
Spoligotype-based Comparative Population Structure of Multidrug Resistant and Isoniazid...
11
JOURNAL OF CLINICAL MICROBIOLOGY, Nov. 2010, p. 3899–3909 Vol. 48, No. 11 0095-1137/10/$12.00 doi:10.1128/JCM.00572-10 Copyright © 2010, American Society for Microbiology. All Rights Reserved. Spoligotype-Based Comparative Population Structure Analysis of Multidrug-Resistant and Isoniazid-Monoresistant Mycobacterium tuberculosis Complex Clinical Isolates in Poland Tomasz Jagielski, 1 Ewa Augustynowicz-Kopec ´, 1 Thierry Zozio, 2 Nalin Rastogi, 2 * and Zofia Zwolska 1 Department of Microbiology, National Tuberculosis and Lung Diseases Research Institute, Warsaw, Poland, 1 and WHO Supranational TB Reference Laboratory, Unite ´ de la Tuberculose et des Mycobacte ´ries, Institut Pasteur de Guadeloupe, Abymes Cedex, Guadeloupe, France 2 Received 17 March 2010/Returned for modification 8 June 2010/Accepted 24 August 2010 The spoligotyping-based population structure of multidrug-resistant (MDR) Mycobacterium tuberculosis strains isolated in Poland (n 46), representing all culture-positive MDR tuberculosis (MDR-TB) cases, was compared to that of isoniazid (INH)-monoresistant strains (n 71) isolated in 2004. The latter data set from a previous study (E. Augustynowicz-Kopec ´, T. Jagielski, and Z. Zwolska, J. Clin. Microbiol. 2008, 46:4041– 4044) represented 87% of all INH-monoresistant strains. The clustering rates and genotypic-diversity indexes for the 2 subpopulations were not significantly different (P 0.05). The results were entered in the SITVIT2 database to assign specific shared type designations, corresponding genotypic lineages, and geographical distributions and compared to available data from neighboring countries (Germany, n 704; Czech Republic, n 530; Sweden, n 379; Kaliningrad, Russia, n 90) and strains from previous studies in Poland (n 317). MDR strains resulted in 27 patterns (20 unique strains within the study and 7 clusters containing 2 to 6 isolates per cluster with a clustering rate of 56.5%) and belonged to the following genotypic lineages: ill-defined T family (28.3%), Haarlem (17.4%), Latin American and Mediterranean (LAM) (13%), Beijing (8.7%), S family (4.35%), and the X clade (2.17%). Comparison of the genetic structure of the MDR strains with that of INH-monoresistant strains showed that a total of 9 patterns were shared by both groups; these represented 1/3 of the MDR strains and 2/3 of the INH-monoresistant strains. Interestingly, 76.1% of the MDR isolates and 71.8% of the INH-resistant isolates yielded spoligotypes that were previously reported from Poland. The observation that nearly half of the spoligotypes identified among both MDR (48.1%) and INH-monoresistant (43.3%) M. tuberculosis isolates were present in Poland’s neighboring countries suggested that a significant proportion of MDR and INH-resistant TB cases in Poland were caused by strains actively circulating in Poland or its neighbors. Our results corroborate the leading role of the T and Haarlem genotypes in the epidemiology of drug-resistant TB in Poland. Nevertheless, the LAM and Beijing family strains that infected, correspond- ingly, 13% and 9% of patients with MDR-TB were absent among the strains from patients with INH- monoresistant TB, suggesting that a proportion of MDR-TB cases in Poland are due to ongoing transmission of MDR clones exhibiting specific genotypes. Study of the population genetic relationships between MDR and INH-monoresistant strains by drawing minimum spanning trees showed that ill-defined T1 sublineage strains (1/3 of all INH-monoresistant strains), represented by its prototype, SIT53, constituted the central node of the tree, followed by strains belonging to the well-defined H3, H1, and S subgroups. However, the MDR group, in addition, contained LAM (n 6) and Beijing (n 4) lineage isolates. With the exception of the 4 Beijing lineage strains in the latter group and a single orphan isolate in the INH-monoresistant group, none of the remaining 112/117 isolates belonged to principal genetic group 1 (PGG1) in our study. Given the high rate of clustering and the near absence of immigrants in the study, the persistence of MDR-TB in Poland seems to result from active transmission of MDR strains within the autochthonous population, the bulk of it caused by evolutionarily recent tubercle bacilli. Tuberculosis (TB) remains a substantial global health prob- lem. With more than 9 million new cases and almost 2 million deaths every year (50), TB holds one of the leading positions among infectious causes of morbidity and mortality worldwide. One of the reasons for the continuing TB prevalence is the emergence and spread of drug-resistant (DR) and, especially, multidrug-resistant (MDR) (i.e., resistant to at least isoni- azid [INH] and rifampin [RMP]) Mycobacterium tuberculosis strains. According to a recent World Health Organization (WHO) report, the global population weighted percentages for any resistance and MDR among all TB cases are 20% and 5.3%, respectively (49). Poland, the largest and most populous country in Central Europe (38,530,000 inhabitants in 2002), has witnessed a dramatic fall in the incidence of TB over the last few decades. While in 1957 the notification rate for all clinical forms was 290.4 per 100,000 population, it had dropped almost 10-fold by the end of 1990s (43). However, the TB notification rate in Poland is still relatively high, nearly twice * Corresponding author. Mailing address: Institut Pasteur de Guade- loupe, Morne Jolivie `re, BP484, F97183 Abymes Cedex, Guadeloupe. Phone: 590-590-897661. Fax: 590-590-896941. E-mail: nrastogi@pasteur -guadeloupe.fr. Published ahead of print on 1 September 2010. 3899
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Spoligotype-based Comparative Population Structure of Multidrug Resistant and Isoniazid...

JOURNAL OF CLINICAL MICROBIOLOGY, Nov. 2010, p. 3899–3909 Vol. 48, No. 110095-1137/10/$12.00 doi:10.1128/JCM.00572-10Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Spoligotype-Based Comparative Population Structure Analysis ofMultidrug-Resistant and Isoniazid-Monoresistant
Mycobacterium tuberculosis Complex ClinicalIsolates in Poland�
Tomasz Jagielski,1 Ewa Augustynowicz-Kopec,1 Thierry Zozio,2 Nalin Rastogi,2* and Zofia Zwolska1
Department of Microbiology, National Tuberculosis and Lung Diseases Research Institute, Warsaw, Poland,1
and WHO Supranational TB Reference Laboratory, Unite de la Tuberculose et des Mycobacteries,Institut Pasteur de Guadeloupe, Abymes Cedex, Guadeloupe, France2
Received 17 March 2010/Returned for modification 8 June 2010/Accepted 24 August 2010
The spoligotyping-based population structure of multidrug-resistant (MDR) Mycobacterium tuberculosisstrains isolated in Poland (n � 46), representing all culture-positive MDR tuberculosis (MDR-TB) cases, wascompared to that of isoniazid (INH)-monoresistant strains (n � 71) isolated in 2004. The latter data set froma previous study (E. Augustynowicz-Kopec, T. Jagielski, and Z. Zwolska, J. Clin. Microbiol. 2008, 46:4041–4044) represented 87% of all INH-monoresistant strains. The clustering rates and genotypic-diversity indexesfor the 2 subpopulations were not significantly different (P � 0.05). The results were entered in the SITVIT2database to assign specific shared type designations, corresponding genotypic lineages, and geographicaldistributions and compared to available data from neighboring countries (Germany, n � 704; Czech Republic,n � 530; Sweden, n � 379; Kaliningrad, Russia, n � 90) and strains from previous studies in Poland (n � 317).MDR strains resulted in 27 patterns (20 unique strains within the study and 7 clusters containing 2 to 6isolates per cluster with a clustering rate of 56.5%) and belonged to the following genotypic lineages: ill-definedT family (28.3%), Haarlem (17.4%), Latin American and Mediterranean (LAM) (13%), Beijing (8.7%), S family(4.35%), and the X clade (2.17%). Comparison of the genetic structure of the MDR strains with that ofINH-monoresistant strains showed that a total of 9 patterns were shared by both groups; these represented 1/3of the MDR strains and 2/3 of the INH-monoresistant strains. Interestingly, 76.1% of the MDR isolates and71.8% of the INH-resistant isolates yielded spoligotypes that were previously reported from Poland. Theobservation that nearly half of the spoligotypes identified among both MDR (48.1%) and INH-monoresistant(43.3%) M. tuberculosis isolates were present in Poland’s neighboring countries suggested that a significantproportion of MDR and INH-resistant TB cases in Poland were caused by strains actively circulating in Polandor its neighbors. Our results corroborate the leading role of the T and Haarlem genotypes in the epidemiologyof drug-resistant TB in Poland. Nevertheless, the LAM and Beijing family strains that infected, correspond-ingly, 13% and 9% of patients with MDR-TB were absent among the strains from patients with INH-monoresistant TB, suggesting that a proportion of MDR-TB cases in Poland are due to ongoing transmissionof MDR clones exhibiting specific genotypes. Study of the population genetic relationships between MDR andINH-monoresistant strains by drawing minimum spanning trees showed that ill-defined T1 sublineage strains(1/3 of all INH-monoresistant strains), represented by its prototype, SIT53, constituted the central node of thetree, followed by strains belonging to the well-defined H3, H1, and S subgroups. However, the MDR group, inaddition, contained LAM (n � 6) and Beijing (n � 4) lineage isolates. With the exception of the 4 Beijinglineage strains in the latter group and a single orphan isolate in the INH-monoresistant group, none of theremaining 112/117 isolates belonged to principal genetic group 1 (PGG1) in our study. Given the high rate ofclustering and the near absence of immigrants in the study, the persistence of MDR-TB in Poland seems toresult from active transmission of MDR strains within the autochthonous population, the bulk of it caused byevolutionarily recent tubercle bacilli.
Tuberculosis (TB) remains a substantial global health prob-lem. With more than 9 million new cases and almost 2 milliondeaths every year (50), TB holds one of the leading positionsamong infectious causes of morbidity and mortality worldwide.One of the reasons for the continuing TB prevalence is theemergence and spread of drug-resistant (DR) and, especially,
multidrug-resistant (MDR) (i.e., resistant to at least isoni-azid [INH] and rifampin [RMP]) Mycobacterium tuberculosisstrains. According to a recent World Health Organization(WHO) report, the global population weighted percentages forany resistance and MDR among all TB cases are 20% and5.3%, respectively (49). Poland, the largest and most populouscountry in Central Europe (38,530,000 inhabitants in 2002),has witnessed a dramatic fall in the incidence of TB over thelast few decades. While in 1957 the notification rate for allclinical forms was 290.4 per 100,000 population, it had droppedalmost 10-fold by the end of 1990s (43). However, the TBnotification rate in Poland is still relatively high, nearly twice
* Corresponding author. Mailing address: Institut Pasteur de Guade-loupe, Morne Joliviere, BP484, F97183 Abymes Cedex, Guadeloupe.Phone: 590-590-897661. Fax: 590-590-896941. E-mail: [email protected].
� Published ahead of print on 1 September 2010.
3899

the mean European Union (EU) rate (24.6/100,000 versus12.8/100,000 in 2004) (14).
Since the early years of chemotherapy for TB, the preva-lence of primary drug-resistant TB in Poland has been closelymonitored. In 1994, Poland joined the WHO/International Unionagainst Tuberculosis and Lung Disease (IUATLD) global projecton anti-TB drug resistance surveillance, and during 1996 and1997, the first national survey on primary and acquired drugresistance was performed. The subsequent study, completed in2001, revealed a 2-fold increase in the primary resistance ratein comparison with that in 1997, i.e., 6.1% versus 3.6%, respec-tively. Likewise, the prevalence of primary multidrug resis-tance doubled, from 0.6% in 1997 to 1.2% in 2000 (3). In 2004,there was a slight decrease to 5.6% in the percentage of TBcases with primary drug resistance and quite a significant fall to0.3% in the share of multidrug-resistant tuberculosis (MDR-TB) cases compared to 2000 (49). These figures place Polandamong the countries with low to moderate DR-TB rates in theworld. However, inadequacies in the registration of TB pa-tients may contribute to an underestimation of drug-resistantcases. The most detrimental impact on the epidemiology of TBin Poland might be the spread of DR-TB from outside thecountry. This is because Poland shares borders with countriesof the former Soviet Union, such as Lithuania, Ukraine, andRussia, where the incidence of DR-TB, including MDR-TB, isexceptionally high (49). Given the geographical epidemiologi-cal context, the surveillance capacity for TB in Poland has to beparticularly well established.
Studying genetic relationships among M. tuberculosis strainshas provided important insights into the spread and transmis-sion of TB (8, 15, 26, 30, 31, 34). Spoligotyping targeting thepolymorphism of the chromosomal direct-repeat locus is oneof the most extensively used techniques for genotypic differ-entiation between M. tuberculosis strains (27). Primarily usedfor first-line screening of outbreaks (22, 40), spoligotyping isideally followed by more discriminatory second-line IS6110 re-striction fragment length polymorphism (RFLP) or mycobac-terial interspersed repetitive-unit–variable-number tandem-re-peat (MIRU-VNTR) typing (9, 24). It has also proven usefulfor tracking laboratory cross-contaminations (36) and describ-ing the global spread of TB (9, 17). Major advantages of thespoligotyping technique are its simplicity, rapidity, reproduc-ibility, and portability, due to the digital nature of the gener-ated results. Of particular importance is the existence of theinternational spoligotype database, which provides useful in-formation on the epidemiology and phylogeny of the circulat-ing genotypes of tubercle bacilli worldwide (9).
This study aimed to investigate the spoligotyping-based pop-ulation structure of MDR M. tuberculosis clinical isolates iso-lated in Poland in 2004 compared to that of INH-monoresis-tant strains isolated in the same year. Indeed, as reviewed byDye and Espinal (12), a high proportion of TB patients carry-ing INH-monoresistant strains acquire RMP resistance aftertreatment failure, suggesting that the development of INHresistance usually precedes that to RMP.
MATERIALS AND METHODS
Patients and specimens. This study included a total of 46 MDR strains of M.tuberculosis isolated from as many TB patients in Poland. The patients (40 menand 6 women; age range, 31 to 79 years; median age, 50.5 years) were recruited
in several TB clinics across Poland during a 1-year period from 1 January 2004to 31 December 2004 (Fig. 1). Primary isolation, species identification, and drugsusceptibility testing (DST) were done at regional mycobacteriology laboratories.The isolates were subcultured and sent to the National TB Reference Laboratory(NTRL) at the National Tuberculosis and Lung Diseases Research Institute inWarsaw, where confirmatory identification and DST were performed. Testing ofsusceptibility to the first-line drugs was carried out according to the 1% propor-tion method in Lowenstein-Jensen medium, with the following critical concen-trations: streptomycin (SM), 4 �g/ml; INH, 0.2 �g/ml; RMP, 40 �g/ml; andethambutol (EMB), 2 �g/ml. Basic demographic data were collected for eachpatient, using a standard questionnaire prepared by the NTRL, and sent to thelocal TB laboratories.
Spoligotyping and database comparison. DNAs extracted by the cetyltrimeth-ylammonium bromide (CTAB) method (47) were stored at �20°C before beingsubjected to spoligotyping using a previously described protocol (27). Briefly, thedirect-repeat region of the TB genome was amplified using primers DRa andDRb, and the amplified biotinylated products were hybridized to a set of 43oligonucleotides covalently bound to a membrane (Isogen Life Science B.V.,Utrecht, Netherlands). The hybridized PCR products were incubated withstreptavidin-peroxidase conjugate, and signal detection was obtained with anenhanced chemiluminescence detection system (Amersham, Little Chalfont, En-gland), followed by exposure to X-ray film (Hyperfilm ECL; Amersham) accord-ing to the manufacturer’s instructions. The X-ray film was developed usingstandard photochemical procedures after overnight exposure. Spoligotypes inbinary format were converted to an octal code for comparison with the SITVIT2proprietary database of the Pasteur Institute of Guadeloupe, which is anupdated version of the previously released SpolDB4 database (9; availableonline at http://www.pasteur-guadeloupe.fr:8081/SITVITDemo). At the timeof the present study, SITVIT2 contained genotyping information on 70,000 M.tuberculosis clinical isolates from 160 countries of origin. In the database, SIT(spoligotype international type) designates a spoligotype shared by 2 or morepatient isolates, as opposed to “orphan,” which designates patterns reported fora single isolate. Major phylogenetic clades were assigned according to signaturesprovided in SpolDB4, which defined 62 genetic lineages/sublineages (9). Theyinclude specific signatures for various M. tuberculosis complex species, as well asmajor lineages/sublineages for M. tuberculosis sensu stricto, e.g., Central Asian(CAS), East African-Indian (EAI), Haarlem (H), Latin American-Mediterra-nean (LAM), Manu, S clade, the IS6110 low-banding X clade, and an ill-definedT clade.
Phylogenetic and statistical analyses. Correlation of spoligotype families andprincipal genetic groups (PGG) based on KatG463-gyrA95 polymorphisms wasinferred from the reported linking of specific spoligotype patterns to PGG1(ancient lineages) and PGG2/3 (modern lineages) (9, 42). The evolutionaryrelationships among the spoligotype patterns obtained were further studied bydrawing a minimum spanning tree (MST) with Bionumerics software (version3.5; Applied Maths, Sint-Martens-Latem, Belgium). The MSTs were drawn forthe INH-monoresistant (n � 71) and MDR (n � 46) populations independently,as well as a combined tree for all the isolates taken together (n � 117). An MSTis an undirected network in which all the samples are linked together with thesmallest possible linkages between nearest neighbors.
The chi-square test or Fisher’s exact test (if the cell count was less than 5) wasperformed to evaluate differences in demographic and epidemiologic character-istics between MDR-TB and INH-monoresistant-TB patients. Differences be-tween the genotypic-diversity indexes were assessed by the two-proportion test.A P value that was less than 0.05 was considered statistically significant.
RESULTS AND DISCUSSION
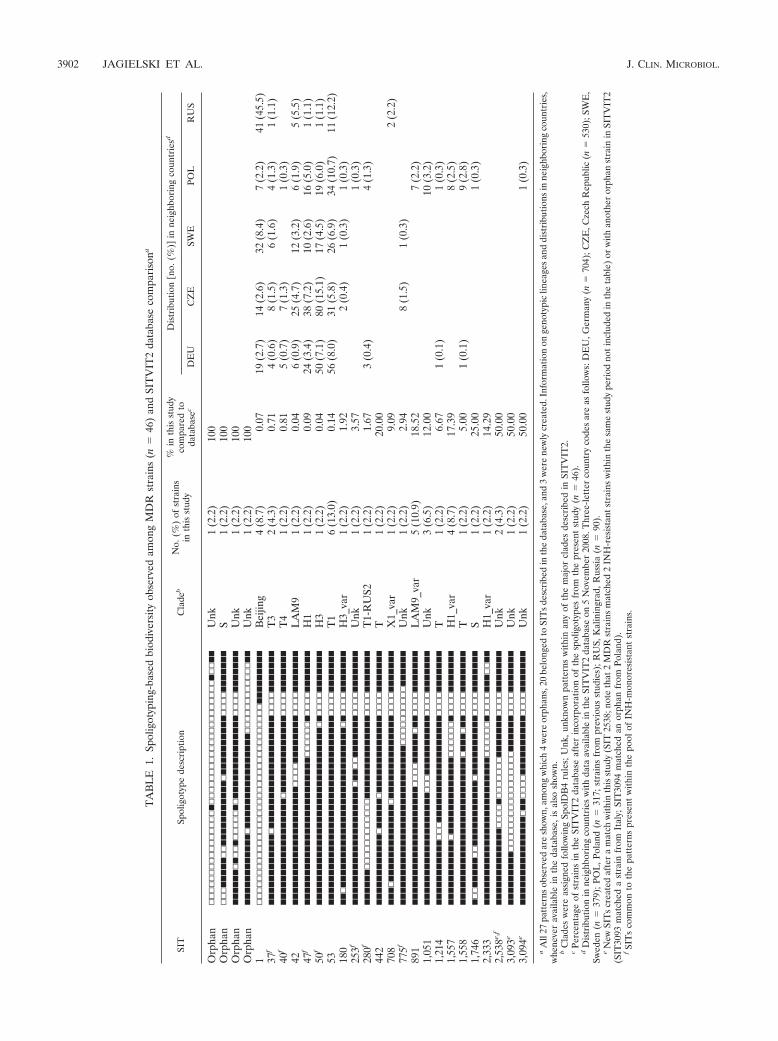
This study included all bacteriologically confirmed MDR-TBcases in Poland over a 1-year period (2004; n � 46 isolatesfrom as many TB patients). These MDR-TB cases represented18.7% of all drug-resistant-TB cases reported in Polandthroughout 2004 (smear-positive TB cases registered [total],n � 9,493; any drug resistance, n � 246; INH-monoresistantisolates, n � 82; MDR-TB isolates, n � 46). The results ob-tained by spoligotyping of the MDR-TB isolates compared tothe INH-monoresistant isolates (n � 71) are summarized inTables 1 and 2. Whenever available, a phylogenetic clade de-scription for each SIT was also provided, as well as their geo-graphical distribution among neighboring countries, limited to
3900 JAGIELSKI ET AL. J. CLIN. MICROBIOL.

Germany (n � 704), Czech Republic (n � 530), Sweden (n �379), and Kaliningrad, Russia (n � 90), as well as Poland(strains from previous studies; n � 317). For the 46 clinicalisolates, spoligotyping produced a total of 27 different patterns(genetic diversity, 58.7%), 4 of which corresponded to orphanpatterns (not found among more than 70,000 strains recordedin the SITVIT2 database). The remaining 42 (91.3%) strainsfrom this study belonged to 23 SITs, including the 3 newlycreated SITs. The latter were new SITs created after either amatch within this study, as in the case of SIT2538, comprising2 MDR isolates, or a match with another orphan strain re-corded in SpolDB4 (SIT3093 matched a strain from Italy,whereas SIT3094 matched a strain from previous studies inPoland). Regarding clustering, a total of 26 (56.5%) clinicalisolates were clustered (7 clusters containing 2 to 6 isolates percluster), and 20 (43.5%) isolates were unique within this study(including the 4 orphan strains).
The clade assignment demonstrated that the highest numberof MDR strains belonged to the ill-defined T family (13 strains,or 28.3% of all strains). The most prevalent shared spoligotypewithin this family was SIT53, representing the T1 sublineage (6strains, or 13% of all strains and 46% of the T-clade strains).The second-largest group, comprising 8 (17.4%) of the ana-lyzed strains, displayed spoligotypes related to those of theHaarlem family, most of them being representative of the H1sublineage (SIT47, SIT1557, and SIT2333). The third-largestgroup comprised members of the LAM family (6 strains, or
13% of all the strains). All these strains displayed spoligotypesthat belonged to the LAM9 subclade (SIT42 and SIT891).Four (8.7%) of the MDR isolates harbored the spoligotypingpattern (SIT1) characteristic of the Beijing family of strains.Two minor clades, namely, X and S, were represented by oneand two isolates, respectively. An important proportion of thestrains (12 strains, or 26.1% of all the strains) could not beassigned to any of the major clades described within the spo-ligotype database. A comparison with INH-monoresistantstrains (n � 71) (Table 2) showed a lower clustering rate (butnot significantly lower; 56.5% versus 70.4%) (Table 3) for theMDR strains, considered a proxy for recent transmission. Theresults also underlined a higher genotypic diversity (58.7%versus 42.2%) and a higher number of unique profiles (43.5%versus 29.6%) of the MDR isolates compared to the INH-monoresistant isolates (4).
Although spoligotyping per se is not an appropriate methodfor assessing the burden of recently transmitted TB (it mayoverestimate the proportion of clustered cases [22–24]), itserves as an efficient preliminary procedure in determining thegenetic relatedness of M. tuberculosis isolates (8, 9, 15, 26, 30,31, 34). Furthermore, the interpretation of the clustering re-sults in the context of recent transmission is fraught with sev-eral ambiguities. This is because the amount of transmissionrepresented by molecular clustering depends on a number ofmethodological factors, such as the methodology employed, asmentioned above; sample size; or study duration. For the last
FIG. 1. Map of Poland depicting the distribution of MDR/INH-monoresistant M. tuberculosis isolates within the 16 provinces of the country.The shaded provinces are the ones from which at least one MDR-TB strain was obtained (administrative map of Poland drawn by the authors;note that boundaries in the map do not attempt to portray exact frontiers between administrative regions in Poland or between Poland and itsneighbors).
VOL. 48, 2010 DRUG-RESISTANT TB IN POLAND 3901
TA
BL
E1.
Spol
igot
ypin
g-ba
sed
biod
iver
sity
obse
rved
amon
gM
DR
stra
ins
(n�
46)
and
SIT
VIT
2da
taba
seco
mpa
riso
na
SIT
Spol
igot
ype
desc
ript
ion
Cla
deb
No.
(%)
ofst
rain
sin
this
stud
y
%in
this
stud
yco
mpa
red
toda
taba
sec
Dis
trib
utio
n�n
o.(%
)�in
neig
hbor
ing
coun
trie
sd
DE
UC
ZE
SWE
POL
RU
S
Orp
han
▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫�▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫�▫▫��
Unk
1(2
.2)
100
Orp
han
▫▫▫�����▫▫�����������▫����������▫▫▫▫�������
S1
(2.2
)10
0O
rpha
n������▫�▫▫�▫▫▫▫▫�▫��������������▫▫▫▫�������
Unk
1(2
.2)
100
Orp
han
������������▫����������������▫▫▫▫▫▫▫▫▫▫▫▫��
Unk
1(2
.2)
100
1▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫���������
Bei
jing
4(8
.7)
0.07
19(2
.7)
14(2
.6)
32(8
.4)
7(2
.2)
41(4
5.5)
37f
������������▫�������������������▫▫▫▫�������
T3
2(4
.3)
0.71
4(0
.6)
8(1
.5)
6(1
.6)
4(1
.3)
1(1
.1)
40f
������������������▫�������������▫▫▫▫�������
T4
1(2
.2)
0.81
5(0
.7)
7(1
.3)
1(0
.3)
42��������������������▫▫▫▫��������▫▫▫▫�������
LA
M9
1(2
.2)
0.04
6(0
.9)
25(4
.7)
12(3
.2)
6(1
.9)
5(5
.5)
47f
�������������������������▫▫▫▫▫▫�▫▫▫▫�������
H1
1(2
.2)
0.09
24(3
.4)
38(7
.2)
10(2
.6)
16(5
.0)
1(1
.1)
50f
������������������������������▫�▫▫▫▫�������
H3
1(2
.2)
0.04
50(7
.1)
80(1
5.1)
17(4
.5)
19(6
.0)
1(1
.1)
53��������������������������������▫▫▫▫�������
T1
6(1
3.0)
0.14
56(8
.0)
31(5
.8)
26(6
.9)
34(1
0.7)
11(1
2.2)
180
��▫���������������������������▫�▫▫▫▫�������
H3_
var
1(2
.2)
1.92
2(0
.4)
1(0
.3)
1(0
.3)
253f
��������������������▫��▫▫�������▫▫▫▫�������
Unk
1(2
.2)
3.57
1(0
.3)
280f
������▫▫▫▫▫▫▫▫▫▫▫▫��������������▫▫▫▫�������
T1-
RU
S21
(2.2
)1.
673
(0.4
)4
(1.3
)44
2������������▫�����▫�������������▫▫▫▫�������
T1
(2.2
)20
.00
708
���▫�������������▫��������������▫▫▫▫�������
X1_
var
1(2
.2)
9.09
2(2
.2)
775f
���������������������������▫▫▫▫▫▫▫▫▫▫������
Unk
1(2
.2)
2.94
8(1
.5)
1(0
.3)
891
��������������������▫▫▫▫�����▫��▫▫▫▫�������
LA
M9_
var
5(1
0.9)
18.5
27
(2.2
)1,
051
�������������������▫▫▫����������▫▫▫▫�������
Unk
3(6
.5)
12.0
010
(3.2
)1,
214
�����������▫▫▫������������������▫▫▫▫�������
T1
(2.2
)6.
671
(0.1
)1
(0.3
)1,
557
�����������▫�������������▫▫▫▫▫▫�▫▫▫▫�������
H1_
var
4(8
.7)
17.3
98
(2.5
)1,
558
�������������������������▫���▫��▫▫▫▫�������
T1
(2.2
)5.
001
(0.1
)9
(2.8
)1,
746
�▫▫�����▫▫�����������▫����������▫▫▫▫�������
S1
(2.2
)25
.00
1(0
.3)
2,33
3�������������������������▫▫▫▫▫▫�▫▫▫▫���▫▫��
H1_
var
1(2
.2)
14.2
92,
538e,
f�������������▫▫▫▫�▫▫▫▫▫▫▫�������▫▫▫▫�������
Unk
2(4
.3)
50.0
03,
093e
���������▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫������▫▫▫▫�������
Unk
1(2
.2)
50.0
03,
094e
���������������▫��▫▫▫▫����������▫▫▫▫�������
Unk
1(2
.2)
50.0
01
(0.3
)
aA
ll27
patt
erns
obse
rved
are
show
n,am
ong
whi
ch4
wer
eor
phan
s,20
belo
nged
toSI
Ts
desc
ribe
din
the
data
base
,and
3w
ere
new
lycr
eate
d.In
form
atio
non
geno
typi
clin
eage
san
ddi
stri
butio
nsin
neig
hbor
ing
coun
trie
s,w
hene
ver
avai
labl
ein
the
data
base
,is
also
show
n.b
Cla
des
wer
eas
sign
edfo
llow
ing
Spol
DB
4ru
les;
Unk
,unk
now
npa
tter
nsw
ithin
any
ofth
em
ajor
clad
esde
scri
bed
inSI
TV
IT2.
cPe
rcen
tage
ofst
rain
sin
the
SIT
VIT
2da
taba
seaf
ter
inco
rpor
atio
nof
the
spol
igot
ypes
from
the
pres
ent
stud
y(n
�46
).d
Dis
trib
utio
nin
neig
hbor
ing
coun
trie
sw
ithda
taav
aila
ble
inth
eSI
TV
IT2
data
base
on5
Nov
embe
r20
08.T
hree
-lett
erco
untr
yco
des
are
asfo
llow
s:D
EU
,Ger
man
y(n
�70
4);C
ZE
,Cze
chR
epub
lic(n
�53
0);S
WE
,Sw
eden
(n�
379)
;PO
L,P
olan
d(n
�31
7;st
rain
sfr
ompr
evio
usst
udie
s);R
US,
Kal
inin
grad
,Rus
sia
(n�
90).
eN
ewSI
Ts
crea
ted
afte
ra
mat
chw
ithin
this
stud
y(S
IT25
38;n
ote
that
2M
DR
stra
ins
mat
ched
2IN
H-r
esis
tant
stra
ins
with
inth
esa
me
stud
ype
riod
noti
nclu
ded
inth
eta
ble)
orw
ithan
othe
ror
phan
stra
inin
SIT
VIT
2(S
IT30
93m
atch
eda
stra
infr
omIt
aly;
SIT
3094
mat
ched
anor
phan
from
Pola
nd).
fSI
Ts
com
mon
toth
epa
tter
nspr
esen
tw
ithin
the
pool
ofIN
H-m
onor
esis
tant
stra
ins.
3902 JAGIELSKI ET AL. J. CLIN. MICROBIOL.

TA
BL
E2.
Spoligotyping-basedbiodiversity
observedam
ongIN
H-m
onoresistantstrains
(n�
71)and
SITV
IT2
databasecom
parisona
SITSpoligotype
descriptionC
ladeb
No.(%
)of
strainsin
thisstudy
%in
thisstudy
compared
todatabase
c
Distribution
�no.(%)�
inneighboring
countries d
DE
UC
ZE
SWE
POL
RU
S
Orphan
��������������▫▫▫�▫▫▫▫▫▫��������▫▫▫▫���▫���
Unk
1(1.4)
100O
rphan������▫▫▫▫▫▫▫▫▫▫▫▫���������▫����▫▫▫▫�������
T1-R
US2_var
1(1.4)
100O
rphan����▫▫▫▫▫�����������������������▫▫▫▫���▫▫▫▫
Unk
1(1.4)
100O
rphan���▫▫▫��������������������������▫▫▫▫���▫▫��
Unk
1(1.4)
100O
rphan▫▫▫▫▫▫▫▫▫▫▫▫▫▫������▫▫▫▫▫▫▫▫▫▫▫▫▫▫���������
Unk
1(1.4)
10037
������������▫�������������������▫▫▫▫�������
T3
1(1.4)
0.44
(0.6)8
(1.5)6
(1.6)4
(1.3)1
(1.1)40
������������������▫�������������▫▫▫▫�������
T4
1(1.4)
0.85
(0.7)7
(1.3)1
(0.3)44
����������������������▫���������▫▫▫▫�������
T5
1(1.4)
0.62
(0.3)8
(1.5)3
(0.9)47
�������������������������▫▫▫▫▫▫�▫▫▫▫�������
H1
5(7.0)
0.424
(3.4)38
(7.2)10
(2.6)16
(5.0)1
(1.1)50
������������������������������▫�▫▫▫▫�������
H3
7(9.9)
0.350
(7.1)80
(15.1)17
(4.5)19
(6.0)1
(1.1)53
��������������������������������▫▫▫▫�������
T1
24(33.8)
0.556
(8)31
(5.8)26
(6.9)34
(10.7)11
(12.2)253
��������������������▫��▫▫�������▫▫▫▫�������
Unk
2(2.8)
7.11
(0.3)262
�������▫▫�������������������▫▫▫�▫▫▫▫�������
H3_var
1(1.4)
0.81
(0.2)1
(0.3)3
(0.9)1
(1.1)280
������▫▫▫▫▫▫▫▫▫▫▫▫��������������▫▫▫▫�������
T1-R
US2
2(2.8)
3.33
(0.4)1
(0.3)4
(1.3)382
����▫��������������������▫▫▫▫▫▫�▫▫▫▫�������
H1
1(1.4)
8.31
(0.3)498
�����������▫��������������������▫▫▫▫�������
T1_var
1(1.4)
5.01
(0.1)1
(0.2)602
������������������������▫▫▫▫▫▫▫▫▫▫▫▫�������
Unk
1(1.4)
1.21
(0.1)1
(0.3)612
��������������������������������▫▫▫▫����▫��
T1_var
1(1.4)
5.3775
���������������������������▫▫▫▫▫▫▫▫▫▫������
Unk
1(1.4)
2.98
(1.5)1
(0.3)1,253
��▫�����▫▫�����������▫����������▫▫▫▫�������
S2
(2.8)12.5
1(0.1)
1,278▫▫▫▫�������▫��������������������▫▫▫▫�������
Unk
3(4.2)
25.01
(0.2)1
(0.3)1,410
���������������������▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫▫
Unk
1(1.4)
5.01
(0.3)1,498
��������������������������▫▫▫▫▫▫▫▫▫▫▫������
Unk
1(1.4)
11.11,640
������������������������������▫�▫▫▫▫����▫▫▫
H3_var
1(1.4)
33.31,821
���������▫���▫▫�����������������▫▫▫▫�������
Unk
1(1.4)
16.72,504
e�������������▫▫▫▫�▫▫▫�����������▫▫▫▫�������
Unk
3(4.2)
75.02,538
e�������������▫▫▫▫�▫▫▫▫▫▫▫�������▫▫▫▫�������
Unk
2(2.8)
50.02,567
e���������������▫▫�▫������▫▫▫▫▫▫�▫▫▫▫�������
H1
1(1.4)
50.01
(0.3)3,091
e�������������������������������▫▫▫▫▫▫▫▫▫���
Unk
1(1.4)
50.03,092
e������������▫�������▫▫▫▫����▫▫▫�▫▫▫▫�������
Unk
1(1.4)
50.01
(0.3)
aA
ll27patterns
observedare
shown,am
ongw
hich5
were
orphans,20belonged
toSIT
sdescribed
inthe
database,and5
were
newly
created.Information
ongenotypic
lineagesand
distributionsin
neighboringcountries,
whenever
availablein
thedatabase,is
alsoshow
n.b
Clades
were
assignedfollow
ingSpolD
B4
rules;Unk,unknow
npatterns
within
anyof
them
ajorclades
describedin
SITV
IT2.
cPercentageof
strainsin
theSIT
VIT
2database
afterincorporation
ofthe
spoligotypesfrom
thepresent
study(n
�71).
dD
istributionin
neighboringcountries
with
dataavailable
inthe
SITV
IT2
databaseon
5N
ovember
2008.Three-letter
countrycodes
areas
follows:D
EU
,Germ
any(n
�704);C
ZE
,Czech
Republic
(n�
530);SWE
,Sw
eden(n
�379);PO
L,Poland
(n�
317;strainsfrom
previousstudies);R
US,K
aliningrad,Russia
(n�
90).eN
ewSIT
screated
aftera
match
within
thisstudy
(SIT2538;note
that2IN
H-resistantstrains
matched
2M
DR
strainsw
ithinthe
same
studyperiod
notincludedin
thetable)
orw
ithanother
orphanstrain
inSIT
VIT
2(SIT
2504m
atcheda
strainfrom
Austria,SIT
2567and
SIT3092
matched
strainsfrom
previousstudies
inPoland,and
SIT3091
matched
astrain
fromT
aiwan).
VOL. 48, 2010 DRUG-RESISTANT TB IN POLAND 3903

two, the larger the number of TB cases included in the study,the higher the proportion of clustered or recently transmittedcases. Likewise, the observation time is positively associatedwith clustering, and it is considered that studies based on lessthan 2 years of sampling underestimate recent transmission(20, 35). Given the small pool of analyzed strains and fact thatthe study period lasted only 1 year, the rate of clustering forMDR-TB and INH-monoresistant isolates (56.5% and70.4%, respectively; the difference was not statistically sig-nificant; P � 0.1252) in our study seems to be high. Never-theless, there are conflicting results in the literature on therelative transmissibility of drug-resistant and drug-sensitiveM. tuberculosis strains. While some studies showed that drug-resistant isolates were less likely to be associated with cluster-ing (46, 48), others identified drug resistance as a strong pre-dictor of being in a cluster (1, 10).
In spite of a high degree of genetic diversity, an importantpart (41.3%), of the analyzed MDR strains belonged to one of
the four clusters SIT53, SIT891, SIT1, and SIT1557. The com-pactness of the MDR M. tuberculosis population structure wasalso apparent at the phylogenetic level, as four major lineages,i.e., the T, Haarlem, LAM, and Beijing families, compristedmore than 67% of the isolates studied (Table 1). With 30% ofthe isolates in the SpolDB4 database (9), the T family containspoorly informative, ubiquitous spoligotypes with obscure ge-nealogies. It is supposed, however, that most of the spoligo-types within the T clade represent relatively old genotypesprevalent in Europe (9). The Haarlem family, of Europeanorigin, comprises nearly 1/4 of the M. tuberculosis population inEurope, although it also accounts for a similar proportion ofstrains in the Caribbean and Central America (9, 17). In boththese regions, the Haarlem family is believed to represent aremnant of the post-Columbian European colonization (17,41). The finding of the Haarlem strains in our MDR popula-tion assumes particular significance in light of previous studiesdemonstrating the ability of the Haarlem genotypes to causeoutbreaks of MDR-TB, as reported in Argentina (37), theCzech Republic (30), and Tunisia (34). The association be-tween drug resistance and the Haarlem family has also beenobserved in other studies. Strains of this family predominatedamong MDR-TB cases in Tehran, Iran, and drug-resistant TBcases in Hungary; the rates of infection by the Haarlem geno-type were 33.5% (15) and 66.2% (28), respectively. The LAMfamily, which was the third-largest genogroup found in thisstudy, has a geographical distribution concentrated in LatinAmerica and the Mediterranean region. Strains of the LAMfamily are hypothesized to be of European descent, and theirhigh rate of occurrence in Latin American countries is sup-posed to be linked to the Portuguese and Spanish colonizationof the New World in the 15th century (8). It is noteworthy thatthe LAM family of strains in our study was representedexclusively by the LAM9 genotype, which is considered to beancestral to the entire LAM lineage (9). This finding mayspeak in favor of the historical presence of the LAM genotypestrains in this part of Europe. Also, the fact that an importantproportion of Polish MDR strains belonged to the LAM familymay be somewhat related to the situation observed in Poland’seastern neighbor, Russia, where the LAM genotypes representas much as 30 to 50% of the MDR strains (26, 33).
Upon review of the literature, the three genotype familiescharacterized above (Haarlem, LAM, and T) appeared to bedominant in many European countries. They represent asmuch as 60% of M. tuberculosis strains isolated in Italy (31),70% of the strains from Sweden (10), and 80% of the strainsfrom France and Portugal (6, 11). Thus, the population struc-ture revealed in our study is characteristic of a Europeancountry. Nonetheless, the finding in this study of four strainsbelonging to the Beijing genotype requires special attention.This is because strains of this genotype possess high epidemi-ological potential due to their enhanced transmissibility, par-ticular propensity for developing drug resistance, and in-creased virulence. The Beijing genotype strains were initiallyfound in China but have spread globally in recent years. Theprevalence of the Beijing family is remarkably high on theAsian continent, and in many countries (China, South Korea,Hong Kong, and Thailand) it far exceeds 50% (21). In otherparts of the world, the proportion of TB attributable to theBeijing genotype is much more variable: low in parts of Africa
TABLE 3. Comparison of demographic and epidemiologiccharacteristics of MDR-TB patients and
INH-monoresistant TB patients
Characteristic
No. (%) of patients with:
PMDR TB INH-monoresistant
TB
SexMale 40 (87) 52 (73.2) 0.0697Female 6 (13) 19 (26.8)
Age group (yr)0–14 0 1 (1.4) 0.724515–39 9 (19.6) 15 (21.1)40–64 28 (60.8) 39 (55)�65 9 (19.6) 16 (22.5)
OriginPolish born 45 (97.8) 70 (98.6) 0.6338Foreign born 1 (2.2) 1 (1.4)
Region of PolandCentral 15 (32.6) 21 (29.6) 0.9261Eastern 8 (17.4) 12 (16.9)Western 23 (50) 38 (53.5)
History of TBYes 39 (84.8) 16 (22.5) �0.0001a
No 7 (15.2) 55 (77.5)
AFB smearPositive 40 (87) 43 (60.6) 0.0014a
Negative 6 (13) 28 (39.4)
Strain clusteringYes 26 (56.5) 50 (70.4) 0.1252No 20 (43.5) 21 (29.6)
Spoligotype familyT 13 (28.3) 32 (45.1) 0.0006a
Haarlem 8 (17.4) 16 (22.5)LAM 6 (13) 0Beijing 4 (8.7) 0Orphan 4 (8.7) 5 (7)
Total 46 (100) 71 (100)
a Statistically significant at a P value of �0.05.
3904 JAGIELSKI ET AL. J. CLIN. MICROBIOL.

(0 to 4%), Latin America (�1%), and Western Europe(�6%); intermediate in North America and the Caribbean (8to 27%); and high in the countries of the former Soviet Union(29 to 56%) (13). Several studies from across the world havedocumented the association between the Beijing genotype anddrug resistance, including multidrug resistance (2, 15, 29, 39,44, 45). This association is most clearly evidenced in the post-Soviet countries. A study performed in the Archangel Oblast inRussia showed that Beijing strains accounted for 77% of alldrug-resistant TB cases and of all MDR-TB cases (44). InEstonia and Latvia, the Beijing strains were the cause of 87%and 58% of the MDR-TB cases, respectively (29, 45). In ourstudy, the proportion of MDR strains belonging to the Beijingfamily (9%) was similar to those reported in Spain (6%) (39)and Denmark (8%) (32). At the same time, it was about half ofthose reported in the Netherlands (17%) (7) and Sweden(22%) (19). In all these countries, most of the patients withBeijing isolates were immigrants from Asia and Eastern Eu-rope. Of the four Beijing strains identified in our study, onlyone was obtained from a foreign-born (Mongolian) patient. Itis noteworthy that the number of MDR-TB cases due to theBeijing genotype in Poland remained stable. In a previousstudy on the molecular epidemiology of drug-resistant M. tu-berculosis strains in Poland in 2000, four MDR and threenon-MDR isolates were found to belong to the Beijing geno-type. Three of those isolates (2 MDR and 1 non-MDR) wereobtained from patients born in countries where the genotype isprevalent (38). All this suggests that even if foreign-born per-sons constitute the main reservoir for the Beijing family strains,these strains are being transmitted in Poland, although ata very low rate. Whether previously proposed variable host-pathogen compatibility in M. tuberculosis (18) could explain thepropensity of specific mycobacterial lineages (such as the T orHaarlem clade) to spread within the Polish population to thedetriment of the Beijing genotype remains purely speculativeand open to discussion.
When comparing the genetic structure of the MDR strainswith that of INH-monoresistant strains, some important fea-tures were observed. First, a total of 9 shared types (SIT37,SIT40, SIT47, SIT50, SIT53, SIT253, SIT280, SIT775, andSIT2538) described for the MDR strains were also foundamong INH-monoresistant strains. These spoligotypes com-prised 34.8% of the former and 60.6% of the latter. Thispercentage difference was due to the fact that, for INH-resis-tant TB, spoligotype SIT53 accounted for 33.8% of all thecases (4). No such predominance of a single genotype wasobserved among MDR-TB cases. This finding might be indic-ative of a greater ability of drug-monoresistant M. tuberculosisstrains to spread within the population compared with that ofMDR strains. Second, 76.1% of the MDR isolates and 71.8%of the INH-resistant isolates yielded spoligotypes that hadbeen reported in Poland previously (including SIT2538, sharedby 2 MDR strains and 2 INH-resistant strains). This observa-tion, together with the fact that nearly half of the spoligotypesidentified among both MDR (48.1%) and INH-monoresistant(43.3%) M. tuberculosis isolates are present in Poland’s neigh-boring countries, indicates that the vast majority of MDR- andINH-resistant-TB cases are due to infections with strains ac-tively circulating in Poland and/or its neighbors. The fact thatall but two patients (one Mongolian, with MDR-TB, and one
Ukrainian, with INH-resistant TB) were Polish underlines theimportance of in-country transmission. Third, the phylogeneticanalysis revealed that the two most prevalent clades amongMDR and INH-resistant M. tuberculosis strains were T andHaarlem, representing 46% and 80% of these groups, respec-tively. These results corroborate previous observations on thepredominance of the T and Haarlem genotypes in the epide-miology of drug-resistant TB in Poland (5, 38). Furthermore,the presence of the T and Haarlem genotype isolates in suchhigh proportions is consistent with the general features of theEuropean population of M. tuberculosis.
Although the distributions of the four major clades (T,Haarlem, LAM, and Beijing) were significantly different in theMDR and INH-monoresistant strains (P � 0.0003), the moststriking feature was the exclusive presence of Beijing and LAMlineages in the MDR group (8.7% and 13%, respectively). Thissuggests that, in addition to the possible emergence of MDRstrains from a pool of INH-monoresistant strains of the T, H1,H3, and S lineages, a proportion of MDR-TB cases could alsoarise due to ongoing transmission of specific genotypes, such asLAM and Beijing. To better recognize the population geneticrelationships between INH-monoresistant (n � 71) and MDR(n � 46) M. tuberculosis isolates, we drew and compared MSTsof these 2 data sets independently (Fig. 2A and B, respec-tively), as well as a combined tree for all the isolates together(n � 117) (Fig. 2C). As mentioned earlier, the greatest con-tributor to INH-monoresistant strains of M. tuberculosis inPoland is the cluster made up of SIT53 strains belonging tothe T1 sublineage (33.8% of all INH-monoresistant strains).The T1 strains, represented by its prototype, SIT53, consti-tute the central node of the MST shown in Fig. 2A. Nonethe-less, there are known difficulties associated with attempts todraw epidemiological inferences from ill-defined T-family spo-ligotypes (representing evolutionarily recent TB strains), whichcorresponded to about 30% of all entries in the SpolDB4database present in all continents (9). Based on single spacerdifferences, they were initially stratified into 5 sublineages (T1to T5), out of which 8 nested clades with robust spoligotypingsignatures have been extracted so far (T-Tuscany, T1-Russia,T2-Uganda, T3-Ethiopia, T3-Osaka, T4-Central Europe, T5-Madrid, and T5-Russia). Consequently, further molecularcharacterization of the isolates designated “T” by default (17)is needed before phylogenetic inferences are drawn from thecentral node made up of spoligotype SIT53. In the MST shownin Fig. 2A, it is immediately followed by SIT50 (9.9%), SIT47(7%), and SIT1253 (2.8%), which are successively placed onthe MST within the well-defined H3, H1, and S subgroups. Thespoligotyping results and clade definitions were also linked tothe distribution of clinical isolates within PGG1 versus PGG2/3(characterized by the lack of spacers 33 to 36). With the ex-ception of a single orphan strain with an unknown signature(the 5th strain from the top in Table 2), isolates causing thebulk of INH monoresistance in Poland were made up of evo-lutionarily modern lineages of tubercle bacilli (T, Haarlem,and S). The T and Haarlem lineages/sublineages were dulyrepresented by prototypes that characterized their descriptionin the SpolDB4 database (9): T1 (SIT53), T1_RUS (SIT280),T3 (SIT37), T4 (SIT40), T5 (SIT44), H1 (SIT47), and H3(SIT50).
The MST of the MDR strains involved five major branches
VOL. 48, 2010 DRUG-RESISTANT TB IN POLAND 3905
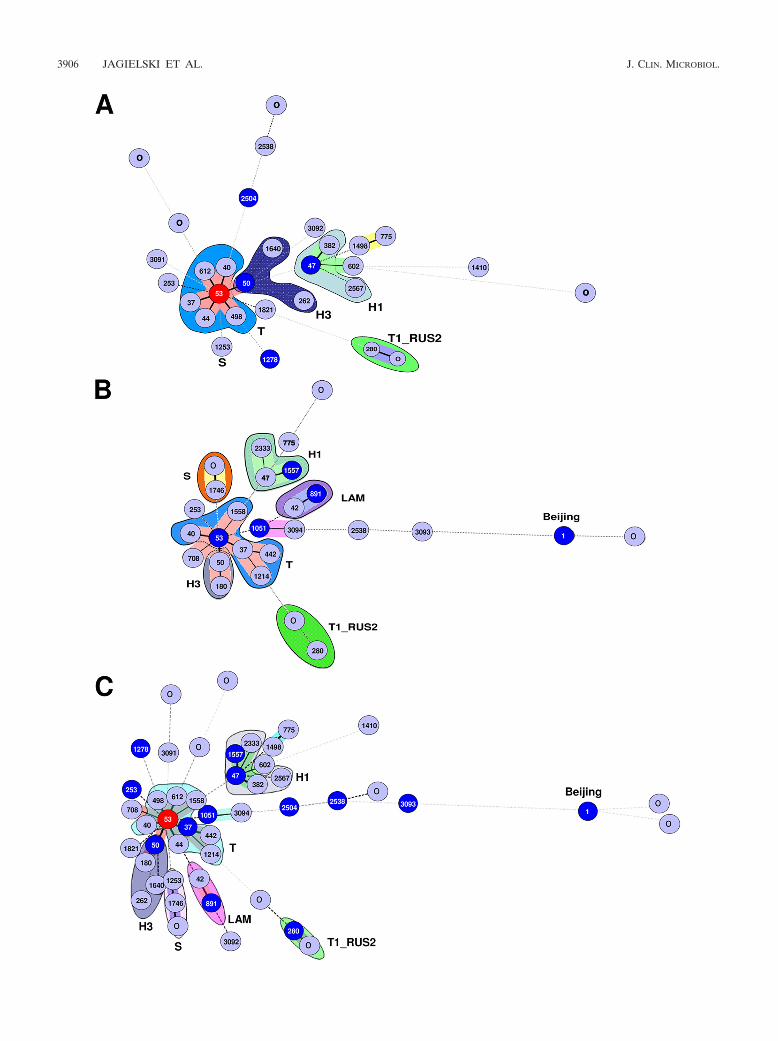
3906 JAGIELSKI ET AL. J. CLIN. MICROBIOL.

(Fig. 2B). The largest essentially contained genotypes from theill-defined T family. The other four branches exclusively orpredominantly included genotypes from the Haarlem (H1 andH3), LAM, Beijing, and S families (Fig. 2B). The genotypeSIT53 had a central position in this MST similar to that of theINH-monoresistant strains. However, as stressed above, furthermolecular characterization of SIT53 node strains is needed be-fore phylogenetic inferences are drawn. Thus, a distinctivefeature of the MDR population was the presence of the Beijingand LAM family strains and a significantly lower proportion ofHaarlem family strains compared with the INH-monoresistantpopulation. This observation suggests that, in addition to ac-quired resistance to RMP occurring within a pool of INH-monoresistant strains, a nonnegligible proportion of MDR-TBin Poland is due both to active transmission within the autoch-thonous population and to frequent contacts with neighboringcountries. A second-line typing using MIRU-VNTRs and de-tailed contact tracing, in conjunction with characterization ofspecific katG alleles shared among INH-monoresistant andMDR strains (33, 38, 46), might be helpful to answer thisquestion.
Interestingly, a combined tree for all the isolates together(Fig. 2C) (n � 117 isolates) reproduced most of the phyloge-netic characteristics mentioned above for the MDR group. Forexample, similar to the MDR tree, the branches leading to the(i) T1_RUS, (ii) H1, and (iii) H3 sublineages were perfectlymaintained, with the same succession of intermediary spoligo-types (Fig. 2C). The lack of insertion of INH-monoresistantstrains within these well-defined branches, in conjunction withthe exclusive presence of LAM and Beijing lineage strainswithin the MDR subset, underlines an active transmission ofMDR strains belonging to the T, H1, T1-RUS2, and H3 lin-eages. On the other hand, strains belonging to the LAM andBeijing genotypic families are apparently transmitted indepen-dently of the prevailing INH-monoresistant pool. At the levelof PGG classification, the only PGG1 strains included 4 Beijingstrains in the MDR-TB sample. Added to the single PGG1orphan strain in the INH-monoresistant group (see above), thetotal study sample (n � 117 strains) contained only 5, or 4.3%,of the PGG1 (ancient-lineage) strains compared to a very highproportion of PGG2/3 modern-lineage strains (112, or 95.7%).In conjunction with the noted absence of the CAS and EAIlineages in our study, this observation undeniably shows thatcirculating M. tuberculosis strains causing the bulk of drugresistance in Poland are made up of evolutionarily moderntubercle bacilli of the T, Haarlem, and S genotypic lineages.
Finally, we compared some of the basic epidemiologicalcharacteristics of the MDR-TB and INH-monoresistant-TBpatients (Table 3). In both these groups, male patients pre-
dominated. The male-to-female ratios among MDR-TB pa-tients and INH-resistant-TB patients were 6.7:1 and 2.7:1, re-spectively. Patients with MDR-TB were only slightly older thanthose with INH-resistant TB (median age, 50.5 versus 48years). All of the patients studied were Polish, except two whowere of Mongolian (MDR-TB group) and Ukrainian (INH-resistant-TB group) origin. There were no differences betweenMDR-TB cases and INH-resistant cases in terms of prevalenceby region of residence. The prevalence of TB, regardless of theresistance phenotype, was highest in the western region ofPoland, followed by the central and eastern regions. The pro-portion of patients who reported a history of TB was signifi-cantly higher among MDR-TB patients than among INH-re-sistant TB patients (84.8% versus 22.5%; P � 0.0001). Patientswith MDR-TB were also more likely to have acid-fast bacillus(AFB) smear-positive sputa (P � 0.0014). The epidemiologicalstatus of MDR-TB in Poland, as inferred from patient demo-graphic and clinical features, fits generally into the epidemio-logical picture of this infection described for other Europeancountries. In a systemic review based on studies from 12 coun-tries, being male, being less than 65 years old, and having ahistory of previous TB treatment were among major risk fac-tors for MDR-TB in Europe (16). A reported history ofanti-TB medication, which, according to the cited study, was thestrongest determinant of MDR-TB, may suggest the acquirednature of drug resistance. A strong correlation betweenMDR-TB and sputum smear positivity in our study is alsonoteworthy. Such a correlation, very seldom encountered insimilar investigations (25), may reflect the fact that patientswith MDR-TB often did not receive adequate treatment at theoutset (until the drug susceptibility results were available), thusremaining infectious for prolonged periods. A separate paperin preparation, devoted to the treatment of TB patients, showsthat most, if not all, MDR-TB patients in Poland belong to thepoorest members of the society; often having associated phys-ical or mental handicaps, they were also characterized withpoorer follow-up and longer delays to diagnosis and treatment(results not shown). Interestingly, there were virtually noMDR-TB cases among foreign-born persons in our study. Thisis in contrast to what has been observed in Western Europeancountries, where the association between MDR-TB and beingforeign born is remarkably consistent. The near absence ofimmigrants among MDR-TB cases can be explained by therelatively small immigrant population living in Poland.
In conclusion, our study is the first to provide a detailedpopulation-based phylogenetic analysis of the MDR M. tuber-culosis isolates compared to prevailing INH-monoresistantstrains in Poland. Although spoligotyping, with its limited dis-criminatory power, may have overlooked finer intraspecies re-
FIG. 2. MST illustrating the potential evolutionary relationships of M. tuberculosis spoligotypes in Poland. The tree summarizes the phyloge-netic links between two spoligotypes differing by genetic changes. The lengths of the branches indicate the levels of changes induced by loss or gainof spoligotype spacers in the 43-oligonucleotide format to induce a shift from one allele to another. Solid lines represent a single spacer change,while dotted lines represent 2 (black) or more (gray) spacer changes. The colors of the circles are proportional to the number of clinical isolatesin our study: sky blue, 1 or 2 isolates; marine blue, 3 to 5 isolates; deep blue, 6 or 7 strains; and red, �8 strains. The remaining colors in the treedrawing denote strain lineages/sublineages for closely related strains such as T1_RUS2, T, H1, S, etc. The tree drawing was set to integrate databased on both single and multiple DVR deletions and to further differentiate between the loss of contiguous and noncontiguous multiple directvariable repeats (DVRs). (A) MST of isoniazid-monoresistant strains. (B) MST of MDR strains. (C) Combined MST for all the isolates together(n � 117). Note that the central node of the unrooted trees is represented by SIT53, which is the prototype of the T1 lineage.
VOL. 48, 2010 DRUG-RESISTANT TB IN POLAND 3907

lationships within the M. tuberculosis complex, our results em-phasize that the persistence of MDR-TB in Poland seeminglyresults from active transmission of well-established clones ofM. tuberculosis historically linked to Poland within the autoch-thonous population. Some of the spoligotype-defined lineagesand sublineages of the MDR group were shared by the INH-monoresistant group, which indicates that further contact-trac-ing epidemiology and a more discriminatory molecular meth-odology are warranted to pinpoint the major contributor to thedevelopment and persistence of MDR-TB in Poland. This isimportant in the context of the known instances of conversionreported in the literature from INH monoresistance to MDR(12), in which 10 out of 12 INH-monoresistant strains con-verted to MDR after treatment failure during therapy. If con-firmed in extended studies, it will constitute a data-supportedinstance of INH monoresistance conversion to MDR status inPoland. Nonetheless, the exclusive presence of LAM and Bei-jing family strains within the MDR group suggests a particularrole of these genotypes in the etiology of MDR-TB emergencein Poland.
ACKNOWLEDGMENTS
The SITVIT2 database project was partially financed by the Re-gional Council of Guadeloupe (CR/08-1612; Biodiversite et RisqueInfectieux dans les Modeles Insulaires).
Tomasz Jagielski is grateful to Łukasz Szafron (Ph.D. student, De-partment of Molecular Pathology, Institute of Oncology, Warsaw) forhelping to draw the administrative map of Poland and prepare the finalversion of Fig. 1 in this article.
REFERENCES
1. Alland, D., G. E. Kalkut, A. R. Moss, R. A. McAdam, J. A. Hahn, W.Bosworth, E. Drucker, and B. R. Bloom. 1994. Transmission of tuberculosisin New York City. An analysis by DNA fingerprinting and conventionalepidemiologic methods. N. Engl. J. Med. 330:1710–1716.
2. Almeida, D., C. Rodrigues, T. F. Ashavaid, A. Lalvani, Z. F. Udwadia, and A.Mehta. 2005. High incidence of the Beijing genotype among multidrug-resistant isolates of Mycobacterium tuberculosis in a tertiary care center inMumbai, India. Clin. Infect. Dis. 40:881–886.
3. Augustynowicz-Kopec E., Z. Zwolska, A. Jaworski, E. Kostrzewa, and M.Klatt. 2003. Drug-resistant tuberculosis in Poland in 2000: second nationalsurvey and comparison with the 1997 survey. Int. J. Tuberc. Lung Dis.7:645–651.
4. Augustynowicz-Kopec, E., T. Jagielski, and Z. Zwolska. 2008. Genetic diver-sity of isoniazid-resistant Mycobacterium tuberculosis isolates collected inPoland and assessed by spoligotyping. J. Clin. Microbiol. 46:4041–4044.
5. Augustynowicz-Kopec, E., T. Jagielski, M. Kozinska, A. Zabost, M. Klatt,and Z. Zwolska. 2008. Molecular analysis of drug-resistant Mycobacteriumtuberculosis isolates collected in central Poland. Clin. Microbiol. Infect. 14:605–607.
6. Bezanahary, H., M. C. Baclet, C. Sola, V. Gazaille, P. Turlure, P. Weinbreck,F. Denis, and C. Martin. 2006. Interet du typage moleculaire pour l’epide-miologie de la tuberculose en Limousin (1998–2006). Med. Mal. Infect.38:309–317.
7. Borgdorff, M. W., P. de Haas, K. Kremer, and D. van Soolingen. 2003.Mycobacterium tuberculosis Beijing genotype, the Netherlands. Emerg. In-fect. Dis. 9:1310–1313.
8. Brudey, K., I. Filliol, S. Ferdinand, V. Guernier, P. Duval, B. Maubert, C.Sola, and N. Rastogi. 2006. Long-term population-based genotyping study ofMycobacterium tuberculosis complex isolates in the French departments ofthe Americas. J. Clin. Microbiol. 44:183–191.
9. Brudey, K., J. R. Driscoll, L. Rigouts, W. M. Prodinger, A. Gori, S. A.Al-Hajoj, C. Allix, L. Aristimuno, J. Arora, V. Baumanis, L. Binder, P.Cafrune, A. Cataldi, S. Cheong, R. Diel, C. Ellermeier, J. T. Evans, M.Fauville-Dufaux, S. Ferdinand, D. Garcia de Viedma, C. Garzelli, L. Gaz-zola, H. M. Gomes, M. C. Guttierez, P. M. Hawkey, P. D. van Helden, G. V.Kadival, B. N. Kreiswirth, K. Kremer, M. Kubin, S. P. Kulkarni, B. Liens,T. Lillebaek, M. L. Ho, C. Martin, I. Mokrousov, O. Narvskaïa, Y. F. Ngeow,L. Naumann, S. Niemann, I. Parwati, Z. Rahim, V. Rasolofo-Razanam-parany, T. Rasolonavalona, M. L. Rossetti, S. Rusch-Gerdes, A. Sajduda, S.Samper, I. G. Shemyakin, U. B. Singh, A. Somoskovi, R. A. Skuce, D. vanSoolingen, E. M. Streicher, P. N. Suffys, E. Tortoli, T. Tracevska, V. Vincent,
T. C. Victor, R. M. Warren, S. F. Yap, K. Zaman, F. Portaels, N. Rastogi, andC. Sola. 2006. Mycobacterium tuberculosis complex genetic diversity: miningthe fourth international spoligotyping database (SpolDB4) for classification,population genetics and epidemiology. BMC Microbiol. 6:23.
10. Brudey, K., M. Gordon, P. Mostrom, L. Svensson, B. Jonsson, C. Sola, M.Ridell, and N. Rastogi. 2004. Molecular epidemiology of Mycobacteriumtuberculosis in Western Sweden. J. Clin. Microbiol. 42:3046–3051.
11. David, S., D. R. Ribeiro, A. Antunes, C. Portugal, L. Sancho, and J. G. deSousa. 2007. Contribution of spoligotyping to the characterization of thepopulation structure of Mycobacterium tuberculosis isolates in Portugal. In-fect. Genet. Evol. 7:609–617.
12. Dye, C., and M. A. Espinal. 2001. Will tuberculosis become resistant to allantibiotics? Proc. R. Soc. Lond. B 268:45–52.
13. European Concerted Action on New Generation Genetic Markers and Tech-niques for the Epidemiology and Control of Tuberculosis. 2006. Beijing/Wgenotype Mycobacterium tuberculosis and drug resistance. Emerg. Infect. Dis.12:736–743.
14. EuroTB. 2006. Surveillance of tuberculosis in Europe: report on tuberculosiscases notified in 2004. EuroTB-Institut de Veille Sanitaire, Saint-Maurice,France.
15. Farnia, P., M. R. Masjedi, M. Mirsaeidi, F. Mohammadi, Jallaledin-Gha-navi, V. Vincent, M. Bahadori, and A. A. Velayati. 2006. Prevalence ofHaarlem I and Beijing types of Mycobacterium tuberculosis strains in Iranianand Afghan MDR-TB patients. J. Infect. 53:331–336.
16. Faustini, A., A. J. Hall, and C. A. Perucci. 2006. Risk factors for multidrugresistant tuberculosis in Europe: a systemic review. Thorax 61:158–163.
17. Filliol, I., J. R. Driscoll, D. van Soolingen, B. N. Kreiswirth, K. Kremer, G.Valetudie, D. A. Dang, R. Barlow, D. Banerjee, P. J. Bifani, K. Brudey, A.Cataldi, R. C. Cooksey, D. V. Cousins, J. W. Dale, O. A. Dellagostin, F.Drobniewski, G. Engelmann, S. Ferdinand, D. Gascoyne-Binzi, M. Gordon,M. C. Gutierrez, W. H. Haas, H. Heersma, E. Kassa-Kelembho, M. L. Ho, A.Makristathis, C. Mammina, G. Martin, P. Mostrom, I. Mokrousov, V. Nar-bonne, O. Narvskaya, A. Nastasi, S. N. Niobe-Eyangoh, J. W. Pape, V.Rasolofo-Razanamparany, M. Ridell, M. L. Rossetti, F. Stauffer, P. N. Suf-fys, H. Takiff, J. Texier-Maugein, V. Vincent, J. H. de Waard, C. Sola, and N.Rastogi. 2003. Snapshot of moving and expanding clones of Mycobacteriumtuberculosis and their global distribution assessed by spoligotyping in aninternational study. J. Clin. Microbiol. 41:1963–1970.
18. Gagneux, S., K. DeRiemer, T. Van, M. Kato-Maeda, B. C. de Jong, S.Narayanan, M. Nicol, S. Niemann, K. Kremer, M. C. Gutierrez, M. Hilty,P. C. Hopewell, and P. M. Small. 2006. Variable host-pathogen compatibilityin Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. U. S. A. 103:2869–2873.
19. Ghebremichael, S., R. Petersson, T. Koivula, A. Pennhag, V. Romanus, I.Berggren, B. Petrini, S. Hoffner, and G. Kallenius. 2008. Molecular epide-miology of drug-resistant tuberculosis in Sweden. Microbes Infect. 10:699–705.
20. Glynn, J. R., J. Bauer, A. S. de Boer, M. W. Borgdorff, P. E. Fine, P.Godfrey-Faussett, and E. Vynnycky. 1999. Interpreting DNA fingerprintclusters of Mycobacterium tuberculosis. European Concerted Action on Mo-lecular Epidemiology and Control of Tuberculosis. Int. J. Tuberc. Lung Dis.3:1055–1060.
21. Glynn, J. R., J. Whiteley, P. J. Bifani, K. Kremer, and D. van Soolingen.2002. Worldwide occurrence of Beijing/W strains of Mycobacterium tubercu-losis: a systematic review. Emerg. Infect. Dis. 8:843–849.
22. Goguet de la Salmoniere, Y. O., H. M. Li, G. Torrea, A. Bunschoten, J. D. A.van Embden, and B. Gicquel. 1997. Evaluation of spoligotyping in a study ofthe transmission of Mycobacterium tuberculosis. J. Clin. Microbiol. 35:2210–2214.
23. Gori, A., A. Bandera, G. Marchetti, A. Degli Esposti, L. Catozzi, G. P. Nardi,L. Gazzola, G. Ferrario, J. D. van Embden, D. van Soolingen, M. Moroni,and F. Franzetti. 2005. Spoligotyping and Mycobacterium tuberculosis.Emerg. Infect. Dis. 11:1242–1248.
24. Goyal, M., N. A. Saunders, J. D. A. van Embden, D. B. Young, and R. J.Shaw. 1997. Differentiation of Mycobacterium tuberculosis isolates byspoligotyping and IS6110 restriction fragment length polymorphism. J. Clin.Microbiol. 35:647–651.
25. Granich, R. M., P. Oh, B. Lewis, T. C. Porco, and J. Flood. 2005. Multidrugresistance among persons with tuberculosis in California, 1994–2003. JAMA293:2732–2739.
26. Ignatova, A., S. Dubiley, V. Stepanshina, and I. Shemyakin. 2006. Predom-inance of multi-drug-resistant LAM and Beijing family strains among Myco-bacterium tuberculosis isolates recovered from prison inmates in Tula Re-gion, Russia. J. Med. Microbiol. 55:1413–1418.
27. Kamerbeek, J., L. Schouls, A. Kolk, M. van Agterveld, D. van Soolingen, S.Kuijper, A. Bunschoten, H. Molhuizen, R. Shaw, M. Goyal, and J. D. A. vanEmbden. 1997. Simultaneous detection and strain differentiation of Myco-bacterium tuberculosis for diagnosis and epidemiology. J. Clin. Microbiol.35:907–914.
28. Kodmon, C., S. Niemann, J. Lukacs, E. Sor, S. David, and A. Somoskovi.2006. Molecular epidemiology of drug-resistant tuberculosis in Hungary.J. Clin. Microbiol. 44:4258–4261.
29. Kruuner, A., S. E. Hoffner, H. Sillastu, M. Danilovits, K. Levina, S. B.
3908 JAGIELSKI ET AL. J. CLIN. MICROBIOL.

Svenson, S. Ghebremichael, T. Koivula, and G. Kallenius. 2001. Spread ofdrug-resistant pulmonary tuberculosis in Estonia. J. Clin. Microbiol. 39:3339–3345.
30. Kubín, M., M. Havelkova, I. Hyncicova, Z. Svecova, J. Kaustova, K. Kremer,and D. van Soolingen. 1999. A multidrug-resistant tuberculosis microepi-demic caused by genetically closely related Mycobacterium tuberculosisstrains. J. Clin. Microbiol. 37:2715–2716.
31. Lari, N., L. Rindi, D. Bonanni, N. Rastogi, C. Sola, E. Tortoli, and C.Garzelli. 2007. Three-year longitudinal study of genotypes of Mycobacteriumtuberculosis isolates in Tuscany, Italy. J. Clin. Microbiol. 45:1851–1857.
32. Lillebaek, T., A. B. Andersen, A. Dirksen, J. R. Glynn, and K. Kremer. 2003.Mycobacterium tuberculosis Beijing genotype. Emerg. Infect. Dis. 9:1553–1557.
33. Lipin, M. Y., V. N. Stepanshina, I. G. Shemyakin, and T. M. Shinnick. 2007.Association of specific mutations in katG, rpoB, rpsL and rrs genes withspoligotypes of multidrug-resistant Mycobacterium tuberculosis isolates inRussia. Clin. Microbiol. Infect. 13:620–626.
34. Mardassi, H., A. Namouchi, R. Haltiti, M. Zarrouk, B. Mhenni, A. Karboul,N. Khabouchi, N. C. Gey van Pittius, E. M. Streicher, J. Rauzier, B. Gicquel,and K. Dellagi. 2005. Tuberculosis due to resistant Haarlem strain, Tunisia.Emerg. Infect. Dis. 11:957–961.
35. Murray, M., and D. Alland. 2002. Methodological problems in the molecularepidemiology of tuberculosis. Am. J. Epidemiol. 155:565–571.
36. Nivin, B., J. Driscoll, T. Glaser, P. Bifani, and S. Munsiff. 2000. Use ofspoligotype analysis to detect laboratory cross-contamination. Infect. Con-trol Hosp. Epidemiol. 21:525–527.
37. Ritacco, V., M. Di Lonardo, A. Reniero, M. Ambroggi, L. Barrera, A. Dam-brosi, B. Lopez, N. Isola, and I. N. de Kantor. 1997. Nosocomial spread ofhuman immunodeficiency virus-related multidrug-resistant tuberculosis inBuenos Aires. J. Infect. Dis. 176:637–642.
38. Sajduda, A., A. Brzostek, M. Popławska, N. Rastogi, C. Sola, E. Augustynow-icz-Kopec, Z. Zwolska, J. Dziadek, and F. Portaels. 2004. Molecular epide-miology of drug-resistant Mycobacterium tuberculosis strains isolated frompatients with pulmonary tuberculosis in Poland: a 1-year study. Int. J. Tu-berc. Lung Dis. 8:1448–1457.
39. Samper, S., M. J. Iglesias, M. J. Rabanaque, L. I. Gomez, M. C. Lafoz, M. S.Jimenez, A. Ortega, M. A. Lezcano, D. van Soolingen, C. Martín, and theSpanish Working Group on MDR-TB. 2005. Systematic molecular charac-terization of multidrug-resistant Mycobacterium tuberculosis complex isolatesfrom Spain. J. Clin. Microbiol. 43:1220–1227.
40. Sola, C., A. Devallois, L. Horgen, J. Maïsetti, I. Filliol, E. Legrand, and N.
Rastogi. 1999. Tuberculosis in the Caribbean: using spacer oligonucleotidetyping to understand strain origin and transmission. Emerg. Infect. Dis.5:404–414.
41. Sola, C., I. Filliol, M. C. Gutierrez, I. Mokrousov, V. Vincent, and N. Rastogi.2001. Spoligotype database of Mycobacterium tuberculosis: biogeographicdistribution of shared types and epidemiologic and phylogenetic perspec-tives. Emerg. Infect. Dis. 7:390–396.
42. Sreevatsan, S., X. Pan, K. E. Stockbauer, N. D. Connell, B. N. Kreiswirth,T. S. Whittam, and J. M. Musser. 1997. Restricted structural gene polymor-phism in the Mycobacterium tuberculosis complex indicates evolutionarilyrecent global dissemination. Proc. Natl. Acad. Sci. U. S. A. 94:9869–9874.
43. Szczuka, I. 2005. Incidence of tuberculosis in Poland 2004 and its trends.Pneumonol. Alergol. Pol. 73:221–227.
44. Toungoussova, O. S., P. Sandven, A. O. Mariandyshev, N. I. Nizovtseva, G.Bjune, and D. A. Caugant. 2002. Spread of drug-resistant Mycobacteriumtuberculosis strains of the Beijing genotype in the Archangel Oblast, Russia.J. Clin. Microbiol. 40:1930–1937.
45. Tracevska, T., I. Jansone, V. Baumanis, O. Marga, and T. Lillebaek. 2003.Prevalence of Beijing genotype in Latvian multidrug-resistant Mycobacte-rium tuberculosis isolates. Int. J. Tuberc. Lung Dis. 7:1097–1103.
46. van Doorn, H. R., P. E. de Haas, K. Kremer, C. M. Vandenbroucke-Grauls,M. W. Borgdorff, and D. van Soolingen. 2006. Public health impact ofisoniazid-resistant Mycobacterium tuberculosis strains with a mutation atamino-acid position 315 of katG: a decade of experience in the Nether-lands. Clin. Microbiol. Infect. 12:769–775.
47. van Embden, J. D. A., M. D. Cave, J. T. Crawford, J. W. Dale, K. D.Eisenach, B. Gicquel, P. Hermans, C. Martin, R. McAdam, T. M. Schinnick,and P. M. Small. 1993. Strain identification of Mycobacterium tuberculosis byDNA fingerprinting: recommendations for a standardized methodology.J. Clin. Microbiol. 31:406–409.
48. van Soolingen, D., M. W. Borgdorff, P. E. de Haas, M. M. Sebek, J. Veen, M.Dessens, K. Kremer, and J. D. van Embden. 1999. Molecular epidemiologyof tuberculosis in the Netherlands: a nationwide study from 1993 through1997. J. Infect. Dis. 180:726–736.
49. World Health Organization and International Union against Tuberculosisand Lung Disease. 2008. The WHO/IUATLD Global Project on Anti-Tuberculosis Drug Resistance Surveillance, 2002–2007: anti-tuberculosisdrug resistance in the world, 4th global report. WHO/HTM/TB/2008.394.WHO, Geneva, Switzerland.
50. World Health Organization. 2006. Global tuberculosis control: surveillance,planning, financing. WHO, Geneva, Switzerland.
VOL. 48, 2010 DRUG-RESISTANT TB IN POLAND 3909




















