
Morphological, Physiological, and Biochemical Changes in Rhodopsin Knockout Mice

Chinese Science Bulletin
© 2009 SCIENCE IN CHINA PRESS
Springer
AR
TIC
LES
M
ED
ICA
L C
ELL
ULA
R B
IOLO
GY
Citation: Sheng B Y, Niu Y, Zhou H, et al. The mitochondrial function was impaired in APP knockout mouse embryo fibroblast cells. Chinese Sci Bull, 2009, 54(10): 1725-1731, doi: 10.1007/s11434-009-0239-7
The mitochondrial function was impaired in APP knockout mouse embryo fibroblast cells
SHENG BaiYang1, NIU Ying1, ZHOU Hui2, YAN JiaXin2, ZHAO NanMing3, ZHANG XiuFang1
& GONG YanDao1† 1 State Key Laboratory of Biomembrane and Membrane Biotechnology, Department of Biological Sciences and Biotechnology, Tsinghua University, Beijing 100084, China;
2 Wuhan Institute of Biological Products, Wuhan 430060, China; 3 Medical School, Tsinghua University, Beijing 100084, China
The amyloid precursor protein (APP) is recognized as the source of Aβ, which plays an important role in Alzheimer’s disease. However, the biological function of APP is obscure. Previous studies showed that mitochondria could be a target of APP. In this work, APP knockout mouse embryo fibroblast (MEF) cells were used to test if APP plays any role in maintaining the mitochondrial function. As the result, APP knockout MEF cells (APP−/− cells) showed the abnormal mitochondrial function, including slower cell proliferation, lower mitochondrial membrane potential, lower intracellular ROS, higher mitochon-drial membrane fluidity and lower cytochrome c oxidase activity than their wild-type counterparts. However, no change was found in the amount of mitochondria in MEF APP−/− cells.
amyloid precursor protein (APP), Alzheimer’s disease, mitochondria, mitochondrial membrane potential, Cytochrome c oxidase activity
As the source of β amyloid peptide (Aβ), the amyloid precursor protein (APP) is believed to play a pivotal part in etiology of Alzheimer’s disease. A great deal of work has been done about this well-known protein, but the overwhelming majority of these studies were based on its proteolytic processing and the notorious role in Alz-heimer’s disease. As a result, the biological function of APP itself remains elusive. It is well known that APP is a member of an evolutionarily conserved family of type I membrane proteins that includes APP-like proteins 1 and 2 (APLP1 and APLP2)[1,2]. APP’s evolutionary con-servation and existence of APP-like isoforms (APLP1 and APLP2) which lack the Aβ sequence, however, suggest that APP may have important physiological functions that are unrelated to Aβ production[3].
APP-deficient (as well as APLP1 and 2-deficient) mice show subtle neurological deficits, including vari-ous abnormalities in the formation of forebrain com-missures, locomotor activity, grip strength, reduction in synaptophysin immunoreactivity and overall dendritic
length in hippocampal CA1 neurons[4,5]. The non-amy- loidogenic sAPP has been implicated in enhancement of synaptogenesis, neurite outgrowth, and neuronal sur-vival[3]. In particular, sAPP has been demonstrated to be able to stimulate the proliferation of neural progenitor cells in culture[6,7]. Meanwhile, it was shown that the N-terminus of APP plays a crucial role in proliferation, adhesion and migration of Keratinocytes in vitro[8,9]. In addition, Han et al.[10] reported that the level of tau phosphorylation in APP deficient cells and brains is sig-nificantly higher than that in wild-type controls, result-ing from activation of cyclin-dependent kinase 5 (CDK5) but not glycogen synthase kinase 3, the two major tau kinases. Together, these findings suggest that APP has
Received January 14, 2009; accepted March 2, 2009 doi: 10.1007/s11434-009-0239-7 †Corresponding author (email: [email protected]) Supported by Tsinghua-Yue-Yuen Medical Sciences Fund (Grant No. 20240000514), Science and Technology Planning Project of Beijing Municipal (Grant No. H060920050430) and State Key Laboratory of Biomembrane and Membrane Bio-technology

1726 www.scichina.com | csb.scichina.com | www.springerlink.com
important physiological functions. Newly synthesized intracellular APP is delivered to
the surface membrane where it is subjected to prote-olytic processing by α-secretase. Uncleaved APP mole-cules can be internalized into endocytic compartments and subsequently cleaved by β-secretase (BACE) and γ-secretase to generate Aβ. A fraction of Aβ peptides is generated in the Golgi apparatus and the endoplasmic reticulum[11]. However, in a recent study, it was found that APP carries both a mitochondrial-targeting domain and an endoplasmic reticulum-targeting domain. The dual leader sequence permits APP targeting to the endo-plasmic reticulum (ER) or to mitochondria[12]. In cul-tured cells overexpressing APP, APP was detected in mitochondria-enriched cell fractions by immunoblotting and APP immunoreactivity in mitochondria by immu-noelectron microscopy. In accord with this result, when the leader sequence was mutated, the mitochondrial as-sociation was not seen. The latter work showed that the nonglycosylated full-length and C-terminal truncated amyloid precursor protein (APP) accumulated exclu-sively in the protein import channels of mitochondria in the brain samples from Alzheimer’s disease patients but not in the age-matched control samples[13]. However, with the leader sequence targeting to mitochondria, whether APP has normal functions within mitochondria remains unknown[14]. In this study, we compared the function of mitochondria of APP knockout (APP−/−) mouse embryonic fibroblasts with that of the wild-type APP (APP+/+) mouse embryonic fibroblasts. It was shown that deletion of APP led to a significantly reduced proliferation in vitro, decreased mitochondrial mem-brane potential and electron transport chain complex IV activity, and impaired mitochondrial membrane fluidity. Interestingly, this deletion also resulted in a significantly reduced level of reactive oxygen species (ROS). There-fore, APP may play an important role in maintaining the normal function of mitochondria.
1 Materials and methods
1.1 Chemicals
1,6-Diphenyl-1,3,5hexatrie (DPH), Rhodamine 123,3- (4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and 2,7-dichlorofluorescin-diacetole (DCFH-DA) were obtained from Sigma, USA. DMEM and trypsin were from Gibco, USA. Penicillin and streptomycin were from Amresco, USA. Fetal bovine
serum was purchased from Hyclone, USA. Mouse an-ti-Aβ1-42 monoclonal antibody 4d10 was prepared by ourselves.
1.2 Cell proliferation
For the analysis of cell proliferation in vitro, APP wild-type (APP+/+) and APP−/− mouse embryonic fibro-blasts were seeded at a density of 5 × 104 cells/mL in a 6-well tissue culture plate, which contained 3 mL DMEM supplemented with 10% FBS. After incubation at 37℃ in a 5% CO2 incubator for 3 or 5 d, the cells were digested by 0.1% trypsinase and then counted by blood counting instrument. The mitochondrial function was evaluated by the 3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide (MTT) reduction assay. Briefly, 1 mL of serum-free medium and 200 μL of MTT solution (5 mg/mL in PBS) were added into each well. After 4 h incubation at 37℃, the MTT solution was re-moved and the insoluble formazane crystal was dissolved in dimethyl sulfoxide (DMSO). About 200 μL of the solu-tion from each well was aspirated and poured into a 96-well plate for absorbance measurement at 570 nm.
1.3 Mitochondrial membrane potential
To determine the mitochondrial membrane potential (ψm), fluorescence dye Rhodamine 123 was added to the cell culture medium to a final concentration of 1 μmol/L for 15 min. Then the cells were washed 5 times with Krebs’ ringer solution (100 mmol/L NaCl, 2.6 mmol/L KCl, 25 mmol/L NaHCO3, 1.2 mmol/L MgSO4, 1.2 mmol/L KH2PO4 and 11 mmol/L glucose) and examined under a confocal fluorescence microscope equipped with an argon laser.
1.4 Intracellular reactive oxygen species (ROS)
The intracellular ROS was measured in APP+/+ and APP−/− MEF cells. The cells were rinsed with Krebs’ ringer solution, and 10 μmol/L DCFH-DA was loaded. After incubation at 37 in a 5% CO℃ 2 incubator for 1 h, the cells were washed 5 times with the same buffer and examined under a confocal fluorescence microscope equipped with an argon laser.
1.5 Preparation of isolated mitochondria
For isolating mitochondrial fractions, adherent cells were collected and washed by buffer 1 (NaCl, 134 mmol/L; KCl, 5 mmol/L; Na2HPO4, 0.7 mmol/L; Tris, 10 mmol/L; pH 7.5). The cells were resuspended in 1.5 mL buffer 2 (NaCl, 10 mmol/L; CaCl2, 1.5 mmol/L; Tris,

Sheng B Y et al. Chinese Science Bulletin | May 2009 | vol. 54 | no. 10 1727
AR
TIC
LES
M
ED
ICA
L C
ELL
ULA
R B
IOLO
GY
10 mmol/L; pH 7.5) and homogenized in a glass ho-mogenizer for 10 min. Then 0.6 mL buffer 3 (sucrose, 2000 mmol/L; EDTA35, mmol/L; Tris, 50 mmol/L; pH 7.5) was added into the glass homogenizer. The resulting homogenate was centrifuged at 1500 × g for 10 min at 4℃. Then, the supernatant fraction was centrifuged at 15000 × g for 5 min at 4℃ to pellet mitochondria. The resulting pellet was washed twice and resuspended in 1 mL of buffer 4 (Mannitol, 210 mmol/L; sucrose, 70 mmol/L; Tris, 5 mmol/L; EDTA, 5 mmol/L; pH 7.5). The whole process was carried out on the ice.
1.6 Mitochondrial Membrane Fluidity
Fluorescence polarizability of the lipophilic probe 1,6-diphenyl-1,3,5-hexatriene (DPH) was used to meas-ure mitochondrial membrane fluidity. 0.1 mg/mL mito-chondria was incubated with DPH (2 μmol/L final) dis-solved in tetrahydrofuran in PBS at 4℃ overnight in the dark. Fluorescence polarizability (P) was measured at 30℃ using a Shimadzu Spectrophotofluorimeter model RF-5000 at excitation and emission wavelengths of 360 and 430 nm, respectively. It was calculated from emis-sion intensities through a polarizer whose polarization axis was oriented parallel (IVV) or perpendicular (IVH) to the vertical polarizer of excitation light. We calculated a correction factor G = IHV/IHH. IHH and IHV are intensities measured with the horizontal polarizer of the excitation beam and the horizontal or vertical polarizer of the emission beam, respectively. The polarization value was thus obtained from the relation: P = (IVV−G·IVH)/(IVV + G·IVH), and the mitochondrial membrane viscosity (η) was calculated by the formula: η = 2P/0.46−P. The mi-tochondrial membrane fluidity was denoted by 1/η.
1.7 Complex IV Activity
Intact isolated mitochondria were used to determine Cytochrome c oxidase activity. The colorimetric assay is based on the observation that a decrease in absorbance at 550 nm of ferrocytochrome c is caused by its oxidation to ferricytochrome c by Cytochrome c oxidase[15].
1.8 Mitochondria amount assay
The amount of mitochondria in MEF APP+/+ and MEF APP−/− cells was measured using the cell-permanent mi-tochondrion-selective dye Mitotracker Red. The probe can accumulate in active mitochondria and stain mito-chondria independently from the mitochondrial mem-brane potential. Mitotracker Red was solubilized in
DMSO. MEF APP+/+ and MEF APP−/− cells were seeded the day before at a density of 1×105 cells/mL. Cells were incubated for 30 min at 37℃ with 50 ng/mL Mitotracker Red or 50 ng/mL Rhodamine 123. Then the cells were washed twice with phosphate-buffered saline and ana-lyzed using the laser-scanning confocal microscope. The integral of fluorescence intensity over the cell area repre-sentes the total amount of mitochondria in MEF cells[16].
1.9 The mitochondrial localization of APP assay
MEF APP+/+ cells were seeded the day before. The cells were incubated for 30 min at 37℃ with cell-permanent mitochondrion-selective dye Mitotracker Red. Then the cells were permeabilized with 0.1% Triton X-100 and labelled with monoclonal antibody 4d10. The cells were then labelled with fluorescence-tagged secondary anti-bodies and stained with DAPI for 10 min. Then the cells were washed up with PBS several times and subjected to confocal microscopic observation.
2 Results
2.1 APP−/− MEF cells showed impaired cell prolif-eration
APP+/+ and APP−/− MEF cells were characterized by immunofluorescence microscopy using the monoclonal antibody 4d10, which was prepared by ourselves. In APP−/− cells, little endogenous APP was detectable (Fig-ure 1(b)), whereas in the wild-type MEF cells it was
Figure 1 Expression of APP detected by immunofluorescence. APP could be found in APP+/+ cells (a), but not in APP−/− cells (b). The red fluorescence denotes APP, and the blue fluorescence de-notes the cell nuclear. Bar: 20 μm.

1728 www.scichina.com | csb.scichina.com | www.springerlink.com
Figure 2 Comparison of cell proliferation of APP+/+ MEF cells and APP−/− cells. (a) The cell number was recorded after 3- and 5-day-culture. Compared with APP−/− MEF: *, P < 0.01; **, P < 0.0001. (b) After 3- and 5-day-culture, cell proliferation was evaluated by MTT reduction assay. Data represent the Mean ± SD for 10 samples. Compared with APP−/− MEF cells: *, P < 0.01; **, P < 0.0001. found that the perinuclear region displayed the typical staining (Figure 1(a)).
When 5×104 APP+/+ and APP−/− MEF cells were seeded on the 6-well tissue culture plate, after 3 or 5 d of culture, the amount of APP+/+ MEF cells was signifi-cantly larger than that of APP−/− MEF cells (Figure 2(a)). Three days later, the density of APP+/+ MEF cells grew to 24.7×104, while that of the APP−/− MEF cells only multiplied to 14×104. Moreover, 5 d later, the density of APP+/+ MEF cells grew to 106.5×104, while that of the APP−/− MEF cells only multiplied to 21.5×104. In accord with these results, a significant decrease of optical den-sity for the APP−/− MEF cells was detected by the MTT reduction assay either after 3- or 5-day culture (Figure 2(b)). These observations indicated that the proliferation of APP−/− MEF cells is abnormal compared to the wild-type controls.
2.2 Mitochondrial membrane potential was reduced in APP−/− MEF cells
To investigate the influence of APP knockout on the function of mitochondria, we first measured the mito-chondrial membrane potential of MEF cells with or without APP (APP+/+ and APP−/−). However, APP−/− MEF cells showed a lower membrane potential than APP−/− MEF cells (Figure 3(c)). These data suggest that APP is very important for maintaining the mitochondrial function.
2.3 ROS was reduced in APP−/− MEF cells
When we used fluorescent probe H2DCF-DA to detect cytosolic ROS, unexpectedly, we found that the level of ROS in APP+/+ MEF cells was significantly higher than
Figure 3 Comparison of mitochondrial membrane potential (MMP) in APP−/− and APP+/+ MEF cells. APP−/− MEF cells (b) showed a significantly lower membrane potential than APP+/+ MEF cells (a). (c) Quantification of mitochondrial membrane potential. *, P < 0.01.
Figure 4 Comparison of intracellular reactive oxygen species (ROS) in APP−/− and APP+/+ MEF cells. The ROS level in APP+/+ cells (a) was significantly higher than that of the APP−/− ones (b). (c) Quantification of reactive oxygen species. *, P < 0.01. that in APP−/− ones. This result was intriguing. It has been reported that APP KO neurons are associated with reduced survival, increased apoptosis, and impaired neurite outgrowth[10]. ROS is considered to be capable of

Sheng B Y et al. Chinese Science Bulletin | May 2009 | vol. 54 | no. 10 1729
AR
TIC
LES
M
ED
ICA
L C
ELL
ULA
R B
IOLO
GY
mediating inhibition of protein phosphatases, contribut-ing to both cellular proliferation and apoptosis suppres-sion[17]. Therefore, reduced ROS may be involved in the abnormal proliferation of APP−/− MEF cells.
2.4 APP−/− MEF cells showed higher mitochondrial membrane fluidity and lower Cytochrome c oxidase activity
The changes of mitochondrial membrane fluidity were determined by measuring fluorescence polarizability changes of the lipophilic probe 1,6-diphenyl-1,3,5- hexatriene (DPH). Mitochondrial membrane fluidity was significantly higher in APP−/− MEF cells compared to that in the control mitochondria (Figure 5(a)). Moreover, APP−/− MEF cells showed a significantly lower Cyto-chrome c oxidase activity compared with APP+/+ MEF cells (Figure 5(b)), which implicated that the mitochon-drial respiratory chain was impaired in APP−/− MEF cells.
Figure 5 (a) Comparison of mitochondrial membrane fluidity in APP−/− MEF cells and the controls; (b) comparison of cytochrome c oxidase activity in APP−/− and APP+/+ MEF cells. *, P < 0.01.
2.5 APP−/− MEF cells showed no change in the amount of mitochondria
To exclude the possibility that Cytochrome c oxidase
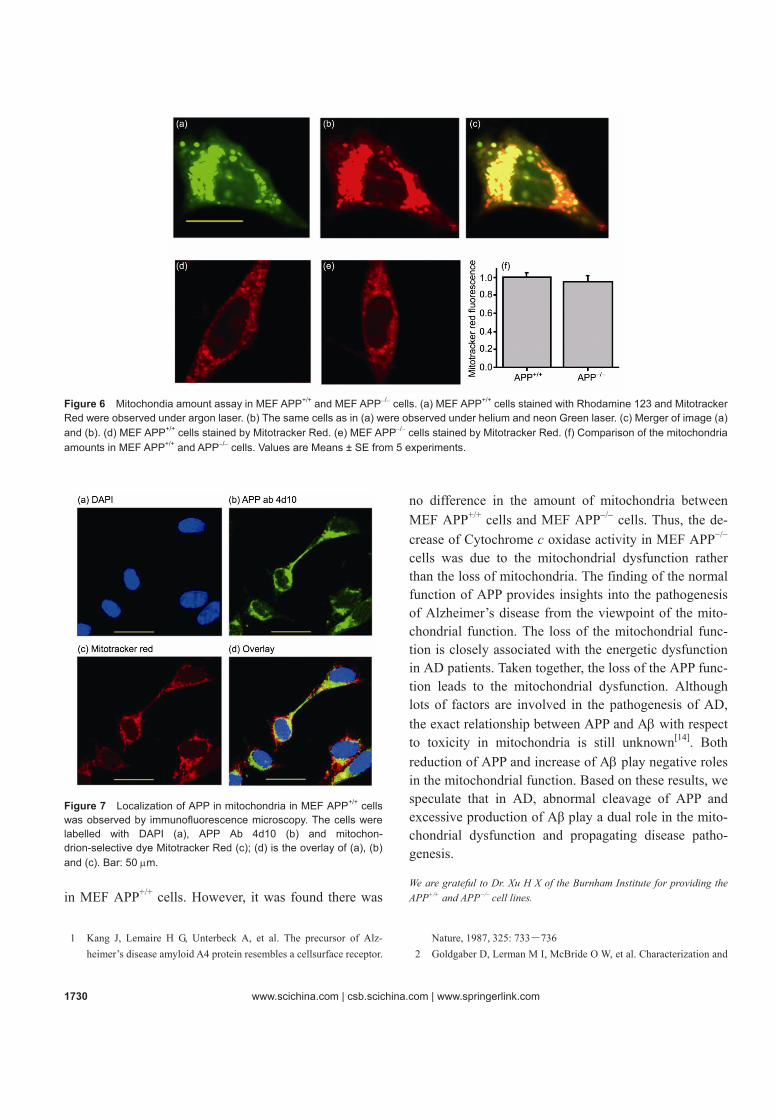
activity was reduced in MEF APP−/− cells due to a loss of mitochondria, we determined the amount of mito-chondria in MEF APP+/+ and MEF APP−/− cells using the mitochondrial dye Mitotracker Red, which selectively stains mitochondria independently from the mitochon-drial membrane potential. Firstly, we observed that Mi-totracker Red was co-localized well with the mitochon-drial membrane potential dye Rhodamine 123 (Figure 6(a)-(c)), which indicated that Mitotracker Red could selectively stain mitochondria as well as Rhodamine 123. As shown in Figure 6 (d)-(f), there was no change in the amount of mitochondria in MEF APP+/+ cells and MEF APP−/− cells.
2.6 The mitochondrial localization of APP in MEF APP+/+ cells
Localization of APP in mitochondria was further inves-tigated by immunobiochemistry. The mitochondria in MEF APP+/+ cells were stained with Mitotracker red. Then the Triton-permeabilized cells were subjected to immunostaining with monoclonal antibody 4d10. Both overlapping and nonoverlapping APP-stained structures with mitochondrial-specific staining were observed (Figure 7(a)-(d)). The result indicated that part of APP was located in mitochondria in MEF APP+/+ cells.
3 Discussion
APP, a type I transmembrane protein that undergoes en-zymatic processing to produce various fragments, is linked to Alzheimer’s disease. The physiological func-tions of APP are still obscure. APP KO mice revealed mild abnormalities such as impaired locomotor activity[5] and defects in synaptic density in the cortex. Previous studies have shown that APP carries a dual leader se-quence, permitting targeting to the endoplasmic reticu-lum (ER) or to mitochondria. When overexpressed in cultured cells, APP was found in mitochondria by im-munoelectron microscopy and Western blot analysis[12]. Thus, we speculate that APP is involved in maintaining the physiological function of mitochondria. The ob-tained results here support our hypothesis.
In this work, APP knockout MEF cells showed the reduced activity of proliferation, lower mitochondrial membrane potential, lower level of intracellular ROS, lower COX IV activity, and higher mitochondrial mem-brane fluidity than APP+/+ MEF cells. Accordingly, it was found that part of APP was located in mitochondria
1730 www.scichina.com | csb.scichina.com | www.springerlink.com
Figure 6 Mitochondia amount assay in MEF APP+/+ and MEF APP−/− cells. (a) MEF APP+/+ cells stained with Rhodamine 123 and Mitotracker Red were observed under argon laser. (b) The same cells as in (a) were observed under helium and neon Green laser. (c) Merger of image (a) and (b). (d) MEF APP+/+ cells stained by Mitotracker Red. (e) MEF APP−/− cells stained by Mitotracker Red. (f) Comparison of the mitochondria amounts in MEF APP+/+ and APP−/− cells. Values are Means ± SE from 5 experiments.
Figure 7 Localization of APP in mitochondria in MEF APP+/+ cells was observed by immunofluorescence microscopy. The cells were labelled with DAPI (a), APP Ab 4d10 (b) and mitochon-drion-selective dye Mitotracker Red (c); (d) is the overlay of (a), (b) and (c). Bar: 50 μm. in MEF APP+/+ cells. However, it was found there was
no difference in the amount of mitochondria between MEF APP+/+ cells and MEF APP−/− cells. Thus, the de-crease of Cytochrome c oxidase activity in MEF APP−/− cells was due to the mitochondrial dysfunction rather than the loss of mitochondria. The finding of the normal function of APP provides insights into the pathogenesis of Alzheimer’s disease from the viewpoint of the mito-chondrial function. The loss of the mitochondrial func-tion is closely associated with the energetic dysfunction in AD patients. Taken together, the loss of the APP func-tion leads to the mitochondrial dysfunction. Although lots of factors are involved in the pathogenesis of AD, the exact relationship between APP and Aβ with respect to toxicity in mitochondria is still unknown[14]. Both reduction of APP and increase of Aβ play negative roles in the mitochondrial function. Based on these results, we speculate that in AD, abnormal cleavage of APP and excessive production of Aβ play a dual role in the mito-chondrial dysfunction and propagating disease patho-genesis.
We are grateful to Dr. Xu H X of the Burnham Institute for providing the APP+/+ and APP−/− cell lines.
1 Kang J, Lemaire H G, Unterbeck A, et al. The precursor of Alz-
heimer’s disease amyloid A4 protein resembles a cellsurface receptor. Nature, 1987, 325: 733―736
2 Goldgaber D, Lerman M I, McBride O W, et al. Characterization and

Sheng B Y et al. Chinese Science Bulletin | May 2009 | vol. 54 | no. 10 1731
AR
TIC
LES
M
ED
ICA
L C
ELL
ULA
R B
IOLO
GY
chromosomal localization of a cDNA encoding brain amyloid of Alzheimer’s disease. Science, 1987, 235: 877―880
3 Chen Y, Tang B L. The amyloid precursor protein and postnatal neu-rogenesis/neuroregeneration. Biochem Biophys Res Commun, 2006, 341(1): 1―5
4 Magara F, Müller U, Li Z W, et al. Genetic background changes the pattern of forebrain commissure defects in transgenic mice underex-pressing the beta amyloid-precursor protein. Proc Natl Acad Sci USA, 1999, 96: 4656―4661
5 Zheng H, Jiang M, Trumbauer M E, et al, Beta-amyloid precursor protein-deficient mice show reactive gliosis and decreased locomotor activity. Cell, 1995, 81: 525―531
6 Hayashi Y, Kashiwagi K, Ohta J, et al. Alzheimer amyloid protein precursor enhances proliferation of neural stem cells from fetal rat brain. Biochem Biophys Res Commun, 1994, 205(1): 936―943
7 Ohsawa I, Takamura C, Morimoto T, et al. Amino-terminal region of secreted form of amyloid precursor protein stimulates proliferation of neural stem cells. Eur J Neurosci, 1999, 11(6): 1907―1913
8 Kirfel G, Borm B, Rigort A, et al. The secretory, beta-amyloid pre-cursor protein is a motogen for human epidermal keratinocytes. Eur J Cell Biol, 2002, 81: 664―676
9 Siemes C, Quast T, Kummer C, et al. Keratinocytes from APP/ APLP2-deficient mice are impaired in proliferation, adhesion and migration in vitro. Exp Cell Res, 2006, 312(11): 1939―1949
10 Han P, Dou F, Li F, Zhang X, et al. Suppression of cyclin-dependent kinase 5 activation by amyloid precursor protein: A novel excitopro-tective mechanism involving modulation of tau phosphorylation. J Neurosci, 2005, 25(50): 11542―11552
11 Kamenetz F, Tomita T, Hsieh H, et al. APP processing and synaptic function. Neuron, 2003, 37: 925―937
12 Anandatheerthavarada H K, Biswas G, Robin M A, et al. Mitochon-drial targeting and a novel transmembrane arrest of Alzheimer’s amyloid precursor protein impairs mitochondrial function in neuronal cells. J Cell Biol, 2003, 161: 41―54
13 Devi L, Prabhu B M, Galati D F, et al. Accumulation of amyloid precursor protein in the mitochondrial import channels of human Alzheimer’s disease brain is associated with mitochondrial dysfunc-tion. J Neurosci, 2006, 26(35): 9057―9068
14 Lin M T, Beal M F. Alzheimer’s APP mangles mitochondria. Nat Med, 2006, 12(11): 1241―1243
15 Vijayasarathy C, Damle S, Lenka N, et al. Tissue variant effects of heme in-hibitors on the mouse Cytochrome c oxidase gene expression and catalytic activity of the enzyme complex. Eur J Biochem, 1999, 266(1): 191―200
16 Keil U, Bonert A, Marques C A, et al. Amyloid β-induced changes in nitric oxide production and mitochondrial activity lead to apoptosis. J Biol Chem. 2004, 279(48): 50310―50320
17 Fruehauf J P, Meyskens F L Jr. Reactive oxygen species: A breath of life or death? Clin Cancer Res, 2007, 13(3): 789―794
Copyright © 2022 FDOKUMEN




















