
Expression profiling of glial genes during Drosophila embryogenesis
Upload
xn--nibge-zhaCategory
view
1download
0
Plant Cell Rep (2006)DOI 10.1007/s00299-006-0135-4
GENETICS AND GENOMICS
Kim E. Nolan · Nasir A. Saeed · Ray J. Rose
The stress kinase gene MtSK1 in Medicago truncatulawith particular reference to somatic embryogenesis
Received: 16 August 2005 / Revised: 11 November 2005 / Accepted: 14 January 2006C© Springer-Verlag 2006
Abstract Medicago truncatula, a model for legume ge-nomics, can be regenerated by somatic embryogensis bythe use of a suitable genotype and an auxin plus cytokinin.The stress response induced by explant wounding and cul-ture is increasingly recognized as an important componentof somatic embryo induction. We have cloned and investi-gated the stress kinase gene MtSK1 in relation to somaticembryogenesis in M. truncatula, using the highly embryo-genic mutant Jemalong 2HA (2HA) and its progenitor Je-malong. The main features of the MtSK1 protein of 351amino acids are an N-terminal kinase domain and a C-terminal glutamic acid-rich region, which is predicted tobe a coiled-coil. MtSK1 is a member of the SnRK2 sub-group of the SnRK group of plant kinases. Members of theSnRK2 kinases play a role in stress responses of plants.MtSKI expression is induced by wounding in the culturedtissue independent of auxin or cytokinin. However, in both2HA and Jemalong, as the callus develops in response toauxin plus cytokinin, MtSK1 expression continues to in-crease. MtSK1 responds to salt stress in vivo, consistentwith its role as a stress kinase. The likely role of MtSK1in stress-induced signaling will facilitate the relating ofstress–response pathways to auxin and cytokinin-inducedsignaling in the understanding of the molecular mecha-nisms involved in the induction of somatic embryogenesisin M. truncatula.
Communicated by P. Lakshmanan
K. E. Nolan · N. A. Saeed · R. J. Rose (�)Australian Research Council Centre for Integrative LegumeResearch and School of Environmental and Life Sciences,The University of Newcastle, Callaghan,New South Wales, 2308 Australiae-mail: [email protected].: + 61-2-49215711Fax: + 61-2-49216923e-mail: [email protected]: [email protected]
Present address:N. A. SaeedNational Institute for Biotechnology and Genetic Engineering,Faisalabad, Pakistan
Keywords Somatic embryogenesis . SnRK kinase .Medicago truncatula . Salt . Stress . Plant hormones
Introduction
The induction of somatic embryogenesis in cultured tis-sue is a multifactorial event. Induction by plant hormones,particularly auxin, is generally considered to be the mostimportant factor (Rose 2004) and is by far the most widelystudied. Other chemical factors, such as the nutritional com-ponents of the culture medium, and physical factors such aslighting or whether liquid or solid culture medium is used,have also been shown to play a role in some species. An-other factor, which has become increasingly recognized inrecent years, is stress. There is now evidence that the stress–response of cultured tissue plays a major role in somaticembryo induction (Feher et al. 2003; Lee et al. 2001). Thisconcept has been highlighted by reports that stressors alone,in the absence of exogenously supplied hormones can in-duce somatic embryo formation in some species (Kiyosueet al. 1989; Kamada et al. 1993, 1994; Touraev et al. 1997).This view is further reinforced by reports of the stress-induced hormone, abscisic acid (ABA) also playing a rolein somatic embryo induction in M. truncatula (Nolan andRose 1998) and in other species (Nishiwaki et al. 2000).
It is clear that excising tissue from the parent plant, sub-jecting it to sterilization procedures, cutting it into piecesand putting it into an artificial culture environment exertsconsiderable stress on the plant tissue. The genetic regu-lation of the stress response and any interaction betweenstress–response and hormone signaling during the induc-tion of somatic embryogenesis has received virtually noattention to date. In light of the increasing evidence of arole of stress in the induction of somatic embryogenesis,it is necessary to take into account the stress–response ofcultured tissue when investigating the genetic regulation ofsomatic embryogenesis.
During the investigation of genes induced early in so-matic embryogenesis of Medicago truncatula Jemalong2HA, we identified genes that were stimulated by auxin

and cytokinin and those which were hormone independent,and likely stress-induced. For example the expression ofthe important somatic embryogenesis gene MtSERK1 is in-duced by auxin and not induced by cytokinin. However,the auxin stimulation is potentiated by cytokinin (Nolanet al. 2003). In this study we have cloned a gene and in-vestigated its role in relation to somatic embryogenesis inM. truncatula, in order to understand more about signalinginduced by stress. This gene, called MtSK1, is a memberof the class of plant kinases called the SNF1-related kinase(SnRK) group (specifically the SnRK2 subgroup). Mem-bers of the SnRK group of kinases are thought to play arole in stress responses of plants, and some of them areinduced by ABA. The SnRK protein kinases belong to therecently categorized CDPK-SnRK superfamily of proteinkinases (Hrabak et al. 2003). The SnRK group was sonamed because of the similarity of the kinase domain ofthese kinases to the kinase domain of the yeast (Saccha-romyces cerevisiae) SNF1 protein kinase, and to anotherclosely related protein, the mammalian AMP-activated pro-tein kinase (AMPK). Both of these kinases are involved inresponses to cellular stresses. The plant SnRK group is fur-ther subdivided into three subgroups: SnRK1, SnRK2 andSnRK3, with the SnRK2 and SnRK3 subgroups apparentlybeing unique to plants (Hrabak et al. 2003).
Medicago truncatula is a model for legume genomics(Frugoli and Harris 2001) and can be regenerated viasomatic embryogenesis by the use of a suitable genotypeand an auxin plus cytokinin (Nolan et al. 1989; Chabaudet al. 1996; Nolan and Rose 1998; das Neves et al. 1999;Iantcheva et al. 2001). The highly embryogenic seed lineof M. truncatula, Jemalong 2HA (2HA) was developedfrom M. truncatula cv Jemalong, which forms very fewsomatic embryos in culture (Nolan et al. 1989; Rose et al.1999). Therefore, 2HA can be considered to be a nearlyisogenic, highly embryogenic mutant of M. truncatula cvJemalong. The MtSK1 gene has been investigated in both2HA and Jemalong which have differences in their proteincomplement during culture (Imin et al. 2005) and providea good system for investigating organogenesis and somaticembryogenesis in vitro. MtSKI is induced by woundingin cultured tissue. However, after the initial woundingresponse, its expression continues to increase over severalweeks of culture, after which it remains elevated. Thismay indicate a sustained stress–response of cultured tissuethroughout the culture process. It shows similar responsesin both Jemalong and 2HA throughout the culture processand also responds to salt stress in vivo. The relationship ofMtSK1 to other members of the SnRK2 group and its rolein the induction of somatic embryogenesis is examined.
Materials and methods
Tissue culture and RNA extraction
Tissue culture was conducted using M. truncatula plantsfrom either the highly regenerable seed line 2HA (Roseet al. 1999) or M. truncatula cv. Jemalong. Seeds of M.
truncatula cv Jemalong were obtained from the NationalMedicago Collection (Northfield Research Laboratories,Adelaide, South Australia). The culture procedure and themedia used have been described (Nolan et al. 2003). Plantsgrown for salt experiments were grown in standard pottingmix under glasshouse conditions and watered normally un-til 10 cm in height. Salt treatments were then applied bywatering the plants with 50 mM or 250 mM NaCl solu-tion. RNA extractions were carried out using the RNeasy r©
Plant Mini Kit (Qiagen) according to manufacturer’sinstructions.
Obtaining the MtSK1 cDNA sequence
We initially cloned a 406 bp sequence from a band from pre-liminary differential RNA display studies comparing cul-tured Jemalong and 2HA and found a 96% homology withthe Arabidopsis serine/threonine kinase ASK1 (P43291on the NCBI database; http://www.ncbi.nlm.nih.gov/). Afull length cDNA sequence was obtained using 3′- and5′-RACE with DNase-treated RNA from one or 2-weekold callus from 2HA. For 3′-RACE first strand cDNAwas obtained using the 3′-RACE adaptor primer (Invit-rogen) and the Superscript Preamplification System forFirst Strand cDNA synthesis kit (Invitrogen) according tothe manufacturer’s instructions. The gene specific primersMF4 (5′-TTTCTTCCAGCAGCTCATC-3′) and MF5 (5′-GAACCCCAGCCTACATAGCA-3′) were used with theAbridged Universal Amplification Primer (AUAP; Invit-rogen) in nested PCR reactions to obtain the 3′ end.The PCR product obtained was cloned and sequencedas described previously (Nolan et al. 2003). 5′-RACEof MtSK1 was conducted using the 5′-RACE Systemfor Rapid Amplification of cDNA ends, Version 2 (In-vitrogen) as per manufacturer’s instructions. The Re-verse transcription was carried out using the gene-specificprimer MR1 (5′-CCAACTTTCCGTCATATTCTC-3′).The nested gene specific primers used were MR2(5′-GCTGGGGTTCCCACTGTAGA-3′) and MR3 (5′-AAGGTGTTCTCGAGCTGCAAGT-3′). The 5′ RACEproduct was cloned and sequenced as described.
Once the full coding sequence of MtSK1 had beenassembled, the coding region of the gene was ampli-fied in a single PCR reaction using the primers MF1(5′-GCACTGAAAGGTGATGGAGAA-3′) and MR4(5′-GAGAGCGAAGGAAACCAACA-3′) and cDNAfrom 2HA 2-week old calli. The full length cDNAsequence from 3-week-old Jemalong callus tissue wasalso amplified in a single PCR reaction using the MF2(5′-CGATCCGGCGTTGAAATAG-3′ and MR4 primersand sequenced.
Genomic sequencing of MtSK1
Genomic DNA used for sequencing was extracted from2HA leaf tissue. Two PCR reactions gave overlappingfragments which were sequenced. The primers used for

the first half of the sequence were MF2 and MR1, and theprimers for the second half were MF5 and MR4 primers.
Sequence comparisons
Database searches were performed at the NationalCentre for Biotechnology Information (http://www.ncbi.nlm.nih.gov/) using the BLAST network service (Altschulet al. 1997) or at the TIGR Medicago truncatula GeneIndex (MtGI) (http://www.tigr.org/tdb/tgi/mtgi/). Se-quence alignments and comparisons were performed usingClustalW (Thompson et al. 1994) on BioManager byANGIS (http://www.angis.org.au). Protein predictionswere performed using PSORT (http://psort.nibb.ac.jp/) (Nakai and Kanehisa 1992), Scan Prosite (http://www.expasy.org/prosite/) (Hofmann et al. 1999), SMART(http://smart.embl-heidelberg.de/), ChloroP (http://www.cbs.dtu.dk/services/ChloroP/) (Emanuelsson et al.1999), SignalP (http://www.cbs.dtu.dk/services/SignalP/),(Bendtsen et al. 2004) and COILS (http://www.ch.embnet.org/ software/COILS form.html) (Lupas et al. 1991).
Phylogenetic analysis
Phylogenetic analysis of the full-length amino acid se-quences was performed on BioManager by ANGIS usingthe programs ClustalW (Thompson et al. 1994), Seqboot,Prodist, Fitch and Consense (Felsenstein 1989). Trees weredrawn using Tree View, Version 1.6.6 (Page 1996). Se-quences used for the alignment were the MtSK1 proteinsequence, 74 sequences listed as SnRKs on the PlantsPdatabase (http://plantsp.sdsc.edu/) and five other sequenceswhich were not listed on the PlantsP database, but were highscoring hits on a BLASTP search of the NCBI database us-ing the MtSK1 amino acid sequence (see Fig. 1).
MtSK1 expression studies using real time PCR
The relative expression of MtSK1 was measured usingReal Time PCR on the Rotorgene (Corbett Research) usinga TaqMan r© probe and primers (Applied Biosystems).A gene coding for a M. truncatula glyceraldehyde3-phosphate dehydrogenase (MtGAPDH) was used asthe endogenous control. To prevent amplification ofany contaminating genomic DNA, the Taq-Man probesfor each of MtSK1 and MtGAPDH were designedover an exon/intron boundary in the cDNA sequence.Taq-Man probe and primers were designed using PrimerExpress software (Applied Biosystems). The primerand probe sequences used were: MtSK1 probe, 5′-FAM-TGTCATTCCATGCAAATCTGCCATCG-3′; MtSK1forward primer MF6 (5′-GCAGCTCATCTCAGGAGTTAGCTA-3′); MtSK1 reverse primer MR6 (5′-ATAAGGTGTTCTCGAGCTTCAAGTC-3′); MtGAPDH MGBprobe, 5′ VIC-CCACCCTTCAAATGA-3′; MGAPDH for-ward primer 5′-ACAAACATGGGAGCATCCTTACTAG-
3′ and MGAPDH reverse primer 5′-GTTTTTACCGACAAGGACAAAGCT-3′. Real time PCR resultswere analyzed using the Comparative CT method withappropriate validation experiments (Applied Biosystems,User Bulletin #2, http://home.appliedbiosystems.com/).
MtSK1 expression studies using Northern hybridisation
10 µg of each RNA sample was run on a denaturingformaldehyde gel and blotted on to a HybondTM-N + ny-lon membrane (Amersham Pharmacia Biotech). RNA wasfixed to the membrane by baking in an 80◦C oven forone to two hours. The hybridisation probe was preparedby PCR from a 406 bp PCR product from the kinase do-main of the MtSK1 gene which had been cloned into pCRScript Amp SK( + ) cloning vector (Stratagene). This se-quence corresponds to the region between 349 and 755 bpon the MtSK1 mRNA sequence (GenBank accession num-ber AY772417). Probe DNA was labeled with [α32P]dCTPusing the Multiprime DNA labelling system (AmershamPharmacia Biotech) as per the manufacturer’s instructions.Hybridisation was carried out overnight in a bottle (Hy-baid) containing 10 ml hybridisation buffer while rotatingin a mini hybridisation oven (Hybaid) at 42◦C.
The membrane was washed twice for 5 min at roomtemperature in 1 × SSPE, 0.1% w/v SDS solution, twice at50◦C in 0.1 × SSPE, 0.1% w/v SDS solution for 30 mineach wash, then rinsed briefly in 0.1 × SSPE, 0.1% w/vSDS solution and blotted dry on Whatmann 3MM paper.The membrane was wrapped in cling wrap and exposedto Kodak BioMax MS X-ray film using an intensifyingscreen at − 80◦C for 20 min to several days, depending onthe intensity of the signal.
Quantification of Northern hybridisation for graphicalrepresentation
The intensity of bands corresponding to the MtSK1transcript was measured using a GS-700 densitometer(Bio-Rad). For each treatment the band density from threeindependent experiments was measured and the mean andstandard error calculated. The level of expression in leaftissue was used as a standard with the assigned value ofone, and relative expression according to the density ofbands in other tissues expressed in comparison to leaftissue.
Results
The MtSK1 sequence
The full-length cDNA sequence of the coding region ofMtSK1 was obtained by applying 3′-RACE and 5′-RACEto a 406 bp cloned sequence obtained from 2-week cul-tured 2HA tissue. Amplification of the full length MtSK1cDNA confirmed that the sequence obtained was the

* 20 * 40 * 60 MtSK1 : MEKYEVVKDIGSGNFGVARLMRHKDTKQLVAMKYIERGHKIDENVAREIINHRSLRHPNIIRF : 63 OSAK : MDKYELVKDIGSGNFGVARLMRHKETKELVAMKYIERGHKIDENVAREIINHRSLRHPNIIRF : 63 SPK-3 : MDKYEAVKDLGAGNFGVARLMRNKETKELVAMKYIERGQKIDENVAREIINHRSLRHPNIIRF : 63 SPK-4 : MDKYEAVKDLGAGNFGVARLMRNKVTKELVAMKYIERGPKIDENVAREIMNHRSLRHPNIIRY : 63 BSK2 : MEKYELVKDIGAGNFGVARLMKVKNSKELVAMKYIERGPKIDENVAREIINHRSLRHPNIIRF : 63 ASK1 : MDKYELVKDIGAGNFGVARLMKVKNSKELVAMKYIERGPKIDENVAREIINHRSLRHPNIIRF : 63 pk4 : MDKYEVVKDLGAGNFGVARLLRHKETKELVAMKYIERGLKIDENVAREIINHRSLRHPNIIRF : 63 pk5 : MEKYELVKDIGSGNFGVARLMRNKESKELVAMKYIERGRKIDENVAREIINHRSLRHPNIIRF : 63 CsSTK1 : --------------------------------------------------------------- : - m kye vkd g gnfgvarl k k lvamkyierg kidenvarei nhrslrhpniir * 80 * 100 * 120 MtSK1 : KEVVLTPTHLGIVMEYAAGGELFDRICSAGRFSEDEARYFFQQLISGVSYCHSMQICHRDLKL : 126 OSAK : KEVLVTPTHLAIVMEYAAGGELFERICNAGRFSEDEARYFFQQLISGVHYCHNMQICHRDLKL : 126 SPK-3 : KEVVLTPTHLAIVMEYAAGGELFERICNAGRFSEDEARYFFQQLISGVHYCHAMQICHRDLKL : 126 SPK-4 : KEVVLTPTHLAIVMEYAAGGELFERICSAGRFSEDEARYFFQQLISGVHFCHTMQICHRDLKL : 126 BSK2 : KEVVLTPTHLAIAMEYAAGGELFERICSAGRFSEDEARYFFQQLISGVSYCHAMQICHRDLKL : 126 ASK1 : KEVVLTPTHLAIAMEYAAGGELFERICSAGRFSEDEARYFFQQLISGVSYCHAMQICHRDLKL : 126 pk4 : KEVVLTPTHLAIVMEYAAGGELFERICTAGKFSEDEARYFFQQLISGVNYCHSMQICHRDLKL : 126 pk5 : KEVVLTPTHLAIVMEYAAGGELFERICAAGRFSEDEARYFFQQLISGVPYCHSMQICHRDLKL : 126 CsSTK1 : ----------------------------------DEARYFFQQLISGVCYCHSMQICHRNLKL : 29 kev tpthlai meyaaggelferic ag fseDEARYFFQQLISGV 5CH MQICHR1LKL * 140 * 160 * 180 MtSK1 : ENTLLDGSPAPRLKICDFGYSKSSLLHSRPKSTVGTPAYIAPEVLSRREYDGKLADVWSCGVT : 189 OSAK : ENTLLDGSPAPRLKICDFGYSKSSLLHSRPKSTVGTPAYIAPEVLSRREYDGKLADVWSCGVT : 189 SPK-3 : ENTLLDGSPAPRLKICDFGYSKSSLLHSRPKSTVGTPAYIAPEVLSRREYDGKLADVWSCGVT : 189 SPK-4 : ENTLLDGSPAPRLKICDFGYSKSSLLHSRPKSTVGTPAYIAPEVLSRREYDGKLADVWSCAVT : 189 BSK2 : ENTLLDGSPAPRLKICDFGYSKSSLLHSRPKSTVGTPAYIAPEVLSRREYDGKMADVWSCGVT : 189 ASK1 : ENTLLDGSPAPRLKICDFGYSKSSLLHSRPKSTVGTPAYIAPEVLSRREYDGKMADVWSCGVT : 189 pk4 : ENTLLDGSPAPRLKICDFGYSKSSLLHSRPKSTVGTPAYIAPEVLSRREYDGKLADVWSCGVT : 189 pk5 : ENTLLDGSPAPRLKICDFGYSKSSLLHSRPKSTVGTPAYIAPEVLSRREYDGKLADVWSCGVT : 189 CsSTK1 : ENTLLDGSPAPRLKICDFGYSKSSLLHSRPKPTVGTPAYIAPEVLSRREYDGKMADVWSCGVT : 92 ENTLLDGSPAPRLKICDFGYSKSSLLHSRPKsTVGTPAYIAPEVLSRREYDGK6ADVWSCgVT * 200 * 220 * 240 * MtSK1 : LYVMLVGAYPFEDQDDPKNFRKTINRIMAVQYKIPDYVHISQECRHLLSRIFVASPARR-ITI : 251 OSAK : LYVMLVGAYPFEDQEDPKNFRKTIQRIMAVQYKIPDYVHISQDCRHLLSRIFVANPARR-ITI : 251 SPK-3 : LYVMLVGAYPFEDQDDPRNFRKTIQRIMAVQYKIPDYVHISQDCRHLLSRIFVANPLRR-ISL : 251 SPK-4 : LYVMLVGAYPFEDQDDPRNFRKTIQRIMAVQYKIPDYVHISQDCRHLLSRIFVANPLRR-ITI : 251 BSK2 : LYVMLVGAYPFEDQEDPKNFRKTIQKIMAVQYKIPDYVHISQDCKHLLSRIFVANSLKR-ITI : 251 ASK1 : LYVMLVGAYPFEDQEDPKNFRKTIQKIMAVQYKIPDYVHISQDCKNLLSRIFVANSLKR-ITI : 251 pk4 : LYVMLVGAYPFEDQDDPRNFRKTIQKIMAVQYKVPDYVHISQDCKHLFSRLFVANPSRR-ITL : 251 pk5 : LYVMLVGAYPFEDPEDPKNFRKPINRIMAVQYKIPDYVHISQECKHLLSRIFAANPSKR-ITI : 251 CsSTK1 : LYVMLVGAYPFEDPADPKNFRKTINRIMAVQYKIPDYVHISQECRHLLSRIFVASSLKEGITI : 155 LYVMLVGAYPFEDq DP4NFRKtI 4IMAVQYK6PDYVHISQ C4hLlSR6FvAn 4r I36 260 * 280 * 300 * MtSK1 : KEIKSHPWFLKNLPRELTEMAQAVYYRKENPTYSLQSIEDIMKIVEEAKNPPQASRSVGGFGW : 314 OSAK : KEIKSHPWFLKNLPRELTEAAQAAYYRRENPTFSLQSVEEIMKIVEEAKTPAPASRSVSGFGW : 314 SPK-3 : KEIKSHPWFLKNLPRELTESAQAVYYQRGNPSFSIQSVEEIMKIVGEARDPPPVSRPVKGFGW : 314 SPK-4 : KEIKNHPWFLRNLPRELTESAQAIYYQRDSPNFHLQSVDEIMKIVGEARNPPPVSRALKGFGW : 314 BSK2 : AEIKKHPWFLKNLPRELTETAQAAYFKKENPTFSPQTAEEIMKIVDDAKTPPPVSRSIGGFGW : 314 ASK1 : AEIKKHSWFLKNLPRELTETAQAAYFKKENPTFSLQTVEEIMKIVADAKTPPPVSRSIGGFGW : 314 pk4 : KEIKNHPWFLKNLPRELTESSQAIYYQRDNPSFSLQSVDEIMKIVGEARNPPPSSKIVRGFSW : 314 pk5 : KEIKNHPWFLKNLPRELTEAAQIMYYKKENPSFSLQSDREITKIVEDAKIPPPTSRSIGGFGW : 314 CsSTK1 : KEIKSHPWFLKTLPRELTEPAQQCTTRKENPSFSLQEFR-IYKNGEEAKFLPTSKR--GGFGW : 215 kEIK HpWFL4nLPRELTE aQa y 4 nP 5s Q eImKiv A4 ppp s4 GFgW 320 * 340 * 360 MtSK1 : GG-----------EEDDDEINEAEAELEE----DEYEKRVKEAQESGEIHVN : 351 OSAK : G----------GEEEEEEKEGDVEEEEEDEEEEDEYEKQVKQAHESGEVRLT : 356 SPK-3 : D------------GEEDEGEEDVEEEEDEE---DEYDKRVKEVHASGEFQIS : 351 SPK-4 : -------------EGEEDLDEEVEEEEDE----DEYDKRVKEVHASGEFQIS : 349 BSK2 : GGKGDEE------EEEVDEEEVVEEEEDEE---DEYDKTVKEAHASGEV--- : 354 ASK1 : GGNGDADGKEEDAEDVEEEEEEVEEEEDDE---DEYDKTVKEVHASGEVRIS : 363 pk4 : GP-------DENEEGNEDIDAEVEEEEEEEDEEDEYDKRVKEVHASGEFHIS : 359 pk5 : GG----------EEDGDGKEEEVNAEDEEE---DEYEKKVKEAQESGDVQVS : 353 CsSTK1 : G-------------GKKKG--------------------------------- : 221 g e ve e dey k vk sge
Fig. 1 Amino acid alignmentof deduced MtSK1 sequencewith other similar SnRK2proteins. GI numbers: 19568098(OSAK), 13430168 (CsSTK1),38228677 (pk4), 38343920(pk5), 1362051 (SPK-3),1161254 (SPK-4), 1097354(BSK2), and 15220251 (ASK1)

Fig. 2 Exon/intron structure ofMtSK1. Dark rectangles areexons (1–9). Light rectanglesare introns
consensus sequence from the overlapping fragments ob-tained earlier. Amplification of the full length MtSK1 cDNAfrom Jemalong 3-week callus confirmed that the sequencefor the coding region of MtSK1 was identical in 2HA and Je-malong. The final MtSK1 mRNA sequence obtained (Gen-Bank accession number, AY772417) is 1382 bp in length,with a predicted amino acid sequence of 351 amino acids(Fig. 1), a calculated molecular mass of 40.305 kD, and apredicted pI of 6.1.
Using a BLASTN search of the NCBI database, se-quences with the highest matches to the MtSK1 mRNA se-quence were identified. These sequences were then alignedusing ClustalW to determine total sequence identity. Thesequences with highest identity to MtSK1 were pk5 fromEuropean beech (Fagus sylvatica) with 67% identity, BSK2from oilseed rape (Brassica napus) with 62% and SPK-3 from soybean (Glycine max) with 60% (GI numbers:38343919, 289373 and 310581 respectively).
A BLASTN search of M. truncatula expressed sequencetags (ESTs) on the TIGR MtGI database revealed a num-ber of expressed sequences with high identity to MtSK1.On this database, MtSK1 is represented by the contig,TC107574. Another contig, TC107575, although not full-length, matches MtSK1 in the coding domain, except foran 8 bp deletion at the end of exon 1. There are also dif-ferences between the two sequences in the 5′ untranslatedregion. Another partial-length tentative contig sequence,TC105431, essentially matches MtSK1, except that it alsocontains the complete sequence of the first intron (Fig. 2).After that it contains sequence corresponding to exons 2,
3, 4 and 5 of MtSK1 with 100% identity to the MtSK1sequence. The part of the TC105431 sequence contain-ing the intron came from a single EST. It is noteworthythat most of the intron sequence codes for an amino acidsequence that is continuous with the identified coding se-quence of exon 1. The extra amino acid sequence is 54residues long and 50% of it is comprised of leucine orisoleucine residues. A BLASTP search of this sequenceon the NCBI database reveals no homology to known pro-teins. This tentative contig may represent an alternativesplice variant of MtSK1, but as the extra sequence is onlysupported by one EST sequence, and shows no homol-ogy to known proteins; it is most likely from an unsplicedsequence and does not represent a biologically activeprotein.
A number of other sequences show significant identityto MtSK1, primarily in the kinase domain. There are ninecontigs (other than those discussed above) which show>65% identity with MtSK1 over a region >900 bp inlength. (Contig numbers: TC95611, TC95687, TC94925,TC107709, TC107708, TC108117, TC107038, TC96356and TC105483). This indicates that the SnRK gene familyis most likely well represented in M. truncatula.
The tentative contig (TC107574), which is identical toMtSK1, is comprised of ESTs from a number of differentcDNA libraries. The source of tissue for these libraries isshown in Table 1. MtSK1 is expressed in a wide varietyof plant tissues including leaves, roots, flowers, pods andseeds.
Table 1 A list of the sourcesof ESTs belonging to MtSK1 onthe TIGR Medicago truncatulaGene Index (MtGI)
cDNA library Description
GPOD ESTs from immature pod wallsMHAM ESTs from roots colonized with Glomus versiformeDeveloping flower ESTs from pooled flowersGESD ESTs from immature seeds6KUG ESTs from mixed tissues at various stagesKV3 ESTs from roots 72 h after inoculation with RhizobiumDeveloping leaf ESTs from developing leavesPhoma-infected ESTs from young trifoliate leaves infected with Phoma medicaginisMtBC ESTs from roots infected with Glomus intraradicesAphid-infected shoots ESTs from aphid-infected shootsGerminating seed ESTs from germinating seedsMethyl jasmonate-elicited rootcell suspension culture
ESTs from methyl jasmonate-elicited root cell suspension culture
The MtSK1 mRNA sequence corresponds to the contig, TC107574 on the TIGR MtGI database. Partof this contig was assembled from Expressed Sequence Tags (ESTs), which came from the 12 cDNAlibraries listed. These show that MtSK1 is expressed in a broad range of tissues within the plant. Furtherinformation on the cDNA libraries is available from TIGR MtGI (http://www.tigr.org/tdb/ tgi/mtgi/)

The MtSK1 amino acid sequence shows high identitywith a number of proteins from different plant species(Fig. 1) including an osmotic stress-activated protein kinase(OSAK) from tobacco (Nicotiana tabacum; 87%), CsSTK1from European chestnut (Castanea sativa; 85%), pk4 andpk5 from European beech (Fagus sylvatica; 80% and 85%),SPK-3 and SPK-4 from soybean (82% and 80%), BSK2from oilseed rape (82%) and ASK1 (SnRK2.4) from Ara-bidopsis thaliana (79%). (GI numbers: 19568098 (OSAK),13430168 (CsSTK1), 38228677 (pk4), 38343920 (pk5),1362051 (SPK-3), 1161254 (SPK-4), 1097354 (BSK2) and15220251 (ASK1)).
It is likely that MtSK1 is a cytoplasmic protein, as anal-ysis of the predicted amino acid sequence reveals no evi-dence of a signal peptide, chloroplast transit peptide, mito-chondrial target sequence or nuclear signal.
The main features of the MtSK1 protein are an N-terminalkinase domain and a C-terminal Glutamic acid-rich region,which is predicted to be a coiled-coil. The kinase domainextends from residues 4–260. The eleven protein kinase
subdomains, as defined by Hanks and Hunter (1995), arepresent. It contains a predicted protein kinase ATP sig-nature pattern from residues 10–33 and a predicted Ser-ine/Threonine active protein kinase signature region fromresidues 119–131. The predicted coiled-coil region extendsfrom residues 316–349.
Genomic sequencing of MtSK1
The assembled genomic sequence (GenBank accessionnumber, AY772418) is 2842 bp in length, extending from4 bp before the ATG start codon to 43 bp after the stopcodon. It contains nine exons (Fig. 2).
Phylogenetic analysis
A number of the proteins which show high amino acidsequence identity with the predicted MtSK1 sequence aredefined members of the SnRK2 subgroup of the SnRK
Fig. 3

protein kinases (PlantsP (http://plantsp.sdsc.edu/). To de-termine if MtSK1 is also a member of this subgroup, aphylogenetic analysis was performed using the full length(or full known) amino acid sequences from MtSK1, all 74SnRK sequences from the PlantsP database, and five othersequences with high sequence identity with MtSK1, butwhich were not included on the PlantsP database. Thesewere OSAK, pk4, pk5 and CsSTK1 (described above) anda sequence from rice (Oryza sativa), GI number 38344935.The phylogenetic tree is shown in Fig. 3. The three SnRKsubgroups form three distinct branches on the unrootedtree, with a bootstrap value of 100% separating the threebranches. MtSK1 (number 1 on tree) and the other unde-fined, highly aligning sequences (numbers 2, 3, 4, 10, and13 on tree) all fall into the region with the known SnRK2subgroup members (Fig. 3).
All of the proteins of the SnRK2 branch of the phyloge-netic tree were checked using the COILS program (Lupaset al. 1991) for the presence of a coiled coil motif. All ofthem except for CsSTK1 contained a predicted coiled coilon the C-terminal end of the protein.
MtSK1 expression studies
2HA and Jemalong leaf tissue was cultured usingour standard culture protocol of P4 10:4 (containing10 µM 1-naphthalene acetic acid (NAA): 4 µM 6-benzylaminopurine (BAP)) medium for 3 weeks, thentransferred to P4 10:4:1 (10 µM NAA: 4 µM BAP: 1 µM
ABA). During the first 5 weeks of culture, tissue from both2HA and Jemalong shows the same response to culture byforming friable callus tissue. At around 5 weeks somaticembryos start to become visible on the 2HA calli, and by8 weeks of culture, each callus has formed multiple em-bryos. In contrast, embryo formation on Jemalong calli isuncommon.
The expression of MtSK1 was measured using Real TimePCR in 2HA and Jemalong cultures at various intervalsover a 5-week culture period, which corresponds to the pe-riod of induction of somatic embryogenesis in 2HA tissue(Fig. 4). The expression of MtSK1 was not significantlydifferent between the two seedlines over the culture periodexamined when calibrated to expression in each seedline’sstarting leaf tissue. Expression increases within 2 days ofculture, and continues to increase for 3 weeks. After 3weeks, MtSK1 expression plateaus, with the expressionat 5 weeks showing no significant difference to 3 weeks(Fig. 4).
The induction of MtSK1 expression is not dependent onhormones supplied in the culture medium. MtSK1 expres-sion is up regulated within 2 days of culture in both 2HAand Jemalong cultures in both the presence and absence ofhormones (results not shown).
The bioinformatics information and the related literatureon homologues clearly indicated a stress related signalingrole for MtSK1 as a member of the SnRK2 subgroup ofprotein kinases. SnRK2 subgroup of protein kinases havebeen shown to respond to a variety of stresses either ata transcription, or at a kinase activation level (Anderberg
� Fig. 3 Phylogenetic analysis of SnRK2 proteins. The bootstrap values at the forks indicate the number of times the groups consistingof the species which are distal to that fork occurred among the trees, out of 100 trees. The proteins fall into three groups (blue circles)corresponding to the SnRK subgroups (SnRK1, SnRK2 and SnRK3) with a bootstrap value of 100 dividing the groups. MtSK1 (number 1 onthe tree) lies in the SnRK2 subgroup. The proteins represented by the numbers on the tree are as follows in the format of: Number on tree. GInumber or Arabidopsis locus, Plant, (Other given name,s); 1 55140608, Medicago truncatula, (MtSK1); 2 38343920, Fagus sylvatica (pk5);3 134301678, Castanea sativa (CsSTK1); 4 19568098, Nicotiana tabacum (OSAK); 5 3046731, Craterostigma plantagineum, (cppk1); 6At1g60940, Arabidopsis thaliana (SnRK2.10); 7 At1g10940, Arabidopsis thaliana, (SnRK2.4, ASK1); 8 1097353, Brassica napus, (BSK1);9 1097354, Brassica napus, (BSK2); 10 38228677, Fagus sylvatica, (pk4); 11 1362051, Glycine max, (SPK-3); 12 1161254, Glycine max,(SPK-4); 13 38344935, Oryza sativa; 14 At5g63650, Arabidopsis thaliana, (SnRK2.5); 15 At5g08590, Arabidopsis thaliana, (SnRK2.1,ASK2); 16 At2g23030, Arabidopsis thaliana, (SnRK2.9); 17 1669573, Oryza sativa, (REK); 18 6739629, Vicia faba, (AAPK); 19 At3g50500,Arabidopsis thaliana, (SnRK2.2, SPK-2); 20 At5g66880, Arabidopsis thaliana, (SnRK2.3); 21 At4g33950, Arabidopsis thaliana, (SnRK2.6);22 2055374, Triticum aestivum, (TaPK3); 23 170664, Triticum aestivum, (PKABA1); 24 At1g78290, Arabidopsis thaliana, (SnRK2.8); 25407057, Mesembryanthemum crystallinum, (MCPK9); 26 At4g40010, Arabidopsis thaliana, (SnRK2.7); 27 1362050, Glycine max, (SPK-1); 28 4107005, Oryza sativa, (OSK2); 29 At5g39440, Arabidopsis thaliana, (SnRK1.3); 30 1216280, Solanum tuberosum, (PKIN1);31 1477684, Oryza sativa, (RSk1); 32 4107003, Oryza sativa, (OSK5); 33 4107001, Oryza sativa, (OSK4); 34 101523, Secale cereale,(RKIN1); 35 100610, Hordeum vulgare, (BKIN12); 36 At3g01090, Arabidopsis thaliana, (SnRK1.1, AKin10); 37 1076633, Nicotianatabacum, (NPK5); 38 7434358, Solanum tuberosum; 39 1743009, Cucumis sativus; 40 4567091, Glycine max; 41 At3g29160, Arabidopsisthaliana, (SnRK1.2, Akin11); 42 4091885, Oryza sativa; 43 4107009, Oryza sativa, (OSK1); 44 575292, Hordeum vulgare, (HVBKIN2);45 At5g57630, Arabidopsis thaliana, (SnRK3.4,CIPK21); 46 At3g17510, Arabidopsis thaliana, (SnRK3.16, CIPK1); 47 At1g48260,Arabidopsis thaliana, (SnRK3.21,CIPK17); 48 At1g48260, Arabidopsis thaliana, (F11A17.18); 49 At5g35410, Arabidopsis thaliana,(SnRK3.11, SOS2); 50 At4g24400, Arabidopsis thaliana, (SnRK3.13, CIPK8); 51 At1g01140, Arabidopsis thaliana, (SnRK3.12, CIPK9);52 At1g30270, Arabidopsis thaliana, (SnRK3.23, CIPK23); 53 2632252, Sorghum bicolor, (SNFL1); 54 At2g26980, Arabidopsis thaliana,(SnRK3.17, CIPK3); 55 At3g23000, Arabidopsis thaliana, (SnRK3.10, CIPK7); 56 At4g14580, Arabidopsis thaliana, (SnRK3.3, CIPK4);57 At5g10930, Arabidopsis thaliana, (SnRK3.24, CIPK5); 58 At5g25110, Arabidopsis thaliana, (SnRK3.25, CIPK25); 59. At2g25090,Arabidopsis thaliana, (SnRK3.18, CIPK16); 60 At5g01820, Arabidopsis thaliana, (SnRK3.15, CIPK14); 61 At2g30360, Arabidopsisthaliana, (SnRK3.22, CIPK11); 62 At2g38490, Arabidopsis thaliana; (SnRK3.19, CIPK22); 63 4884974, Mesembryanthemum crystallinum;64 At4g30960, Arabidopsis thaliana, (SnRK3.14, CIPK6); 65 At5g01810, Arabidopsis thaliana, (SnRK3.1, CIPK15); 66 2246625, Oryzasativa; 67 3256035, Sorghum bicolor, (SNFL3); 68 At5g58380, Arabidopsis thaliana, (SnRK3.8, CIPK10); 69 At5g07070, Arabidopsisthaliana, (SnRK3.2, CIPK2); 70 At5g45820, Arabidopsis thaliana, (SnRK3.6, CIPK20); 71 7340882, Oryza sativa; 72 7340880, Oryzasativa; 73 5821069, Oryza sativa, (OsPK7); 74 6651177, Zea mays, (ZmPK4); 75 626040, Triticum aestivum, (wpk4-p58); 76 3928519,Triticum aestivum, (wpk4); 77 At2g34180, Arabidopsis thaliana, (SnRK3.7, CIPK13); 78 At1g29230, Arabidopsis thaliana, (SnRK3.20,CIPK18); 79 At4g18700, Arabidopsis thaliana, (SnRK3.9, CIPK12); 80 At5g45810, Arabidopsis thaliana, (SnRK3.5, CIPK19). Note:Numbers 47 and 48 appear to be splice variants encoded by the same gene
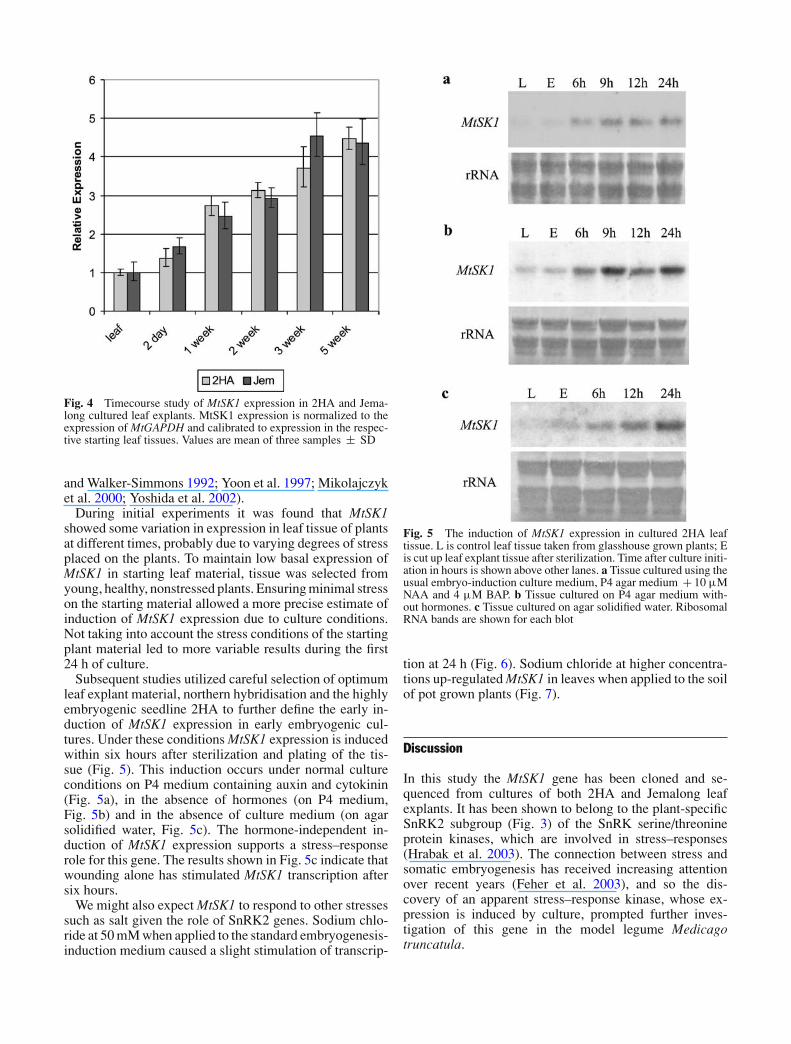
Fig. 4 Timecourse study of MtSK1 expression in 2HA and Jema-long cultured leaf explants. MtSK1 expression is normalized to theexpression of MtGAPDH and calibrated to expression in the respec-tive starting leaf tissues. Values are mean of three samples ± SD
and Walker-Simmons 1992; Yoon et al. 1997; Mikolajczyket al. 2000; Yoshida et al. 2002).
During initial experiments it was found that MtSK1showed some variation in expression in leaf tissue of plantsat different times, probably due to varying degrees of stressplaced on the plants. To maintain low basal expression ofMtSK1 in starting leaf material, tissue was selected fromyoung, healthy, nonstressed plants. Ensuring minimal stresson the starting material allowed a more precise estimate ofinduction of MtSK1 expression due to culture conditions.Not taking into account the stress conditions of the startingplant material led to more variable results during the first24 h of culture.
Subsequent studies utilized careful selection of optimumleaf explant material, northern hybridisation and the highlyembryogenic seedline 2HA to further define the early in-duction of MtSK1 expression in early embryogenic cul-tures. Under these conditions MtSK1 expression is inducedwithin six hours after sterilization and plating of the tis-sue (Fig. 5). This induction occurs under normal cultureconditions on P4 medium containing auxin and cytokinin(Fig. 5a), in the absence of hormones (on P4 medium,Fig. 5b) and in the absence of culture medium (on agarsolidified water, Fig. 5c). The hormone-independent in-duction of MtSK1 expression supports a stress–responserole for this gene. The results shown in Fig. 5c indicate thatwounding alone has stimulated MtSK1 transcription aftersix hours.
We might also expect MtSK1 to respond to other stressessuch as salt given the role of SnRK2 genes. Sodium chlo-ride at 50 mM when applied to the standard embryogenesis-induction medium caused a slight stimulation of transcrip-
Fig. 5 The induction of MtSK1 expression in cultured 2HA leaftissue. L is control leaf tissue taken from glasshouse grown plants; Eis cut up leaf explant tissue after sterilization. Time after culture initi-ation in hours is shown above other lanes. a Tissue cultured using theusual embryo-induction culture medium, P4 agar medium + 10 µMNAA and 4 µM BAP. b Tissue cultured on P4 agar medium with-out hormones. c Tissue cultured on agar solidified water. RibosomalRNA bands are shown for each blot
tion at 24 h (Fig. 6). Sodium chloride at higher concentra-tions up-regulated MtSK1 in leaves when applied to the soilof pot grown plants (Fig. 7).
Discussion
In this study the MtSK1 gene has been cloned and se-quenced from cultures of both 2HA and Jemalong leafexplants. It has been shown to belong to the plant-specificSnRK2 subgroup (Fig. 3) of the SnRK serine/threonineprotein kinases, which are involved in stress–responses(Hrabak et al. 2003). The connection between stress andsomatic embryogenesis has received increasing attentionover recent years (Feher et al. 2003), and so the dis-covery of an apparent stress–response kinase, whose ex-pression is induced by culture, prompted further inves-tigation of this gene in the model legume Medicagotruncatula.

Fig. 6 Northern analysis ofMtSK1 expression in responseto culture using normal culturemedium (no salt) and normalculture medium plus 50 mMNaCl (50 mM salt). L is controlleaf tissue taken fromglasshouse grown plants; E iscut up leaf explant tissue aftersterilization. Time after cultureinitiation is shown in hours. aNorthern hybridisation usingMtSK1 probe (top) andribosomal RNA bands from thesame blot (bottom). b Relativeexpression of MtSK1 calibratedto expression in leaf tissue.Results are the mean ±standard error of densitometerscans of three Northernexperiments
The MtSK1 gene
The amino acid sequence of MtSK1 shows high homol-ogy with other SnRK2 protein sequences (Fig. 1). SnRK2proteins share a high level of homology over their kinasedomain, which comprises most of the protein. The major di-vergence between the different proteins is in the C-terminal,acidic-rich region, which may also be part of a coiled coilregion. This region probably controls the specificity of theprotein and is also thought to function in protein–protein in-teractions (Hrabak et al. 1996). Two closely related SnRK2proteins of wheat illustrate the control of specificity by thisregion. The PKABA1 and TaPK3 proteins show 99–100%amino acid identity over most of the protein except fordivergence at the C-terminal, but respond to different envi-ronmental factors. PKABA1 responds to cold temperature,dehydration and ABA. TaPK3 does not respond to any ofthese stimuli, but is induced in greening tissue (Holappaand Walker-Simmons 1995, 1997).
Protein–protein interactions are a function of the coiledcoil motif. A coiled coil is a bundle of α-helices thatare wound into a superhelix and constitute a proteinoligomerization motif (Lupas 1996; Burkhard et al. 2001).The assembly of coiled coils can be modulated bypH, phosphorylation or by interaction with ions, mak-ing the system highly versatile (Burkhard et al. 2001).Of the SnRK2 proteins shown in Fig. 3, all exceptCsSTK1 contain a predicted coiled coil on the C-terminalend.
All of the SnRKs have a kinase domain close to the N-terminus. The SnRK1 subgroup is the most similar to theyeast SNF1 protein kinase with some similarity in the C-terminal regulatory domain as well as in the kinase domain(Hardie 1999). The SnRK1 proteins are global regulators ofcarbon metabolism in plants, by directly regulating enzymeactivity and through control of gene expression (Halfordand Hardie 1998; Laurie and Halford 2001; Halford et al.2003). The SnRK3 subgroup has an autoinhibitor domainadjacent to the C-terminal end of the kinase domain. Theautoinhibitor domain binds to a calcium binding proteinresulting in the activation of the kinase. No specific autoin-hibitor domain has been identified in SnRK1 and SnRK2subgroups (Hrabak et al. 2003).
Characterized SnRK2s function either in stress–responses (Anderberg and Walker-Simmons 1992; Yoonet al. 1997; Mikolajczyk et al. 2000; Yoshida et al. 2002)or are light inducible (Park et al. 1993; Holappa andWalker-Simmons 1997). The highest match to MtSK1 ona BLASTP search is the OSAK protein from tobacco(N. tabacum, GI:19568098). Although this work is un-published, the protein is described on the NCBI Pro-tein database as an osmotic stress-activated protein kinaseisolated from a BY-2 cell line (http://www.ncbi.nlm.nih.gov/). MtSK1 does not show the light-inducible expres-sion observed in ASK1 (SnRK2.4) and ASK2 (SnRK2.1) ofArabidopsis, and TaPK3 of wheat. Expression of ASK1and ASK2 is switched off after 24 h of dark treatment(Park et al. 1993). TaPK3 is expressed in greening seedlings

Fig. 7 Northern analysis of MtSK1 expression in leaf tissue fromplants grown in the presence of 50 mM (50) and 250 mM (250) NaClsolution. Salt solution was applied to plants (root zone) for 2, 4 and8 d and leaves were taken for total RNA isolation. a Northern hy-bridisation using MtSK1 probe (top) and ribosomal RNA bands fromthe same blot (bottom). C is control leaf tissue from plants withoutsalt treatment. 50 and 250 represent plants watered with 50 mM and250 mM NaCl solution respectively. b Relative expression of MtSK1calibrated to expression in leaf tissue from plants without salt treat-ment. Results are the mean ± standard error of densitometer scansof three Northern experiments
(Holappa and Walker-Simmons 1997). MtSK1 expressionis induced in the absence of light as cultures are maintainedin complete darkness.
MtSK1 expression in cultured 2HA and Jemalong
The induction of expression of MtSK1 in culture is not de-pendent on hormones in the culture medium with elevatedexpression on both hormone-containing and hormone-freemedia. The expression of MtSK1 gradually increases overthe first 3-weeks of culture. The changes in MtSK1 expres-sion are the same in 2HA and Jemalong. This suggests thatthe stress-related signaling of which MtSK1 is part is notrelated to the greatly enhanced capacity for embryogene-sis in 2HA. Clearly, however, the stress induces a majormetabolic shift which must interact with the auxin and cy-tokinin in promoting embryogenesis in 2HA. Our previouswork has implicated an inability of Jemalong to respondto cytokinin in the same way as 2HA (Nolan et al. 2003).What is of interest is that the cultured tissue, whether ornot embryos are being formed, appears to be continuouslyunder stress.
How is MtSK1 induced?
Excising the tissue has up-regulated MtSK1 expression.Expression of the CsSTK1 (an SnRK2 gene from chestnut)
was induced three hours after wounding, with the level ofexpression declining sharply after 24 h (Schafleitner andWilhelm 2002). However, with MtSK1, the continuing cul-ture has increased the expression still further. This may re-flect the nature of in vitro culture in a nutrient rich mediumand some osmotic stress. Consistent with this, MtSK1 ex-pression is up-regulated by sodium chloride. A connectionbetween these types of stresses and induction of somaticembryogenesis has been shown in carrot cultures, whereboth osmotic stress and salt stress have been shown to in-duce somatic embryogenesis (Kiyosue et al. 1989; Kamadaet al. 1993).
Most of the SnRK2s characterized to date are influencedby stresses related to water availability, and/or respond tothe stress-hormone ABA. Initial experiments have indi-cated some subtle changes in MtSK1 expression in responseto ABA in the culture medium, but further work is requiredto properly investigate this response. ABA in the culturemedium has been shown to increase the number of em-bryos that form on 2HA calli, and for this reason 1 µMABA is added to the medium after the first 3 weeks of cul-ture (Nolan and Rose 1998). Also, it has been shown thatSnRK2 regulation by ABA can occur either at the transcrip-tion level or at the protein activation level. For example,SnRK2.6 does not show an ABA-induced upregulation oftranscription, but SnRK2.6 kinase activity is stimulated byABA (Mustilli et al. 2002; Yoshida et al. 2002), whereastranscript levels of the SnRK2 genes, SPK-3 and PKABA1are upregulated by ABA (Anderberg and Walker-Simmons1992; Yoon et al. 1997). At this point in time there isno information on what may affect the kinase activity ofMtSK1.
Stress and development
Although the expression of MtSK1 cannot be directly re-lated to the induction of somatic embryogenesis in thissystem, the continued elevated expression of this gene isan indication of a prolonged stress–response that is presentin cultured tissue long after the stresses invoked by theinitiation of culture (such as wounding and chemical as-sault with sterilizing solutions) have subsided. This pro-longed stress–response occurring during the time of induc-tion of somatic embryogenesis may constitute an importantfoundation pathway without which, somatic embryogene-sis would not occur. Fig. 8 illustrates the stress responsepathway occurring in both Jemalong and 2HA and illus-trates a parallel hormone (auxin plus cytokinin) responsepathway in the somatic embryogenesis induction period.It is in the latter part of this process that 2HA diverges toproduce somatic embryos.
It is clear from proteomic studies that there are manyproteins produced early in culture in M. truncatula that arerelated to biotic and abiotic stress (Imin et al. 2004, 2005).For example using these culture systems it was shown thatthe level of the antioxidant protein, 1-Cys peroxiredoxin,is elevated in 2HA compared with Jemalong after 8 weeksof culture, whereas the reverse is true of the reactive

Fig. 8 Model of the development in culture of the near isogenic lines2HA and Jemalong. At the time of culture initiation, two parallel butpossibly interacting pathways, the stress–response pathway and thehormone-response pathway are invoked (yellow boxes). These path-ways in combination lead to the development of somatic embryos.The visible effects of these pathways are the similar growth anddevelopment of tissue from 2HA and Jemalong during culture up un-til the time of induction of somatic embryogenesis (blue boxes andphotographs). Around this point in time, the two near-isogenic seed-
lines diverge in their development. This becomes apparent at around5 weeks when somatic embryos become visible on 2HA cultures. By8 weeks, 2HA exhibits multiple embryo formation, in comparison tovery rare embryo formation exhibited on Jemalong tissue. It is notknown whether Jemalong tissue undergoes the process of inductionof somatic embryogenesis and further development of the embryosis blocked before they become visible, or if the process of inductiondoes not occur at all in Jemalong
oxygen species-scavenging proteins, ascorbate peroxidaseand a thioredoxin H-type protein (Imin et al. 2005).Among the most abundant proteins in 2HA and Jemalongcultures induced from both leaf explants and protoplastsare the pathogenesis related protein, PR10-1, and anABA-responsive protein that shows high homology toPR10-1 (Imin et al. 2004, 2005).
There are differences between Jemalong and 2HA, but theconnection between these differences and somatic embryo-genesis remains unknown. Any relationship of MtSK1 toABA-enhancement of somatic embryogenesis is unknownat this time, though further investigation is warranted. Theestablished role of ABA in stress–responses (Quatrano et al.1997; Bonetta and McCourt 1998; Seo and Koshiba 2002),and the fact that it stimulates somatic embryogenesis inM. truncatula (Nolan and Rose 1998), suggests that thishormone is probably instrumental in both pathways, andmay indicate points of crossover between the two. AnotherSnRK2 kinase (OST1/SnRK2.6) has been linked to signal-ing between ABA perception and hydrogen peroxide pro-duced via NADPH oxidase (Mustilli et al. 2002). Unpub-lished data in our laboratory (Saeed and Rose) have showninhibition of somatic embryogenesis using the NADPH ox-idase inhibitor DPI (diphenylene iodonium). The likely roleof MtSK1 in stress-induced signaling provides a way for-ward in relating the stress–response pathway to the auxin
and cytokinin-induced pathways involved in the inductionof somatic embryogenesis in the M. truncatula culture sys-tem.
Acknowledgements The authors would like to thank Dr ThomasKaiser of Corbett Research for his assistance with Real Time PCRand Professor Barry Rolfe (Australian National University, Canberra)for helpful discussions. This work was supported by an AustralianResearch Council Centre of Excellence grant to the University ofNewcastle node of the Centre of Excellence for Integrative LegumeResearch (to RJR).
References
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, MillerW, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: A newgeneration of protein database search programs. Nucleic AcidsRes 25:3389–3402
Anderberg RJ, Walker-Simmons MK (1992) Isolation of a wheatcDNA clone for an abscisic acid-inducible transcript with ho-mology to protein kinases. PNAS 89:10183–10187
Bendtsen JD, Nielsen H, von Heijne G, Brunak S (2004) Improvedprediction of signal peptides: SignalP 3.0. J Mol Biol 340:783–795
Bonetta D, McCourt P (1998) Genetic analysis of ABA signal trans-duction pathways. Trends Plant Sci 3:231–235
Burkhard P, Stetefeld J, Strelkov SV (2001) Coiled coils: Ahighly versatile protein folding motif. Trends Cell Biol 11:82–88

Chabaud M, Larsonneau C, Marmouget C, Huguet T (1996) Trans-formation of Barrel Medic (Medicago truncatula Gaertn) ByAgrobacterium tumefaciens and regeneration via somatic em-bryogenesis of transgenic plants with the Mtenod12 nodulinpromoter fused to the Gus reporter gene. Plant Cell Rep 15:305–310
das Neves LO, Duque SRL, de Almeida JS, Fevereiro PS (1999)Repetitive somatic embryogenesis in Medicago truncatula ssp.Narbonensis and M-truncatula Gaertn cv. Jemalong. Plant CellRep 18:398–405
Emanuelsson O, Nielsen H, Von Heijne G (1999) ChloroP, a neuralnetwork-based method for predicting chloroplast transit peptidesand their cleavage sites. Protein Sci 8:978–984
Feher A, Pasternak TP, Dudits D (2003) Transition of somatic plantcells to an embryogenic state. Plant Cell Tissue Organ Cult74:201–228
Felsenstein J (1989) PHYLIP—Phylogeny inference package (Ver-sion 3.2). Cladistics 5:164–166
Frugoli J, Harris J (2001) Medicago truncatula on the move! PlantCell 13:458–463
Halford NG, Hardie DG (1998) SNF1-related protein kinases: Globalregulators of carbon metabolism in plants? Plant Mol Biol37:735–748
Halford NG, Hey S, Jhurreea D, Laurie S, McKibbin RS, Paul M,Zhang Y (2003) Metabolic signalling and carbon partitioning:Role of Snf1-related (SnRK1) protein kinase. J Exp Bot 54:467–475
Hanks SK, Hunter T (1995) The eukaryotic protein kinase super-family: Kinase (catalytic) domain structure and classification.FASEB J 9:576–596
Hardie DG (1999) Plant protein serine threonine kinases: Classifi-cation and functions. Annu Rev Plant Physiol Plant Molec Biol50:97–131
Hofmann K, Bucher P, Falquet L, Bairoch A (1999) The PROSITEdatabase, its status in 1999. Nucleic Acids Res 27:215–219
Holappa LD, Walker-Simmons MK (1995) The wheat abscisic acid-responsive protein kinase mRNA, PKABA1, is up-regulated bydehydration, cold temperature, and osmotic stress. Plant Physiol108:1203–1210
Holappa LD, Walker-Simmons MK (1997) The wheat protein kinasegene, TaPK3, of the PKABA1 subfamily is differentially regu-lated in greening wheat seedlings. Plant Mol Biol 33:935–941
Hrabak EM, Chan CWM, Gribskov M, Harper JF, Choi JH, HalfordN, Kudla J, Luan S, Nimmo HG, Sussman MR, Thomas M,Walker-Simmons K, Zhu J-K, Harmon AC (2003) The Ara-bidopsis CDPK-SnRK superfamily of protein kinases. PlantPhysiol 132:666–680
Hrabak EM, Dickmann LJ, Satterlee JS, Sussman MR (1996) Charac-terization of eight new members of the calmodulin-like domainprotein kinase gene family from Arabidopsis thaliana. Plant MolBiol 31:405–412
Iantcheva A, Vlahova M, Trinh TH, Brown SC, Slater A, ElliotMC, Atanassov A (2001) Assessment of polysomaty, embryoformation and regeneration in liquid media for various speciesof diploid annual Medicago. Plant Sci 160:621–627
Imin N, De Jong F, Mathesius U, van Noorden G, Saeed NA, WangX-D, Rose RJ, Rolfe BG (2004) Proteome reference maps ofMedicago truncatula embryogenic cell cultures generated fromsingle protoplasts. Proteomics 4:1883–1896
Imin N, Nizamidin M, Daniher D, Nolan KE, Rose RJ, Rolfe BG(2005) Proteomic analysis of somatic embryogenesis in Med-icago truncatula. Plant Physiol 137:1250–1260
Kamada H, Ishikawa K, Saga H, Harada H (1993) Induction ofsomatic embryogenesis in carrot by osmotic stress. Plant TissueCult Lett 10:38–44
Kamada H, Tachikawa Y, Saitou T, Harada H (1994) Heat stressesinduction of carrot somatic embryogenesis. Plant Tissue CultLett 11:229–232
Kiyosue T, Kamada H, Harada H (1989) Induction of somatic em-bryogenesis by salt stress in carrot. Plant Tissue Cult Lett 6:162–164
Laurie S, Halford NG (2001) The role of protein kinases in theregulation of plant growth and development. Plant Growth Regul34:253–265
Lee EK, Cho DY, Soh WY (2001) Enhanced production and ger-mination of somatic embryos by temporary starvation in tissuecultures of Daucus carota. Plant Cell Rep 20:408–415
Lupas A (1996) Coiled coils: New structures and new functions.Trends Biochem Sci 21:375–382
Lupas A, Vandyke M, Stock J (1991) Predicting coiled coils fromprotein sequences. Science 252:1162–1164
Mikolajczyk M, Awotunde OS, Muszynska G, Klessig DF, Do-browolska G (2000) Osmotic stress induces rapid activation of asalicylic acid-induced protein kinase and a homolog of proteinkinase ASK1 in tobacco cells. Plant Cell 12:165–178
Mustilli A-C, Merlot S, Vavasseur A, Fenzi F, Giraudat J (2002)Arabidopsis OST1 protein kinase mediates the regulation ofstomatal aperture by abscisic acid and acts upstream of reactiveoxygen species production. Plant Cell 14:3089–3099
Nakai K, Kanehisa M (1992) A knowledge base for predicting pro-tein localization sites in eukaryotic cells. Genomics 14:897–911
Nishiwaki M, Fujino K, Koda Y, Masuda K, Kikuta Y (2000) Somaticembryogenesis induced by the simple application of abscisicacid to carrot (Daucus carota L.) seedlings in culture. Planta211:756–759
Nolan KE, Irwanto RR, Rose RJ (2003) Auxin up-regulatesMtSERK1 expression in both medicago truncatula root-forming and embryogenic cultures. Plant Physiol 133:218–230
Nolan KE, Rose RJ (1998) Plant regeneration from cultured Med-icago truncatula with particular reference to abscisic acid andlight treatments. Aust J Bot 46:151–160
Nolan KE, Rose RJ, Gorst JE (1989) Regeneration of Medicagotruncatula from tissue culture: increased somatic embryogenesisfrom regenerated plants. Plant Cell Rep 8:278–281
Page RDM (1996) TREEVIEW: An application to display phyloge-netic trees on personal computers. Comput Appl Biosci 12:357–358
Park YS, Hong SW, Oh SA, Kwak JM, Lee HH, Nam HG (1993)Two putative protein kinases from Arabidopsis thaliana containhighly acidic domains. Plant Mol Biol 22:615–624
Quatrano RS, Bartels D, Ho THD, Pages M (1997) New insights intoABA-mediated processes. Plant Cell 9:470–475
Rose RJ (2004) Somatic embryogenesis in plants. In: Goodman RM(ed) Encyclopedia of plant and crop science. Marcel DekkerInc., New York, pp 1165–1168
Rose RJ, Nolan KE, Bicego L (1999) The development of the highlyregenerable seed line Jemalong 2HA for transformation of Med-icago truncatula—Implications for regenerability via somaticembryogenesis. J Plant Physiol 155:788–791
Schafleitner R, Wilhelm E (2002) Isolation of wound inducible genesfrom Castanea sativa stems and expression analysis in the barktissue. Plant Physiol Biochem 40:235–245
Seo M, Koshiba T (2002) Complex regulation of ABA biosynthesisin plants. Trends Plant Sci 7:41–48
Thompson MR, Johnson LB, Gibson TJ (1994) CLUSTAL W: Im-proving the sensitivity of progressive multiple sequence align-ment through sequence weighting, position-specific gap penal-ties and weight matrix choice. Nucleic Acids Res 22:4673–4680
Touraev A, Vicente O, Heberlebors E (1997) Initiation of microsporeembryogenesis by stress. Trends Plant Sci 2:297–302
Yoon HW, Kim MC, Shin PG, Kim JS, Kim CY, Lee SY, HwangI, Bahk JD, Hong JC, Han C, Cho MJ (1997) Differential ex-pression of two functional serine/threonine protein kinases fromsoyabean that have an unusual acidic domain at the carboxyterminus. Mol Gen Genet 255:359–371
Yoshida R, Hobo T, Ichimura K, Mizoguchi T, Takahashi F, Aronso J,Ecker JR, Shinozaki K (2002) ABA-activated SnRK2 protein ki-nase is required for dehydration stress signaling in Arabidopsis.Plant Cell Physiol 43:1473–1483
Copyright © 2022 FDOKUMEN




















