
Histamine, Histamine Receptors, and their Role in Immunomodulation: An Updated Systematic Review
Upload
independentCategory
view
0download
0
of August 20, 2014.This information is current as
Cross-Presentation by Dendritic CellsHistamine Improves Antigen Uptake and
VermeulenSalamone, Cristian Cañones, Jorge Geffner and Mónica Maria Marta Amaral, Carlos Davio, Ana Ceballos, Gabriela
http://www.jimmunol.org/content/179/6/3425doi: 10.4049/jimmunol.179.6.3425
2007; 179:3425-3433; ;J Immunol
Referenceshttp://www.jimmunol.org/content/179/6/3425.full#ref-list-1
, 21 of which you can access for free at: cites 59 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2007 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on August 20, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on August 20, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

Histamine Improves Antigen Uptake and Cross-Presentationby Dendritic Cells1
Maria Marta Amaral,* Carlos Davio,† Ana Ceballos,* Gabriela Salamone,* Cristian Canones,*Jorge Geffner,* and Monica Vermeulen2*
Previous studies have shown that histamine is able to modulate the function of dendritic cells (DCs). Histamine seems to berequired for the normal differentiation of DCs. Moreover, it is capable of stimulating the chemotaxis of immature DCs and ofpromoting the differentiation of T CD4� cells into a Th2 profile. In this study, we analyzed whether histamine was able to modulateendocytosis and cross-presentation mediated by immature DCs. Our results show that both functions are stimulated by histamine.Endocytosis of soluble HRP and FITC-OVA and cross-presentation of soluble OVA were markedly increased by histamine.Interestingly, stimulation of endocytosis and cross-presentation appeared to be mediated through different histamine receptors. Infact, the enhancement of endocytosis was prevented by the histamine2 receptor (H2R) antagonist cimetidine, whereas the stimu-lation of cross-presentation was prevented by the H3R/H4R antagonist thioperamide. Of note, contrasting with the observationsmade with soluble Ags, we found that histamine did not increase either the uptake of OVA-attached to latex beads, or thecross-presentation of OVA immobilized on latex beads. This suggests that the ability of histamine to increase endocytosis andcross-presentation is dependent on the Ag form and/or the mechanisms through which the Ag is internalized by DCs. Our resultssupport that histamine may favor cross-presentation of soluble allergens by DCs enabling the activation of allergen-specific TCD8� cells, which appears to play an important role in the development of allergic responses in the airway. The Journal ofImmunology, 2007, 179: 3425–3433.
D endritic cells (DCs)3 are highly specialized APCs foundin almost all peripheral tissues as well as in primary andsecondary lymphoid organs (1, 2). They have the unique
ability to activate resting T lymphocytes and play a critical role notonly in the priming of adaptive immune responses, but also in theinduction of self-tolerance (3, 4). Upon encountering inflammatorystimuli or pathogens in the periphery, DCs become activated andundergo a number of changes, leading to their terminal differen-tiation or maturation. These changes enable DCs to activate T cellsand to direct the differentiation of CD4� T cells into distinct pro-files (5–7).
Studies on the mechanisms involved in the regulation of DCactivity are mostly restricted to cytokines, chemokines, and mi-crobial products. However, DCs are also able to recognize a largevariety of stressors generated or released during the course of dan-gerous processes, including protons (8), angiotensin-II (9), bradi-
kinin (10), the complement anaphylotoxin C5a (11), oxidants (12),and histamine (13). Histamine (2-[4-imidazole]-ethylamine) is alow m.w. amine synthesized by decarboxylation of histidine by theenzyme L-histidine decarboxylase (14). Mast cells and basophilsare the major sources of histamine, whereas certain leukocyte pop-ulations such as neutrophils (15), macrophages (16), and T lym-phocytes (17) do not store histamine, but are capable of producingand releasing high amounts of histamine. Histamine plays an im-portant role in a variety of processes, such as neurotransmission,gastrointestinal and circulatory functions, and inflammatory re-sponses responsible for allergic reactions (14, 18). More recentstudies have shown that histamine is also able to modulate thefunction of DCs. Histamine appears to be involved in the nor-mal differentiation of human DCs (19), and it is also capable ofinducing the chemotaxis of immature DCs (20) and promotingthe differentiation of T CD4� cells into a Th2 profile (21, 22),a mechanism that seems to contribute to the severity of atopicdiseases.
Recently, it has been shown that the development of airwayhyperresponsiveness and lung inflammation in experimental mod-els as well as in patients with asthma is dependent, not only onCD4� Th2 cells, but also on CD8� T cells (23, 24). These cellsshow a TC2 phenotype; produce IL-4, IL-5, and IL-13; and expressthe high-affinity receptor for leukotriene B4. This receptor appearsto play an important role in the recruitment of allergen-specificCD8� T cells in the lung (25).
Because APCs commonly find allergens in the extracellularspace, the participation of T CD8� cells in allergic reactions opensthe question of how allergens could be routed for presentation onMHC class I molecules. In this study, we show that histaminestimulates both the uptake and the presentation of Ags by MHCclass I molecules (cross-presentation), supporting a novel mecha-nism through which histamine may exacerbate the course of aller-gic reactions.
*Institute of Hematologic Research, National Academy of Medicine and NationalReference Center for AIDS, Department of Microbiology, Buenos Aires UniversitySchool of Medicine, Buenos Aires, Argentina; and †Laboratory of Radioisotopes,Buenos Aires University School of Pharmacology and Biochemestry, Buenos Aires,Argentina
Received for publication December 18, 2006. Accepted for publication June 26, 2007.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by grants from the Consejo Nacional de InvestigacionesCientıficas y Tecnicas and Agencia Nacional de Promocion Cientıfica y Tecnologica,Argentina.2 Address correspondence and reprint requests to Dr. Monica Vermeulen, Departa-mento de Inmunologıa, Instituto de Investigaciones Hematologicas, Academia Na-cional de Medicina, Pacheco de Melo 3081, 1425 Buenos Aires, Argentina. E-mailaddress: [email protected] Abbreviations used in this paper: DC, dendritic cell; ER, endoplasmic reticulum;HR, histamine receptor; MFI, mean fluorescence intensity.
Copyright © 2007 by The American Association of Immunologists, Inc. 0022-1767/07/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on August 20, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

Materials and MethodsMice
All experiments were conducted using 2-mo-old virgin female C57BL/6mice raised at the National Academy of Medicine (Buenos Aires, Argen-tina). They were housed six per cage and kept at 20 � 2°C under automatic12-h light-dark schedule. Animal care was in accordance with institutionalguidelines.
Reagents
HRP, OVA, wortmannin, cytochalasin B, and mannan from Saccharomy-ces cerevisiae were from Sigma-Aldrich. RPMI 1640, FCS, penicillin, andstreptomycin were from Invitrogen Life Technologies; histamine base, ke-totifene (histamine1 receptor (H1R) antagonist), cimetidine (H2R antago-nist), thioperamide maleate salt (H3R/H4R antagonist), and LPS from Esch-erichia coli 0111:B4 were from Sigma-Aldrich. The 257–264 OVA peptide(SIINFEKL) was provided by S. Amigorena (Institut Curie, Paris, France).
DC generation from bone marrow cultures
The procedure used in this study was described by Inaba et al. (26), withminor modifications (8). Briefly, bone marrow was flushed from the longbones of the limbs using 2 ml of RPMI 1640 with a syringe and 25-gaugeneedle. Red cells were lysed with ammonium chloride. After washing, cellswere suspended at a concentration of 1.5 � 106 cells/ml in 70% RPMI1640 medium supplemented with 10% FCS, 5.5 � 10–5 2-ME (Sigma-Aldrich) (complete medium), and 30% J588-GM cell line supernatant. Thecultures were fed every 2 days by gently swirling the plates, aspirating 50%of the medium, and adding back fresh medium with J588-GM cell linesupernatant. At day 9 of the culture, �90% of the harvested cells expressedMHC class II, CD40, and CD11c, but not Gr-1 (data not shown).
Conjugation of OVA to latex beads
A total of 200 �g/ml FITC-OVA or OVA was coupled to the surface of3-�m carboxylated polystyrene (latex) beads by passive absorption, ac-cording to the manufacturer’s instructions (Polysciences), followed by ex-tensive washing.
Endocytosis of FITC-OVA
Cells were suspended at 1.5 � 106/ml in complete medium. FITC-OVAwas added at a final concentration of 100 �g/ml, and cells were incubatedfor 30 min at 37°C. Cells were then washed three times with coldPBS containing 1% FCS and 0.01% NaN3 and were analyzed using aFACS flow cytometer and CellQuest software (BD Biosciences). The flu-orescence background (cells incubated with FITC-OVA at 4°C) was al-ways subtracted. In some experiments, we used the dye trypan blue toquench extracellular fluorescence, as described (8, 27). Endocytosis assayswere performed, as previously indicated, but acquisition of samples wasconducted in the presence of 200 �g/ml trypan blue. The efficacy of trypanblue to quench extracellular fluorescence was controlled in experiments inwhich DCs were stained with FITC mAb directed to cell surface Ags (30min at 4°C). In all cases, we observed that fluorescence intensity wasdiminished by �85% when the acquisition of the samples was performedin the presence of trypan blue.
Endocytosis of HRP
Endocytosis of HRP was performed, as previously described (28). Briefly,DCs (1.5 � 106/ml) were suspended in complete medium. HRP was addedat a final concentration of 150 �g/ml, and cells were cultured for 30 minat 37°C. Then DCs were collected, washed four times in PBS containing1% FCS and four times in cold PBS alone, and lysed with 0.05% TritonX-100 in 10 mM Tris buffer (pH 7.4) for 30 min, and the enzyme activityof the lysate was measured using o-phenylenediamine and H2O2 as sub-strates with reference to a standard curve, at 492 nm. The amount of HRPtaken up by DCs was determined as the difference between HRP activitiesin disrupted and nondisrupted cells. HRP activity in nondisrupted DCs wasalways �15% compared with disrupted cells.
Phagocytosis of FITC-OVA-coated latex beads
A total of 1 � 106/ml DCs in 200 �l of complete culture medium wasincubated with 50 �l of FITC-OVA-coated latex beads (109 beads/ml) for3 h at 37°C. The uptake of FITC-OVA-coated latex beads was quantifiedby flow cytometry. Acquisition of the samples was performed in the pres-ence of trypan blue (200 �g/ml) to quench extracellular fluorescence.
Analysis of the phenotype of DCs
Cell staining was performed using the following mAbs: anti-CD11c FITC,anti-CD40 FITC, anti-CD86 FITC, anti-I-Ab PE (MHC class II), anti-CD11b PE, anti-GR-1 PE, and anti-H-2Kb (MHC class I) (BD Pharmin-gen). Cell surface Ag expression was evaluated by single staining, andanalysis was performed using a FACS flow cytometer and CellQuest soft-ware (BD Biosciences).
Radioligand-binding assays for the expression of H1R and H2R
[3H]Mepyramine-binding assay. Duplicate assays were per-formed in polyethylene tubes in 50 mM Tris-HCl (pH 7.4), aspreviously described (29). For saturation studies, increasing con-centrations of [3H]mepyramine, ranging from 0.01 to 15 nM, wereincubated with 106 cells/tube, in the absence or presence of 10 �Mmepyramine, in a total volume of 100 �l. After 40 min at 4°C,incubation was stopped by dilution with 3 ml of ice-cold 50 mMTris-HCl (pH 7.4) and rapid filtration under reduced pressure ontoWhatman GF/B glass-fiber filters, followed by three washes with3 ml of ice-cold buffer. Experiments were conducted at 4°C toavoid internalization of the ligand.
[3H]Tiotidine-binding assay. Duplicate assays were performed,as previously described (30), in polyethylene tubes in 50 mM Tris-HCl (pH 7.4). For saturation studies, increasing concentrations of[3H]tiotidine, ranging from 0.4 to 130 nM, were incubated with106 cells/tube, in the absence or presence of 10 �M tiotidine, in atotal volume of 100 �l. After 40 min at 4°C, incubation wasstopped by dilution with 3 ml of ice-cold 50 mM Tris-HCl (pH 7.4)and rapid filtration under reduced pressure onto Whatman GF/Bglass-fiber filters, followed by three washes with 3 ml of ice-coldbuffer. Experiments were conducted at 4°C.
In all cases, specific binding was calculated by subtraction ofnonspecific from total binding. Analysis of radioligand-bindingdata was done using GraphPad Prism 3.00 for Windows. One-wayANOVA with Dunnett’s posttest was performed using GraphPadInStat version 3.01.
Analysis of the expression of H3R and H4R by RT-PCR
Total RNA was extracted from brain (positive control for H3R), bone mar-row (positive control for H4R), and mouse DCs, using TRIzol reagent(Invitrogen Life Technologies). The reverse-transcription reaction con-tained 3 �g of total RNA, and it was performed using the M-MLV reverse-transcriptase enzyme (Promega). The primers were provided by InvitrogenLife Technologies. Forward primers for the H3R and H4R are as follows:TAC AAG GGC CTG GCC GTA GAA GG and CGT GTT GTT TAACTG GAA TTT TGG AAG TGG AAT CTG CAT G, respectively. Re-verse primers for the H3R and H4R are as follows: GCT GTC GCG GGACAA GAA GG and ACC AAG AAA GCC AGT ATC CAA ACA GCCACC ATT TGA GC, respectively. A GeneAmp PCR system (PerkinElmer/Applied Biosystems) was used. PCR products were separated on a 1.5%agarose gel, stained with ethidium bromide, and visualized by an UVtransilluminator.
Histamine production
It was measured in cell supernatants using a commercial histamine ELISAkit (IBL).
Quantification of cellular apoptosis and viability by fluorescencemicroscopy
Quantification was conducted, as described (31), using the fluorescentDNA-binding dyes acridine orange (100 �g/ml), to determine the percent-age of cells that had undergone apoptosis, and ethidium bromide (100�g/ml), to differentiate between viable and nonviable cells. With thismethod, nonapoptotic cell nuclei show structure, variations in fluorescenceintensity that reflect the distribution of euchromatin and heterochromatin.By contrast, apoptotic nuclei exhibit highly condensed chromatin that isuniformly stained by acridine orange. In fact, the entire apoptotic nucleusis present as bright spherical beads. To assess the percentage of cells show-ing morphologic features of apoptosis, at least 200 cells were scored ineach experiment.
3426 HISTAMINE STIMULATES CROSS-PRESENTATION BY DCs
by guest on August 20, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
Ag cross-presentation assay
Presentation of OVA epitope 257–264 on Kb was detected using the T cellhybridoma B3Z, which carries a �-galactosidase construct driven byNF-AT elements from the IL-2 promoter (8). For Ag presentation assays,DCs were exposed to different concentrations of soluble OVA or OVA-coated latex beads (DC:bead ratio: 1:50) during 3 h at 37°C. Then cellswere washed, suspended in complete medium, and cultured in the presenceof the T cell hybridoma B3Z. After 18 h of culture, cells were washed withPBS, and a colorimetric assay using o-nitrophenyl-p-D-galactoside (ONPG)(Sigma-Aldrich) as a substrate was used to detect LacZ activity in B3Z lysates.Pulse-chase experiments were conducted by incubating DCs with solubleOVA for 3 h at 37°C. Cells were then washed and cultured for various chaseperiods, after which B3Z cells were added and cross-presentation was as-sessed, as described above.
Statistical analysis
The significance between means was assessed by Student’s t test. Value ofp � 0.05 was taken as indicating statistical significance.
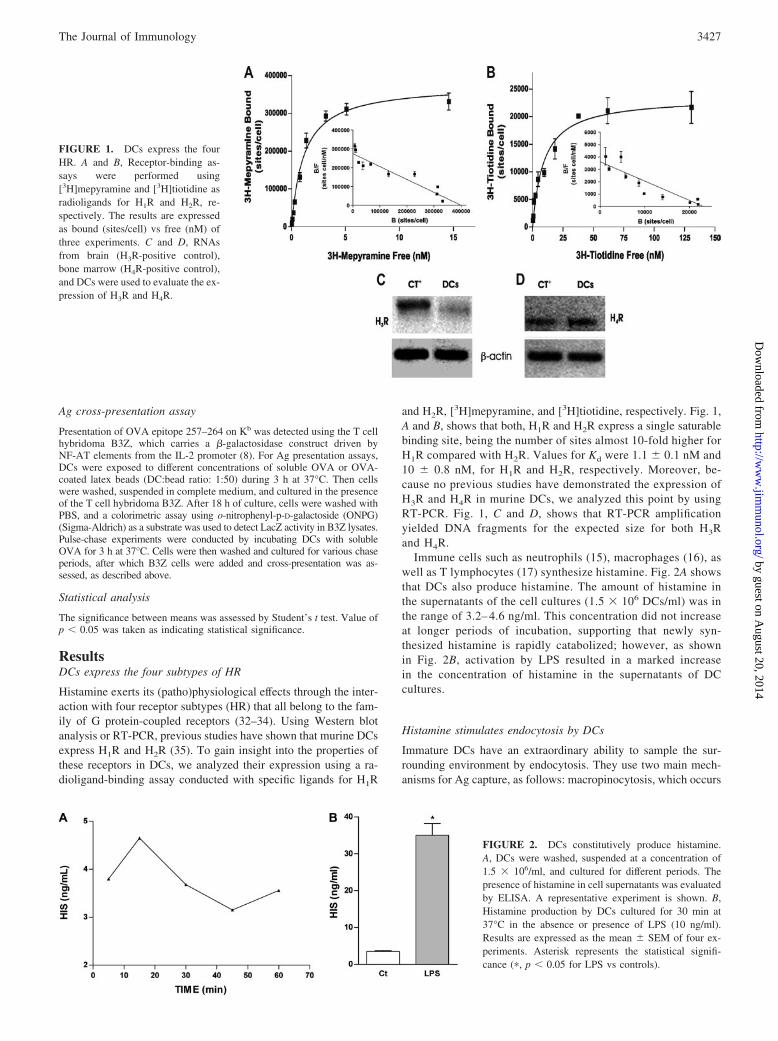
ResultsDCs express the four subtypes of HR
Histamine exerts its (patho)physiological effects through the inter-action with four receptor subtypes (HR) that all belong to the fam-ily of G protein-coupled receptors (32–34). Using Western blotanalysis or RT-PCR, previous studies have shown that murine DCsexpress H1R and H2R (35). To gain insight into the properties ofthese receptors in DCs, we analyzed their expression using a ra-dioligand-binding assay conducted with specific ligands for H1R
and H2R, [3H]mepyramine, and [3H]tiotidine, respectively. Fig. 1,A and B, shows that both, H1R and H2R express a single saturablebinding site, being the number of sites almost 10-fold higher forH1R compared with H2R. Values for Kd were 1.1 � 0.1 nM and10 � 0.8 nM, for H1R and H2R, respectively. Moreover, be-cause no previous studies have demonstrated the expression ofH3R and H4R in murine DCs, we analyzed this point by usingRT-PCR. Fig. 1, C and D, shows that RT-PCR amplificationyielded DNA fragments for the expected size for both H3Rand H4R.
Immune cells such as neutrophils (15), macrophages (16), aswell as T lymphocytes (17) synthesize histamine. Fig. 2A showsthat DCs also produce histamine. The amount of histamine inthe supernatants of the cell cultures (1.5 � 106 DCs/ml) was inthe range of 3.2– 4.6 ng/ml. This concentration did not increaseat longer periods of incubation, supporting that newly syn-thesized histamine is rapidly catabolized; however, as shownin Fig. 2B, activation by LPS resulted in a marked increasein the concentration of histamine in the supernatants of DCcultures.
Histamine stimulates endocytosis by DCs
Immature DCs have an extraordinary ability to sample the sur-rounding environment by endocytosis. They use two main mech-anisms for Ag capture, as follows: macropinocytosis, which occurs
FIGURE 1. DCs express the fourHR. A and B, Receptor-binding as-says were performed using[3H]mepyramine and [3H]tiotidine asradioligands for H1R and H2R, re-spectively. The results are expressedas bound (sites/cell) vs free (nM) ofthree experiments. C and D, RNAsfrom brain (H3R-positive control),bone marrow (H4R-positive control),and DCs were used to evaluate the ex-pression of H3R and H4R.
FIGURE 2. DCs constitutively produce histamine.A, DCs were washed, suspended at a concentration of1.5 � 106/ml, and cultured for different periods. Thepresence of histamine in cell supernatants was evaluatedby ELISA. A representative experiment is shown. B,Histamine production by DCs cultured for 30 min at37°C in the absence or presence of LPS (10 ng/ml).Results are expressed as the mean � SEM of four ex-periments. Asterisk represents the statistical signifi-cance (�, p � 0.05 for LPS vs controls).
3427The Journal of Immunology
by guest on August 20, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
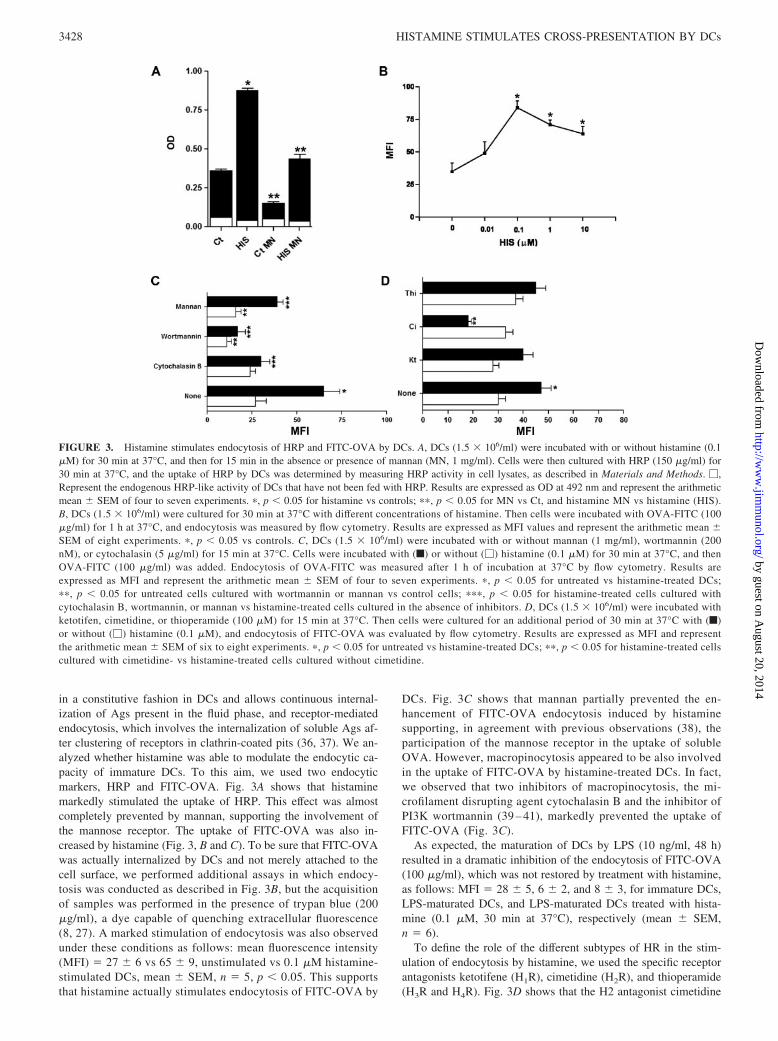
in a constitutive fashion in DCs and allows continuous internal-ization of Ags present in the fluid phase, and receptor-mediatedendocytosis, which involves the internalization of soluble Ags af-ter clustering of receptors in clathrin-coated pits (36, 37). We an-alyzed whether histamine was able to modulate the endocytic ca-pacity of immature DCs. To this aim, we used two endocyticmarkers, HRP and FITC-OVA. Fig. 3A shows that histaminemarkedly stimulated the uptake of HRP. This effect was almostcompletely prevented by mannan, supporting the involvement ofthe mannose receptor. The uptake of FITC-OVA was also in-creased by histamine (Fig. 3, B and C). To be sure that FITC-OVAwas actually internalized by DCs and not merely attached to thecell surface, we performed additional assays in which endocy-tosis was conducted as described in Fig. 3B, but the acquisitionof samples was performed in the presence of trypan blue (200�g/ml), a dye capable of quenching extracellular fluorescence(8, 27). A marked stimulation of endocytosis was also observedunder these conditions as follows: mean fluorescence intensity(MFI) � 27 � 6 vs 65 � 9, unstimulated vs 0.1 �M histamine-stimulated DCs, mean � SEM, n � 5, p � 0.05. This supportsthat histamine actually stimulates endocytosis of FITC-OVA by
DCs. Fig. 3C shows that mannan partially prevented the en-hancement of FITC-OVA endocytosis induced by histaminesupporting, in agreement with previous observations (38), theparticipation of the mannose receptor in the uptake of solubleOVA. However, macropinocytosis appeared to be also involvedin the uptake of FITC-OVA by histamine-treated DCs. In fact,we observed that two inhibitors of macropinocytosis, the mi-crofilament disrupting agent cytochalasin B and the inhibitor ofPI3K wortmannin (39 – 41), markedly prevented the uptake ofFITC-OVA (Fig. 3C).
As expected, the maturation of DCs by LPS (10 ng/ml, 48 h)resulted in a dramatic inhibition of the endocytosis of FITC-OVA(100 �g/ml), which was not restored by treatment with histamine,as follows: MFI � 28 � 5, 6 � 2, and 8 � 3, for immature DCs,LPS-maturated DCs, and LPS-maturated DCs treated with hista-mine (0.1 �M, 30 min at 37°C), respectively (mean � SEM,n � 6).
To define the role of the different subtypes of HR in the stim-ulation of endocytosis by histamine, we used the specific receptorantagonists ketotifene (H1R), cimetidine (H2R), and thioperamide(H3R and H4R). Fig. 3D shows that the H2 antagonist cimetidine
FIGURE 3. Histamine stimulates endocytosis of HRP and FITC-OVA by DCs. A, DCs (1.5 � 106/ml) were incubated with or without histamine (0.1�M) for 30 min at 37°C, and then for 15 min in the absence or presence of mannan (MN, 1 mg/ml). Cells were then cultured with HRP (150 �g/ml) for30 min at 37°C, and the uptake of HRP by DCs was determined by measuring HRP activity in cell lysates, as described in Materials and Methods. �,Represent the endogenous HRP-like activity of DCs that have not been fed with HRP. Results are expressed as OD at 492 nm and represent the arithmeticmean � SEM of four to seven experiments. �, p � 0.05 for histamine vs controls; ��, p � 0.05 for MN vs Ct, and histamine MN vs histamine (HIS).B, DCs (1.5 � 106/ml) were cultured for 30 min at 37°C with different concentrations of histamine. Then cells were incubated with OVA-FITC (100�g/ml) for 1 h at 37°C, and endocytosis was measured by flow cytometry. Results are expressed as MFI values and represent the arithmetic mean �SEM of eight experiments. �, p � 0.05 vs controls. C, DCs (1.5 � 106/ml) were incubated with or without mannan (1 mg/ml), wortmannin (200nM), or cytochalasin (5 �g/ml) for 15 min at 37°C. Cells were incubated with (f) or without (�) histamine (0.1 �M) for 30 min at 37°C, and thenOVA-FITC (100 �g/ml) was added. Endocytosis of OVA-FITC was measured after 1 h of incubation at 37°C by flow cytometry. Results areexpressed as MFI and represent the arithmetic mean � SEM of four to seven experiments. �, p � 0.05 for untreated vs histamine-treated DCs;��, p � 0.05 for untreated cells cultured with wortmannin or mannan vs control cells; ���, p � 0.05 for histamine-treated cells cultured withcytochalasin B, wortmannin, or mannan vs histamine-treated cells cultured in the absence of inhibitors. D, DCs (1.5 � 106/ml) were incubated withketotifen, cimetidine, or thioperamide (100 �M) for 15 min at 37°C. Then cells were cultured for an additional period of 30 min at 37°C with (f)or without (�) histamine (0.1 �M), and endocytosis of FITC-OVA was evaluated by flow cytometry. Results are expressed as MFI and representthe arithmetic mean � SEM of six to eight experiments. �, p � 0.05 for untreated vs histamine-treated DCs; ��, p � 0.05 for histamine-treated cellscultured with cimetidine- vs histamine-treated cells cultured without cimetidine.
3428 HISTAMINE STIMULATES CROSS-PRESENTATION BY DCs
by guest on August 20, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
completely suppressed the stimulation of endocytosis by hista-mine, whereas the H1 and the H3/H4 antagonists, used atconcentrations able to block the activity of these receptors in DCs
(42–44), did not exert any effect. It should be noted that none ofthe antagonists used modified the endocytic ability of DCs whenthey were cultured in the absence of exogenous histamine,
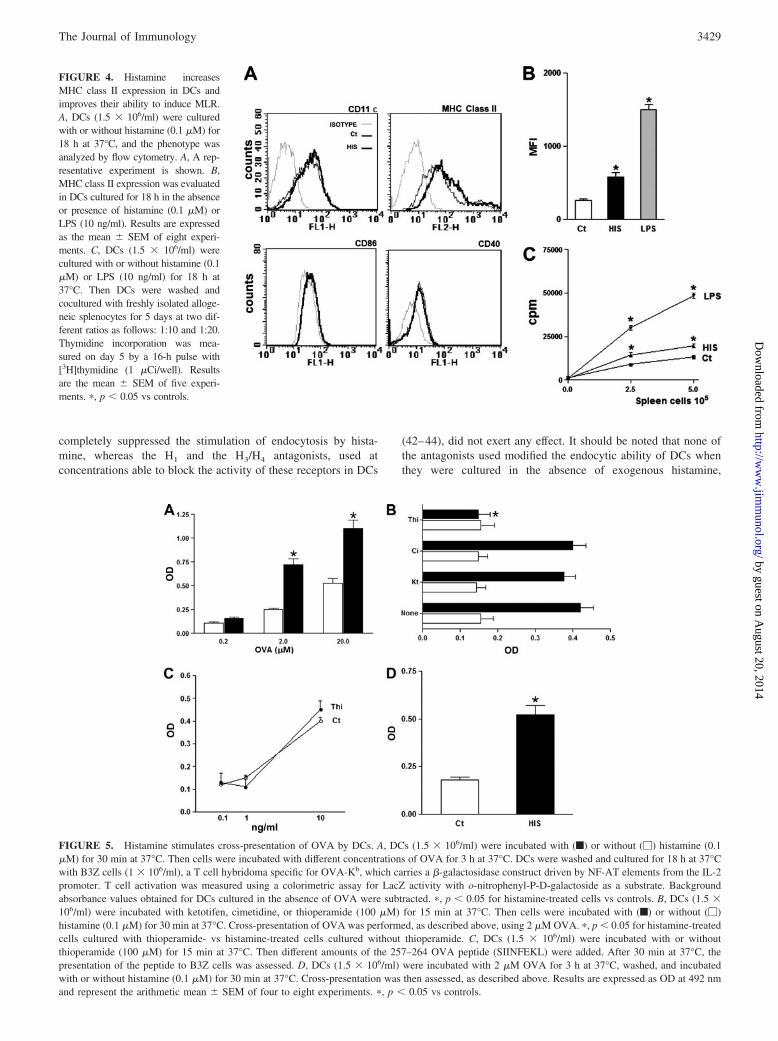
FIGURE 4. Histamine increasesMHC class II expression in DCs andimproves their ability to induce MLR.A, DCs (1.5 � 106/ml) were culturedwith or without histamine (0.1 �M) for18 h at 37°C, and the phenotype wasanalyzed by flow cytometry. A, A rep-resentative experiment is shown. B,MHC class II expression was evaluatedin DCs cultured for 18 h in the absenceor presence of histamine (0.1 �M) orLPS (10 ng/ml). Results are expressedas the mean � SEM of eight experi-ments. C, DCs (1.5 � 106/ml) werecultured with or without histamine (0.1�M) or LPS (10 ng/ml) for 18 h at37°C. Then DCs were washed andcocultured with freshly isolated alloge-neic splenocytes for 5 days at two dif-ferent ratios as follows: 1:10 and 1:20.Thymidine incorporation was mea-sured on day 5 by a 16-h pulse with[3H]thymidine (1 �Ci/well). Resultsare the mean � SEM of five experi-ments. �, p � 0.05 vs controls.
FIGURE 5. Histamine stimulates cross-presentation of OVA by DCs. A, DCs (1.5 � 106/ml) were incubated with (f) or without (�) histamine (0.1�M) for 30 min at 37°C. Then cells were incubated with different concentrations of OVA for 3 h at 37°C. DCs were washed and cultured for 18 h at 37°Cwith B3Z cells (1 � 106/ml), a T cell hybridoma specific for OVA-Kb, which carries a �-galactosidase construct driven by NF-AT elements from the IL-2promoter. T cell activation was measured using a colorimetric assay for LacZ activity with o-nitrophenyl-P-D-galactoside as a substrate. Backgroundabsorbance values obtained for DCs cultured in the absence of OVA were subtracted. �, p � 0.05 for histamine-treated cells vs controls. B, DCs (1.5 �106/ml) were incubated with ketotifen, cimetidine, or thioperamide (100 �M) for 15 min at 37°C. Then cells were incubated with (f) or without (�)histamine (0.1 �M) for 30 min at 37°C. Cross-presentation of OVA was performed, as described above, using 2 �M OVA. �, p � 0.05 for histamine-treatedcells cultured with thioperamide- vs histamine-treated cells cultured without thioperamide. C, DCs (1.5 � 106/ml) were incubated with or withoutthioperamide (100 �M) for 15 min at 37°C. Then different amounts of the 257–264 OVA peptide (SIINFEKL) were added. After 30 min at 37°C, thepresentation of the peptide to B3Z cells was assessed. D, DCs (1.5 � 106/ml) were incubated with 2 �M OVA for 3 h at 37°C, washed, and incubatedwith or without histamine (0.1 �M) for 30 min at 37°C. Cross-presentation was then assessed, as described above. Results are expressed as OD at 492 nmand represent the arithmetic mean � SEM of four to eight experiments. �, p � 0.05 vs controls.
3429The Journal of Immunology
by guest on August 20, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

supporting that this function is not under the control of an auto-crine loop involving histamine.
Histamine induces only minor effects on the phenotype andallostimulatory capacity of immature DCs
Contrasting results have been reported regarding the ability of his-tamine to induce the activation of DCs (13, 14, 34, 45). To re-examine this point, DCs were cultured for 18 h in the absence orpresence of 0.1 �M histamine, and their phenotype was analyzedby flow cytometry. Fig. 4, A and B, shows that histamine induceda low, but significant increase in the expression of MHC class II,without modifying the expression of CD11c, CD40, and CD86.Similar results were observed using 1 �M histamine (data notshown). As shown in Fig. 4C, the enhanced expression of MHCclass II induced by histamine correlated with the T cell-primingactivity of DCs, as indicated by the ability of histamine to exert alow, but significant increase in the allostimulatory response medi-ated by DCs.
Histamine stimulates cross-presentation by DCs
Cross-presentation is the process by which extracellular Ags,which are normally presented in association with MHC class IImolecules, are instead routed for presentation on MHC class Imolecules, enabling extracellular Ags to activate T CD8� cells (4).This pathway is mainly mediated by DCs and can lead either to thetolerization or activation of Ag-specific CD8� T cells (3, 46). Be-cause allergen-specific T CD8� cells appear to play an importantrole in the development of allergic reactions in the lung, and alsoconsidering that allergens are usually found at the extracellularspace, we examined whether histamine may be able to stimulateAg cross-presentation. This point was studied by analyzing thepresentation of OVA to a CD8� T cell hybridoma called B3Z,which carries a �-galactosidase construct driven by NF-AT ele-ments from the IL-2 promoter, enabling the analysis of T cell
activation by measuring �-galactosidase activity in cell lysates (8,39). DCs were incubated with or without 0.1 �M histamine for 30min at 37°C. They were then exposed to different concentrations ofOVA during 3 h at 37°C, and presentation of the OVA257–264
epitope/H-2Kb to B3Z cells was assessed. Fig. 5A shows that his-tamine markedly improved MHC class I presentation. In fact, toallow similar levels of Ag presentation, untreated DCs requiredconcentrations of Ag in the extracellular medium at least 10-foldhigher compared with histamine-treated DCs. As expected, no Agpresentation was observed when DCs were fixed with glutaralde-hyde before the addition of OVA (data not shown).
Using the specific receptor antagonists ketotifene (H1R), cime-tidine (H2R), and thioperamide (H3R and H4R), we then analyzedthe role of HR in the stimulation of cross-presentation. Fig. 5Bshows that ketotifene and cimetidine did not mediate any effect,whereas thioperamide completely abrogated the stimulation ofcross-presentation by histamine. As shown in Fig. 5B, none of theantagonists assessed modulated cross-presentation of DCs culturedin the absence of histamine. To rule out any toxic effect mediatedby thioperamide, we performed additional experiments in whichthe effect of thioperamide on the direct presentation of theOVA257–264 peptide to B3Z cells was analyzed. Fig. 5C shows thatthioperamide did not disturb this process. Additional experimentswere also performed to be sure that thioperamide treatment did notresult in a loss of DC viability. To this aim, DCs were culturedwith 100 �M thioperamide for 48 h, in the absence or presence of0.1 �M histamine, and the percentages of cell viability were de-termined by fluorescence microscopy, as described in Materialsand Methods. The percentages of cell viability were, in all cases,higher than 85%, whereas the percentages of apoptotic cells werelower than 12%. We also determined the absolute number of DCsrecovered after 48 h of culture, and found that cell recovery wasalways higher than 80%. Together, these data indicate that thiop-eramide does not induce deleterious effects on DCs.
FIGURE 6. Histamine does not in-crease the expression of MHC class I,but prolongs the expression of MHCclass I-257–264 OVA peptide com-plexes in DCs pulsed with solubleOVA. A, DCs (1.5 � 106/ml) were cul-tured with or without histamine (0.1�M) for 18 h at 37°C, and expressionof MHC class I was analyzed by flowcytometry. A representative experimentis shown. B, DCs (1.5 � 106/ml) werecultured with (F) or without (E) hista-mine (0.1 �M) for different periods at37°C, and the expression of MHC classI was analyzed by flow cytometry. Re-sults are expressed as MFI and repre-sent the arithmetic mean � SEM ofthree experiments. C, Pulse-chase ex-periments were performed by incubat-ing histamine-pretreated (0.1 �M, 30min at 37°C) or untreated DCs with orwithout OVA for 3 h at 37°C. OVAwas then removed from the culture me-dium, and DCs were cultured for vari-ous chase periods, after which B3Zcells were added and cross-presentationwas assessed, as described in Materialsand Methods. A representative experi-ment (n � 3) is shown.
3430 HISTAMINE STIMULATES CROSS-PRESENTATION BY DCs
by guest on August 20, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

We then performed a new set of experiments directed to analyzethe mechanisms involved in the stimulation of cross-presentationby histamine. First, we studied whether histamine was also able tostimulate cross-presentation when added after Ag pulsing. DCswere first cultured with OVA (2 �M) for 3 h at 37°C in the absenceof histamine. Then cells were washed and cultured for an addi-tional period of 30 min at 37°C with or without 0.1 �M histamine,in the absence of exogenous OVA. Cross-presentation was thenassessed, as described in Fig. 5A. The results obtained (Fig. 5D)showed that histamine stimulated cross-presentation of OVA evenwhen it was added to DC cultures after extracellular OVA wasremoved from the culture medium. We then analyzed whether his-tamine may be able to increase the expression of MHC class Imolecules on DCs. No changes were observed in the expression ofMHC class I in DCs cultured for 0.5, 2, 4, and 18 h in the presenceof histamine (Fig. 6, A and B).
Additional studies were then performed to determine whetherhistamine may enable OVA-pulsed DCs to express the MCH-Ipeptide complexes for longer periods compared with control DCs.This point was analyzed using a functional assay in pulse-chaseexperiments. Histamine-pretreated or untreated DCs were culturedwith OVA for 3 h at 37°C. OVA was then removed from theculture medium and DCs were cultured for various chase periods,after which B3Z cells were added and cross-presentation was as-sessed, as described above. Fig. 6C shows that the ability of un-treated DCs to cross-present OVA declined after a chase period of48 h. By contrast, the ability of histamine-treated DCs only de-clined after chase periods of 72–96 h.
It is well known that proteins internalized by either macropino-cytosis or phagocytosis can be cross-presented (47, 48). To deter-mine whether histamine was also able to enhance cross-presenta-tion of Ags internalized by phagocytosis, experiments wereperformed using OVA-coated latex beads. As shown in Fig. 7A,and contrasting with the observations made with the soluble AgsHRP and FITC-OVA, histamine did not increase the uptake ofFITC-OVA-coated latex beads, suggesting that it is unable to stim-ulate phagocytosis. Interestingly, Fig. 7B shows that histamine didnot improve cross-presentation of OVA immobilized on latexbeads, indicating that the ability of histamine to increase cross-presentation is strongly dependent on the Ag form and/or themechanisms through which the Ag is internalized by DCs.
DiscussionHistamine is an important mediator of immediate-type allergic re-actions, and also modulates the function of DCs, the only APCcapable of activating naive T lymphocytes (14). Previous studiesdirected to analyze the effect of histamine on DCs have mainlyfocused on the capacity of DCs to regulate the differentiation of TCD4� cells into a Th1 or Th2 profile. Histamine has been shownto inhibit IL-12 and to increase IL-10 production by activated DCs,favoring the differentiation of T CD4� cells into a Th2 profile (21,49). This effect involves H2R and H4R and represents a positivefeedback loop that seems to contribute to the severity of atopicdiseases (20, 31, 50). It has also been demonstrated that histamine,acting through the H1R and H3R, induces the chemotaxis of im-mature DCs (45). Moreover, Szeberenyi et al. (19) have shown thathistamine is actively synthesized during the differentiation of DCsand that inhibition of histamine synthesis disturbs this process.
In the present study, we observed that immature DCs express thefour subtypes of HR. Despite that in most systems H1R and H2Rare able to mediate opposite effects (45, 51), no previous studieshave examined the density and the affinity of these receptors inDCs. Using a radioligand-binding assay conducted with specificligands for the H1R and H2R, we were able to define that each ofthese receptors expresses a single and saturable binding site ofhigh affinity, and that the number of sites was almost 10-foldhigher for H1R compared with H2R. Moreover, using RT-PCR, wefound that DCs express not only the H4R, which is primarily ex-pressed in the spleen, thymus, bone marrow, and peripheral bloodleukocytes (14, 34, 52), but also the H3R, a receptor mainly ex-pressed in the CNS (13, 33). We were unable, however, to validatethe gene expression of H3R and H4R at the protein level by West-ern blot analysis because no specific Abs are available.
Immature DCs constitutively macropinocytose extracellularfluid (37), and also express a large variety of receptors mediatingendocytosis and phagocytosis of Ags and pathogens (53). Our re-sults show that histamine stimulates endocytosis by immatureDCs. Because immature DCs and mast cells are strategically lo-calized in close proximity at anatomic sites with high antigenicexposure such as skin and mucosal surfaces, we hypothesize thatactivation of endocytosis by histamine may be able to improve theAg-sampling capacity of immature DCs in vivo. In this regard, itshould be noted that concentrations of histamine similar to that
FIGURE 7. Histamine does not stimulate phagocytosis nor cross-presentation of OVA immobilized on latex beads. A, DCs (1.5 � 106/ml) were culturedwith (histograms B and D) or without 0.1 �M histamine (histograms A and C) for 30 min at 37°C, and phagocytosis of FITC-OVA-coated latex beads wasperformed, as described in Materials and Methods. A representative experiment is shown (n � 4). Histograms A and B, Cells incubated without FITC-OVA-coated latex beads. Histograms C and D, Cells incubated with FITC-OVA-coated latex beads. B, DCs (1.5 � 106/ml) were incubated without (�)or with (f) histamine (0.1 �M) for 30 min at 37°C. Then cells were incubated with OVA (2 �M) or OVA-coated latex beads, at a DC:bead ratio of 1:50,for 3 h at 37°C. DCs were then washed, and the cross-presentation assay was performed. Results are expressed as OD at 492 nm and represent the arithmeticmean � SEM of three to five experiments. �, p � 0.05 vs controls.
3431The Journal of Immunology
by guest on August 20, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

used in our study (0.1 �M) were described in vivo at inflammatorysites during the course of allergic reactions (54–56).
In this study, we report, for the first time, that histamine is ca-pable of stimulating cross-presentation of extracellular Ags. Cross-presentation enables DCs to present captured Ags through MHCclass I molecules, thus providing a mechanism by which extracel-lular Ags could activate T CD8� cell-mediated responses (44, 57).Interestingly, cross-presentation appears to play an important role,not only in antiviral and antitumor immunity, but also in the in-duction of tolerance (3, 58, 59). The ability of histamine to stim-ulate cross-presentation via H3R and/or H4R may represent a novelmechanism through which mast cells, the major cellular source ofhistamine, modulate the course of the adaptive immune response.
We have yet not defined the mechanisms through which hista-mine increases cross-presentation of soluble OVA. The enhance-ment of cross-presentation could not be merely explained by in-creased Ag uptake, because stimulation of cross-presentation andendocytosis appears to be mediated through distinct HR. Pulse-chase experiments, in contrast, suggested that histamine enablesAg-pulsed DCs to express MHC I-peptide complexes at the cellsurface for longer periods. An enhanced expression of MHC I-peptide complexes as well as a more persistent expression of thesecomplexes in Ag-pulsed DCs could be explained through differentmechanisms, such as the facilitation of endosomal escape of theAg to the cytosol or by changes in the mechanisms operatingdownstream this step. Of note, histamine was completely unable toincrease cross-presentation of OVA bound to latex beads, used asa model of particulate Ag. It is well known that peptides derivedfrom particulate Ags are cross-presented more efficiently than sol-uble Ags. This effect cannot be explained by increased Ag uptake,because cross-presentation of soluble Ags is enhanced by the ad-dition of unrelated particles (47, 48), suggesting that phagocytosisitself favors Ag delivery into the MHC class I pathway. It has alsobeen shown that particulate and soluble Ags use different transportpathways (47). Particulate Ags access to peripheral endoplasmicreticulum (ER)-like phagosomes, which are competent for cross-presentation, whereas soluble proteins may enter the lumen of theER, being then translocated to the citosol. Finally, the generatedpeptides were transported back into the ER, where they bind toMHC class I molecules. We speculate that the ability of histamineto stimulate cross-presentation of soluble, but not particulate Agscould reflect an action exerted by histamine on the intracellulartransport pathways specifically used by soluble, but not by partic-ulate Ags. Interestingly, we also found that histamine efficientlystimulated the uptake of soluble, but not particulate Ags by DCs,supporting the view that the overall handling of soluble, but notparticulate Ags by DCs might be under the regulation of histaminein peripheral tissues during the course of inflammatory allergicreactions.
It is now becoming clear that allergen-specific CD8� T cells areimportant contributors to the development of allergic responses inthe lung. They appear to be required for the development of bothairway hyperresponsiveness and eosinophilic airway inflammation(23, 60). We hypothesize that stimulation of cross-presentation ofallergens by DCs induced by histamine may be a novel mechanismthrough which mast cells contribute to the development of allergicresponses via activation of allergen-specific T CD8� cells.
Previous studies have shown that DCs produce histamine (19,50). In fact, the intracellular levels of both histidine decarboxylaseand histamine appear to increase during the differentiation of hu-man monocytes into DCs (14, 52). Moreover, Dunford et al. (22)have reported the presence of histamine in cocultures of murineDCs and spleen T CD4� cells, and also observed that the amountsof histamine in cell supernatants markedly increased as a conse-
quence of LPS stimulation. Consistent with these results, we re-ported in this study that immature DCs constitutively producehistamine. However, the fact that neither the stimulation of endo-cytosis nor the promotion of cross-presentation was modulated byHR antagonists in DCs cultured in the absence of exogenous his-tamine supports that the autocrine production of histamine doesnot exert a modulatory effect on DC function under resting con-ditions. However, upon activation by LPS, the amount of hista-mine released by DCs markedly increased reaching 0.1 �M, aconcentration able to increase endocytosis and cross-presentationby DCs (Figs. 3 and 5). This result supports the hypothesis that anautocrine loop involving histamine may modulate the function ofDCs under inflammatory conditions.
AcknowledgmentsWe thank Selma Tolosa and Evelia Lopez for their technical assistance,and Maria Rita Furnkorn for her secretarial assistance.
DisclosuresThe authors have no financial conflict of interest.
References1. Grannucci, F., M. Foti, and P. Ricciardi-Castagnoli. 2005. Dendritic cell biology.
Adv. Immunol. 88: 193–233.2. Steinman, R. M. 2003. Some interferences of dendritic cell biology. APMIS 111:
675–697.3. Steinman, R. M., D. Hawiger, and M. C. Nussenzweig. 2003. Tolerogenic den-
dritic cells. Annu. Rev. Immunol. 21: 685–711.4. Carbone, F. R., G. T. Belz, and W. R. Heath. 2004. Transfer of antigen between
migrating and lymph node-resident DCs in peripheral T-cell tolerance and im-munity. Trends Immunol. 25: 655–658.
5. Cop, E., and R. Medzhitov. 2003. Recognition of microbial infection by Toll-likereceptors. Curr. Opin. Immunol. 15: 396–401.
6. Sporri, R., and C. Reis e Sousa. 2005. Inflammatory mediators are insufficient forfull dendritic cell activation and promote expansion of CD4� T cell populationslacking helper function. Nat. Immunol. 6: 163–170.
7. Reis e Sousa, C. 2004. Activation of dendritic cells: translating innate into ad-aptative immunity. Curr. Opin. Immunol. 16: 21–25.
8. Vermeulen, M., M. Giordano, A. S. Trevani, C. Sedlik, R. Gamberale,P. Fernandez-Calotti, G. Salamone, S. Raiden, J. Sanjurjo, and J. R. Geffner.2004. Acidosis improves uptake of antigens and MHC class I-restricted presen-tation by dendritic cells. J. Immunol. 172: 3196–3204.
9. Nahmod, K., M. E. Vermeulen, S. Raiden, G. Salamone, R. Gamberale,P. Fernandez-Calotti, A. Alvarez, V. Nahmod, M. Giordano, and J. R. Geffner.2003. Control of dendritic cell differentiation by angiotensin II. FASEB J. 17:491–493.
10. Aliberti, J., J. P. Viola, A. Vieira-de-Abreu, P. T. Bozza, A. Sher, andJ. Scharfstein. 2003. Cutting edge: bradykinin induces IL-12 production by den-dritic cells: a danger signal that drives Th1 polarization. J. Immunol. 70:5349–5353.
11. Kohl, J., R. Baelder, I. P. Lewkowich, M. K. Pandey, H. Hawlisch, L. Wang,J. Best, N. S. Herman, A. A. Sproles, J. Zwirner, et al. 2006. A regulatory rolefor the C5a anaphylatoxin in type 2 immunity in asthma. J. Clin. Invest. 116:783–796.
12. Handley, M. E., M. Thakker, G. Pollara, B. M. Chain, and D. R. Katz. 2005. JNKactivation limits dendritic cell maturation in response to reactive oxygen speciesby the induction of apoptosis. Free Radical Biol. Med. 38: 1637–1652.
13. Schneider, E., M. Rolli-Derkinderen, M. Arock, and M. Dy. 2002. Trends inhistamine research: new functions during immune responses and hematopoiesis.Trends Immunol. 23: 255–263.
14. Jutel, M., T. Watanabe, M. Akdis, K. Blaser, and C. A. Akdis. 2002. Immuneregulation by histamine opinion. Curr. Opin. Immunol. 14: 735–740.
15. Tanaka, S., K. Dear, A. Konomi, K. Takahashi, H. Yamame, and Y. Sugimoto.2004. Expression of L-histidine decarboxylase in granules of elicited mouse poly-morphonuclear leukocytes. Eur. J. Immunol. 34: 1472–1482.
16. Takamatsu, S., I. Nakashima, and K. Nakano. 1996. Modulation of endotoxin-induced histamine synthesis by cytokines in mouse bone marrow-derived mac-rophages. J. Immunol. 156: 778–785.
17. Kubo, Y., and K. Nakano. 1999. Regulation of histamine synthesis in mouseCD4� and CD8� T lymphocytes. Inflamm. Res. 48: 149–153.
18. Jutel, M., K. Blaser, and C. A. Akdis. 2005. Histamine in chronic allergic re-sponses. J. Invest. Allergy Clin. Immunol. 15: 1–8.
19. Szeberenyi, J. B., E. Pallinger, M. Zsinko, Z. Pos, G. Rothe, E. Orso,S. Szeberenyi, G. Schmitz, A. Falus, and V. Lazlo. 2001. Inhibition of effects ofendogenously synthesized histamine disturbs in vitro human dendritic cell dif-ferentiation. Immunol. Lett. 76: 175–182.
20. Gutzmer, R. C., S. Diestel, B. Mommert, H. Koether, H. Stark, M. Witmann, andT. Werfel. 2005. Histamine H4 receptor stimulation suppresses IL-12p70 pro-duction and mediates chemotaxis in human monocyte-derived dendritic cells.J. Immunol. 174: 5224–5232.
3432 HISTAMINE STIMULATES CROSS-PRESENTATION BY DCs
by guest on August 20, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

21. Caron, G., Y. Delneste, E. Roelants, C. Duez, J.-Y. Bonnefoy, J. Pestel, andP. Jeannin. 2001. Histamine polarizes human dendritic cells into Th2 cell-pro-moting effector dendritic cells. J. Immunol. 167: 3682–3686.
22. Dunford, P. J., N. O. O’Donnell, J. P. Riley, K. N. Williams, L. Karlsson, andR. L. Thurmond. 2006. The histamine H4 receptor mediates allergic airway in-flammation by regulating the activation of CD4� T cells. J. Immunol. 176:7062–7070.
23. Gelfand, E. W., and A. Dakhama. 2006. CD8� T lymphocytes and leukotrieneB4: novel interactions in the persistence and progression of asthma.J. Allergy Clin. Immunol. 117: 577–582.
24. Miyahara, N., K. Takeda, T. Kodama, A. Joetham, C. Taube, J. W. Park,S. Miyahara, A. Balhorn, A. Dakhama, and E. W. Gelfand. 2004. Contribution ofantigen-primed CD8� T cells to the development of airway hyperresponsivenessand inflammation is associated with IL-13. J. Immunol. 172: 2549–2558.
25. Miyahara, N., K. Takeda, S. Miyahara, C. Taube, A. Joetham, T. Koya,S. Matsubara, A. Dakhama, A. M. Tager, A. D. Luster, and E. W. Gelfand. 2005.Leukotriene B4 receptor-1 is essential for allergen-mediated recruitment ofCD8� T cells and airway hyperresponsiveness. J. Immunol. 174: 4789–4784.
26. Inaba, K., M. Inaba, N. Romani, H. Aya, M. Deguchi, S. Ikehara, S. Muramatsu,and R. M. Steinman. 1992. Generation of large numbers of dendritic cells frommouse bone marrow cultures supplemented with granulocyte/macrophage colo-ny-stimulating factor. J. Exp. Med. 176: 1693–1697.
27. Sahlin, S., J. Hed, and I. Rundquist. 1983. Differentiation between attached andingested immune complexes by a fluorescence quenching cytofluorometric assay.J. Immunol. Methods 60: 115–118.
28. Sallusto, F., M. Cella, C. Danieli, and A. Lanzavecchia. 1995. Dendritic cells usemacropynocitosis and the mannose receptor to concentrate macromolecules in themajor histocompatibility complex class II compartment: down-regulation by cy-tokines and bacterial products. J. Exp. Med. 182: 389–393.
29. Matsuyama, K., T. Ichikawa, Y. Nitta, Y. Ikoma, K. Ishimura, S. Horio, andH. Fukui. 2006. Localized expression of histamine H1 receptors in syncytiotro-phoblast cells of human placenta. J. Pharmacol. Sci. 102: 331–337.
30. Monczor, F., N. Fernandez, B. L. Legnazzi, M. E. Riveiro, A. Baldi, C. Shayo,and C. Davio. 2003. Tiotidine, a histamine H2 receptor inverse agonist that bindswith high affinity to an inactive G-protein-coupled form of the receptor: exper-imental support for the cubic ternary complex model. Mol. Pharmacol. 64:512–520.
31. Coligan, J. E., A. M. Kruibeek, D. H. Margulies, E. M. Shevach, and W. Strober.1994. Morphological and biochemical assays of apoptosis. In Current Protocolsin Immunology. J. E. Coligan, ed. Wiley, New York, p. 3.17.
32. Akdis, C. A., and F. E. R. Simons. 2006. Histamine receptors are hot in immu-nopharmacology. Eur. J. Pharmacol. 533: 69–76.
33. Chen, J., C. Liu, and W. T. W. Lovenberg. 2003. Molecular and pharmacologicalcharacterization of the mouse histamine H3 receptor. Eur. J. Pharmacol. 467:57–65.
34. Liu, C., S. J. Wilson, C. Kuei, and T. W. Lovenberg. 2001. Comparison ofhuman, mouse, rat, and guinea pig histamine H4 receptors reveals substantialpharmacological species variation. J. Pharmacol. Exp. Ther. 299: 121–130.
35. Pavlinkova, G., Y. Yanagawa, K. Kikuchi, K. Iwabuchi, and K. Onoe. 2003.Effects of histamine on functional maturation of dendritic cells. Immunobiology207: 315–325.
36. Guermonprez, P., J. Valladeau, L. Zitvogel, C. Thery, and S. Amigorena. 2002.Antigen presentation and T cell stimulation by dendritic cells. Annu. Rev. Immu-nol. 20: 621–667.
37. Banchereau, J., F. Briere, C. Caux, J. Davoust, S. Lebecque, Y. Liu,B. Pulendram, and K. Palucka. 2000. Immunobiology of dendritic cells. Annu.Rev. Immunol. 18: 767–811.
38. Burgdorf, S., V. Lukacs-Kornek, and C. Kurtz. 2006. The mannose receptormediates uptake of soluble but not of cell-associated antigen for cross-presenta-tion. J. Immunol. 176: 6770–6776.
39. Thery, C., and S. Amigorena. 2001. The cell biology of antigen presentation indendritic cells. Curr. Opin. Immunol. 13: 43–48.
40. Terebiznik, M. R., O. V. Viera, S. L. Marcus, A. Slade, C. M. Yip, W. S. Trimble,T. Mayer, B. B. Finlay, and S. Grinstein. 2002. Elimination of host cellPtdIns(4,5)P(2) by bacterial SigD promotes membrane fission during invasion bySalmonella. Nat. Cell. Biol. 4: 766–782.
41. Araki, N., N. T. Jonson, and S. J. Swanson. 1996. A role for phosphoinositide3-kinase in the completion of macropinocytosis and phagocytosis by macro-phages. J. Cell Biol. 135: 1249–1252.
42. Gantner, F., K. Sakai, M. W. Tusche, W. W. Cruikshank, D. M. Center, andK. B. Bacon. 2002. Histamine H4 and H2 receptors control histamine-inducedinterleukin-16 release from human CD8� T cells. J. Pharmacol. Exp. Ther. 303:300–307.
43. Kubota, T., H. Fujiwara, T. Ueda, T. Itoh, T. Yamashita, T. Yoshimura,K. Okugawa, Y. Yano, and H. Yamagishi. 2002. Cimetidine modulates the an-tigen presenting capacity of dendritic cells from colorectal cancer patients. Br. J.Cancer 86: 1257–1261.
44. Idzko, M., A. Sala, D. Ferrari, E. Panther, Y. Herouy, S. Dichmann,M. Mockenhaupt, F. Di Virgilio, G. Girolomoni, and J. Norgauer. 2002. Expres-sion and function of histamine receptors in human monocyte-derived dendriticcells. J. Allergy Clin. Immunol. 109: 839–846.
45. Caron, G., Y. Delneste, E. Roelandts, C. Duez, N. Herbault, G. Magistrelli, J.-Y.Bonnefoy, J. Pestel, and P. Jeannin. 2001. Histamine induces CD86 expressionand chemokine production by human immature dendritic cells. J. Immunol. 166:6000–6006.
46. Freigang, S., D. Egger, K. Bienz, H. Hengartner, and R. M. Zinkernagel. 2003.Endogenous neosynthesis vs cross-presentation of viral antigens for cytotoxic Tcell priming. Proc. Natl. Acad. Sci. USA 100: 13477–13482.
47. Ackerman, A. L., C. Kyritsis, R. Tampe, and P. Cresswell. 2005. Access ofsoluble antigens to the endoplasmic reticulum can explain cross-presentation bydendritic cells. Nat. Immunol. 6: 107–113.
48. Schnorrer, P., G. M. Behrens, N. S. Wilson, J. L. Pooley, C. M. Smith,D. El-Sukkari, G. Davey, F. Kupresanin, M. Li, E. Maraskovsky, et al. 2006. Thedominant role of CD8� dendritic cells in cross-presentation is not dictated byantigen capture. Proc. Natl. Acad. Sci. USA 103: 10729–10734.
49. Igaz, P., I. Novak, E. Lazar, B. Horvath, E. Heninger, and A. Falus. 2000. Bi-directional communication between histamine and cytokines. Inflamm. Res. 50:123–128.
50. Tanaka, S., and A. Ichikawa. 2006. Recent advances in molecular pharmacologyof the histamine systems: immune regulatory roles of histamine produced byleukocytes. J. Pharmacol. Sci. 101: 19–23.
51. Mazzoni, A., H. A. Young, J. H. Spitzer, A. Visitin, and D. M. Segal. 2001.Histamine regulates cytokine production in maturating dendritic cells, resulting inaltered T cell polarization. J. Clin. Invest. 108: 1865–1873.
52. Szeberenyi, J. B., B. Lazlo, E. Pallinger, E. Orso, G. Rothe, G. Schmitz, andA. Falus. 2001. Intracellular histamine content increases during in vitro dendriticcell differentiation. Inflamm. Res. 2: S112–S113.
53. Vermeulen, E. M., A. S. Trevani, R. Gamberale, D. Martınez, A. Cevallos,J. Sabatte, M. Giordano, and J. R. Geffner. 2004. The impact of extracellularacidosis on dendritic cell function. Crit. Rev. Immunol. 24: 363–384.
54. Sirois, J., G. Menard, A. S. Moses, and E. Y. Bissonnette. 2000. Importance ofhistamine in the cytokine network in the lung through H2 and H3 receptors:stimulation of IL-10 production. J. Immunol. 164: 2964–2970.
55. Calhoun, W. J., B. J. Lavins, R. Minkwittz, G. J. Evans, J. Gleich, and J. Cohn.1998. Effect of Zafirlukast (Accolate) on cellular mediators of inflammation.Am. J. Respir. Crit. Care Med. 157: 1381–1389.
56. Hannon, J. P., B. Tigani, I. Williams, L. Mazzoni, and J. R. Fozard. 2001. Mech-anism of airway hyperresponsiveness to adenosine induced by allergen challengein actively sensitized Brown Norway rats. Br. J. Pharmacol. 132: 1509–1523.
57. Rock, K. L., and L. Shen. 2005. Cross-presentation: mechanisms and immuno-biological significance. Immunol. Rev. 207: 166–183.
58. Heath, W. R., and F. R. Carbone. 2001. Cross-presentation in viral immunity andself-tolerance. Nat. Rev. Immunol. 1: 126–135.
59. Kurts, C., H. Kosaka, F. R. Carbone, J. F. Miller, and W. R. Heath. 1997. ClassI-restricted presentation of exogenous self-antigens leads to deletion of autore-active CD8� T cells. J. Exp. Med. 186: 239–245.
60. Miyahara, N., K. Takeda, T. Kodama, A. Joetham, C. Taube, J. W. Park,S. Miyahara, A. Balhorn, A. Dakhama, and E. W. Gelfand. 2004. Contribution ofantigen-primed CD8� T cells to development of airway hyperresponsiveness andinflammation is associated with IL-13. J. Immunol. 172: 2549–2558.
3433The Journal of Immunology
by guest on August 20, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















