
Antioxidant mechanisms in apolipoprotein E deficient mice prior to and following closed head injury
Upload
independentCategory
view
1download
0
European Journal of Pharmacology 670 (2011) 519–526
Contents lists available at SciVerse ScienceDirect
European Journal of Pharmacology
j ourna l homepage: www.e lsev ie r .com/ locate /e jphar
Cardiovascular Pharmacology
Antiatherogenic effects of oleanolic acid in apolipoprotein E knockout mice
Niels Henrik Buus a,b,⁎, Nicolaj C. Hansson a, Rosalia Rodriguez-Rodriguez a,c, Edgaras Stankevicius a,Malene Rohr Andersen d, Ulf Simonsen a
a Department of Pharmacology, Institute of Biomedicine, Aarhus University, Aarhus, Denmarkb Department of Renal Medicine, Aarhus University Hospital, Aarhus, Denmarkc Department of Pharmacology, University of Sevilla, Spaind Department of Clinical Biochemistry, Copenhagen University Hospital, Gentofte, Denmark
⁎ Corresponding author at: Department of Renal MedicBrendstrupgaardsvej, 8200 Aarhus N, Denmark. Tel.: +45
E-mail address: [email protected] (N.H. Buus).
0014-2999/$ – see front matter © 2011 Elsevier B.V. Alldoi:10.1016/j.ejphar.2011.09.037
a b s t r a c t
a r t i c l e i n f oArticle history:Received 14 June 2011Received in revised form 8 September 2011Accepted 11 September 2011Available online 21 September 2011
Keywords:AtherosclerosisApolipoprotein E knockout mouseOleanolic acidNitric oxide
Oleanolic acid (OA) is a plant triterpenoid steroid with potentially antiatherogenic properties. We investigatedwhether OA affected atherosclerosis development and vascular function in apolipoprotein E knockout (ApoE−/−)mice. ApoE−/− mice were fed a high cholesterol Western-type diet in combination with OA (100 mg/kg/day),fluvastatin (5 mg/kg/day) or vehicle, with wild type (WT) mice serving as controls. After 8 weeks of treatmentatherosclerotic plaque areas in the aortic arch andplasma lipid concentrationswere determined. Vasoconstrictionand relaxation of the proximal part of aorta were investigated in vitro. Inducible nitric oxide synthase (iNOS)wasvisualized using immunoblotting. As opposed to WT and fluvastatin- and vehicle-treated mice, OA-fed ApoE−/−
mice gained no weight during the treatment period. Plasma concentrations of total-cholesterol and triglyceridewere not significantly reduced by OA- or fluvastatin treatment. Plaque area of vehicle-treated mice was 25%,but only 14% in OA- and 19% in fluvastatin-treated mice. As compared to WT, vasoconstriction to phenylephrinewas attenuated in ApoE−/−mice. TheNOS inhibitor asymmetric dimethylarginine (ADMA) enhancedphenyleph-rine constriction, but significantly more so in vehicle- and fluvastatin-treated than in OA-treated and WT mice.Relaxation to acetylcholine was only slightly attenuated in ApoE−/− mice and not affected by OA or fluvastatintreatment. ADMA abolished acetylcholine relaxation almost completely. In ApoE−/− mice iNOS expression wasreduced by OA treatment. In conclusion OA exerts potent antiatherogenic effects independent of plasma lipidlevels andwithoutmajor changes in eNOS-mediated acetylcholine relaxation. However, OA reduced iNOS expres-sion possibly altering vascular reactivity to phenylephrine.
ine, Aarhus University Hospital,8949 5566.
rights reserved.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
Cardiovascular disease is a major contributor to morbidity andmortality and despite antihypertensive, antithrombotic and cholesterollowering treatments, atherosclerosis remains prevalent and further in-terventions to delay disease progression are warranted. Both epidemio-logical and interventional studies have documented the consumptionof a traditional Mediterranean diet, with olive oil as the principal sourceof fat, to be associated with a reduction of cardiovascular risk factors andincidence of coronary heart disease (de Lorgeril et al., 1994; Trichopoulouet al., 2003). These effects havemainly been ascribed to the high contentof monounsaturated fatty acids combined with a low content of saturat-ed fatty acids in olive oil. Evidence suggests though, that the beneficialeffects of olive oil cannot be fully explained by the favorable fatty acidcomposition (Perona et al., 2006). Hence, research has been performedto elucidate the cardiovascular effects of some minor constituents inolive oil, particularly phenols and tocopherols (El and Karakaya, 2009).
However, during the recent years also the triterpenoids, such as oleanolicacid (OA, Fig. 1), have received increased attention.
Triterpenoids are ubiquitous in the plant kingdom and possess awide range of biological effects. Hepatoprotective and antitumoral activ-ities of OA have long been recognized (Liu, 1995), but also evidence ofpotentially antiatherogenic effects is now emerging. Chronic adminis-tration of OA to Dahl salt-sensitive rats, demonstrated antihyperlipi-demic, antioxidant and antihypertensive effects (Somova et al., 2003a).Furthermore, in vitro studies on isolated rat aortic segments showed en-dothelium-dependent, nitric oxide (NO)-mediated vasorelaxing proper-ties of OA, presumably due to enhanced phosphorylation of endothelialNO synthase (eNOS) (Rodriguez-Rodriguez et al., 2008). Cholesterol-lowering statins are traditionally considered to exert their effectsthrough inhibition of cholesterol synthesis, but they may also enhancethe bioavailability of NO through increased eNOS expression or antioxi-dant effects (Mason et al., 2004). Thus, the pleiotropic actions of statinscorrespond to some of the expectedmechanisms, bywhichOAmay alsoexert its antiatherogenic effects.
The homozygous apolipoprotein E-deficient (ApoE−/−) mouse is awidely used animalmodel of atherosclerosis, due to the spontaneous de-velopment of severe hypercholesterolemia and atherosclerotic lesions,
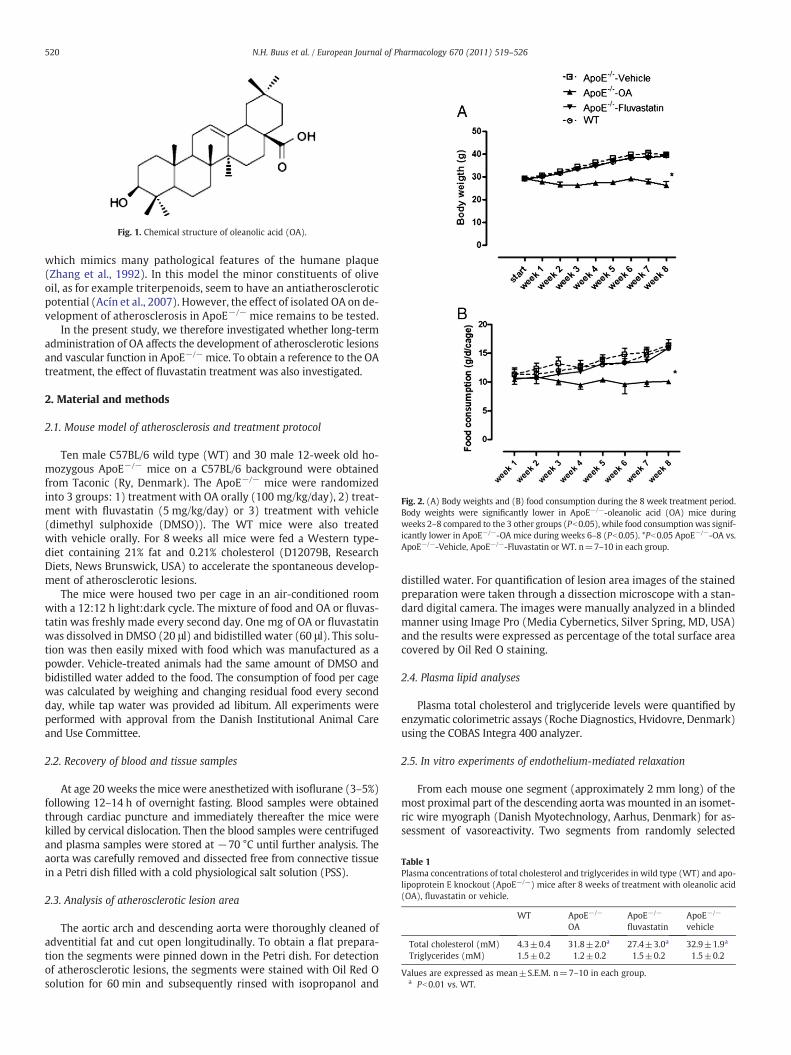
Fig. 1. Chemical structure of oleanolic acid (OA).
Fig. 2. (A) Body weights and (B) food consumption during the 8 week treatment period.Body weights were significantly lower in ApoE−/−-oleanolic acid (OA) mice duringweeks 2–8 compared to the 3 other groups (Pb0.05), while food consumption was signif-icantly lower in ApoE−/−-OA mice during weeks 6–8 (Pb0.05). *Pb0.05 ApoE−/−-OA vs.ApoE−/−-Vehicle, ApoE−/−-Fluvastatin or WT. n=7–10 in each group.
Table 1Plasma concentrations of total cholesterol and triglycerides in wild type (WT) and apo-lipoprotein E knockout (ApoE−/−) mice after 8 weeks of treatment with oleanolic acid(OA), fluvastatin or vehicle.
WT ApoE−/−
OAApoE−/−
fluvastatinApoE−/−
vehicle
Total cholesterol (mM) 4.3±0.4 31.8±2.0a 27.4±3.0a 32.9±1.9a
Triglycerides (mM) 1.5±0.2 1.2±0.2 1.5±0.2 1.5±0.2
Values are expressed as mean±S.E.M. n=7–10 in each group.a Pb0.01 vs. WT.
520 N.H. Buus et al. / European Journal of Pharmacology 670 (2011) 519–526
which mimics many pathological features of the humane plaque(Zhang et al., 1992). In this model the minor constituents of oliveoil, as for example triterpenoids, seem to have an antiatheroscleroticpotential (Acín et al., 2007). However, the effect of isolated OA on de-velopment of atherosclerosis in ApoE−/− mice remains to be tested.
In the present study, we therefore investigated whether long-termadministration of OA affects the development of atherosclerotic lesionsand vascular function in ApoE−/− mice. To obtain a reference to the OAtreatment, the effect of fluvastatin treatment was also investigated.
2. Material and methods
2.1. Mouse model of atherosclerosis and treatment protocol
Ten male C57BL/6 wild type (WT) and 30 male 12-week old ho-mozygous ApoE−/− mice on a C57BL/6 background were obtainedfrom Taconic (Ry, Denmark). The ApoE−/− mice were randomizedinto 3 groups: 1) treatment with OA orally (100 mg/kg/day), 2) treat-ment with fluvastatin (5 mg/kg/day) or 3) treatment with vehicle(dimethyl sulphoxide (DMSO)). The WT mice were also treatedwith vehicle orally. For 8 weeks all mice were fed a Western type-diet containing 21% fat and 0.21% cholesterol (D12079B, ResearchDiets, News Brunswick, USA) to accelerate the spontaneous develop-ment of atherosclerotic lesions.
The mice were housed two per cage in an air-conditioned roomwith a 12:12 h light:dark cycle. The mixture of food and OA or fluvas-tatin was freshly made every second day. One mg of OA or fluvastatinwas dissolved in DMSO (20 μl) and bidistilled water (60 μl). This solu-tion was then easily mixed with food which was manufactured as apowder. Vehicle-treated animals had the same amount of DMSO andbidistilled water added to the food. The consumption of food per cagewas calculated by weighing and changing residual food every secondday, while tap water was provided ad libitum. All experiments wereperformed with approval from the Danish Institutional Animal Careand Use Committee.
2.2. Recovery of blood and tissue samples
At age 20 weeks themice were anesthetized with isoflurane (3–5%)following 12–14 h of overnight fasting. Blood samples were obtainedthrough cardiac puncture and immediately thereafter the mice werekilled by cervical dislocation. Then the blood samples were centrifugedand plasma samples were stored at −70 °C until further analysis. Theaorta was carefully removed and dissected free from connective tissuein a Petri dish filled with a cold physiological salt solution (PSS).
2.3. Analysis of atherosclerotic lesion area
The aortic arch and descending aorta were thoroughly cleaned ofadventitial fat and cut open longitudinally. To obtain a flat prepara-tion the segments were pinned down in the Petri dish. For detectionof atherosclerotic lesions, the segments were stained with Oil Red Osolution for 60 min and subsequently rinsed with isopropanol and
distilled water. For quantification of lesion area images of the stainedpreparation were taken through a dissection microscope with a stan-dard digital camera. The images were manually analyzed in a blindedmanner using Image Pro (Media Cybernetics, Silver Spring, MD, USA)and the results were expressed as percentage of the total surface areacovered by Oil Red O staining.
2.4. Plasma lipid analyses
Plasma total cholesterol and triglyceride levels were quantified byenzymatic colorimetric assays (Roche Diagnostics, Hvidovre, Denmark)using the COBAS Integra 400 analyzer.
2.5. In vitro experiments of endothelium-mediated relaxation
From each mouse one segment (approximately 2 mm long) of themost proximal part of the descending aorta wasmounted in an isomet-ric wire myograph (Danish Myotechnology, Aarhus, Denmark) for as-sessment of vasoreactivity. Two segments from randomly selected

Fig. 3. (A) Representative examples of atherosclerotic lesion formation in the ascending thoracic aorta, as evaluated by en face Oil Red O staining from WT mice, ApoE−/−-Vehicle,ApoE−/−-oleanolic acid (OA), and ApoE−/−-Fluvastatin-treated mice. Mean values of atherosclerotic lesion formation in the aortic arch (B) and descending thoracic aorta (C).*Pb0.05 for ApoE−/−-Fluvastatin vs. ApoE−/−-Vehicle, **Pb0.01 for ApoE−/−-OA vs. ApoE−/−-Vehicle, ***Pb0.001 for ApoE−/−-Vehicle vs. ApoE−/−-OA. n=7–10 in each group.
521N.H. Buus et al. / European Journal of Pharmacology 670 (2011) 519–526
micewere studied in parallel. In brief the vessel segmentsweremountedon two 100 μm thin steel wires for recordings of tension as previouslydescribed for other types of arteries (Simonsen et al., 1992). After anequilibration period of 30 min, vessel diameter was adjusted to thatresulting in themaximal active tension development. To check vessel vi-ability a standard start involving stimulationwith PSS containing 60 mMof potassium chloride (KPSS) and 1 μM phenylephrine was performed.Only segments with a tension development of 1.5 N/m were acceptedfor further studies.
A cumulative concentration–response curve for phenylephrine wasthen obtained and the concentration resulting in a response of 60–70%of the maximal contraction was used for preconstriction of the vesselsegment during the subsequent relaxation experiments. First a cumula-tive concentration–response curve for acetylcholine was performed.After thoroughwash-out the vesselwas incubatedwith theNO synthaseinhibitor NG, NG-dimethyl-L-arginine (ADMA 300 μM) and after 30 minthe concentration–response curve for acetylcholine was repeated.Finally, a cumulative concentration–response curve for the NO donorS-nitroso-N-acetylpenicillamine (SNAP) was constructed.
Acute effects of OA on vascular tone were tested in isolated aortasegments constricted with phenylephrine (1 μM).
2.6. Immunostaining
To study the localization of inducible NOS (iNOS), glucose-relatedprotein 78 (GRP78), and 3-nitrotyrosine thoracic aorta segments werefixedwith cold (4 °C) 4% paraformaldehyde, pH 7.0, for 1 h, and embed-ded in paraffin. Longitudinal sections 5 μmthickwere obtained andpro-cessed following the avidin–biotin–peroxidase method as previouslydescribed (Hernanz et al., 2004). Thus, the sections were incubatedovernight with rabbit polyclonal anti-iNOS (1:1000, TransductionLaboratories, Lexington, UK), anti-GRP78 (1:400, ABR Bioreagents, AHDiagnostic, Copenhagen, Denmark), or rabbit polyclonal antibodyanti-3-nitrotyrosine (1:400) diluted in 1% bovine serum albumin.Then, the sections were reacted with a biotinylated antirabbit immuno-globulins followed by incubation with streptavidin which was conju-gated to horseradish peroxidase (LSAB 2 kit for rat tissue, DAKO,
Denmark), and the immunocomplex was visualized as a brown prod-uct after incubating with 0.05% 3,3-diamino-benzidine and 0.0225%H2O2. Controls were obtained using arterial sections incubated with-out the corresponding primary antibody.
2.7. Immunoblotting
From each animal one proximal abdominal segment was quick-frozen in liquid nitrogen and kept at −70 °C until protein expressionanalysis. Western blots for iNOS and GRP78 were performed as previ-ously described (Hernanz et al., 2004; Østergaard et al., 2009). Immu-noblotting for 3-nitrotyrosine was also intended, but required to largeamounts of proteins. Panactin was used as a comparative control anddid not change in the aorta samples from wild type versus vehicle,oleanolic acid, and fluvastatin-treated ApoE−/− mice.
2.8. Drugs and solutions
The PSS usedwas of the following composition (mM): NaCl 119, KCl4.7, KH2PO4 1.18, MgSO4 1.17, NaHCO3 25, CaCl2 2.5, ethylen diaminetetraacetate (EDTA) 0.027 and glucose 5.5. KPSS was similar to PSSexcept thatNaClwas substituted for KCl to yield a potassium concentra-tion of 60 mM. Phenylephrine, acetylcholine, SNAP, ADMA, Red Oil O,and DMSO were purchased from Sigma-Aldrich (Brøndby, Denmark).OAwas obtained from Extrasynthese (Genay, France), while fluvastatinwas obtained from the pharmacy at Aarhus University Hospital. Stocksolutions of SNAP, OA, and fluvastatin were prepared using DMSO as asolvent and further dilution was made in bidistilled water. Oil Red Owas dissolved in 60% isopropanol and subsequently dextrin wasadded to reduce precipitation. All other drugswere diluted in bidistilledwater.
2.9. Calculations and statistical analysis
All results are expressed asmeans±S.E.M.,wheren equals the num-ber of animals per group. The concentration–response curves and thedata concerning changes in body weight and food consumption during

Fig. 4. The contractile responses of aortic segments to phenylephrine (PE) (A) and the ratiobetween the responses obtained to the PE concentration inducing 60–70% of maximal con-traction after and before incubation with asymmetric dimethylarginine (ADMA; 0.3 mM)(B). The contractile responses to PE were increased in WT and ApoE−/−-oleanolic acid(OA) mice as compared to ApoE−/−-Vehicle and ApoE−/−-Fluvastatin (*Pb0.05). **Pb0.01for ApoE−/−-OA vs. ApoE−/−-Vehicle. n=6–8 in each group.
Fig. 5. Average relaxations of aortic segments to acetylcholine (ACh) before (A) andafter (B) incubation with asymmetric dimethylarginine (ADMA). ACh-induced vasore-laxation was significantly impaired in the three groups of ApoE−/− mice compared toWT mice (Pb0.05). Incubation with ADMA had similar effects in all groups, and nearlyblocked the response to ACh. n=6–8 in each group.
522 N.H. Buus et al. / European Journal of Pharmacology 670 (2011) 519–526
the treatment period were compared by two-way ANOVA followed byBonferroni post-test. Data on atherosclerotic lesion formation, plasmalipids and ratio between tension development to phenylephrine withandwithout ADMAwere statistically evaluated by one-way ANOVA fol-lowed by Bonferroni post-test. A P-value of 0.05 was considered statis-tically significant. All statistical calculations were performed usingGraph Pad Prism (version 4.00, Graph Pad Software Inc., San Diego,CA, USA).
3. Results
3.1. Body weight and food consumption
During the 8-week treatment period ApoE−/−-OA mice had con-stant food consumption and kept a stable body weight of about 30 g,while ApoE−/−-Vehicle, ApoE−/−-Fluvastatin, andWTmice had signif-icantly increased daily food intake and increased body weight to about40 g (Fig. 2A and B).
3.2. Plasma lipids
ApoE−/− mice had pronouncedly elevated plasma levels of choles-terol, which remained unaffected by OA and was only slightly reduced
by fluvastatin (Table 1). The levels of plasma triglyceride were similarin all 4 groups of mice (Table 1).
3.3. Atherosclerotic lesion area
As expected WTmice expressed no signs of atherosclerotic lesionsin neither the aortic arch nor in the descending aorta. In contrastApoE−/−-Vehicle mice developed extensive lesions in the aorticarch, whereas formation in the descending aorta was less pronounced(Fig. 3A). Atherosclerotic lesion formation in the aortic arch was sig-nificantly reduced in OA- and fluvastatin treated ApoE−/− mice(Fig. 3B), but only significantly decreased in the thoracic aorta inthe OA treated group (Fig. 3C).
3.4. Phenylephrine contraction
The contractile response to increased concentrations of phenyl-ephrine was significantly higher in aorta segments from WT andApoE−/−-OA mice as compared to segments from ApoE−/−-Vehicleand ApoE−/−-Fluvastatin mice (Fig. 4A). The phenylephrine doseresulting in 60–70% of themaximal contraction to the initial stimulation

Fig. 6. Representative relaxations in thoracic aortic segments constricted with phenyl-ephrine (PE) followed by addition of increasing concentrations of (A) acetylcholine(ACh) or (B) oleanolic acid (OA). In contrast to ACh, OA did not induce significant re-laxations. The traces are representative of 6 experiments.
Fig. 7. Relaxation of aortic segments to S-nitroso-N-acetylpenicillamine (SNAP).ApoE−/−-oleanolic acid (OA) and ApoE−/−-Fluvastatin mice had a slightly increasedrelaxation to SNAP comparedwith ApoE−/−-Vehicle andWTmice (Pb0.05). The relaxationsof ApoE−/−-OA mice were significantly enhanced compared to ApoE−/−-Fluvastatin mice(Pb0.05). n=6–8 in each group.
523N.H. Buus et al. / European Journal of Pharmacology 670 (2011) 519–526
with KPSS and phenylephrine (10 μM)was between 0.3 and 3 μM. Afterincubation with ADMA the same concentration of phenylephrineresulted in a significantly enhanced contractile response in all groups.The amplification of this response was however significantly smallerin OA-treated and WT mice as compared to fluvastatin- and vehicle-treated ApoE−/− mice (Fig. 4B).
3.5. Responses to acetylcholine and SNAP
In phenylephrine-contracted aorta segments, acetylcholine inducedconcentration-dependent relaxations. Compared to WT all the ApoE−/−
groups displayed a small, but significant, reduction in acetylcholine relax-ation (Fig. 5A). After incubationwith ADMA acetylcholine relaxationwasnearly abolished in all 4 groups of mice (Fig. 5B). In contrast, there wereno effects of OA on vascular tone in phenylephrine-constricted aorta seg-ments fromApoE−/−mice (Fig. 6). However, the same solution of OA didinduce relaxation in phenylephrine-constricted smallmesenteric arteriesfrom Wistar rats as previously described (Rodriguez-Rodriguez et al.,2008).
Relaxation to SNAP was slightly more pronounced in OA- andfluvastatin-treated mice, although at the highest concentration ofSNAP no difference could be detected between any of the groups (Fig. 7).
3.6. Immunostaining and immunoblotting
Representative examples of immunostaining with iNOS, GRP78 and3-nitrotyrosine are shown in Fig. 8. iNOS was mainly expressed in themedia of aorta (Fig. 8A-D), and immunoblotting revealed a slightly,but not significantly, increased expression of iNOS in vehicle-treatedApoE−/− mice, whereas OA treatment reduced iNOS expression withno effect offluvastatin (Fig. 9A). GRP78was expressed in all three layersof aorta (Fig. 8E–H), but there were no differences in the expression ofGRP78 between any of the groups (Fig. 9B). Immunoreaction to 3-nitrotyrosine was found in media and adventitia of ApoE−/− mice(Fig. 8I-L), and scoring of the immunostainings confirmed that 3-nitrotyrosine was markedly upregulated in aortas from ApoE−/−
mice treated with vehicle or fluvastatin, whereas this was not the casein aortas from OA-treated animals (Fig. 10).
4. Discussion
Themain finding of the present study is that oral treatmentwith OAeffectively reduces atherosclerosis development in the aortic arch anddescending aorta of ApoE−/− mice. The mechanism behind this is ap-parently independent of plasma total cholesterol and triglyceride levelsbut seems associated with alterations in vascular contractile reactivitywithout directly influencing eNOS-dependent acetylcholine-inducedrelaxation. Instead OA seems to reduce iNOS activity.
4.1. Effects of OA on body weight
ApoE−/− mice fed a diet mixed with OA, had considerably lowerfood consumption and body weights after 2 weeks of treatment. Apossible explanation could be that OA has a repellent taste or smellto mice, although we did not discover such effects in an initial pilotstudy. Potential toxic effects of OA have to be considered, especiallygiven the fact that this compound appear to exert a variety of biolog-ical effects. Data on toxicity of isolated OA in vivo is sparse, but theacute toxicity of OA is not likely to have any implication in thisstudy, as the oral LD50 value in mice has been reported to be greaterthan 2 g/kg (Singh et al., 1992). The chronic toxicity of OA in mice hasnot yet been studied, but in rats treated daily with 60 mg/kg i.p. for6 weeks, there were no reports of adverse effects on animal health(Somova et al., 2003b). Interestingly, a recent study suggests thatthe effect of OA on body weight in mice could be related to improvedglucose tolerance and reduced visceral fat deposition (de Melo et al.,2010).
4.2. Antiatherogenic effects of OA
This is the first report on OA treatment in atherosclerotic mice andour results demonstrate a pronounced effect on atherosclerosis de-velopment in aorta. The effect is completely independent on totalcholesterol levels which remained very high despite OA treatment.Previous studies of OA treatment have mainly been carried out in rats(Somova et al., 2003a; Rodriguez-Rodriguez et al., 2007; Rodriguez-Rodriguez et al., 2009). However, despite feeding rats a high-fat dietthere is only minimal development of atherosclerosis and instead

Fig. 8. Immunostaining for (A–D) inducible nitric oxide synthase (iNOS); (E–H) glucose-related protein (GRP78); and (I–L) 3-nitrotyrosine in thoracic aortic segments from vehicle-treated wild type (WT) mice, ApoE−/− mice which were treated with either vehicle, oleanolic acid (OA), or fluvastatin. The brown precipitations indicate a positive immunoreac-tion. Immunoreaction for 3-nitrotyrosine was markedly increased in all ApoE−/− mice compared to WT with no differences between the different treatments of ApoE−/− mice. Thepictures are representative for 8–10 mice in each group.
524 N.H. Buus et al. / European Journal of Pharmacology 670 (2011) 519–526
focus have been on the antioxidant properties of olive oil and its variouscomponents (Fitó et al., 2007).
The aortic wall of female ApoE−/−mice has a high level of activationof enzyme systems involved in the production of reactive oxygen spe-cies thought to contribute to plaque formation (Husain et al., 2010).Feeding of ApoE−/− mice with olive oil enriched in minor components,including triterpenoids, decreased induced lipid peroxidation and 8-isoprostaglandin F2α plasma levels with less formation of atherosclerot-ic lesions (Acín et al., 2007; Rosenblat et al., 2008). In the present studyexpression of GRP78, which is a chaperone and can protect against cel-lular oxidative stress in the endoplasmic reticulum (Xu et al., 2004),wasmeasured and foundunaltered. However, another indicator of oxidativestress and peroxynitrite formation (Chew et al., 2009), 3-nitrotyrosine,was markedly upregulated in aorta from ApoE−/− mice compared toaorta from control mice and as opposed to treatment with vehicle orfluvastatin OA reduced immunoreaction to 3-nitrotyrosine to the levelof WT mice.
The precise antiatherogenic effects of OA are still unknown and sev-eral mechanisms can be of importance. Thus OA may protect againstlow density lipoprotein oxidation (Andrikopoulos et al., 2002) or re-duce pro-inflammatory cytokine production (Marquez-Martin et al.,2006). Whether OA affects macrophage and foam cell apoptosis is not
known, but a structural analog to OA, betulinic acid, is a potent antipro-liferative and proapoptotic agent (Yun et al., 2003). Another potentialmechanism that may delay development of atherosclerosis could be areduction in blood pressure level. ApoE knockout mice often developa minor increase in blood pressure as compared to their WT controls(Husain et al., 2010). The effect of OA on blood pressure in ApoE knock-out mice has not been investigated, but in the Dahl salt-sensitive ratmodel of hypertension long termOA treatment to some extent preventsthe increase in blood pressure (Somova et al., 2003b) while oral admin-istration to spontaneously hypertensive rats did not change blood pres-sure (Rodriguez-Rodriguez et al., 2007). Although we cannot exclude apotential effect of OA on blood pressure that may contribute to the ben-eficial effects on atherosclerosis, it seems an unlikely explanation.
We foundfluvastatin to have an antiatherogenic effect despite almostunchanged levels of plasma lipids supporting the concept of “pleiotro-pic” effects of statins (Davignon, 2004) and the findings of other studiesinvestigating the effect of statins in ApoE−/− mice (Johnson et al., 2005;Sparrow et al., 2001). It is notably though, that fluvastatin in one studyhad no effect on atherosclerosis even though it decreased plasma choles-terol (Gervais et al., 2003). The discrepancy between this study and ourfinding may relate to the 10-fold smaller dose and shorter duration oftreatment used in our study.

Fig. 9. (A) Expression of inducible nitric oxide synthase (iNOS) and (B) and glucose-regulated protein (GRP78) in aorta from wild-type mice (WT), and ApoE −/− micetreated with vehicle, oleanolic acid (OA), or fluvastatin. n=5–6 mice in each group.*Pb0.05 OA-treated vs. vehicle-treated ApoE−/− mice.
525N.H. Buus et al. / European Journal of Pharmacology 670 (2011) 519–526
4.3. Vascular function and OA
The effects of hypercholesterolemia and atherosclerosis on vascularcontractile function are controversial, as both decreased and increasedcontractions have been reported. This difference may depend on thecontracting agent used (Ibengwe and Suzuki, 1986). We demonstrateda significantly decreased response to phenylephrine in aortic segmentsfrom ApoE−/− mice as compared to WT mice, a finding which has notpreviously been reported in this animal model. Moreover, inhibitionof NO synthesis by ADMA had greater impact on phenylephrine-induced contractions in arteries from vehicle-treated ApoE−/− mice,
Fig. 10. Immunoreaction scoring of 3-nitrotyrosine in wild type mice (WT) andApoE−/− mice treated with vehicle, oleanolic acid (OA), or fluvastatin. n=5–6 micein each group. *Pb0.05 as compared to WT.
indicating that the decreased contractility in atherosclerotic vesselsmight be due to a higher basal release of NO. However, as acetylcho-line-induced relaxation was only minimally changed in aorta fromApoE−/− mice, the results suggest alterations in non-eNOS derivedNO production. This observation is supported by findings of inductionof a non-eNOSNO synthase in atherosclerotic aortic segments from rab-bits (Verbeuren et al., 1993). Although not evident in the present study,upregulation of iNOS has previously been detected in atheroscleroticvessels of ApoE−/− mice (Detmers et al., 2000; Wilcox et al., 1997;Husain et al., 2010) and, could be partly responsible for the increasedNO synthesis. OA treatment normalized both the contractile responseto phenylephrine and the effect of ADMA on contractility, to the levelof WTmice, suggesting that OA has an inhibitory effect on iNOS. In mu-rine macrophages treated with lipopolysaccharide to induce iNOS ex-pression, OA has in fact been shown to suppress NO release (Ha et al.,2006). Furthermore, OA can inhibit production of tumor necrosis fac-tor-α and interleukin-1β inmurinemacrophages, thus providing a pos-sible mechanistic linkage to iNOS inhibition (Ha et al., 2006).
It can be speculated whether iNOS has a causative role in the path-ophysiology of atherosclerosis. This issue has been elucidated throughthe investigation of iNOS and ApoE double knockoutmice. Thesemice,when fed aWestern-type diet, were found to develop significantly lessatherosclerosis compared to ApoE−/− control mice, and this impact ofiNOS knockout was shown to be more pronounced in older animalswith severe lesions (Detmers et al., 2000; Kuhlencordt et al., 2001).As these data indicate an important role of iNOS in atherosclerosis,the potential iNOS-suppressing effect of OA could account for the anti-atherogenic properties of this compound observed in the presentstudy. In aortae from fluvastatin-treated ApoE−/− mice, the responseto phenylephrine, both with and without ADMA present, was notaltered compared to aortae from vehicle-treated ApoE−/− mice, indi-cating that themechanism behind the antiatherogenic effect of fluvas-tatin was different from that of OA.
4.4. Effects of OA on endothelial function
One of the hallmarks of atherosclerosis is an impaired endothelialfunction, due to a decreased bioavailability of NO. Indeed, in ApoE−/−
mice, impaired endothelium-dependent vasorelaxations in responseto acetylcholine has been observed in segments of the atherosclerosis-prone, proximal descending aorta (Crauwels et al., 2003; d'Uscio et al.,2001). In the present study though, the endothelial dysfunction in theproximal aorta of ApoE−/− mice was not especially pronounced com-pared to the aforementioned studies. These discrepancies are likely tobe due to the shorter period of Western diet feeding and the youngerage of our mice.
As opposed to previous observations obtained in rat aorta and resis-tance arteries (Rodriguez-Rodriguez et al., 2004; Rodriguez-Rodriguezet al., 2008), OA did not relax the isolated mouse aorta. In rat vesselsthis effect of OA involves phosphorylation and activation of eNOS(Rodriguez-Rodriguez et al., 2008). In the present studywe did not per-formmeasurements of eNOS and thus cannot rule out an effect of OA onthis enzyme. However, we could not demonstrate any functional conse-quence of such an effect. Moreover, in ApoE−/− mice the abundance ofiNOS seems far to outweigh the amount of eNOS (Husain et al., 2010)rendering the iNOS a much more important contributor to the basalNO level in this animal model. Taken together our results suggest thatOA treatment has no major effect on eNOS-responsible vasorelaxationbut instead attenuates iNOS induction in the ApoE−/− model ofatherosclerosis.
In conclusion, our study demonstrates for the first time that OA hasbeneficial effects, exceeding those of fluvastatin, on development ofatherosclerosis in ApoE−/− mice. The mechanism by which OA exertsits antiatherogenic effects is lipid-independent, and may be mediatedby an inhibition of iNOS, rather than effects on the bioavailability of

526 N.H. Buus et al. / European Journal of Pharmacology 670 (2011) 519–526
eNOS-derived NO. Further investigation is needed though, to clarify theexact effect of OA on iNOS.
Role of the funding source
The study was financially supported by the Danish Research Councilfor Health and Disease and the Danish Heart Foundation.
Conflict of interest
The authors have no conflicts of interest to disclosure.
References
Acín, S., Navarro,M.A., Perona, J.S., Arbonés-Mainar, J.M., Surra, J.C., Guzmán,M.A., Carnicer,R., Arnal, C., Orman, I., Segovia, J.C., Osada, J., Ruiz-Gutiérrez, V., 2007. Olive oil prepara-tion determines the atherosclerotic protection in apolipoprotein E knockout mice.J. Nutr. Biochem. 18, 418–424.
Andrikopoulos, N.K., Kaliora, A.C., Assimopoulou, A.N., Papageorgiou, V.P., 2002. Inhib-itory activity of minor polyphenolic and nonpolyphenolic constituents of olive oilagainst in vitro low-density lipoprotein oxidation. J. Med. Food 5, 1–7.
Chew, P., Derek, Y.C., Koh, P., Stefanovic, N., Febbraio, M.A., Kola, I., Cooper, M.E., deHaan, J.B., 2009. Site-specific antiatherogenic effect of the antioxidant ebselen inthe diabetic apolipoprotein E-deficient mouse. Arterioscler. Thromb. Vasc. Biol.29, 823–830.
Crauwels, H.M., VanHove, C.E., Holvoet, P., Herman, A.G., Bult, H., 2003. Plaque-associatedendothelial dysfunction in apolipoprotein E-deficient mice on a regular diet. Effect ofhuman apolipoprotein AI. Cardiovasc. Res. 59, 189–199.
d'Uscio, L.V., Baker, T.A., Mantilla, C.B., Smith, L., Weiler, D., Sieck, G.C., Katusic, Z.S.,2001. Mechanisms of endothelial dysfunction in apolipoprotein E-deficient mice.Arterioscler. Thromb. Vasc. Biol. 21, 1017–1022.
Davignon, J., 2004. Beneficial cardiovascular pleiotropic effects of statins. Circulation109, III39–III43.
de Lorgeril, M., Renaud, S., Salen, P., Monjaud, I., Mamelle, N., Martin, J.L., Guidollet, J.,Touboul, P., Delaye, J., 1994. Mediterranean alpha-linolenic acid-rich diet in sec-ondary prevention of coronary heart disease. Lancet 343, 1454–1459.
deMelo, C.L., Queiroz, M.G., Fonseca, S.G., Bizerra, A.M., Lemos, T.L., Melo, T.S., Santos, F.A.,Rao, V.S., 2010. Oleanolic acid, a natural triterpenoid improves blood glucose toler-ance in normal mice and ameliorates visceral obesity in mice fed a high-fat diet.Chem. Biol. Interact. 185, 59–65.
Detmers, P.A., Hernandez,M.,Mudgett, J., Hassing,H., Burton, C.,Mundt, S., Chun, S., Fletcher,D., Card, D.J., Lisnock, J., Weikel, R., Bergstrom, J.D., Shevell, D.E., Hermanowski-Vosatka,A., Sparrow, C.P., Chao, Y.S., Rader, D.J., Wright, S.D., Pure, E., 2000. Deficiency in in-ducible nitric oxide synthase results in reduced atherosclerosis in apolipoproteinE-deficient mice. J. Immunol. 165, 3430–3435.
El, S.N., Karakaya, S., 2009. Olive tree (Olea europaea) leaves: potential beneficial effectson human health. Nutr. Rev. 67, 632–638.
Fitó, M., de la Torre, R., Covas, M.I., 2007. Olive oil and oxidative stress. Mol. Nutr. FoodRes. 51, 1215–1224.
Gervais, M., Pons, S., Nicoletti, A., Cosson, C., Giudicelli, J.F., Richer, C., 2003. Fluvastatinprevents renal dysfunction and vascular NO deficit in apolipoprotein E-deficientmice. Arterioscler. Thromb. Vasc. Biol. 23, 183–189.
Ha, C.L., Weng, C.Y., Wang, L., Lian, T.W., Wu, M.J., 2006. Immunomodulatory effect ofGlossogyne tenuifolia in murine peritoneal macrophages and splenocytes. J. Ethno-pharmacol. 107, 116–125.
Hernanz, R., Alonso, M.J., Zibrandtsen, H., Alvarez, Y., Salaices, M., Simonsen, U., 2004.Measurements of nitric oxide concentrations and hyporeactivity in rat superiormesenteric artery exposed to endotoxin. Cardiovasc. Res. 62, 202–211.
Husain, K., Suarez, E., Isidro, A., Ferder, L., 2010. Effects of paricalcitol and enalapril onatherosclerotic injury in mouse aortas. Am. J. Nephrol. 32, 296–304.
Ibengwe, J.K., Suzuki, H., 1986. Changes in mechanical responses of vascular smoothmuscles to acetylcholine, noradrenaline and high potassium solution in hypercho-lesterolemic rabbits. Br. J. Pharmacol. 87, 395–402.
Johnson, J., Carson, K., Williams, H., Karanam, S., Newby, A., Angelini, G., George, S.,Jackson, C., 2005. Plaque rupture after short periods of fat feeding in the apolipo-protein E-knockout mouse: model characterization and effects of pravastatin treat-ment. Circulation 111, 1422–1430.
Kuhlencordt, P.J., Chen, J., Han, F., Astern, J., Huang, P.L., 2001. Genetic deficiency ofinducible nitric oxide synthase reduces atherosclerosis and lowers plasma lipidperoxides in apolipoprotein E-knockout mice. Circulation 103, 3099–3104.
Liu, J., 1995. Pharmacology of oleanolic acid and ursolic acid. J. Ethnopharmacol. 49,57–68.
Marquez-Martin, M., De La Puerta, R., Fernandez-Arche, A., Ruiz-Gutierrez, V., Yaqoob,P., 2006. Modulation of cytokine secretion by pentacyclic triterpenes from olivepomace oil in human mononuclear cells. Cytokine 36, 211–217.
Mason, R.P., Walter, M.F., Jacob, R.F., 2004. Effects of HMG-CoA reductase inhibitors onendothelial function: role of microdomains and oxidative stress. Circulation 109,II34–II41.
Østergaard, L., Simonsen, U., Eskildsen-Helmond, Y., Vorum, H., Uldbjerg, N., Honoré, B.,Mulvany, M.J., 2009. Proteomics reveals lowering oxygen alters cytoskeletal andendoplasmatic stress proteins in human endothelial cells. Proteomics 9,4457–4467.
Perona, J.S., Cabello-Moruno, R., Ruiz-Gutierrez, V., 2006. The role of virgin olive oilcomponents in the modulation of endothelial function. J. Nutr. Biochem. 17,429–445.
Rodriguez-Rodriguez, R., Herrera, M.D., Perona, J.S., Ruiz-Gutiérrez, V., 2004. Potentialvasorelaxant effects of oleanolic acid and erythrodiol, two triterpenoids containedin 'orojo' olive oil, on rat aorta. Br. J. Nutr. 92, 635–642.
Rodriguez-Rodriguez, R., Herrera, M.D., de Sotamayor, M.A., Ruiz-Gutierrez, V., 2007.Pomace olive oil improves endothelial function in spontaneously hypertensiverats by increasing endothelial nitric oxide synthase expression. Am. J. Hypertens.20, 728–734.
Rodriguez-Rodriguez, R., Stankevicius, E., Herrera, M.D., Østergaard, L., Andersen, M.R.,Ruiz-Gutierrez, V., Simonsen, U., 2008. Oleanolic acid induces relaxation and calci-um-independent release of endothelium-derived nitric oxide. Br. J. Pharmacol.155, 535–546.
Rodriguez-Rodriguez, R., Herrera, M.D., de Sotomayor, M.A., Ruiz-Gutierrez, V., 2009.Effects of pomace olive oil-enriched diets on endothelial function of small mesen-teric arteries from spontaneously hypertensive rats. Br. J. Nutr. 102, 1435–1444.
Rosenblat, M., Volkova, N., Coleman, R., Almagor, Y., Aviram, M., 2008. Antiatherogeni-city of extra virgin olive oil and its enrichment with green tea polyphenols in theatherosclerotic apolipoprotein-E-deficient mice: enhanced macrophage cholester-ol efflux. J. Nutr. Biochem. 19, 514–523.
Simonsen, U., Prieto, D., Mulvany, M.J., Ehrnrooth, E., Korsgaard, N., Nyborg, N.C., 1992.Effect of induced hypercholesterolemia in rabbits on functional responses of isolat-ed large proximal and small distal coronary arteries. Arterioscler. Thromb. 12,380–392.
Singh, G.B., Singh, S., Bani, S., Gupta, B.D., Banerjee, S.K., 1992. Anti-inflammatory activ-ity of oleanolic acid in rats and mice. J. Pharm. Pharmacol. 44, 456–458.
Somova, L.O., Nadar, A., Rammanan, P., Shode, F.O., 2003a. Cardiovascular, antihyperli-pidemic and antioxidant effects of oleanolic and ursolic acids in experimentalhypertension. Phytomedicine 10, 115–121.
Somova, L.I., Shode, F.O., Ramnanan, P., Nadar, A., 2003b. Antihypertensive, antiathero-sclerotic and antioxidant activity of triterpenoids isolated from Olea europaea, sub-species africana leaves. J. Ethnopharmacol. 84, 299–305.
Sparrow, C.P., Burton, C.A., Hernandez, M., Mundt, S., Hassing, H., Patel, S., Rosa, R.,Hermanowski-Vosatka, A., Wang, P.R., Zhang, D., Peterson, L., Detmers, P.A., Chao,Y.S., Wright, S.D., 2001. Simvastatin has anti-inflammatory and antiatherosclero-tic activities independent of plasma cholesterol lowering. Arterioscler. Thromb.Vasc. Biol. 21, 115–121.
Trichopoulou, A., Costacou, T., Bamia, C., Trichopoulos, D., 2003. Adherence to a Medi-terranean diet and survival in a Greek population. N. Engl. J. Med. 348, 2599–2608.
Verbeuren, T.J., Bonhomme, E., Laubie, M., Simonet, S., 1993. Evidence for induction ofnonendothelial NO synthase in aortas of cholesterol-fed rabbits. J. Cardiovasc.Pharmacol. 21, 841–845.
Wilcox, J.N., Subramanian, R.R., Sundell, C.L., Tracey, W.R., Pollock, J.S., Harrison, D.G.,Marsden, P.A., 1997. Expression of multiple isoforms of nitric oxide synthase in nor-mal and atherosclerotic vessels. Arterioscler. Thromb. Vasc. Biol. 17, 2479–2488.
Xu, W., Liu, L., Charles, I.G., Moncada, S., 2004. Nitric oxide induces coupling of mito-chondrial signalling with the endoplasmatic reticulum stress response. NatureCell Biol. 6, 1129–1134.
Yun, Y., Han, S., Park, E., Yim, D., Lee, S., Lee, C.K., Cho, K., Kim, K., 2003. Immunomod-ulatory activity of betulinic acid by producing pro-inflammatory cytokines andactivation of macrophages. Arch. Pharm. Res. 26, 1087–1095.
Zhang, S.H., Reddick, R.L., Piedrahita, J.A., Maeda, N., 1992. Spontaneous hypercholesterol-emia and arterial lesions in mice lacking apolipoprotein E. Science 258, 468–471.
Copyright © 2022 FDOKUMEN




















