationships.
J Bacteriol 174: 3843–3849.
Lorenz MG & Wackernagel W (1994) Bacterial gene transfer by
natural genetic transformation in the environment. Microbiol
Rev 58: 563–602.
Maguin E, Prevost H, Ehrlich SD & Gruss A (1996) Efficient
insertional mutagenesis in lactococci and other gram-positive
bacteria. J Bacteriol 178: 931–935.
Mortier-Barriere I, de Saizieu A, Claverys JP & Martin B
(1998) Competence-specific induction of recA is required
for full recombination proficiency during transformation
in Streptococcus pneumoniae. Mol Microbiol 27:
159–170.
Ochman H, Elwyn S & Moran NA (1999) Calibrating bacterial
evolution. Proc Natl Acad Sci USA 96: 12638–12643.
Ogura M, Yamaguchi H, Kobayashi K, Ogasawara N, Fujita Y &
Tanaka T (2002) Whole-genome analysis of genes regulated by
the Bacillus subtilis competence transcription factor ComK.
J Bacteriol 184: 2344–2351.
Opdyke JA, Scott JR & Moran CP Jr. (2001) A secondary RNA
polymerase sigma factor from Streptococcus pyogenes. Mol
Microbiol 42: 495–502.
Page RD (1996) TreeView: an application to display phylogenetic
trees on personal computers. Comput Appl Biosci 12: 357–358.
Pestova EV, Havarstein LS & Morrison DA (1996) Regulation of
competence for genetic transformation in Streptococcus
pneumoniae by an auto-induced peptide pheromone and a
two-component regulatory system. Mol Microbiol 21: 853–862.
FEMS Microbiol Lett 257 (2006) 32–42 c� 2006 Federation of European Microbiological SocietiesNo claim to original French government works
41Natural competence in L. lactis

Redon E, Loubiere P & Cocaign-Bousquet M (2005)
Transcriptome analysis of the progressive adaptation of
Lactococcus lactis to carbon starvation. J Bacteriol 187:
3589–3592.
Renault P, Corthier G, Goupil N, Delorme C & Ehrlich SD (1996)
Plasmid vectors for gram-positive bacteria switching from
high to low copy number. Gene 183: 175–182.
Sambrook J, Fritsch EF & Maniatis T (1989) Molecular Cloning:
a Laboratory Manual, 2nd edn. Cold Spring Harbor,
New York.
Smeets LC, Bijlsma JJ, Kuipers EJ, Vandenbroucke-Grauls CM &
Kusters JG (2000) The dprA gene is required for natural
transformation of Helicobacter pylori. FEMS Immunol Med
Microbiol 27: 99–102.
Stiles ME & Holzapfel WH (1997) Lactic acid bacteria of
foods and their current taxonomy. Int J Food Microbiol 36:
1–29.
Tailliez P, Tremblay J, Ehrlich SD & Chopin A (1998) Molecular
diversity and relationship within Lactococcus lactis, as revealed
by randomly amplified polymorphic DNA (RAPD). Syst Appl
Microbiol 21: 530–538.
Terzaghi BE & Sandine WE (1975) Inproved medium for lactic
streptococci and their bacteriophages. Appl Microbiol 29:
807–813.
Thompson JD, Higgins DG & Gibson TJ (1994) CLUSTAL W:
improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, position-specific gap
penalties and weight matrix choice. Nucleic Acids Res 22:
4673–4680.
Ward LJ, Brown JC & Davey GP (1998) Two methods for the
genetic differentiation of Lactococcus lactis ssp. lactis and
cremoris based on differences in the 16S rRNA gene sequence.
FEMS Microbiol Lett 166: 15–20.
Wosten MM (1998) Eubacterial sigma-factors. FEMS Microbiol
Rev 22: 127–150.
FEMS Microbiol Lett 257 (2006) 32–42c� 2006 Federation of European Microbiological SocietiesNo claim to original French government works
42 S. Wydau et al.




















