Spatial and temporal expression of Zimp7 and Zimp10 PIAS-like proteins in the developing mouse...
8
Spatial and temporal expression of Zimp7 and Zimp10 PIAS-like proteins in the developing mouse embryo Hector Rodriguez-Magada ´n, Enrique Merino, Denhı ´ Schnabel, Laura Ramı ´rez, Hilda Lomelı ´ * Departamento de Gene ´tica del Desarrollo y Fisiologı ´a Molecular, Instituto de Biotecnologı ´a, Universidad Nacional Auto ´noma de Me ´xico, Cuernavaca, Morelos 62210, Me ´xico Received 10 May 2007; received in revised form 13 October 2007; accepted 15 October 2007 Available online 24 October 2007 Abstract ZIMP7 and ZIMP10 are two novel human PIAS-like proteins that share a similarity beyond the SP-RING Zn-finger domain that characterizes the PIAS family. This extended similarity is conserved in proteins of several other species and define an independent sub- family. ZIMP10 has been shown to increase the sumoylation of the androgen receptor (AR) leading to a stimulation of AR-mediated transcription. The Drosophila tonalli (tna) is the ortholog gene of ZIMP7 and ZIMP10 and presents genetic interactions with the SWI–SNF complex. Mutations in the tna gene produce flies with homeotic phenotypes. In this study, we determined the spatial–temporal expression pattern of Zimp7 and Zimp10 in mouse embryos from embryonic day 7.5 (E7.5), to mid-gestation. We found that these two genes are extensively expressed during these embryonic days and present partially overlapping patterns with a predomination of the tran- scripts in the neural tissues at early stages and a drop of expression at E12.5. Unlike other PIAS proteins, the tonalli-related Zimp genes might be essential for development. Comparison of conserved motifs in Zimp7 and Zimp10 protein sequences identified characteristic family domains that might be related to their specific biological roles, besides their common role previously identified in the sumoylation pathway. Ó 2007 Elsevier B.V. All rights reserved. Keywords: Zimp proteins; Sumoylation; X-SPRING; Mouse embryos; Tonalli; Phylogenetic analysis; SP-RING; Zn-finger; PIAS-like proteins 1. Results and discussion ZMIZ2 (ZIMP7) and ZMIZ1 (ZIMP10) are PIAS-like proteins that have been described as androgen receptor (AR) co-regulators (Beliakoff and Sun, 2006). Members of the PIAS [protein inhibitor of activated STAT (signal transducer and activator of transcription)] family are char- acterized by the presence of a zinc finger domain termed SP-RING, which has been associated with the sumoylation process. The PIAS proteins were first identified as negative regulators of the JAK-STAT pathway and are evolution- arily conserved from flies to humans. Now, it is established that they participate in a wide variety of signaling path- ways including P53, Wnt and steroid hormone receptor sig- naling (reviewed in Schmidt and Muller, 2003). In Drosophila, TnaA protein encoded by the tonalli (tna) gene shows important similarity with ZIMP7 and ZIMP10 in a region that extends beyond the SP-RING, suggesting that these Zimp proteins could be orthologs of TnaA. The common domain shared by these three proteins was called X-SPRING (eXtended SP-RING) domain and defines a subfamily of the PIAS group (Gutierrez et al., 2003). The tna gene had been previously identified by its genetic interactions with the SWI/SNF chromatin-remod- eling complex (Gutierrez et al., 2003). Recent studies have also demonstrated that ZIMP7 physically interacts with essential components of the SWI/SNF complex, including the catalytic subunit Brg1 (Huang et al., 2005). On the 1567-133X/$ - see front matter Ó 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.modgep.2007.10.005 * Corresponding author. Tel.: +52 73 29 1663; fax: +52 73 17 2388. E-mail address: [email protected] (H. Lomelı ´). www.elsevier.com/locate/gep Gene Expression Patterns 8 (2008) 206–213
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Spatial and temporal expression of Zimp7 and Zimp10 PIAS-like proteins in the developing mouse...

www.elsevier.com/locate/gep
Gene Expression Patterns 8 (2008) 206–213
Spatial and temporal expression of Zimp7 and Zimp10PIAS-like proteins in the developing mouse embryo
Hector Rodriguez-Magadan, Enrique Merino, Denhı Schnabel,Laura Ramırez, Hilda Lomelı *
Departamento de Genetica del Desarrollo y Fisiologıa Molecular, Instituto de Biotecnologıa, Universidad Nacional Autonoma de Mexico,
Cuernavaca, Morelos 62210, Mexico
Received 10 May 2007; received in revised form 13 October 2007; accepted 15 October 2007Available online 24 October 2007
Abstract
ZIMP7 and ZIMP10 are two novel human PIAS-like proteins that share a similarity beyond the SP-RING Zn-finger domain thatcharacterizes the PIAS family. This extended similarity is conserved in proteins of several other species and define an independent sub-family. ZIMP10 has been shown to increase the sumoylation of the androgen receptor (AR) leading to a stimulation of AR-mediatedtranscription. The Drosophila tonalli (tna) is the ortholog gene of ZIMP7 and ZIMP10 and presents genetic interactions with theSWI–SNF complex. Mutations in the tna gene produce flies with homeotic phenotypes. In this study, we determined the spatial–temporalexpression pattern of Zimp7 and Zimp10 in mouse embryos from embryonic day 7.5 (E7.5), to mid-gestation. We found that these twogenes are extensively expressed during these embryonic days and present partially overlapping patterns with a predomination of the tran-scripts in the neural tissues at early stages and a drop of expression at E12.5. Unlike other PIAS proteins, the tonalli-related Zimp genesmight be essential for development. Comparison of conserved motifs in Zimp7 and Zimp10 protein sequences identified characteristicfamily domains that might be related to their specific biological roles, besides their common role previously identified in the sumoylationpathway.� 2007 Elsevier B.V. All rights reserved.
Keywords: Zimp proteins; Sumoylation; X-SPRING; Mouse embryos; Tonalli; Phylogenetic analysis; SP-RING; Zn-finger; PIAS-like proteins
1. Results and discussion
ZMIZ2 (ZIMP7) and ZMIZ1 (ZIMP10) are PIAS-likeproteins that have been described as androgen receptor(AR) co-regulators (Beliakoff and Sun, 2006). Membersof the PIAS [protein inhibitor of activated STAT (signaltransducer and activator of transcription)] family are char-acterized by the presence of a zinc finger domain termedSP-RING, which has been associated with the sumoylationprocess. The PIAS proteins were first identified as negativeregulators of the JAK-STAT pathway and are evolution-arily conserved from flies to humans. Now, it is establishedthat they participate in a wide variety of signaling path-
1567-133X/$ - see front matter � 2007 Elsevier B.V. All rights reserved.
doi:10.1016/j.modgep.2007.10.005
* Corresponding author. Tel.: +52 73 29 1663; fax: +52 73 17 2388.E-mail address: [email protected] (H. Lomelı).
ways including P53, Wnt and steroid hormone receptor sig-naling (reviewed in Schmidt and Muller, 2003).
In Drosophila, TnaA protein encoded by the tonalli (tna)gene shows important similarity with ZIMP7 and ZIMP10in a region that extends beyond the SP-RING, suggestingthat these Zimp proteins could be orthologs of TnaA.The common domain shared by these three proteins wascalled X-SPRING (eXtended SP-RING) domain anddefines a subfamily of the PIAS group (Gutierrez et al.,2003).
The tna gene had been previously identified by itsgenetic interactions with the SWI/SNF chromatin-remod-eling complex (Gutierrez et al., 2003). Recent studies havealso demonstrated that ZIMP7 physically interacts withessential components of the SWI/SNF complex, includingthe catalytic subunit Brg1 (Huang et al., 2005). On the
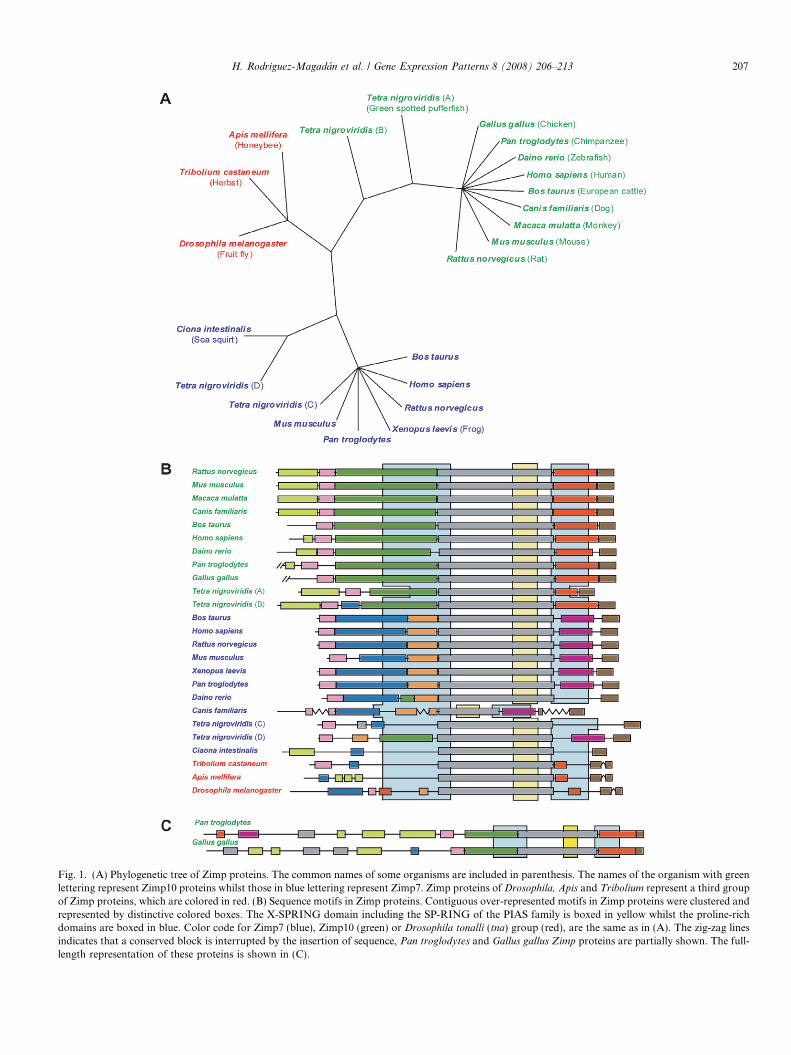
Fig. 1. (A) Phylogenetic tree of Zimp proteins. The common names of some organisms are included in parenthesis. The names of the organism with greenlettering represent Zimp10 proteins whilst those in blue lettering represent Zimp7. Zimp proteins of Drosophila, Apis and Tribolium represent a third groupof Zimp proteins, which are colored in red. (B) Sequence motifs in Zimp proteins. Contiguous over-represented motifs in Zimp proteins were clustered andrepresented by distinctive colored boxes. The X-SPRING domain including the SP-RING of the PIAS family is boxed in yellow whilst the proline-richdomains are boxed in blue. Color code for Zimp7 (blue), Zimp10 (green) or Drosophila tonalli (tna) group (red), are the same as in (A). The zig-zag linesindicates that a conserved block is interrupted by the insertion of sequence, Pan troglodytes and Gallus gallus Zimp proteins are partially shown. The full-length representation of these proteins is shown in (C).
H. Rodriguez-Magadan et al. / Gene Expression Patterns 8 (2008) 206–213 207

Fig. 2. Logo representation of conserved motifs common to all Zimp proteins. Aligned blocks of contiguous sequences present in Zimp proteins were usedas input to the LOGO web server at http://weblogo.berkeley.edu/ (Crooks et al., 2004) to obtain the representation of the relative frequency of each aminoacid in the motifs. Note that many positions in these blocks are highly conserved, thus they have the maximum possible frequency value of 100%. Colorcode representation for Proline-rich region is boxed in blue whilst the X-SPRING domain is boxed in red. The consensus bases of the SP-RING aremarked by asterisks. Polar amino acids (G, S, T, Y, C, Q, N) are shown in blue; basic amino acids (K, R, H) are shown in green; acidic residues (D, E) areshown in purple whilst hydrophobic amino acids (A, V, L, I, P, W, F, M) are shown in black. Proline residues are marked in red.
208 H. Rodriguez-Magadan et al. / Gene Expression Patterns 8 (2008) 206–213
other hand, Sharma et al., 2003 showed that both ZIMP7and ZIMP10 colocalise with SUMO in the nucleus ofhuman prostate epithelial cells and form a replication com-plex at replication foci (Sharma et al., 2003). In addition,ZIMP10 was also shown to enhance the sumoylation ofthe AR and augment its transcriptional activity in severalcell lines. These data suggest that ZIMP proteins couldbe regulating the transcription of specific target genes byaltering their chromatin structure through the sumoylationof specific subunits of the SWI/SNF complex.
In Drosophila, mutations in the tna gene affect theexpression of homeotic genes like Antennapedia, Ultrabi-
thorax and Sex combs reduced (Gutierrez et al., 2003).Also, it was recently reported that ZIMP10 enhanced thetranscriptional activity of Smad3 suggesting a connectionbetween ZIMP10 and the TGFb/Smad signaling pathway(Li et al., 2006). These observations together with theinvolvement of ZIMP7 and ZIMP10 in the transcriptional
regulation of nuclear receptors predict that the mammalianZimp7 and Zimp10 proteins will show important roles dur-ing embryonic development.
In the mouse genome Zimp7 and Zimp10 are presentand share 87% and 97% of similarity with their correspond-ing human counterparts. In this study, we analyzed Zimp7
and Zimp10 mRNA expression patterns in mice embryos.We found that the expression patterns of these genes arepartially overlapping: at early stages, the transcripts forboth of these proteins were strongly enriched in the neuraltissue, by mid-gestation the sharpest expression was foundwith a broad distribution throughout the embryo of bothtranscripts; after this stage the expression of Zimp7 andZimp10 mRNAs started to decline and nearly turned offby E12.5 where both transcripts were present in the limbs,somites and few brain regions. This pattern is consistentwith a participation of the mouse Zimp proteins duringembryonic development.
H. Rodriguez-Magadan et al. / Gene Expression Patterns 8 (2008) 206–213 209
1.1. Phylogenetic and structural relationships among
X-SPRING subfamily genes of different species
In order to have an overall view of the primary struc-ture of the Zimp proteins, a set of 25 non-redundantsequences were aligned using the MUSCLE program(Edgar, 2004). Three main kind of regions in the alignedsequences were identified: (a) Highly conserved regionspresent in all the set of Zimp7, Zimp10 or the Drosophila
melanogaster TnaA protein homologs; (b) Highly con-served regions shared only by Zimp7 or Zimp10 homo-logs; (c) Not conserved sequence regions. To identifymore precisely over-represented motifs in Zimp proteins,we used the public domain motif discovery tool MEME(Bailey and Elkan, 1994). Although many of the con-served blocks identified in our analysis were found to bepresent in all Zimp proteins, distinctive blocks predomi-nantly found in Zimp7 or Zimp10 were also found. Thepresence (value 1) or the absence (value 0) of these motifswas utilized in the CLIQUE program (Felsenstein, 1995)to construct a phylogenetic tree (Fig. 1A). Interestingly,this simplified representation of the information carriedby Zimp sequences (only 30 binary data per sequence)was sufficient to obtain a congruent tree of this family
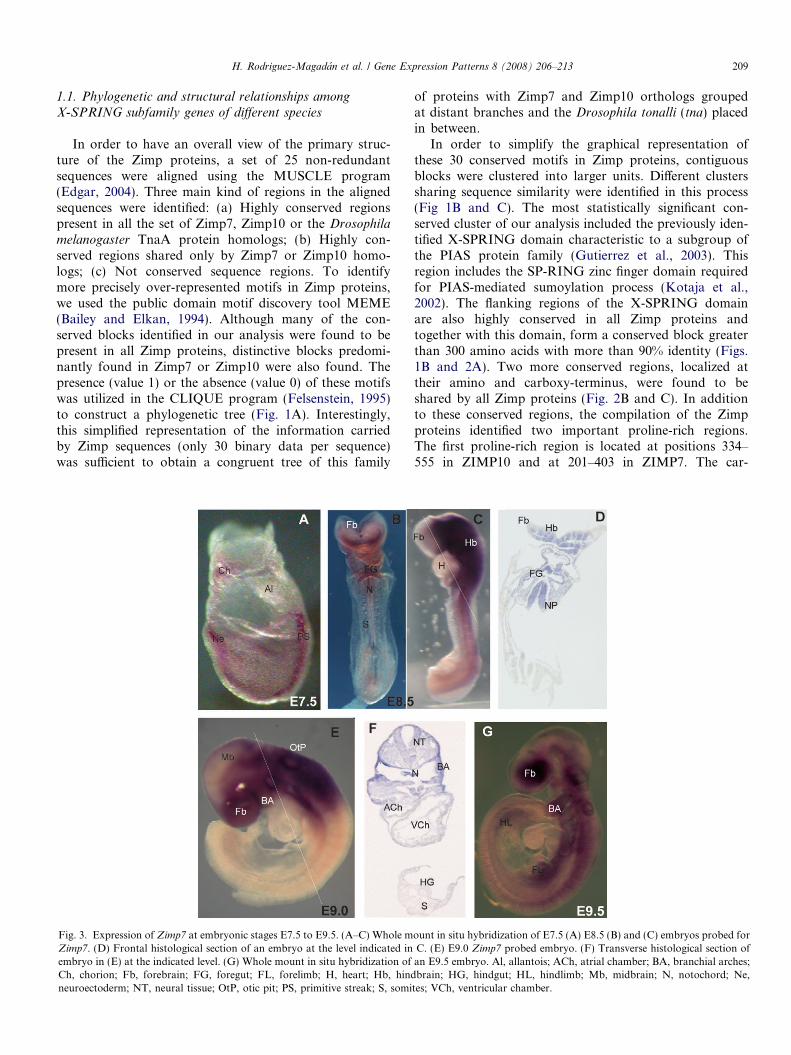
Fig. 3. Expression of Zimp7 at embryonic stages E7.5 to E9.5. (A–C) Whole mZimp7. (D) Frontal histological section of an embryo at the level indicated inembryo in (E) at the indicated level. (G) Whole mount in situ hybridization ofCh, chorion; Fb, forebrain; FG, foregut; FL, forelimb; H, heart; Hb, hindneuroectoderm; NT, neural tissue; OtP, otic pit; PS, primitive streak; S, somi
of proteins with Zimp7 and Zimp10 orthologs groupedat distant branches and the Drosophila tonalli (tna) placedin between.
In order to simplify the graphical representation ofthese 30 conserved motifs in Zimp proteins, contiguousblocks were clustered into larger units. Different clusterssharing sequence similarity were identified in this process(Fig 1B and C). The most statistically significant con-served cluster of our analysis included the previously iden-tified X-SPRING domain characteristic to a subgroup ofthe PIAS protein family (Gutierrez et al., 2003). Thisregion includes the SP-RING zinc finger domain requiredfor PIAS-mediated sumoylation process (Kotaja et al.,2002). The flanking regions of the X-SPRING domainare also highly conserved in all Zimp proteins andtogether with this domain, form a conserved block greaterthan 300 amino acids with more than 90% identity (Figs.1B and 2A). Two more conserved regions, localized attheir amino and carboxy-terminus, were found to beshared by all Zimp proteins (Fig. 2B and C). In additionto these conserved regions, the compilation of the Zimpproteins identified two important proline-rich regions.The first proline-rich region is located at positions 334–555 in ZIMP10 and at 201–403 in ZIMP7. The car-
ount in situ hybridization of E7.5 (A) E8.5 (B) and (C) embryos probed forC. (E) E9.0 Zimp7 probed embryo. (F) Transverse histological section ofan E9.5 embryo. Al, allantois; ACh, atrial chamber; BA, branchial arches;brain; HG, hindgut; HL, hindlimb; Mb, midbrain; N, notochord; Ne,
tes; VCh, ventricular chamber.
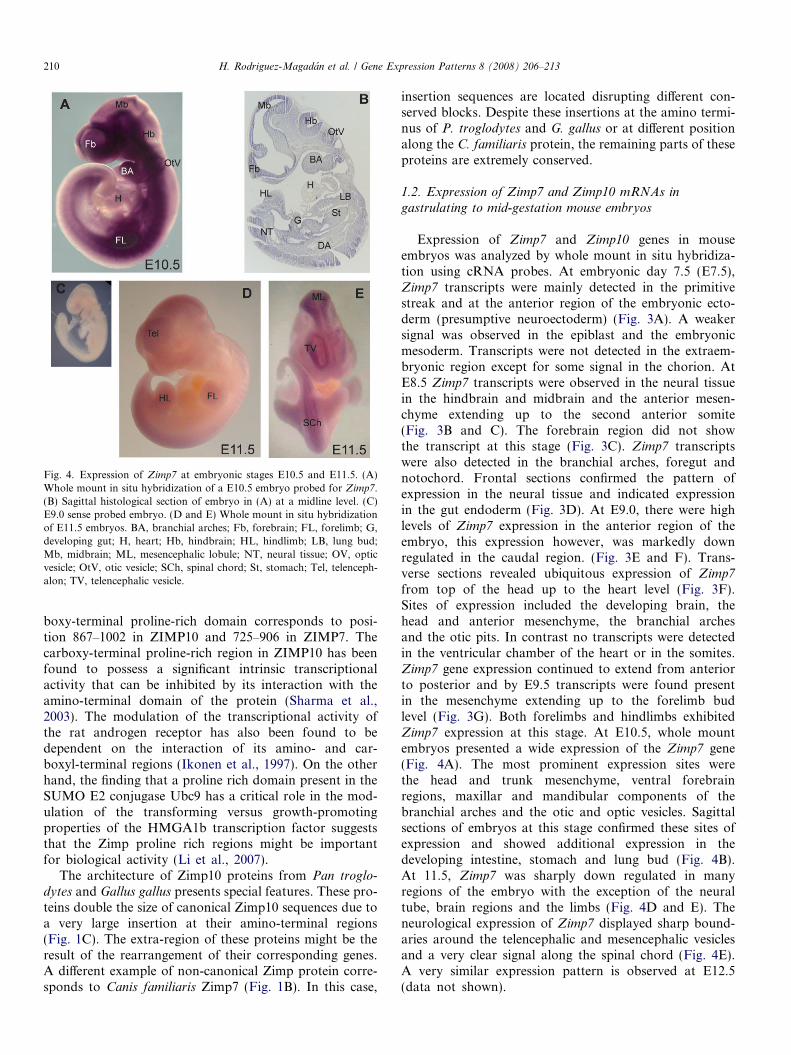
Fig. 4. Expression of Zimp7 at embryonic stages E10.5 and E11.5. (A)Whole mount in situ hybridization of a E10.5 embryo probed for Zimp7.(B) Sagittal histological section of embryo in (A) at a midline level. (C)E9.0 sense probed embryo. (D and E) Whole mount in situ hybridizationof E11.5 embryos. BA, branchial arches; Fb, forebrain; FL, forelimb; G,developing gut; H, heart; Hb, hindbrain; HL, hindlimb; LB, lung bud;Mb, midbrain; ML, mesencephalic lobule; NT, neural tissue; OV, opticvesicle; OtV, otic vesicle; SCh, spinal chord; St, stomach; Tel, telenceph-alon; TV, telencephalic vesicle.
210 H. Rodriguez-Magadan et al. / Gene Expression Patterns 8 (2008) 206–213
boxy-terminal proline-rich domain corresponds to posi-tion 867–1002 in ZIMP10 and 725–906 in ZIMP7. Thecarboxy-terminal proline-rich region in ZIMP10 has beenfound to possess a significant intrinsic transcriptionalactivity that can be inhibited by its interaction with theamino-terminal domain of the protein (Sharma et al.,2003). The modulation of the transcriptional activity ofthe rat androgen receptor has also been found to bedependent on the interaction of its amino- and car-boxyl-terminal regions (Ikonen et al., 1997). On the otherhand, the finding that a proline rich domain present in theSUMO E2 conjugase Ubc9 has a critical role in the mod-ulation of the transforming versus growth-promotingproperties of the HMGA1b transcription factor suggeststhat the Zimp proline rich regions might be importantfor biological activity (Li et al., 2007).
The architecture of Zimp10 proteins from Pan troglo-
dytes and Gallus gallus presents special features. These pro-teins double the size of canonical Zimp10 sequences due toa very large insertion at their amino-terminal regions(Fig. 1C). The extra-region of these proteins might be theresult of the rearrangement of their corresponding genes.A different example of non-canonical Zimp protein corre-sponds to Canis familiaris Zimp7 (Fig. 1B). In this case,
insertion sequences are located disrupting different con-served blocks. Despite these insertions at the amino termi-nus of P. troglodytes and G. gallus or at different positionalong the C. familiaris protein, the remaining parts of theseproteins are extremely conserved.
1.2. Expression of Zimp7 and Zimp10 mRNAs in
gastrulating to mid-gestation mouse embryos
Expression of Zimp7 and Zimp10 genes in mouseembryos was analyzed by whole mount in situ hybridiza-tion using cRNA probes. At embryonic day 7.5 (E7.5),Zimp7 transcripts were mainly detected in the primitivestreak and at the anterior region of the embryonic ecto-derm (presumptive neuroectoderm) (Fig. 3A). A weakersignal was observed in the epiblast and the embryonicmesoderm. Transcripts were not detected in the extraem-bryonic region except for some signal in the chorion. AtE8.5 Zimp7 transcripts were observed in the neural tissuein the hindbrain and midbrain and the anterior mesen-chyme extending up to the second anterior somite(Fig. 3B and C). The forebrain region did not showthe transcript at this stage (Fig. 3C). Zimp7 transcriptswere also detected in the branchial arches, foregut andnotochord. Frontal sections confirmed the pattern ofexpression in the neural tissue and indicated expressionin the gut endoderm (Fig. 3D). At E9.0, there were highlevels of Zimp7 expression in the anterior region of theembryo, this expression however, was markedly downregulated in the caudal region. (Fig. 3E and F). Trans-verse sections revealed ubiquitous expression of Zimp7
from top of the head up to the heart level (Fig. 3F).Sites of expression included the developing brain, thehead and anterior mesenchyme, the branchial archesand the otic pits. In contrast no transcripts were detectedin the ventricular chamber of the heart or in the somites.Zimp7 gene expression continued to extend from anteriorto posterior and by E9.5 transcripts were found presentin the mesenchyme extending up to the forelimb budlevel (Fig. 3G). Both forelimbs and hindlimbs exhibitedZimp7 expression at this stage. At E10.5, whole mountembryos presented a wide expression of the Zimp7 gene(Fig. 4A). The most prominent expression sites werethe head and trunk mesenchyme, ventral forebrainregions, maxillar and mandibular components of thebranchial arches and the otic and optic vesicles. Sagittalsections of embryos at this stage confirmed these sites ofexpression and showed additional expression in thedeveloping intestine, stomach and lung bud (Fig. 4B).At 11.5, Zimp7 was sharply down regulated in manyregions of the embryo with the exception of the neuraltube, brain regions and the limbs (Fig. 4D and E). Theneurological expression of Zimp7 displayed sharp bound-aries around the telencephalic and mesencephalic vesiclesand a very clear signal along the spinal chord (Fig. 4E).A very similar expression pattern is observed at E12.5(data not shown).
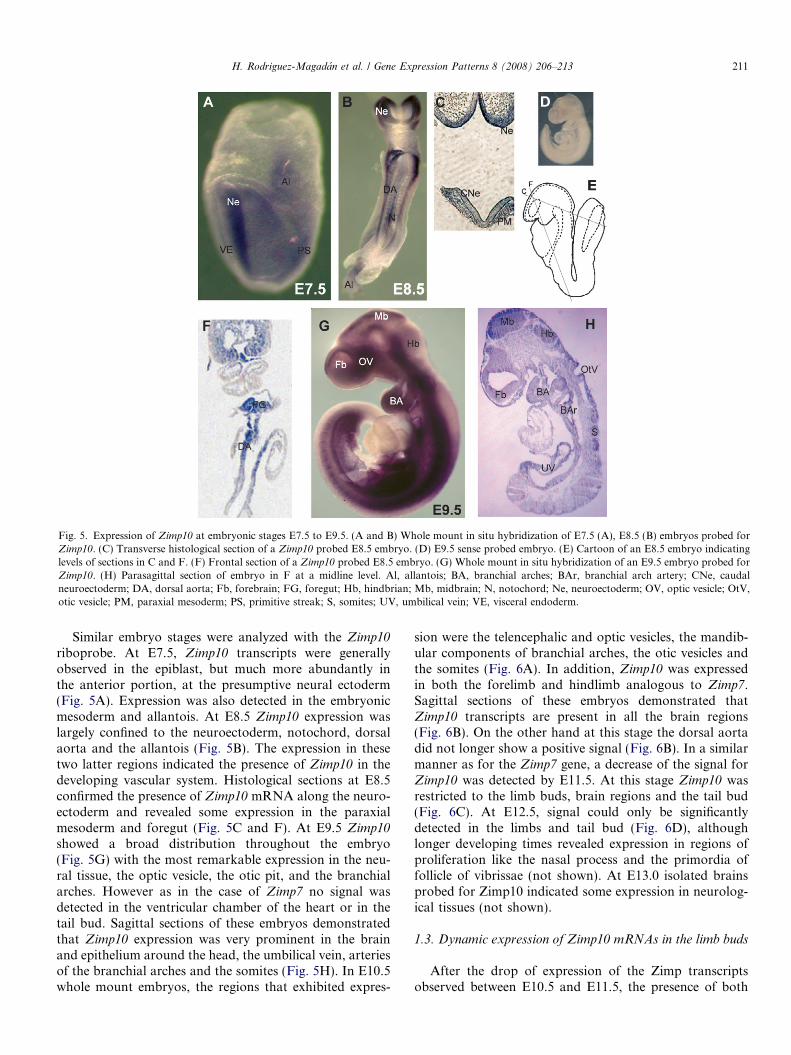
Fig. 5. Expression of Zimp10 at embryonic stages E7.5 to E9.5. (A and B) Whole mount in situ hybridization of E7.5 (A), E8.5 (B) embryos probed forZimp10. (C) Transverse histological section of a Zimp10 probed E8.5 embryo. (D) E9.5 sense probed embryo. (E) Cartoon of an E8.5 embryo indicatinglevels of sections in C and F. (F) Frontal section of a Zimp10 probed E8.5 embryo. (G) Whole mount in situ hybridization of an E9.5 embryo probed forZimp10. (H) Parasagittal section of embryo in F at a midline level. Al, allantois; BA, branchial arches; BAr, branchial arch artery; CNe, caudalneuroectoderm; DA, dorsal aorta; Fb, forebrain; FG, foregut; Hb, hindbrian; Mb, midbrain; N, notochord; Ne, neuroectoderm; OV, optic vesicle; OtV,otic vesicle; PM, paraxial mesoderm; PS, primitive streak; S, somites; UV, umbilical vein; VE, visceral endoderm.
H. Rodriguez-Magadan et al. / Gene Expression Patterns 8 (2008) 206–213 211
Similar embryo stages were analyzed with the Zimp10
riboprobe. At E7.5, Zimp10 transcripts were generallyobserved in the epiblast, but much more abundantly inthe anterior portion, at the presumptive neural ectoderm(Fig. 5A). Expression was also detected in the embryonicmesoderm and allantois. At E8.5 Zimp10 expression waslargely confined to the neuroectoderm, notochord, dorsalaorta and the allantois (Fig. 5B). The expression in thesetwo latter regions indicated the presence of Zimp10 in thedeveloping vascular system. Histological sections at E8.5confirmed the presence of Zimp10 mRNA along the neuro-ectoderm and revealed some expression in the paraxialmesoderm and foregut (Fig. 5C and F). At E9.5 Zimp10showed a broad distribution throughout the embryo(Fig. 5G) with the most remarkable expression in the neu-ral tissue, the optic vesicle, the otic pit, and the branchialarches. However as in the case of Zimp7 no signal wasdetected in the ventricular chamber of the heart or in thetail bud. Sagittal sections of these embryos demonstratedthat Zimp10 expression was very prominent in the brainand epithelium around the head, the umbilical vein, arteriesof the branchial arches and the somites (Fig. 5H). In E10.5whole mount embryos, the regions that exhibited expres-
sion were the telencephalic and optic vesicles, the mandib-ular components of branchial arches, the otic vesicles andthe somites (Fig. 6A). In addition, Zimp10 was expressedin both the forelimb and hindlimb analogous to Zimp7.Sagittal sections of these embryos demonstrated thatZimp10 transcripts are present in all the brain regions(Fig. 6B). On the other hand at this stage the dorsal aortadid not longer show a positive signal (Fig. 6B). In a similarmanner as for the Zimp7 gene, a decrease of the signal forZimp10 was detected by E11.5. At this stage Zimp10 wasrestricted to the limb buds, brain regions and the tail bud(Fig. 6C). At E12.5, signal could only be significantlydetected in the limbs and tail bud (Fig. 6D), althoughlonger developing times revealed expression in regions ofproliferation like the nasal process and the primordia offollicle of vibrissae (not shown). At E13.0 isolated brainsprobed for Zimp10 indicated some expression in neurolog-ical tissues (not shown).
1.3. Dynamic expression of Zimp10 mRNAs in the limb buds
After the drop of expression of the Zimp transcriptsobserved between E10.5 and E11.5, the presence of both
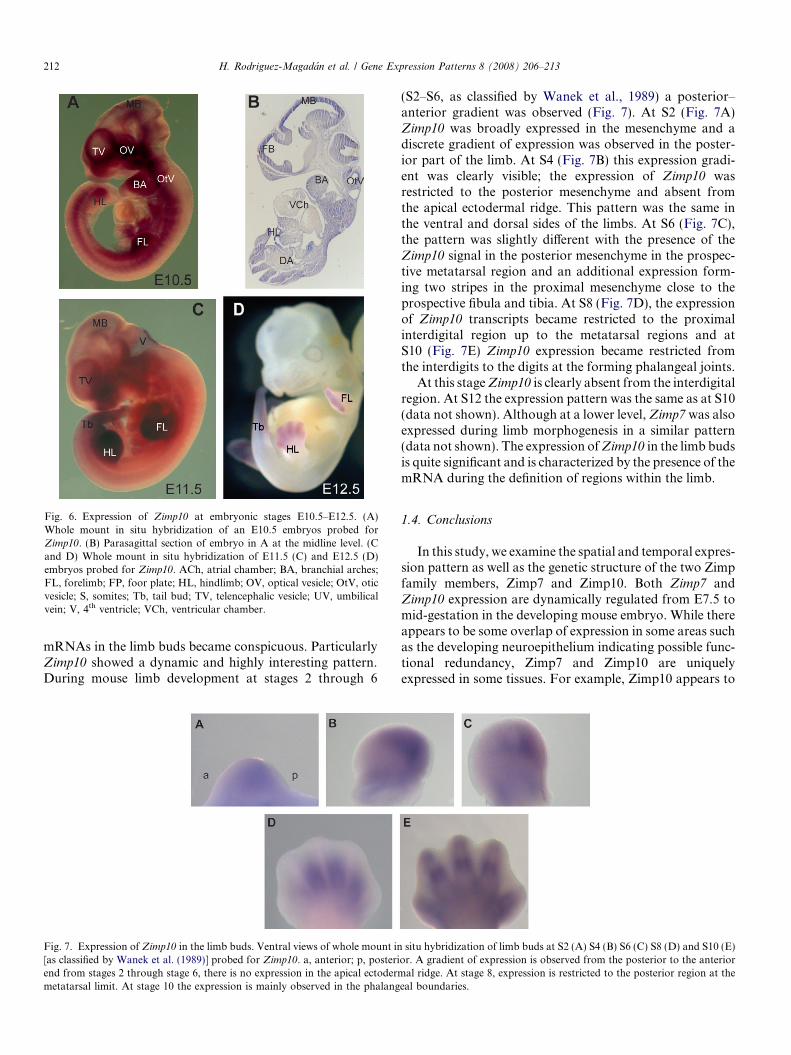
Fig. 6. Expression of Zimp10 at embryonic stages E10.5–E12.5. (A)Whole mount in situ hybridization of an E10.5 embryos probed forZimp10. (B) Parasagittal section of embryo in A at the midline level. (Cand D) Whole mount in situ hybridization of E11.5 (C) and E12.5 (D)embryos probed for Zimp10. ACh, atrial chamber; BA, branchial arches;FL, forelimb; FP, foor plate; HL, hindlimb; OV, optical vesicle; OtV, oticvesicle; S, somites; Tb, tail bud; TV, telencephalic vesicle; UV, umbilicalvein; V, 4th ventricle; VCh, ventricular chamber.
212 H. Rodriguez-Magadan et al. / Gene Expression Patterns 8 (2008) 206–213
mRNAs in the limb buds became conspicuous. ParticularlyZimp10 showed a dynamic and highly interesting pattern.During mouse limb development at stages 2 through 6
Fig. 7. Expression of Zimp10 in the limb buds. Ventral views of whole mount in[as classified by Wanek et al. (1989)] probed for Zimp10. a, anterior; p, posteriend from stages 2 through stage 6, there is no expression in the apical ectodermmetatarsal limit. At stage 10 the expression is mainly observed in the phalang
(S2–S6, as classified by Wanek et al., 1989) a posterior–anterior gradient was observed (Fig. 7). At S2 (Fig. 7A)Zimp10 was broadly expressed in the mesenchyme and adiscrete gradient of expression was observed in the poster-ior part of the limb. At S4 (Fig. 7B) this expression gradi-ent was clearly visible; the expression of Zimp10 wasrestricted to the posterior mesenchyme and absent fromthe apical ectodermal ridge. This pattern was the same inthe ventral and dorsal sides of the limbs. At S6 (Fig. 7C),the pattern was slightly different with the presence of theZimp10 signal in the posterior mesenchyme in the prospec-tive metatarsal region and an additional expression form-ing two stripes in the proximal mesenchyme close to theprospective fibula and tibia. At S8 (Fig. 7D), the expressionof Zimp10 transcripts became restricted to the proximalinterdigital region up to the metatarsal regions and atS10 (Fig. 7E) Zimp10 expression became restricted fromthe interdigits to the digits at the forming phalangeal joints.
At this stage Zimp10 is clearly absent from the interdigitalregion. At S12 the expression pattern was the same as at S10(data not shown). Although at a lower level, Zimp7 was alsoexpressed during limb morphogenesis in a similar pattern(data not shown). The expression of Zimp10 in the limb budsis quite significant and is characterized by the presence of themRNA during the definition of regions within the limb.
1.4. Conclusions
In this study, we examine the spatial and temporal expres-sion pattern as well as the genetic structure of the two Zimpfamily members, Zimp7 and Zimp10. Both Zimp7 andZimp10 expression are dynamically regulated from E7.5 tomid-gestation in the developing mouse embryo. While thereappears to be some overlap of expression in some areas suchas the developing neuroepithelium indicating possible func-tional redundancy, Zimp7 and Zimp10 are uniquelyexpressed in some tissues. For example, Zimp10 appears to
situ hybridization of limb buds at S2 (A) S4 (B) S6 (C) S8 (D) and S10 (E)or. A gradient of expression is observed from the posterior to the anterior
al ridge. At stage 8, expression is restricted to the posterior region at theeal boundaries.

H. Rodriguez-Magadan et al. / Gene Expression Patterns 8 (2008) 206–213 213
be present in the developing vascular system at early stageswith expression in the dorsal aorta and the allantois. UnlikeZimp7, Zimp10 is also expressed in the paraxial mesodermand somites. At E7.5, we show that Zimp7 possess restrictedexpression in the primitive streak, while at mid-gestation weobserve expression in the spinal chord and brain lobules. Ofinterest is the dynamic of expression of Zimp7 that extendsfrom anterior to posterior as development proceeds and sug-gests a particular transcriptional organization. Functionalwork done for the Zimp proteins indicates that at leastZimp10 has specific roles during transcriptional regulation(Li et al., 2006). Therefore, considering the strong activityof both genes during embryogenesis it is possible that thesetwo genes are important for development. The analysis ofZimp7 and Zimp10 sequences shows that these proteinsshare common regions of highly conserved sequence in addi-tion to the SP-RING domain that characterizes the PIASfamily. Besides these common conserved regions to all Zimpproteins, Zimp7 and Zimp10 proteins present other sequencemotifs specific to each of these variants. The phylogeneticanalysis of Zimp proteins locates the previously character-ized D. tonalli (tna) between the Zimp7 and Zimp10 groups.
2. Experimental procedures
2.1. Computer analyses
Using the D. melanogaster TnaA protein sequence and those from itshuman homologs, ZIMP7 and ZIMP10, all the available Zimp sequencesfrom the GenBank database were identified and retrieved using the BLASTprogram (http://www.ncbi.nlm.nih.gov/blast/). Partially sequenced orredundant versions of Zimp proteins were eliminated. After this process,the sequences of 23 Zimp proteins were obtained. In order to identifyover-represented ‘signatures’ in our set of Zimp protein sequences, we usedthe public domain motif discovery tool Multiple EM for Motif Elicitation(MEME) (Bailey and Elkan, 1994). Thirty non-overlapping motifs of aminoacid sequence carrying no-indels were identified. The presence (value 1) ofabsence (value 0) of these motifs in the sequences were used as input ofthe CLIQUE program of J. Felsentein’s PHYLIP 3.57c phylogeny inferencepackage program (Felsenstein, 1995) to obtain the largest cliques of charac-ters and the suggested tree. For simplicity, cluster of contiguous motifs weregrouped and represented by continuous boxes (Fig. 1 B). The alignment ofZimp protein sequences was performed using the MUSCLE program(Edgar, 2004). The SP-RING domain was identified using the Pfam web ser-ver at http://www.sanger.ac.uk/Software/Pfam/search.shtml.
2.2. Probe preparation
The Zimp10 probe was obtained by subcloning into pKS vector a frag-ment corresponding to bases 1–500 of the coding sequence from the com-plete cDNA clone. This vector was linearized with HinDIII for theantisense probe or NotI for the sense probe. The Zimp7 probe, corre-sponded to bases 532–1098 of the coding sequence subcloned into pKS vec-tor and linearized with HinDIII for the antisense probe and NotI for thesense probe. Both antisense riboprobes were in vitro transcribed with digox-igenin RNA labeling mix and T7 RNA polymerase (Roche), and the senseriboprobes were in vitro transcribed with T3 RNA polymerase (Roche).
2.3. Whole-mount RNA in situ hybridization
Whole embryos were fixed overnight in 4% paraformaldehyde (PFA)at 4 �C and dehydrated in a graded methanol series prior to storage at�20 �C. Whole-mount in situ hybridization was performed as described
previously (Hogan et al., 1994). Briefly, embryos were rehydrated in agraded methanol series and treated with proteinase K (10 lg/ml) (Invitro-gen). Embryos were re-fixed with 0.2% glutaraldehyde/ 4% PFA andwashed in PBT (0.1% Tween-20/PBS) prior to hybridization overnightat 68 �C. Embryos were washed twice in 2· SSC/0.2% SDS at 70 �C, fol-lowed by two identical washes in 0.2· SSC/0.2% SDS. Embryos were incu-bated overnight with anti-digoxigenin antibody (Roche) at 4 �C and afterseveral washes with TBST color was developed using NBT/BCIP (Roche).
Acknowledgements
We thank Dr. Luke Krebs for critical comments on themanuscript. We also thank Claudia Lomelı for assistancein the artwork and Marcela Ramirez and Sergio Gonzalesfor assistance with mice. This work was supported by DGAP-A-UNAM Grants IN213602-3 and CONACyT 49114 andby a sabbatical fellowship to HL by DGAPA-UNAM.
References
Bailey, T.L., Elkan, C., 1994. Fitting a mixture model by expectationmaximization to discover motifs in biopolymers. Proc. Int. Conf.Intell. Syst. Mol. Biol 2, 28–36.
Beliakoff, J., Sun, Z., 2006. Zimp7 and Zimp10, two novel PIAS-likeproteins, function as androgen receptor coregulators. Nucl. Recep.Signal 4, e017.
Crooks, G.E., Hon, G., Chandonia, J.M., Brenner, S.E., 2004. WebLogo:a sequence logo generator. Genome Res. 14, 1188–1190.
Edgar, R.C., 2004. MUSCLE: a multiple sequence alignment method withreduced time and space complexity. BMC Bioinformatics 5, 113.
Felsenstein, J., 1995. PHYLIP (Phylogeny Inference Package) version3.57c. Department of Genetics, University of Washington, Seattle.
Gutierrez, L., Zurita, M., Kennison, J.A., Vazquez, M., 2003. TheDrosophila trithorax group gene tonalli (tna) interacts genetically withthe Brahma remodeling complex and encodes an SP-RING fingerprotein. Development 130, 343–354.
Hogan, B., Beddington, R., Costantini, F., Lacy, E., 1994. Manipulatingthe Mouse Embryo: a Laboratory Manual. Cold Spring Harbor, ColdSpring Harbor Laboratory Press, New York.
Huang, C.Y., Beliakoff, J., Li, X., Lee, J., Li, X., Sharma, M., et al., 2005.hZimp7, a novel PIAS-like protein, enhances androgen receptor-mediated transcription and interacts with SWI/SNF-like BAF com-plexes. Mol. Endocrinol. 19, 2915–2929.
Ikonen, T., Palvimo, J.J., Janne, O.A., 1997. Interaction between theamino- and carboxyl-terminal regions of the rat androgen receptormodulates transcriptional activity and is influenced by nuclear receptorcoactivators. J. Biol. Chem. 272, 29821–29828.
Kotaja, N., Karvonen, U., Janne, O.A., Palvimo, J.J., 2002. PIAS proteinsmodulate transcription factors by functioning as SUMO-1 ligases.Mol. Cell. Biol. 22, 5222–5234.
Li, X., Thyssen, G., Beliakoff, J., Sun, Z., 2006. The novel PIAS-likeprotein hZimp10 enhances Smad transcriptional activity. J. Biol.Chem. 281, 23748–23756.
Li, Y., Lu, J., Prochownik, E.V., 2007. Dual role for sumo E2 conjugaseUBC9 in modulating the transforming and growth-promoting prop-erties of the HMGA1B architectural transcription factor. J. Biol.Chem. 282, 13363–13371.
Schmidt, D., Muller, S., 2003. PIAS/SUMO: new partners in transcrip-tional regulation. Cell Mol. Life Sci. 60, 2561–2574.
Sharma, M., Li, X., Wang, Y., Zarnegar, M., Huang, C.Y., Palvimo, J.J.,Lim, B., Sun, Z., 2003. hZimp10 is an androgen receptor co-activatorand forms a complex with SUMO-1 at replication foci. EMBO J. 17,6101–6114.
Wanek, N., Muneoka, K., Holler-Dinsmore, G., Burton, R., Bryant, S.V.,1989. A staging system for mouse limb development. J. Exp. Zool. 249,41–49.