
Liquid overlaying improves somatic embryogenesis in Catharanthus roseus
10
ORIGINAL PAPER Liquid overlaying improves somatic embryogenesis in Catharanthus roseus Zahid H. Siddiqui • A. Mujib • Mehpara Maqsood Received: 31 May 2010 / Accepted: 16 August 2010 / Published online: 27 August 2010 Ó Springer Science+Business Media B.V. 2010 Abstract Somatic embryos were induced from in vitro germinating seed-hypocotyls of Catharanthus roseus. The process of embryogenesis has been categorized into a few distinct stages (induction, proliferation, maturation and germination) in which liquid overlaying at varying levels 0 ml (T 0 ), 0.25 ml (T 1 ), 0.5 ml (T 2 ) , 0.75 ml (T 3 ) and 1.0 ml (T 4 ) was applied on solid medium. It was found that liquid overlaying improved proliferation; maturation, ger- mination of embryos in C. roseus. In proliferation stage, particularly in T 2 , torpedo embryo number increased sig- nificantly (i.e. 129.6%) as compared to control. Liquid overlaying (T 2 ,T 3 and T 4 ) also improved embryo matu- ration and showed early germination even in maturation medium. It also accelerated normal embryo germination frequency particularly in treatment with T 2 and shortened ‘embryo—plantlet’ recovery time. Biochemical analyses revealed more proline, protein and amino acid with increasing level of liquid overlaying as it improved embryo induction, development and faster germination. Keywords Somatic Embryogenesis Liquid overlaying Catharanthus roseus Abbreviations ANOVA Analysis of variance BA N 6 -Benzyladenine 2,4-D 2,4-Dichlorophenoxyacetic acid GA 3 Gibberellic acid KIN Kinetin NAA Naphthalene acetic acid MS Murashige and Skoog (1962) medium TCA Trichloroacetic acid Introduction The family Apocynaceae consists of several genera, and Catharanthus roseus is one of the important members of this family. It contains more than 130 alkaloids (Van der Heijden et al. 2004), out of which vincristine and vin- blastine are very precious (Miura et al. 1988; Moreno et al. 1995). Both these compounds are well known anticancer drugs and are administered against different kinds of car- cinomas (leukemia, Hodgkin’s disease, Wilm tumor, neu- roblastoma, rhabdosarcoma and reticulum cell sarcoma). Beside antileukemic properties, the total alkaloids and chloroform fractions of crude drug also showed sedative and tranquilizing properties (Wealth of India 1992). C. roseus is also a well known ornamental plant with pink and white flowers, it is commonly known as Madagascar periwinkle or ‘‘Sadabahar’’. C. roseus has been studied extensively for several years in order to synthesize anti- cancerous compounds in culture and to improve the yield particularly vincristine and vinblastine content. (Moreno et al. 1995; Mujib et al. 2003). Somatic embryogenesis (SE) is an important biological process in which embryos are formed in culture. Although this in vitro embryogenesis is known for quite some times, the technique is relatively new in this genus and was described recently (Junaid et al. 2006). Various novel and innovative ideas like optimization of plant growth regulator levels, adjustment of osmotic conditions by using various sugars (sucrose, mannitol, glucose, sorbitol), gelling agents, media condition (solid, liquid, immersion, liquid Z. H. Siddiqui A. Mujib (&) M. Maqsood Cellular Differentiation and Molecular Genetics Section, Department of Botany, Hamdard University, New Delhi 110062, India e-mail: [email protected] 123 Plant Cell Tiss Organ Cult (2011) 104:247–256 DOI 10.1007/s11240-010-9828-z
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Liquid overlaying improves somatic embryogenesis in Catharanthus roseus

ORIGINAL PAPER
Liquid overlaying improves somatic embryogenesisin Catharanthus roseus
Zahid H. Siddiqui • A. Mujib • Mehpara Maqsood
Received: 31 May 2010 / Accepted: 16 August 2010 / Published online: 27 August 2010
� Springer Science+Business Media B.V. 2010
Abstract Somatic embryos were induced from in vitro
germinating seed-hypocotyls of Catharanthus roseus. The
process of embryogenesis has been categorized into a few
distinct stages (induction, proliferation, maturation and
germination) in which liquid overlaying at varying levels
0 ml (T0), 0.25 ml (T1), 0.5 ml (T2), 0.75 ml (T3) and
1.0 ml (T4) was applied on solid medium. It was found that
liquid overlaying improved proliferation; maturation, ger-
mination of embryos in C. roseus. In proliferation stage,
particularly in T2, torpedo embryo number increased sig-
nificantly (i.e. 129.6%) as compared to control. Liquid
overlaying (T2, T3 and T4) also improved embryo matu-
ration and showed early germination even in maturation
medium. It also accelerated normal embryo germination
frequency particularly in treatment with T2 and shortened
‘embryo—plantlet’ recovery time. Biochemical analyses
revealed more proline, protein and amino acid with
increasing level of liquid overlaying as it improved embryo
induction, development and faster germination.
Keywords Somatic Embryogenesis � Liquid overlaying �Catharanthus roseus
Abbreviations
ANOVA Analysis of variance
BA N6-Benzyladenine
2,4-D 2,4-Dichlorophenoxyacetic acid
GA3 Gibberellic acid
KIN Kinetin
NAA Naphthalene acetic acid
MS Murashige and Skoog (1962) medium
TCA Trichloroacetic acid
Introduction
The family Apocynaceae consists of several genera, and
Catharanthus roseus is one of the important members of
this family. It contains more than 130 alkaloids (Van der
Heijden et al. 2004), out of which vincristine and vin-
blastine are very precious (Miura et al. 1988; Moreno et al.
1995). Both these compounds are well known anticancer
drugs and are administered against different kinds of car-
cinomas (leukemia, Hodgkin’s disease, Wilm tumor, neu-
roblastoma, rhabdosarcoma and reticulum cell sarcoma).
Beside antileukemic properties, the total alkaloids and
chloroform fractions of crude drug also showed sedative
and tranquilizing properties (Wealth of India 1992).
C. roseus is also a well known ornamental plant with pink
and white flowers, it is commonly known as Madagascar
periwinkle or ‘‘Sadabahar’’. C. roseus has been studied
extensively for several years in order to synthesize anti-
cancerous compounds in culture and to improve the yield
particularly vincristine and vinblastine content. (Moreno
et al. 1995; Mujib et al. 2003).
Somatic embryogenesis (SE) is an important biological
process in which embryos are formed in culture. Although
this in vitro embryogenesis is known for quite some times,
the technique is relatively new in this genus and was
described recently (Junaid et al. 2006). Various novel and
innovative ideas like optimization of plant growth regulator
levels, adjustment of osmotic conditions by using various
sugars (sucrose, mannitol, glucose, sorbitol), gelling
agents, media condition (solid, liquid, immersion, liquid
Z. H. Siddiqui � A. Mujib (&) � M. Maqsood
Cellular Differentiation and Molecular Genetics Section,
Department of Botany, Hamdard University,
New Delhi 110062, India
e-mail: [email protected]
123
Plant Cell Tiss Organ Cult (2011) 104:247–256
DOI 10.1007/s11240-010-9828-z

overlaying) have recently been trialed to improve the
embryogenesis efficiency in plants (Te-chato and Lim 1999;
Pullman and Skryabina 2007). Liquid overlaying can be
defined as a method in which liquid medium is put over solid
medium. Depending upon the need and requirement, dif-
ferent workers overlaid liquid medium which may be of
similar composition as that of solid medium as was observed
in Hevea brasiliensis (Sushamakumari et al. 2000), half-
strength liquid Murashige and Skoog (1962) medium (MS
medium) with plant growth regulators, used in Garcinia
mangostana (Te-chato and Lim 1999), in Anthurium, med-
ium without plant growth regulators was also added
(Nitayadatpat and Te-chato 2005). Pullman and Skryabina
(2007) used liquid overlaying treatment in medium 1,081 or
its modification with varying concentrations of NAA.
Anthony et al. (1995) reported using B5 medium as solid and
ammonium-free liquid MS medium for overlaying during
the protoplast culture of Mannihot esculenta. Mineral oil
overlaying was also applied by Mathur (1991) in Selenium
candolii which showed increased somatic embryo numbers
per gram of fresh weight. In Pseudotsuga menziesii,
embryogenic tissue initiation was successfully made by
gelled medium or in a gelled-liquid medium overlay system
(Pullman et al. 2009). Volume of liquid overlaying was,
however, varied in different plants, in Pinus taeda 0.25 to
0.75 ml was added (Pullman and Skryabina 2007) but in
Garcinia mangostana cultures, 10–15 ml of half- strength
liquid MS medium was used (Te-chato and Lim 1999). To
improve the selection of citrus transformation, a liquid
overlay system was used which decreased the growth of GUS
negative- shoots without inducing Agrobacterium over-
growth (Gutierrez-E et al. 1997; Yang et al. 2000). Liquid
overlaying also brings down the cost of plant production was
reported by Maene and Debergh (1985); temporary immer-
sion, another related technique involving liquid was earlier
estimated to reduce the cost by about 50–60% (Sluis 2006).
Gupta and Timmis (2005) noted a faster rate of growth in
liquid medium compared with gelled medium, where culture
growth was slow.
However, liquid medium also has a few limitations; the
transfer of the tissue to a semi-solid medium is often
essential at later stages, as continuous agitation hinders
plants’ development and induces vitrification, which is
considered to be a physiological disorder (Etienne et al.
1997). Semi-solid medium has some other disadvantages
too; some of the agars used in culture contain inhibitory
substances which prevent in vitro morphogenesis, and rates
of growth can be slow as the toxic exudates from explants
do not diffuse away quickly (Powell and Uhrig 1987). In
solid medium, explants are implanted above the medium
and do not require special means of aeration to proliferate,
shoots and roots grow in a more orderly fashion in a semi-
solid culture, even explants may be retrieved safely if not
responded in culture. Liquid overlay system in gelled
medium wisely utilizes the advantages of solid and liquid
medium and eliminates the disadvantages present in the
two systems. In this present communication, the impor-
tance of liquid overlaying in C. roseus in vitro embryogeny
has been analyzed and discussed.
Materials and methods
In vitro germination of seeds and culture condition
The basic process of somatic embryogenesis that was
described earlier in Catharanthus roseus cv. Nirmal (Junaid
et al. 2006) was followed. In short, seeds were collected from
in vivo grown plants of Jamia Hamdard (Hamdard Univer-
sity) herbal garden, which is located within the University
campus. Twelve to twenty-five surface disinfected seeds
were placed in Magenta 7 vessels containing 50 ml of MS
solid medium (Murashige and Skoog 1962) without any
growth regulator. Germinated seedlings were grown until
they had attained 2–4 cm length. Various parts (nodal stem,
leaf, root, and hypocotyl) were used and inoculated in test
tubes (Borosil, India) as explants. For embryogenic callus
initiation, MS medium was supplemented with 4.52 lM 2,4-
Dichlorophenoxyacetic acid (2,4-D). For somatic embryo
proliferation, medium was amended with 6.72 lM N6-
Benzyladenine (BA) and 5.37 lM Naphthalene acetic acid
(NAA); embryo maturation and germination medium con-
tained earlier optimized 2.60 lM Gibberellic acid (GA3) and
2.24 lM BA, respectively (Junaid et al. 2006). The pH of the
medium was adjusted to 5.6 before autoclaving at 121�C for
20 min. All cultures were incubated under a 16 h photope-
riod by cool white (Fluorescent F40 T12/CW/EG) lamp at a
photon flux density of 100 lmol m-2 s-1 at 25�C light/20�C
dark temperature.
Embryogenesis has been categorized into following
distinct stages induction and proliferation; maturation; and
germination or plantlet conversion. Liquid overlaying of
four different volumes i.e. 0.25 ml (T1), 0.50 ml (T2),
0.75 ml (T3) and 1.0 ml (T4) was applied to tissues with an
average area of 30-40 sq.mm and a control (T0) was also
maintained for the comparative study. The composition of
liquid medium is the same as that of MS solid medium,
which was used throughout the developmental stages of
embryogenesis. Morphogenetic and biochemical studies
were conducted after 4 weeks of culture. For estimation of
fresh—and dry weight, 4 weeks old calli were used. For
estimation of fresh weight, calli were weighed immediately
after isolation of culture media. Calli were dried at 60�C
for 18 h, their dry weights were measured and finally the
absolute dry mass was calculated using the following for-
mula (Winkelmann et al. 2004).
248 Plant Cell Tiss Organ Cult (2011) 104:247–256
123

Absolutedry mass %ð Þ ¼ Dry weight=Fresh weight� 100:
Estimation of total sugar
Estimation of total sugar was made according to Dey
(1990). Different stages of developing embryos (0.5 g)
were extracted twice with 90% ethanol, and the extracts
were pooled. The final volume of the pooled extract was
made up to 25 ml with double distilled water. To an aliquot
of 1.0 ml, 1.0 ml 5% phenol and 5.0 ml of concentrated
analytical-grade sulphuric acid were added, and cooled in
air. The optical density was measured at 485 nm. A solu-
tion containing 1.5 ml of 55% glycerol, 0.5 ml ninhydrin
and 4.0 ml double distilled water was used as a calibration
standard.
Estimation of proline
For measurement of proline content, 0.2 g of specific
stages of embryos were homogenized in 5 ml 3% aqueous
sulfosalicylic acid and filtered by Whatman filter paper
(No. 1). To 1 ml extract, 1 ml acid ninhydrin and 1 ml of
glacial acetic acid were added and the reaction mixture
incubated at 100�C for 1 h. The reaction mixture was
placed on ice and extracted with 2 ml toluene. Proline
content in the extract was subjected to the spectrophoto-
metric assay of Bates et al. (1973).
Estimation of protein
Protein was estimated by Bradford method (1976), 0.5 g
tissue was ground in a pre-cooled mortar and pestle with
1.5 ml (0.1 M) phosphate buffer (pH 7.0), placed on ice
and centrifuged at 5,000 rpm for 10 min. With 0.5 ml
Trichloroacetic acid (TCA), the sample was again cen-
trifuged at 5,000 rpm for 10 min. The supernatant was
discarded, and the pellet was washed with chilled acetone
and dissolved in 1.0 ml of 0.1 N Sodium hydroxide
(NaOH). Then 0.5 ml aliquot was added with 5.0 ml of
Bradford reagent, the optical density was measured at
595 nm.
Estimation of free amino acids
Free amino acids were estimated by the method developed
by Lee and Takahashi (1966). In short, 0.1 g tissue was
incubated overnight in 70% ethanol followed by washing
with double distilled water. Then 1.5 ml of 55% glycerol
and 0.5 ml ninhydrin solution were added, boiled at 100�C
for 20 min and cooled down. The final volume was made
up to 6 ml with double distilled water, and the optical
density was measured at 570 nm.
Statistical analysis
The data on the effects of plant growth regulators and
liquid overlaying on different stages of embryo and other
biochemical parameters were analyzed by one-way analy-
sis of variance (ANOVAs). Values are means of five rep-
licates from two experiments, and the presented mean
values were separated using Duncan’s Multiple Range Test
(DMRT) at P B 0.05.
Results
Overlaying improves induction of embryogenic tissue
The hypocotyls of in vitro grown plantlet were placed in
medium containing 4.52 lM 2,4-D, callus was friable, light
yellow and fast growing; and proved to be embryogenic.
Calli from other sources were non-embryogenic, therefore
was discontinued in future sets of experiments. Hypocotyl-
calli was routinely maintained on the same medium or in
combination with cytokinins, BA 2.22–8.9 lM or KIN
2.32–9.30 lM. We observed that in liquid overlaying
condition the embryogenic callus was more faster com-
pared to control (Fig. 1); the calli appeared to be more
friable and white (Fig. 1b) specially in treatment with T1
and T2. Higher volumes of liquid overlaying i.e. T3 and T4
(Fig. 1c) were less responsive, callus became compact and
light brown and showed poor growth.
Overlaying induced somatic embryo proliferation
Embryogenic cultures with developing embryos were
placed on medium supplemented with 5.37 lM (NAA) and
6.72 lM BA with different volumes of liquid overlaying
(Fig. 2a, b, c). It was found that the embryogenic calli
differentiated into embryos, and maximum number of
embryos was formed in T2 with an increase value of
23.89% (Table 1) and in the same treatment, the number
of torpedo embryo (129.61%) was more as compared to
control. After T2, in T1 treatment, the number of embryo
was also improved with an increase value of 14.17%, in T3,
the number of cotyledonary stage embryo was increased
with a value of 80.74%. With different volumes of liquid
overlaying, the dry weight and absolute dry mass level
increased, specially in treatment with T2 (Table 2); fresh
weight, however, was decreased.
Overlaying improves embryo maturation
White cotyledonary somatic embryos (5–6 mm) were
placed on medium supplemented with 2.60 lM GA3 with
different volumes of liquid overlaying. It was found that in
Plant Cell Tiss Organ Cult (2011) 104:247–256 249
123
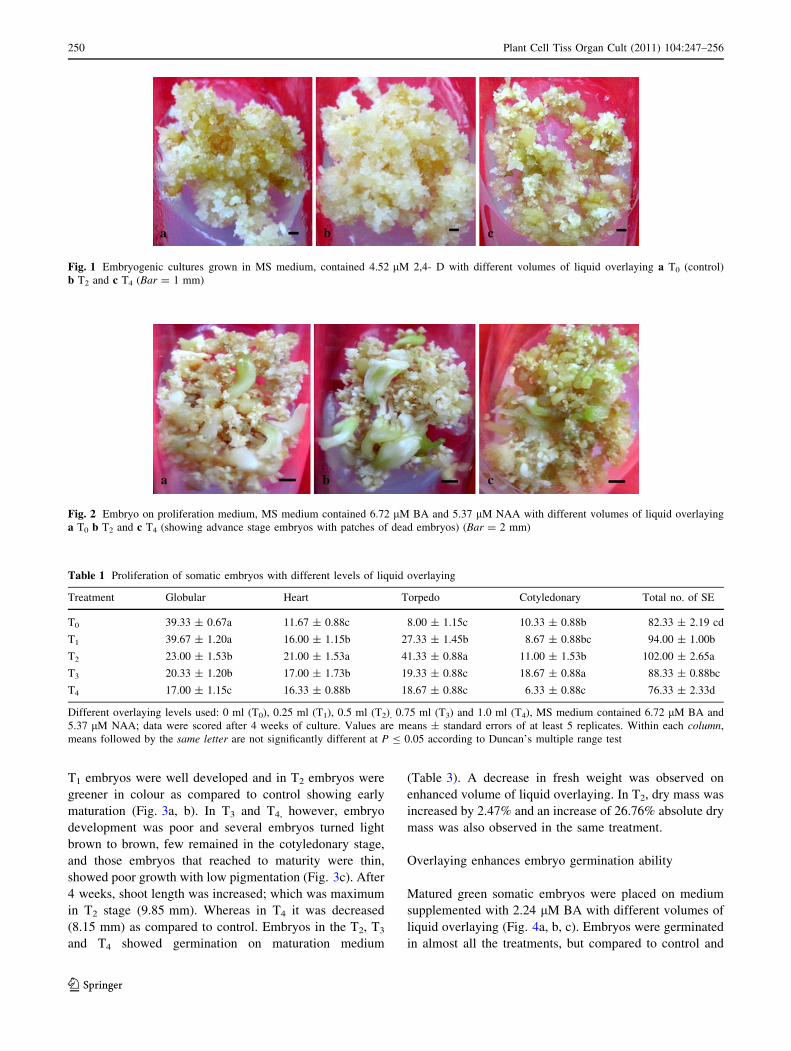
T1 embryos were well developed and in T2 embryos were
greener in colour as compared to control showing early
maturation (Fig. 3a, b). In T3 and T4, however, embryo
development was poor and several embryos turned light
brown to brown, few remained in the cotyledonary stage,
and those embryos that reached to maturity were thin,
showed poor growth with low pigmentation (Fig. 3c). After
4 weeks, shoot length was increased; which was maximum
in T2 stage (9.85 mm). Whereas in T4 it was decreased
(8.15 mm) as compared to control. Embryos in the T2, T3
and T4 showed germination on maturation medium
(Table 3). A decrease in fresh weight was observed on
enhanced volume of liquid overlaying. In T2, dry mass was
increased by 2.47% and an increase of 26.76% absolute dry
mass was also observed in the same treatment.
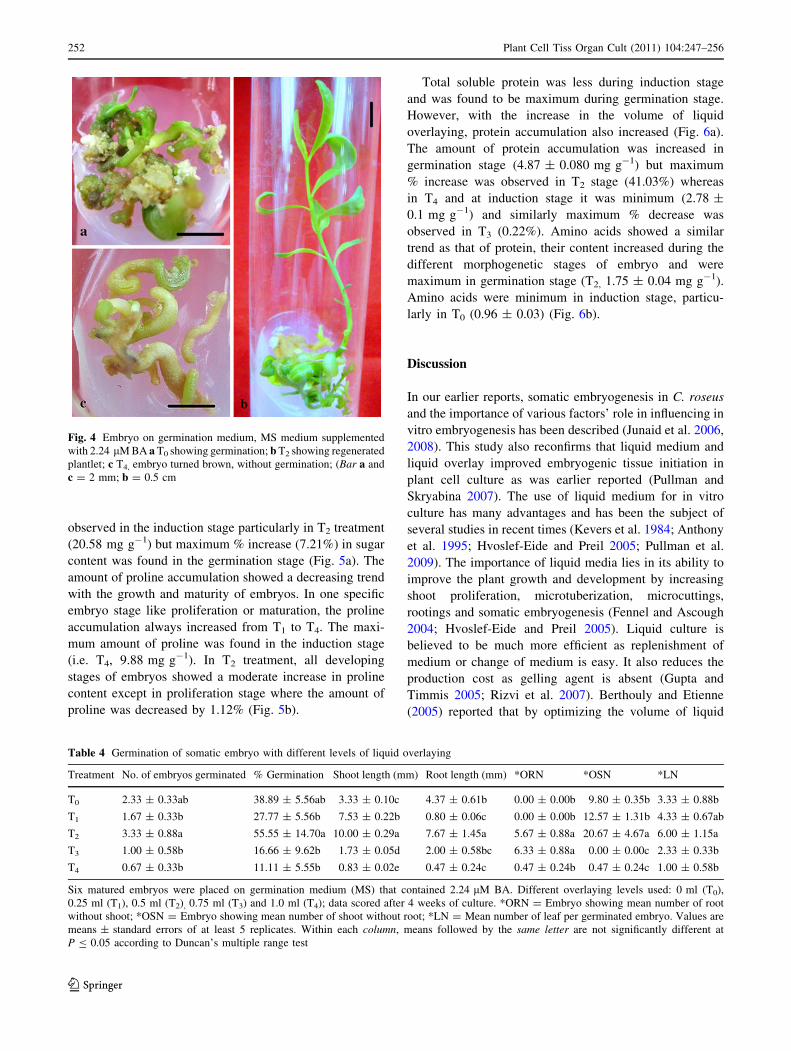
Overlaying enhances embryo germination ability
Matured green somatic embryos were placed on medium
supplemented with 2.24 lM BA with different volumes of
liquid overlaying (Fig. 4a, b, c). Embryos were germinated
in almost all the treatments, but compared to control and
Fig. 1 Embryogenic cultures grown in MS medium, contained 4.52 lM 2,4- D with different volumes of liquid overlaying a T0 (control)
b T2 and c T4 (Bar = 1 mm)
Fig. 2 Embryo on proliferation medium, MS medium contained 6.72 lM BA and 5.37 lM NAA with different volumes of liquid overlaying
a T0 b T2 and c T4 (showing advance stage embryos with patches of dead embryos) (Bar = 2 mm)
Table 1 Proliferation of somatic embryos with different levels of liquid overlaying
Treatment Globular Heart Torpedo Cotyledonary Total no. of SE
T0 39.33 ± 0.67a 11.67 ± 0.88c 8.00 ± 1.15c 10.33 ± 0.88b 82.33 ± 2.19 cd
T1 39.67 ± 1.20a 16.00 ± 1.15b 27.33 ± 1.45b 8.67 ± 0.88bc 94.00 ± 1.00b
T2 23.00 ± 1.53b 21.00 ± 1.53a 41.33 ± 0.88a 11.00 ± 1.53b 102.00 ± 2.65a
T3 20.33 ± 1.20b 17.00 ± 1.73b 19.33 ± 0.88c 18.67 ± 0.88a 88.33 ± 0.88bc
T4 17.00 ± 1.15c 16.33 ± 0.88b 18.67 ± 0.88c 6.33 ± 0.88c 76.33 ± 2.33d
Different overlaying levels used: 0 ml (T0), 0.25 ml (T1), 0.5 ml (T2), 0.75 ml (T3) and 1.0 ml (T4), MS medium contained 6.72 lM BA and
5.37 lM NAA; data were scored after 4 weeks of culture. Values are means ± standard errors of at least 5 replicates. Within each column,
means followed by the same letter are not significantly different at P B 0.05 according to Duncan’s multiple range test
250 Plant Cell Tiss Organ Cult (2011) 104:247–256
123

other treatments, T2 showed maximum germination
(55.55%), and in T4 minimum germination was minimum
(11.11%). In T2, there were plantlets showing shoots and
roots, shoots without roots and embryos with roots without
any visible shoot (Table 4). Length of shoot was found
maximum in T2 (10.00 ± 0.29 mm). In T2, fresh weight
increase was 76.15% and absolute dry mass increased by
11.69% in the same treatment.
Biochemical profiles during embryogenesis stages
as influenced by overlaying
Sugar content was observed to be maximum in induction
stage which was decreased during proliferation, maturation
and germination stage of embryos. Opon increasing the
volume of liquid overlaying, sugar level declined in all
growing stages. Maximum amount of sugar was however,
Table 2 Growth indexes of
somatic embryos in
proliferation, maturation and
germination stages with varying
levels of liquid overlaying
Different overlaying levels
used: 0 ml (T0), 0.25 ml (T1),
0.5 ml (T2), 0.75 ml (T3) and
1.0 ml (T4), and data were
scored after 4 weeks of culture.
Values are means ± standard
errors of at least 5 replicates.
Means within a column for each
of proliferation, maturation and
germination with differentletters are significantly different
at P B 0.05 according to
Duncan’s multiple range test
Treatment Fresh weight (g) Dry weight (g) Absolute dry mass (%)
Proliferation T0 1.39 ± 0.13c 0.118 ± 0.011c 8.54 ± 0.56b
T1 1.79 ± 0.11b 0.151 ± 0.011bc 8.61 ± 0.17b
T2 2.68 ± 0.14a 0.282 ± 0.032a 10.47 ± 0.64a
T3 1.97 ± 0.16b 0.183 ± 0.019b 9.26 ± 0.23b
T4 1.18 ± 0.09c 0.101 ± 0.010c 8.38 ± 0.25b
Maturation T0 1.43 ± 0.016c 0.128 ± 0.009c 9.01 ± 0.38a
T1 1.64 ± 0.11c 0.153 ± 0.003c 9.40 ± 0.72a
T2 2.53 ± 0.09a 0.250 ± 0.016a 9.87 ± 0.43a
T3 2.19 ± 0.11b 0.199 ± 0.010b 9.10 ± 0.14a
T4 2.06 ± 0.10b 0.186 ± 0.017b 9.02 ± 0.47a
Germination T0 1.30 ± 0.15b 0.18 ± 0.03b 13.26 ± 0.38ab
T1 1.63 ± 0.10b 0.22 ± 0.02b 13.78 ± 0.40a
T2 2.29 ± 0.26a 0.33 ± 0.03a 14.81 ± 0.43a
T3 1.54 ± 0.10b 0.22 ± 0.02b 14.36 ± 0.41a
T4 1.56 ± 0.08b 0.19 ± 0.00b 12.18 ± 0.35b
Fig. 3 Embryos on maturation
medium, MS medium contained
2.60 lM GA3 with different
volumes of liquid overlaying
a T0 (showing matured green
embryos) b T1 (showing
germination on maturation
medium) and c T4 (brown mass
of dead embryos with a few
mature embryo) (Bar = 5 mm)
Table 3 Maturation of somatic embryos with different levels of liquid overlaying
Treatment Initial length After 2 weeks After 4 weeks Germination (%)
T0 5.67 ± 0.08a 7.74 ± 0.22a 8.81 ± 0.08b 0.00 ± 0.00c
T1 5.56 ± 0.06a 7.01 ± 0.20b 8.80 ± 0.05b 0.00 ± 0.00c
T2 5.68 ± 0.03a 7.88 ± 0.23a 9.85 ± 0.12a 2.67 ± 0.33a
T3 5.73 ± 0.06a 7.00 ± 0.20b 8.16 ± 0.33c 1.33 ± 0.33b
T4 5.70 ± 0.04a 6.63 ± 0.19b 8.15 ± 0.27c 0.33 ± 0.33c
Different overlaying levels used: 0 ml (T0), 0.25 ml (T1), 0.5 ml (T2), 0.75 ml (T3) and 1.0 ml (T4), MS medium contained 2.60 lM GA3, data
scored after 4 weeks of culture. Values are means ± standard errors of at least 5 replicates. Within each column, means followed by the sameletter are not significantly different at P B 0.05 according to Duncan’s multiple range test
Plant Cell Tiss Organ Cult (2011) 104:247–256 251
123
observed in the induction stage particularly in T2 treatment
(20.58 mg g-1) but maximum % increase (7.21%) in sugar
content was found in the germination stage (Fig. 5a). The
amount of proline accumulation showed a decreasing trend
with the growth and maturity of embryos. In one specific
embryo stage like proliferation or maturation, the proline
accumulation always increased from T1 to T4. The maxi-
mum amount of proline was found in the induction stage
(i.e. T4, 9.88 mg g-1). In T2 treatment, all developing
stages of embryos showed a moderate increase in proline
content except in proliferation stage where the amount of
proline was decreased by 1.12% (Fig. 5b).
Total soluble protein was less during induction stage
and was found to be maximum during germination stage.
However, with the increase in the volume of liquid
overlaying, protein accumulation also increased (Fig. 6a).
The amount of protein accumulation was increased in
germination stage (4.87 ± 0.080 mg g-1) but maximum
% increase was observed in T2 stage (41.03%) whereas
in T4 and at induction stage it was minimum (2.78 ±
0.1 mg g-1) and similarly maximum % decrease was
observed in T3 (0.22%). Amino acids showed a similar
trend as that of protein, their content increased during the
different morphogenetic stages of embryo and were
maximum in germination stage (T2, 1.75 ± 0.04 mg g-1).
Amino acids were minimum in induction stage, particu-
larly in T0 (0.96 ± 0.03) (Fig. 6b).
Discussion
In our earlier reports, somatic embryogenesis in C. roseus
and the importance of various factors’ role in influencing in
vitro embryogenesis has been described (Junaid et al. 2006,
2008). This study also reconfirms that liquid medium and
liquid overlay improved embryogenic tissue initiation in
plant cell culture as was earlier reported (Pullman and
Skryabina 2007). The use of liquid medium for in vitro
culture has many advantages and has been the subject of
several studies in recent times (Kevers et al. 1984; Anthony
et al. 1995; Hvoslef-Eide and Preil 2005; Pullman et al.
2009). The importance of liquid media lies in its ability to
improve the plant growth and development by increasing
shoot proliferation, microtuberization, microcuttings,
rootings and somatic embryogenesis (Fennel and Ascough
2004; Hvoslef-Eide and Preil 2005). Liquid culture is
believed to be much more efficient as replenishment of
medium or change of medium is easy. It also reduces the
production cost as gelling agent is absent (Gupta and
Timmis 2005; Rizvi et al. 2007). Berthouly and Etienne
(2005) reported that by optimizing the volume of liquid
Fig. 4 Embryo on germination medium, MS medium supplemented
with 2.24 lM BA a T0 showing germination; b T2 showing regenerated
plantlet; c T4, embryo turned brown, without germination; (Bar a and
c = 2 mm; b = 0.5 cm
Table 4 Germination of somatic embryo with different levels of liquid overlaying
Treatment No. of embryos germinated % Germination Shoot length (mm) Root length (mm) *ORN *OSN *LN
T0 2.33 ± 0.33ab 38.89 ± 5.56ab 3.33 ± 0.10c 4.37 ± 0.61b 0.00 ± 0.00b 9.80 ± 0.35b 3.33 ± 0.88b
T1 1.67 ± 0.33b 27.77 ± 5.56b 7.53 ± 0.22b 0.80 ± 0.06c 0.00 ± 0.00b 12.57 ± 1.31b 4.33 ± 0.67ab
T2 3.33 ± 0.88a 55.55 ± 14.70a 10.00 ± 0.29a 7.67 ± 1.45a 5.67 ± 0.88a 20.67 ± 4.67a 6.00 ± 1.15a
T3 1.00 ± 0.58b 16.66 ± 9.62b 1.73 ± 0.05d 2.00 ± 0.58bc 6.33 ± 0.88a 0.00 ± 0.00c 2.33 ± 0.33b
T4 0.67 ± 0.33b 11.11 ± 5.55b 0.83 ± 0.02e 0.47 ± 0.24c 0.47 ± 0.24b 0.47 ± 0.24c 1.00 ± 0.58b
Six matured embryos were placed on germination medium (MS) that contained 2.24 lM BA. Different overlaying levels used: 0 ml (T0),
0.25 ml (T1), 0.5 ml (T2), 0.75 ml (T3) and 1.0 ml (T4); data scored after 4 weeks of culture. *ORN = Embryo showing mean number of root
without shoot; *OSN = Embryo showing mean number of shoot without root; *LN = Mean number of leaf per germinated embryo. Values are
means ± standard errors of at least 5 replicates. Within each column, means followed by the same letter are not significantly different at
P B 0.05 according to Duncan’s multiple range test
252 Plant Cell Tiss Organ Cult (2011) 104:247–256
123

medium shoot proliferation efficiency improved substan-
tially in plant tissue culture system. In our study, T2 (0.5 ml
of liquid medium) was found to be most effective as
compared to T1, T3 and T4. It has been suggested that
liquid overlaying as temporary immersion, generally
improved plant tissue quality, and also promoted the
number of somatic embryos as well (Berthouly and Etienne
2005). We also observed that the number of embryos
increased significantly in T2 treatment during proliferation
and that the induced embryos were more distinct and
showed fast growth and development. Liquid overlaying in
this case was considered to be temporary because a film of
medium was retained on the tissue, which was earlier
reported not to hinder gaseous exchange. However, within
3–4 days, growing tissues absorbed the nutrient mist that
prevented plant tissue from hyperhydricity (vitrification).
This is a common incidence in tissue culture employing
liquid culture (Kevers et al. 1984; Gasper et al. 1987;
Debergh et al. 1992). In rubber plant, embryogenic pro-
toplasts showed fairly good growth on KPR semi-solid
medium when overlaid with same liquid medium
(Sushamakumari et al. 2000). Similar results were obtained
by Bell and Reed (2002) in Pyrus on solid medium with
two- phase liquid overlay or intermittent liquid immersion
techniques that increased shoot proliferation markedly.
Etienne et al. (1997) reported that somatic embryo devel-
opment was considerably improved when temporary
immersion was done, and frequency of germination was
0
5
10
15
20
25
T0 T1 T2 T3 T4
Su
gar
co
nte
nt
(mg
g-1
fre
sh w
eig
ht)
of
tiss
ue
afte
r 4
wee
ks
0
2
4
6
8
10
12
T0 T1 T2 T3 T4Pro
line
con
ten
t (
mg
g-1
fre
sh w
eig
ht)
of
tiss
ue
afte
r 4
wee
ks
Induction Prolifreation Maturation Germination
Induction Prolifreation Maturation Germination
b
a Fig. 5 (a) and (b) Sugar
content (top) and proline
content (bottom) content at
different stages of
embryogenesis after various
liquid overlaying treatments
Plant Cell Tiss Organ Cult (2011) 104:247–256 253
123

also enhanced by improving polarization of two distinct
cotyledons. In our studied plant material, we observed
that medium added with 2.89 lM GA3 facilitate embryo
maturation but with liquid overlaying the same medium
promoted early embryo germination, thus the use of
liquid overlaying improved embryogenesis by shortening
‘embryo—plantlet’ recovery period which was just over
3 months. Earlier, Pullman and Gupta (1994) reported that
an increased osmolality of media found to be effective for
development and maturation of embryos.
In this study, we found that the sugar content decreased
with increased level of liquid overlaying, being maximum in
induction stage and decreased later on during proliferation,
maturation and germination stages of embryos, which is in
agreement with earlier work (Junaid et al. 2007). The amount
of protein and amino acid gradually increased with the
developing stages of embryo and with the use of increasing
level of liquid overlaying. Association of specific protein
with different developing stages (i.e. induction, development
and germination) of embryos was earlier reported in some
studied materials (Fellers et al. 1997; Roja Rani et al. 2005).
In contrast, the content of proline decreased with the devel-
oping stages of somatic embryo while some recovery in
proline level was noticed with the increasing amount of
liquid overlaying, which might be due to the immersion
effect of the liquid overlaying. The analysis of proline has
often been conducted as to study cellular stress conditions in
plants and during somatic embryogenesis. Recently, Kiran
Ghanti et al. (2009) observed increased and steady proline
accumulation during early embryo development that
declined sharply at later embryo stages and in germination
time. Proline is considered to act as an osmotic balance
agent, a reservoir of nitrogen pool or the source of energy and
NADP?, necessary for the fast growing tissues like embryos
(Kiran Ghanti et al. 2009). How liquid overlaying improves
the performance of the embryos is still not known clearly but
0
1
2
3
4
5
6
T0 T1 T2 T3 T4
Pro
tein
co
nte
nt
(mg
g-1
fre
sh w
eig
ht)
o
f ti
ssu
e af
ter
4 w
eeks
Induction Prolifreation Maturation Germination
Induction Prolifreation Maturation Germination
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
T0 T1 T2 T3 T4
Am
ino
aci
d c
on
ten
t (m
g g
-1 F
resh
wei
gh
t)
of
tiss
ue
afte
r 4
wee
ks
b
a Fig. 6 (a) and (b) Protein
content (top) and Amino acid
content (bottom) content at
different stages of
embryogenesis after various
liquid overlaying treatments
254 Plant Cell Tiss Organ Cult (2011) 104:247–256
123

liquid overlaying has been proved to be very effective in
plant regeneration in other studied plant species (Te-chato
and Lim 1999). It also improved the growth and development
of plants like Anthurium (Nitayadatpat and Te-chato 2005).
We, therefore, believe that the liquid overlaying causes faster
embryogenic growth in C. roseus, which may be due to direct
contact with the medium that facilitate rapid and fast
absorption of nutrients by the growing tissues. Liquid over-
laying provides an excellent tool to adjust the pH, nutritional
and hormonal environments of tissues growing in vitro
(Pullman et al. 2009). Solid medium alone shows several
disadvantages which ultimately reduce growth as in agar-
added medium water potential reduces linearly (Ghashghaie
et al. 1991). Gelling agents also contribute elemental content
to medium (Van Winkle and Pullman 2003) and in some
cases Ca, Mg, S, Mn, Fe and Al are completely bound to agar
(Scholten and Pierik 1998). Similarly, it generates concen-
tration gradients for each nutrient in the zone of gel, next to
cell that slows cellular growth (Gupta and Timmis 2005).
Conclusion
Callus mediated embryogenesis shows a prominent possi-
bility to increase the propagation rate as embryogenic
cultures can easily be developed from fast growing cell
suspensions (Caligari and Shohet 1993). In C. roseus,
embryogeny was further improved by using liquid over-
laying technique where induced embryos showed fast
growth and development. Liquid overlaying also improved
early germination in C. roseus thus shortening the ‘embryo
to plantlet’ recovery time. Liquid overlaying based
embryogenesis may provide a valuable tool to enhance the
pace of genetic improvement. This technique could be
utilized in other potential applications of biotechnology
such as artificial seeds, micropropagation and gemplasm
conservation. The origin of embryos in overlaid condition
may also help in our understanding of the cellular switches
that keep the cell totipotent while undergoing cell division,
growth and reorganization.
Acknowledgments The first author is thankful to Council of Sci-
entific and Industrial Research (CSIR) for receiving Senior Research
Fellowship (SRF). Thanks are also due to Dr. Saumitra Banerjee for
his help with this manuscript.
References
Anthony P, Davey MR, Power JB, Lowe KC (1995) An improved
protocol for the culture of cassava leaf protoplasts. Plant Cell
Tissue Org Cult 42:299–302
Bates L, Waldren PP, Teare JD (1973) Rapid determination of free
proline of water stress studies. Plant Soil 39:205–207
Bell RL, Reed BM (2002) In vitro tissue culture of Pear: advances in
techniques for micropropagation and germplasm preservation.
Acta Hort 596:412–418
Berthouly M, Etienne H (2005) Temporary immersion system: a new
concept for use liquid medium in mass propagation. In: Hvoslef-
Eide AK, Preil W (eds) Liquid culture systems for in vitro plant
propagation. Springer, Dordrecht, pp 165–195
Bradford MM (1976) A rapid and sensitive method for quantification
of microgram quantities of protein, utilizing the principle of
protein dye binding. Anal Biochem 72:248–541
Caligari PDS, Shohet S (1993) Variability in somatic embryos. In:
Redenbaugh K (ed) Synseeds: application of synthetic seeds to
crop improvement. CRC Press, Boca Raton, pp 163–174
Debergh PC, Aitken-Christic J, Cohen B, Von Arnold S, Zimmerman
R, Ziv M (1992) Reconsideration of the term ‘‘vitrification’’ as
used in micropropagation. Plant Cell Tissue Org Cult
30:135–140
Dey PM (1990) Methods in plant biochemistry. Carbohydrates, vol 2.
Academic Press, London
Etienne H, Lartaud M, Michaux-ferriere N, Carron ME, Berthouly M,
Teisson C (1997) Improvement of somatic embryogenesis in
Hevea brasiliensis (MULL. ARG.) Using the temporary immer-
sion technique. In Vitro Cell Dev Biol -Plant 33:81–87
Fellers JP, Guenzi AC, Porter DR (1997) Marker proteins associated
with somatic embryogenesis of wheat callus cultures. J Plant
Physiol 151:201–208
Fennel CW, Ascough GD (2004) The regulation of plant growth and
development in liquid culture. S Afr J Bot 70:181–190
Gasper TH, Kevers C, Debergh P, Maene L, Paques M, Boxus PH
(1987) Vitrification: morphological, physiological and ecologi-
cal aspects. In: Bonga JM, Duran DJ (eds) Cell and tissue culture
in forestry, vol 1. General principles and Biotechnology,
Martinus Nijhoff, Dordrecht, pp 152–166
Ghashghaie F, Brenckmann F, Saugier B (1991) Effects of agar
concentration on water status and growth of rose plants cultured
in vitro. Physiol Plant 82:73–78
Gupta PK, Timmis R (2005) Mass propagation of conifer trees in
liquid cultures—progress towards commercialization. Plant Cell
Tissue Org Cult 81:339–346
Gutierrez-E MA, Luth D, Moore GA (1997) Factors affecting
Agrobacterium-mediated transformation in Citrus and production
of sour orange (Citrus aurantium L.) plants expressing the coat
protein gene of Citrus tristeza virus. Plant Cell Rep 16:745–753
Hvoslef-Eide AK, Preil W (2005) Liquid culture systems for in vitroplant propagation. Springer, Dordrecht
Junaid A, Mujib A, Bhat MA, Sharma MP (2006) Somatic embryo
proliferation, maturation and germination in Catharanthusroseus. Plant Cell Tissue Org Cult 84:325–332
Junaid A, Mujib A, Sharma MP, Samaj J (2007) Somatic embryo-
genesis and plant regeneration in Catharanthus roseus. Biol
Plant 51(4):641–646
Junaid A, Mujib A, Fatima S, Sharma MP (2008) Cultural conditions
affect somatic embryogenesis in Catharanthus roseus L (G.)
Don. Plant Biotechn Rep 2:179–189
Kevers C, Comans M, Coumans-Gilles ME, Gasper TH (1984)
Physiological and biochemical events leading to vitrification in
plant cultured in vitro. Physiol Plantarum 61:69–74
Kiran Ghanti S, Sujata KG, Rao S, Udayakumar M, Kavi Kishor PB
(2009) Role of enzymes and identification of stage-specific
proteins in developing somatic embryos of chickpea (Cicer
arietinum L.). In Vitro Cell Dev Biol Plant 45:667–672
Lee YP, Takahashi T (1966) Improved calorimetric determination of
amino acids with the use of ninhydrin. Anal Biochem 24:71–77
Maene L, Debergh PC (1985) Liquid medium additions to established
tissue cultures to improve elongation and rooting in vivo. Plant
Cell Tissue Org Cult 5:23–33
Plant Cell Tiss Organ Cult (2011) 104:247–256 255
123

Mathur J (1991) Enhanced somatic embryogenesis in Selinumcandolii DC. Under a mineral oil overlay. Plant Cell Tissue
Org Cult 27:23–26
Miura Y, Hirata K, Kurano N, Miyamoto K, Chiida K (1988)
Formation of vinblastine in multiple shoot culture of Catharan-
thus roseus. Planta Med 54:18–20
Moreno PRH, van der Heijden R, Verpoorte HR, ten Hoopen JG
(1995) Cell and tissue culture of Catharanthus roseus: a literature
survey II. Updating from 1988–1993. Plant cell Tissue Org Cult
42:1–25
Mujib A, Ilah A, Gandotra N, Abdin MZ (2003) In vitro application to
improve alkaloids yield in Catharanthus roseus. In: Govil JN,
Kumar PA, Singh VK (eds) Biotechnology and genetic engi-
neering, vol IV. SciTech Publishing, Texas, pp 415–439
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bioassay with tobacco tissue cultures. Physiol Plant 15:473–497
Nitayadatpat R, Te-chato S (2005) Enhanced efficiency for propaga-
tion of anthurium by tissue culture technique. Songklanakarin J
Sci Technol 27(5):1003–1008
Powell W, Uhrig H (1987) Anther culture of Solanum gemotypes.
Plant Cell Tissue Org Cult 11:13–24
Pullman GS, Gupta PK (1994) Method for reproducing conifers by
somatic embryogenesis using mixed growth hormones for
embryo culture. US Patent 5294549
Pullman GS, Skryabina A (2007) Liquid medium and liquid overlays
improve embryogenic tissue initiation in conifers. Plant Cell Rep
26:873–887
Pullman GS, Johnson S, Bucalo K (2009) Douglas fir embryogenic
tissue initiation. Plant Cell Tissue Org Cult 96:75–84
Rizvi MZ, Kukreja AK, Khanuja SPS (2007) I vitro culture of
Chlorophytum borivilianum Sant. et Fernand. in liquid culture
medium as a cost-effective measure. Curr Sci 92(1):87–90
Roja Rani A, Reddy VD, Prakash Babu P, Padmaja G (2005) Changes
in protein profiles associated with somatic embryogenesis in
peanut. Biol Plant 49:347–354
Scholten HJ, Pierik RLM (1998) Agar as a gelling agent: chemical
and physical analysis. Plant Cell Rep 17:230–235
Sluis CJ (2006) Integrating automation technologies with commercial
micropropagation: an economic perspective. In: Gupta SD,
Ibaraki Y (eds) Plant tissue culture engineering. Springer,
Netherlands, pp 231–251
Sushamakumari S, Asokan MP, Anthony P, Lowe KC, Power JB,
Davey MR (2000) Plant regeneration from embryogenic cell
suspension-derived protoplasts of rubber. Plant Cell Tissue Org
Cult 61:81–85
Te-chato S, Lim M (1999) Plant regeneration of mangosteen via
nodular callus formation. Plant Cell Tissue Org Cult 59:89–93
Van Der Heijden R, Jacobs DI, Snoeijer W, Hallared D, Verpoorte R
(2004) The Catharanthus alkaloids: pharmacognosy and bio-
technology. Curr Med Chem 11:607–628
Van Winkle SC, Pullman GS (2003) The combined impact of pH and
activated carbon on the elemental composition of plant tissue
culture media. Plant Cell Rep 22:303–311
Wealth of India (1992) A dictionary of Indian raw materials and
industrial products. Vol-3 (Revised) Ca- Ci. Publication and
information directorate, Council of Scientific and Industrial
Research, New Delhi
Winkelmann T, Mußmann V, Serek M (2004) Cryopreservation of
embryogenic cell suspension cultures of Cyclamen persicumMill. Plant Cell Rep 23:1–8
Yang ZN, Ingelbrecht IL, Louzada E, Skaria M, Mirkov TE (2000)
Agrobacterium-mediated transformation of the commercially
important grapefruit cultivar Rio Red (Citrus paradisi Macf).
Plant Cell Rep 19:1203–1211
256 Plant Cell Tiss Organ Cult (2011) 104:247–256
123




















