
Histamine Improves Antigen Uptake and Cross-Presentation by Dendritic Cells1
Upload
independentCategory
view
3download
0
of February 13, 2015.This information is current as
T Cells and NK Cells+CD8DAP12 Reduces Cytotoxicity of Activated Silencing Human NKG2D, DAP10, and
Verneris, Luis Soares and Robert S. NegrinMobin Karimi, Thai M. Cao, Jeanette A. Baker, Michael R.
http://www.jimmunol.org/content/175/12/7819doi: 10.4049/jimmunol.175.12.7819
2005; 175:7819-7828; ;J Immunol
Referenceshttp://www.jimmunol.org/content/175/12/7819.full#ref-list-1
, 29 of which you can access for free at: cites 56 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2005 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on February 13, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on February 13, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

Silencing Human NKG2D, DAP10, and DAP12 ReducesCytotoxicity of Activated CD8� T Cells and NK Cells1
Mobin Karimi,* Thai M. Cao,* Jeanette A. Baker,* Michael R. Verneris,‡ Luis Soares,†§ andRobert S. Negrin2*
Human CD8� T cells activated and expanded by TCR cross-linking and high-dose IL-2 acquire potent cytolytic ability againsttumors and are a promising approach for immunotherapy of malignant diseases. We have recently reported that in vitro killingby these activated cells, which share phenotypic and functional characteristics with NK cells, is mediated principally by NKG2D.NKG2D is a surface receptor that is expressed by all NK cells and transmits an activating signal via the DAP10 adaptor molecule.Using stable RNA interference induced by lentiviral transduction, we show that NKG2D is required for cytolysis of tumor cells,including autologous tumor cells from patients with ovarian cancer. We also demonstrated that NKG2D is required for in vivoantitumor activity. Furthermore, both activated and expanded CD8� T cells and NK cells use DAP10. In addition, direct killingwas partially dependent on the DAP12 signaling pathway. This requirement by activated and expanded CD8� T cells for DAP12,and hence stimulus from a putative DAP12-partnered activating surface receptor, persisted when assayed by anti-NKG2D Ab-mediated redirected cytolysis. These studies demonstrated the importance of NKG2D, DAP10, and DAP12 in human effector cellfunction. The Journal of Immunology, 2005, 175: 7819–7828.
N atural killer cells are frontline guardians in surveillanceof pathogens and tumors, and their activation is regu-lated by the integration of excitatory and inhibitory sig-
nals initiated by membrane-bound receptor molecules (1, 2). In-hibitory signals are mediated by monomeric receptors possessingintracytoplasmic immunoreceptor tyrosine-based inhibition motifsthat recruit tyrosine protein phosphatases (3). Activating signalsare generated from more diverse and complementary groups ofmembrane molecules that, upon ligand binding, can oligomerizewith different cytoplasmic adaptor polypeptides to form distinctsignaling complexes. In human NK cells, these types of activatingreceptors include NKG2D, which associate with the adaptor mol-ecule disulphide adaptor molecule (DAP)3-10, and the low-affinityIgG FcR CD16, which noncovalently couples with the adaptormolecules CD3� and FcR�. CD3�, FcR�, and another adaptor mol-ecule found in NK cells, named DAP12 (4, 5), have in common amembrane-bound structure with a cytoplasmic domain bearing oneor more ITAM motifs. Human DAP12 is known to mediate acti-vating signals from diverse binding partners that include killer cellIg-like receptors S such as killer cell Ig-like receptor 2DS2, theactivation-induced NKp44 receptor, and HLA-E binding CD94/NKG2C heterodimers (5–8). DAP12 has been shown to associatewith a host of other NK cell surface proteins that have yet to be
identified, and it is possible that these interactions may yield ad-ditional activating signals (9). CD3�, FcR�, and DAP12 intracel-lular ITAM motifs initiate protein tyrosine kinase-dependent sig-naling pathways (10), whereas DAP10 contains a YxxM activationmotif that triggers the lipid kinase cascade (11–13).
In human NK cells, DAP10 is the exclusive binding partner andsignaling intermediate for NKG2D (14). NKG2D is a potent acti-vating receptor whose ligands include proteins induced by cellularstress and malignant transformation, such as MHC class I-relatedprotein (MICA) and -B and members of the UL16-binding pro-teins, and is thought to have a particularly important role in anti-tumor immunity (15).
The expression and function of these activating signaling com-plexes are not limited to NK cells. DAP12 is expressed in granu-locytes, monocytes, as well as CD4� T cells, and DAP12-deficientmice have been shown to have defects in dendritic cells and NKcells (16–19). NKG2D has been identified on �� TCR� T cellsand human CD8� T cells, where it appears to have costimulatoryactivity and can be induced on activated mouse CD8� T cells(20–22). We have recently reported that NKG2D expression isup-regulated by human CD8� T cells activated and expanded exvivo in the presence of IFN-�, high-dose IL-2, and TCR cross-linking with an anti-CD3 mAb (23). Activated and expanded Tcells up-regulate cytotoxic effector molecules, acquire functionaland phenotypic properties that resemble NK cells, and developbroad cytotoxicity against a variety of malignant cell targets, in-cluding autologous leukemic blasts (24–28). This acquisition ofcytotoxicity, which is MHC unrestricted, coincides with the induc-tion of DAP10 expression, and the majority of killing ability isaccounted for by NKG2D signaling.
Activated and expanded CD8� T cells may have considerabletherapeutic utility as an immune-based strategy aimed at eradica-tion or suppression of malignant diseases. These T cells have broadin vitro and in vivo biological activity against inoculated tumorsafter both syngeneic and allogeneic bone marrow transplantationin rodent models (29, 30). Autologous infusion of cells of this typein patients has been associated with minimal toxicity, can reduce
*Division of Blood and Marrow Transplantation and †Division of Immunology andRheumatology, Department of Medicine, Stanford University School of Medicine,Stanford, CA 94305; ‡Division of Pediatric Blood and Marrow Diseases, Departmentof Pediatrics, University of Minnesota, Minneapolis, MN 55455; and §Institute ofBiomedical Pharmacology, Curitiba, Brazil
Received for publication May 18, 2005. Accepted for publication September21, 2005.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported in part by Grants KO8HL04505, P01CA049605, andR01CA080006 from the National Institutes of Health.2 Address correspondence and reprint requests to Dr. Robert S. Negrin, Division ofBlood and Marrow Transplantation, Stanford University, CCSR Room 2205, 269West Campus Drive, Stanford, CA 94305-5170. E-mail: [email protected]
The Journal of Immunology
Copyright © 2005 by The American Association of Immunologists, Inc. 0022-1767/05/$02.00
by guest on February 13, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

the risk of tumor recurrence in patients with hepatocellular carci-noma after surgical resection, and may induce partial remissions inpatients with relapse of lymphoma after bone marrow transplan-tation (31, 32). In this study, we extended our evaluation of thesignaling requirements for induction of cytotoxicity by activatedand expanded human CD8� T cells. We used a combination of invitro and in vivo assays against tumor cell lines and autologoustumor targets and gene targeting by RNA interference (RNAi). Weshow that both DAP10 and DAP12, in addition to NKG2D, con-tribute major activating signals in regulating the cytotoxicity ofactivated and expanded human CD8� T cells. We show, further-more, that each of these molecules, including DAP12, has a non-redundant and partially obligate signaling role in triggering tumorkilling.
Materials and MethodsmAb reagents and flow cytometry
The mAbs used in this study were the following: CD3 (UCHT1), CD8(RPA-T8), CD16 (3G8), CD56 (NCAM16-2), and isotype control (IgG)were purchased from BD Biosciences as either FITC or PE conjugates.Purified NKG2D were purchased from R&D Systems, and PE NKG2D-PEand NKG2D-allophycoyanin were purchased from Beckman Coulter. La-beled cells were analyzed on either a FACScan or an LSR-II instrument(BD Biosciences) maintained by the Stanford Shared FACS Facility (Stan-ford University). For micromagnetic cell separation, human CD8 (BW135/80) MicroBeads, human CD16 MicroBeads (130-045-701), or human HEAMicroBeads (130-061-101) were used for positive selection using an Au-toMACS device according to the vendor’s instructions (Miltenyi Biotec).Where stated, additional cell isolation by FACS sorting was performedwith a MoFlo instrument (DakoCytomation).
Isolation and generation of ex vivo expanded and activatedCD8� T cells
CD8� T cells were activated and expanded as previously described (23).Briefly, mononuclear cells were isolated from healthy donors by Ficoll-Hypaque density centrifugation, washed three times with PBS, and sepa-rated by positive selection with CD8 MicroBeads. Cells were then labeledand further purified by FACS sorting for the CD3�CD8� population. Thefinal product was resuspended at 2 � 106 cells/ml in complete RPMI 1640(cRPMI) medium supplemented with 10% FCS, 2 mM L-glutamine, 100U/ml penicillin, 100 �g/ml streptomycin, and 50 �g/ml 2-ME at 37°C in5% CO2. On day 0, cells were activated with human IFN-� (1000 U/ml;Genentech). On day 1, cells were stimulated with OKT-3 (25 ng/ml; Or-thoBioTech) and human rIL-2 (300 U/ml; Chiron). On day 4, cells weredivided into flasks with new medium and not restimulated with OKT-3.Every 3–5 days thereafter, fresh medium was added supplemented withIL-2 (300 U/ml) to maintain a cell density of 1.5–2 � 106/ml for a total of14–28 days. Bulk populations of cells have been referred to as cytokine-induced killer cells. Twenty-four to 48 h before use, cells were labeled andfurther purified by FACS sorting for the CD3�CD8� population. For someexperiments CD8� T cells were also activated and expanded from patientswith ovarian cancer according to protocols approved by the Stanford Uni-versity administration panel on human subjects.
Isolation of NK cells
Mononuclear cells from healthy donors freshly isolated with Ficoll-Hypaque were purified for NK cells by a two-step procedure. Cells werefirst positively selected with CD16 MicroBeads, followed by additionalpurification with FACS sorting for the CD3�D56� cell population. NK cellpurity was typically �98% as determined by reanalysis of an aliquot ofseparated cells. During lentiviral infection for RNAi, isolated NK cellswere maintained in DMEM/Ham’s F-12 medium (2/1 mixture) for 7 dayssupplemented with 10% human type AB serum, 600 U/ml IL-2, 100 U/mlpenicillin, 100 �g/ml streptomycin, 50 �M ME, 10 ng/ml ethanolamine, 10�g/ml ascorbate, and 50 ng/ml selenium.
NKG2D, DAP10, and DAP12 RNAi
Nucleotide sequences for short hairpin RNA (shRNA) are summarized inTable I. A previously reported RNAi sequence specific for murine CD8�(mCD8) was used as a nonspecific control (33). Oligonucleotides weredesigned that incorporated these sequences within a short hairpin structure,using the stem loop sequence 5�-TCAAGAGA-3�, which were then clonedbetween MluI and ClaI sites downstream of an H1 promoter in the plasmidpLVTHM as previously described (34). Plasmid pLVTHM, derived frompSUPERn which contains a GFP expression cassette upstream of the H1promoter (35), was a gift from D. Trono (University of Geneva, Geneva,Switzerland). For some experiments we replaced the GFP gene onpLVTHM with a DsRed reporter gene via PmeI and SpeI linkers. 293 cellswere transfected using the calcium phosphate method with 10 �g pLVThMlentivirus, 3.5 �g of vesicular stomatitis virus G plasmid, and 6.5 �g ofCMV�R8.74 according to standard protocols (36). After 16 h, the mediumwas changed, and recombinant lentivirus vectors were harvested 24–48 hlater. CD8� T cell or NK cell infection was performed three times at 24-hintervals. For each infection, cells were plated in 48-well plates at 1 � 105
cells/well and infected in the presence of protamine and hexadimethrin.Spin infection was performed at 1200 rpm for 90 min at 37°C. Four daysafter the first infection, transduced cells were isolated by FACS sorting forGFP� cells to �99% purity.
Cell lines and autologous tumor cell isolation
RPMI 1640, a human myeloma cell line cultured in cRPMI, and P815, amurine mastocytoma cell line cultured in complete DMEM, were bothpurchased from American Type Culture Collection. UCI101 (a gift fromS. Y. Liao, University of California, Irvine, CA) is a human ovarian car-cinoma cell line cultured in complete IDMEM. Luciferase expressingUCI101 (UCI101-luc) was generated according to a modified protocol thatwe have previously described (37). In brief, retrovirus generated in Phoe-nix A producer cells transfected with the pMSCVpuroBA-L2G plasmid (agift from Dr. C. Contag, Stanford University, Stanford, CA) containing afirefly luciferase insert cloned into the SalI and NotI sites, was used toinfect UCI101 cells plated in the presence of polybrene and protamine.
For preparation of tumor targets used for evaluation of autologous ac-tivated and expanded CD8� T cell cytolysis, fresh tumor was collectedfrom six ovarian cancer patients at the time of surgical resection and di-gested and homogenized to single-cell suspensions using a metal cellstrainer. The tumor cells were incubated on a shaker at 1200 rpm for 30min at 37°C with collagenase 2 (10 �g/ml) or collagenase D (10 �g/ml) incRPMI. The cells were then selected in T25 flasks and subsequently pos-itively enriched using HEA MicroBeads to �80% purity. At the time ofsurgery, PBMC were concurrently obtained from each patient and used forthe generation of autologous activated and expanded CD8� T cells as notedabove. The protocol for collection of ovarian and peripheral blood cellswas approved by the institutional review board at Stanford University.
3 Abbreviations used in this paper: DAP, disulphide adaptor molecule; MICA, MHCclass I-related protein; RNAi, RNA interference; cRPMI, complete RPMI 1640; m,murine; shRNA, short hairpin RNA; siRNA, small interfering RNA.
Table I. shRNA sequences for human NKG2D, DAP10, and DAP12RNAI a
shRNA nt Positionb Sequence (5�33�)
NKG2D-sh1 67–85 AGAGTTTGCTGGCCTGTGCNKG2D-sh2 254–272 GATCTTCCCTCTCTGAGCANKG2D-sh3 618–636 AGTTACTGTGGCCCATGTCNKG2D-sh4 846–864 GATGGCTCCATTCTCTCACNKG2D-sh5 880–898 TAATTGAAATGCAGAAGGGNKG2D-sh6 1468–1486 CGGGGTCAGGGAGGTGGTG
DAP10-sh1 332–350 GAAGATGGCAAAGTCTACADAP10-sh2 335–353 GATGGCAAAGTCTACATCADAP10-sh3 343–361 AGTCTACATCAACATGCCGDAP12-sh1 83–101 CCCTGCAGCAGGCTCCTGCDAP12-sh2 126–144 GTGGTCTCCGTCCTGTCCADAP12-sh3 310–328 ACAGCGTATCACTGAGACCDAP12-sh4 385–403 CACACAGAGGCCGTATTAC
mCD8-sh 888–906 GCTACAACTACTACATGAC
a Abbreviations: mCD8, murine CD8�; nt, nucleotide.b Nucleotide position numbered according to the following GenBank nucleotide
sequence: NKG2D, X54870; DAP10, AF172929; DAP12, AF019562; and murineCD8�, XM132621.
7820 NKG2D, DAP10, AND DAP12 IN CD8� T AND NK CELL CYTOTOXICITY
by guest on February 13, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

51Cr release cytotoxicity assay
Tumor targets were labeled with 51Cr (DuPont-NEN) by incubating 1 �106 cells in 300 �Ci (11.1 MBq) of 51Cr for at 37°C for 2 h in 5% CO2.The labeled cells were washed three times with PBS, resuspended incRPMI, and plated in 96-well plates at a concentration of 1 � 104 cells/mlin triplicate. Effector cells were added at specified ratios (10:1 or 40:1) andincubated at 37°C for 4 h in 5% CO2. At the completion of each assay,supernatant was collected and counted using a gamma counter (Cobra/AII;Packard). The percent-specific 51Cr lysis was calculated with the followingequation: percent-specific lysis � 100 � (test release) � (spontaneousrelease)/(maximal release) � (spontaneous release).
Redirected cytotoxicity assay
For redirected killing assay, 1 � 106 P815 cells were labeled with 51Cr asdescribed above. Excess 51Cr was removed by washing, and the cells wereresuspended in 500 �l of medium. Anti-NKG2D (5 �g) was added andincubated for 30 min. The P815 target cells were then used as a target inthe 51Cr release assay as described.
Cytokine production
Activated and expanded CD8� T cells at 21 days of culture were removed,washed, and incubated with 20 ng/ml PMA (Sigma-Aldrich) and 1 �Mionomycin or medium alone in a total volume of 200 �l/well in a 96-wellplate and cultured for 48 h. The supernatant was harvested and assayed forcytokine production by ELISA for IL-2 and IFN-� (eBioscience) accordingto the vendor’s instructions.
Northern blotting
RNA was prepared with the RNeasy kit (Qiagen) according to the vendor’sinstructions, and RNA blotting was performed according to standard pro-tocols (38). Three �-32P-labeled probes were constructed with the follow-ing PCR primers: NKG2D, 5�-CTGGGAGATGAGTGAATTTCATA-3�and 5�-GACTTCACCAGTTTAAGTAAATC-3� (417-bp fragment);DAP10, 5�-CATCTGGGTTCACATCCTCTT-3� and 5�-CAGAAGTCAAAGGTCCAAGC-3� (306-bp fragment); and DAP12, 5�-CCGCAAAGACCTGTACGCCA-3� and 5�-TGGACTTGGGAGAGGACTGG-3� (650-pbfragment). PCR products were cloned by TA cloning into pCR2.1 (Invitro-gen Life Technologies), digested with EcoRI, separated by 1.5% agarosegel electrophoresis, and purified with the QiaQick gel extraction kit(Qiagen).
Immunoprecipitations and Western blotting
CD8� T cells or NK cells were lysed using freshly prepared lysis buffer (10mM Tris (pH 8.0), 150 mM NaCl, 1 mM EDTA, 1% Nonidet P-40, 0.5%deoxycholate, 0.1% SDS, complete Protease Inhibitor Cocktail (Roche),and 500 �M PMSF) and clarification at 12,000 � g for 10 min at 4°C.Seventy micrograms of each cell lysate was separated on an 18% denatur-ing polyacrylamide gel and transferred to polyvinylidene difluoride mem-branes for immunoblot analysis. Membranes were blocked with 5% nonfatmilk and probed with monoclonal anti-NKG2D (M585, mouse IgG) orpolyclonal DAP10 (C-20, goat IgG; Santa Cruz Biotechnology), followedby incubation with a species-specific, secondary, HRP-conjugated Ab(Santa Cruz Biotechnology). Blots were developed using ECL reagent(Amersham Biosciences).
For immunoprecipitation, clarified lysate was labeled with 1 �g of anti-DAP12 polyclonal rat IgG Ab (FL-113; Santa Cruz Biotechnology) for 1 hat 4°C, followed by cross-linking to protein G-agarose beads (Pierce) pre-equilibrated with lysis buffer, for 1 h at 4°C on a rotator. After washing,immunoprecipitated lysate was resuspended in Laemmli buffer with 5%2-ME and boiled for 5 min before separation and immunoblotting as de-scribed above.
SCID/Hu tumor model and bioluminescent imaging
C.B-17 SCID mice, 8–12 wk old, were obtained from the colony at theStanford University Research Animal Facility and irradiated with a single2-Gy fraction using a Phillips Unit Irradiator (250 kV, 15 mA) at a doserate of 100 rad/min. After irradiation, mice were maintained on antibioticwater containing trimethoprin/sulfmethoxole. Recipient mice received 1 �106 luciferase-expressing UCI101-luc cells (1 ml/injection) via i.p. injec-tion. Intraperitoneal injection with UCI101-luc into immune-deficient miceestablishes tumors with peritoneal implants within 1 wk and eventuallymassive ascites and death (39). On days 6 and 12 after tumor inoculation,each mouse was given an injection with 40 � 106 activated and expandedCD8� T cells. Mice were evaluated every second day from the time oftumor injection for 30 days by bioluminescence imaging with the IVIS 200Imaging System (Xenogen) as previously described (37).
Statistics
Differences in the mean observations between experimental groups weredetermined with Student’s t test. Statistical significance was conferred atthe p � 0.05 level.
ResultsRNAi with NKG2D-specific shRNA suppresses cytotoxicity ofactivated and expanded CD8� T cells
We have recently demonstrated that human CD8� T cells activatedand expanded during culture in the presence of IFN-�, IL-2, andOKT3 lose cytolytic potential when NKG2D is silenced by tran-sient transfection with a small interfering RNA (siRNA) duplexoligonucleotide (23). To complement these studies and to developa model for in vivo analysis of NKG2D function, we sought toestablish stable NKG2D knockdown by RNAi in activated CD8�
T cells via transduction of shRNA with the pLVTHM lentiviralvector. NKG2D-sh6, which incorporates nucleotide sequences tar-geting a portion of the NKG2D gene transcript previously shownfunctional for silencing NKG2D by siRNA transfection, was eval-uated along with five other NKG2D-specific shRNA. NKG2D-sh6was again found to most effectively silence NKG2D gene expres-sion in CD8� T cells activated and expanded after 14–28 days inculture (Fig. 1, A and B). FACS analyses of transduced cells, iden-tified by positive GFP reporter gene expression, demonstrate thatinfection with empty pLVTHM vector does not affect NKG2Dsurface levels (Fig. 1, C and D). At the same time, NKG2D RNAiwith NKG2D-sh6 is specific insofar as it has no effect on othersurface receptors, including CD3, CD8, CD16, and CD56. Acti-vated and expanded CD8� T cells can lyse RPMI tumor targets(Fig. 1E) and retain this ability when RNAi is attempted withNKG2D-specific shRNA that do not effectively silence NKG2D.Cytolysis is similarly potent after RNAi with mCD8-sh, an shRNAspecific for murine CD8� (33), and after transduction by emptypLVTHM vector without RNAi. Cytolysis is suppressed to a levelcomparable with background cytolysis of P815 targets, however, ifNKG2D RNAi is induced by NKG2D-sh6 ( p � 0.0001). P815 isa murine mastocytoma cell line known to be resistant to killing byactivated and expanded human CD8� T cells. Thus, we confirmedour prior observations that NKG2D is required for in vitro tumorlysis by activated CD8� T cells.
NKG2D is required for cytolysis of autologous tumor byactivated and expanded CD8� T cells
We and others have shown that human plasmacytoma cell lines,such as U266, and plasmacytoma cells from patients with multiplemyeloma express MICA and ULBP3, and that cytotoxicity by ac-tivated CD8� T cells and NK cells can correlate with the expres-sion of these known NKG2D ligands (23, 40). We sought to extendthese observations by examining requirements for NKG2D signal-ing in killing of autologous tumor by CD8� T cells activated andexpanded from human patients undergoing cancer therapy. Anovel NKG2D ligand, termed Letal, has recently been found to beexpressed by ovarian carcinoma patients (41). Patients with ovar-ian cancer undergoing surgical resection of their tumor were se-lected as sources of tumor targets for in vitro cytolysis studies. APBMC sample was simultaneously obtained at the time of tumorcollection for activation and expansion of CD8� T cells. Afterexpansion, NKG2D expression in CD8� T cells from these pa-tients can be functionally silenced using NKG2D-sh6 andpLVTHM lentiviral transduction (Fig. 2, A and B). In 51Cr releaseassays, we then observed that activated CD8� T cells can effec-tively lyse labeled tumor cell targets isolated from the same patientwhen transduced without RNAi or with nonspecific mCD8-sh (Fig.2C). In contrast, cytolysis of each ovarian tumor target paired with
7821The Journal of Immunology
by guest on February 13, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

patient activated CD8� T cells was strongly suppressed whenNKG2D was silenced by NKG2D-sh6 ( p � 0.0001).
NK cells can kill tumor targets through a combination of cell-mediated and humoral cytotoxicity mechanisms. We previouslyshowed that activated human CD8� T cells require cell-cell con-tact and rely on granule release to effect cytolysis (42, 43). Weevaluated whether cytokine production may contribute to killingby activated CD8� T cells by ELISA for IL-2 and IFN-�(Fig. 2D). After 21 days of activation and expansion in culture,these cells constitutively produce IFN-�. Stimulation with PMA
and ionomycin failed to augment levels of IFN-�, and productionof these cytokines was not influenced by NKG2D silencing.Similar amounts were obtained for IL-2. These results suggest thatthe production of cytotoxic cytokines does not contribute to killingby activated and expanded CD8� T cells.
DAP10 RNAi in activated and expanded human CD8� T cellssuppresses in vitro cytolysis
NKG2D is a type II homodimeric, C-type, lectin-like transmem-brane molecule with a short cytoplasmic tail and by itself lacks
FIGURE 2. NKG2D RNAi suppresses cytolysis ofautologous ovarian tumor by activated and expandedCD8� T cells. CD8� T cells activated and expandedfrom patients were evaluated for cytolysis of autolo-gous ovarian tumors isolated at the time of surgical re-section. RNA (A) and Western blots (B) from a repre-sentative patient show that RNAi with NKG2D-sh6 bypLVTHM lentivirus effectively silences NKG2D ex-pression. C, Cytolysis of 51Cr-labeled ovarian tumortargets by paired patient activated and expanded CD8�
T cells (E:T cell ratio, 10:1 or 40:1) is significantlyreduced by NKG2D RNAi (p � 0.0001) compared withnonspecific RNAi and mock pLVTHM lentiviral trans-duction. Shown are representative results from one ex-periment repeated four times with other ovarian cancerautologous T cells paired samples. ELISA for IFN-�(D) by activated CD8� T cells showing that productionof this cytokine is not influenced by NKG2D silencingand does not significantly increase with stimulation byPMA and ionomycin.
FIGURE 1. NKG2D RNAi with shRNA by lentiviraltransduction in activated and expanded human CD8� Tcells. Human CD8� T cells activated and expanded after14–28 days of culture in the presence of IFN-�, IL-2, andOKT3 were infected with pLVTHM lentivirus expressingone of six shRNA-targeting NKG2D. As shown by RNA(A) and Western blotting (B) of transduced cells, NKG2Dsilencing is most effective with NKG2D-sh6, which tar-gets a sequence (nt 1468–1486) in the 3� untranslatedregion of the NKG2D transcript. C and D, FACS analysisof transduced cells. NKG2D RNAi with NKG2D-sh6 in-duces complete silencing of NKG2D while preservingsurface expression of CD3, CD8, CD16, and CD56. E,Cytolysis of 51Cr-labeled RPMI human plasmacytomacell line by activated and expanded CD8� T cells is sup-pressed �90% by NKG2D RNAi with NKG2D-sh6 (p �0.0001), whereas RNAi with nonspecific mCD8 shRNA(mCD8-sh) and transduction with empty pLVTHM (NoRNAi) had no effect. Cytolysis of murine P815 mastocy-toma was not observed. Shown are representative resultsfrom one experiment repeated three times.
7822 NKG2D, DAP10, AND DAP12 IN CD8� T AND NK CELL CYTOTOXICITY
by guest on February 13, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

intrinsic signaling capabilities (44). Instead, NKG2D forms a sta-ble complex with the predominantly intracellular DAP10 adaptorprotein to deliver activating signals via recruitment of the p85subunit of the PI3K. We therefore used RNAi to evaluate the roleof DAP10 signaling in activated and expanded CD8� T cells. Wefirst determined that DAP10 is expressed by activated T cells andthe CD8� T cell subset (Fig. 3A). We then generated three DAP10-specific shRNA and found that DAP10-sh3, which targets exon 4of the DAP10 mRNA near the translation stop codon, most effec-tively silenced DAP10 gene expression when transduced with len-tivirus (Fig. 3, B and C). Activated CD8� T cells subjected toDAP10 RNAi using the DAP10-sh3 lost 75% of cytolytic ac-tivity ( p � 0.0003) compared with transduction using ineffectiveDAP10 shRNA, nonspecific mCD8-sh, and empty pLVTHMvector.
DAP12 silencing also suppresses cytolysis by activated andexpanded CD8� T cells
DAP10 gene knockdown suppressed cytolysis by activated andexpanded CD8� T cells less potently than did silencing ofNGK2D, suggesting that additional signaling pathways contributeto maximum cytolysis triggered by NKG2D-mediated activation.No association between NKG2D and DAP12, another intracyto-plasmic activating adaptor protein, has been observed in humans(9, 14). DAP12 is a promiscuous binding partner, however, andthus may contribute signals important for cytolysis in activatedCD8� T cells through pairing with other activating surface recep-
tors. Similar to DAP10, we found levels of DAP12 expression inactivated and expanded CD8� T cells comparable to those ob-served in freshly isolated NK cells (Fig. 4A). Of four DAP12-specific shRNA evaluated, we determined that DAP12-sh3 in-duced the most suppression of DAP12 gene expression in activatedCD8� T cells (Fig. 4B). The DAP12-sh3 nucleotide sequence iscomplementary to an entirely translated portion of exon 4 of theDAP12-coding sequence. In accordance with our speculation, weobserved a partial and significant reduction in cytotoxicity by ac-tivated and expanded CD8� T cells when DAP12 was silenced byRNAi using DAP12-sh3 ( p � 0.002; Fig. 4C). Although less pro-found than requirements for NKG2D- and DAP10-mediated sig-naling, these results suggest that activated human CD8� T cellsalso rely on DAP12-dependent signaling pathways for optimal tu-mor cytolysis.
Double-knockdown of DAP10 and DAP12 in activated CD8� Tcells
The proximal signaling pathway for DAP10 includes Grb-2, phos-pholipase C-2, SLP-76, and PI3K (45). For DAP12, initial signal-ing involves activation of Src protein kinases, which phosphorylatethe ITAM on tandem tyrosine residues, followed by second-linerecruitment of Syk family protein kinases. It is thought that thesedisparate signals then culminate in a common pathway leading toactivation of Rac, MAPK kinase, and ERK and, ultimately, cytol-ysis. To gain insight into whether the integration of DAP10 andDAP12 signaling contributes to induction of optimal tumor killing,we generated activated CD8� T cells silenced for both DAP10 and
FIGURE 3. DAP10 RNAi in activated and expanded CD8� T cells. A,RNA blot showing that DAP10 is constitutively expressed by freshly iso-lated human NK cells and all activated and expanded T cells including theCD8� subset. By RNA (B) and Western blotting (C), DAP10 silencing bythree DAP10 shRNA in activated CD8� T cells by pLVTHM lentiviruswas most effective with DAP10-sh3. DAP10-sh3 (nt 343–361) targets exon4 of the DAP10 transcript, nine nucleotides upstream of the translation stopcodon. D, Cytolysis of 51Cr-labeled RPMI 1640 targets by activated andexpanded CD8� T cells is suppressed by 75% (p � 0.003) after DAP10RNAi with DAP10-sh3. Shown are representative results from one exper-iment repeated three times.
FIGURE 4. DAP12 RNAi in activated and expanded CD8� T cells. A,RNA blot showing that DAP12 is expressed by activated and expandedhuman T cells at a level comparable to that in freshly isolated NK cells. B,RNA blot; DAP12 silencing using four DAP12 shRNA in activated CD8� Tcells by pLVTHM lentivirus was functionally achieved with DAP12-sh3.DAP12-sh3 (nt 310–328) targets exon 4, an entirely translated portion of theDAP12 coding sequence. C, Cytolysis of RPMI 1640 targets in a 51Cr releaseassay by activated and expanded CD8� T cells is partially suppressed byDAP12 RNAi with DAP12-sh3 (p � 0.002) and to a lesser extent by DAP12-sh4 at the higher E:T cell ratio of 40:1 compared with negative mCD8-sh andempty pLVTHM vector controls. Shown are representative results from oneexperiment repeated three times.
7823The Journal of Immunology
by guest on February 13, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

DAP12. To isolate double transductants for in vitro cytolysis as-says, we replaced the GFP reporter gene on the pLVTHM lenti-viral vector with DsRed for transduction of DAP10-sh3. Thus,DAP10-sh3-transduced cells can be FACS sorted based on DsRedfluorescence, as opposed to GFP fluorescence for successfulDAP12-sh3 transduction, and fluorescence of both DsRed andGFP in DAP10-sh3 and DAP12-sh3 double transductants (Fig. 5,A–C). NKG2D expression was preserved even with simultaneoussilencing of DAP10 and DAP12 after activation in 14–28 days ofculture, as determined by FACS. Suppression of in vitro cytolysis
by DAP10 and DAP12 double knockdown was significant, with a�80% reduction ( p � 0.0001) in cytotoxicity compared with neg-ative controls (Fig. 5D), reflecting more potent suppression thansilencing of DAP10 or DAP12 alone.
Human NKG2D has been shown to assemble as a hexamericcomplex consisting of the NKG2D homodimer interacting withfour DAP10 molecules via a three-helix interface in the transmem-brane domains (46). In DAP10-deficient mice, NKG2D surfaceassembly and signaling can be delivered through noncovalentbinding with DAP12 via expression of an alternative splice variant
FIGURE 5. Double transduction with DAP10-sh3 andDAP12-sh3 for RNAi. A, The GFP cassette in pLVTHMwas replaced by a DsRed reporter gene for transductionof DAP10-sh3 to confirm by double DAP10 and DAP12knockdown FACS analysis in activated T cells as shownby GFP and DsRed fluorescence, also indicating no re-duction in NKG2D expression. B, RNAi of DAP10 andDAP12 is specifically mediated by, respectively, DAP10-sh3 and DAP12-sh3, as shown in this RNA blot of acti-vated and expanded CD8� T cells. C, Western blot forDAP10 and DAP12 after transduction for DAP10-sh3and DAP12-sh3. D, Suppression of 51Cr-labeled RPMI1640 cytolysis by activated and expanded T cells in thepresence of simultaneous DAP10 and DAP12 RNAi (p �0.001) compared with mCD8-sh and empty pLVTHMvector negative controls as shown. Shown are represen-tative results from one experiment repeated three times.
FIGURE 6. NKG2D pairs with DAP12 in activatedCD8� T cells. Western blot of whole cell lysates fromactivated and expanded CD8� T cells (A) or purifiedNK cells (B) confirming silencing of DAP10 byDAP10-sh3, whereas NKG2D protein expression ispreserved. C, Receptor complexes were immunopre-cipitated with a polyclonal anti-DAP12 Ab and blottedfor NKG2D, showing pairing of DAP12 with NKG2Dregardless of DAP10 silencing in activated and ex-panded CD8� T cells but not in purified NK cells (D).E, Binding of NKG2D and DAP12 in activated CD8�
T cells was confirmed by immunoprecipitation with anti-NKG2D, followed by blotting for DAP12.
7824 NKG2D, DAP10, AND DAP12 IN CD8� T AND NK CELL CYTOTOXICITY
by guest on February 13, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

(22). Because no human NKG2D splice variant has been identifiedto date, we were surprised to find that NKG2D surface expressionis maintained in activated CD8� T cells despite DAP10 silencing.It is possible that human NKG2D isoforms may exist at low levelsor only in certain conditions or cell types, however, and thus weexamined whether NKG2D can interact with DAP12 in activatedand expanded CD8� T cells. We first confirmed by Western blot-ting that DAP10-sh3 effectively silences DAP10 expression inboth CD8� T cells and NK cells (Fig. 6, A and B). Activated CD8�
T cells differ from NK cells, however, in that we observed thatNKG2D can be coimmunoprecipitated with DAP12 regardless ofwhether DAP10 is silenced, which is lost when DAP12 RNAi isinduced (Fig. 6, C–E). These results indicate that human NKG2Dexpression can be accounted for by pairing with DAP12 whenDAP10 is not present in activated and expanded CD8� T cells.
NKG2D, DAP10, and DAP12 RNAi in human NK cells
Using the NKG2D-sh6, DAP10-sh3, and DAP12-sh3 constructs,we examined the roles of these signaling molecules in primary NKcells isolated from normal human donors. These three shRNA andthe pLVTHM lentivirus were able to elicit high level functionalsilencing of NKG2D, DAP10, and DAP12 gene expression in NKcells (Fig. 7, A–C). NK cell killing of RPMI 1640 target cells wasnot compromised by lentiviral transduction, but was potently sup-pressed by NKG2D silencing, which was not seen with RNAi bycontrol shRNA (Fig. 7D). This finding is in agreement with ob-servations by others who have established the general dependencyby NK cells on NKG2D for in vitro killing of NKG2D ligand-bearing tumor targets (15). Because NKG2D-activating signals ap-pear to be exclusively transduced by DAP10 in human NK cells(9), we were not surprised to find a similar degree of NK cellcytolysis suppression with DAP10 gene silencing (Fig. 7E). Un-expectedly, however, we observed that DAP12 RNAi also sub-stantially suppressed NK cell killing compared with control lenti-viral transductions.
NKG2D, DAP10, and DAP12 RNAi in redirected cytolysis ofNK cells and activated CD8� T cells
Elegant studies conducted by Rosen et al. (14) provided compel-ling evidence suggesting that DAP10 is entirely sufficient, and,hence, DAP12 dispensable, for NKG2D function in human NKcells insofar as direct killing of MICA-transduced BaF/3 cell tar-gets. The surface density of NKG2D ligands where natively ex-pressed by tumor cell lines can be at low levels, however, whichmay account for conflicting observations of only partial depen-
dency on NKG2D for NK tumor cytolysis in these different ex-perimental circumstances (23, 47). We tested whether a strongNKG2D stimulus, serving as the dominant activating signal, canovercome suppression of killing due to DAP12 knockdown byperforming redirected cytolysis of Fc�R-bearing P815 cell targetspreincubated with an agonistic anti-NKG2D mAb. Vigorous Ab-mediated redirected killing was observed by freshly isolated hu-man NK cells when transduced with empty pLVTHM vector orwith control mCD8-sh RNAi, but not after transduction withNKG2D-sh6 or DAP10-sh3 (Fig. 8A). In contrast to cytolysis ofRPMI targets, we observed little suppression of NK cell-redirectedkilling with DAP12-sh3-mediated RNAi. Suppression of redi-rected cytolysis by activated and expanded CD8� T cell persisted,however, when DAP12 was silenced by RNAi (Fig. 8B).
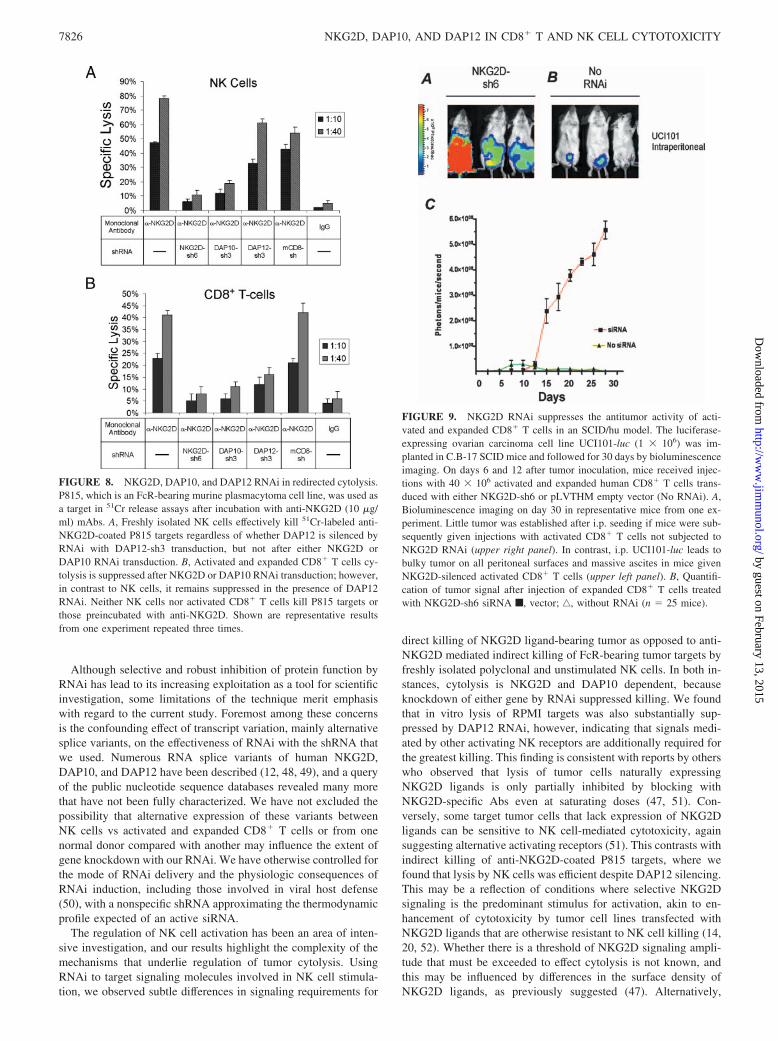
NKG2D RNAi in an SCID/Hu tumor model
Using NKG2D-sh6 and the lentiviral vector to induce stable genesilencing, we evaluated the role of NKG2D signaling in an in vivotumor model. We generated a luciferase-expressing human ovariancarcinoma cell line, UCI101-luc, and inoculated these cells inC.B-17 SCID mice, followed by i.v. injections of activated andexpanded human CD8� T cells on days 6 and 12 after tumor seed-ing. Tracking tumor growth by bioluminescence imaging, we ob-served that mice treated with expanded and activated CD8� T cellstransduced with NKG2D-sh6 had markedly faster tumor progres-sion than animals treated with CD8� T cells not containing theNKG2D-sh6 siRNA (Fig. 9). These results indicate that suppres-sion of NKG2D expression significantly impacts the in vivo bio-logical capability of expanded CD8� T cells in controlling tumorgrowth and progression.
DiscussionIn this report we dissect the molecular signaling events leading tocytolysis of tumor cells by activated and expanded human CD8�
T cells and NK cells. We developed shRNA specific for humanNKG2D, DAP10, and DAP12 for lentiviral transduction that wereeffective for stable gene expression silencing. Using these shRNAin RNAi experiments, we confirmed our previous observation thatactivated and expanded human CD8� T cells depend on NKG2Dfor in vitro killing and demonstrated that NKG2D is also requiredfor in vivo antitumor activity. This dependency on NKG2D andconsequently DAP10 signaling, although obligate, is not sufficientfor optimal cytolysis, because partial suppression of killing solelyby DAP12 RNAi suggests a nonredundant role for activation ini-tiated by the DAP12 pathway.
FIGURE 7. NKG2D, DAP10, and DAP12 RNAi inhuman NK cells. CD3�CD56� NK cells freshly iso-lated from normal human donors were evaluated forcytolysis after RNAi for NKG2D, DAP10, and DAP12.RNA (A) and Western blots (B) demonstrate that func-tional silencing of NKG2D can be achieved in freshpolyclonal human NK cells using NKG2D-sh6 andpLVTHM lentiviral vector. Similar DAP10 and DAP12RNAi was observed by RNA blot (C) after transductionwith DAP10-sh3 and DAP12-sh3. Cytolysis of 51Cr-labeled RPMI 1640 targets by fresh NK cells is sup-pressed with NKG2D RNAi (D; p � 0.002) and to asimilar extent with either DAP10 (p � 0.005) orDAP12 (p � 0.009) RNAi (E). Shown are representa-tive results from one experiment repeated three times.
7825The Journal of Immunology
by guest on February 13, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
Although selective and robust inhibition of protein function byRNAi has lead to its increasing exploitation as a tool for scientificinvestigation, some limitations of the technique merit emphasiswith regard to the current study. Foremost among these concernsis the confounding effect of transcript variation, mainly alternativesplice variants, on the effectiveness of RNAi with the shRNA thatwe used. Numerous RNA splice variants of human NKG2D,DAP10, and DAP12 have been described (12, 48, 49), and a queryof the public nucleotide sequence databases revealed many morethat have not been fully characterized. We have not excluded thepossibility that alternative expression of these variants betweenNK cells vs activated and expanded CD8� T cells or from onenormal donor compared with another may influence the extent ofgene knockdown with our RNAi. We have otherwise controlled forthe mode of RNAi delivery and the physiologic consequences ofRNAi induction, including those involved in viral host defense(50), with a nonspecific shRNA approximating the thermodynamicprofile expected of an active siRNA.
The regulation of NK cell activation has been an area of inten-sive investigation, and our results highlight the complexity of themechanisms that underlie regulation of tumor cytolysis. UsingRNAi to target signaling molecules involved in NK cell stimula-tion, we observed subtle differences in signaling requirements for
direct killing of NKG2D ligand-bearing tumor as opposed to anti-NKG2D mediated indirect killing of FcR-bearing tumor targets byfreshly isolated polyclonal and unstimulated NK cells. In both in-stances, cytolysis is NKG2D and DAP10 dependent, becauseknockdown of either gene by RNAi suppressed killing. We foundthat in vitro lysis of RPMI targets was also substantially sup-pressed by DAP12 RNAi, however, indicating that signals medi-ated by other activating NK receptors are additionally required forthe greatest killing. This finding is consistent with reports by otherswho observed that lysis of tumor cells naturally expressingNKG2D ligands is only partially inhibited by blocking withNKG2D-specific Abs even at saturating doses (47, 51). Con-versely, some target tumor cells that lack expression of NKG2Dligands can be sensitive to NK cell-mediated cytotoxicity, againsuggesting alternative activating receptors (51). This contrasts withindirect killing of anti-NKG2D-coated P815 targets, where wefound that lysis by NK cells was efficient despite DAP12 silencing.This may be a reflection of conditions where selective NKG2Dsignaling is the predominant stimulus for activation, akin to en-hancement of cytotoxicity by tumor cell lines transfected withNKG2D ligands that are otherwise resistant to NK cell killing (14,20, 52). Whether there is a threshold of NKG2D signaling ampli-tude that must be exceeded to effect cytolysis is not known, andthis may be influenced by differences in the surface density ofNKG2D ligands, as previously suggested (47). Alternatively,
FIGURE 8. NKG2D, DAP10, and DAP12 RNAi in redirected cytolysis.P815, which is an FcR-bearing murine plasmacytoma cell line, was used asa target in 51Cr release assays after incubation with anti-NKG2D (10 �g/ml) mAbs. A, Freshly isolated NK cells effectively kill 51Cr-labeled anti-NKG2D-coated P815 targets regardless of whether DAP12 is silenced byRNAi with DAP12-sh3 transduction, but not after either NKG2D orDAP10 RNAi transduction. B, Activated and expanded CD8� T cells cy-tolysis is suppressed after NKG2D or DAP10 RNAi transduction; however,in contrast to NK cells, it remains suppressed in the presence of DAP12RNAi. Neither NK cells nor activated CD8� T cells kill P815 targets orthose preincubated with anti-NKG2D. Shown are representative resultsfrom one experiment repeated three times.
FIGURE 9. NKG2D RNAi suppresses the antitumor activity of acti-vated and expanded CD8� T cells in an SCID/hu model. The luciferase-expressing ovarian carcinoma cell line UCI101-luc (1 � 106) was im-planted in C.B-17 SCID mice and followed for 30 days by bioluminescenceimaging. On days 6 and 12 after tumor inoculation, mice received injec-tions with 40 � 106 activated and expanded human CD8� T cells trans-duced with either NKG2D-sh6 or pLVTHM empty vector (No RNAi). A,Bioluminescence imaging on day 30 in representative mice from one ex-periment. Little tumor was established after i.p. seeding if mice were sub-sequently given injections with activated CD8� T cells not subjected toNKG2D RNAi (upper right panel). In contrast, i.p. UCI101-luc leads tobulky tumor on all peritoneal surfaces and massive ascites in mice givenNKG2D-silenced activated CD8� T cells (upper left panel). B, Quantifi-cation of tumor signal after injection of expanded CD8� T cells treatedwith NKG2D-sh6 siRNA f, vector; ‚, without RNAi (n � 25 mice).
7826 NKG2D, DAP10, AND DAP12 IN CD8� T AND NK CELL CYTOTOXICITY
by guest on February 13, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

NKG2D binds with its various ligands at markedly different bind-ing affinities (44); thus, heterogeneous tumor cell expression ofNKG2D ligands may influence signaling.
We unexpectedly observed that NKG2D expression is main-tained in activated and expanded CD8� T cells despite DAP10silencing and showed that binding with DAP12 may account forthis phenomenon. The receptor specificity of DAP10 and DAP12resides entirely in their transmembrane domains (9, 14, 46). It ispossible that there are structural variants in the transmembranedomain of either NKG2D or the adaptor proteins permissive forpairing of NKG2D with DAP12 in activated CD8� T cells, al-though evidence supporting this has not been found in studies ofpolyclonal NK cells and IL-2-activated blood mononuclear cells(53). This consideration is further complicated by our observationthat NKG2D is expressed by activated CD8� T cells even whenboth DAP10 and DAP12 are simultaneously silenced. Similaritiescan be drawn to observations made in Jurkat cell lines, which donot express DAP12 and express DAP10 only at low levels (5, 12),whereby NKG2D expression can be induced by transfection withan NKG2D expression vector alone (20). Although binding ofNKG2D with adaptor proteins other than DAP10 and DAP12 hasnot been reported, we can speculate on novel associations betweenNKG2D and yet to be identified receptor complex subunits. Dou-ble knockdown of DAP10 and DAP12 abolishes much of the kill-ing by activated and expanded CD8� T cells, however, implyingthat these novel NKG2D receptor complexes are unlikely to beimportant in signaling for activation of cytolysis.
Unlike NK cells, activated and expanded CD8� T cells requiresimultaneous stimulation mediated by DAP10 as well as DAP12pathways for all aspects of in vitro killing evaluated in this study.From previous studies we had characterized cytolysis by activatedand expanded T cells as being MHC unrestricted and TCR inde-pendent, requiring cell-cell contact, and relying on an intact gran-ule release pathway; more recently, we identified NKG2D as thecritical activating intermediary (23, 29, 42, 43). Our finding thatkilling by activated and expanded CD8� T cells is sensitive toDAP10 silencing by RNAi is predictable given the exquisite rela-tionship between cytolysis and Ab-mediated NKG2D blockadethat we had observed from previous studies. That cytolysis canalso be inhibited by DAP12 RNAi was unexpected and raisesquestions about whether there are key DAP12-binding receptorpartners expressed by activated and expanded CD8� T cells in-fluencing cytolysis. Of clinical relevance, DAP12 loss-of-functionmutations have been identified (49, 54), and it would be importantto understand whether CD8� T cells activated and expanded fromsuch individuals would be impaired with regard to tumor killing.
Many emerging lines of evidence now point toward remarkablyconvergent evolution between NK cells and T cells, and our resultsin this study lend additional support to this idea. Although NKcells used a strategy of balancing selection to diversify its reper-toire of recognition receptors as opposed to T cells, which acquireddiversity through germline recombination of the TCR, both centeron interaction with ligands bearing the general structure of MHCmolecules for activation (1, 55). NK cells constitutively expressgranzyme, perforin, and IFN-�, whereas cytotoxic T cells mustundergo proliferation and de novo transcription of these gene prod-ucts to become competent effector cells, yet both ultimately use thesame defense mechanisms to defend against invading pathogens ortumor (56). It is interesting to speculate about whether physiologicconditions mimicking our in vivo activation and expansion proto-col exist and how these could be beneficial for effective immunefunction, or why a T cell would maintain, but keep dormant, path-ways for activation of cytolysis shared with NK cells and, more-over, use it preferentially over its native TCR. Our results in this
study clearly show that activated and expanded CD8� T cells relyon NKG2D and DAP10 signaling to acquire cytolysis of tumortargets, but we have yet to understand the role of DAP12 in theactivation cascade. We have also not clarified the distinction be-tween cytolysis as opposed to proliferation and cytokine secretionand the dependency on each of these proximal signaling pathways,although our results suggest that cytokine production does not con-tribute to killing in activated CD8� T cells. We have observed thatactivated and expanded murine CD8� T cells efficiently eradicatetumor, but do not cause graft-vs-host disease (30), and it is pos-sible that subtle differences in these activation pathways, comparedwith those used by unmanipulated alloreactive T cells, may becrucial for avoidance of deleterious immune effects.
Ultimately, we aim to test the antitumor activity of activated andexpanded CD8� T cells in rationally designed clinical studies in-volving patients with malignant diseases. We provide additionalevidence that CD8� T cells activated and expanded from normaldonors have in vivo tumor activity using an SCID/hu model. Wefurthermore show that these activated cells can be generated frompatients with ovarian cancer, which gain the ability to lyse autol-ogous tumor targets isolated from the same patient. Findings fromthis study provide us with a better understanding of the broadantitumor potential of these cells, enhance our understanding ofregulatory mechanisms supporting cytolysis, and will aid in theimplementation of activated and expanded CD8� T cells as a po-tentially efficacious form of cellular immunotherapy.
AcknowledgmentsWe thank Dr. John Chan for assistance with acquisition of PBMC andtumor samples from ovarian cancer patients, and the patients themselvesfor participating in our study. We also thank Dr. Tim Doyle for providingus with the pMSCVpuroBA-L2G retroviral vector, and Dr. Christopher H.Contag for his thoughtful advice. Many thanks to Dr. Leon Su for help withvector cloning and discussion of experimental results. We thank Dr. RubyWong for assistance with statistical analyses.
DisclosuresThe authors have no financial conflict of interest.
References1. Lanier, L. L. 2005. NK cell recognition. Annu. Rev. Immunol. 23: 225–274.2. Yokoyama, W. M., S. Kim, and A. R. French. 2004. The dynamic life of natural
killer cells. Annu. Rev. Immunol. 22: 405–429.3. Vely, F., and E. Vivier. 1997. Conservation of structural features reveals the
existence of a large family of inhibitory cell surface receptors and noninhibitory/activatory counterparts. J. Immunol. 159: 2075–2077.
4. Wirthmueller, U., T. Kurosaki, M. S. Murakami, and J. V. Ravetch. 1992. Signaltransduction by Fc�RIII (CD16) is mediated through the � chain. J. Exp. Med.175: 1381–1390.
5. Lanier, L. L., B. C. Corliss, J. Wu, C. Leong, and J. H. Phillips. 1998. Immu-noreceptor DAP12 bearing a tyrosine-based activation motif is involved in acti-vating NK cells. Nature 391: 703–707.
6. Lanier, L. L., B. Corliss, J. Wu, and J. H. Phillips. 1998. Association of DAP12with activating CD94/NKG2C NK cell receptors. Immunity 8: 693–701.
7. Vitale, M., C. Bottino, S. Sivori, L. Sanseverino, R. Castriconi, E. Marcenaro,R. Augugliaro, L. Moretta, and A. Moretta. 1998. NKp44, a novel triggeringsurface molecule specifically expressed by activated natural killer cells, is in-volved in non-major histocompatibility complex-restricted tumor cell lysis.J. Exp. Med. 187: 2065–2072.
8. Cantoni, C., C. Bottino, M. Vitale, A. Pessino, R. Augugliaro, A. Malaspina,S. Parolini, L. Moretta, A. Moretta, and R. Biassoni. 1999. NKp44, a triggeringreceptor involved in tumor cell lysis by activated human natural killer cells, is anovel member of the immunoglobulin superfamily. J. Exp. Med. 189: 787–796.
9. Wu, J., H. Cherwinski, T. Spies, J. H. Phillips, and L. L. Lanier. 2000. DAP10and DAP12 form distinct, but functionally cooperative, receptor complexes innatural killer cells. J. Exp. Med. 192: 1059–1068.
10. McVicar, D. W., L. S. Taylor, P. Gosselin, J. Willette-Brown, A. I. Mikhael,R. L. Geahlen, M. C. Nakamura, P. Linnemeyer, W. E. Seaman, S. K. Anderson,et al. 1998. DAP12-mediated signal transduction in natural killer cells: a domi-nant role for the Syk protein-tyrosine kinase. J. Biol. Chem. 273: 32934–32942.
11. Chang, C., J. Dietrich, A. G. Harpur, J. A. Lindquist, A. Haude, Y. W. Loke,A. King, M. Colonna, J. Trowsdale, and M. J. Wilson. 1999. Cutting edge:KAP10, a novel transmembrane adapter protein genetically linked to DAP12 butwith unique signaling properties. J. Immunol. 163: 4651–4654.
7827The Journal of Immunology
by guest on February 13, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

12. Wu, J., Y. Song, A. B. Bakker, S. Bauer, T. Spies, L. L. Lanier, and J. H. Phillips.1999. An activating immunoreceptor complex formed by NKG2D and DAP10.Science 285: 730–732.
13. Billadeau, D. D., J. L. Upshaw, R. A. Schoon, C. J. Dick, and P. J. Leibson. 2003.NKG2D-DAP10 triggers human NK cell-mediated killing via a Syk-independentregulatory pathway. Nat. Immunol. 4: 557–564.
14. Rosen, D. B., M. Araki, J. A. Hamerman, T. Chen, T. Yamamura, andL. L. Lanier. 2004. A structural basis for the association of DAP12 with mouse,but not human, NKG2D. J. Immunol. 173: 2470–2478.
15. Raulet, D. H. 2003. Roles of the NKG2D immunoreceptor and its ligands. Nat.Rev. Immunol. 3: 781–790.
16. Bakker, A. B., E. Baker, G. R. Sutherland, J. H. Phillips, and L. L. Lanier. 1999.Myeloid DAP12-associating lectin (MDL)-1 is a cell surface receptor involved inthe activation of myeloid cells. Proc. Natl. Acad. Sci. USA 96: 9792–9796.
17. Aoki, N., S. Kimura, Y. Takiyama, Y. Atsuta, A. Abe, K. Sato, and M. Katagiri.2000. The role of the DAP12 signal in mouse myeloid differentiation. J. Immunol.165: 3790–3796.
18. Tomasello, E., P. O. Desmoulins, K. Chemin, S. Guia, H. Cremer, J. Ortaldo,P. Love, D. Kaiserlian, and E. Vivier. 2000. Combined natural killer cell anddendritic cell functional deficiency in KARAP/DAP12 loss-of-function mutantmice. Immunity 13: 355–364.
19. Snyder, M. R., M. Lucas, E. Vivier, C. M. Weyand, and J. J. Goronzy. 2003.Selective activation of the c-Jun NH2-terminal protein kinase signaling pathwayby stimulatory KIR in the absence of KARAP/DAP12 in CD4� T cells. J. Exp.Med. 197: 437–449.
20. Bauer, S., V. Groh, J. Wu, A. Steinle, J. H. Phillips, L. L. Lanier, and T. Spies.1999. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science 285: 727–729.
21. Groh, V., R. Rhinehart, J. Randolph-Habecker, M. S. Topp, S. R. Riddell, andT. Spies. 2001. Costimulation of CD8�� T cells by NKG2D via engagement byMIC induced on virus-infected cells. Nat. Immunol. 2: 255–260.
22. Diefenbach, A., E. Tomasello, M. Lucas, A. M. Jamieson, J. K. Hsia, E. Vivier,and D. H. Raulet. 2002. Selective associations with signaling proteins determinestimulatory versus costimulatory activity of NKG2D. Nat. Immunol. 3:1142–1149.
23. Verneris, M. R., M. Karami, J. Baker, A. Jayaswal, and R. S. Negrin. 2004. Roleof NKG2D signaling in the cytotoxicity of activated and expanded CD8� T cells.Blood 103: 3065–3072.
24. Brooks, C. G., D. L. Urdal, and C. S. Henney. 1983. Lymphokine-driven “dif-ferentiation” of cytotoxic T-cell clones into cells with NK-like specificity: cor-relations with display of membrane macromolecules. Immunol. Rev. 72: 43–72.
25. Schmidt-Wolf, I. G., R. S. Negrin, H. P. Kiem, K. G. Blume, and I. L. Weissman.1991. Use of a SCID mouse/human lymphoma model to evaluate cytokine-in-duced killer cells with potent antitumor cell activity. J. Exp. Med. 174: 139–149.
26. Lu, P. H., and R. S. Negrin. 1994. A novel population of expanded humanCD3�CD56� cells derived from T cells with potent in vivo antitumor activity inmice with severe combined immunodeficiency. J. Immunol. 153: 1687–1696.
27. Hoyle, C., C. D. Bangs, P. Chang, O. Kamel, B. Mehta, and R. S. Negrin. 1998.Expansion of Philadelphia chromosome-negative CD3�CD56� cytotoxic cellsfrom chronic myeloid leukemia patients: in vitro and in vivo efficacy in severecombined immunodeficiency disease mice. Blood 92: 3318–3327.
28. Alvarnas, J. C., Y. C. Linn, E. G. Hope, and R. S. Negrin. 2001. Expansion ofcytotoxic CD3�CD56� cells from peripheral blood progenitor cells of patientsundergoing autologous hematopoietic cell transplantation. Biol. Blood MarrowTransplant. 7: 216–222.
29. Verneris, M. R., M. Ito, J. Baker, A. Arshi, R. S. Negrin, and J. A. Shizuru. 2001.Engineering hematopoietic grafts: purified allogeneic hematopoietic stem cellsplus expanded CD8� NK-T cells in the treatment of lymphoma. Biol. BloodMarrow Transplant. 7: 532–542.
30. Baker, J., M. R. Verneris, M. Ito, J. A. Shizuru, and R. S. Negrin. 2001. Expan-sion of cytolytic CD8� natural killer T cells with limited capacity for graft-versus-host disease induction due to interferon � production. Blood 97:2923–2931.
31. Takayama, T., T. Sekine, M. Makuuchi, S. Yamasaki, T. Kosuge, J. Yamamoto,K. Shimada, M. Sakamoto, S. Hirohashi, Y. Ohashi, et al. 2000. Adoptive im-munotherapy to lower postsurgical recurrence rates of hepatocellular carcinoma:a randomised trial. Lancet 356: 802–807.
32. Leemhuis, T., S. Wells, C. Scheffold, M. Edinger, and R. S. Negrin. 2005. Aphase I trial of autologous cytokine-induced killer cells for the treatment of re-lapsed Hodgkin disease and non-Hodgkin lymphoma. Biol. Blood Marrow Trans-plant. 11: 181–187.
33. Rubinson, D. A., C. P. Dillon, A. V. Kwiatkowski, C. Sievers, L. Yang,J. Kopinja, D. L. Rooney, M. M. Ihrig, M. T. McManus, F. B. Gertler, et al. 2003.A lentivirus-based system to functionally silence genes in primary mammaliancells, stem cells and transgenic mice by RNA interference. Nat. Genet. 33:401–406.
34. Wiznerowicz, M., and D. Trono. 2003. Conditional suppression of cellular genes:lentivirus vector-mediated drug-inducible RNA interference. J. Virol. 77:8957–8961.
35. Brummelkamp, T. R., R. Bernards, and R. Agami. 2002. A system for stableexpression of short interfering RNAs in mammalian cells. Science 296: 550–553.
36. Zufferey, R., D. Nagy, R. J. Mandel, L. Naldini, and D. Trono. 1997. Multiplyattenuated lentiviral vector achieves efficient gene delivery in vivo. Nat. Biotech-nol. 15: 871–875.
37. Edinger, M., Y. A. Cao, M. R. Verneris, M. H. Bachmann, C. H. Contag, andR. S. Negrin. 2003. Revealing lymphoma growth and the efficacy of immune celltherapies using in vivo bioluminescence imaging. Blood 101: 640–648.
38. Brown, T. 2004. Analysis of RNA by Northern and slot-blot hybridization. InCurrent Protocols in Immunology, Vol. 3. J. E. Coligan, B. E. Bierer,D. H. Margulies, E. M. Shevach, and W. Strober, eds. Wiley & Sons, Hobokin,pp. 10.12.11–10.12.14.
39. Fuchtner, C., D. A. Emma, A. Manetta, G. Gamboa, R. Bernstein, and S. Y. Liao.1993. Characterization of a human ovarian carcinoma cell line: UCI 101. Gy-necol. Oncol. 48: 203–209.
40. Carbone, E., P. Neri, M. Mesuraca, M. T. Fulciniti, T. Otsuki, D. Pende, V. Groh,T. Spies, G. Pollio, D. Cosman, et al. 2005. HLA class I, NKG2D, and naturalcytotoxicity receptors regulate multiple myeloma cell recognition by naturalkiller cells. Blood 105: 251–258.
41. Conejo-Garcia, J. R., F. Benencia, M. C. Courreges, P. A. Gimotty, E. Khang,R. J. Buckanovich, K. A. Frauwirth, L. Zhang, D. Katsaros, C. B. Thompson, etal. 2004. Ovarian carcinoma expresses the NKG2D ligand Letal and promotes thesurvival and expansion of CD28� antitumor T cells. Cancer Res. 64: 2175–2182.
42. Schmidt-Wolf, I. G., P. Lefterova, B. A. Mehta, L. P. Fernandez, D. Huhn,K. G. Blume, I. L. Weissman, and R. S. Negrin. 1993. Phenotypic characteriza-tion and identification of effector cells involved in tumor cell recognition ofcytokine-induced killer cells. Exp. Hematol. 21: 1673–1679.
43. Mehta, B. A., I. G. Schmidt-Wolf, I. L. Weissman, and R. S. Negrin. 1995. Twopathways of exocytosis of cytoplasmic granule contents and target cell killing bycytokine-induced CD3�CD56� killer cells. Blood 86: 3493–3499.
44. O’Callaghan, C. A., A. Cerwenka, B. E. Willcox, L. L. Lanier, andP. J. Bjorkman. 2001. Molecular competition for NKG2D: H60 and RAE1 com-pete unequally for NKG2D with dominance of H60. Immunity 15: 201–211.
45. Vivier, E., J. A. Nunes, and F. Vely. 2004. Natural killer cell signaling pathways.Science 306: 1517–1519.
46. Garrity, D., M. E. Call, J. Feng, and K. W. Wucherpfennig. 2005. The activatingNKG2D receptor assembles in the membrane with two signaling dimers into ahexameric structure. Proc. Natl. Acad. Sci. USA 102: 7641–7646.
47. Pende, D., P. Rivera, S. Marcenaro, C. C. Chang, R. Biassoni, R. Conte,M. Kubin, D. Cosman, S. Ferrone, L. Moretta, et al. 2002. Major histocompat-ibility complex class I-related chain A and UL16-binding protein expression ontumor cell lines of different histotypes: analysis of tumor susceptibility toNKG2D-dependent natural killer cell cytotoxicity. Cancer Res. 62: 6178–6186.
48. Shum, B. P., L. R. Flodin, D. G. Muir, R. Rajalingam, S. I. Khakoo, S. Cleland,L. A. Guethlein, M. Uhrberg, and P. Parham. 2002. Conservation and variationin human and common chimpanzee CD94 and NKG2 genes. J. Immunol. 168:240–252.
49. Toyabe, S., U. Kaneko, and M. Uchiyama. 2004. Decreased DAP12 expressionin natural killer lymphocytes from patients with systemic lupus erythematosus isassociated with increased transcript mutations. J. Autoimmun. 23: 371–378.
50. Huppi, K., S. E. Martin, and N. J. Caplen. 2005. Defining and assaying RNAi inmammalian cells. Mol. Cell 17: 1–10.
51. Jamieson, A. M., A. Diefenbach, C. W. McMahon, N. Xiong, J. R. Carlyle, andD. H. Raulet. 2002. The role of the NKG2D immunoreceptor in immune cellactivation and natural killing. Immunity 17: 19–29.
52. Diefenbach, A., A. M. Jamieson, S. D. Liu, N. Shastri, and D. H. Raulet. 2000.Ligands for the murine NKG2D receptor: expression by tumor cells and activa-tion of NK cells and macrophages. Nat. Immunol. 1: 119–126.
53. Andre, P., R. Castriconi, M. Espeli, N. Anfossi, T. Juarez, S. Hue, H. Conway,F. Romagne, A. Dondero, M. Nanni, et al. 2004. Comparative analysis of humanNK cell activation induced by NKG2D and natural cytotoxicity receptors. Eur.J. Immunol. 34: 961–971.
54. Paloneva, J., M. Kestila, J. Wu, A. Salminen, T. Bohling, V. Ruotsalainen,P. Hakola, A. B. Bakker, J. H. Phillips, P. Pekkarinen, et al. 2000. Loss-of-function mutations in TYROBP (DAP12) result in a presenile dementia with bonecysts. Nat. Genet. 25: 357–361.
55. Norman, P. J., M. A. Cook, B. S. Carey, C. V. Carrington, D. H. Verity,K. Hameed, D. D. Ramdath, D. Chandanayingyong, M. Leppert, H. A. Stephens,et al. 2004. SNP haplotypes and allele frequencies show evidence for disruptiveand balancing selection in the human leukocyte receptor complex. Immunoge-netics 56: 225–237.
56. Stetson, D. B., M. Mohrs, R. L. Reinhardt, J. L. Baron, Z. E. Wang, L. Gapin,M. Kronenberg, and R. M. Locksley. 2003. Constitutive cytokine mRNAs marknatural killer (NK) and NK T cells poised for rapid effector function. J. Exp. Med.198: 1069–1076.
7828 NKG2D, DAP10, AND DAP12 IN CD8� T AND NK CELL CYTOTOXICITY
by guest on February 13, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















