Specific Fibrinogen and Thrombin Concentrations Promote Neuronal Rather Than Glial Growth When...
14
Specific Fibrinogen and Thrombin Concentrations Promote Neuronal Rather Than Glial Growth When Primary Neural Cells Are Seeded Within Plasma-Derived Fibrin Gels Rachael Mooney, Ph.D., 1 Bill Tawil, Ph.D., 2 and Melissa Mahoney, Ph.D. 3 Fibrin gels are attractive scaffolds useful for neural tissue engineering applications. The objective of this work was to investigate the apoptotic activity, survival, proliferation, and differentiation of a mixed population of primary neural cells composed of neurons and multipotent precursor cells when cultured in fibrin gels prepared with varying concentrations of fibrinogen (5–25 mg=mL fibrinogen) and thrombin (1–125 U=mL thrombin). Within all fibrin gel formulations tested, the level of apoptosis on day 1 was low and cell survival was equivalent to levels in monolayer culture (67%). Proliferation in gels made from 5 to 12.5 mg=mL fibrinogen was also similar to that observed in monolayer culture, though a lower proliferative response was observed in 25 mg=mL fi- brinogen formulations. Relative to monolayer culture, cholinergic and dopaminergic neuronal presence was enhanced, whereas glial cell growth was reduced in fibrin gel cultures. The extent to which levels were altered depended on fibrinogen and thrombin concentration. The findings here suggest the importance of fibrinogen and thrombin concentration in differentially regulating the growth and composition of neural cell populations and are of importance for neural tissue engineering strategies focused on the development of implantable scaffolds for treating neurodegenerative disorders. Introduction F ibrin gels are an attractive material for three-dimen- sional (3D) tissue engineering scaffolds because they contain both adhesive and proteolytic sequences that enable entrapped cells to remodel their environment as new tissue is formed. 1 In addition, cell viability is generally high in gels prepared from a wide range of fibrinogen and thrombin concentrations (5–50 mg=mL fibrinogen and 1–250 U=mL thrombin). 2,3 Although cell survival is relatively unaffected by different fibrin formulations, cell proliferation, cell elon- gation, and cell differentiation appear to be influenced by the concentrations of fibrinogen and thrombin. 2–4 In general, cell proliferation is higher in gels prepared with low fibrinogen concentrations. 3,4 It is also commonly observed that cells will grow in either a rounded or an elongated morphology de- pending on whether the fibrin gel is prepared with high or low fibrinogen concentrations, respectively. 2–4 Stem cell differentiation may also be impacted by fibrinogen and thrombin concentration, as human mesenchymal stem cell differentiation has been shown to be enhanced in gels pre- pared with high levels of fibrinogen. 3 Various fibrin formu- lations may impact cell behavior via alterations in exposure to bioactive ligands and=or the changing mechanical prop- erties and structure of the fibrin network. 5 Taken together, these studies highlight the importance of understanding how a range of fibrin formulations may impact the behavior of encapsulated cells, particularly when the cell population has not yet been explored in fibrin gels. Toward this end, the focus of this work is to characterize the growth and differentiation of fetal rat cortical cells in fibrin gels prepared with different concentrations of fibrin- ogen and thrombin to facilitate the development of a fibrin- based tissue engineering therapeutic capable of improving cell transplantation therapy for the treatment of neurode- generative diseases. The fibrin used in this study is a com- mercially available sealant called Tisseel Ò , a material that has been shown to be nonimmunogenic when injected into pri- mate brains. 6 The fetal cells used in this study are isolated at a developmental stage (E14–E15) that closely correlates with the human tissue (16–20 weeks) that is typically used for human transplantation procedures. 7 This rat cell popu- lation has demonstrated curative potential in animal mod- els of neurodegenerative disorders. 8–10 The fetal tissue is composed of a mixed population of neurons and multipotent neural precursor cells (NPCs). Fibrin has previously been 1 Department of Biochemistry, University of Colorado, Boulder, Colorado. 2 Bioengineering Department, University of California at Los Angeles, Los Angeles, California. 3 Department of Chemical and Biological Engineering, University of Colorado, Boulder, Colorado. TISSUE ENGINEERING: Part A Volume 16, Number 5, 2010 ª Mary Ann Liebert, Inc. DOI: 10.1089=ten.tea.2009.0372 1607
-
Upload
cityofhope -
Category
Documents
-
view
4 -
download
0
Transcript of Specific Fibrinogen and Thrombin Concentrations Promote Neuronal Rather Than Glial Growth When...

Specific Fibrinogen and Thrombin Concentrations PromoteNeuronal Rather Than Glial Growth When Primary Neural
Cells Are Seeded Within Plasma-Derived Fibrin Gels
Rachael Mooney, Ph.D.,1 Bill Tawil, Ph.D.,2 and Melissa Mahoney, Ph.D.3
Fibrin gels are attractive scaffolds useful for neural tissue engineering applications. The objective of this workwas to investigate the apoptotic activity, survival, proliferation, and differentiation of a mixed population ofprimary neural cells composed of neurons and multipotent precursor cells when cultured in fibrin gels preparedwith varying concentrations of fibrinogen (5–25mg=mL fibrinogen) and thrombin (1–125U=mL thrombin).Within all fibrin gel formulations tested, the level of apoptosis on day 1 was low and cell survival was equivalentto levels in monolayer culture (67%). Proliferation in gels made from 5 to 12.5mg=mL fibrinogen was also similarto that observed in monolayer culture, though a lower proliferative response was observed in 25mg=mL fi-brinogen formulations. Relative to monolayer culture, cholinergic and dopaminergic neuronal presence wasenhanced, whereas glial cell growth was reduced in fibrin gel cultures. The extent to which levels were altereddepended on fibrinogen and thrombin concentration. The findings here suggest the importance of fibrinogenand thrombin concentration in differentially regulating the growth and composition of neural cell populationsand are of importance for neural tissue engineering strategies focused on the development of implantablescaffolds for treating neurodegenerative disorders.
Introduction
Fibrin gels are an attractive material for three-dimen-sional (3D) tissue engineering scaffolds because they
contain both adhesive and proteolytic sequences that enableentrapped cells to remodel their environment as new tissue isformed.1 In addition, cell viability is generally high in gelsprepared from a wide range of fibrinogen and thrombinconcentrations (5–50mg=mL fibrinogen and 1–250U=mLthrombin).2,3 Although cell survival is relatively unaffectedby different fibrin formulations, cell proliferation, cell elon-gation, and cell differentiation appear to be influenced by theconcentrations of fibrinogen and thrombin.2–4 In general, cellproliferation is higher in gels prepared with low fibrinogenconcentrations.3,4 It is also commonly observed that cells willgrow in either a rounded or an elongated morphology de-pending on whether the fibrin gel is prepared with highor low fibrinogen concentrations, respectively.2–4 Stem celldifferentiation may also be impacted by fibrinogen andthrombin concentration, as human mesenchymal stem celldifferentiation has been shown to be enhanced in gels pre-pared with high levels of fibrinogen.3 Various fibrin formu-lations may impact cell behavior via alterations in exposure
to bioactive ligands and=or the changing mechanical prop-erties and structure of the fibrin network.5 Taken together,these studies highlight the importance of understanding howa range of fibrin formulations may impact the behavior ofencapsulated cells, particularly when the cell population hasnot yet been explored in fibrin gels.
Toward this end, the focus of this work is to characterizethe growth and differentiation of fetal rat cortical cells infibrin gels prepared with different concentrations of fibrin-ogen and thrombin to facilitate the development of a fibrin-based tissue engineering therapeutic capable of improvingcell transplantation therapy for the treatment of neurode-generative diseases. The fibrin used in this study is a com-mercially available sealant called Tisseel�, a material that hasbeen shown to be nonimmunogenic when injected into pri-mate brains.6 The fetal cells used in this study are isolatedat a developmental stage (E14–E15) that closely correlateswith the human tissue (16–20 weeks) that is typically usedfor human transplantation procedures.7 This rat cell popu-lation has demonstrated curative potential in animal mod-els of neurodegenerative disorders.8–10 The fetal tissue iscomposed of a mixed population of neurons and multipotentneural precursor cells (NPCs). Fibrin has previously been
1Department of Biochemistry, University of Colorado, Boulder, Colorado.2Bioengineering Department, University of California at Los Angeles, Los Angeles, California.3Department of Chemical and Biological Engineering, University of Colorado, Boulder, Colorado.
TISSUE ENGINEERING: Part AVolume 16, Number 5, 2010ª Mary Ann Liebert, Inc.DOI: 10.1089=ten.tea.2009.0372
1607

shown to be a permissive matrix for neurons and is capableof supporting the repair of peripheral and central nerveinjuries,11–14 but the behavior of undifferentiated NPC pop-ulations in fibrin gel cultures is less widely studied. Fetal-derived NPCs are of considerable interest because they havebeen shown to integrate well with host tissue upon im-plantation within the brain, and they have the potential todifferentiate into neuronal phenotypes that die during theprogression of Alzheimer’s (cholinergic neurons) or Parkin-son’s (dopaminergic neurons) disease, though they can alsodifferentiate into astrocytes that may contribute to glial scarformation.15 Although purer sources of NPCs such as im-mortalized clones are now available, their curative potentialis still not concretely established.15
Identification of 3D culture conditions that can maximizethe growth of key neuronal phenotypes from dissociatedfetal tissue while minimizing the growth of astrocytes is apriority for the field. Mixed cortical cultures have been usedpreviously in two-dimensional fibrin culture to help eluci-date matrix properties that select for neuronal over glial cellgrowth,16 and expanding this type of study to three dimen-sions is useful for neural tissue engineering applications.There is evidence to suggest that 3D fibrin gels would besupportive of neuronal differentiation as demonstrated inrecent publications involving murine embryonic stem-cell-derived NPCs.17,18 Here we build on these promising find-ings and comprehensively explore the influence of fibrinogenand thrombin concentration on the growth and compositionof fetal cells in three dimensions. Studies were designed tocharacterize the effect of fibrinogen and thrombin concen-tration on gel properties (pore size, compressive modulus,and degradation rate) and on cellular behavior (apoptoticactivity, survival, proliferation, and cell composition) using acombination of biochemical assays and confocal microscopy.To identify potential advantages of culturing this cell pop-ulation in a fibrin gel, cell behavior in gels was also directlycompared to that in monolayer culture.
Materials and Methods
Neural cell isolation and culture
Neural tissue used for all experiments was isolated fromE14–E15 fetal rat (Charles River) forebrains and dissociatedenzymatically as previously described.19 The single-cellsuspension was encapsulated into fibrin gels or plated asmonolayer cultures as described in the section titled ‘‘Pre-paration of cell-seeded fibrin gels and monolayer cultures,’’and then cultured at 378C and 5% CO2 in serum-free mediaconsisting of 50:50 Dulbecco’s modified Eagle’s medium:F12 (MediaTech, Manassas, VA) supplemented with 1�N2(Invitrogen, Carlsbad, CA), 100U=mL penicillin–streptomycin(Hyclone, Logan, UT), 1mM L-glutamine (Invitrogen), and10 ng=mL basic fibroblast growth factor (Sigma, St. Louis,MO). The growth medium was changed every other day for1–7 days depending on the experiment.
Preparation of cell-seeded fibrin gelsand monolayer cultures
Fibrin gel culture. Cells were encapsulated within fibringels by mixing two solutions in a 96-well plate, (1) 50 mL of afibrinogen solution and (2) 50mL of a thrombin-cell solution
(Tisseel; Baxter Biosciences, Deerfield, IL), where the con-centration of each component was adjusted using 1� trisbuffered saline (TBS) and 30mM CaCl2, respectively, toachieve the final gel concentrations listed in Table 1. Cellswere suspended in the thrombin solution to avoid sponta-neous polymerization of the fibrinogen solution caused bythe endogenous secretion of thrombin by neural cells.20
Upon mixing, the fibrinogen and thrombin coagulated toform a fibrin network that encapsulated the cells at a finalconcentration of 1�107 cells=mL. Polymerization was al-lowed to progress for 1 h in the incubator. One hundredmicroliters of the medium was then added, and gels wereincubated for long-term culture.
Monolayer culture. Twenty-four-well plates were coatedovernight in phosphate-buffered saline containing 4mg=mLpoly(ornithine) (Sigma). Wells were rinsed, and cells wereseeded at a final cell density of 2�105 cells=cm2.
Estimation of pore diameter and compressive modulusof fibrin gels
Pore diameter. Fibrin gels were made as describedabove (see section titled ‘‘Preparation of cell-seeded fibringels and monolayer cultures’’) except without cells and withthe additional incorporation of fibrinogen Alexafluor-488(Invitrogen; 50:1 unconjugated:conjugated fibrinogen). Fibringels were prepared on microscope slides to allow for directimaging of the network structure. Freshly prepared fibringels were imaged using the 40�water immersion objective ofa confocal microscope (Zeiss, Oberkochen, Germany) with a280 nm lateral resolution limit and a 1.2 mm axial resolutionlimit. The average pore diameter was estimated by manuallymeasuring n¼ 100 pores present in three different imagesacquired from three different gels using NIH ImageJ soft-ware (Table 1).
Compressive modulus. The compressive modulus ofn¼ 9 fibrin gels from three different experiments was de-termined 24 h after fabrication as previously described.21
Modulus was assessed after at least 24 h had elapsed basedon previous reports that the stiffness of fibrin gels can takeup to 24 h to reach a maximum.22 The compressive moduluswas measured for only one thrombin concentration (1U=mL)per fibrinogen concentration based on previous reportsshowing that increasing thrombin concentrations in therange tested in this study has a minimal effect on compres-sive modulus.23,24
Table 1. Pore Size and Modulus of Tested Fibrin Gels
Fibrinform
Fibrinogen(mg=mL)
Thrombin(U=mL)
Porediameter (mm)
Compressivemodulus (kPa)
1 5 1 9.7� 7.1 3.4� 0.52 5 62.5 5.4� 3.33 5 125 4.0� 2.04 12.5 1 8.1� 5.3 5.4� 1.15 12.5 62.5 4.7� 2.56 12.5 125 3.6� 1.47 25 1 6.4� 3.4 10.9� 1.98 25 62.5 4.0� 1.39 25 125 3.2� 0.8
1608 MOONEY ET AL.

Quantification of gel degradation
The rate of cell-mediated fibrin gel degradation wasquantified as previously described.18 Briefly, fibrin gels weremade as described above (see section titled ‘‘Preparation ofcell-seeded fibrin gels and monolayer cultures’’) except withthe additional incorporation of fibrinogen Alexafluor-488(see section titled ‘‘Estimation of pore diameter and com-pressive modulus of fibrin gels’’). The total amount offluorescently labeled degradation product released into theculture medium was quantified using a spectrofluorometer(Fluostar, Alexandria, VA). A linear standard curve is ob-tainable as long as fibrinogen concentrations greater than0.025mg=mL are being assessed. Twelve fibrin gels from twodifferent experiments were assessed. The extent of gel deg-radation is expressed as the total amount of fluorescentdegradation product released over a given time interval (1, 5,or 9 days) divided by the total amount of fluorescent mate-rial initially present in the gel as estimated by direct mea-surement of freshly made fibrin gels enzymatically digestedusing 10�trypsin. Control gels were also prepared in theabsence of cells.
Measurement of total DNA, ATP,and caspase activity in fibrin cultures
On the day of harvest, fibrin gel cultures were enzymati-cally degraded using 100 mL of collagenase (25U=mL) (Sigma)to dissolve the fibrin gels, and then 100mL of cell lysis buffer(20mM Tris [Bio-Rad, Hercules, CA], 2mM EDTA [Bio-Rad],150mMNaCl [Sigma], and 0.5% Triton X-100 [Fisher Biotech,Wembley, Australia] in diH2O)was added to lyse the cells. Allassays were carried out according to the manufacturer’s in-structions. Total DNA, ATP, and caspase levels were quanti-fied via the PicoGreen assay (Invitrogen), the Cell Glo Assay(Promega, Madison, WI), and the Apo-One assay (Promega),respectively.Whenmeasuring caspase levels, collagenasewasnot used to avoid unintentional cleavage of the assay’sfluorogenic substrate. Instead, cell lysis buffer was directlyapplied to intact fibrin gels to lyse cells. Data represent theaverage values of n¼ 16 samples collected from four differentexperiments. As a negative control and to verify assay func-tionality, an additional polyethylene glycol (PEG) hydrogelculture condition also accompanied each ATP and caspaseassay. Some degree of apoptosis is commonly observed dur-ing the first 24 h when this cell population is encapsulatedwithin a PEG gel. (For details on PEG hydrogel culture, seeMahoney and Anseth.25)
Assessment of cell composition within fibrin gels
Immunocytochemistry. On days 0, 1, and 7, fibrin gel andmonolayer samples were fixed in 4% paraformaldehyde
(Sigma). Fibrin gels were cryosectioned into 40-mm-thicksections and mounted on slides. Using standard immunocy-tochemical techniques, samples were processed for stainingwith antibodies against nestin, b-tubulin, and glial fibrillaryacidic protein (GFAP). These cell markers are commonly usedfor characterization of E14–E15 brain tissue,26 though later indevelopment GFAP and nestin are known to begin to co-labela population of radial glial cells.27 Antibodies were obtainedfrom Sigma. Secondary antibodies conjugated to Alexafluorprobes were obtained from Invitrogen. 40,6-diamidino-2-phenylindole (DAPI) was used as a nuclear counter stain.Samples were imaged via confocal microscopy using the 40�objective, and the percentage of positive nuclei was manuallydetermined in n¼ 15 images=condition=cell type collectedfrom three different experiments.
Quantitative real-time reversetranscriptase-polymerase chain reaction
Standard quantitative real-time reverse transcriptase-polymerase chain reaction (qRT-PCR) was performed to as-sess the relative expression level of cell-type-specific geneson days 0 and 7 of culture in n¼ 12 samples obtained fromthree different experiments. Primers were developed to de-tect gene expression levels for nestin, b-tubulin, choline acet-yltransferase (ChAT), tyrosine hydroxylase (TH), and GFAP.Primer sequences listed in Table 2 were obtained from In-vitrogen. Isolated RNA was DNAse treated and then accu-rately quantified using RiboGreen Quantification Reagent(Invitrogen). Equal amounts of RNA (100 ng) were placed ineach reverse transcription reaction, thus alleviating the needfor inclusion of house-keeping genes.28 cDNA synthesis andSYBRgreen PCR was carried out per the manufacturer’srecommendation (Applied Biosystems, Foster City, CA).
Characterization of cell distributionand morphology within fibrin gels
Cell morphology within the fibrin gels was observed viaconfocal microscopy after labeling live cells on day 7 ofculture with calcein-AM (Invitrogen). Each analyzed imageacquired using a 10�water immersion objective is a projec-tion of 10 optical slices acquired along the z-axis to reflect a100-mm-thick portion of the fibrin gel.
Statistical analysis
For all data, statistical significance was determined usinga two-tailed Student’s t-test. p< 0.05 is considered signifi-cant and is denoted with an asterisk unless otherwisestated. Data are presented as mean� standard error of themean.
Table 2. Quantitative RT-Polymerase Chain Reaction Primer Sequences
Cell type Gene Fwd (50?30) Rev (50?30)
NPC Nestin gtggcctctgggatgatg ttgaccttcctccccctcImmature neuron b-tubulin gtttgtgatgggtgtgaacc tcttctgagtggcagtgatgDopaminergic neuron TH cagggctgctgtcttcctac gggctgtccagtacgtcaatCholinergic neuron ChAT atggccattgacaaccatcttctg ccttgaactgcagaggtctctcatGlial cell GFAP agggacaatctcacacagg gactcaaccttcctctcca
NPC, neural precursor cell; TH, tyrosine hydroxylase; ChAT, choline acetyltransferase; GFAP, glial fibrillary acidic protein.
PRIMARY NEURAL CELL GROWTH IN PLASMA-DERIVED FIBRIN GELS 1609
Results
Fibrin gel structure
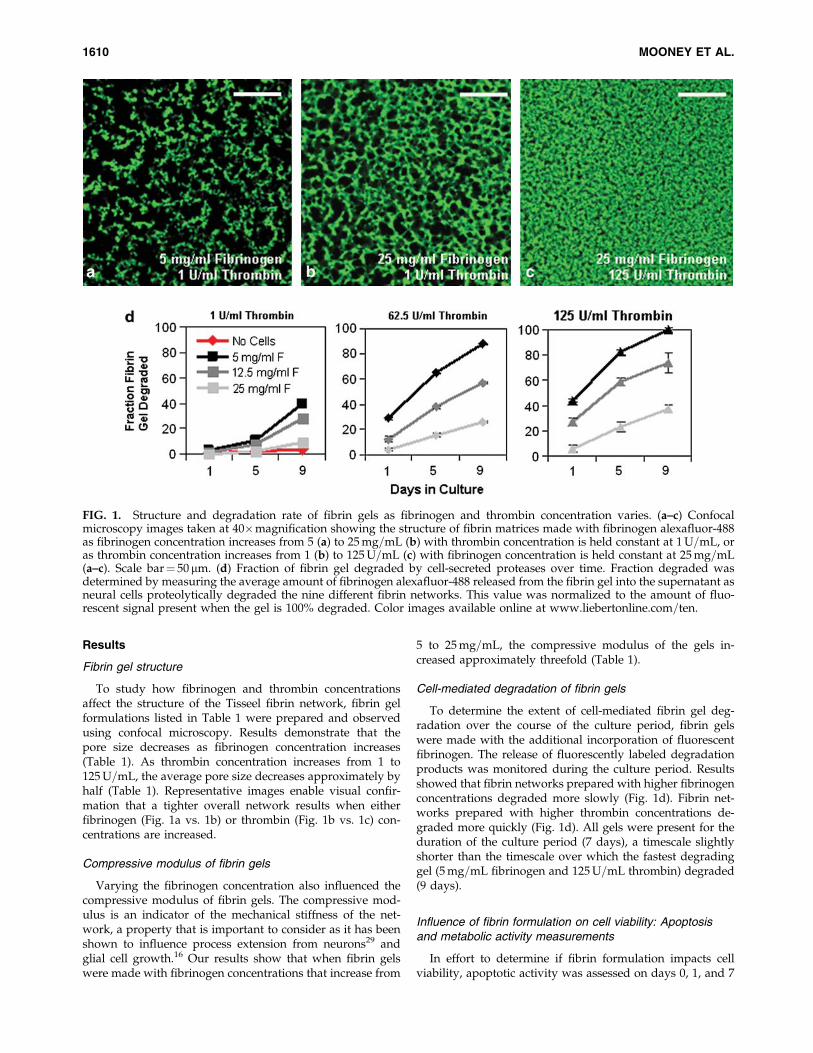
To study how fibrinogen and thrombin concentrationsaffect the structure of the Tisseel fibrin network, fibrin gelformulations listed in Table 1 were prepared and observedusing confocal microscopy. Results demonstrate that thepore size decreases as fibrinogen concentration increases(Table 1). As thrombin concentration increases from 1 to125U=mL, the average pore size decreases approximately byhalf (Table 1). Representative images enable visual confir-mation that a tighter overall network results when eitherfibrinogen (Fig. 1a vs. 1b) or thrombin (Fig. 1b vs. 1c) con-centrations are increased.
Compressive modulus of fibrin gels
Varying the fibrinogen concentration also influenced thecompressive modulus of fibrin gels. The compressive mod-ulus is an indicator of the mechanical stiffness of the net-work, a property that is important to consider as it has beenshown to influence process extension from neurons29 andglial cell growth.16 Our results show that when fibrin gelswere made with fibrinogen concentrations that increase from
5 to 25mg=mL, the compressive modulus of the gels in-creased approximately threefold (Table 1).
Cell-mediated degradation of fibrin gels
To determine the extent of cell-mediated fibrin gel deg-radation over the course of the culture period, fibrin gelswere made with the additional incorporation of fluorescentfibrinogen. The release of fluorescently labeled degradationproducts was monitored during the culture period. Resultsshowed that fibrin networks prepared with higher fibrinogenconcentrations degraded more slowly (Fig. 1d). Fibrin net-works prepared with higher thrombin concentrations de-graded more quickly (Fig. 1d). All gels were present for theduration of the culture period (7 days), a timescale slightlyshorter than the timescale over which the fastest degradinggel (5mg=mL fibrinogen and 125U=mL thrombin) degraded(9 days).
Influence of fibrin formulation on cell viability: Apoptosisand metabolic activity measurements
In effort to determine if fibrin formulation impacts cellviability, apoptotic activity was assessed on days 0, 1, and 7
FIG. 1. Structure and degradation rate of fibrin gels as fibrinogen and thrombin concentration varies. (a–c) Confocalmicroscopy images taken at 40�magnification showing the structure of fibrin matrices made with fibrinogen alexafluor-488as fibrinogen concentration increases from 5 (a) to 25mg=mL (b) with thrombin concentration is held constant at 1U=mL, oras thrombin concentration increases from 1 (b) to 125U=mL (c) with fibrinogen concentration is held constant at 25mg=mL(a–c). Scale bar¼ 50 mm. (d) Fraction of fibrin gel degraded by cell-secreted proteases over time. Fraction degraded wasdetermined by measuring the average amount of fibrinogen alexafluor-488 released from the fibrin gel into the supernatant asneural cells proteolytically degraded the nine different fibrin networks. This value was normalized to the amount of fluo-rescent signal present when the gel is 100% degraded. Color images available online at www.liebertonline.com=ten.
1610 MOONEY ET AL.
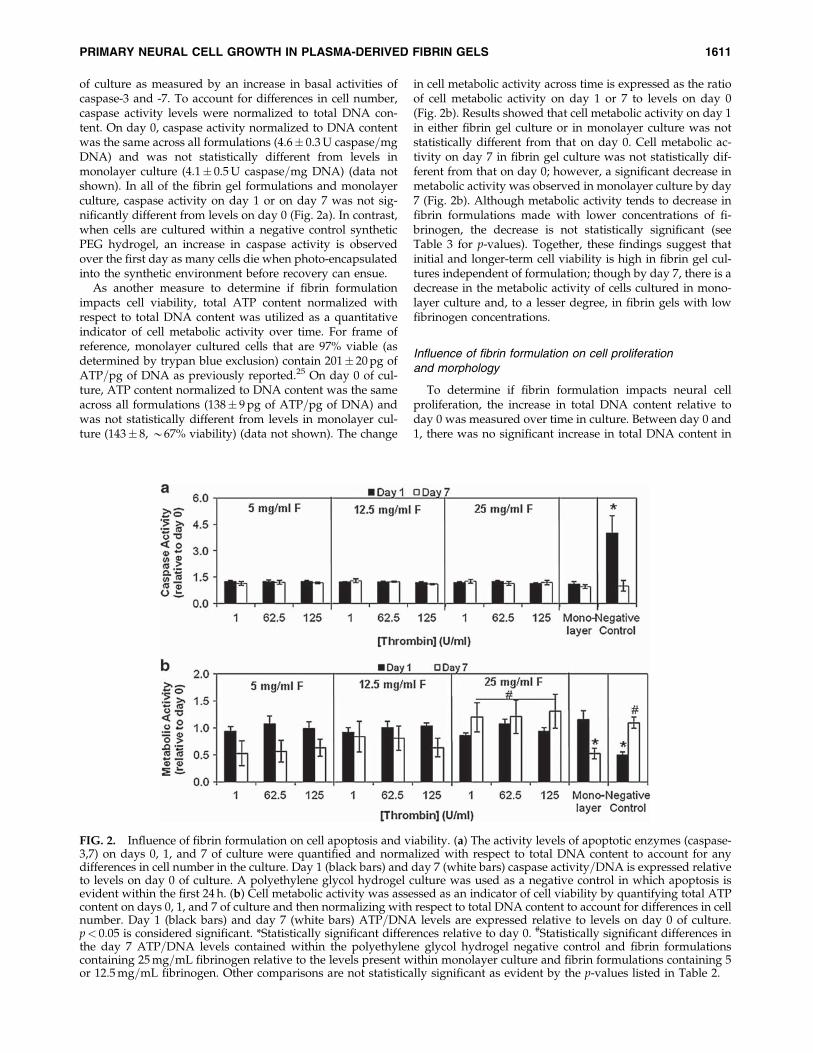
of culture as measured by an increase in basal activities ofcaspase-3 and -7. To account for differences in cell number,caspase activity levels were normalized to total DNA con-tent. On day 0, caspase activity normalized to DNA contentwas the same across all formulations (4.6� 0.3U caspase=mgDNA) and was not statistically different from levels inmonolayer culture (4.1� 0.5U caspase=mg DNA) (data notshown). In all of the fibrin gel formulations and monolayerculture, caspase activity on day 1 or on day 7 was not sig-nificantly different from levels on day 0 (Fig. 2a). In contrast,when cells are cultured within a negative control syntheticPEG hydrogel, an increase in caspase activity is observedover the first day as many cells die when photo-encapsulatedinto the synthetic environment before recovery can ensue.
As another measure to determine if fibrin formulationimpacts cell viability, total ATP content normalized withrespect to total DNA content was utilized as a quantitativeindicator of cell metabolic activity over time. For frame ofreference, monolayer cultured cells that are 97% viable (asdetermined by trypan blue exclusion) contain 201� 20pg ofATP=pg of DNA as previously reported.25 On day 0 of cul-ture, ATP content normalized to DNA content was the sameacross all formulations (138� 9pg of ATP=pg of DNA) andwas not statistically different from levels in monolayer cul-ture (143� 8, *67% viability) (data not shown). The change
in cell metabolic activity across time is expressed as the ratioof cell metabolic activity on day 1 or 7 to levels on day 0(Fig. 2b). Results showed that cell metabolic activity on day 1in either fibrin gel culture or in monolayer culture was notstatistically different from that on day 0. Cell metabolic ac-tivity on day 7 in fibrin gel culture was not statistically dif-ferent from that on day 0; however, a significant decrease inmetabolic activity was observed in monolayer culture by day7 (Fig. 2b). Although metabolic activity tends to decrease infibrin formulations made with lower concentrations of fi-brinogen, the decrease is not statistically significant (seeTable 3 for p-values). Together, these findings suggest thatinitial and longer-term cell viability is high in fibrin gel cul-tures independent of formulation; though by day 7, there is adecrease in the metabolic activity of cells cultured in mono-layer culture and, to a lesser degree, in fibrin gels with lowfibrinogen concentrations.
Influence of fibrin formulation on cell proliferationand morphology
To determine if fibrin formulation impacts neural cellproliferation, the increase in total DNA content relative today 0 was measured over time in culture. Between day 0 and1, there was no significant increase in total DNA content in
FIG. 2. Influence of fibrin formulation on cell apoptosis and viability. (a) The activity levels of apoptotic enzymes (caspase-3,7) on days 0, 1, and 7 of culture were quantified and normalized with respect to total DNA content to account for anydifferences in cell number in the culture. Day 1 (black bars) and day 7 (white bars) caspase activity=DNA is expressed relativeto levels on day 0 of culture. A polyethylene glycol hydrogel culture was used as a negative control in which apoptosis isevident within the first 24 h. (b) Cell metabolic activity was assessed as an indicator of cell viability by quantifying total ATPcontent on days 0, 1, and 7 of culture and then normalizing with respect to total DNA content to account for differences in cellnumber. Day 1 (black bars) and day 7 (white bars) ATP=DNA levels are expressed relative to levels on day 0 of culture.p< 0.05 is considered significant. *Statistically significant differences relative to day 0. #Statistically significant differences inthe day 7 ATP=DNA levels contained within the polyethylene glycol hydrogel negative control and fibrin formulationscontaining 25mg=mL fibrinogen relative to the levels present within monolayer culture and fibrin formulations containing 5or 12.5mg=mL fibrinogen. Other comparisons are not statistically significant as evident by the p-values listed in Table 2.
PRIMARY NEURAL CELL GROWTH IN PLASMA-DERIVED FIBRIN GELS 1611
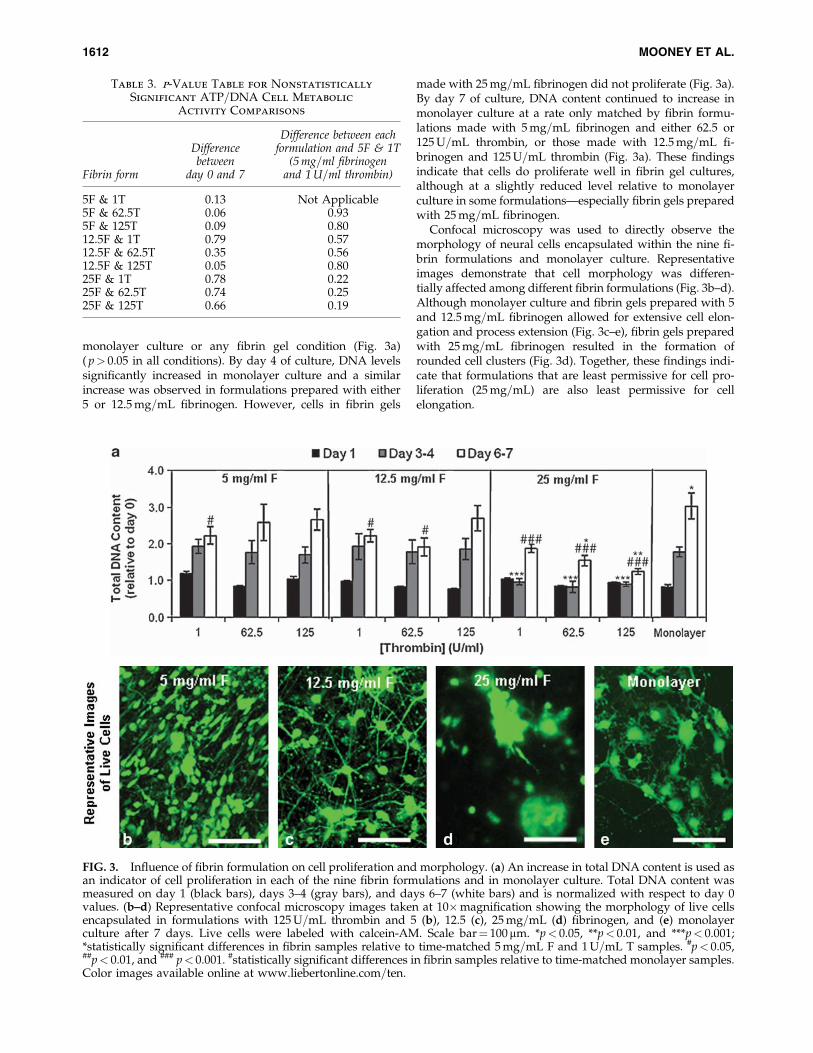
monolayer culture or any fibrin gel condition (Fig. 3a)( p> 0.05 in all conditions). By day 4 of culture, DNA levelssignificantly increased in monolayer culture and a similarincrease was observed in formulations prepared with either5 or 12.5mg=mL fibrinogen. However, cells in fibrin gels
made with 25mg=mL fibrinogen did not proliferate (Fig. 3a).By day 7 of culture, DNA content continued to increase inmonolayer culture at a rate only matched by fibrin formu-lations made with 5mg=mL fibrinogen and either 62.5 or125U=mL thrombin, or those made with 12.5mg=mL fi-brinogen and 125U=mL thrombin (Fig. 3a). These findingsindicate that cells do proliferate well in fibrin gel cultures,although at a slightly reduced level relative to monolayerculture in some formulations—especially fibrin gels preparedwith 25mg=mL fibrinogen.
Confocal microscopy was used to directly observe themorphology of neural cells encapsulated within the nine fi-brin formulations and monolayer culture. Representativeimages demonstrate that cell morphology was differen-tially affected among different fibrin formulations (Fig. 3b–d).Although monolayer culture and fibrin gels prepared with 5and 12.5mg=mL fibrinogen allowed for extensive cell elon-gation and process extension (Fig. 3c–e), fibrin gels preparedwith 25mg=mL fibrinogen resulted in the formation ofrounded cell clusters (Fig. 3d). Together, these findings indi-cate that formulations that are least permissive for cell pro-liferation (25mg=mL) are also least permissive for cellelongation.
Table 3. p-Value Table for Nonstatistically
Significant ATP=DNA Cell Metabolic
Activity Comparisons
Fibrin form
Differencebetween
day 0 and 7
Difference between eachformulation and 5F & 1T
(5mg=ml fibrinogenand 1U=ml thrombin)
5F & 1T 0.13 Not Applicable5F & 62.5T 0.06 0.935F & 125T 0.09 0.8012.5F & 1T 0.79 0.5712.5F & 62.5T 0.35 0.5612.5F & 125T 0.05 0.8025F & 1T 0.78 0.2225F & 62.5T 0.74 0.2525F & 125T 0.66 0.19
FIG. 3. Influence of fibrin formulation on cell proliferation and morphology. (a) An increase in total DNA content is used asan indicator of cell proliferation in each of the nine fibrin formulations and in monolayer culture. Total DNA content wasmeasured on day 1 (black bars), days 3–4 (gray bars), and days 6–7 (white bars) and is normalized with respect to day 0values. (b–d) Representative confocal microscopy images taken at 10�magnification showing the morphology of live cellsencapsulated in formulations with 125U=mL thrombin and 5 (b), 12.5 (c), 25mg=mL (d) fibrinogen, and (e) monolayerculture after 7 days. Live cells were labeled with calcein-AM. Scale bar¼ 100mm. *p< 0.05, **p< 0.01, and ***p< 0.001;*statistically significant differences in fibrin samples relative to time-matched 5mg=mL F and 1U=mL T samples. #p< 0.05,##p< 0.01, and ### p< 0.001. #statistically significant differences in fibrin samples relative to time-matched monolayer samples.Color images available online at www.liebertonline.com=ten.
1612 MOONEY ET AL.
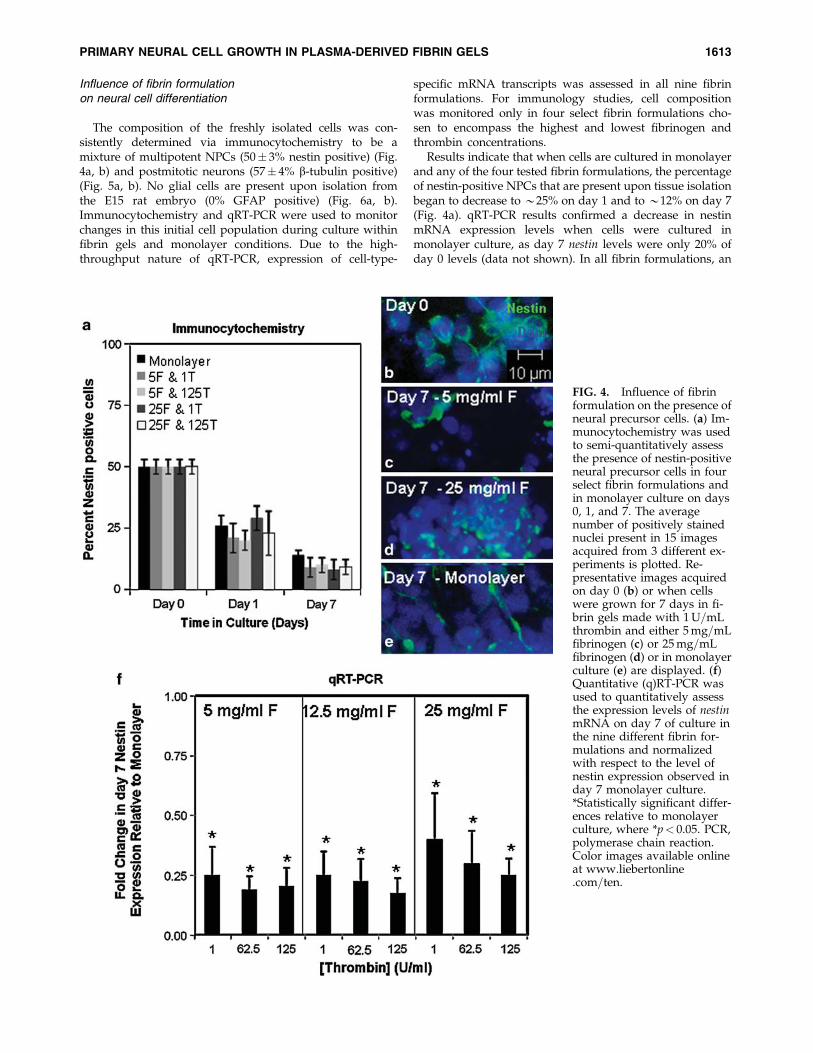
Influence of fibrin formulationon neural cell differentiation
The composition of the freshly isolated cells was con-sistently determined via immunocytochemistry to be amixture of multipotent NPCs (50� 3% nestin positive) (Fig.4a, b) and postmitotic neurons (57� 4% b-tubulin positive)(Fig. 5a, b). No glial cells are present upon isolation fromthe E15 rat embryo (0% GFAP positive) (Fig. 6a, b).Immunocytochemistry and qRT-PCR were used to monitorchanges in this initial cell population during culture withinfibrin gels and monolayer conditions. Due to the high-throughput nature of qRT-PCR, expression of cell-type-
specific mRNA transcripts was assessed in all nine fibrinformulations. For immunology studies, cell compositionwas monitored only in four select fibrin formulations cho-sen to encompass the highest and lowest fibrinogen andthrombin concentrations.
Results indicate that when cells are cultured in monolayerand any of the four tested fibrin formulations, the percentageof nestin-positive NPCs that are present upon tissue isolationbegan to decrease to *25% on day 1 and to *12% on day 7(Fig. 4a). qRT-PCR results confirmed a decrease in nestinmRNA expression levels when cells were cultured inmonolayer culture, as day 7 nestin levels were only 20% ofday 0 levels (data not shown). In all fibrin formulations, an
FIG. 4. Influence of fibrinformulation on the presence ofneural precursor cells. (a) Im-munocytochemistry was usedto semi-quantitatively assessthe presence of nestin-positiveneural precursor cells in fourselect fibrin formulations andin monolayer culture on days0, 1, and 7. The averagenumber of positively stainednuclei present in 15 imagesacquired from 3 different ex-periments is plotted. Re-presentative images acquiredon day 0 (b) or when cellswere grown for 7 days in fi-brin gels made with 1U=mLthrombin and either 5mg=mLfibrinogen (c) or 25mg=mLfibrinogen (d) or in monolayerculture (e) are displayed. (f)Quantitative (q)RT-PCR wasused to quantitatively assessthe expression levels of nestinmRNA on day 7 of culture inthe nine different fibrin for-mulations and normalizedwith respect to the level ofnestin expression observed inday 7 monolayer culture.*Statistically significant differ-ences relative to monolayerculture, where *p< 0.05. PCR,polymerase chain reaction.Color images available onlineat www.liebertonline.com=ten.
PRIMARY NEURAL CELL GROWTH IN PLASMA-DERIVED FIBRIN GELS 1613
even larger decrease in nestin gene expression was observed(Fig. 4f).
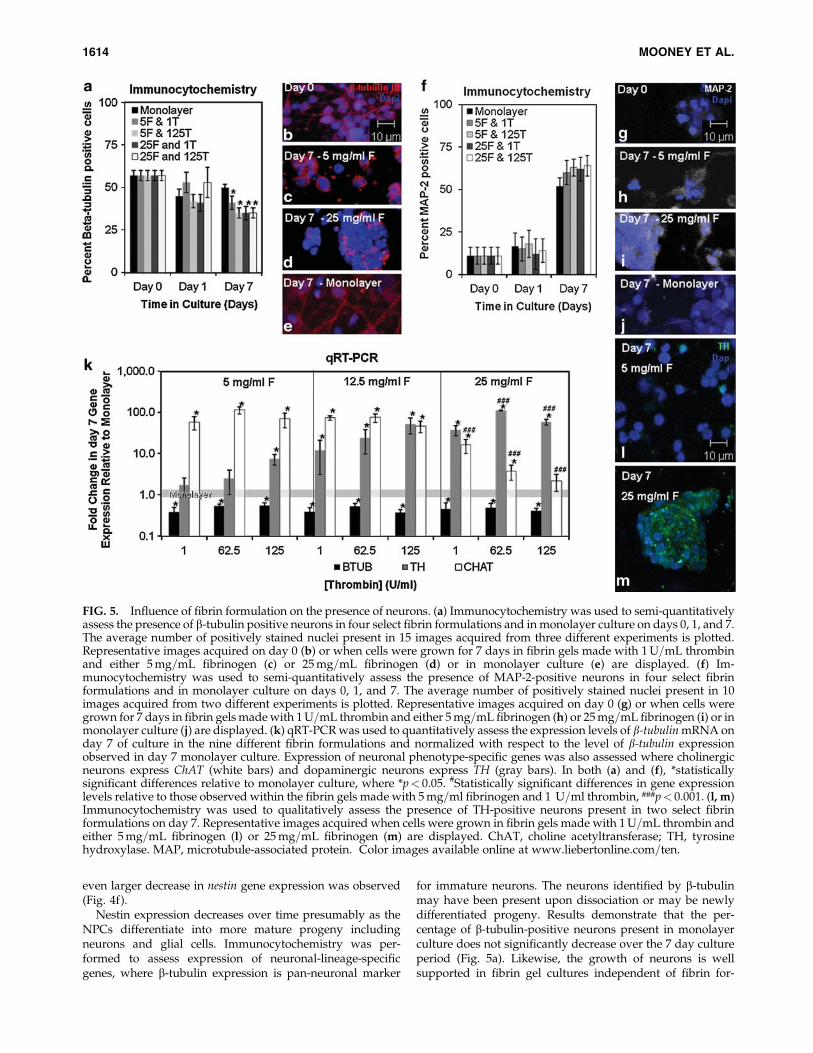
Nestin expression decreases over time presumably as theNPCs differentiate into more mature progeny includingneurons and glial cells. Immunocytochemistry was per-formed to assess expression of neuronal-lineage-specificgenes, where b-tubulin expression is pan-neuronal marker
for immature neurons. The neurons identified by b-tubulinmay have been present upon dissociation or may be newlydifferentiated progeny. Results demonstrate that the per-centage of b-tubulin-positive neurons present in monolayerculture does not significantly decrease over the 7 day cultureperiod (Fig. 5a). Likewise, the growth of neurons is wellsupported in fibrin gel cultures independent of fibrin for-
FIG. 5. Influence of fibrin formulation on the presence of neurons. (a) Immunocytochemistry was used to semi-quantitativelyassess the presence of b-tubulin positive neurons in four select fibrin formulations and in monolayer culture on days 0, 1, and 7.The average number of positively stained nuclei present in 15 images acquired from three different experiments is plotted.Representative images acquired on day 0 (b) or when cells were grown for 7 days in fibrin gels made with 1U=mL thrombinand either 5mg=mL fibrinogen (c) or 25mg=mL fibrinogen (d) or in monolayer culture (e) are displayed. (f) Im-munocytochemistry was used to semi-quantitatively assess the presence of MAP-2-positive neurons in four select fibrinformulations and in monolayer culture on days 0, 1, and 7. The average number of positively stained nuclei present in 10images acquired from two different experiments is plotted. Representative images acquired on day 0 (g) or when cells weregrown for 7 days in fibrin gels madewith 1U=mL thrombin and either 5mg=mL fibrinogen (h) or 25mg=mL fibrinogen (i) or inmonolayer culture (j) are displayed. (k) qRT-PCRwas used to quantitatively assess the expression levels of b-tubulinmRNA onday 7 of culture in the nine different fibrin formulations and normalized with respect to the level of b-tubulin expressionobserved in day 7 monolayer culture. Expression of neuronal phenotype-specific genes was also assessed where cholinergicneurons express ChAT (white bars) and dopaminergic neurons express TH (gray bars). In both (a) and (f), *statisticallysignificant differences relative to monolayer culture, where *p< 0.05. #Statistically significant differences in gene expressionlevels relative to those observed within the fibrin gels made with 5mg=ml fibrinogen and 1 U=ml thrombin, ###p< 0.001. (l, m)Immunocytochemistry was used to qualitatively assess the presence of TH-positive neurons present in two select fibrinformulations on day 7. Representative images acquired when cells were grown in fibrin gels made with 1U=mL thrombin andeither 5mg=mL fibrinogen (l) or 25mg=mL fibrinogen (m) are displayed. ChAT, choline acetyltransferase; TH, tyrosinehydroxylase. MAP, microtubule-associated protein. Color images available online at www.liebertonline.com=ten.
1614 MOONEY ET AL.
mulation as evidenced by the robust staining evident in fi-brin gels on day 7 (Fig. 5c, d), though the percentage of b-tubulin-positive cells present on day 7 does decrease to*30% of day 0 levels (Fig. 5a). qRT-PCR results confirmed asupportive environment for neurons as b-tubulin mRNAexpression levels remain at 80% of day 0 levels when cellswere cultured in monolayer culture (data not shown). qRT-PCR results also confirm a significantly decreased level of
b-tubulin expression when cells are cultured in fibrin gelsrelative to monolayer culture independent of the fibrin for-mulation (Fig. 5f).
As b-tubulin is expressed predominantly in immatureneurons, the consistent decrease in b-tubulin-positive cellsand mRNA expression observed in culture over the 7 dayculture period may be the result of the progression of neu-rons to a more mature phenotype. To test this possibility,microtubule-associated protein-2 (MAP-2) expression wasalso assessed as a pan-neuronal marker expressed in matureneurons. Results confirm an increase in the percentage ofMAP-2-positive cells over the duration of the culture period(Fig. 5f). Although there was no significant difference in thepercentage of MAP-2-positive cells present in monolayerculture and those present in any of the tested fibrin formu-lations, representative images demonstrate more intensestaining in the fibrin gel cultures (Fig. 5h, i) when comparedwith monolayer culture (Fig. 5j).
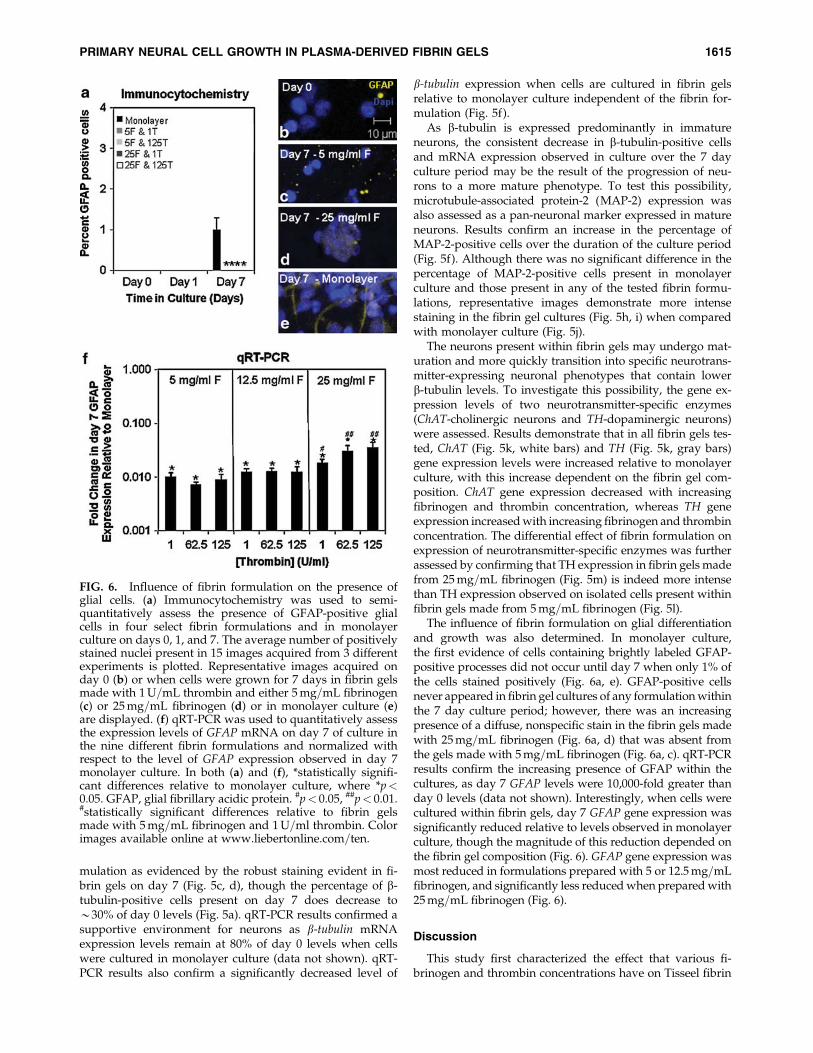
The neurons present within fibrin gels may undergo mat-uration and more quickly transition into specific neurotrans-mitter-expressing neuronal phenotypes that contain lowerb-tubulin levels. To investigate this possibility, the gene ex-pression levels of two neurotransmitter-specific enzymes(ChAT-cholinergic neurons and TH-dopaminergic neurons)were assessed. Results demonstrate that in all fibrin gels tes-ted, ChAT (Fig. 5k, white bars) and TH (Fig. 5k, gray bars)gene expression levels were increased relative to monolayerculture, with this increase dependent on the fibrin gel com-position. ChAT gene expression decreased with increasingfibrinogen and thrombin concentration, whereas TH geneexpression increasedwith increasing fibrinogen and thrombinconcentration. The differential effect of fibrin formulation onexpression of neurotransmitter-specific enzymes was furtherassessed by confirming that TH expression in fibrin gels madefrom 25mg=mL fibrinogen (Fig. 5m) is indeed more intensethan TH expression observed on isolated cells present withinfibrin gels made from 5mg=mL fibrinogen (Fig. 5l).
The influence of fibrin formulation on glial differentiationand growth was also determined. In monolayer culture,the first evidence of cells containing brightly labeled GFAP-positive processes did not occur until day 7 when only 1% ofthe cells stained positively (Fig. 6a, e). GFAP-positive cellsnever appeared in fibrin gel cultures of any formulationwithinthe 7 day culture period; however, there was an increasingpresence of a diffuse, nonspecific stain in the fibrin gels madewith 25mg=mL fibrinogen (Fig. 6a, d) that was absent fromthe gels made with 5mg=mL fibrinogen (Fig. 6a, c). qRT-PCRresults confirm the increasing presence of GFAP within thecultures, as day 7 GFAP levels were 10,000-fold greater thanday 0 levels (data not shown). Interestingly, when cells werecultured within fibrin gels, day 7 GFAP gene expression wassignificantly reduced relative to levels observed in monolayerculture, though the magnitude of this reduction depended onthe fibrin gel composition (Fig. 6). GFAP gene expression wasmost reduced in formulations prepared with 5 or 12.5mg=mLfibrinogen, and significantly less reducedwhen prepared with25mg=mL fibrinogen (Fig. 6).
Discussion
This study first characterized the effect that various fi-brinogen and thrombin concentrations have on Tisseel fibrin
FIG. 6. Influence of fibrin formulation on the presence ofglial cells. (a) Immunocytochemistry was used to semi-quantitatively assess the presence of GFAP-positive glialcells in four select fibrin formulations and in monolayerculture on days 0, 1, and 7. The average number of positivelystained nuclei present in 15 images acquired from 3 differentexperiments is plotted. Representative images acquired onday 0 (b) or when cells were grown for 7 days in fibrin gelsmade with 1U=mL thrombin and either 5mg=mL fibrinogen(c) or 25mg=mL fibrinogen (d) or in monolayer culture (e)are displayed. (f) qRT-PCR was used to quantitatively assessthe expression levels of GFAP mRNA on day 7 of culture inthe nine different fibrin formulations and normalized withrespect to the level of GFAP expression observed in day 7monolayer culture. In both (a) and (f), *statistically signifi-cant differences relative to monolayer culture, where *p<0.05. GFAP, glial fibrillary acidic protein. #p< 0.05, ##p< 0.01.#statistically significant differences relative to fibrin gelsmade with 5mg=mL fibrinogen and 1U=ml thrombin. Colorimages available online at www.liebertonline.com=ten.
PRIMARY NEURAL CELL GROWTH IN PLASMA-DERIVED FIBRIN GELS 1615

gel stiffness, structure, and degradation rate. Tisseel containsunique salt concentrations and aprotinin that may alter thefibrin material properties relative to other studies.30 Com-pressive modulus results were consistent with previousfindings in that as fibrinogen concentration increases, themodulus of the resulting gels increases.31–33 Our structuralresults are also consistent with previous findings in that theporosity of the gel matrix decreases with increasing fibrino-gen and thrombin concentrations.34 Our degradation resultsdemonstrate rapid cell-mediated degradation of fibrin gelsprepared using high thrombin concentrations (62.5 or125U=mL). Although thrombin concentration greatly influ-ences the degree of degradation when making gels usingboth high (25mg=mL) or low (5mg=mL) fibrinogen, fibrin-ogen concentration is also a major determining factor in fi-brin degradation rates, where formulations with lowerfibrinogen concentrations are degraded most quickly byneural cells. These results are consistent with studies show-ing that fibrin gels made using low fibrinogen and highthrombin concentrations contain thin fibers and that thin fi-bers are lysed faster than thick fibers.35–37 Interestingly, thedegradation timescales observed in the present study areslightly prolonged relative to those achieved in studies per-formed using purified human fibrinogen, validating the needfor a separate study exploring the degradation of Tissel fibringels.38 It is pertinent to note the presence of a recent studythat reports results contradictory to those reported herein that increased neurite outgrowth was observed in fibringels that contained greater fibrinogen concentrations.39 Byour measurements, greater fibrinogen concentrations create agel that is less susceptible to proteolysis and should intui-tively inhibit neurite outgrowth. This apparent discrepancymay be attributable to the very low thrombin concentrationsemployed in the previous study (0.25U=mL), which, wehave demonstrated in our lab, leads to very inhomogeneousfibrin gel structures (data not shown) that contain structuralattributes (fiber size, fibril bundle diameter, etc.) that aredifficult to control and that can substantially influenceneurite outgrowth.
Differences in fibrin material properties may variablyimpact cell function. For example, cells encapsulated withinfibrin scaffolds that are very susceptible to proteolytic deg-radation remain as single cells and extend processes, butwhen encapsulated within fibrin scaffolds resistant to pro-teolytic degradation, cells form rounded cell clusters ratherthan extend processes. This result is consistent with previousfindings showing that other cell types remain rounded whengels are prepared with higher fibrinogen, whereas theyelongate in gels prepared with lower fibrinogen concentra-tions.2–4,40
The susceptibility of the fibrin matrix to proteolytic deg-radation may also impact cell proliferation rates. This studydemonstrates that in fibrin formulations prepared with lowlevels of fibrinogen, cells generally proliferated at a ratecomparable to that observed in monolayer culture, suggest-ing that easily degradable matrices do not impede cell pro-liferation rates to a high degree. However, when fibrin gelswere prepared with 25mg=mL fibrinogen, proliferation wasdelayed and was lower in magnitude on day 7, indicatingthat the matrix imparted a proliferation hindrance. Theseresults are consistent with previous findings using other celltypes.3–4
We provide evidence that the decreased proliferation ob-served in gels prepared with high fibrinogen concentrationsis not simply due to variations in the number of survivingcells, as fibrin formulation has no effect on apoptotic activitylevels. This result is consistent with previous studies usingother cell types.2–4 Likewise, cell metabolic activity levelsremain at least as high as those achieved in monolayer cul-ture (67%) for the duration of the experiment. Interestingly,despite good cell viability, there is a reduction in metabolicactivity observed in monolayer culture by day 7. A similartrend toward reduced metabolic activity is also observed infibrin formulations prepared with low fibrinogen concen-trations. Incidentally, these are the same culture conditions inwhich the greatest increases in cell proliferation and processextension are observed. It is possible that the decrease in cellmetabolic activity in these conditions is due to confluency, ascontact inhibition has been previously reported to result inincreased enzyme-mediated hydrolysis of ATP when neuralcells are cultured in vitro.41,42 The increased ATP hydrolysisthat occurs when cells contact each other in confluent cultureconditions would not be associated with an increase in cas-pase activity levels.43
The influence of fibrin formulation on cell compositionwas also assessed in this work. Although the susceptibility ofthe fibrin matrix to cell-mediated degradation influenceshow readily entrapped cells extend processes and proliferate,the results presented here demonstrate that the biggest dif-ferences in cell composition correlate more with fibrinogenconcentration and are more or less independent of degra-dation rate. Immunological characterization of the initial cellpopulation indicated that *50% of the initial cell populationwas nestin-positive, *57% of the initial cell population wasb-tubulin-positive, and 0% was GFAP-positive. The numberof positive cells present on day 0 exceeds 100%, and this islikely due to some co-labeling of nestin and b-tubulin asNPCs transition into neuronal differentiation. Althoughdouble immunostaining was not performed in this work, co-labeling of these two markers during the transition betweenthe precursor cell state and the postmitotic neuronal state hasbeen reported previously.44 The percentage of b-tubulin,nestin, and GFAP-positive cells is very consistent betweencell isolations and agrees well with previously reportedstudies of this cell population from other groups using othertechniques such as fluorescence activated cell sorting (FACs)analysis.45
Fibrin formulation did not seem to influence the presenceof nestin-positive NPCs or b-tubulin-positive neurons,though the presence of these immature cell types were de-creased in fibrin gels relative to monolayer culture. In con-trast, fibrin gel culture resulted in increased expressionintensity of MAP-2, a marker of mature neurons that ispresent on increasing percentages of neurons in both fibringel and monolayer culture throughout the duration of theculture period. The increase in MAP-2 expression over timeis consistent with other studies using neural cells isolatedfrom rat embryos at 14–15 days of gestation.46,47 Levels ofneurotransmitter-specific enzymes produced by mature,therapeutically relevant dopaminergic and cholinergic neu-ronal phenotypes were also increased in fibrin gel culturerelative to monolayer culture. Interestingly, as fibrinogenconcentration increased, the improvement in ChAT expres-sion decreased, whereas the improvement in TH expression
1616 MOONEY ET AL.

increased. We speculate that this may be related to differ-ences in these two neuronal phenotype’s need for glial-cell-derived trophic factors and aggregated cell growth, whichare beneficial for dopaminergic survival and would increasein fibrin gels made with higher fibrinogen concentra-tions.48,49 This hypothesis is supported by the increased in-tensity of TH staining in the cell aggregates that grow withinfibrin gels made from 25mg=mL fibrinogen relative to theisolated cells that grow when fibrin gels are made with5mg=mL fibrinogen.
In contrast, GFAP mRNA that is expressed by glial cellswhich can contribute to glial scars that inhibit nerve regen-eration was significantly lower in all fibrin formulationsrelative to monolayer culture. These robust differences in theneuronal and glial cell mRNA expression may be related tothe large difference in the stiffness of fibrin gels compared tomonolayer culture. It has been previously demonstrated thatsoft fibrin substrates do not negatively affect neuronal cellgrowth, but reduce actin-filament polymerization in glialcells, thereby discouraging glial cell growth.16 Our studyprovides additional evidence that substrate stiffness influ-ences the extent of glial cell growth, in that fibrin formula-tions with increased compressive moduli (higher fibrinogenconcentration) resulted in higher GFAP gene expression thanthat observed in softer fibrin gels.
Although this work focused on the potential impact ofmechanical properties of various fibrin formulations onneural cell behavior, another possible reason for the differ-ences in neural cell behavior observed when cultured invarious fibrin formulations and=or monolayer culture is thevarying levels of fibrin ligand densities. Bioactive or adhe-sive sites present on fibrin can bind to receptors on cellsurfaces, so the instructive signaling that the matrix providescould be ligand-receptor based rather than mechanical. Inaddition, the fibrinogen and thrombin solutions in the Tisseelkit are prepared by cryoprecipitation from blood plasma, anthus contain other proteins including transforming growthfactor b-1, basic fibroblast growth factor, epidermal growthfactor, and vascular endothelial growth factor.48 Each ofthese proteins has been shown to impact the function ofmultipotent precursor cells or postmitotic neurons,30,50–58
and may be at least partially responsible for the observedchanges in cell behavior in the different fibrin formulationsand in monolayer culture.
In conclusion, the studies described here characterize theeffect of fibrinogen and thrombin on the growth and com-position of a mixed population of postmitotic neurons andmultipotent NPCs in fibrin gels. This cell population is usefulfor neural cell transplant therapy, and the effect of fibrinformulation on this cell type has not yet been explored. Al-though the cells survive well in all formulations tested, im-portant differences in proliferation and resulting cellcomposition were observed. Easily degradable fibrin gelsprepared with lower fibrinogen concentrations facilitatedgreater cell proliferation and process extension. Relative tostiff monolayer culture, glial cell growth was suppressed,and the differentiation of two clinically relevant neuro-transmitter-expressing neurons (dopaminergic and cholin-ergic neurons) was promoted in a manner that wasdependent on the concentration of fibrinogen or thrombin.The findings here suggest the importance of fibrinogen andthrombin concentration in differentially regulating the
growth and differentiation of neural cell populations con-taining NPCs and are of importance for neural tissue engi-neering strategies focused on developing implantable cellscaffolds enriched in neurons that tend to be lost during theprogression of Parkinson’s disease and Alzheimer’s disease.
Disclosure Statement
No competing financial interests exist.
References
1. Mosesson, M.W., and Doolittle, R.F. Molecular Biology ofFibrinogen and Fibrin. Annals of the New York Academy ofSciences, New York, NY, 1983.
2. Bensaid, W., Triffitt, J.T., Blanchat, C., Oudina, K., Sedel, L.,and Petite, H. A biodegradable fibrin scaffold for mesen-chymal stem cell transplantation. Biomaterials 24, 2497,2003.
3. Catelas, I., Sese, N., Wu, B.M., Dunn, J.C.Y., Helgerson, S.,and Tawil, B. Human mesenchymal stem cell proliferationand osteogenic differentiation in fibrin gels in vitro. TissueEng 12, 2385, 2006.
4. Cox, S., Cole, M., and Tawil, B. Behavior of human dermalfibroblasts in 3D fibrin clots: dependence on fibrinogen andthrombin concentration. Tissue Eng 10, 942, 2004.
5. Rowe, S.L., Lee, S.Y., and Stegemann, J.P. Influence ofthrombin concentration on the mechanical and morpholog-ical properties of cell-seeded fibrin hydrogels. Acta Biomater3, 59, 2007.
6. Kassam, A., Nemoto, E., Balzer, J., Rao, G., Welch, W.C.,Kuwabara, H., Boada, F., and Horowitz, M. Effects of Tisseelfibrin glue on the CNS of nonhuman primates. Ear NoseThroat J 83, 246, 2004.
7. Freeman, T.B., and Widner, H. Cell Transplantation forNeurological Disorders: Toward Reconstruction of theHuman Central Nervous System. Totowa, NJ: Humana PressInc., 1998, pp. 155–158.
8. Isacson, O., Brundin, P., Kelly, P.A., Gage, F.H., and Bjork-lund, A. Functional neuronal replacement by grafted striatalneurons in the ibotenic acid-lesioned striatum. Nature 311,
458, 1984.9. Schmidt, R.H., Bjorklund, A., and Stenevi, U. Intracerebral
grafting of dissociated CNS tissue suspensions a new ap-proach for neuronal transplantation to deep brain sites.Brain Res 218, 347,1981.
10. Nishino, H., Aihara, N., Czurko, A., Hashitani, T., Isobe, Y.,Ichikawa, O., and Watari, H. Reconstruction of GABAergictransmission and behavior by striatl cell grafts in rats withischemic infarcts in the middle cerebral artery. J NeuralTransplant Plast 4, 147, 1993.
11. Lee, A.C., Yu, V.M., Lowe, J.B., Brenner, M.J., Hunter, D.A.,Mackinnon, S.E., and Sakiyama-Elbert, S.E. Controlled re-lease of nerve growth factor enhances sciatic nerve regener-ation. Exp Neurol 184, 295, 2003.
12. Taylor, S.J., McDonald, J.W., and Sakiyama-Elbert, S.E.Controlled release of neurotrophin-3 from fibrin gels forspinal cord injury. J Control Release 98, 281, 2004.
13. Cheng, H., Cao, Y., and Olson, L. Spinal cord repair in adultparaplegic rats: partial restoration of hind limb function.Science 273, 510, 1996.
14. Taylor, S.J., Rosenzweig, E.S., McDonald, J.W., and Sakiyama-Elbert, S.E. Delivery of neurotrophin-3 from fibrin enhancesneuronal fiber sprouting after spinal cord injury. J ControlRelease 113, 226, 2006.
PRIMARY NEURAL CELL GROWTH IN PLASMA-DERIVED FIBRIN GELS 1617

15. Bjorkland, A., and Lindvall, O. Cell replacement therapiesfor central nervous system disorders. Nat Neurosci 3, 537–544, 2000.
16. Georges, P., Miller, W., Meaney, D., Sawyer, E., and Janmey,P. Matrices with compliance comparable to that of braintissue select neuronal over glial growth in mixed corticalcultures. Biophys J 90, 3012, 2006.
17. Willerth, S.M., Faxel, T.E., Gottlieb, D.I., and Sakiyama-Elbert, S.E. The effects of soluble growth factors on embry-onic stem cell differentiation inside of fibrin scaffolds. StemCells 9, 2235, 2007.
18. Willerth, S.M., Arendas, K.J., Gottlieb, D.I., and Sakiyama-Elbert, S.E. Optimization of fibrin scaffolds for differentia-tion of murine embryonic stem cells into neural lineage cells.Biomaterials 27, 5990, 2006.
19. Namba R.M., Cole, A.A., Bjugstad, K.B., and Mahoney, M.J.Development of porous PEG hydrogels that enable efficient,uniform cell-seeding and permit early neural process ex-tension. Acta Biomater 5, 1884, 2009.
20. Dihanich, M., Kaser, M., Reinhard, E., Cunningham, D., andMonard, D. Prothrombin mRNA is expressed by cells of thenervous system. Neuron 4, 575, 1991.
21. Mooney, R.G., Costales, C.A., Curtin, J.M., Tawil, B.,and Shaw, M.C. Indentation micromechanics of fibroblast-populated fibrin constructs. Mater Res Soc Symp Proc 874,
205, 2005.22. Nelb, G.W., Gerth, C., Ferry, J.D., and Lorand, L. Rheology
of fibrin clots: III. Sheer creep and creep recovery of fineligated and coarse unligated clots. Biophys Chem 5, 377,1976.
23. Carr, M.E., Martin, E.J., and Carr, S.L. Delayed, reduced orinhibited thrombin production reduces platelet contractileforce and results in weaker clot formation. Blood CoagulFibrinolysis 13, 193, 2002.
24. Duong, H., Wu, B., and Tawil, B. Modulation of 3D fibrinmatrix stiffness by intrinsic fibrinogen–thrombin composi-tions and by extrinsic cellular activity. Tissue Eng Part A 15,
1865, 2008.25. Mahoney, M.J., and Anseth, K.S. 3D growth and function of
neural tissue in degradable polyethylene glycol hydrogels.Biomaterials 27, 2265, 2006.
26. Levers, T.E. The fates of cells generated at the end of neu-rogenesis in developing mouse cortex. J Neurobiol 48, 265,2001.
27. Fukuda, S., Kato, F., Tozuka, Y., Yamaguchi, M., Miyamoto,Y., and Hisatsune, T. Two distinct subpopulations of nestin-positive cells in adult mouse dentate gyrus. J Neurosci 23,9357, 2003.
28. Hashimoto, J.G., Beadles-Bohling, A.S., and Wiren, K.M.Comparison of RiboGreen and 18S rRNA quantitation fornormalizing real-time RT-PCR expression analysis. Bio-techniques 1, 58, 2004.
29. Leach, J.B., Brown, X.Q., Jacot, J.G., DiMilla, P.A., andWong, J.Y. Neurite outgrowth and branching of PC12 cellson very soft substrates sharply decreases below a thresholdof substrate rigidity. J Neural Eng 4, 26, 2007.
30. Ho, W., Tawil, B., Dunn, J.C.Y., and Wu, B.M. The behaviorof human mesenchymal stem cells in 3D Fibrin Clots: de-pendence on fibrinogen concentration and clot structure.Tissue Eng 12, 1587, 2006.
31. Blomback, B., and Okada, M. Fibrin gel structure and clot-ting time. Thromb Res 25, 51, 1982.
32. Roberts, W.W., Kramer, O., Rosser, R.W., Nestler, F.H.M.,and Ferry, J.D. Rheology of fibrin clots. I: dynamic visco-
elastic properties and fluid permeation. Biophys Chem 1,
152, 1974.33. Carr, M.E., Shen, L.L., and Hermans, J. A physical standard
of fibrinogen: measurement of the elastic modulus of dilutefibrin gels with a new elastometer. Anal Biochem 72, 202,1976.
34. Blomback, B., Carlsson, K., Hessel, B., Liljeborg, A., Procyk,R., and Aslund, N. Native fibrin gel networks observed by3D microscopy, permeation and turbidity. Biochim BiophysActa 997, 96, 1989.
35. Blomback, B., Carlsson, K., Fatah, K., Hessel, B., and Procyk,R. Fibrin in human plasma: gel architectures governed byrate and nature of fibrinogen activation. Thromb Res 75, 521,1994.
36. Carr, M.E., and Alving, B.M. Effect of fibrin structure onplasmin-mediated dissolution of plasma clots. Blood CoagulFibrinolysis 6, 567,1995.
37. Gabriel, D.A., Muga, K., and Boothroyd, E.M. The effect offibrin structure on fibrinolysis. J Biol Chem 267, 24259, 1992.
38. Ju, Y., Jamney, P.A., McCormick, M.E., Sawyer, E.S., andFlanagan, L.A. Enhanced neurite growth from mammalianneurons in three-dimensional salmon fibrin gels. Biomater-ials 28, 2097, 2007.
39. Sarig-Nadir, O., and Seliktar, D. Compositional alterationsof fibrin-based materials for regulating in vitro neural out-growth. Tissue Eng Part A 14, 401, 2008.
40. Dikovsky, D., Bianco-Peled, H., and Seliktar, D. Defining therole of matrix compliance and proteolysis in 3D cellspreading and remodeling. Biophys J 94, 2914, 2008.
41. Ohnishi, T., and Kimura, S. Contact-mediated changes inATPase activity at the surface of primary cultured hepatomacells. Histochem Cell Biol 49, 107, 1976.
42. Stefanovic, V., Crestelski-Treska, J., Ledig, M., and Mandel,P. (Naþ, -Kþ)-activated ATPase and Kþ-activated p-nitrophenyl-phosphatase activities of the nervous systemcells in tissue culture. Experientia 31, 807, 1975.
43. Hneino, M., Bouazza, L., Bricca, G., Li, J., and Langlois, D.Density-dependant shift of transforming growth factor-b-1from inhibition to stimulation of vascular smooth muscle cellgrowth is based on unconventional regulation of prolifera-tion, apoptosis, and contact inhibition. J Vasc Res 46, 85, 2009.
44. Zimmerman, L., Lendah, U., Cunningham, M., McKay, R.,Parr, B., Gavin, B., Mann, J., Vassileva, G., and McMahon, A.Independent regulatory elements in the nestin gene directtransgene expression to neural stem cells or muscle precur-sors. Neuron 12, 11, 1994.
45. Hartfuss, E. Characterization of subtypes of precursor cellsin the development of the central nervous system [Thesis].Max Planck Institute for Neurobiology, Munich, Germany,2003, pp. 132–140.
46. Batesm, C.A., Trinh, N., and Meyer, R.L. Distribution ofmicrotubule-associated proteins (MAPs) in adult and em-bryonic mouse retinal explants: presence of the embryonicmap, MAP5=1B, in regenerating adult retinal axons. DevBiol 155, 533, 1993.
47. Erlandsson, A., Enarsson, M., and Forsberg-Nilsson, K. Im-mature neurons from CNS stem cells proliferate in responseto platelet-derived growth factor. J Neurosci 21, 3483, 2001.
48. Peter, E., Batchelor, P.E., Liberatore, G.T., Wong, J.Y.F.,Porritt, M.J., Frerichs, F., Donnan, G.A., and Howells, D.W.Activated macrophages and microglia induce dopaminergicsprouting in the injured striatum and express brain-derivedneurotrophic factor and glial cell line-derived neurotrophicfactor. J Neurosci 19, 1708, 1999.
1618 MOONEY ET AL.

49. Schulz, T.C., Noggle, S.A., Palmarini, G.M., Weiler, D.A.,Lyons, I.G., Pensa, K.A., Meedeniya, A.C.B., Davidson, B.P.,Lambert, N.A., and Condie, B.G. Differentiation of humanembryonic stem cells to dopaminergic neurons in serum-freesuspension culture. Stem Cells 22, 1218, 2004.
50. Gensburger, C., Labourdette, G., and Sensenbrenner, M.Brain basic fibroblast growth factor stimulates the prolifer-ation of rat neuronal precursor cells in vitro. FEBS Lett 217, 1,1987.
51. Richards, L.J., Kilpatrick, T.J., and Bartlett, P.F. De novogeneration of neuronal cells from the adult mouse brain.Proc Natl Acad Sci 89, 8591, 1992.
52. Ray, J., Peterson, D.A., Schinstine, M., and Gage, F.H. Pro-liferation, differentiation, and long-term culture of primaryhippocampal neurons. Proc Natl Acad Sci 90, 3602, 1993.
53. Davis, A.A., and Temple, S. A self-renewing multipotentialstem cell in embryonic rat cerebral cortex. Nature 372, 263,1994.
54. Reynolds, B.A., and Weiss, S. Generation of neurons andastrocytes from isolated cells of the adult mammalian centralnervous system. Science 255, 1707, 1992.
55. Reynolds, B.A., Tetzlaff, W., and Weiss, S. A multipotentEGF-responsive striatal embryonic progenitor cell producesneurons and astrocytes. J Neurosci 12, 4565, 1992.
56. Misumi, S., Kim, T.S., Jung, C.G., Masuda, T., Urakawa, S.,Isobe, Y., Furuyama, F., Nishino, H., and Hida, H. Enhanced
neurogenesis from neural progenitor cells with G1=S-phasecell cycle arrest is mediated by transforming growth factorb1. Eur J Neurosci 28, 1049, 2008.
57. Qian, L., Wei, S.J., Zhang, D., Hu, X., Xu, Z., Wilson, B.,El-Benna, J., Hong, J.S., and Flood, P.M. Potent anti-inflammatory and neuroprotective effects of TGF-b1 aremediated through the inhibition of ERK and p47phox-Ser345phosphorylation and translocation in microglia. J Immunol181, 660, 2008.
58. Lee, H.J., Kim, K.S., Park, I.H., and Kim, S.U. Human neuralstem cells over-expressing VEGF provide neuroprotection,angiogenesis and functional recovery in mouse strokemodel. PLoS ONE 2, e156, 2007.
Address correspondence to:Melissa Mahoney, Ph.D.
Department of Chemical and Biological EngineeringUniversity of ColoradoBox 424, ECCH 111Boulder, CO 80309
E-mail: [email protected]
Received: June 3, 2009Accepted: December 21, 2009
Online Publication Date: February 3, 2010
PRIMARY NEURAL CELL GROWTH IN PLASMA-DERIVED FIBRIN GELS 1619