
Single Molecule Studies of Antibody–Antigen Interaction Strength Versus Intramolecular Antigen...
10
Single Molecule Studies of Antibody–Antigen Interaction Strength Versus Intra-molecular Antigen Stability Ferry Kienberger 1 , Gerald Kada 1 , Harald Mueller 2 and Peter Hinterdorfer 1 * 1 Institute for Biophysics University of Linz Altenbergerstr. 69, A-4040 Linz Austria 2 Department of Microbiology University of Kassel, D-34132 Kassel, Germany We investigated molecular recognition of antibodies to membrane-antigens and extraction of the antigens out of membranes at the single molecule level. Using dynamic force microscopy imaging and enzyme immuno- assay, binding of anti-sendai antibodies to sendai-epitopes genetically fused into bacteriorhodopsin molecules from purple membranes were detected under physiological conditions. The antibody/antigen interaction strength of 70–170 pN at loading rates of 2–50 nN/second yielded a barrier width of xZ0.12 nm and a kinetic off-rate (corresponding to the barrier height) of k off Z6s K1 , respectively. Bacteriorhodopsin unfolding revealed a characteristic intra-molecular force pattern, in which wild-type and sendai- bacteriorhodopsin molecules were clearly distinguishable in their length distributions, originating from the additional 13 amino acid residues epitope in sendai purple membranes. The inter-molecular antibody/anti- gen unbinding force was significantly lower than the force required to mechanically extract the binding epitope-containing helix pair out of the membrane and unfold it (126 pN compared to 204 pN at the same loading rate), meeting the expectation that inter-molecular unbinding forces are weaker than intra-molecular unfolding forces responsible for stabilizing native conformations of proteins. q 2005 Elsevier Ltd. All rights reserved. Keywords: atomic force microscopy; bacteriorhodopsin; purple membranes; antigen stability; force spectroscopy *Corresponding author Introduction Molecular forces between (i.e. inter-molecular) and within (i.e. intra-molecular) bio-molecules determine their structure, dynamics and function. 1 The inter-molecular binding strength of various ligand–receptor pairs is pivotal for many biological processes such as receptor signaling (e.g. binding of drugs to G-protein coupled receptors) and immune system protection (e.g. antibody–antigen interaction). 2 The underlying forces are not only essential for binding activities but also important for the stability of the native molecular structure. 3 To sustain the native structure of bio-molecules during function, intra-molecular forces need to sufficiently stabilize the three-dimensional confor- mation. Since intra-molecular stability is mandatory during function, the stabilizing free energies have to establish a significant energetic barrier between the native and unfolded states. 4 It is therefore expected that the intra-molecular forces maintaining the native structure of bio-molecules are larger than inter-molecular forces acting during biological processes, an assumption that is not yet proven. Besides its capability of imaging bio-molecules with sub-molecular resolution, 5 the atomic force microscope (AFM) 6 is optimally suited for measur- ing molecular forces at the single-molecule level. 7 AFM was successfully employed to measure inter- molecular forces between various ligand and receptor pairs, 8 and intra-molecular unfolding 0022-2836/$ - see front matter q 2005 Elsevier Ltd. All rights reserved. Present address: G. Kada, Molecular Imaging Corporation, 4666 S. Ash Ave., Tempe, AZ 85282, USA. Abbreviations used: DFM, dynamic force microscopy; EIA, enzyme immunoassay; AFM, atomic force microscope; NTA-His6, nitrilotriacetate-histidine 6; aa, amino acid; sendai-PM, Sendai-purple membrane; BR, bacteriorhodopsin; PBS, phospate buffered saline; BSA, bovine serum albumin; MACmode, magnetic ACmode; pdf, probability density functions. E-mail address of the corresponding author: [email protected] doi:10.1016/j.jmb.2005.01.042 J. Mol. Biol. (2005) 347, 597–606
Transcript of Single Molecule Studies of Antibody–Antigen Interaction Strength Versus Intramolecular Antigen...

doi:10.1016/j.jmb.2005.01.042 J. Mol. Biol. (2005) 347, 597–606
Single Molecule Studies of Antibody–AntigenInteraction Strength Versus Intra-molecular AntigenStability
Ferry Kienberger1, Gerald Kada1, Harald Mueller2
and Peter Hinterdorfer1*
1Institute for BiophysicsUniversity of LinzAltenbergerstr. 69, A-4040 LinzAustria
2Department of MicrobiologyUniversity of Kassel, D-34132Kassel, Germany
0022-2836/$ - see front matter q 2005 E
Present address: G. Kada, MolecuCorporation, 4666 S. Ash Ave., Tem
Abbreviations used: DFM, dynamEIA, enzyme immunoassay; AFM, amicroscope; NTA-His6, nitrilotriaceamino acid; sendai-PM, Sendai-purbacteriorhodopsin; PBS, phospate bbovine serum albumin; MACmode,pdf, probability density functions.
E-mail address of the [email protected]
We investigated molecular recognition of antibodies to membrane-antigensand extraction of the antigens out of membranes at the single moleculelevel. Using dynamic force microscopy imaging and enzyme immuno-assay, binding of anti-sendai antibodies to sendai-epitopes geneticallyfused into bacteriorhodopsin molecules from purple membranes weredetected under physiological conditions. The antibody/antigen interactionstrength of 70–170 pN at loading rates of 2–50 nN/second yielded a barrierwidth of xZ0.12 nm and a kinetic off-rate (corresponding to the barrierheight) of koffZ6 sK1, respectively. Bacteriorhodopsin unfolding revealed acharacteristic intra-molecular force pattern, in which wild-type and sendai-bacteriorhodopsin molecules were clearly distinguishable in their lengthdistributions, originating from the additional 13 amino acid residuesepitope in sendai purple membranes. The inter-molecular antibody/anti-gen unbinding force was significantly lower than the force required tomechanically extract the binding epitope-containing helix pair out of themembrane and unfold it (126 pN compared to 204 pN at the same loadingrate), meeting the expectation that inter-molecular unbinding forces areweaker than intra-molecular unfolding forces responsible for stabilizingnative conformations of proteins.
q 2005 Elsevier Ltd. All rights reserved.
Keywords: atomic force microscopy; bacteriorhodopsin; purple membranes;antigen stability; force spectroscopy
*Corresponding authorIntroduction
Molecular forces between (i.e. inter-molecular)and within (i.e. intra-molecular) bio-moleculesdetermine their structure, dynamics and function.1
The inter-molecular binding strength of variousligand–receptor pairs is pivotal for many biologicalprocesses such as receptor signaling (e.g. binding ofdrugs to G-protein coupled receptors) and immunesystem protection (e.g. antibody–antigen
lsevier Ltd. All rights reserve
lar Imagingpe, AZ 85282, USA.ic force microscopy;tomic force
tate-histidine 6; aa,ple membrane; BR,uffered saline; BSA,magnetic ACmode;
ing author:
interaction).2 The underlying forces are not onlyessential for binding activities but also importantfor the stability of the native molecular structure.3
To sustain the native structure of bio-moleculesduring function, intra-molecular forces need tosufficiently stabilize the three-dimensional confor-mation. Since intra-molecular stability is mandatoryduring function, the stabilizing free energies have toestablish a significant energetic barrier between thenative and unfolded states.4 It is therefore expectedthat the intra-molecular forces maintaining thenative structure of bio-molecules are larger thaninter-molecular forces acting during biologicalprocesses, an assumption that is not yet proven.
Besides its capability of imaging bio-moleculeswith sub-molecular resolution,5 the atomic forcemicroscope (AFM)6 is optimally suited for measur-ing molecular forces at the single-molecule level.7
AFM was successfully employed to measure inter-molecular forces between various ligand andreceptor pairs,8 and intra-molecular unfolding
d.

598 Single Molecule Inter and Intra-molecular Forces
forces of individual proteins.9 For measurements ofinter-molecular receptor/ligand binding strengths,ligands are attached to AFM tips and receptors toprobe surfaces (or vice versa). The AFM tip isbrought into contact with the surface so as to formreceptor–ligand complexes. Upon the subsequentcontinuous retraction of the tip from the surface, asteadily increasing force is applied to the ligand–receptor connection until the bond breaks at acritical force (termed unbinding force). Such experi-ments allow for estimation of affinity, rate constants,and structural data of the binding pocket.10 Inaddition, they provide insights into the moleculardynamics of the receptor–ligand recognition pro-cess11 by varying the loading rate of force appliance(termed as force-spectroscopy).12 Applicationsinclude ligand–receptor interactions like avidin-biotin13 and nitrilotriacetate-histidine 6 (NTA-His6;14)antibody–antigen15 and cellular proteins, eitherisolated10 or in cell membranes.16
For single molecule unfolding experiments, abare AFM tip is heavily pushed into a layer ofproteins or other molecules attached to a probesurface, followed by retraction of the tip. Even-tually, the tip picks up a molecule from the surfaceby unspecific adsorption and the retraction ofthe tip will consequently induce a stretching of thesuspended molecule. These studies elucidate themolecular determinants of mechanical stability andthe role of force-induced conformational changes.17
Force-induced domain unfolding in proteins,18
length transitions caused by conformationalchanges in the sugar rings of polysaccharides,19
and transitions in the secondary structure of DNAmolecules20 were investigated in great detail usingatomic force microscopy.
In this study, we investigated the molecularbinding strength of antibodies to their respectiveantigenic sites and compared it to the intra-molecular stability of the antigenic moleculesusing single molecule force spectroscopy. Theantigenic site consisted of a 13 amino acidresidues (aa) long linear recognition epitopethat was appended between the E and F loopof bacteriorhodopsin, a seven-helix membranespanning protein of purple membranes. Com-parison of the forces yielded direct insight intothe mechanical stability of the antibody–antigencomplex, and antigen resistance against extrac-tion from the membrane and unfolding. Theresults obtained in this study demonstrate thepotential of force microscopy as a novel tech-nique to explore fundamental mechanisms ofantibody recognition versus intra-molecularprotein stability.
Results
Antibody binding to sendai-purple membrane
Sendai-purple membrane (sendai-PM) patchesisolated from Halobacterium salinarum were
adsorbed on mica with a preference of the cyto-plasmic side facing towards the subphase.21 Topo-graphy images were acquired using magneticACmode (MACmode) AFM, where an alternatingmagnetic field drives the oscillation of a magneti-cally coated cantilever22,23 (Figure 1(a)–(c)). sendai-PM were 5(G1) nm in height and exhibiteda hexagonal arrangement of trimeric bacterio-rhodopsin (BR) molecules, similar to the appear-ance of wild-type PM.24 In contrast to wild-typePM, sendai-PM have a 13 aa long sendai epitopegenetically fused in the cytoplasmatic E-F loop ofBR (Figure 1(e)). The latter peptide sequencerepresents a specific high-affinity binding targetfor monoclonal anti-sendai-antibodies.25 The speci-ficity of anti-sendai-antibody binding to sendai-PMwas shown using biochemical techniques and AFMmeasurements.
Enzyme immunoassays (EIA) yielded specificbinding of anti-sendai-antibodies to sendai-PM(Figure 1(d)). A strong binding signal was obtainedafter the sendai-PM were incubated with anti-sendai-antibodies (open circles) and subsequentlymarked with secondary enzyme-linked antibodies(cf. Materials and Methods). In contrast, the bindingsignal was significantly lower when incubationwith the primary antibody on sendai-PM wasomitted (filled circles) or wild-type PM was used(diamonds). Topographical images of sendai-PMprior to antibody incubation revealed smooth andflat membrane patches with diameters of w1 mm(Figures 1(a) and 2(a)) and a vertical surfaceroughness of w0.5 nm (Figure 2(a), lower panel).Subsequently, a low concentration of anti-sendai-antibodies was injected into the fluid cell of theAFM and images were taken at different times ofincubation (Figure 2). The sendai-PM patch inFigure 2(b) revealed clearly resolved individualanti-sendai-antibodies on top of the membranesurface.26 Antibodies appeared as bright dots withw15 nm in diameter and 2–3 nm in height. Thelower panel of Figure 2(b) shows the cross-sectionprofile of the continuous line in the topographicalimage. Four antibodies were detected along this linewith a high signal-to-noise ratio (see arrows).Increasing the incubation time from 30 minutes to60 minutes led to a higher coverage of antibodies onsendai-PM (Figure 2(c), upper panel). The cross-section profile (Figure 2(c), lower panel) stillexhibits individual antibodies, although their sur-face coverage is already w60%. At 90 minutesincubation time, most of the sendai-PM patcheswere fully covered with antibodies (Figure 2(d),upper panel). Individual antibodies were notresolved anymore, however, their presence wasevident due to a height increase from 5 nm (baresendai-PM, Figure 2(a), lower panel) to 7.5 nm(fully covered sendai-PM, Figure 2(d), lower panel).No antibodies were detected on wild-type PM evenafter prolonged times of antibody incubation time(data not shown), confirming the specificity ofantibody binding.
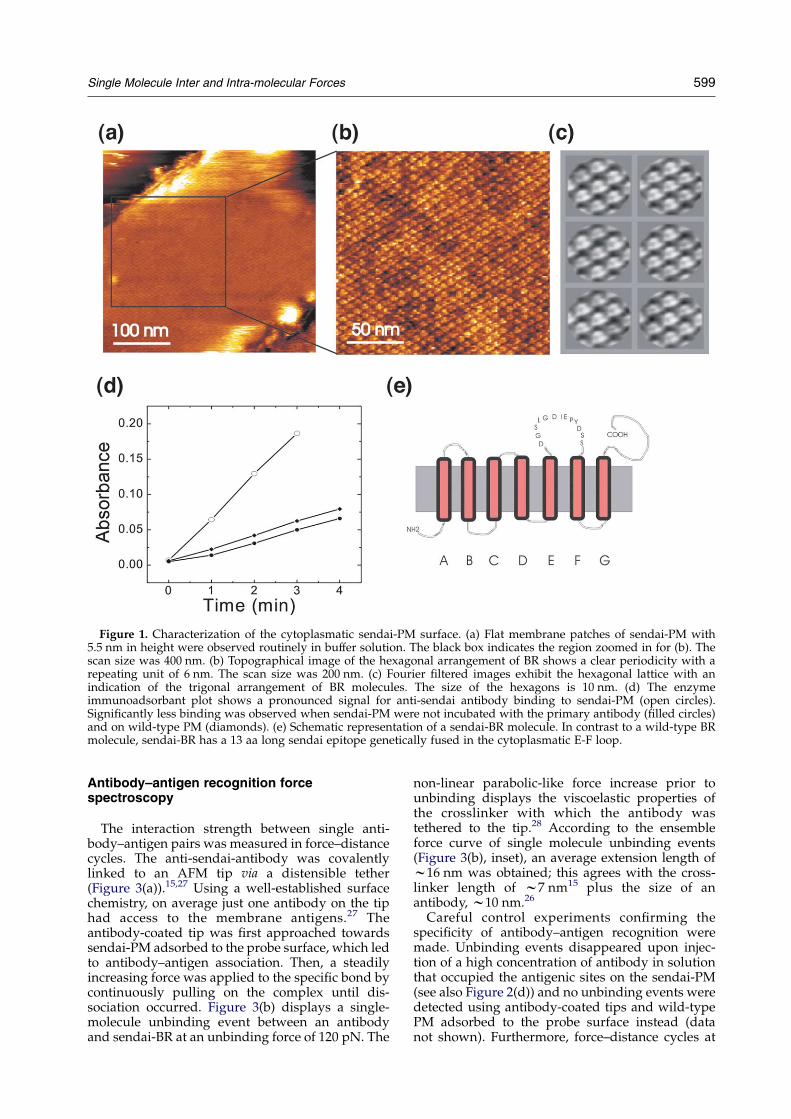
Figure 1. Characterization of the cytoplasmatic sendai-PM surface. (a) Flat membrane patches of sendai-PM with5.5 nm in height were observed routinely in buffer solution. The black box indicates the region zoomed in for (b). Thescan size was 400 nm. (b) Topographical image of the hexagonal arrangement of BR shows a clear periodicity with arepeating unit of 6 nm. The scan size was 200 nm. (c) Fourier filtered images exhibit the hexagonal lattice with anindication of the trigonal arrangement of BR molecules. The size of the hexagons is 10 nm. (d) The enzymeimmunoadsorbant plot shows a pronounced signal for anti-sendai antibody binding to sendai-PM (open circles).Significantly less binding was observed when sendai-PM were not incubated with the primary antibody (filled circles)and on wild-type PM (diamonds). (e) Schematic representation of a sendai-BR molecule. In contrast to a wild-type BRmolecule, sendai-BR has a 13 aa long sendai epitope genetically fused in the cytoplasmatic E-F loop.
Single Molecule Inter and Intra-molecular Forces 599
Antibody–antigen recognition forcespectroscopy
The interaction strength between single anti-body–antigen pairs was measured in force–distancecycles. The anti-sendai-antibody was covalentlylinked to an AFM tip via a distensible tether(Figure 3(a)).15,27 Using a well-established surfacechemistry, on average just one antibody on the tiphad access to the membrane antigens.27 Theantibody-coated tip was first approached towardssendai-PM adsorbed to the probe surface, which ledto antibody–antigen association. Then, a steadilyincreasing force was applied to the specific bond bycontinuously pulling on the complex until dis-sociation occurred. Figure 3(b) displays a single-molecule unbinding event between an antibodyand sendai-BR at an unbinding force of 120 pN. The
non-linear parabolic-like force increase prior tounbinding displays the viscoelastic properties ofthe crosslinker with which the antibody wastethered to the tip.28 According to the ensembleforce curve of single molecule unbinding events(Figure 3(b), inset), an average extension length ofw16 nm was obtained; this agrees with the cross-linker length of w7 nm15 plus the size of anantibody, w10 nm.26
Careful control experiments confirming thespecificity of antibody–antigen recognition weremade. Unbinding events disappeared upon injec-tion of a high concentration of antibody in solutionthat occupied the antigenic sites on the sendai-PM(see also Figure 2(d)) and no unbinding events weredetected using antibody-coated tips and wild-typePM adsorbed to the probe surface instead (datanot shown). Furthermore, force–distance cycles at

Figure 2. Antibodies specifically bound to sendai-PM at different incubation times. (a) Flat membrane patches of sendai-PM were observed prior to antibody incubation (upper panel) with a surface roughness of 0.5 nm, measured in a cross-section profile (lower panel) along the continuous line in (a). Scan size was 900 nm. (b) 30 minutes after anti-sendai antibodyinjection into the liquid cell of the AFM, individual antibodies were observed as small dots with 10–15 nm in diameter and 2–3 nm in height (upper panel). The cross-section profile (lower panel) along the continuous line in (b) shows four antibodieswith a high signal-to-noise ratio (arrows). The scan size was 600 nm. (c) 60 minutes after antibody injection, a high coveragewas observed on the cytoplasmatic side of sendai-PM. Individual antibodies were detected on both the topographical image(upper panel) and on the cross-section profile (lower panel). The scan size was 1000 nm. (d) A full coverage of the membraneswas obtained after 90 minutes of incubation (upper panel). The layer of antibodies increased the height of the membranes,from 5 nm without antibodies to 7.5 nm with a full layer of antibodies (lower panel). The scan size was 800 nm.
600 Single Molecule Inter and Intra-molecular Forces

Figure 3. Single molecule antibody–antigen recognition. (a) Scheme depicting an AFM tip to which an antibody iscovalently tethered via a crosslinker. (b) Force–distance cycle. A single molecule unbinding event resulting in anunbinding force of 120 pN and an unbinding length of 15 nm is shown. The force profile reflects the increasing forceapplied to the antibody–antigen bond until it suddenly breaks at a certain unbinding force. Inset: Superposition ofantibody–antigen unbinding curves exhibiting the stretching of the crosslinker and molecular unbinding events at anaverage extension of 16 nm; 17 single force profiles were used for the superposition. (c) Probability density function. Theplot shows a pdf for the antibody–antigen interaction at a loading rate of 9 nN/second. The Gaussian fit (dotted line)reveals the most probable unbinding force fu (126 pN at the corresponding loading rate) and the respective standarddeviation. The pdf is constructed from 70 single molecule unbinding forces and their respective standard deviations.(d) Unbinding force fu in dependence of the loading rate r (force spectroscopy). The most probable unbinding force fu isplotted against the loading rate r on a semi-logarithmic scale. The continuous line shows the fit using equation (1),resulting in the separation of the energetic barrier to the equilibrium position, xZ0.12 nm, and the dissociation constantat zero force, koffZ6 sK1.
Single Molecule Inter and Intra-molecular Forces 601
standard conditions on sendai-PM with a bare tipdid also not reveal unbinding force signals. Fromsingle molecule force measurements, empiricalprobability density functions (pdf) of unbindingforces were constructed.29 A pdf consisting of 70single unbinding force values and their respectivestandard deviations is depicted in Figure 3(c).The pdf was fitted using a Gaussian function(Figure 3(c), dotted line), yielding a most probableunbinding force fu (maximum of distribution) of126 pN.
Since receptor–ligand interactions are dependenton the dynamics of the experiment,30 we measuredthe unbinding force fu as a function of the loadingrate r. For several different retract velocities in therange of 200 nm/second to 5000 nm/second, pdfsof typically 70 unbinding events were calculatedand the most probable unbinding forces fu wereplotted against the respective loading rate r. Theloading rate r (pN/second) was calculated from theretract velocity (nm/second) times the total elas-ticity (pN/nm) (tether plus cantilever spring

602 Single Molecule Inter and Intra-molecular Forces
constant) upon unbinding of the ligand–receptorcomplex. According to theory, a linear rise of theunbinding force with respect to a logarithmicallyincreasing loading rate is characteristic for asingle-energy barrier in the thermally activatedregime.12,30,31 Hereby, the unbinding force fu isrelated to the loading rate r through:32
fu ¼ ðkBT=xÞ ln ðrx=ðkBTkoffÞÞ (1)
where x is the separation of the energetic barrierto the equilibrium position, koff is the disso-ciation constant at zero force, and kBT is the thermalenergy. fu and r are the most probable unbindingforce and the loading rate, respectively. Theantibody–antigen unbinding force varied in therange of 70–170 pN, depending on the appliedloading rate. A linear increase of the most probableunbinding force versus loading rate was indeedfound for the anti-sendai–antibody/sendai-BRinteraction shown in the semi-logarithmic plot inFigure 3(d). From the force spectroscopy plot, theseparation of the energetic barrier from the equili-brium position x and the koff rate were calculatedusing equation (1), resulting in xZ0.12 nm andkoffZ6 sK1, respectively.
Bacteriorhodopsin unfolding
Force–distance cycles under standard conditionson sendai-PM with a bare tip did not reveal anyattractive force signals. However, when the inter-action time of the tip on the sendai-PM wasincreased to one second at a compression force of1 nN, characteristic force patterns due to thesequential extraction and subsequent unfolding ofsendai-BR were observed (Figure 4(a)). In most ofthe force–distance cycles a characteristic pattern offour peaks was obtained because the tip mainlypicked up the BR at its C terminus.33 A super-imposed force spectrum of sendai-BR unfolding isshown in Figure 4(b). With each peak in the force–distance cycles, the length of the chain increased bythe respective helix pair that was pulled out of themembrane. Using a worm-like chain model with apersistence length of 0.4 nm,34 and the contourlengths derived from the model of pair-wise helixextraction depicted in Figure 4(c),33 an almostperfect overlap between the model and themeasurements was revealed (Figure 4(b), continu-ous lines). Apparently, the BR molecule was pulledout of the membrane by a sequential extraction andsimultaneous unfolding of single helix pairs. Ananalysis of the force distribution of respectiveextraction force peaks is shown in Figure 4(d). Thefour peaks (first, continuous line; second, brokenline; third, dotted line; fourth, dash-dot line)unfolded consecutively at lower forces, from164 pN for the first peak to 80 pN for the fourthpeak.
Unfolding experiments were also performed withwild-type PM. The superimposed force-spectrum ofwild-type BR exhibited an unfolding pattern con-sisting of four peaks; however, shorter characteristic
lengths compared to sendai-BR. Both for wild-typeand sendai-BR the lengths of the force peaks takenfrom the respective force spectra exactly matchedthe theoretical lengths (Figure 4(e)). While thelength of the first peak of wild-type and sendai-BR practically overlap, the succeeding lengths ofwild-type BR were about 5 nm shorter (Figure 4(e)),due to the missing sendai sequence (13 aa) inthe E-F loop.
Discussion
DFM permits in vivo studies of dynamic bio-logical processes in physiological conditions withhigh lateral resolution. Individual antibodies boundto sendai-PM were imaged at different densities.Depending on the incubation time, either singleantibodies or a dense layer of bound antibodieswere observed. These experiments demonstratethat upon injection of a high concentration ofantibodies in solution the antigenic sites on sendai-PM are occupied, which corroborates the blockingmethodology in our single molecule force spectro-scopy experiments. The strength of antibody bind-ing was investigated in force spectroscopyexperiments, yielding loading rate-dependent inter-action forces in the range of 70–170 pN. Since theforces showed a strictly mono-logarithmical depen-dence on the loading rate, only one single sharpenergy barrier was probed in the time-rangeaccessible in this study.30 The linear fit of the forcespectroscopy curve yielded the length dimension ofthe energy barrier along the dissociation path, x,and the kinetic off-rate of dissociation, koff. Thelatter value corresponds to the off-rate in solution ifthe energy barrier dominating the behavior of thebond in force experiments is also the most promin-ent barrier in solution. Since the energy landscapeof receptor–ligand bonds can be rugged terrains,several energy barriers were often found using atechnique that allows variation of the loading rateby many orders of magnitudes.30 The high kineticoff-rate of the antibody–sendai-BR interactiongained from the force spectroscopy curve maytherefore reflect a different energy barrier than insolution.35
BR molecules were pulled out of the membraneby sequential extraction and simultaneous unfold-ing of single helix pairs. The respective extractionforces revealed that the helix pairs unfoldedconsecutively at lower forces, most likely causedby a subsequent loss of BR stability during the step-wise extraction process.36 From the length distri-butions of the intra-molecular unfolding forcepattern the active antigen, sendai-BR, could bedistinguished from the wild-type BR. Since the 13aa long sendai epitope resulted in 5 nm extension ofthe unfolding length, almost identical trans-membrane molecules were differentiated bymeasuring their unfolding force pattern.
The force required for pulling out the two helixpairs that are connected to the sendai epitope was
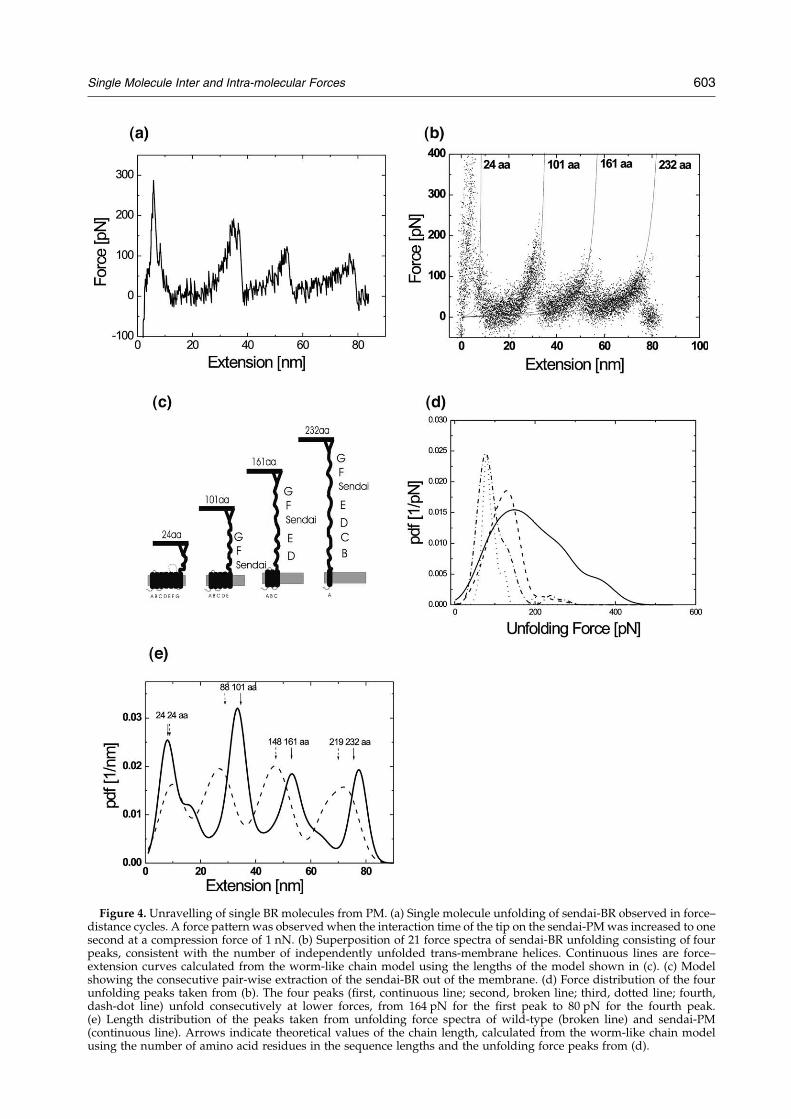
Figure 4. Unravelling of single BR molecules from PM. (a) Single molecule unfolding of sendai-BR observed in force–distance cycles. A force pattern was observed when the interaction time of the tip on the sendai-PM was increased to onesecond at a compression force of 1 nN. (b) Superposition of 21 force spectra of sendai-BR unfolding consisting of fourpeaks, consistent with the number of independently unfolded trans-membrane helices. Continuous lines are force–extension curves calculated from the worm-like chain model using the lengths of the model shown in (c). (c) Modelshowing the consecutive pair-wise extraction of the sendai-BR out of the membrane. (d) Force distribution of the fourunfolding peaks taken from (b). The four peaks (first, continuous line; second, broken line; third, dotted line; fourth,dash-dot line) unfold consecutively at lower forces, from 164 pN for the first peak to 80 pN for the fourth peak.(e) Length distribution of the peaks taken from unfolding force spectra of wild-type (broken line) and sendai-PM(continuous line). Arrows indicate theoretical values of the chain length, calculated from the worm-like chain modelusing the number of amino acid residues in the sequence lengths and the unfolding force peaks from (d).
Single Molecule Inter and Intra-molecular Forces 603

604 Single Molecule Inter and Intra-molecular Forces
204 pN (force of second plus force of third peak inFigure 4(d)). Therefore, the mechanical stability ofextracting and unfolding a single sendai-BR mol-ecule out of the membrane was found to besignificantly larger than the mechanically inducedexertion arising from a single antibody–antigenunbinding event (126 pN; compared at the sameloading rate). Thus, the intra-molecular stability ofthe antigenic molecule was guaranteed during anantibody–antigen unbinding event and the BRmolecules remained folded despite the mechanicalloading throughout the experiment.
Materials and Methods
Construction and expression of sendai-BR
The derivative bacteriorhodopsingen with the addi-tional nucleotide sequence coding for the C-terminal epi-tope of the sendai virus L-protein (DGSLGDIEPYDSS) inthe EF-loop of BR was obtained with PCR-mutagenesis,recloned into the wild-type bacteriorhodopsingen (bop) ofthe halobacterial expression vector pFG6,37 transformedin H. salinarum L-33, and cultured as described.38
Enzyme immuno assay (EIA)
The specificity and density of antibody-binding sites ofsendai-PM adsorbed on mica was determined with anEIA similar to that mentioned in the work done byHinterdorfer et al.27 First, sendai-PM were incubated onmica in phospate-buffered saline (PBS) in order to get asurface density of sendai-PM of about 10%. This step wasfollowed by incubation for 15 minutes with PBA (5 mg/mlof bovine serum albumin (BSA; Sigma, Vienna, Austria) inPBS). Then a solution of 55 mg/ml of anti-sendai antibodyin PBA was applied to the probes for 50 minutes, washedwith PBA, incubated in 60 mg/ml of horseradish peroxi-dase-goat-anti-rabbit (HRP-GAR) antibody (Biomedica,Vienna, Austria) in PBA for another 50 minutes, andwashed with PBA and PBS. For blank values, the bindingstep with the primary anti-sendai antibody was omitted.Enzyme density measurements were carried out byinserting the substrates in 3 ml photometer cuvettescontaining 3 ml of 0.8 mg/ml o-phenylendiamine$2 HCl(Sigma, Deisenhofen, Germany) in 50 mM citric acid–NaOH (pH 5.5) plus 30 ml of 3% H2O2 (v/v). The solutionin the cuvettes was continuously stirred and the cuvetteswere placed every minute in a spectrophotometer(Shimadzu UV-160) to measure E450. Enzyme densitieswere calculated by comparing the observed dE450/dtvalues with calibration experiments in solution.
AFM imaging
Purple membranes prepared as described39 wereadsorbed in a buffer solution (300 mM KCl, 20 mM Tris,pH 8.8) onto freshly cleaved mica and subsequentlywashed with the same buffer at pH 7.6. Images wereacquired in this buffer with a magnetically drivendynamic force microscope (MOLECULAR IMAGING,Tempe, AZ) using magnetically coated MacLeverse(MOLECULAR IMAGING, Tempe, AZ) at 5 nm oscil-lation amplitude and a nominal spring constant of100 pN/nm. Images were taken with the lowest possibleamplitude reduction at a line scan rate of 1.3 Hz.
Figure 1(c) was Fourier filtered using IMAGIC (ImageScience Software GmbH, Berlin, Germany). Anti-sendai-antibodies were injected into the fluid cell of the AFMresulting in a final concentration of 3 mg/ml.
Force spectroscopy
Coupling of antibodies to AFM tips was done asdescribed.27 Briefly, antibodies were covalently coupledto AFM-tips using a three-step binding protocol. Bindingof 2-aminoethanol-HCl to Si3N4 tips (Veeco Instruments,Santa Barbara, CA), was followed by coupling of aheterobifunctional tether (NHS-PEG18-PDP, N-hydroxy-succinimide ester-poly(ethylene glycol)18-2-pyridyl-dithiopropionyl) to the amine-containing tip surface insolvent. Prior to conjugation of anti-sendai-antibodies tothe thiol-reactive end of the tip-bound tether in buffersolution, anti-sendai-antibodies were labeled withN-succinimidyl acetylthiopropionate (SATP). The such-like modified tips were washed with the same buffer andstored in the cold room until use. For antibody–antigenforce measurements conventional force–distance cycleswere used to detect single molecule antibody–antigenunbinding events for different force velocities. Force–distance cycles were performed with velocities rangingfrom 20 nm/second to 700 nm/second. Each individualcantilever spring constant was calibrated in solutionusing the equi-partition theorem.40 The recognition forceswere quantified using a transition detection algorithmfrom which empirical probability density functions (pdfs)of forces and lengths were constructed.29 The mostprobable unbinding force and respective standard devi-ations were fitted from the pdfs and the most probableunbinding forces were plotted in a half logarithmic forcespectroscopy plot.
Bacteriorhodopsin unfolding
The contact time between unmodified Si3N4 tips andsendai-PM was increased to one second and BR unfoldingevents were observed with about 30% probability inconventional force–distance cycles.33 Force–distancecycles were carried out at a fixed force velocity of90 nm/second. For Figure 4(b), 21 single moleculeunfolding curves were superimposed and fitted using aworm-like chain model.
Acknowledgements
This work was supported by the Austrian ScienceFoundation Projects 12802/14549. We thankChristian Rankl for helpful discussions.
References
1. Sinha, N. & Smith-Gill, S. J. (2002). Protein structure tofunction via dynamics. Protein Pept. Letters, 9, 367–377.
2. Allison, D. P., Hinterdorfer, P. & Han, W. (2002).Biomolecular force measurements and the atomicforce microscope. Curr. Opin. Biotechnol. 13, 47–51.
3. Muller, D. J., Janovjak, H., Lehto, T., Kuerschner, L. &Anderson, K. (2002). Observing structure, functionand assembly of single proteins by AFM. Prog.Biophys. Mol. Biol. 79, 1–43.

Single Molecule Inter and Intra-molecular Forces 605
4. White, S. H. & Wimley, W. C. (1999). Membraneprotein folding and stability: physical principles.Annu. Rev. Biophys. Biomol. Struct. 28, 319–365.
5. Engel, A. & Muller, D. J. (2000). Observing singlebiomolecules at work with the atomic force micro-scope. Nature Struct. Biol. 7, 715–718.
6. Binnig, G., Quate, C. F. & Gerber, C. (1986). Atomicforce microscope. Phys. Rev. Letters, 56, 930–933.
7. Willemsen, O. H., Snel, M. M., Cambi, A., Greve, J., DeGrooth, B. G. & Figdor, C. G. (2000). Biomolecularinteractions measured by atomic force microscopy.Biophys. J. 79, 3267–3281.
8. Hinterdorfer, P. (2002). Molecular recognition studiesusing the atomic force microscope. Methods Cell. Biol.68, 115–139.
9. Fisher, T. E., Oberhauser, A. F., Carrion-Vazquez, M.,Marszalek, P. E. & Fernandez, J. M. (1999). The studyof protein mechanics with the atomic force micro-scope. Trends Biochem. Sci. 24, 379–384.
10. Baumgartner, W., Hinterdorfer, P., Ness, W., Raab, A.,Vestweber, D., Schindler, H. & Drenckhahn, D. (2000).Cadherin interaction probed by atomic forcemicroscopy. Proc. Natl Acad. Sci. USA, 97, 4005–4010.
11. Nevo, R., Stroh, C., Kienberger, F., Kaftan, D.,Brumfeld, V., Elbaum, M. et al. (2003). A molecularswitch between alternative conformational states inthe complex of Ran and importin beta1. Nature Struct.Biol. 10, 553–557.
12. Evans, E. & Ritchie, K. (1997). Dynamic strength ofmolecular adhesion bonds. Biophys. J. 72, 1541–1555.
13. Lee, G. U., Chrisey, A. C. & Colton, R. J. (1994).Sensing discrete streptavidin-biotin interactions withatomic force microscopy. Langmuir, 10, 354–357.
14. Kienberger, F., Kada, G., Gruber, H. J., Pastushenko,V., Riener, C., Trieb, M. et al. (2000). Recognition forcespectroscopy studies of the NTA-His6 bond. SingleMol. 1, 59–65.
15. Hinterdorfer, P., Baumgartner, W., Gruber, H. J.,Schilcher, K. & Schindler, H. (1996). Detection andlocalization of individual antibody-antigen recog-nition events by atomic force microscopy. Proc. NatlAcad. Sci. USA, 93, 3477–3481.
16. Lehenkari, P. P. & Horton, M. A. (1999). Single integrinmolecule adhesion forces in intact cells measured byatomic force microscopy. Biochem. Biophys. Res. Com-mun. 259, 645–650.
17. Fisher, T. E., Marszalek, P. E. & Fernandez, J. M.(2000). Stretching single molecules into novel confor-mations using the atomic force microscope. NatureStruct. Biol. 7, 719–724.
18. Rief, M., Gautel, M., Oesterhelt, F., Fernandez, J. M. &Gaub, H. E. (1997). Reversible unfolding of individualtitin immunoglobulin domains by AFM. Science, 276,1109–1112.
18. Marszalek, P. E., Pang, Y. P., Li, H., El Yazal, J.,Oberhauser, A. F. & Fernandez, J. M. (1999). Atomiclevers control pyranose ring conformations. Proc. NatlAcad. Sci. USA, 96, 7894–7898.
20. Rief, M., Clausen-Schaumann, H. & Gaub, H. E.(1999). Sequence-dependent mechanics of singleDNA molecules. Nature Struct. Biol. 6, 346–349.
21. Muller, D. J., Schabert, F. A., Buldt, G. & Engel, A.(1995). Imaging purple membranes in aqueoussolutions at sub-nanometer resolution by atomicforce microscopy. Biophys. J. 68, 1681–1686.
22. Han, W., Lindsay, S. M. & Jing, T. (1996).A magnetically driven oscillating probe microscopefor operation in liquid. Appl. Phys. Letters, 69, 1–3.
23. Kienberger, F., Stroh, C., Kada, G., Moser, R.,
Baumgartner, W., Pastushenko, V. et al. (2003).Dynamic force microscopy imaging of native mem-branes. Ultramicroscopy, 97, 229–237.
24. Muller, D. J., Heymann, J. B., Oesterhelt, F., Moller, C.,Gaub, H., Buldt, G. & Engel, A. (2000). Atomic forcemicroscopy of native purple membrane. Biochim.Biophys. Acta, 1460, 27–38.
25. Einberger, H., Mertz, R., Hofschneider, P. H. &Neubert, W. J. (1990). Purification, renaturation, andreconstituted protein kinase activity of the Sendaivirus large (L) protein: L protein phosphorylates theNP and P proteins in vitro. J. Virol. 64, 4274–4280.
26. Kienberger, F., Muller, H., Pastushenko, V. &Hinterdorfer, P. (2004). Following single antibodybinding to purple membranes in real time. EMBORep.5, 579–583.
27. Hinterdorfer, P., Schilcher, K., Baumgartner, W.,Gruber, H. J. & Schindler, H. (1998). A mechanisticstudy of the dissociation of individual antibody-antigen pairs by atomic force microscopy. Nanobiology,4, 177–188.
28. Kienberger, F., Pastushenko, V., Kada, G., Gruber,H. J., Riener, C., Schindler, H. & Hinterdorfer, P.(2000). Static and dynamical properties of singlepoly(ethylene glycol) molecules investigated byforce spectroscopy. Single Mol. 1, 123–128.
29. Baumgartner, W., Hinterdorfer, P. & Schindler, H.(2000). Data analysis of interaction forces measuredwith the atomic force microscope. Ultramicroscopy, 82,85–95.
30. Merkel, R., Nassoy, P., Leung, A., Ritchie, K. & Evans,E. (1999). Energy landscapes of receptor-ligand bondsexplored with dynamic force spectroscopy. Nature,397, 50–53.
31. Evans, E. & Ritchie, K. (1999). Strength of a weak bondconnecting flexible polymer chains. Biophys. J. 76,2439–2447.
32. Strunz, T., Oroszlan, K., Schumakovitch, I.,Guntherodt, H. & Hegner, M. (2000). Model energylandscapes and the force-induced dissociation ofligand-receptor bonds. Biophys. J. 79, 1206–1212.
33. Oesterhelt, F., Oesterhelt, D., Pfeiffer, M., Engel, A.,Gaub, H. E. & Muller, D. J. (2000). Unfoldingpathways of individual bacteriorhodopsins. Science,288, 143–146.
34. Carrion-Vazquez, M., Oberhauser, A. F., Fowler, S. B.,Marszalek, P. E., Broedel, S. E., Clarke, J. & Fernandez,J. M. (1999). Mechanical and chemical unfolding of asingle protein: a comparison. Proc. Natl Acad. Sci.USA, 96, 3694–3699.
35. Schwesinger, F., Ros, R., Strunz, T., Anselmetti, D.,Guntherodt, H. J., Honegger, A. et al. (2000). Unbind-ing forces of single antibody-antigen complexescorrelate with their thermal dissociation rates. Proc.Natl Acad. Sci. USA, 97, 9972–9977.
36. Muller, D. J., Kessler, M., Oesterhelt, F., Moller, C.,Oesterhelt, D. & Gaub, H. (2002). Stability ofbacteriorhodopsin alpha-helices and loops analyzedby single-molecule force spectroscopy. Biophys. J. 83,3578–3588.
37. Gropp, F., Gropp, R. & Betlach, M. C. (1995). Effects ofupstream deletions on light- and oxygen-regulatedbacterio-opsin gene expression in Halobacterium halo-bium. Mol. Microbiol. 16, 357–364.
38. Oesterhelt, D. & Stoeckenius, W. (1974). Isolation ofthe cell membrane of Halobacterium halobium and itsfractionation into red and purple membrane. MethodsEnzymol. 31, 667–678.

606 Single Molecule Inter and Intra-molecular Forces
39. Wagner, G., Oesterhelt, D., Kripphal, G. & Lanyi, J. K.(1981). Bioenergetic role of halorhodopsin in Halobac-terium halobium cells. FEBS Letters, 131, 341–345.
40. Butt, H. J. & Jaschke, M. (1995). Calculation of thermalnoise in atomic force microscopy. Nanotechnology,6, 1–7.
Edited by W. Baumeister
(Received 13 December 2004; accepted 18 January 2005)




















