
Somatic embryo proliferation, maturation and germination in Catharanthus roseus
8
Somatic embryo proliferation, maturation and germination in Catharanthus roseus A. Junaid, A. Mujib*, M.A. Bhat & M.P. Sharma Cellular Differentiation and Molecular Genetics Section, Department of Botany, Faculty of Science, Hamdard University, New Delhi, India (*requests for offprints: E-mail: [email protected]) Received 14 March 2005; accepted in revised form 4 October 2005 Key words: carbon sources, Catharanthus roseus, somatic embryogenesis Abstract In the present study an efficient somatic embryogenesis method has been developed in Catharanthus roseus. Friable embryogenic callus was induced from hypocotyl of in vitro germinated seeds on Murashige and Skoog basal nutrient media supplemented with various auxins particularly 2,4-D (1.0 mg l )1 ). However, only NAA (1.0 mg l )1 ) produced somatic embryos in cultures. Embryo proliferation was even high on the same medium added with BAP. Cotyledonary somatic embryo germinated and converted into plantlets in BAP (0.5 mg l )1 ) added medium following a treatment with gibberellic acid (1.0 mg l )1 ) for maturation. Carbon sources and concentrations had a marked influence on maturation process. Plantlet conversion was better achieved when embryos were matured on 3% fructose or 3–6% maltose. The result discussed in this paper indicates that somatic embryos were produced in numbers and converted plantlets can be used as raw material, genetic modification to embryo precursor cell may improve alkaloid yield further. Abbreviations: ANOVA – analysis of variance; BAP – 6-benzylaminopurine; CPA – chlorophenoxyacetic acid; 2,4-D – 2,4-dichlorophenoxyacetic acid; GA 3 – gibberellic acid; IAA – indole acetic acid; MS – Murashige and Skoog; NAA – naphthalene acetic acid; 2,4,5-T – 2,4,5-trihydroxy-butyrophenone Introduction Catharanthus roseus is an important medicinal plant, known worldwide for its anticancerous property. Amongst the numerous compounds that have been reported so far from this herb vincris- tine and vinblastine are perhaps the most impor- tant alkaloids. In vitro studies aiming to improve the low yield have been attempted for a long time in which various plant parts are used. Information on the role of cell/suspension culture, cultural conditions, target site regulation and other in vitro optimization were also previously discussed and reviewed (Moreno et al., 1995; Mujib et al., 2003). However, the use of somatic embryo and the development of transgenic Catharanthus using effective embryogenic system are yet to be explored since there has been no published report of somatic embryogenesis except a preliminary report by Junaid et al. (2004). It is a unique and efficient method for rapid clonal propagation of selected genotypes. The main advantage of somatic embryogenesis over other methods is its unlimited potentiality in producing clones, to be used as medicinal raw material for secondary product (Fowler, 1983). Although there is a risk of somacl- onal variation in long-term indirect embryogenesis (Varga and Badea, 1992), in vitro embryogenesis has been widely used in plant production and transgenic development (Rommens et al., 2004; Walter, 2004). However, in many plant systems, somatic embryo germination and subsequent plant recovery are relatively poor and difficult. The poor somatic embryo quality, lack of maturation Plant Cell, Tissue and Organ Culture (2006) 84: 325–332 Ó Springer 2006 DOI 10.1007/s11240-005-9041-7
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Somatic embryo proliferation, maturation and germination in Catharanthus roseus

Somatic embryo proliferation, maturation and germination in Catharanthusroseus
A. Junaid, A. Mujib*, M.A. Bhat & M.P. SharmaCellular Differentiation and Molecular Genetics Section, Department of Botany, Faculty of Science, HamdardUniversity, New Delhi, India (*requests for offprints: E-mail: [email protected])
Received 14 March 2005; accepted in revised form 4 October 2005
Key words: carbon sources, Catharanthus roseus, somatic embryogenesis
Abstract
In the present study an efficient somatic embryogenesis method has been developed in Catharanthus roseus.Friable embryogenic callus was induced from hypocotyl of in vitro germinated seeds on Murashige andSkoog basal nutrient media supplemented with various auxins particularly 2,4-D (1.0 mg l)1). However,only NAA (1.0 mg l)1) produced somatic embryos in cultures. Embryo proliferation was even high on thesame medium added with BAP. Cotyledonary somatic embryo germinated and converted into plantlets inBAP (0.5 mg l)1) added medium following a treatment with gibberellic acid (1.0 mg l)1) for maturation.Carbon sources and concentrations had a marked influence on maturation process. Plantlet conversion wasbetter achieved when embryos were matured on 3% fructose or 3–6% maltose. The result discussed in thispaper indicates that somatic embryos were produced in numbers and converted plantlets can be used as rawmaterial, genetic modification to embryo precursor cell may improve alkaloid yield further.
Abbreviations: ANOVA – analysis of variance; BAP – 6-benzylaminopurine; CPA – chlorophenoxyaceticacid; 2,4-D – 2,4-dichlorophenoxyacetic acid; GA3 – gibberellic acid; IAA – indole acetic acid; MS –Murashige and Skoog; NAA – naphthalene acetic acid; 2,4,5-T – 2,4,5-trihydroxy-butyrophenone
Introduction
Catharanthus roseus is an important medicinalplant, known worldwide for its anticancerousproperty. Amongst the numerous compounds thathave been reported so far from this herb vincris-tine and vinblastine are perhaps the most impor-tant alkaloids. In vitro studies aiming to improvethe low yield have been attempted for a long timein which various plant parts are used. Informationon the role of cell/suspension culture, culturalconditions, target site regulation and other in vitrooptimization were also previously discussed andreviewed (Moreno et al., 1995; Mujib et al., 2003).However, the use of somatic embryo and thedevelopment of transgenic Catharanthus usingeffective embryogenic system are yet to be explored
since there has been no published report ofsomatic embryogenesis except a preliminary reportby Junaid et al. (2004). It is a unique and efficientmethod for rapid clonal propagation of selectedgenotypes. The main advantage of somaticembryogenesis over other methods is its unlimitedpotentiality in producing clones, to be used asmedicinal raw material for secondary product(Fowler, 1983). Although there is a risk of somacl-onal variation in long-term indirect embryogenesis(Varga and Badea, 1992), in vitro embryogenesishas been widely used in plant production andtransgenic development (Rommens et al., 2004;Walter, 2004). However, in many plant systems,somatic embryo germination and subsequent plantrecovery are relatively poor and difficult. Thepoor somatic embryo quality, lack of maturation
Plant Cell, Tissue and Organ Culture (2006) 84: 325–332 � Springer 2006DOI 10.1007/s11240-005-9041-7

dessication tolerance and dormancy or inhibi-tors(s) within the embryo are observed to be thereasons for low embryo germination (Rtienneet al., 1993; Mujib et al., 1998). In recent yearsseveral innovated methods, such as addition ofosmotic agents, carbohydrates, sugar alcohols etc.,have been used to improve embryo quality beforegermination (Xing et al., 1999; Lipavska andKonradova, 2004; Robichaud et al., 2004). In thispresent paper, we report a reliable and effi-cient method of plant production via somaticembryogenesis from hypocotyl-derived callus inCatharanthus roseus. The role of various carbohy-drates on embryogenesis has also been evaluated.This embryogenic system can be used in futuregenetic modification program in Catharanthusroseus.
Material and methods
In vitro germination of seeds and culture condition
Catharanthus roseus seeds were collected fromin vivo grown plants of Jamia Hamdard herbalgarden. Twelve to 25 surface disinfested seeds wereplaced in Maganta 7 vessels containing 50 ml ofMS medium (Murashige and Skoog, 1962) with nogrowth regulator. Germinated seedlings weregrown until they had attained 2–4 cm length.Various parts (nodal stem, leaf, root, and hypo-cotyl) were used as explants. For embryogeniccallus initiation, MS medium was supplementedwith different plant growth regulators like 2,4-D,2,4,5-T, NAA, IAA and CPA alone or in combi-nations with different concentrations of cytoki-nins. The pH of the medium was adjusted to 5.6before autoclaving at 121 �C for 20 min. Allcultures were incubated under a 16-h photoperiodby cool white (Fluorescent F40 T12/CW/EG)lamp at a photon flux density of 100 lmol m)2 s)1
at 25 �C light/20 �C dark temperature.
Induction of callus and somatic embryos
The initial procedure of establishment of embryo-genic callus and embryo induction were earlierdescribed (Junaid et al., 2004) which was followed.In short, to examine the effect of 2,4-D, CPA,2,4,5-T and other auxins on embryogenic callusformation, hypocotyl of 5–7 days old germinated
seeds were placed on MS medium supplementedwith a range of concentrations (0.25, 0.5, 1.0, 1.5,2.0 mg l)1). The percentage of hypocotyl produc-ing calluses (calli) and somatic embryos weredetermined after 4 weeks onwards. The efficiencyof plant growth regulators and their concentra-tions were analyzed on the basis of visual obser-vation (callusing percentage, callus growth andcallus necrosis etc.), the ineffective treatments werediscontinued.
Proliferation of somatic embryos
For somatic embryo proliferation, 40–50 mg cal-lus was cultured on MS proliferation mediumsupplemented with different concentrations ofBAP and NAA. The percentage of culture showingsomatic embryogenesis (embryogenesis percent-age), somatic embryo number/culture (total andat different developmental stages) have beenrecorded.
Somatic embryo maturation and germination
White opaque cotyledonary somatic embryos(5–6 mm in length) were placed on MS mediumwith varying carbohydrate sources and concentra-tions to evaluate the maturation process. GA3
(1.0 mg l)1) was always added to these maturationmedia since it was earlier noted to be necessaryduring maturation (unpublished). Somatic embryobecame green and elongated that were monitoredafter 5th and 7th weeks. Finally, matured greenembryos were cultured on MS supplemented withoptimized BAP (0.5 mg l)1) for germination(Junaid et al., 2004). The same carbon sourceswere again supplemented to see the influence ongermination. Somatic embryo proliferation interms of root, shoot development, plant conver-sion etc. were observed.
Statistical analysis
Each treatment had 6 somatic embryos with 5replicates, for confirmation, each of the exp-eriment was repeated twice. Somatic embryomaturation and subsequent germination weredetermined using ANOVA followed by least sig-nificant difference (LSD) test at p = 0.05.
326

Result
Embryogenic callus induction and maintenance
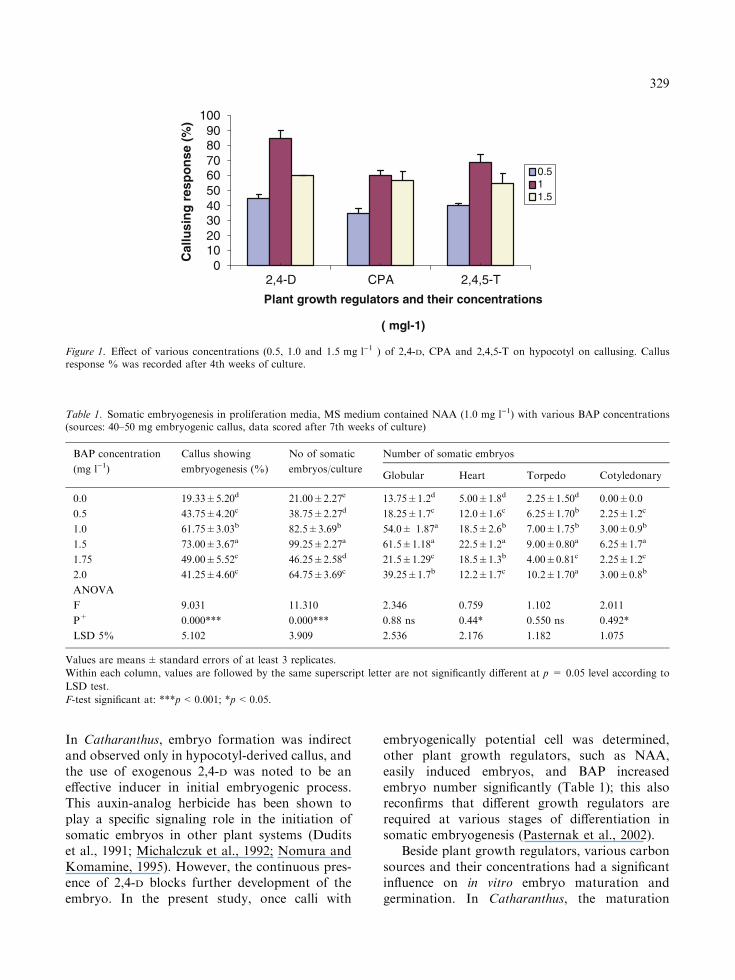
The plant part (hypocotyl) responded well inculture on which three tested auxins had profoundinfluence to produce callus (Figure 1). The maxi-mum callusing response percentage (%) was 85 in2,4-D at 1.0 mg l)1. The effective concentration,however, varied and generally lied within 0.5–1.5 mg l)1 range; higher concentration inhibitedcallus induction and growth. The hypocotyl calluswas friable, light yellow and fast growing, laterproved to be embryogenic (Plate 1a) while other
sources induced calli of non-embryogenic naturecharacterized by their compact and nodularappearance, grown relatively slow. The embryo-genic calli of hypocotyl were routinely maintainedon media supplemented with various auxins aloneor in combinations with cytokinins.
Induction and proliferation of somatic embryos
Although embryogenic callus was induced onseveral auxins treated media, in 1.0 mg l)1 NAA,embryogenic calli rapidly differentiated intoembryos (Plate 1b). The same concentration of2,4-D, CPA and 2,4,5-T were observed to be less
Plate 1. Somatic embryogenesis and plant regeneration from hypocotyl-derived callus in Catharanthus roseus. (a) Friable embryo-genic callus from hypocotyl (�10). (b) Initiation of somatic embryos on 1.0 mg l)1 NAA (�10). (c) Embryos in MS maturationmedium containing 1.0 mg l)1 GA3 (�10). (d ) Advanced stages of somatic embryos (�10). (e) Plant conversion from somaticembryos (�1). ( f ) Normal somatic embryos from sucrose (3%) containing medium (First two from left) and maltose 6% inducedblack-spot on shoot–root junction (Next three) of plantlets (�1). ( g) Complete plant conversion in BAP (0.5 mg l)1) containingmedium (�1).
327

effective. In higher concentrations (1.5 mg l)1 orabove) of 2,4-D and CPA, induced embryogeniccallus showed necrosis after 5th weeks onwards, insome cases, re-callusing started at lower concen-trations (0.5 mg l)1 or below). Induced embryoswere observed in masses and could not be sepa-rated easily. Addition of BAP in NAA (1.0 mg l)1)supplemented media was noted to be more pro-ductive in embryo proliferation process. Embryo-genesis percentage and somatic embryo number/culture were very high on 1.0 and 1.5 mg l)1 ofBAP. Table 1 summarizes and compares the
efficiency of media combinations in differentiatingembryos and subsequent growth.
Maturation and germination of somatic embryos
Morphologically advanced individual somatic em-bryos were cultured on MS medium supplementedwith 1.0 mg l)1 GA3 for maturation (Plate 1c andd). Various carbon sources and concentrationswere also added and performance was evaluated.In all the treatments, somatic embryos grew in size(Table 2) with significant difference (p>0.001),and it maintained a steady growth up to 7th weeks.
The 3% level of carbohydrate was more activethan 6% in which embryo growth was relativelyslow and was applied to all carbon sources i.e.,maltose, glucose, fructose and sucrose. In thesematuration media, somatic embryos became darkgreen, coiled occasionally without any distinctgrowth in embryo morphology. However, in mediasupplemented with BAP (0.5 mg l)1), somaticembryo germinated well (Plate 1e and g). Around70% conversion of the somatic embryos could berecorded in media containing 3% sucrose, maltoseand fructose (Figure 2) except glucose. Only rootformation was observed on media containing 6%of these sugars (Table 3). Glucose did not supportconversion and root formation, only the shootgrowth was observed. The further growth ofconverted embryos was much stronger on mediawith maltose than on media with sucrose orfructose. Better shoot and root development wasnoted on 3–6% maltose followed by 3% fructose.The carbohydrate type had little effect on leafnumber. The plantlets developed on maltosecontaining media had black spots in their shoot–root junction (Plate 1f). The generated somaticembryos and the process of embryogeny have beenused in plantlet multiplication and other researchpurposes.
Discussion
Two different somatic embryogenesis processeshave been observed in plant systems:– Direct embryogenesis (i.e., directly on the
explant), and– Indirect embryogenesis via an intervening
callus phase.
Plate 1. (Continued)
328
In Catharanthus, embryo formation was indirectand observed only in hypocotyl-derived callus, andthe use of exogenous 2,4-D was noted to be aneffective inducer in initial embryogenic process.This auxin-analog herbicide has been shown toplay a specific signaling role in the initiation ofsomatic embryos in other plant systems (Duditset al., 1991; Michalczuk et al., 1992; Nomura andKomamine, 1995). However, the continuous pres-ence of 2,4-D blocks further development of theembryo. In the present study, once calli with
embryogenically potential cell was determined,other plant growth regulators, such as NAA,easily induced embryos, and BAP increasedembryo number significantly (Table 1); this alsoreconfirms that different growth regulators arerequired at various stages of differentiation insomatic embryogenesis (Pasternak et al., 2002).
Beside plant growth regulators, various carbonsources and their concentrations had a significantinfluence on in vitro embryo maturation andgermination. In Catharanthus, the maturation
0102030405060708090
100
2,4-D CPA 2,4,5-T
Plant growth regulators and their concentrations
( mgl-1)
Cal
lusi
ng
res
po
nse
(%
)0.511.5
Figure 1. Effect of various concentrations (0.5, 1.0 and 1.5 mg l)1 ) of 2,4-D, CPA and 2,4,5-T on hypocotyl on callusing. Callusresponse % was recorded after 4th weeks of culture.
Table 1. Somatic embryogenesis in proliferation media, MS medium contained NAA (1.0 mg l)1) with various BAP concentrations(sources: 40–50 mg embryogenic callus, data scored after 7th weeks of culture)
BAP concentration
(mg l)1)
Callus showing
embryogenesis (%)
No of somatic
embryos/culture
Number of somatic embryos
Globular Heart Torpedo Cotyledonary
0.0 19.33±5.20d 21.00±2.27e 13.75±1.2d 5.00±1.8d 2.25±1.50d 0.00±0.0
0.5 43.75±4.20c 38.75±2.27d 18.25±1.7c 12.0±1.6c 6.25±1.70b 2.25±1.2c
1.0 61.75±3.03b 82.5±3.69b 54.0± 1.87a 18.5±2.6b 7.00±1.75b 3.00±0.9b
1.5 73.00±3.67a 99.25±2.27a 61.5±1.18a 22.5±1.2a 9.00±0.80a 6.25±1.7a
1.75 49.00±5.52c 46.25±2.58d 21.5±1.29c 18.5±1.3b 4.00±0.81c 2.25±1.2c
2.0 41.25±4.60c 64.75±3.69c 39.25±1.7b 12.2±1.7c 10.2±1.70a 3.00±0.8b
ANOVA
F 9.031 11.310 2.346 0.759 1.102 2.011
P+ 0.000*** 0.000*** 0.88 ns 0.44* 0.550 ns 0.492*
LSD 5% 5.102 3.909 2.536 2.176 1.182 1.075
Values are means±standard errors of at least 3 replicates.
Within each column, values are followed by the same superscript letter are not significantly different at p = 0.05 level according to
LSD test.
F-test significant at: ***p<0.001; *p<0.05.
329

and germination of the somatic embryos wereinhibited by 3–6% sucrose or glucose; however,6% maltose and 3% fructose individually werevery effective. Similar results i.e., rapid somaticembryo maturation with 3% sucrose or maltose inMS medium was earlier reported by Corredoriaet al. (2003) in Castenia sativa. Although themechanism of action is not clearly known, therole of maltose in improving somatic embryomaturation has been noted in several woodyspecies (Druart, 1990; Alemanno et al., 1997; Liet al., 1998). In maltose-enriched medium, earlyembryo maturation and low hydrolysis was
recently reported in Abies (Norgaard, 1997). InHevea brasiliensis, maltose induced a nutri-tional stress by causing low absorption frommedium, which eventually favored embryo germi-nation (Blanc et al., 1999). Other than maltose, inCatharanthus, 3% fructose also showed a positiveeffect on maturation and germination of thesomatic embryos. Though no sound evidence isavailable in Catharanthus roseus, a similar mech-anism might have played a role during maturationand subsequent germination of somatic embryos.Thus it seems that, in Catharanthus roseus, plantgrowth regulators and carbon sources (maltose
Table 2. Somatic embryo in maturation media (MS + 1.0 mg l)1 GA3), added with different carbohydrates
Treatment Initial length of embryos (mm) Length after 5 weeks (mm) Length after 7 weeks (mm)
Sucrose 3% 5.70±0.5b 8.50±0.3b 10.05±0.2b
Sucrose 6% 5.52±0.2bc 7.275±0.5c 9.05±0.3cd
Maltose 3% 6.57±0.3a 9.675±0.2a 11.47±0.3a
Maltose 6% 5.80±0.6b 8.300±0.2b 9.54±0.3c
Glucose 3% 6.37±0.2a 8.750±0.2b 10.725±0.5b
Glucose 6% 6.00±0.4ab 7.533±0.2c 8.625±0.4d
Fructose 3% 5.725±0.5b 8.675±0.3b 10.625±0.4b
Fructose 6% 5.175±0.3c 7.42±0.8c 9.70±0.2c
ANOVA
F 8.416 12.262 13.12
P+ 0.000*** 0.000*** 0.002 ns
LSD 5% 0.542 0.657 0.559
Values are means±standard errors of 5 replicates with 6 embryos in each replicate.
Within each column, values are followed by the same superscript letters are not significantly different at p = 0.05 level according to
LSD test.
F-test significant at: ***p<0.001; p<0.05.
Figure 2. Effect of various carbon sources on C. roseus somatic embryos showing shoot, root development and plantlet conversion.
330

and fructose) have influenced embryogenesis,which will facilitate mass production of plantletsthat can be used as medicinal raw materials.
References
Alemanno L, Berthouly M & Michaux-Ferriere N (1997) A
comparison between Theobroma cacao L. zygotic embryo-
genesis and somatic embryogenesis from floral explants.
In vitro Cell. Dev. Biol. Plant 33: 163–172
Blanc G, Michaux-Ferriere N, Teisson C, Lardet L & Carron
MP (1999) Effect of carbohydrate addition on the induction
of somatic embryogenesis in Hevea brasiliensis. Plant Cell
Tiss. Org. Cult. 59: 103–112
Corredoira E, Ballester A & Vieitez AM (2003) Proliferation,
maturation and germination of Castanea sativa Mill. somatic
embryos originated from leaf explants. Ann. Bot. 92: 129–136
Druart P (1990) Improvement of somatic embryogenesis of
cherry dwarf root stocks INMIL/GM, by use of different
carbon sources. Acta Horticult. 280: 125–129
Dudits D, Bogre L & Gyorgyey J (1991) Molecular and cellular
approaches to the analysis of plant embryo development
from somatic cells in vitro. J. Cell Sci. 99: 475–484
Fowler MW (1983) Commercial application and economic as-
pects of mass plant cell culture. In: Mantell SH & Smith H
(eds) Plant Biotechnology. Cambridge University Press,
Cambridge (pp 3–38)
Junaid A, Bhat MA, Mujib A & Sharma MP (2004) Somatic
embryogenesis study in Catharanthus roseus (L.) G.Don: an
anticancerous plant. In: Khan IA & Khanum A (eds) Role of
Biotechnology in Medicinal and Aromatic Plants. Vol. XI.
Ukaaz Publication, Hyderabad (pp 98–106)
Li XY , Feng H, Huang H, Murphy JB & Gbur EE Jr (1998)
Polyethylene glycol and maltose enhance somatic embryo
maturation in loblolly pine (Pinus teada L.). In vitro Cell.
Dev. Biol. Plant 34: 22–26
Lipavska H & Konradova H (2004) Somatic embryogenesis in
conifers: the role of carbohydrate metabolism. In Vitro Cell.
Dev. Biol. Plant 40: 23–30
Michalczuk L, Cook TJ & Cohen JD (1992) Auxins levels at
different stages of carrot somatic embryogenesis. Phyto-
chemistry 31: 1097–1103
Moreno PRH, Van der Heijden R, Verpoorte R & Vander
Heijden R (1995) Cell and tissue cultures of Catharanthus
roseus, literatures survey II, updated from 1988–1993. Plant
Cell Tiss. Org. Cult. 42: 1–25
Mujib A, Bandyopadhyay S, Jana BK & Ghosh PD (1998)
Direct and in vitro plant regeneration in Hippeastrum
hybridum. Plant Tiss. Cult. 8(1): 19–25
Mujib A, Ilah A, Gandotra N & Abdin MZ (2003) In vitro
application to improve alkaloids yield in Catharanthus roseus.
In: Govil JN, Kumar PA & Singh VK (eds) Biotechnology
and Genetic Engineering. Vol IV. Sci. Tech Publishing, Texas
(pp 415–439)
Murashige T & Skoog F (1962) A revised medium of rapid
growth and bioassay with tobacco tissue culture. Physiol.
Plant 15: 473–497
Table 3. Somatic embryo germination (plantlet conversion) in BAP (0.5 mg l)1), MS medium also contained different sugars andconcentrations
Treatment Plant conversion (RL
+ SL)
Only root RL (mm) Only shoot SL (mm) Leaf number (LN)
RL (mm) SL (mm)
Sucrose 3% 5.80±0.7d 8.57±0.4d – 11.80±0.4b 3.5±1.2a
Sucrose 6% – – 11.92±0.5a 9.275±0.4c 2.0±1.8b
Maltose 3 % 8.17±0.2b 11.6±0.4b – 11.00±0.1bb 2.5±1.2b
Maltose 6% 9.36±0.4a 12.80±0.1a 9.35±0.5b 12.77±0.7a 3.0±2.1a
Glucose 3% – – – 9.625±0.6c 1.5±1.2c
Glucose 6% 5.30±0.4d 8.55±0.5d – 11.00±0.4b 2.5±2.3b
Fructose 3% 7.26±0.3c 9.55±0.5c – 11.75±0.1b 2.5±1.2b
Fructose 6% – – 7.30±2.6c 7.925±0.3d 2.2±2.6b
ANOVA
F 0.752 0.267 2.646 0.379 0.567
P+ 0.043* 0.0674 ns 0.048* 0.820 ns 0.345*
LSD 5% 0.536 0.554 0.436 0.700 2.455
Data were recorded after 10 weeks on germinating medium following 7 weeks in maturation media.
Values are means±standard errors of 5 replicates with 6 embryos per replicate per treatment.
Values are followed by the same superscript letter are not significantly different at p = 0.05 level according to LSD test.
– There was no converted plantlets and root development.
F-test significant at: *p<0.001; p<0.05.
RL, Root length; SL, Shoot length.
331

Norgaard JV (1997) Somatic embryo maturation and plant
regeneration in Abies nordmanniana LK. Plant Sci. 124: 211–
221
Nomura K & Komamine A (1995) Physiological and bio-
chemical aspects of somatic embryogenesis. In: Thorpe TA
(ed) In Vitro Embryogenesis in Plants. Kluwer Academic
Publishers, Dordrecht (pp 249–266)
Pasternak TP, Prinsen E, Ayaydin F, Miskolezi P, Potters G,
Asard H, Van Onckelen A, Dudits D & Feher A (2002) The
role of auxin, pH, and stress in the activation of embryogenic
cell division in leaf protoplast derived cells of alfalfa. Plant
Physiol. 129: 1807–1819
Robichaud RL, Lessard VC & Merkle SA (2004) Treatments
affecting maturation and germination of American chestnut
somatic embryos. J. Plant Physiol. 161: 957–969
Rommens CM, Humara JM, Ye J, Yan H, Richael C, Zhang L,
Perry R & Swords K (2004) Crop improvement through
modification of the plant’s own genome. Plant Physiol. 135:
421–431
Rtienne H, Montoro P, Michaux-Ferriere N & Carron MP
(1993) Effect of desication medium osmolarity and abscisic
acid on the maturation ofHivea brasiliensis somatic embryos.
J. Exp. Bot. 44: 1613–1619
Varga P & Badea EM (1992) In vitro plant regeneration
methods in alfalfa breeding. Euphytica 59: 119–123
Walter C (2004) Genetic engineering in conifer forestry: tech-
nical and social consideration. In Vitro Cell. Dev. Biol. Plant
40: 434–441
Xing Z, Powell WA & Maynard CA (1999) Development and
germination of American chestnut somatic embryos. Plant
Cell Tiss. Org. Cult. 57: 47–55
332




















