
The expression of FGF receptors in the implanting mouse embryo.
11
Expression of Fibroblast Growth Factor Receptors in Peri-Implantation Mouse Embryos DANIEL A. RAPPOLEE, 1,2,3 * YOGESH PATEL, 1 AND KRISTINE JACOBSON 1 1 Department of Cell and Molecular Biology, and Department of Obstetrics and Gynecology, Northwestern University Medical School, Chicago, Illinois 2 Lurie Cancer Center, and Feinberg Cardiovascular Research Institute, Northwestern University Medical School, Chicago, Illinois 3 Center for Reproductive Sciences, Northwestern University, Evanston, Illinois ABSTRACT FGF receptor (FGFR) function is essential during peri-implantation mouse development. To understand which receptors are functioning, we tested for the expression of all four FGF receptors in peri-implantation blastocysts. By RT-PCR, FGFR-3 and FGFR-4 were detected at high levels, FGFR-2 at lower levels, and FGFR-1 was detected at background levels compared to control tissues. Because FGFR-3 and FGFR-4 were detected at the highest levels, we studied these in detail. Between 3.5 days after fertilization (E3.5) and E6.0, FGFR-4 mRNA was detected ubiqui- tously in the peri-implantation embryo, restricted to the inner cell mass (ICM) and its derivatives and primitive endoderm by E6.0, and was not detected at E6.5. FGFR-3 mRNA was detected ubiquitously in the peri- implantation embryo with a tendency towards extraem- bryonic cells. We tested blastocyst outgrowths, a model for implantation, for FGFR-3 and FGFR-4 protein. FGFR-3 protein was detected in all cells early during the outgrowth. Later, FGFR-3 was detected in the extraem- bryonic endoderm and trophoblast giant cells (TGC), but not in the ICM. FGFR-4 protein was detected in all cells of the implanting embryo, but was restricted to the ICM/primitive endoderm in later stage outgrowths. The distribution of the receptor proteins in the blastocyst outgrowths is similar to the distribution of the mRNA detected by in situ hybridization of sections of em- bryos. The data suggest roles for FGFR-3 and FGFR-4 in peri-implantation development. Mol. Reprod. Dev. 51: 254–264, 1998. r 1998 Wiley-Liss, Inc. Key Words: FGF receptors; RT-PCR; immunocyto- chemistry; in situ hybridization; blastocyst; Northern blot INTRODUCTION Fibroblast growth factor (FGF) signaling is essential for embryonic development in mice. Suppression of FGF signal reception results in a fifth cell cycle block in preimplantation mouse embryos. FGF-4 is expressed in the ICM, is required for its survival, and regulates the outgrowth of parietal endoderm and TGC in vitro (Rappolee et al., 1994; Feldman et al., 1995; Chai et al., 1998). FGF receptor expression has not been character- ized in detail in the peri-implantation embryo. There are four high-affinity, ligand-dependent FGF receptors (FGFR-1 through FGFR-4; K d 5 20–480 pM). FGFR-1 through FGFR-4 are related transmembrane receptors with intracellular split-tyrosine kinase do- mains and three immunoglobulin-like (Ig) extracellular domains. FGFR-1 through FGFR-3 have alternate spliced forms that yield alternate third Ig ectodomains, IIIb and IIIc, with the IIIb domain having narrower ligand specificity (reviewed in Ornitz et al., 1996). An FGF ligand, the FGF receptor, and a heparin sulfate proteoglycan (HSPG) form a ternary complex where dimerized tyrosine kinase domains activate the enzy- matic function of the receptors (Rappolee, 1998). Candi- date HSPG molecules, such as syndecan and perlecan, have been detected in the mouse blastocyst (Rappolee, 1998). FGF ligands also bind with high affinity to a nontyrosine kinase containing cysteine-rich FGF recep- tor (CFR) of unknown function (Burrus et al., 1992). Isoforms of all four high-affinity FGF receptors bind FGF-4 (Ornitz et al., 1996). FGFR-1 mRNA has been detected by RT-PCR, and FGFR-3 and FGFR-4 mRNA have been detected by in situ hybridization in the blastocyst (Rappolee et al,. 1994, reviewed in Deng et al., 1996; Rappolee, 1998). FGFR-2 mRNA is detected in the early postimplantation embryo (Orr-Urtreger et al., 1991), but FGFR-1 is not detected by in situ hybridiza- tion until gastrulation (Yamaguchi et al., 1992). FGFR-2 mRNA is detected in the primitive ectoderm at 5.5 days after fertilization (E5.5) through day E7.5 (Orr-Urt- reger et al. 1991). FGFR-3 and FGFR-4 mRNA have Grant sponsor: National Institutes of Health; Grant number NICHD HD30739–02; Grant sponsor: Feinberg Cardiovascular Research Insti- tute at Northwestern University, AHA USA; Grant number: 96014080; Grant sponsor: ACS Illinois division; Grant number: 95–10; Grant sponsor: The Markey Charitable Trust. K. Jacobson’s current address: Vysis Inc., 3100 Woodcreek Dr., Down- er’s Grove, Illinois 60515. *Correspondence to: Daniel A. Rappolee, Ph.D., Department of Obstet- rics and Gynecology, Northwestern University Medical School, 303 E. Chicago Avenue, Tarry Building 4–725, Chicago, Illinois 60611. E-mail: [email protected] Received 7 April 1998; Accepted 9 June 1998 MOLECULAR REPRODUCTION AND DEVELOPMENT 51:254–264 (1998) r 1998 WILEY-LISS, INC.
Transcript of The expression of FGF receptors in the implanting mouse embryo.

Expression of Fibroblast Growth Factor Receptorsin Peri-Implantation Mouse EmbryosDANIEL A. RAPPOLEE,1,2,3* YOGESH PATEL,1 AND KRISTINE JACOBSON1
1Department of Cell and Molecular Biology, and Department of Obstetrics and Gynecology, Northwestern UniversityMedical School, Chicago, Illinois2Lurie Cancer Center, and Feinberg Cardiovascular Research Institute, Northwestern University Medical School,Chicago, Illinois3Center for Reproductive Sciences, Northwestern University, Evanston, Illinois
ABSTRACT FGF receptor (FGFR) function isessential during peri-implantation mouse development.To understand which receptors are functioning, wetested for the expression of all four FGF receptors inperi-implantation blastocysts. By RT-PCR, FGFR-3 andFGFR-4 were detected at high levels, FGFR-2 at lowerlevels, and FGFR-1 was detected at background levelscompared to control tissues. Because FGFR-3 andFGFR-4 were detected at the highest levels, we studiedthese in detail. Between 3.5 days after fertilization(E3.5) and E6.0, FGFR-4 mRNA was detected ubiqui-tously in the peri-implantation embryo, restricted to theinner cell mass (ICM) and its derivatives and primitiveendoderm by E6.0, and was not detected at E6.5.FGFR-3 mRNA was detected ubiquitously in the peri-implantation embryo with a tendency towards extraem-bryonic cells. We tested blastocyst outgrowths, amodel for implantation, for FGFR-3 and FGFR-4 protein.FGFR-3 protein was detected in all cells early during theoutgrowth. Later, FGFR-3 was detected in the extraem-bryonic endoderm and trophoblast giant cells (TGC),but not in the ICM. FGFR-4 protein was detected in allcells of the implanting embryo, but was restricted to theICM/primitive endoderm in later stage outgrowths. Thedistribution of the receptor proteins in the blastocystoutgrowths is similar to the distribution of the mRNAdetected by in situ hybridization of sections of em-bryos. The data suggest roles for FGFR-3 and FGFR-4 inperi-implantation development. Mol. Reprod. Dev. 51:254–264, 1998. r 1998 Wiley-Liss, Inc.
Key Words: FGF receptors; RT-PCR; immunocyto-chemistry; in situ hybridization; blastocyst; Northernblot
INTRODUCTIONFibroblast growth factor (FGF) signaling is essential
for embryonic development in mice. Suppression ofFGF signal reception results in a fifth cell cycle block inpreimplantation mouse embryos. FGF-4 is expressed inthe ICM, is required for its survival, and regulates theoutgrowth of parietal endoderm and TGC in vitro(Rappolee et al., 1994; Feldman et al., 1995; Chai et al.,
1998). FGF receptor expression has not been character-ized in detail in the peri-implantation embryo.
There are four high-affinity, ligand-dependent FGFreceptors (FGFR-1 through FGFR-4; Kd 5 20–480 pM).FGFR-1 through FGFR-4 are related transmembranereceptors with intracellular split-tyrosine kinase do-mains and three immunoglobulin-like (Ig) extracellulardomains. FGFR-1 through FGFR-3 have alternatespliced forms that yield alternate third Ig ectodomains,IIIb and IIIc, with the IIIb domain having narrowerligand specificity (reviewed in Ornitz et al., 1996). AnFGF ligand, the FGF receptor, and a heparin sulfateproteoglycan (HSPG) form a ternary complex wheredimerized tyrosine kinase domains activate the enzy-matic function of the receptors (Rappolee, 1998). Candi-date HSPG molecules, such as syndecan and perlecan,have been detected in the mouse blastocyst (Rappolee,1998). FGF ligands also bind with high affinity to anontyrosine kinase containing cysteine-rich FGF recep-tor (CFR) of unknown function (Burrus et al., 1992).Isoforms of all four high-affinity FGF receptors bindFGF-4 (Ornitz et al., 1996). FGFR-1 mRNA has beendetected by RT-PCR, and FGFR-3 and FGFR-4 mRNAhave been detected by in situ hybridization in theblastocyst (Rappolee et al,. 1994, reviewed in Deng etal., 1996; Rappolee, 1998). FGFR-2 mRNA is detected inthe early postimplantation embryo (Orr-Urtreger et al.,1991), but FGFR-1 is not detected by in situ hybridiza-tion until gastrulation (Yamaguchi et al., 1992). FGFR-2mRNA is detected in the primitive ectoderm at 5.5 daysafter fertilization (E5.5) through day E7.5 (Orr-Urt-reger et al. 1991). FGFR-3 and FGFR-4 mRNA have
Grant sponsor: National Institutes of Health; Grant number NICHDHD30739–02; Grant sponsor: Feinberg Cardiovascular Research Insti-tute at Northwestern University, AHA USA; Grant number: 96014080;Grant sponsor: ACS Illinois division; Grant number: 95–10; Grantsponsor: The Markey Charitable Trust.K. Jacobson’s current address: Vysis Inc., 3100 Woodcreek Dr., Down-er’s Grove, Illinois 60515.*Correspondence to: Daniel A. Rappolee, Ph.D., Department of Obstet-rics and Gynecology, Northwestern University Medical School, 303 E.Chicago Avenue, Tarry Building 4–725, Chicago, Illinois 60611.E-mail: [email protected] 7 April 1998; Accepted 9 June 1998
MOLECULAR REPRODUCTION AND DEVELOPMENT 51:254–264 (1998)
r 1998 WILEY-LISS, INC.

unique expression patterns after gastrulation (Peterset al., 1992; Stark et al., 1991). FGFR-3 mRNA tran-scripts are detected in resting cartilage in the endochon-dral model for long bone and in the germinal epitheliumof the neural tube between E9.5 and E16.5 (Peters etal., 1992). FGFR-4 transcripts are expressed in defini-tive endoderm (E8.5–E9.5), in skeletal muscle, and inendodermal derivatives such as liver, pancreas, andlung (Stark et al., 1991; Partanen et al., 1992). The CFRis detected ubiquitously in chick (Burrus et al., 1992),but the functional fucosylated form of CFR is restrictedto neutrophils in mouse (Steegmaier et al., 1995); CFRhas not been studied during development. FGFR-3 andFGFR-4 mRNA and protein in the peri-implantationmouse embryo have not been studied in detail.
The FGF family consists of 15 FGF ligand (FGF-1through FGF-15) genes of related sequence (McWhirteret al., 1997; Rappolee 1998, reviewed in Chai et al.,1998). The first nine ligands are the best characterized.These heparin-binding factors are between 14 kDa and28 kDa. Some are secreted (FGF-4 through FGF-9),while some lack the signal sequences required forsecretion (FGF-1 through FGF-3). FGF-10, FGF-8, andFGF-4 mRNA, but not FGF-2, FGF-3, and FGF-7mRNA, have been detected in the pre-implantationmouse embryo (Rappolee et al., 1988, 1994; Rappolee,1998; Rappolee and Changyaleket, unpublished).
To understand the essential role of FGF signaling inmouse development, we report here the expression ofFGFR-3 and FGFR-4 mRNA and protein in the implant-ing mouse embryo.
MATERIALS AND METHODSMaterials
Amplitaq DNA polymerase was purchased from Per-kin-Elmer Cetus (Norwalk, CT). Restriction enzymesand T4 polynucleotide kinase were purchased fromNew England BioLabs (Beverly, MA) and Gibco/BRL(Gaithersburg, MD). Radioisotopes were obtained fromNew England Nuclear (Boston, MA). Moloney murineleukemia virus (MMLV) reverse transcriptase was ob-tained from Gibco/BRL. Oligonucleotides for the poly-merase chain reaction (PCR) were synthesized on anApplied Biosystems (Foster City, CA) PCR-Mate oligo-synthesizer. Digoxygenin-UTP Genius RNA labelingkit, antidigoxygenin-alkaline phosphatase conjugate,nitro blue tetrazolium (NBT), and X-Phosphate (5-bromo-4-chloro-3-indolyl phosphate) were all from Boeh-ringer-Mannheim (Indianapolis, IN). Hoechst 33258was purchased from Eastman Kodak (Rochester, NY).All other chemicals were purchased from Sigma Chemi-cal (St. Louis, MO) or VWR (Chicago, IL). The agarosewas purchased from FMC (Rockland, ME).
DNACFR was a 1,600 bp XhoI-Eco RI fragment of the
chicken CFR (from 2,253–3,857 bp; Burrus et al., 1992)excised from pBSSK. Actin was a 1.1 kb Pst I fragmentof chicken b-actin (from 2,271–3,295 bp; Kost et al.,
1983). FGFR-4 was a 133 bp fragment in the 38untranslated region (UTR) (2,888–3,020 bp, Stark etal., 1991) and FGFR-3 was a 146 bp fragment from the38 UTR of mouse FGFR-3 (2,677–2,833 bp; Avivi et al.,1991). The FGFR-3 and -4 probes were constructed andused previously (Rappolee et al., 1994; Puscheck et al.,1997).
Mouse Eggs and EmbryosStandard techniques were used for obtaining eggs
and zygotes (Hogan et al., 1994). Female CD-1 (6–10weeks old, Charles River, Wilmington, MA) or MF-1(Harlan Sprague Dawley, Indianapolis, IN) mice weresuperovulated and embryos obtained as previouslydescribed (Rappolee et al., 1994; Puscheck et al., 1997).Blastocyst outgrowth culture was described previously(Puscheck et al., 1998).
Cell CultureEmbryonal carcinoma F9 cells, obtained from the
ATCC, were cultured to obtain undifferentiated anddifferentiated cells as described (Robertson, 1987; Rap-polee et al., 1994; Puscheck et al., 1998). D3 embryonicstem cells were obtained from Dr. Doestchman anddifferentiated in monolayer culture after removal ofleukemia inhibitory factor (LIF) as described(Doetschman et al., 1985; Mummery et al., 1990, 1993).C3H/NE trophectoderm carcinoma cells were obtainedfrom Dr. Damjanov (Wistar Institute) and cultured togive cystic bods spontaneously expressing trophectoder-mal characteristics (Damjanov and Damjanov, 1985).
RNA PreparationFor RT-PCR, total RNA from 300 embryos was iso-
lated as previously described (Rappolee et al., 1994).Twenty µg of E. coli rRNA was added as a carrier beforeCsCl gradient ultracentrifugation and the concentra-tion of RNA was determined by measurement of absor-bance at 260 nm. For Northern blot hybridization, RNAwas collected from cells or tissues using standardtechniques (Ausubel et al., 1993).
Reverse Transcription PolymeraseChain Reaction (RT-PCR)
RT-PCR was done essentially as described previously(Rappolee et al., 1994; Puscheck et al., 1997) with thefollowing modifications: In early studies on FGF recep-tor expression, reaction mixtures were heat-denaturedand flash-cooled; 50 units of additional buffered MMLVreverse transcriptase (RT) was added, and the RTmixture was reincubated. The proportion of the RTmixture that was equivalent to 30 embryos was addedto sequence-specific primed (see Table 1; design ofprimers) PCR reaction mixtures (Rappolee et al., 1994).The mixture was amplified for 50 cycles on a Perkin-Elmer Cetus Thermocycler 9600 programmable heat-ing block. One tenth of this first PCR was reamplifiedusing a pair of primers within the first set for another50 cycles (Table 1). All RT reactions were checked withprimers to actin (which span introns) for contamination
FGF RECEPTORS IN MOUSE EMBRYOS 255
of the RNA prep with DNA. RT-PCR performed onRNase-treated embryo RNA was always negative. Frag-ments were verified by size, restriction enzyme map-ping, and all amplimers were subcloned into PCRII asdescribed previously (Rappolee et al., 1988, 1994; Pus-check et al., 1997).
Northern Blot HybridizationWhole RNA was purified as described previously
(Puscheck et al., 1997) and purified by oligo dT cellulosecolumn (Life technologies, Grand Island, NY), 5 µg ofpoly A positive RNA was fractionated on a 1.0% agarosegel, blotted onto a nylon membrane, hybridized withrandom primed antisense probe at 109 cpm/µg(RadPrime kit, Life technologies, Grand Island NY),and exposed to Kodak Xmat film for 5 hours (actin), 72hours (CFR), or 16 hours (FGFR-4) (Rappolee, et al.,1988; Ausubel, 1993). The Xmat film was scanned usinga Biorad Scanning densitometer and the intensity ofbands was calculated after subtraction of backgroundabove and below the band. The intensities of the bandsfor FGFR-4 or CFR were compared with those of theactin loading control for each cell line and for organs.The size of the bands was computed from an analysis onsemilog paper on the migration of a 0.24–95 kb RNAladder (Life technologies) in a similar way to thatreported for analysis of proteins in polyacrylamide gels(Dunkert and Rueckert, 1969).
In Situ HybridizationAmplimers of FGFR-1–4 receptors were cloned into
PcrII vectors according to manufacturers’ protocols(Invitrogen; Ausubel, 1993). Transformants werescreened using PCR and the identity of the amplimers
was verified as described (Rappolee et al., 1994, 1996;Puscheck et al., 1997). Amplimers for FGFR-4 andFGFR-3 were derived from sequences in the 38 UTR.Digoxygenin cRNA was prepared from plasmid DNA bylinearization and runoff transcription with T7 Polymer-ase and SP6 Polymerase in sense and antisense direc-tions for each plasmid as previously described (Rap-polee et al., 1994, 1996; Puscheck et al., 1997).Blastocysts were pipetted into the oviduct, or embryosin utero were fixed and embedded as previously. Afterdehydration the slides were air-dried and stored at220°C until use. In situ hybridization was done aspreviously described. FGFR-3 probes detected mRNAin E10.5 telencephalon and FGFR-4 probes detectedmRNA in E13.5 myotome as previously reported (Starket al., 1991; Peters et al., 1992).
Indirect Immunocytochemistryand Nuclear Staining
For immunocytochemical analysis, blastocyst out-growths were fixed and permeabilized as previouslydescribed (Puscheck et al., 1998). The blastocysts werestained with immunopurified rabbit polyclonal antise-rum to mouse FGFR-4 antibody (diluted at 1:50) orimmunonpurified rabbit polyclonal anti-FGFR-3 oligo-peptide (1:200–1:400; Santa Cruz Biotech, Santa Cruz,CA), or polyclonal anti-laminin (1:200; Sigma). Rabbitantisera were raised by hyperimmunization to theC terminus 14 mer oligopeptide of FGFR-4 or to theC terminus 15 mer oligopeptide of FGFR-3 and purifiedby affinity chromatography to the immunogen (Chai etal., 1998; Santa Cruz Biotech). The FGFR-4 antiserawere tested for specificity on cos cells transiently trans-fected with FGFR-4. Antisera coincubated with excess
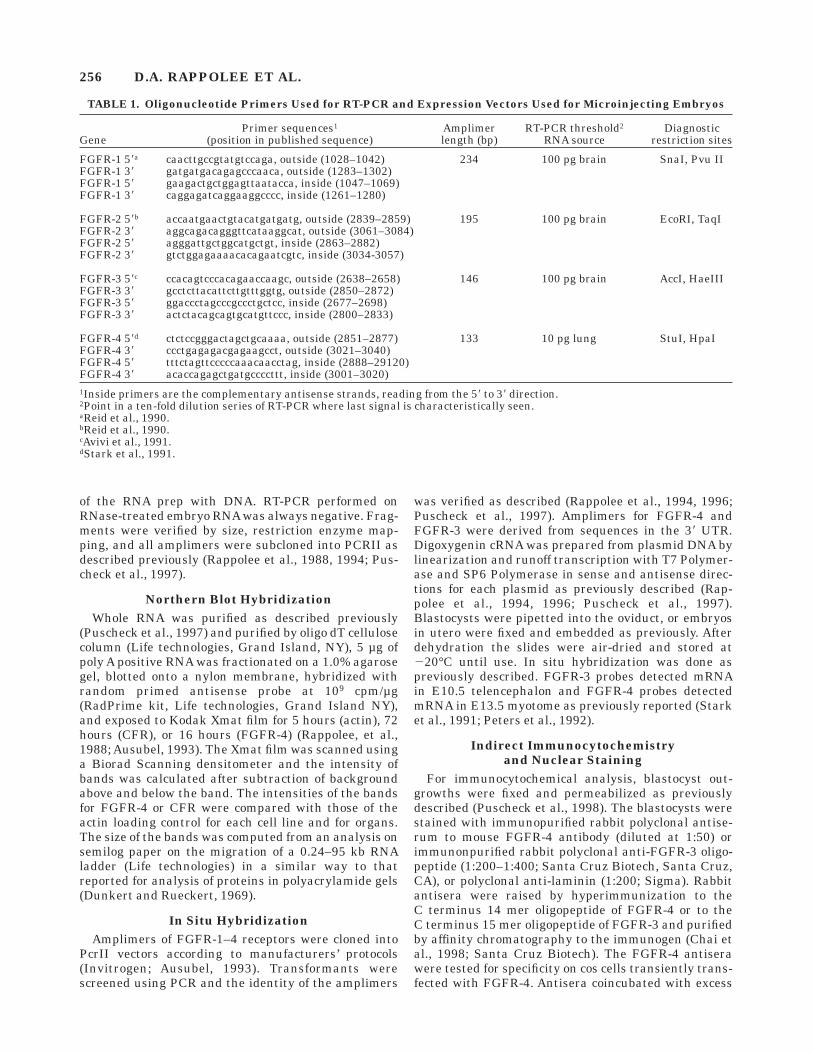
TABLE 1. Oligonucleotide Primers Used for RT-PCR and Expression Vectors Used for Microinjecting Embryos
GenePrimer sequences1
(position in published sequence)Amplimerlength (bp)
RT-PCR threshold2
RNA sourceDiagnostic
restriction sites
FGFR-1 58a caacttgccgtatgtccaga, outside (1028–1042) 234 100 pg brain SnaI, Pvu IIFGFR-1 38 gatgatgacagagcccaaca, outside (1283–1302)FGFR-1 58 gaagactgctggagttaatacca, inside (1047–1069)FGFR-1 38 caggagatcaggaaggcccc, inside (1261–1280)
FGFR-2 58b accaatgaactgtacatgatgatg, outside (2839–2859) 195 100 pg brain EcoRI, TaqIFGFR-2 38 aggcagacagggttcataaggcat, outside (3061–3084)FGFR-2 58 agggattgctggcatgctgt, inside (2863–2882)FGFR-2 38 gtctggagaaaacacagaatcgtc, inside (3034-3057)
FGFR-3 58c ccacagtcccacagaaccaagc, outside (2638–2658) 146 100 pg brain AccI, HaeIIIFGFR-3 38 gcctcttacattcttgtttggtg, outside (2850–2872)FGFR-3 58 ggaccctagcccgccctgctcc, inside (2677–2698)FGFR-3 38 actctacagcagtgcatgttccc, inside (2800–2833)
FGFR-4 58d ctctccgggactagctgcaaaa, outside (2851–2877) 133 10 pg lung StuI, HpaIFGFR-4 38 ccctgagagacgagaagcct, outside (3021–3040)FGFR-4 58 tttctagttcccccaaacaacctag, inside (2888–29120)FGFR-4 38 acaccagagctgatgccccttt, inside (3001–3020)1Inside primers are the complementary antisense strands, reading from the 58 to 38 direction.2Point in a ten-fold dilution series of RT-PCR where last signal is characteristically seen.aReid et al., 1990.bReid et al., 1990.cAvivi et al., 1991.dStark et al., 1991.
256 D.A. RAPPOLEE ET AL.

(0.5 µg) immunizing oligopeptide were used as a nega-tive control. The primary antibody was followed bybiotinylated anti–rabbit IgG and then by alkaline phos-phatase-coupled streptavidin (Rappolee et al., 1994).FGFR-3 and laminin staining was developed withstreptavidin-Texas Red or streptavidin-FITC (1:500;Vector). All specimens were micrographed and format-ted in Photoshop 3.0 as done previously (Chai et al.,1998; Puscheck et al., 1998).
RESULTSExpression of mRNA for FGF
Receptors in Mouse BlastocystsRT-PCR was used to detect the presence of mRNA for
the four high-affinity FGF receptors in mouse blasto-cysts. Total RNA was purified from freshly isolated,washed blastocysts in the presence of carrier rRNA andthe number of blastocysts was normalized to the amountof carrier rRNA recovered. Groups of three blastocystswere assayed for actin transcripts to preclude DNAcontamination and to normalize the amount of cDNA.FGFR-3 and FGFR-4 were detected at relatively highlevels, FGFR-2 at lower levels, and FGFR-1 was de-tected at background levels (Fig. 1). In each 10-folddilution series, the first lane was equivalent to threeblastocysts or approximately 4.5 ng of whole RNA (Pikoand Clegg, 1982). The thresholds for detection in thedilution analysis of blastocysts were 4.5 ng, 450 pg, 45pg, and 0.45 pg for FGFR-1, -2, -3, and -4, respectively.The positive controls for each FGF receptor had thresh-olds of 100 pg for FGFR-1, -2, and -3 and 10 pg forFGFR-4 (Table 1). Each analysis of control and blasto-cyst was repeated three times with similar results. Intissues reported to lack FGFR-1 receptor expression asdetected Northern blot hybridization (Partanen et al.,1992), FGFR-1 receptor transcripts were detected atthresholds of 2–200 ng of RNA (data not shown). Thesedata suggest that FGFR-3 and FGFR-4 are expressed
at high levels, FGFR-2 at lower levels and FGFR-1 atbackground levels in the blastocyst.
We also tested for the expression of FGFR-4 receptorin embryonic stem cell and embryonal carcinoma celllines. This receptor has been reported to be either 3.0 or3.5 kb in the mouse and to increase with differentiationof embryonic stem cells (Stark et al., 1991; Partanen etal., 1992; McDonald and Heath, 1994). The cells differ-entiating in monolayer after LIF removal are parietalendoderm with a nondefined mesoderm-type cell (Mum-mery et al., 1990). We confirmed one previous reportthat the FGFR-4 is a single 3.5 kb transcript (Fig. 2A,C; Stark et al., 1991) in the mouse. In addition, weanalyzed the CFR that was detected in the chick as asingle 8.6 kb transcript coding for a cytoplasmic, nonen-zymatic receptor (Burrus et al., 1992; Zuber et al.,1997). Interestingly, it was also cloned in the mouse asthe cell surface ligand for the E-selectin receptor (Steeg-maier et al., 1995). The mouse homologue contains aunique amino-terminus domain of 70 amino acids andmay be a splice variant of the chick CFR. As in thechick, the mouse CFR is a single mRNA at 8.6 kb (Fig.2C). It has an expression pattern similar to FGFR-4 indifferentiating embryonic stem cells and F9 teratocarci-noma cells, but with a lower expression in a cell linewith characteristics of TGC (Fig. 2A, B; Damjanov andDamjanov, 1985).
Expression of FGFR-3 mRNA in MousePeri-Implantation Embryos
Since FGFR-3 and FGFR-4 mRNA were detected atthe highest levels by RT-PCR, we used in situ hybridiza-tion to localize the cells that express FGFR-3 mRNA.We detected FGFR-3 mRNA in TGC and ICM of theblastocyst at E3.5 and E5.0 by in situ hybridization(Fig. 3). It appeared that FGFR-3 was detected athigher levels in extraembryonic cell lineages than in
Fig. 1. mRNA transcripts for FGFR-3 and FGFR-4 are expressed athigh levels in the E3.5 mouse blastocyst. FGFR-2 transcripts areexpressed at lower levels, and FGFR-1 transcripts are expressed atbackground levels. Each series was an RT-PCR representing 3 CD-1blastocysts in lane (1) at the left, followed by a 10-fold dilution series in
the RT-PCR reactions to the right (2, 3, 4, 5, 6). Each preparation ofblastocysts was first normalized for an equal amount of b-actin mRNAin the RT-PCR. M 5 marker. FGFR-1 5 234 bp, FGFR-2 5 195 bp,FGFR-3 5 146 bp, and FGFR-4 5 133 bp. Arrowheads show the bandof interest for each receptor.
FGF RECEPTORS IN MOUSE EMBRYOS 257
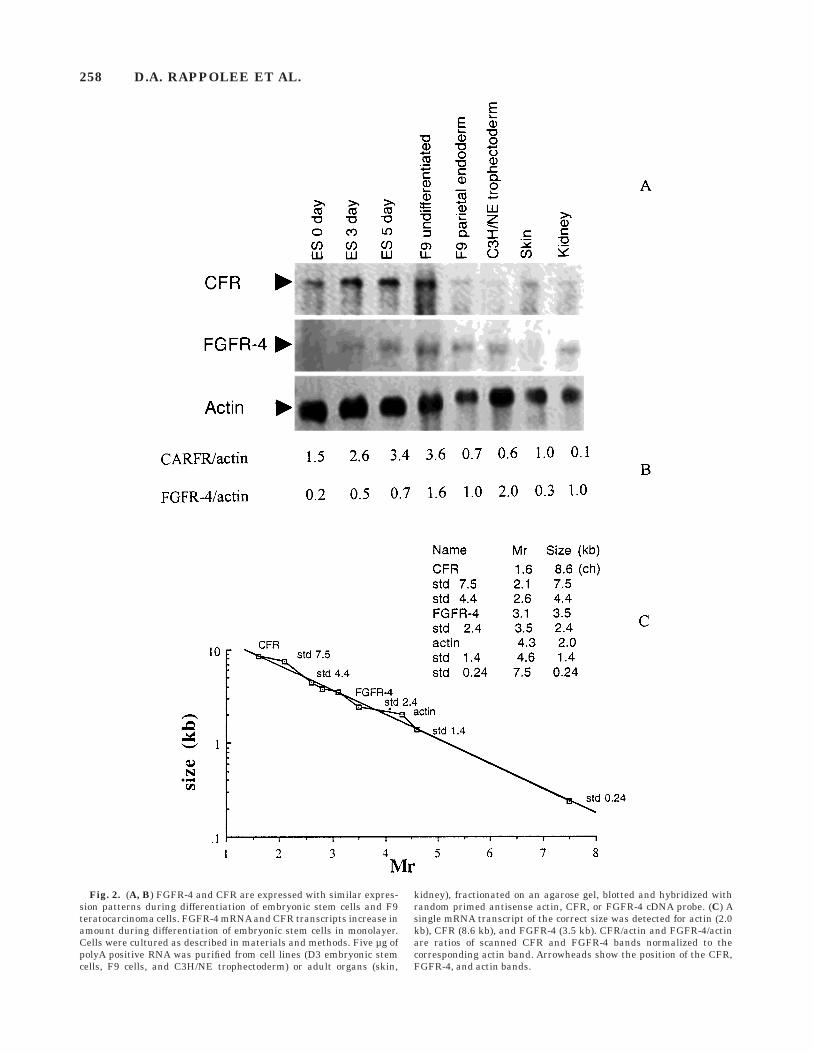
Fig. 2. (A, B) FGFR-4 and CFR are expressed with similar expres-sion patterns during differentiation of embryonic stem cells and F9teratocarcinoma cells. FGFR-4 mRNA and CFR transcripts increase inamount during differentiation of embryonic stem cells in monolayer.Cells were cultured as described in materials and methods. Five µg ofpolyA positive RNA was purified from cell lines (D3 embryonic stemcells, F9 cells, and C3H/NE trophectoderm) or adult organs (skin,
kidney), fractionated on an agarose gel, blotted and hybridized withrandom primed antisense actin, CFR, or FGFR-4 cDNA probe. (C) Asingle mRNA transcript of the correct size was detected for actin (2.0kb), CFR (8.6 kb), and FGFR-4 (3.5 kb). CFR/actin and FGFR-4/actinare ratios of scanned CFR and FGFR-4 bands normalized to thecorresponding actin band. Arrowheads show the position of the CFR,FGFR-4, and actin bands.
258 D.A. RAPPOLEE ET AL.

Fig. 3. FGFR-3 mRNA transcripts are detected in mouse embryos on day E3.5 and E5.0. (A, B) In situhybridization of antisense FGFR-3 to a section of day E3.5 blastocysts. (C, D) In situ hybridization ofantisense FGFR-3 to a section of an implanting day E5.0 blastocyst. (E, F) In situ hybridization of senseFGFR-3 to a section of implanting day E5.0 blastocyst (lower magnification than C and D). Hoechst-stained nuclei of sections in B, D, and F correspond to those in A, C, and E, respectively. Arrowheads showexpression of FGFR-3 mRNA (A–D). These sections are not to the same scale.
FG
FR
EC
EP
TO
RS
INM
OU
SE
EM
BR
YO
S259

the ICM and its derivatives, and it was not detected inmaternal tissue.
Expression of FGFR-4 in MousePeri-Implantation Embryos
By in situ hybridization, FGFR-4 mRNA was de-tected in the mural and polar trophectoderm as well asthe ICM of the blastocyst at E3.5, but not in theoviductal epithelium and submucosa (Fig. 4A, B).FGFR-4 mRNA was highly expressed and developmentof the alkaline phosphatase-mediated stain was alwaysrapid. At E5.0, the implanting embryo FGFR-4 mRNAwas detected in mural and polar trophectoderm and inembryonic ectoderm (Fig. 4C, D). In E5.5 embryos,FGFR-4 mRNA was detected in the primitive ectoderm,
but at not in parietal endoderm (Fig. 4E, F). FGFR-4mRNA was detected in E6.0 embryos at high levels inthe primitive ectoderm, at low levels in extraembryonicectoderm, but not in primitive endoderm (Fig. 4G, I);and FGFR-4 mRNA was not detected in E6.5 embryos(Fig. 4H, J). These data suggest that FGFR-4 mRNA isdetectable ubiquitously, but it is restricted to certaincell types after implantation.
Expression of FGFR-3 and FGFR-4 Proteinin Blastocyst Outgrowths
Blastocyst outgrowths are used as an in vitro modelfor studying implantation. Although the morphology isnot the same as the embryo in utero, many of theprocesses of yolk sac and placental formation occur
Fig. 4. FGFR-4 transcripts are expressed in all cells of preimplanta-tion mouse blastocysts at E3.5 and E5.0. (A, B) In situ hybridization ofantisense FGFR-4 to a section of E3.5 mouse embryos. (C, D) In situhybridization of antisense FGFR-4 to a section of an implanting E5.0blastocyst. (E, F) In situ hybridization of antisense FGFR-4 to asection of an E5.5 embryo. (G, H) In situ hybridization of antisenseFGFR-4 to a section of an E6.0 embryo. (I, J) In situ hybridization of
antisense FGFR-4 to a section of an E6.5 embryo. (K, L) In situhybridization of sense FGFR-4 to a section of an implanting E5.0blastocyst (lower magnification than C and D). Hoechst-stained nucleiof sections in (B, D, F, H, J, and L) correspond to antisense and senseFGFR-4 hybridizations in (A, C, E, G, I, and K), respectively. Arrow-heads show detection of FGFR-3 mRNA and arrows show cells that donot express FGFR-3 mRNA. The sections are not to the same scale.
260 D.A. RAPPOLEE ET AL.

Fig. 5. FGFR-3 protein is detected in ICM, extraembryonic ectoderm andprimitive endoderm in the early blastocyst outgrowth and in extraembryoniccell lineages in later outgrowths. Blastocysts were cultured for three days,fixed and stained for FGFR-3 (A, C) and counterstained with Hoechst. In (A,B) FGFR-3 protein is detected in ICM (arrowhead). In (C, D) FGFR-3 proteinis detected in primitive endoderm at the top of the ICM/endoderm mound(arrowhead). E3.5 blastocysts were cultured for 6 days, fixed and stained forFGFR-3 and Hoechst. In (E–H), the focal plane of the micrograph was at thecoverslip where the arrowhead shows that FGFR-3 protein was detected inTGC (E, F) and parietal endoderm (G, H). In E, the arrow shows the positionof the nucleus. In (I, J) blastocyst outgrowths were micrographed at a focalplane at the midpoint of the ICM/endoderm mound; arrowheads show thatFGFR-3 is detected in primitive endoderm and arrows show that FGFR-3 isnot detected in ICM cells. In (K, L) no FGFR-3 is detected when excessquantities of immunizing FGFR-3 oligopeptide is coincubated with theantisera. The diagram shows early blastocyst outgrowths with ICM, primi-tive endoderm (PRIM E), and TGC (TGC), and late outgrowths with the samecell types and parietal endoderm (PE). The arrows at the right show theplanes of focus for the micrographs.
FGF RECEPTORS IN MOUSE EMBRYOS 261
similarly (diagram in Fig. 5; Gonda and Hsu, 1980).Under these culture conditions, TGC grow out from theblastocyst at 1–3 days of culture. Blastocysts culturedfor 4–6 days produced TGC and a smaller population ofparietal endoderm cells that were positive for laminin(Puscheck et al., 1998). At this stage, ICM and tropho-blasts from these embryos respond to exogenous FGF-4added to the culture (Rappolee et al., 1994; Chai et al.,1998). Blastocyst outgrowths were tested for the expres-sion of FGFR-3 and FGFR-4 protein. At 3 days culture,FGFR-3 was detected by immunocytochemical meansin embryonic and extraembryonic cells of the blastocystoutgrowth (Fig. 5A–D). On a focal plane at the cover-slip, FGFR-3 protein was detected in outgrowing TGCcells (Fig. 5A, B). On a focal plane at the top of the ICMmound, FGFR-3 protein was detected in the primitive
endoderm. In later stage outgrowths, on a focal plane atthe coverslip, FGFR-3 protein was detected in out-growing TGC (Fig. 5E, F) and parietal endoderm(Fig. 5G, H). At a focal plane at midway up theICM/primitive endoderm mound, FGFR-3 protein isdetected in primitive endoderm, but not ICM (Fig. 5I,J). These data suggest that FGFR-3 is detected inprimitive and parietal endoderm and TGC, but not inthe ICM.
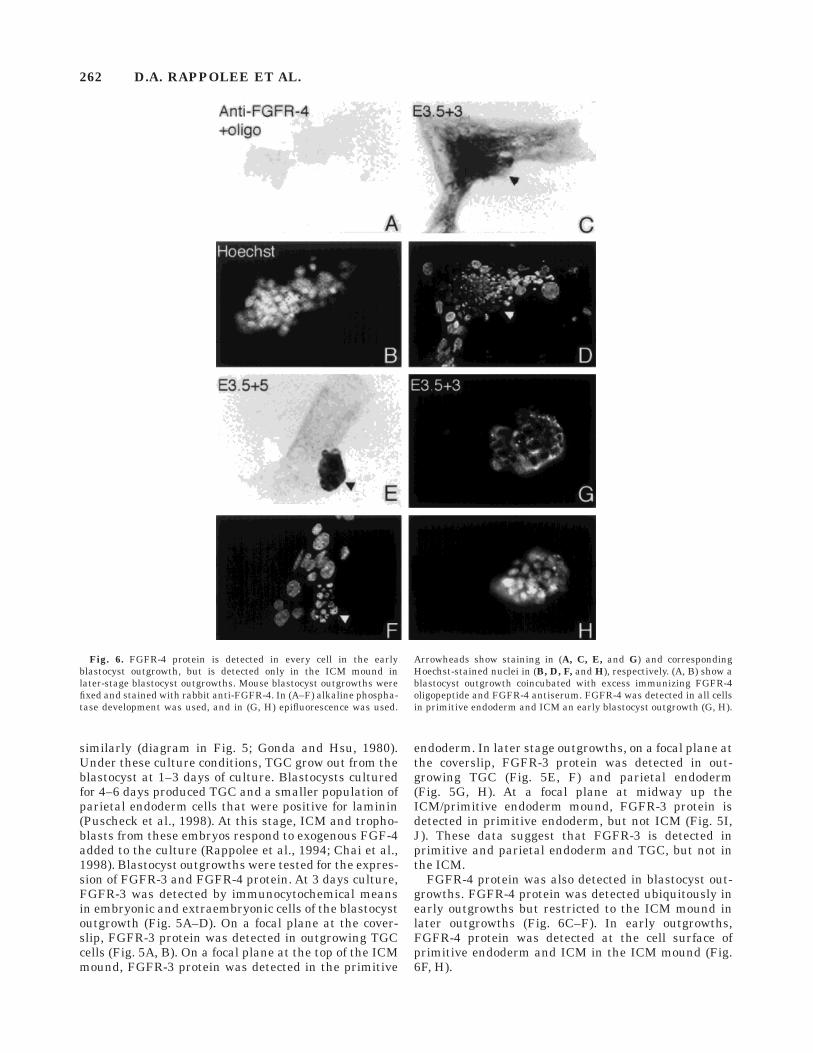
FGFR-4 protein was also detected in blastocyst out-growths. FGFR-4 protein was detected ubiquitously inearly outgrowths but restricted to the ICM mound inlater outgrowths (Fig. 6C–F). In early outgrowths,FGFR-4 protein was detected at the cell surface ofprimitive endoderm and ICM in the ICM mound (Fig.6F, H).
Fig. 6. FGFR-4 protein is detected in every cell in the earlyblastocyst outgrowth, but is detected only in the ICM mound inlater-stage blastocyst outgrowths. Mouse blastocyst outgrowths werefixed and stained with rabbit anti-FGFR-4. In (A–F) alkaline phospha-tase development was used, and in (G, H) epifluorescence was used.
Arrowheads show staining in (A, C, E, and G) and correspondingHoechst-stained nuclei in (B, D, F, and H), respectively. (A, B) show ablastocyst outgrowth coincubated with excess immunizing FGFR-4oligopeptide and FGFR-4 antiserum. FGFR-4 was detected in all cellsin primitive endoderm and ICM an early blastocyst outgrowth (G, H).
262 D.A. RAPPOLEE ET AL.

DISCUSSIONWe report here that FGFR-3 and FGFR-4 mRNA
transcripts are detected at high levels in the implantingblastocyst, and that FGFR-2 is expressed at lowerlevels. Contrary to a previous report (reviewed in Denget al., 1996), FGFR-1 was detected at backgroundlevels. In blastocyst outgrowths, FGFR-3 protein isdetected in extraembryonic cells that give rise to theyolk sac and placenta, but not in the embryo proper.Conversely, FGFR-4 protein is detected throughout theblastocyst, but localizes to the embryonic precursors inblastocyst outgrowths. In implanting embryos, ubiqui-tous FGFR-4 mRNA becomes restricted to the embry-onic ectoderm and is not detected at gastrulation.FGFR-3 mRNA is detected ubiquitously, with higherexpression in the TGC.
What is the evidence for a role for FGFR-3 inimplantation? FGFR-3 is mitogenic when overex-pressed in the preimplantation embryo so it may trans-duce some of the signal of FGF ligands (Chai et al.,1998). The null mutant for fgfr-3 has no lethal peri-implantation phenotype, suggesting that the FGFR-3 isnot essential at this time (Deng et al., 1996). FGFR-3mRNA and protein are detected in the implantingembryo as described above and FGFR-3 is a candidatefor transducing some of the signal of FGF in extraembry-onic membranes such as endoderm and TGC. The IIIcsplice form of FGFR-3 transduces a mitogenic signal inthe embryo (Chai et al., 1998), emphasizing the need forfurther studies to detect this isoform.
What is the evidence for a role for FGFR-4 inimplantation? FGFR-4 is not mitogenic when overex-pressed in the preimplantation embryo (Chai et al.,1998) so it does not transduce the essential mitogenicsignal of FGF ligands in this period. The murine form ofFGFR-4 is not thought to be mitogenic in cell lines(Wang et al., 1994). It is expressed ubiquitously atE3.5–4.5 and may mediate nonmitogenic effects. It ispossible that the fgf-4 null phenotype is different thanthat of the dnFGFR and may be due to nonmitoticeffects (Feldman et al., 1995; Chai et al., 1998). Underthese circumstances, FGFR-4 might transduce some ofthe signal of FGF-4. Since we detect FGF-8 mRNA inthe blastocyst (Rappolee and Changyaleket, unpub-lished data), it is interesting to note that FGFR-4 hasthe most potent affinity for FGF-8 of any FGF receptor(Ornitz et al., 1996). This emphasizes the need for thefurther testing for non-mitogenic roles for FGFR-4 indevelopment.
What is the evidence for a role for FGFR-2 inimplantation? Null mutants of FGFR-2 develop a phe-notype that is similar to that of fgf-4 null (Arman et al.,1998), and the FGFR2 null has a continuing tropho-blast defect (Xu et al., 1998). FGFR-2 mRNA is detectedat low levels in the E3.5 blastocyst and in the ICM ofthe blastocyst outgrowth (data not shown). The proteinfor FGFR-2 is detected ubiquitously (Arman et al.,1998). Taken together, these data suggest a major rolefor FGFR-2 in the peri-implantation embryo.
What is the evidence for a role for FGFR-1 inimplantation? The FGFR-1 null has a later lethalitythan the FGF-4 null, suggesting that FGFR-1 is notresponsible for transducing the signal of FGF-4. FGFR-1mRNA is detected at near-background levels in theE3.5 blastocyst, suggesting a limited role.
Taken together with functional data for the role ofFGFs in early embryos, the data reported here suggestroles for FGFR-3 in peri-implantation development.FGFR-4 mRNA and protein are detected and mighthave a nonmitogenic role. No role for FGFR-1 is indi-cated by these data. Although detected at low levels,FGFR-2 appears to transduce the necessary signal ofFGF-4 at implantation.
ACKNOWLEDGMENTSWe thank Drs. Pergament, Puscheck, and Ghosh for
reading the manuscript and Crystal Beecher for editingthe manuscript. We thank Dr. Lonai for sharing databefore publication. We thank Benjarat Changyaleketand Tess Kim for excellent technical assistance. A grantfrom the Markey Charitable Trust supported researchby D.A.R.
REFERENCESArman E, Haffner-Krausz R, Chen Y, Heath JK, Lonai P (1998):
Targeted disruption of FGFR2 suggests a role for FGF signaling inpre-gastrulation mammalian development. Proc Natl Acad Sci USA95:5062–5065.
Avivi A, Zimmer Y, Yayon Y, Givol D (1991): Flg-2, a new member of thefamily of fibroblast growth factor receptors. Oncogene 6:1089–1092.
Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman GJ, SmithJA, Struhl K (1993): ‘‘Current Protocols in Molecular Biology.’’ NewYork: Wiley.
Burrus LW, Zuber ME, Lueddecke BA, Olwin BB (1992): Identificationof a cysteine-rich receptor for fibroblast growth factors. Mol Cell Biol12:5600–5609.
Chai N, Patel Y, Jacobson K, McMahon J, McMahon A, Rappolee DA(1998): FGF is an essential regulator of the fifth cell division in thepreimplantation mouse embryo. Dev Biol 196:105–115.
Damjanov L, Damjanov A (1985): Trophectodermal carcinoma: Mouseteratocarcinoma-derived tumour stem cells differentiating into tro-phoblastic and yolk sac elements. J Embryol Exp Morp 86:125–141.
Deng C, Wynshaw-Boris A, Zhou F, Kuo A, Leder P (1996): Fibroblastgrowth factor receptor 3 is a negative regulator of bone growth. Cell84:911–921.
Doetschman TC, Eistetter H, Katz M, Schmidt W, Kemler R (1985):The in vitro development of blastocyst-derived embryonic stem celllines: Formation of visceral yolk sac, blood islands, and myocar-dium. J Embryol Exp Morph 87:27–45.
Dunkert AK, Rueckert RR (1969): Observations on molecular weightdeterminations on polyacrylamide gels. J Biol Chem 244:5074–5080.
Feldman B, Poueymirou W, Papaioannou VE, DeChiara TM, GoldfarbM (1995): Requirement of FGF-4 for postimplantation mouse devel-opment. Science 267:246–249.
Gonda MA, Hsu Y-C (1980): Correlative scanning electron, transmis-sion electron, and light microscopic studies of the in vitro develop-ment of mouse embryos on a plastic substrate at the implantationstage. J Embryol Exp Morph 56:23–39.
Hogan B, Beddington R, Constantini F, Lacy B (1994): Manipulatingthe mouse embryo, a laboratory manual. In: ‘‘Cold Spring HarborLaboratory,’’ 2nd edition. New York: Cold Spring Harbor Laborato-ries Press.
Kost TA, Thoedorakis N, Hughes SH (1983): The nucleotide sequenceof the chick cytoplasmic beta-actin gene. Nuc Acid Res 11:8287–8301.
FGF RECEPTORS IN MOUSE EMBRYOS 263

McDonald FJ, Heath JK (1994): Developmentally regulated expres-sion of fibroblast growth factor receptor genes and splice variants bymurine embryonic stem and embryonal carcinoma cells. Dev Gen15:148–154.
McWhirter JR, Goulding M, Weiner JA, Chun J, Murre C (1997): Anovel fibroblast growth factor gene expressed in the developingnervous system is a downstream target of the chimeric homeodo-main oncoprotein E2A-Pbx1. Development 124:3221–3232.
Mummery CL, Van Den Eijnden-Van Raaij AJM, Feijen A, Freund E,Hulskotte E, Schoorlemmer J, Kruijer, W (1990): Expression ofgrowth factors during the differentiation of embryonic stem cells inmonolayer. Dev Biol 142:406–413.
Mummery CL, van Rooyen M, Bracke M, van den Eijnden-van Raaij J,Joop van Zoelen E, Alitalo K (1993): Fibroblast growth factor-mediated growth regulation and receptor expression in embryonalcarcinoma and embryonic stem cells and human germ cell tumours.Biochem Biophys Res Comm 191:188–195.
Ornitz DM, Xu J, Colvin JS, McEwen DG, MacArthur CA, Coulier F,Gao G, Goldfarb M (1996): Receptor specificity of the fibroblastgrowth factor family. J Biol Chem 271:15292–15297.
Orr-Urtreger A, Givol D, Yayon A, Yarden Y, Lonai P (1991): Develop-mental expression of 2 murine FGF receptors flg and bek. Develop-ment 113:1419–1434.
Partanen J, Vainikka S, Korhonen J, Armstrong E, Alitalo K (1992):Diverse receptors for FGFs. Prog Growth Factor Res 4:69–83.
Peters KG, Werner S, Chen G, Williams LT (1992): Two FGF receptorgenes are differently expressed in epithelial and mesenchymaltissues during limb formation and organogenesis in the mouse.Development 114:233–243.
Piko L, Clegg KB (1982): Quantitative changes in total RNA, totalpoly(A) and ribosomes in early mouse embryos. Dev Biol 89:362–78.
Puscheck EE, Yogesh P, Rappolee DA (1997): The expression offibroblast growth factor receptor (FGFR)-4 mRNA, but not FGFR-3in the pregnant ovary. Mol Cell Endocrinol 132:169–176.
Puscheck EE, Dreschler J, Patel Y, Pergament E, Rappolee DA (1998):Expression of insulin receptor substrate-1 in the mouse blastocyst.Mol Reprod Dev 49:386–393.
Rappolee DA, Brenner CA, Schultz R, Mark D, and Werb Z (1988):
Developmental expression of PDGF, TGF-alpha, and TGF-betagenes in preimplantation mouse embryos. Science 241:1823–1825.
Rappolee DA, Basilico CA, Patel Y, Werb Z (1994): Expression andfunction of FGF-4 in peri-implantation development of mouseembryos. Development 120:2259–2269.
Rappolee DA (1998): Growth factors and hormones in mammaliandevelopment. Growth factors in the mammalian pre- and post-implantation embryo. In: R Dickson and D Salomon (eds): ‘‘Hor-mones and Growth Factors in Development and Neoplasia.’’ NewYork: Wiley, pp 93–115.
Rappolee DA, Iyer A, Patel Y (1996): Hepatocyte growth factor and itsreceptor are expressed in cardiac myocytes during early cardiogen-esis. Circ Res 78:1028–1036.
Reid HH, Wilks AF, Bernard O (1990): Two forms of the bFGFreceptor-like mRNA are expressed in the developing mouse brain.Proc Natl Acad Sci 87:1596–1600.
Robertson, EJ (1987): ‘‘Teratocarcinomas and Embryonic Stem Cells: APractical Approach. Oxford: IRL Press.
Stark KL, McMahon JA, McMahon AP (1991): FGFR-4 and newmember of the FGF receptor family, expressed in the definitiveendoderm and skeletal muscle lineages of the mouse. Development113: 641–651.
Steegmaier,M, Levinovitz A, Isenmann S, Borges E, Lenter M, KocherHP, Kleuser B, Vestweber D (1995): The E-selectin-ligand ESL-1 is avariant of a receptor for fibroblast growth factor. Nature 373:615–620.
Wang J-K, Gao G, Goldfarb M (1994): Fibroblast growth factorreceptors have different signaling and mitogenic potentials. MolCell Biol 14:181–184.
Yamaguchi TP, Conlon RA, Rossant J (1992): Expression of FGFR-1/flgduring gastrulation and segmentation in the mouse embryo. DevBiol 152:75–88.
Xu X, Weinstein M, Li C, Naski M, Cohen R, Ornitz D, Leder P, Deng C(1998): Fibroblast growth factor receptor 2 (FGFR2)–mediatedreciprocal regulation loop between FGF8 and FGF10 is essential forlimb induction. Development 125:753–765.
Zuber ME, Zhou Z, Burrus LW, Olwin BB (1997): Cysteine-rich FGFreceptor regulates intracellular FGF-1 and FGF-2 levels. J CellPhys 170:217–227.
264 D.A. RAPPOLEE ET AL.




















