
P-Glycoprotein and Multidrug Resistance - NCBI
13
Oliver Bähr 1 ; Johannes Rieger 1 ; Frank Duffner 2 ; Richard Meyermann 3 ; Michael Weller 1 ; Wolfgang Wick 1 1 Laboratory of Molecular Neuro-Oncology, Department of Neurology, 2 Department of Neurosurgery, 3 Institute for Brain Research, University of Tübingen, Germany. Understanding and overcoming multidrug resist- ance (MDR) may be a promising strategy to develop more effective pharmacotherapies for malignant gliomas. In the present study, human malignant glioma cell lines (n = 12) exhibited heterogeneous mRNA and protein expression and functional activ- ity of the mdr gene-encoded P-glycoprotein (PGP) and MDR-associated protein (MRP). Correlation between mRNA expression, protein levels and functional activity was strong. Inhibition of PGP activity by ver- apamil or PSC 833 enhanced the cytotoxic effects of vincristine, doxorubicin, teniposide and taxol. Inhi- bition of MRP activity by indomethacin or probenecid enhanced the cytotoxic effects of vin- cristine, doxorubicin and teniposide. The human cerebral endothelial cell line, SV-HCEC, exhibited the strongest PGP activity of all cell lines. Five primary human glioblastomas and one anaplastic astrocy- toma displayed heterogenous protein levels of PGP and MRP-1 in tumor cells and of PGP in biopsy specimens in vivo, but no functional activity of these proteins upon ex vivo culturing. These data sug- gest that the glioma cell line-associated MDR-type drug resistance is a result of long-term culturing and that cerebral endothelial, but not glioma cells, may contribute to MDR-type drug resistance of gliomas in vivo. Introduction Multidrug resistance resulting from the expression of the mdr-1 and mrp-1 gene products is one of the major obstacles to successful chemotherapy in several types of cancer (6, 7, 15). The corresponding proteins, PGP and MRP, belong to the family of ATP binding cassette (ABC) transporters which function as ATP-dependent efflux pumps. Amongst others, PGP and MRP transport various common cancer chemotherapeutics. Malignant gliomas, notably glioblastomas, are large- ly resistant to current treatment strategies which include cytoreductive surgery, involved-field radiotherapy and adjuvant chemotherapy. Less than 30% of patients with glioblastoma benefit from adjuvant chemotherapy (9, 14). It would be a great clinical advance if these patients could be identified prospectively by examining specif- ic biological parameters. We have previously reported that several candidate genes for drug sensitivity, including p53, p16, cyclin-dependent kinase (CDK)-4, murine double minute (MDM)-2 and various B cell lymphoma (BCL)-2 family proteins fail to predict response to chemotherapy in human malignant glioma cell lines (30). The clinical significance of the MDR phenotype in glioblastoma has remained controversial (2). Immuno- histochemical analyses of human glioma sections indi- cate that both the tumor cells proper and the endothelial cells express MDR in human gliomas (2, 4, 8, 16, 19, 21, 28, 29). The role of endothelial cells has also been strengthened by an in vivo xenograft study (27). Further, disruption of the mdr-1a gene in mice resulted in enhanced vinblastine accumulation in the normal brain (25). The effects of adjuvant cytotoxic therapies on the MDR phenotype in gliomas in vivo have been examined in 2 immunohistochemical studies: a Japanese study observed an increase in MRP/PGP expression after chemotherapy (2) whereas no such effect was observed in our own series (23). Few studies have addressed the functional activity and the drug specificity of MDR and MRP in glioma cells (1, 10, 18, 23, 24). High levels of mrp-1 mRNA expression in glioma cell lines were linked to reduced etoposide accumulation and to resist- ance to etoposide, vincristine and doxorubicin (1). Kiwit and colleagues observed that the calcium antagonist, verapamil, enhanced the sensitivity of PGP-expressing glioma cells to ACNU, cisplatin and vincristine (18). We previously noted that the chemosensitizing properties of RESEARCH ARTICLE P-Glycoprotein and Multidrug Resistance- associated Protein Mediate Specific Patterns of Multidrug Resistance in Malignant Glioma Cell Lines, but not in Primary Glioma Cells Corresponding author: Wolfgang Wick, MD, Department of Neurology, University of Tübingen, Medical School, Hoppe-Seyler-Strasse 3, 72076 Tübingen, Germany (E-mail: [email protected])
-
Upload
khangminh22 -
Category
Documents
-
view
5 -
download
0
Transcript of P-Glycoprotein and Multidrug Resistance - NCBI

Oliver Bähr1; Johannes Rieger1; Frank Duffner2;Richard Meyermann3; Michael Weller1; WolfgangWick1
1 Laboratory of Molecular Neuro-Oncology, Department ofNeurology, 2Department of Neurosurgery, 3Institute for BrainResearch, University of Tübingen, Germany.
Understanding and overcoming multidrug resist-ance (MDR) may be a promising strategy to developmore effective pharmacotherapies for malignantgliomas. In the present study, human malignantglioma cell lines (n = 12) exhibited heterogeneousmRNA and protein expression and functional activ-ity of the mdr gene-encoded P-glycoprotein (PGP) andMDR-associated protein (MRP). Correlation betweenmRNA expression, protein levels and functionalactivity was strong. Inhibition of PGP activity by ver-apamil or PSC 833 enhanced the cytotoxic effects ofvincristine, doxorubicin, teniposide and taxol. Inhi-bition of MRP activity by indomethacin orprobenecid enhanced the cytotoxic effects of vin-cristine, doxorubicin and teniposide. The humancerebral endothelial cell line, SV-HCEC, exhibitedthe strongest PGP activity of all cell lines. Five primaryhuman glioblastomas and one anaplastic astrocy-toma displayed heterogenous protein levels of PGPand MRP-1 in tumor cells and of PGP in biopsyspecimens in vivo, but no functional activity ofthese proteins upon ex vivo culturing. These data sug-gest that the glioma cell line-associated MDR-typedrug resistance is a result of long-term culturingand that cerebral endothelial, but not glioma cells,may contribute to MDR-type drug resistance ofgliomas in vivo.
IntroductionMultidrug resistance resulting from the expression of
the mdr-1 and mrp-1 gene products is one of the majorobstacles to successful chemotherapy in several types ofcancer (6, 7, 15). The corresponding proteins, PGP andMRP, belong to the family of ATP binding cassette(ABC) transporters which function as ATP-dependent
efflux pumps. Amongst others, PGP and MRP transportvarious common cancer chemotherapeutics.
Malignant gliomas, notably glioblastomas, are large-ly resistant to current treatment strategies which includecytoreductive surgery, involved-field radiotherapy andadjuvant chemotherapy. Less than 30% of patients withglioblastoma benefit from adjuvant chemotherapy (9, 14).It would be a great clinical advance if these patientscould be identified prospectively by examining specif-ic biological parameters. We have previously reported thatseveral candidate genes for drug sensitivity, includingp53, p16, cyclin-dependent kinase (CDK)-4, murinedouble minute (MDM)-2 and various B cell lymphoma(BCL)-2 family proteins fail to predict response tochemotherapy in human malignant glioma cell lines(30).
The clinical significance of the MDR phenotype inglioblastoma has remained controversial (2). Immuno-histochemical analyses of human glioma sections indi-cate that both the tumor cells proper and the endothelialcells express MDR in human gliomas (2, 4, 8, 16, 19, 21,28, 29). The role of endothelial cells has also beenstrengthened by an in vivo xenograft study (27). Further,disruption of the mdr-1a gene in mice resulted inenhanced vinblastine accumulation in the normal brain(25). The effects of adjuvant cytotoxic therapies on theMDR phenotype in gliomas in vivo have been examinedin 2 immunohistochemical studies: a Japanese studyobserved an increase in MRP/PGP expression afterchemotherapy (2) whereas no such effect was observedin our own series (23). Few studies have addressed thefunctional activity and the drug specificity of MDR andMRP in glioma cells (1, 10, 18, 23, 24). High levels ofmrp-1 mRNA expression in glioma cell lines werelinked to reduced etoposide accumulation and to resist-ance to etoposide, vincristine and doxorubicin (1).Kiwit and colleagues observed that the calcium antagonist,verapamil, enhanced the sensitivity of PGP-expressingglioma cells to ACNU, cisplatin and vincristine (18). Wepreviously noted that the chemosensitizing properties of
RESEARCH ARTICLE
P-Glycoprotein and Multidrug Resistance-associated Protein Mediate Specific Patterns ofMultidrug Resistance in Malignant Glioma CellLines, but not in Primary Glioma Cells
Corresponding author: Wolfgang Wick, MD, Department of Neurology, University of Tübingen, Medical School, Hoppe-Seyler-Strasse 3, 72076 Tübingen,Germany (E-mail: [email protected])

483Multidrug Resistance in Human Malignant Glioma—Bähr et al
the cyclooxygenase inhibitor, indomethacin, were notrelated to the inhibition of cyclooxygenase, but to theMRP-inhibitory properties of this drug (24). Further, weobserved that human glioma cell lines exhibited a vera-pamil-sensitive PGP transport activity in vitro, but, cor-responding to our analysis of paired pre- and post-radiochemotherapy clinical glioma specimens, chronicdrug exposure did not produce major alterations in theconstitutive MDR phenotype in vitro (23).
Here, we performed a detailed study on the expressionand functional activity of MDR-associated proteins inglioma cell lines and examined their role in drug resist-ance, studying a panel of cancer chemotherapeuticswhich are commonly administered to glioma patients orwhich are currently evaluated for efficacy in that disease.Further, given the probable role of endothelial cells inmediating MDR in malignant gliomas (5, 21, 26), we per-formed these experiments with the human cerebralendothelial cell line SV-HCEC in parallel (20). It has beendemonstrated that endothelial cells from experimentalglioma differ with respect to MDR expression and met-alloproteinase-9 activity from regular brain vasculature(22). To assess the clinical relevance of PGP/MRP-1 inthe glioma cells in vivo, we stained biopsy specimens forPGP/MRP-1, generated ex vivo (primary) cultures of thesame tumors and examined PGP/MRP-1 expressionand functional activity.
Materials and Methods
Chemicals. Cytotoxic drugs were obtained from thefollowing sources: 1-(2-chlorethyl)-3-cyclohexyl-1-nitrosourea (CCNU, lomustine) (Medac, Hamburg,Germany), topotecan (SmithKline Beecham, Munich,Germany), gemcitabine (Lilly, Bad Homburg, Ger-many), teniposide, taxol, cisplatin (Bristol, Munich,Germany), vincristine, doxorubicin (Sigma, St. Louis,Mo). Verapamil, indomethacin and probenecid werealso obtained from Sigma. PSC 833 was kindly provid-ed by Novartis (Basel, Switzerland).
Cell culture and cytotoxicity assay. The malignantglioma cell lines were kindly provided by Prof N. de Tri-bolet (Lausanne, Switzerland). The cells were main-tained in DMEM containing 10% fetal calf serum, 2mM glutamine and penicillin (100 IU/ml)/streptomycin(100 �g/ml) (30). For cytotoxicity assays, the cellswere seeded at a density of 104 cells per well in 96 wellplates, adhered for 24 hours and then coexposed toMDR or MRP inhibitors and one of several chemother-apeutic drugs for 72 hours. Survival was assessed by crys-
tal violet staining. The human cerebral endothelial cellline, SV-HCEC, was kindly provided by Dr. A. Muru-ganandam (Ottawa, Ontario) and cultured as described(20).
Primary human glioma cultures. Biopsy samples ofhuman gliomas (5 glioblastomas and one anaplasticastrocytoma-T132-) were obtained during brain tumorsurgery, and short term cultures were established fromsamples with high tumor content as assessed by mor-phological analysis of tumor sections (T57, T107,T113, T132, T140 and T159). In some samples, there wassmall contamination by reactive astrocytes andendothelial cells. Briefly, tissue was washed severaltimes with sterile PBS, transferred to a 35-mm tissue cul-ture dish and cut into small pieces using a sterilescalpel. The tissue was transferred to a 50 ml centrifugetube, enzymatically dissociated with trypsin (0.25%)and 50 �g/ml DNase for 30 minutes at 37°C, filteredthrough a 70-�m mesh and washed with DMEM medi-um containing 10% FCS. Cells were pelleted for 10minutes at 1800 rpm at room temperature, resuspendedin 10% FCS in DMEM and plated in a 75-cm2 tissue cul-ture flask. Cells from passage numbers less than 15were used for each individual assay.
Immunohistochemistry. Tumor specimens of 6patients were analyzed for the expression of PGP andMRP-1 using the DAKO CSA HRP-System(Dakopatts, Copenhagen, Denmark). Anti-PGP-mouse-MRK-16 and anti-MRP-1-rat-MRPr1 (Hölzel Diagnos-tics, Cologne, Germany) were used as primary antibod-ies. Human liver specimens served as positive controls.The fraction of immunopositive tumor cells was evalu-ated semiquantitatively according to the followingscore: -, no positive tumor cells; +, <10% positivetumor cells; ++, 10 to 50% positive tumor cells; +++, 50to 90% positive tumor cells; ++++, >90% positivetumor cells. All sections were counterstained withhemalum (magnification �200).
RT-PCR. RNA was isolated using the RNeasy Minikit (Quiagen, Hilden, Germany). First strand synthesiswas performed using Superscript II reverse transcriptase(Gibco, Gaithersburg, MD). Sense and antisense PCRprimers for mdr-1 and mrp-1 were kindly provided byJames Beck (University Children’s Hospital, Tübingen,Germany). Primers for mrp-2 ,-3, -4 and -5 wereobtained from MWG Biotech (Ebersberg, Germany).Human �-actin primers were obtained from Biosource(Fleurus, Belgium). The primer sequences were as fol-

lows: mdr-1 sense 5�-GGAGA GATCC TCACCAAGCG-3� (nucleotides 2765-2784), mdr-1 antisense 5�-GTTGC CAACC ATAGA TGAAG G-3� (nucleotides2973-2993), yielding a fragment of 229 base pairs (bp);human mrp-1 sense 5�-CGTGT ACTCC AACGCTGAC-3� (nucleotides 2540-2558), human mrp-1 anti-sense 5�-CTGGA CCGCT GACGC CCGTG AC-3�(nucleotides 2844-2865), yielding a fragment of 326bp; human cMOAT/mrp-2 sense 5�-CTGCC TCTTCAGAAT CTTAG-3� (nucleotides 4136-4155), humancMOAT/mrp-2 antisense 5�-CCCAA GTTGC AGGCTGGCC-3� (nucleotides 4358-4376), yielding a fragmentof 241 bp; human mrp-3 sense 5�-GATAC GCTCGCCACA GTCC-3� (nucleotides 3229-3247), humanmrp-3 antisense 5�-CAGTT GGCCG TGATG TGGCTG-3� (nucleotides 3425-3439), yielding a fragment of 211bp; human mrp-4 sense 5�-CCATT GAAGA TCTTCCTGG-3� (nucleotides 12-30), human mrp-4 antisense5�-GGTGT TCAAT CTGTG TGC-3� (nucleotides 233-250), yielding a fragment of 239 bp; human mrp-5sense 5�-GGATA ACTTC TCAGT GGG-3�(nucleotides 4033-4050), human mrp-5 antisense 5�-GGAAT GGCAA TGCTC TAAAG-3� (nucleotides4461-4480), yielding a fragment of 448 bp; human �-actin sense 5�-TGTTT GAGAC CTTCA ACACC C-3�(nucleotides 409-429), human �-actin antisense 5�-AGCAC TGTGT TGGCG TACAG-3� (nucleotides918-937), yielding a fragment of 529 bp. The PCR con-ditions included pre-denaturation 95°C per 5 minutes and35 cycles consisting each of 95°C per 45 seconds denat-uration, 55°C per 45 seconds annealing, 72°C per 1minute synthesis, and a post-synthesis run of 72°C per7 minutes. The PCR products were separated in 2%agarose gels and visualized by ethidium bromide stain-ing. The �-actin cDNA fragment was amplified as aninternal control for equal amplification. A water controlwas run in each amplification to control for cross-con-tamination between tubes.
Flow cytometry. The expression of PGP and MRP-1proteins was quantified by flow cytometry using thespecific monoclonal antibodies MRK-16 and MRPr1. Todetect PGP, 0.5 �106 cells were incubated with MRK-16(25 �g/ml) or with mouse IgG2a isotype control (25�g/ml) for one hour at 4°C. After washing with cold phos-phate-buffered saline (PBS)/ bovine serum albumin(0.1%), the cells were stained with a phycoerythrin-labeled goat anti-mouse IgG antibody (Sigma) foranother 30 minutes at 4°C. Since the MRPr1 antibody rec-ognizes an intracellularly exposed epitope of MRP, thecells were permeabilized prior to labelling. This was
done by incubating the cells with 100% methanol at -20°Cfor 15 minutes (11). The cells were incubated with thespecific MRPr1 antibody (2 �g/ml) or the correspondingisotype antibody (rat IgG2a, 2 �g/ml) for one hour,washed and stained with fluorescein isothiocyanate-labeled anti-rat IgG antibody (Sigma). Fluorescenceintensity was measured using a Becton Dickinson FAC-SCalibur (Heidelberg, Germany). The specific fluores-cence index (SFI) was calculated by dividing the meanfluorescence intensity of cells stained with the specificantibody by the mean fluorescence intensity obtainedusing an isotype-matched control antibody (equal con-centrations).
Rhodamine-123 efflux assay. PGP functional activ-ity was assessed by measuring efflux of the fluorescentprobe, rhodamine-123. Rhodamine-123 is a specificPGP substrate that will be actively eliminated from thecell by PGP. It is a measure for PGP function. Five � 105
cells were loaded with rhodamine-123 (200 ng/ml) forone hour at 37°C. Thereafter, cells were allowed toexport the dye for 60 or 120 minutes in the absence orpresence of the PGP inhibitor, verapamil (2 or 20 �M).Changes in fluorescence were monitored with a BectonDickinson FACScalibur. Results are expressed as ratiosof mean fluorescences (rhodamine-123 plus verapamilover rhodamine-123 alone). PSC 833 had similareffects as verapamil in this assay whereas indomethacinor probenecid had no effect (data not shown).
Fluorescein accumulation assay. MRP functionalactivity was assessed by measuring cellular accumula-tion of the fluorescent dye, fluorescein, in the absence orpresence of indomethacin (17). Fluorescein is a specif-ic MRP substrate that will be actively eliminated fromthe cell by MRP. It is a measure for MRP function. Thecells were seeded on 24 well plates and allowed toattach for 24 hours. Fluorescein (100 �M) was added tothe confluent monolayers alone or in the presence of theMRP inhibitor, indomethacin (4, 20 or 100 �M). Aftera 2-hour incubation period at 37°C, dye and inhibitor wereremoved, and the cells were washed 3 times with ice-coldPBS. To determine fluorescence, cells were lysed inlysis buffer (10 mM TRIS-HCl, 10 mM EDTA, 0.2 % Tri-ton X-100, pH = 7.5) for 20 minutes on ice and analyzedusing a CytoFluor 2350 plate reader (Millipore, Bedford,Mass) at 485 nm excitation and 530 nm emmissionwavelengths. Raw data were absolute fluorescence val-ues. Data were expressed, after background subtraction,as ratios of fluorescence in the presence versus theabsence of indomethacin. The PGP inhibitors verapamil
484 Multidrug Resistance in Human Malignant Glioma—Bähr et al

and PSC 833 had no effect in this assay whereasprobenecid had similar effects as indomethacin (datanot shown).
Statistical analysis. PGP and MRP mRNA expression,PGP and MRP protein levels, and PGP and MRP func-tional activity measured by flow cytometry were corre-lated by ANOVA. PGP and MRP mRNA expression andp53 status, p16/CDK-4 status, PTEN status or BCL-2family protein expression were correlated by Spearmanrank order correlation. PGP and MRP protein levels andfunctional activity and ratios of effective concentration(EC)50 values were correlated by Pearson product-moment correlation. The overall effect on an inhibitor onEC50 values of cytotoxic drugs on all cell lines wasdetermined by comparing EC50 values in the absence ofinhibitor and EC50 values in the presence of inhibitor usinga paired t-test.
Results
Characterization of the mdr/mrp phenotype inhuman malignant glioma cell lines and primaryhuman glioma cultures. Mdr-1 and mrp mRNAexpression were assessed by RT-PCR (Figure 1A). LN-18, T98G, LN-229 and LN-308 showed strong mdr-1mRNA expression whereas LN-428 and U373MGshowed faint mdr-1 mRNA expression. Mdr-1 mRNAlevels in U138MG, U87MG, D247MG, LN-319, A172and U251MG cells were below detection limit. T98G,LN-319, LN-229, A172 and LN-308 cells showedstrong mrp-1 mRNA expression. Mrp-1 mRNA expres-sion was moderate in LN-428, D247MG, U251MG andU373MG cells and low in LN-18, U138MG andU87MG cells. mRNA for mrp-2, mrp-3, mrp-4 andmrp-5 was detected in 4, 10, 6 and 10 of the 12 cell lines,respectively.
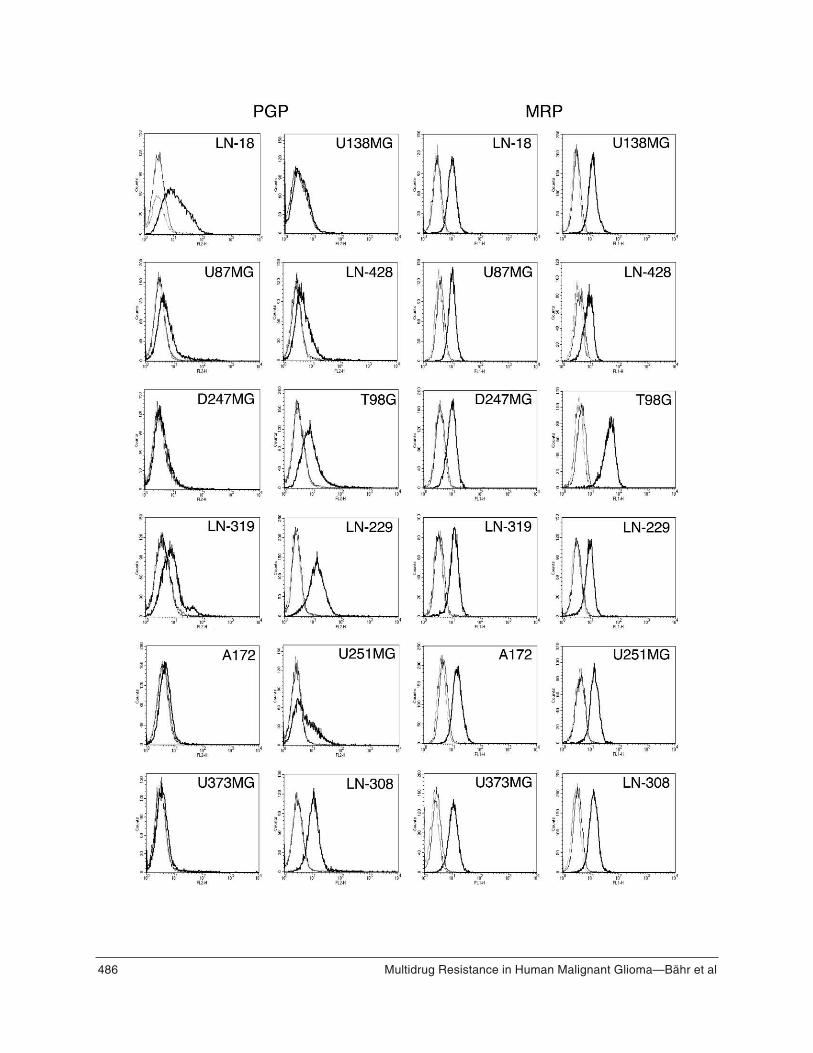
As predicted from the mRNA data, PGP proteinexpression assessed by flow cytometry was strongest inthe LN-18, T98G, LN-229 and LN-308 cell lines (Fig-ure 1B). Flow cytometry as performed here was rathersensitive in that it picked up some signal in cell lineswhere the mRNA levels measured by RT-PCR werebelow detection limit. Alternatively, the PGP antibodymay recognize a second protein other than PGP but thisis unlikely because of the good overall correlationbetween the RT-PCR and flow cytometry data for PGP.Representative flow cytometry profiles for each cellline are shown in Figure 2. As for PGP, there was alsogood correlation between mrp-1 mRNA expression(Figure 1A) and MRP-1 protein expression (Figures
485Multidrug Resistance in Human Malignant Glioma—Bähr et al
Figure 1. Mdr-1/PGP and MRP expression. A. The expressionof mdr-1, mrp-1, -2, -3, -4, -5 or �-actin mRNA was analysed byRT-PCR. The predicted fragment sizes were 229 bp for mdr-1,326 bp for mrp-1, 241 bp for mrp-2, 211 bp for mrp-3, 239 bp formrp-4, 448 bp for mrp-5 and 529 bp for �-actin. B. PGP (openbars) and MRP (grey bars) protein levels were analysed byflow cytometry. Data are expressed as SFI values and SD (n =3).
486 Multidrug Resistance in Human Malignant Glioma—Bähr et al

1B, 2). Note that the RT-PCR as performed here was per-formed chiefly to detect the presence of mRNA, not toquantify mRNA expression. For instance, the mrp-1mRNA signal for U87MG was commonly low althoughflow cytometry and transport activity (see below)always demonstrated MRP activity. Moreover, noexperimental tools to define the impact of the mrp-2-5-encoded genes on the transport activities measured here(see below, Figure 4) are available at present.
In addition to the analyses of established long termglioma cell lines, PGP and MRP-1 protein expressionwere assessed in 6 ex vivo primary human malignantglioma cultures. First, immunohistochemistry of thebiopsy specimens of the respective tumors for PGPrevealed staining patterns of +++ in T57 and T107, ++in T113, + in T132 and T140 and - in T159 (Figure 3A).Tumor endothelial cells were also positive (++) forPGP. Immunohistochemistry for MRP-1 failed to detectspecific signals in the glioma biopsies and liver sec-tions that should serve as positive controls, indicating thatthe available antibody (MRK-16) is not suitable forimmunohistochemistry (data not shown). Flow cytom-etry detected a strong signal for PGP in T57, T107 andT113 and for MRP-1 in T57, T107, T113, T132 andT140 whereas T159 was negative for both PGP andMRP-1, and T132 and T140 were weakly positive forPGP (Figure 3B).
The functional activities of PGP and MRP wereassessed by previously established substrate transportassays (23, 24). Figure 4 provides representative exam-ples for the measurement of PGP (Figure 4A) and MRP(Figure 4B) activity. For illustration, we show a PGP-highcell line (A, LN-18) and a MRP-high cell line (B,T98G). The transport data on all 12 glioma cell lines aresummarized in Figure 4C. Similarly, transport activitiesfor the 6 primary cultures were analysed. Except for
487Multidrug Resistance in Human Malignant Glioma—Bähr et al
Figure 2. (Opposing page) PGP and MRP-1 protein expression.All cell lines were analyzed for PGP and MRP-1 proteinexpression by flow cytometry. Representative profiles areshown. Bold lines are profiles obtained with the specific anti-bodies, thin lines represent the signals obtained with the isotypecontrol antibodies, dotted lines represent unstained cells.
Figure 3. PGP and MRP-1 protein expression in primary ex vivoglioma cell cultures. A. Biopsy specimens of five glioblastomaand one anaplastic astrocytoma (T132) were stained for PGP.Specific immunoreactivity is recognized as the brown color,blue color represents the counterstain magnification �200). B.Primary cultures of the gliomas shown in A were analyzed forPGP (open bars) and MRP-1 (grey bars) protein expression byflow cytometry. Data are expressed as SFI values and SD (n =3).

488 Multidrug Resistance in Human Malignant Glioma—Bähr et al
Vin
cris
tin
e [n
M]
Do
xoru
bic
in [
nM
]Te
nip
osi
de
[nM
]Ta
xol [
nM
]C
isp
lati
n [
µM]
Top
ote
can
[n
M]
Gem
cita
bin
e [n
M]
CC
NU
[n
M]
LN-1
875
±7
140
±17
580
±43
230
±18
4.2
±0.
224
0 ±
221
±4
710
±80
+ V
PM
2 ±0
.2*
42 ±
3*18
0 ±
13*
31 ±
2*3.
1 ±
0.3*
260
±2*
22 ±
559
0 ±
75+
Indo
63 ±
889
±8*
390
±41
*23
0 ±
173.
8 ±
0.1
200
±2*
21 ±
264
0 ±
72
U13
8MG
50 ±
417
0 ±
1254
0 ±
4023
0 ±
235.
2 ±
0.2
84 ±
812
00 ±
8340
0 ±
40+
VP
M22
±2*
73 ±
6*39
0 ±
32*
260
±12
5.1
±0.
510
2 ±
1019
00 ±
420*
420
±44
+ In
do25
±2*
80 ±
5*39
0 ±
39*
211
±19
5.1
±0.
957
±4*
1200
±16
039
0 ±
56
U87
MG
28 ±
319
0 ±
1684
0 ±
7684
±9
15 ±
1.5
78 ±
518
±3
740
±36
+ V
PM
10 ±
1*80
±9*
640
±45
*85
±8
19 ±
2.1*
87 ±
620
±2
860
±66
+ In
do12
±2*
68 ±
8*38
0 ±
29*
83 ±
716
±2.
079
±4
18 ±
274
0 ±
89
LN-4
2828
±3
270
±3
550
±46
88 ±
1010
±0.
930
0 ±
328
±4
590
±62
+ V
PM
6 ±
0.5*
170
±19
*39
0 ±
23*
68 ±
5*9
±0.
632
0 ±
2*32
±3
490
±31
+ In
do25
±3
240
±22
500
±25
92 ±
119.
9 ±
0.5
280
±3*
29 ±
541
0 ±
40*
D24
7MG
45 ±
329
0 ±
2776
00 ±
530
67 ±
719
±0.
970
00 ±
650
1300
0 ±
900
530
±40
+ V
PM
19 ±
2*23
0 ±
22*
9500
±56
0*68
±7
21 ±
1.0
6600
±51
015
000
±20
0059
0 ±
69+
Indo
37 ±
416
0 ±
19*
8000
±70
066
±7
20 ±
1.8
6700
±54
012
000
±15
0055
0 ±
64
T98
G63
±6
430
±32
1100
±97
44 ±
352
±5.
630
0 ±
2463
±10
1000
±99
+ V
PM
3 ±
3*74
±5*
290
±22
*36
±4
45 ±
3.4
230
±16
*75
±12
1100
±10
6+
Indo
12 ±
2*21
0 ±
24*
510
±49
*42
±3
55 ±
4.7
910
±15
0*85
±10
1000
±88
LN-3
1931
±4
375
±40
1000
±11
016
0 ±
189
±0.
565
00 ±
530
9300
±88
055
0 ±
48+
VP
M3
±0.
4*22
0 ±
18*
660
±88
*10
7 ±
10*
6.7
±0.
3*72
00 ±
630
9300
±12
0054
0 ±
57+
Indo
21 ±
2*29
0 ±
13*
950
±77
130
±15
9.7
±0.
766
00 ±
460
1200
0 ±
1600
570
±31
LN-2
2921
±2
230
±16
1400
±13
010
0 ±
86
±0.
275
±6
20 ±
128
0 ±
31+
VP
M1
±0.
3*14
0 ±
16*
1400
±13
043
±5*
10 ±
1.1*
86 ±
662
±30
250
±40
+ In
do17
±2
130
±13
*76
0 ±
56*
97 ±
98.
2 ±
0.9*
75 ±
720
±5
260
±37
A17
230
±4
120
±9
1500
±14
076
±5
26 ±
2.1
190
±19
69 ±
1050
0 ±
41+
VP
M12
±1*
56 ±
5*13
00 ±
170
81 ±
426
±1.
922
0 ±
2686
±16
500
±50
+ In
do16
±2*
58 ±
8*80
0 ±
59*
58 ±
4*30
±2.
515
0 ±
1258
±8
500
±50
U25
1MG
22 ±
184
±8
250
±21
40 ±
31.
3 ±
0.2
20 ±
218
±3
120
±10
+ V
PM
6 ±
1*42
±6*
120
±9*
31 ±
2*1.
2 ±
0.1
19 ±
217
±2
180
±22
*+
Indo
8 ±
1*29
±4*
140
±10
*40
±2
1.1
±0.
120
±2
21 ±
214
0 ±
14
U37
3MG
34 ±
318
0 ±
1843
0 ±
4314
0 ±
101.
5 ±
0.2
37 ±
253
0 ±
6147
0 ±
79+
VP
M5
±1*
68 ±
5*31
0 ±
23*
140
±11
1.5
±0.
138
±4
1700
±52
0*48
0 ±
79+
Indo
14 ±
2*98
±10
*33
0 ±
23*
160
±12
1.5
±0.
236
±4
590
±79
500
±56
LN-3
0814
0 ±
957
0 ±
612
00 ±
100
620
±45
14 ±
1.5
1300
±95
550
±50
330
±56
+ V
PM
7 ±
1*16
0 ±
13*
540
±47
*11
0 ±
7*15
±1.
312
00 ±
100
2000
±69
0*29
0 ±
21+
Indo
89 ±
9*44
0 ±
27*
540
±29
*61
0 ±
4913
±1.
187
0 ±
9657
0 ±
6333
0 ±
28
Tab
le 1
.Pat
tern
s of
dru
g se
nsiti
vity
in th
e ab
senc
e or
pre
senc
e of
PG
Por
MR
Pan
tago
nist
s.T
he c
ells
wer
e tr
eate
d as
in F
igur
e 5.
EC
50va
lues
for
each
dru
g w
ere
deriv
edin
the
abse
nce
of a
res
pons
e m
odifi
er (
naiv
e) o
r in
the
pres
ence
of v
erap
amil
(VP
M, 2
0 �
M)
or in
dom
etha
cin
(Ind
o, 2
0 �
M)
(mea
n an
d S
D, n
=3,
*p<
0.05
, t-t
est,
com
pare
dw
ith c
ytot
oxic
dru
g tr
eatm
ent o
nly)
.

T132 which showed medium MRP-1 activity, the primarycells showed weak MRP-1 activity and no PGP activity(Figure 4D). Hence, in the glioma cell lines there wasstrong correlation between PGP protein expression andactivity (r = 0.78, p = 0.003) and MRP protein expres-sion and activity (r = 0.78, p = 0.003). In contrast, PGP orMRP-1 expression in vivo and ex vivo did not translateinto PGP or MRP-1 activity in the primary cultures(Figures 3A, B, 4D). Further, there was no correlationbetween PGP or MRP-1 expression and activity (p =0.64) or PGP or MRP-1 immunohistochemical stainingand transport activity. There was no correlation amongthe cell lines between i) mdr and mrp-1 mRNA expres-sion (Figure 1A), ii) PGP and MRP-1 protein expression(Figures 1B, 2), or iii) PGP- and MRP-1 transport activ-ity (Figure 4).
Modulation of drug cytotoxicity by PGP or MRPantagonists. To assess the functional role of PGP andMRP activity, we treated the glioma cells with increas-ing concentrations of various cancer chemotherapeuticsin the absence or presence of the PGP inhibitors, vera-pamil or PSC 833, or the MRP inhibitors, indomethacinor probenecid, and determined changes in drug-inducedcell death. Typical concentration response curves areshown in Figure 5. Such data were then used to deriveEC50 values for drug cytotoxicity in all cell lines in theabsence or presence of PGP or MRP inhibitors (Table 1).A paired t-test analysis of the overall influence of vera-pamil on drug sensitivity on all cell lines examinedrevealed a significant reduction in EC50 values by vera-pamil for vincristine (p = 0.003) and doxorubicin (p =0.002). Sensitizing effects were also seen for tenipo-
side and taxol in many cell lines but these effects failedto achieve statistical significance in the overall analysisof all cell lines. Verapamil (Table 1) or PSC 833 (data notshown) had no consistent effect on the cytotoxicity oftopotecan, cisplatin, gemcitabine or CCNU. In fact, sin-gle cell lines were even moderately protected from thecytotoxicity of these drugs by verapamil. There wasgood correlation between PGP expression and activity ina given cell line and the sensitivity to verapamil-
489Multidrug Resistance in Human Malignant Glioma—Bähr et al
Figure 4. PGP and MRP transport activity. A. LN-18 cells wereexamined for PGP activity in the rhodamine-123 expulsionassay (2 h/37°C). Bold lines indicate fluorescence (rhodamine)retention in the presence of verapamil, thin lines (middle in A)show fluorescence retention in the absence of verapamil. Theprofile on the left represents the autofluorescence of the cells.B. T98G cells were examined for MRP function using the fluo-rescein accumulation assay (2 h/37°C). The bars represent theintracellular fluorescence (mean and SD, n=3) of unstained cells,cells treated with fluorescein alone (control) or cells whichadditionally received the MRP inhibitor, indomethacin, at 4, 20or 100 �M. C. Synopsis of PGP and MRP transport activitydetermined as in A and B. Data are expressed as mean ratiosand SD (n = 3) of fluorescence in the presence versus absenceof inhibitor (open bars, PGP, using verapamil at 20 �M; grey bars,MRP, using indomethacin at 100 �M). A value of 1 indicates theabsence of PGP or MRP activity. D. PGP and MRP transportactivities in primary cultures of 5 human glioblastoma and oneanaplastic astrocytoma were examined as in A-C. Data areexpressed as in C.

enhanced augmentation of drug cytotoxicity, especiallyfor vincristine (Table 2).
A paired t-test analysis for MRP inhibition revealedthat indomethacin enhanced glioma cell sensitivity to vin-cristine (p = 0.002), doxorubicin (p = 0.00004) and teni-poside (p = 0.01). The effects of taxol, cisplatin, topote-
can, gemcitabine and CCNU were essentially unaffect-ed by indomethacin. The correlation between MRPexpression and activity and the sensitizing effect ofindomethacin is summarized in Table 2. Probenecid hadsimilar effects on drug sensitivity as indomethacin butproved to be rather toxic on its own in the glioma cells(data not shown).
If PGP or MRP were the most important determi-nants for the differential sensitivity of glioma cell linesto a given drug, the EC50 values in the presence of ver-apamil or indomethacin should be more homogeneousthan in the absence of the inhibitors. However, this wasnot the case for vincristine, doxorubicin, teniposide ortaxol. For instance, the range of EC50 values for vin-cristine in the absence of verapamil was 0.021 to 0.143�M and the ratio of highest versus lowest EC50 value was6.8. In the presence of verapamil, the range of EC50 val-ues was 0.0013 to 0.022 �M and the ratio of highest ver-sus lowest EC50 assumed a value of 17. Thus, althoughthe EC50 values were shifted towards a lower range,they were not more homogeneous in the presence com-pared with the absence of verapamil.
Similarly, we treated the primary cultures withincreasing concentrations of vincristine, doxorubicin,
490 Multidrug Resistance in Human Malignant Glioma—Bähr et al
Vincristine Doxorubicin Teniposide Taxol
PGP 0.83* 0.29 0.41 0.73*expression
PGP 0.92* 0.44 0.62* 0.96*activity
MRP 0.89* 0.14 0.28 0.19expression
MRP 0.78* 0.64* 0.46 0.27activity
Table 2. Correlation between PGP and MRP protein expressionand transport activity with the modulation of drug cytotoxicity byverapamil and indomethacin. A ratio of EC50 with inhibitor overEC50 without inhibitor was determined for each cell line and eachdrug, to quantify the degree of sensitization. PGP and MRP pro-tein levels and transport activity were correlated with theseratios (*p<0.05, Pearson product-moment correlation).
Figure 5. Modulation of drug cytotoxicity by PGP or MRPantagonists. LN-18, U87MG, T98G and LN-229 cells weretreated with vincristine in the absence (open circles) or presenceof verapamil (VPM) at 2 �M (filled squares) or 20 �M (filled rhom-boids) (A, B) or in the absence (open circles) or presence ofindomethacin (Indo) at 4 �M (filled squares) or 20 �M (filledrhomboids) for 72 hours. Survival was assessed by crystal vio-let staining. Data are expressed as mean percentages of sur-vival and SEM (n = 3).

teniposide and taxol in the absence or presence of ver-apamil or indomethacin, and determined changes indrug-induced cell death. As expected from the substratetransport assays the toxicity of neither drug wasenhanced by verapamil or indomethacin in any of the pri-mary cultures (data not shown).
Eventually, we correlated the data on multidrugresistance reported here with our previously publisheddata on apoptosis-regulatory genes in the same celllines (30, 31). Specifically, we asked whether PGP orMRP expression or activity as shown in Figures 1B and4C and the modulation of drug cytotoxicity by vera-pamil or indomethacin showed a relation to genetic orfunctional p53 status, p16/CDK-4 status, PTEN status orBCL-2 family protein expression. These data wereoverall negative except for an association of geneticwild-type PTEN status with PGP protein expression inthat SFI values were higher for wild-type cell lines(3.42 ± 1.46) than mutant cell lines (1.95 ± 0.73) (p =0.04). In the absence of a link of PTEN status to PGPactivity or the effects of verapamil on drug sensitivity,this association may have arisen by chance and isunlikely to be of biological significance.
Multidrug resistance of human cerebral endothelialcells. SV-HCEC cells were used as an in vitro model toassess a contribution of blood-brain barrier endothelialcells to the MDR phenotype of gliomas in vivo. RT-PCR revealed that SV-HCEC cells expressed mdr-1,mrp-1, mrp-2, mrp-4 but no mrp-3 or mrp-5 mRNA(data not shown). Flow cytometry showed moderatePGP and MRP expression (Figure 6A,B). In contrast, PGPactivity assessed by rhodamine-123 transport achieveda value of 4.3 and was thus higher than in any of the 12glioma cell lines (Figure 6C). MRP activity was in theupper range of the values observed in the glioma cell lines(Figure 6D). As expected, coexposure to verapamil sig-nificantly enhanced the cytotoxic effects of vincristine,doxorubicin and teniposide, but not of the other drugs,and indomethacin failed to modulate drug cytotoxicityin SV-HCEC cells (data not shown).
Discussion
491Multidrug Resistance in Human Malignant Glioma—Bähr et al
Figure 6. Human cerebral endothelial cells exhibit strong PGPactivity. SV-HCEC cells were analysed for PGP (A) or MRP (B)expression by flow cytometry as in Figure 2 and for PGP (C) orMRP (D) transport activity as in Figure 4. The SFI values cal-culated as in Figure 1B were 2.13 for PGP and 2.04 for MRP andtransport indexes (compare with Figure 4C) were 4.3 for PGPand 2.12 for MRP.

The molecular mechanisms of chemoresistance inhuman malignant glioma have not been elucidated. Noparameter has been demonstrated to identify the subsetof glioma patients that will benefit from chemotherapy.Although a subject of cancer research for many years, thepossible role of MDR in human gliomas remains poor-ly defined. It is yet undefined whether glioma cellsproper or endothelial cells or both represent the mainsource of MDR in gliomas. The present work providesa systematic analysis of multidrug resistance in a largepanel of human malignant glioma cell lines and the firstanalysis of multidrug resistance in paired in vivo tissuesand primary ex vivo glioma cell cultures. The expressionof mdr-1/PGP and mrp-1/MRP-1 was studied at mRNAand protein level by RT-PCR and flow cytometry (Fig-ures 1 and 2) and at a functional level by transport (Fig-ure 4) and PGP/MRP inhibitor-enhanced chemotherapyassays (Figure 5; Table 1). We find that most humanmalignant glioma cells exhibit PGP or MRP activity orboth (Figures 1, 2, 4) and respond to inhibition ofPGP/MRP activity with enhanced sensitivity to specif-ic drugs (Table 1). The spectrum of drugs whose actionis potentiated by PGP or MRP-1 inhibitors in glioma cellsconfirmed previous studies in nonglial cell lines (6, 15).Of note, the augmentation of sensitivity to vincristine anddoxorubicin by PGP/MDR inhibition appears to be suf-ficiently substantial to be clinically relevant. We did notconfirm the previous observation (16) of a modulationof nitrosourea cytotoxicity by verapamil (Table 1). Fur-ther, we present novel data on the expression of themrp-2, mrp-3, mrp-4 and mrp-5 genes in glioma cells(Figure 1). At present, we do not know how the geneproducts encoded by the mrp-2-5 genes influenced theMRP-1 protein expression and activity data reportedhere. Previously the specificity of various MDR anti-bodies and therefore the significance of the MDR phe-nomenon in gliomas have been questioned (2). Thegood correlation between mRNA and protein expres-sion as well as transport activity, together with thepotent effects of pharmacological PGP/MRP inhibitionon drug sensitivity, reported here could serve as con-vincing evidence for a biological role of MDR inhuman glioma cell lines.
However, the extension of such analyses from long-term cell lines to fresh glioma tissues resulted in datastrikingly different from the cell line data. Six malignantglioma biopsies were stained for PGP and showed adiverse pattern of slight to intense staining in theglioma cells (Figure 3A). In addition, samples of thesetumors taken into culture were positive for PGP andMRP-1 in flow cytometry (Figure 3B). However, the
expression of PGP and MRP-1 did not translate intofunctional activity in the transport assays. Consequent-ly, the effects of cytotoxic drugs were not potentiated byPGP or MRP-1 inhibitors in these primary cells. Weassume that as yet unknown negative regulators of PGPand MRP-1 activity that are effective in primary cellseventually loose their effects during the process of longterm culturing.
In contrast to this discrepancy and consistent with ear-lier immunochemical analyses of malignant gliomaspecimens, brain endothelial cells may play an importantrole in the MDR phenotype of these tumors (2, 8, 23, 28,29). Endothelial cells within the tumors exhibit PGPstaining in the biopsy samples shown in Figure 3A, eg,in the center of T140. Both PGP and MRP transportactivity were high in a human brain endothelial cellline, SV-HCEC (Figure 6). Functionality for PGP in exvivo isolated rat brain endothelial cells has been shownrecently (13). In this work, inhibition of PGP with val-spodar (SDZ PSC 833) allowed paclitaxel to reacheffective concentrations in a nude mouse U-118MGglioma xenograft model. Therefore, brain endothelialcells may well represent the first and central barrier forthe penetration of drugs such as vincristine, doxoru-bicin or paclitaxel into malignant gliomas.
In conclusion, MDR-type drug resistance may inter-fere with the efficacy of many drugs currently exploredfor their activity in malignant gliomas in vivo. Thepresent work provides a basis to further explore thepossible perspective of enhancing response tochemotherapy by modulating PGP/MRP activity invivo. It suggests that tumor blood vessel-formingendothelial cells but not glioma cells proper will be tar-gets of such an approach. The clinical use of PGP/MRPinhibitors in human cancer patients has been limited bythe apparent failure to achieve therapeutic levels ofinhibitors in vivo in the absence of intolerable sideeffects. Novel multidrug resistance inhibitors such as thecyclosporin derivative, PSC 833 may overcome thisproblem. Of note, lessons from the mdr-1a knockoutmice tell that the exposure of human glioma patients toMDR/MRP modulators may also enhance drug deliveryto tumor-free brain, raising as yet unresolved potentialproblems of neurotoxicity (11, 23).
AcknowledgmentsThis study was supported by grants from the German
Cancer Council (Deutsche Krebshilfe, 10-1802-We 5) andthe Fortüne-Program of the University of Tübingen toWW (1020-0-0) and to MW (766-0-0).
492 Multidrug Resistance in Human Malignant Glioma—Bähr et al

References
1. Abe T, Hasegawa S, Taniguchi K, Yokomizo A, Kuwano T,Ono M, Mori T, Hori S, Kohno K, Kuwano M (1994) Pos-sible involvement of multidrug-resistance-associatedprotein (MRP) gene expression in spontaneous drugresistance to vincristine, etoposide and adriamycin inhuman glioma cells. Int J Cancer 58:860-864.
2. Abe T, Mori T, Wakabayashi Y, Nakagawa M, Cole SP,Koike K, Kuwano M, Hori S (1998) Expression of multidrugresistance protein gene in patients with glioma afterchemotherapy. J Neuro-Oncol 40:11-18.
3. Ashmore SM, Thomas DG, Darling JL (1999) Does P-glycoprotein play a role in clinical resistance of malignantastrocytoma? Anticancer Drugs 10:861-872.
4. Becker I, Becker KF, Meyermann R, Hollt V (1991) The mul-tidrug-resistance gene MDR1 is expressed in human glialtumors. Acta Neuropathol 82:516-519.
5. Borst P, Schinkel AH (1996) What have we learnt thus farfrom mice with disrupted P-glycoprotein genes? Eur JCancer 32:985-990.
6. Borst P, Evers R, Kool M, Wijnholds J (1999) The multidrugresistance protein family. Biochim Biophys Acta1461:347-357.
7. Cole SP, Bhardwaj G, Gerlach JH, Mackie JE, Grant CE,Almquist KC, Stewart AJ, Kurz EU, Duncan AM, Deeley RG(1992) Overexpression of a transporter gene in a multidrug-resistant human lung cancer cell line. Science 258:1650-1654.
8. Cordon-Cardo C, O´Brien JP, Casals D, Rittman-GrauerL, Biedler JL, Melamed MR, Bertino JR (1989) Multidrug-resistance gene (P-glycoprotein) is expressed byendothelial cells at blood-brain barrier sites. Proc NatlAcad Sci U S A 86:695-698.
9. De Angelis LM, Burger PC, Green SB, Cairncross JG(1998) Malignant glioma: who benefits from adjuvantchemotherapy? Ann Neurol 44:691-695.
10. Decleves X, Fajac A, Lehmann-Che J, Tardy M, MercierC, Hurbain I, Laplanche JL, Bernaudin JF, Scherrmann JM(2002) Molecular and functional MDR1-PGP and MRPSexpression in human glioblastoma multiforme cell lines. IntJ Cancer 98:173-180.
11. Den Boer ML, Zwaan CM, Pieters R, Kazemier KM, Rot-tier MM, Flens MJ, Scheper RJ, Veerman AJ (1997) Opti-mal immunocytochemical and flow cytometric detection ofP-gp, MRP and LRP in childhood acute lymphoblasticleukemia Leukemia 11:1078-1085.
12. Den Boer ML, Pieters R, Kazemier KM, Janka-Schaub GE,Henze G, Veerman AJ (1998) The modulating effect of PSC833, cyclosporin A, verapamil and genistein on in vitro cyto-toxicity and intracellular content of daunorubicin in child-hood acute lymphoblastic leukemia. Leukemia 12:912-920.
13. Fellner S, Bauer B, Miller DS, Schaffrik M, Fankhänel M,Spruß T, Bernhardt G, Graeff C, Färber L, GschaidmeierH, Buschauer A, Fricker G (2002) Transport of paclitaxel(Taxol) across the blood-brain barrier in vitro and in vivo.J Clin Invest 110:1309-1318.
14. Fine HA, Dear KB, Loeffler JS, Black PM, Canellos GP(1993) Meta-analysis of radiation therapy with and with-out adjuvant chemotherapy for malignant gliomas inadults. Cancer 71:2585-2597.
15. Gottesman MM, Pastan I, Ambudkar SV (1996) P-glyco-protein and multidrug resistance. Curr Opin Genet Dev6:610-617.
16. Henson JW, Cordon-Cardo C, Posner JB (1992) P-gly-coprotein expression in brain tumors. J Neuro-Oncol14:37-43.
17. Huai-Yun H, Secrest DT, Mark KS, Carney D, BrandquistC, Elmquist WF, Miller DW (1998) Expression of mul-tidrug resistance-associated protein (MRP) in brainmicrovessel endothelial cells. Biochem Biophys ResCommun 243:816-820.
18. Kiwit JCW, Hertel A, Matuschek AE (1994) Reversal ofchemoresistance in malignant gliomas by calcium antag-onists: correlation with the expression of multidrug-resist-ant p-glycoprotein. J Neurosurg 81:587-594.
19. Matsumoto T, Tani E, Kaba K, Shindo H, Miyaji K (1991)Expression of P-glycoprotein in human glioma cell lines andsurgical glioma specimens. J Neurosurg 74:460-466.
20. Muruganandam A, Herx LM, Monette R, Durkin JP, Stan-imirovic DB (1997) Development of immortalized humancerebromicrovascular endothelial cell line as an in vitromodel of the human blood-brain barrier. FASEB J11:1187-1197.
21. Regina A, Koman A, Piciotti M, El Hafny B, Center MS,Bergmann R, Couraud PO, Roux F (1998) Mrp1 mul-tidrug resistance-associated protein and P-glycoproteinexpression in rat brain microvessel endothelial cells. JNeurochem 71:705-715.
22. Regina A, Demeule M, Berube A, Moumdjian R, BertheletF, Beliveau R (2003) Differences in multidrug resistancephenotype and matrix metalloproteinase activity betweenendothelial cells from normal brain and glioma. J Neu-rochem 84:316-324.
23. Rieger L, Rieger J, Winter S, Streffer J, Esser P, DichgansJ, Meyermann R, Weller M (2000) Evidence for a consti-tutive, verapamil-sensitive, non-P-glycoprotein multidrugresistance phenotype in malignant glioma that is unal-tered by radiochemotherapy in vivo. Acta Neuropathol99:555-562.
24. Roller A, Bähr O, Streffer J, Winter S, Heneka M,Deininger M, Meyermann R, Naumann U, Gulbins E,Weller M (1999) Selective potentiation of drug cytotoxic-ity by NSAID in human glioma cells: the role of COX-1 andMRP. Biochem Biophys Res Commun 259:600-605.
25. Schinkel AH, Smit JJ, van Tellingen O, Beijnen JH,Wagenaar E, van Deemter L, Mol CA, van der Valk MA,Robanus-Maandag EC, te Riele HP (1994) Disruption ofthe mouse mdr1a P-glycoprotein gene leads to a deficiencyin the blood-brain barrier and to increased sensitivity todrugs. Cell 77:491-502.
493Multidrug Resistance in Human Malignant Glioma—Bähr et al

26. Schinkel AH, Wagenaar E, Mol CA, Mol CA, Borst P(1996) P-glycoprotein in the blood-brain barrier of miceinfluences the brain penetration and pharmacologicalactivity of many drugs. J Clin Invest 97:2517-2524.
27. Takamiya Y, Abe Y, Tanaka Y, Tsugu A, Kazuno M, Oshi-ka Y, Maruo K, Ohnishi Y, Sato O, Yamazaki H, Kijima H,Ueyama Y, Tamaoki N, Nakamura M (1997) Murine P-gly-coprotein on stromal vessels mediates multidrug resistancein intracerebral human glioma xenografts. Br J Cancer76:445-450.
28. Tanaka Y, Abe T, Tsugu A, Takamiya Y, Akatsuka A, Tsu-ruo T, Yamazaki H, Ueyama Y, Sato O, Tamaoki N (1994)Ultrastructural localization of P-glycoprotein on capillaryendothelial cells in human gliomas. Virchows Arch425:133-138.
29. Tóth K, Vaughan MM, Peress NS, Slocum HK, Rustum YM(1996) MDR1 P-glycoprotein is expressed by endothelialcells of newly formed capillaries in human gliomas but isnot expressed in the neovasculature of other primarytumors. Am J Pathol 149:853-858.
30. Weller M, Rieger J, Grimmel C, Van Meir EG, De TriboletN, Krajewski S, Reed JC, von Deimling A, Dichgans J(1998) Predicting chemoresistance in human malignantglioma cells: the role of molecular genetic analyses. Int JCancer 79:640-644.
31. Wick W, Furnari FB, Naumann U, Cavenee WK, Weller M(1999) PTEN gene transfer in human malignant glioma:sensitization to irradiation and CD95L-induced apopto-sis. Oncogene 18:3936-3943.
494 Multidrug Resistance in Human Malignant Glioma—Bähr et al




















