
NK cells recognize and lyse Ewing sarcoma cells through NKG2D and DNAM-1 receptor dependent pathways
9
Molecular Immunology 45 (2008) 3917–3925 Contents lists available at ScienceDirect Molecular Immunology journal homepage: www.elsevier.com/locate/molimm NK cells recognize and lyse Ewing sarcoma cells through NKG2D and DNAM-1 receptor dependent pathways Dirk H.J. Verhoeven a , Alfons S.K. de Hooge a , Esther C.K. Mooiman a , Susy Justo Santos a , Monique M. ten Dam a , Hans Gelderblom b , Cornelis J.M. Melief c , Pancras C.W. Hogendoorn d , R. Maarten Egeler a , Maarten J.D. van Tol a , Marco W. Schilham a , Arjan C. Lankester a,∗ a Division of Immunology, Hematology, Oncology, Bone Marrow Transplantation and Autoimmune Diseases, Department of Pediatrics, Leiden University Medical Center, PO Box 9600, 2300 RC Leiden, The Netherlands b Department of Clinical Oncology, Leiden University Medical Center, PO Box 9600, 2300 RC Leiden, The Netherlands c Department of Immunohematology and Blood Transfusion, Leiden University Medical Center, PO Box 9600, 2300 RC Leiden, The Netherlands d Department of Pathology, Leiden University Medical Center, PO Box 9600, 2300 RC Leiden, The Netherlands article info Article history: Received 17 May 2008 Received in revised form 16 June 2008 Accepted 19 June 2008 Available online 26 July 2008 Keywords: NK cells Ewing sarcoma NKG2D DNAM-1 Cellular immunotherapy abstract Introduction: Ewing sarcoma (EWS) is a malignant bone-associated sarcoma, with poor prognosis in case of metastasis or relapse. To explore the feasibility of natural killer (NK) cell mediated immunotherapy and to identify molecular mechanisms involved, the susceptibility of EWS to NK cells was investigated. Methods and results: All EWS cell lines tested (n = 7) were lysed by purified allogeneic NK cells from healthy donors, and the efficacy of lysis was increased by activating NK cells with interleukin-15 (IL-15). FACS analysis and immunohistochemistry revealed that EWS cell lines as well as primary tumor cells expressed ligands for the activating NK cell receptors NKG2D and DNAM-1. NK cell cytotoxicity to EWS cells critically depended on the combination of NKG2D and DNAM-1 signaling, since blocking either of these receptors abrogated lysis by resting NK cells. Cytokine-activated NK cells more efficiently recognized EWS cells, since only combined, but not single blockade of NKG2D and DNAM-1 by antibodies inhibited lysis of EWS cells. Induction or blockade of HLA class I on EWS cells did not significantly influence lysis. This suggests that predominantly activating, rather than inhibitory signals on EWS cells determined susceptibility to NK cell cytotoxicity. NK cell cytotoxicity to EWS cells and K562 was reduced in EWS patients at diagnosis (n = 11) compared to age matched controls, despite normal NK cell numbers and increased expression of NKG2D. The impaired function of these NK cells was restored after activation with IL-15 in vitro. Conclusion: These results demonstrate that EWS cells are potentially susceptible to NK cell cytotoxicity due to the expression of activating NK cell receptor ligands. The use of cytokine-activated NK cells rather than resting NK cells in immunotherapy may be instrumental to optimize NK cell reactivity to EWS. © 2008 Elsevier Ltd. All rights reserved. 1. Introduction Ewing sarcoma (EWS) is the second most common malignant bone tumor occurring in children and young adults. Most patients suffering from EWS have a medullary-centered osseous localiza- tion, but a minority of patients presents with a primary soft tissue localization. EWS tumor cells typically bear the CD99 marker on their cell surface, and, in 95% of the cases, the tumor is character- ized by EWS/FLI-1 or EWS/ERG gene fusion (Burchill, 2003; Delattre et al., 1994). Treatment of EWS consists of chemotherapy and sur- ∗ Corresponding author. Tel.: +31 71 526 2824; fax: +31 71 526 6876. E-mail address: [email protected] (A.C. Lankester). gical resection, sometimes in combination with radiotherapy. This treatment strategy results in an overall survival rate of 60–65% in localized disease. However, approximately 25% of children with EWS have detectable metastases at the time of diagnosis, and these patients have a poor prognosis with a 5-year relapse free survival of less than 30%. In case of relapse, this prognosis may even be worse (Cotterill et al., 2000; Bernstein et al., 2006). Since the prognosis of patients presenting with metastasized or relapsed Ewing sarcoma is poor despite current chemotherapeutic and surgical treatment, immunotherapy may provide an additional or adjuvant treatment modality. Besides dendritic cells (DC) and T cells, NK cells may serve as effector or mediator cells of an anti-tumor immune response. Natural killer (NK) cells are bone marrow derived lymphocytes, which do not bear clonally rearranged antigen-specific receptors. 0161-5890/$ – see front matter © 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.molimm.2008.06.016
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of NK cells recognize and lyse Ewing sarcoma cells through NKG2D and DNAM-1 receptor dependent pathways

Molecular Immunology 45 (2008) 3917–3925
Contents lists available at ScienceDirect
Molecular Immunology
journa l homepage: www.e lsev ier .com/ locate /mol imm
NK cells recognize and lyse Ewing sarcoma cells through NKG2Dand DNAM-1 receptor dependent pathways
Dirk H.J. Verhoevena, Alfons S.K. de Hoogea, Esther C.K. Mooimana, Susy Justo Santosa,Monique M. ten Dama, Hans Gelderblomb, Cornelis J.M. Melief c, Pancras C.W. Hogendoornd,R. Maarten Egelera, Maarten J.D. van Tola, Marco W. Schilhama, Arjan C. Lankestera,∗
a Division of Immunology, Hematology, Oncology, Bone Marrow Transplantation and Autoimmune Diseases, Department of Pediatrics,Leiden University Medical Center, PO Box 9600, 2300 RC Leiden, The Netherlandsb Department of Clinical Oncology, Leiden University Medical Center, PO Box 9600, 2300 RC Leiden, The Netherlandsc Department of Immunohematology and Blood Transfusion, Leiden University Medical Center, PO Box 9600, 2300 RC Leiden, The Netherlandsd Department of Pathology, Leiden University Medical Center, PO Box 9600, 2300 RC Leiden, The Netherlands
a r t i c l e i n f o
Article history:
a b s t r a c t
a (Eexpl
hanisS celf lyschemK celtionNK
not sie of Hting,
Received 17 May 2008Received in revised form 16 June 2008Accepted 19 June 2008Available online 26 July 2008
Keywords:NK cellsEwing sarcomaNKG2DDNAM-1Cellular immunotherapy
Introduction: Ewing sarcomof metastasis or relapse. Toto identify molecular mecMethods and results: All EWdonors, and the efficacy oanalysis and immunohistoligands for the activating Ndepended on the combinaabrogated lysis by restingsince only combined, butcells. Induction or blockadthat predominantly activa
NK cell cytotoxicity.NK cell cytotoxicity to EWS ceage matched controls, despite nfunction of these NK cells was rConclusion: These results demodue to the expression of activatthan resting NK cells in immun
1. Introduction
Ewing sarcoma (EWS) is the second most common malignantbone tumor occurring in children and young adults. Most patientssuffering from EWS have a medullary-centered osseous localiza-tion, but a minority of patients presents with a primary soft tissuelocalization. EWS tumor cells typically bear the CD99 marker ontheir cell surface, and, in 95% of the cases, the tumor is character-ized by EWS/FLI-1 or EWS/ERG gene fusion (Burchill, 2003; Delattreet al., 1994). Treatment of EWS consists of chemotherapy and sur-
∗ Corresponding author. Tel.: +31 71 526 2824; fax: +31 71 526 6876.E-mail address: [email protected] (A.C. Lankester).
0161-5890/$ – see front matter © 2008 Elsevier Ltd. All rights reserved.doi:10.1016/j.molimm.2008.06.016
WS) is a malignant bone-associated sarcoma, with poor prognosis in caseore the feasibility of natural killer (NK) cell mediated immunotherapy andms involved, the susceptibility of EWS to NK cells was investigated.l lines tested (n = 7) were lysed by purified allogeneic NK cells from healthyis was increased by activating NK cells with interleukin-15 (IL-15). FACSistry revealed that EWS cell lines as well as primary tumor cells expressedl receptors NKG2D and DNAM-1. NK cell cytotoxicity to EWS cells criticallyof NKG2D and DNAM-1 signaling, since blocking either of these receptorscells. Cytokine-activated NK cells more efficiently recognized EWS cells,ngle blockade of NKG2D and DNAM-1 by antibodies inhibited lysis of EWS
LA class I on EWS cells did not significantly influence lysis. This suggestsrather than inhibitory signals on EWS cells determined susceptibility to
lls and K562 was reduced in EWS patients at diagnosis (n = 11) compared toormal NK cell numbers and increased expression of NKG2D. The impaired
estored after activation with IL-15 in vitro.nstrate that EWS cells are potentially susceptible to NK cell cytotoxicitying NK cell receptor ligands. The use of cytokine-activated NK cells ratherotherapy may be instrumental to optimize NK cell reactivity to EWS.© 2008 Elsevier Ltd. All rights reserved.
gical resection, sometimes in combination with radiotherapy. Thistreatment strategy results in an overall survival rate of 60–65% inlocalized disease. However, approximately 25% of children withEWS have detectable metastases at the time of diagnosis, and thesepatients have a poor prognosis with a 5-year relapse free survival ofless than 30%. In case of relapse, this prognosis may even be worse(Cotterill et al., 2000; Bernstein et al., 2006). Since the prognosis ofpatients presenting with metastasized or relapsed Ewing sarcomais poor despite current chemotherapeutic and surgical treatment,immunotherapy may provide an additional or adjuvant treatmentmodality. Besides dendritic cells (DC) and T cells, NK cells may serveas effector or mediator cells of an anti-tumor immune response.
Natural killer (NK) cells are bone marrow derived lymphocytes,which do not bear clonally rearranged antigen-specific receptors.

ar Imm
3918 D.H.J. Verhoeven et al. / MoleculThey are cytotoxic against tumor cells and virus infected cells. Activ-ity of NK cells is controlled by a balance between inhibitory andactivating signals (Carayannopoulos and Yokoyama, 2004; Morettaet al., 2006). Inhibitory signals are provided by classical as wellas non-classical human leukocyte antigen (HLA) class I molecules,which either bind to killer immunoglobulin-like receptors (KIR) orNKG2A/CD94 on NK cells (Parham, 2005; Moretta et al., 2006).These inhibitory signals are counteracted by activating signals,provided by several receptor–ligand combinations. The best char-acterized activating NK cell receptor is NKG2D, a homodimer witha C-type lectin-like extracellular domain. The activating signal istransduced through recruitment of the adaptor molecule DAP10and the subsequent activation of the phosphatidylinositol 3-kinase(PI3-K) pathway. NKG2D binds the MIC (MICA/B) and ULBP (ULBP1-4) family of proteins (Raulet, 2003; Hayakawa and Smyth, 2006).These ligands are expressed on tumor cells and virus infected cellsand can amongst others be induced by DNA damage (Gasser andRaulet, 2006).
DNAX accessory molecule-1 (DNAM-1, CD226) is an adhesionreceptor with two Ig-like domains in the extracellular part, whichis expressed on the majority of NK cells, T cells and monocytes(Shibuya et al., 1996). It is associated with LFA-1 and transfers anactivating signal through Fyn mediated tyrosine phosphorylationand protein kinase C (PKC) mediated serine phosphorylation of thecytoplasmic domain of DNAM-1 (Shibuya et al., 1998, 1999). Thisactivating receptor recognizes the poliovirus receptor (PVR, CD155)and Nectin-2 (CD112) (Bottino et al., 2003). These two membersof the Nectin family are not only expressed on healthy neuronal,epithelial, endothelial cells and fibroblasts, but they are also highlyexpressed on several tumor cells (Campadelli-Fiume et al., 2000).Although the precise role of DNAM-1 in NK and T cell biology isstill under investigation, NK cell mediated lysis of neuroblastomacells and ovarian carcinoma cells was shown to be dependent onthe interaction of DNAM-1 with its ligands (Castriconi et al., 2004;Carlsten et al., 2007).
Other activating receptors include the natural cytotoxicityreceptors (NCRs): NKp30 and NKp46, which are expressed on allNK cells; NKp44, which can be detected on cytokine-activated NKcells. The ligands for these receptors are largely unknown, althoughHLA-B-associated transcript 3 (BAT3) was reported to be an acti-vating ligand for NKp30 (Pogge von Strandmann et al., 2007) andinfluenza hemagglutinin has been demonstrated to be a ligand forNKp46 (Moretta et al., 2006).
We report here that EWS tumor cells are recognized and lysed
by resting and with higher efficacy by activated NK cells throughNKG2D and DNAM-1 dependent pathways. Furthermore, we reportthat NK cell cytotoxicity is impaired in patients with EWS by a yetunknown mechanism.2. Patients, materials and methods
2.1. Patients
During 2004–2006, 11 children and adolescents with newlydiagnosed Ewing sarcoma with an osseous localization (age aver-age 15.0 years, range 4.9–20.5 years) were enrolled in this studyat the Leiden University Medical Center (Departments of Pedi-atrics and Clinical Oncology) after informed written consent. Thestudy was approved by the Institutional Review Board on MedicalEthics. Peripheral blood mononuclear cells (PBMC) were isolatedfrom blood collected prior to start of chemotherapy by Ficoll den-sity gradient separation and were stored in liquid nitrogen untilfurther investigation. Decalcified, paraffin embedded tissue andcryosections from tumor biopsies obtained at diagnosis were avail-
unology 45 (2008) 3917–3925
able for research purposes. PBMC, obtained from 15 age matchedhealthy controls (age average 13.9 years, range 3.9–28.8 years) afterinformed consent, were used as a reference in experiments.
2.2. Cell lines
The Ewing sarcoma cell lines A673, RD-ES, SKES-1, SK-N-MC,TC32 (American Type Culture Collection, Manassas, VA, USA), EW3(Urano et al., 1998), and L1062 (Szuhai et al., 2007) and the humanerythroleukemia cell line K562 were cultured in RPMI 1640 (Invit-rogen, Carlsbad, CA, USA), supplemented with 10% fetal bovineserum (Greiner Bio-One, Alphen a/d Rijn, Netherlands) and peni-cillin/streptomycin (Invitrogen). The EWS cell lines have beenintensely characterized at the genomic level (Szuhai et al., 2006).The EBV B-LCL cell line 107 (HLA-A11/33, HLA-B35/58, HLA-Cw3/4)was generated from a healthy blood bank donor expressing HLAalleles which are ligands to all inhibitory KIR receptors [HLA-A3 or-A11 (ligands to KIR3DL2), the Bw4 group (KIR3DL1), the HLA-CLys80
group (KIR2DL1) and the HLA-CAsn80 group (KIR2DL2/3)]. The EWScell line SKES-1 has a HLA class I genotype with similar characteris-tics (HLA-A2/11, HLA-B7/44, HLA-Cw5/7). Cell lines were routinelyscreened for mycoplasma contamination. For upregulation of HLAclass I, the SKES-1 cell line was cultured for 48 h in the presence of100 IU IFN�/ml (Boehringer, Ingelheim, Germany).
2.3. Antibodies
EWS cell lines were stained with the following monoclonal anti-bodies (mAbs): anti-MICA (clone 159227), MICB (236511), ULBP-1(170181), ULBP-2 (165903), ULBP-3 (166510), all from R&D Sys-tems Europe (Abingdon, UK); CD48 (clone J4.57), CD112 (R2.477.1),CD155 (PV.404), all from Beckman Coulter Immunotech (Marseille,France). APC conjugated goat IgG anti-mouse was used as a sec-ondary antibody in stainings with the mAbs mentioned above. Inaddition the following fluorochrome conjugated mAbs were used:anti-HLA-A/B/C-FITC (clone G46-2.6), CD54-PE (HA58), CD58-PE(IC3), CD95-PE (DX2), all from BD Pharmingen (San Diego, CA, USA),and CD11a-PE (clone TB-133) from Sanquin (Amsterdam, Nether-lands).
PBMC from EWS patients were stained with the following fluo-rochrome conjugated mAbs: anti-CD16-FITC (clone 3G8), CD19-PE(J4.119), CD56-APC (N901), CD244/2B4-PE (C1.7), NKp30-PE (Z25),NKp46-PE (BAB281), NKG2A-PE (Z199), NKG2D-PE (ON172), anti-KIR mAbs clone EB6-PE (KIR2DS1/KIR2DL1), FES172-PE (KIR2DS4),
GL183-PE (KIR2DS2 and KIR2DL2/3) and anti-Glycophorin A-PE (KC16), all from Beckman Coulter Immunotech (Marseille,France); CD3-PerCP-Cy5.5 (SK7), CD14/45-FITC/PE (lymphogate),anti-KIR mAbs clone DX9-PE (KIR3DL1), HP3E4-PE (KIR2DS1/S4and KIR2DL1) and CD33-FITC (P67.6), all from Becton-Dickinson(Franklin Lakes, NJ, USA).2.4. Purification and culture of NK cells
PBMC were obtained from healthy blood bank donors afterinformed consent and were isolated using Ficoll density gradi-ent separation. NK cells were enriched either using the MACSNK enrichment kit II and LS columns (Miltenyi Biotec, BergischGladbach, Germany) or by monocyte adherence for 2 h at 37◦ andsubsequent T cell depletion using the MACS system in combina-tion with anti-CD3 microbeads and LD columns (Miltenyi Biotec)according to the manufacturer’s protocol.
T cell contamination was always less than 0.2% as determined byFACS analysis. NK cell purity was 88–96% using the NK enrichmentkit II, which was applied to purify NK cells for use in experimentswith resting NK cells.
ar Imm
determined as: (measured value-spontaneous release)/(maximumrelease-spontaneous release) × 100%. The E:T ratios were correctedfor the percentage of NK cells as was determined by FACS analysis.Cytotoxicity was expressed as the amount of lytic units (LU-25) per106 NK cells. One LU is defined as the number of NK cells requiredto achieve 25% specific lysis in 2500 target cells.
In blocking experiments, NK cells were incubated for 20 minprior to chromium release assays with blocking anti-NKG2D (R&Dsystems, clone 149810, IgG1), blocking anti-DNAM-1 (BD Pharmin-gen, clone DX11, IgG1) or non-blocking anti-CD56 monoclonalantibodies (Coulter Immunotech, clone N901, IgG1) at a concen-tration of 20 �g/mL.
HLA class I was blocked on the target cell lines by pre-incubatingthem with the masking pan-HLA class I antibody w6/32 (IgG2a) ata concentration of 5 �g/mL.
2.8. Statistical analysis
Values obtained from patients and age matched healthy con-trols were statistically evaluated using the non-parametrical
D.H.J. Verhoeven et al. / Molecul
NK cells, enriched by monocyte adherence and T cell depletion,were cultured in AIM-V medium (Invitrogen), supplemented with10% pooled human AB serum (Sanquin, Rotterdam, Netherlands),penicillin/streptomycin, glutamine (Glutamax I, Invitrogen). Cul-tured NK cells were stimulated with 10 ng/mL recombinant humaninterleukin-15 (IL-15) (Bender Medical Systems, Vienna, Austria)once or twice a week. Cytokine-stimulated NK cells were usedbetween 2 and 4 weeks of culture. NK cell cultures were routinelyanalyzed for purity by FACS analysis. After 14 days of culture, >97%of the viable cells were NK cells.
2.5. FACS staining
Four-color flow cytometry was performed on a FACScalibur(Becton Dickinson), and results were analyzed using Cellquestsoftware (Becton Dickinson). Lymphocytes were defined asCD14−/CD33−/glycophorin A−/CD45+ cells, NK cells were definedas CD3−/CD56+ lymphocytes, T cells were defined as CD3+ lympho-cytes, and B cells were defined as CD19+ lymphocytes. Expression ofreceptors on NK cells was determined using a CD3/56/16 staining incombination with the specific NK cell receptor staining. CD56bright
NK cells were defined as CD3−/CD56high/CD16− PBMC, and CD56dim
NK cells were defined as CD3−/CD56intermediate/CD16+ PBMC.
2.6. Staining of biopsies
Immunohistochemistry staining for CD112 and CD155 was per-formed on 4-�m cryostat sections fixed in 4% formaldehyde (Merck,Amsterdam, Netherlands) since the epitopes of these mAbs did notsurvive decalcification. Staining for MICA and CD99 was performedon formalin-fixed, paraffin embedded sections. Decalcified slides(14 days in formic acid) were processed after removal of paraf-fin and after antigen retrieval in citrate buffer. All sections weresubsequently exposed to 0.3% H2O2 to quench the endogenous per-oxidase activity and to 10% normal rabbit serum (Dako, Glostrup,Denmark).
Mouse IgM anti-human CD99 (clone #H036-1.1, Abcam, Cam-bridge, UK), mouse IgG2a anti-human CD112 (clone #L14, agenerous gift from Dr. D. Pende (IST, Genova, Italy)), mouse IgG1anti-human CD155 (clone #D171, Lab Vision Corporation, Fremont,CA, USA) and polyclonal goat IgG anti-human MICA (#AF1300,R&D Systems, Abingdon, UK) were used. After overnight incubationwith one of these primary antibodies, sections were treated withbiotinylated rabbit anti-mouse or rabbit anti-goat immunoglob-
ulin antibody (Dako) and SABC (biotinylated horseradish per-oxidase/streptavidin complex, Dako). Binding was visualizedwith diamino-benzidine-tetrahydrochloride (Sigma–Aldrich, Zwi-jndrecht, Netherlands)/0.002% H2O2. Slides were mounted afterMayer’s hematoxylin nucleus counterstaining.2.7. Chromium release assays
Cytotoxicity was determined in a standard 4-h chromiumrelease assay. Briefly, 2 × 106 target cells were incubated with100 �Ci sodium-51-chromate (PerkinElmer, Wellesley, MA, USA)for 1 h. PBMC obtained from patients or controls were thawed fromstorage in liquid nitrogen and were allowed to recover for 16 hin RPMI supplemented with 10% FCS and penicillin/streptomycin.Effector cells (PBMC, unstimulated purified NK cells or stim-ulated NK cells) were incubated for 4 h with 2500 targetcells (either K562, EWS cell lines or EBV-BLCL) at eight effec-tor:target (E:T) ratios in triplicate. Maximum and spontaneousrelease was determined by incubating targets in 2N HCl ormedium, respectively. Supernatants were harvested and mea-sured in a gamma-counter (Wallac, PerkinElmer). Specific lysis was
unology 45 (2008) 3917–3925 3919
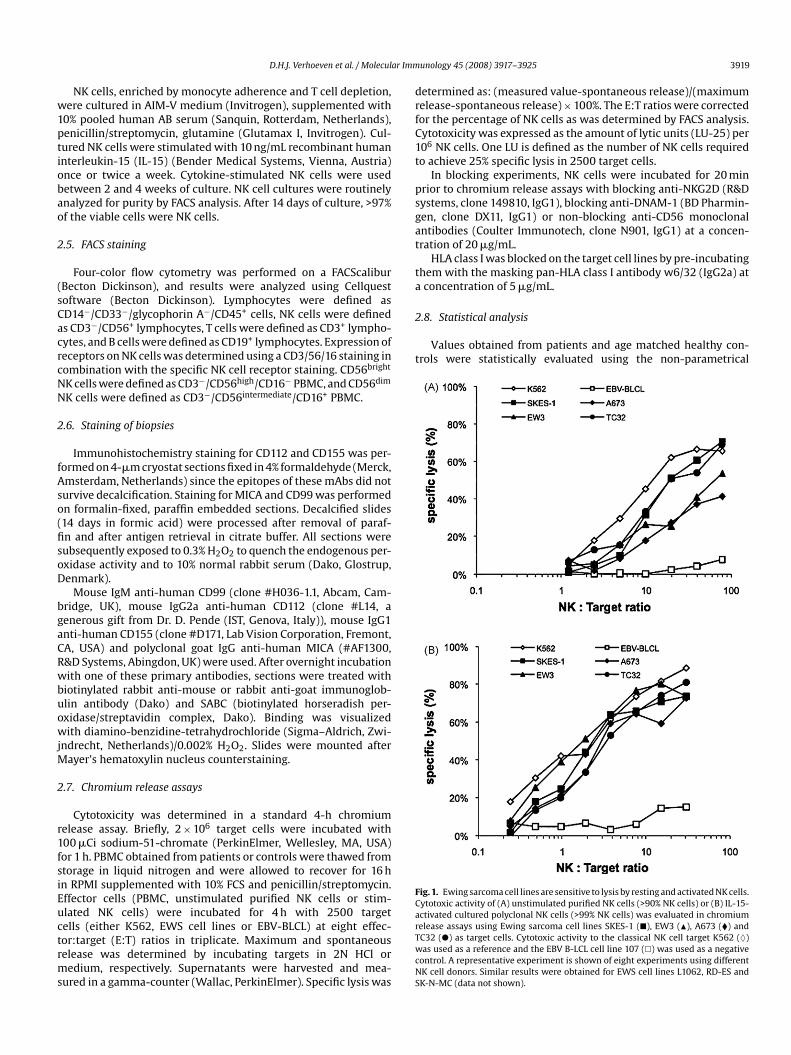
Fig. 1. Ewing sarcoma cell lines are sensitive to lysis by resting and activated NK cells.Cytotoxic activity of (A) unstimulated purified NK cells (>90% NK cells) or (B) IL-15-activated cultured polyclonal NK cells (>99% NK cells) was evaluated in chromiumrelease assays using Ewing sarcoma cell lines SKES-1 (�), EW3 (�), A673 (�) andTC32 (�) as target cells. Cytotoxic activity to the classical NK cell target K562 (♦)was used as a reference and the EBV B-LCL cell line 107 (�) was used as a negativecontrol. A representative experiment is shown of eight experiments using differentNK cell donors. Similar results were obtained for EWS cell lines L1062, RD-ES andSK-N-MC (data not shown).

3920 D.H.J. Verhoeven et al. / Molecular Imm
Table 1Ewing sarcoma cell lines express ligands for the activating NK cell receptors NKG2Dand DNAM-1
SKES-1 SKNMC L1062 A673 EW3 TC32 RD-ES
MHC class I − ++ + ++ + ++ ++CD48 (2B4 ligand) − − − − − − −MICA + +/− ++ ++ − + +/−MICB +/− +/− + + +/− + −ULBP-1 ++ ++ + + ++ + ++ULBP-2 + + ++ ++ − − +ULBP-3 − − − +/− − − +CD112 (Nectin-2) ++ ++ ++ ++ + ++ +CD155 (PVR) ++ ++ ++ ++ ++ ++ +CD54 (ICAM-1) − − − − + − ++CD58 (LFA-3) ++ ++ ++ + − + +
Expression of ligands for inhibitory NK cell receptors KIR and 2B4, ligands for acti-vating NK cell receptors NKG2D and DNAM-1 and of adhesion molecules (CD54 andCD58) was determined on EWS cell lines using flow cytometry. (−) mean fluores-cence intensity (MFI) ratio of specific staining versus isotype control <2; (+/−) MFIratio between 2 and 5; (+) MFI ratio between 5 and 10; (++) MFI ratio >10.
Mann–Whitney U-test and the SPSS software package (SPSS,Chicago, IL, USA).
3. Results
3.1. Ewing sarcoma cell lines are susceptible to lysis by restingand activated NK cells
Resting NK cells have cytotoxic activity to a limited num-ber of human tumor cell lines. To evaluate the sensitivity ofEwing sarcoma to NK cell mediated lysis, chromium release assayswere performed using EWS cell lines as targets. As illustratedin Fig. 1A, EWS cells were lysed by freshly isolated resting NKcells obtained from healthy donors. Activation of NK cells withrecombinant human IL-15 increased the efficacy of lysis of Ewingsarcoma tumor cells by NK cells (Fig. 1B). Consistent results wereobtained using eight different healthy NK cell donors, indicatingthat EWS tumor cells can be recognized and lysed by both rest-ing and activated NK cells. NK cell activation with interleukin-2resulted in a comparable increase in NK cell cytotoxicity (data notshown).
3.2. Ewing sarcoma tumors and cell lines express ligands foractivating NK cell receptors
Since activation of NK cells is determined by the balancebetween activating and inhibitory signals delivered by the targetcell, the expression of some of the ligands for NK cell receptorswas determined on EWS cell lines and tissue biopsy sections. EWScell lines displayed a heterogeneous HLA class I expression, whichdid not clearly correlate with their sensitivity to lysis by NK cells.In addition, EWS cell lines displayed heterogeneous expression ofMICA, MICB, ULBP-1, ULBP-2 and ULBP-3, which are ligands for theactivating receptor NKG2D on NK cells (Fig. 2A, Table 1). All cell linesexpressed at least one member of the MIC/ULBP family of NKG2Dligands with high intensity. Furthermore, EWS cell lines expressedhigh levels of CD112 (Nectin-2) and CD155 (PVR), which are ligandsfor the activating adhesion receptor DNAM-1 on NK cells (Fig. 2A,Table 1). EWS cell lines were CD48 (the ligand for 2B4) negative andexpressed variable amounts of the adhesion molecules CD54 andCD58 (Table 1).
The expression of NKG2D and DNAM-1 ligands in vivo was con-firmed by positive staining of Ewing sarcoma tumor biopsy sectionsobtained at diagnosis with antibodies against MICA, CD112 andCD155 (Fig. 2B). These results indicate that primary Ewing sarcoma
unology 45 (2008) 3917–3925
tumor cells express activating NK cell receptor ligands in vivo, whichmay provide activating signals for NK cells.
3.3. Cytotoxicity of resting NK cells to EWS tumor cells isdependent on the activating receptors NKG2D and DNAM-1
Cytotoxic activity of NK cells is regulated by the balance betweenthe inhibitory and activating signals that the target cell provides.Induction of HLA class I, which binds to inhibitory KIR receptors,on the HLA class I negative cell line SKES-1 by stimulation withinterferon-� did not result in a significant decrease of cytotoxicactivity of resting and activated NK cells to this cell line (Fig. 3A/B)and blocking HLA class I did not result in increased lysis of EWS cells(Fig. 3C). SKES-1 has an HLA genotype containing HLA-A, HLA-Bw4,HLA-C group 1 as well as HLA C group 2 ligands for inhibitory KIRreceptors, suggesting that not loss of inhibitory signals, but that thepresence of activating signals could be a predominant factor in therecognition of EWS cells by NK cells.
The contribution of signals provided by the activating NK cellreceptors NKG2D and DNAM-1, whose ligands are expressed onEWS tumor cells, was studied using blocking antibodies againstthese receptors. Blocking of either NKG2D or DNAM-1 on restingNK cells efficiently reduced NK cell mediated lysis of EWS cell lineswith 70–90% compared to NK cells incubated with a non-blockingCD56 isotype matched antibody (Fig. 4A). These results suggest thatsimultaneous NKG2D and DNAM-1 mediated signals are critical inthe recognition and lysis of EWS cells by unactivated NK cells.
When IL-15-activated NK cells were used as effector cells, cyto-toxic activity was reduced minimally by single blocking of NKG2Dor DNAM-1 receptors on NK cells. However, blocking NKG2D andDNAM-1 simultaneously, efficiently reduced NK cell cytotoxicity(Fig. 4B). These observations were consistent using nine differenthealthy NK cell donors. Concluding, in both resting and activatedNK cells, DNAM-1 and NKG2D mediated signals are relevant for therecognition and lysis of EWS cells, but activated NK cells dependless on multiple activating signals compared to resting NK cells.
3.4. NK cell cytotoxicity is reduced in Ewing sarcoma patients attime of diagnosis
Since non-activated NK cells from healthy donors recognizeEWS cells, NK cell function and phenotype were investigated inEWS patients at diagnosis (n = 11). Lymphocyte counts (average:2046/�L blood), the number of NK cells (average: 230/�L blood)
and the percentage of the CD56dim NK cell subset (average: 89%of NK cells) were within the range of healthy age matched con-trols (n = 15; average: 2445/�L blood; 265/�L blood and 90% of NKcells, respectively). However, when PBMC from these patients weretested directly ex vivo against the EWS cell line A673, a significantlyreduced cytotoxic activity was observed (Fig. 5A, p = 0.001). Thisdecrease in NK cell cytotoxicity was also observed using the HLAclass I negative NK cell target K562 (Fig. 5A, p = 0.005) and the HLAclass I negative EWS cell line SKES-1 (Fig. 5B, left panel, p = 0.039).The impaired NK cell cytotoxicity was restored by stimulating PBMCfrom EWS patients with IL-15 for 3 days (Fig. 5B). This suggeststhat cytokine activation either restored efficient transduction ofactivating signals or restored impaired effector functions.3.5. NK cells from EWS patients have intact NKG2D expression
In several tumor models, tumor immune escape mechanismshave been described, including downregulation of NKG2D andNCRs, which are expressed on all NK cells (Hayakawa and Smyth,2006; Costello et al., 2002). Therefore, the NK cell receptor expres-sion on NK cells of EWS patients was investigated. As shown in

D.H.J. Verhoeven et al. / Molecular Immunology 45 (2008) 3917–3925 3921
ssionwn forn sectligand
d 155cifiedvo one for t
Fig. 2. Ewing sarcoma cells express ligands for activating NK cell receptors. (A) ExpreCD112 and CD155) was evaluated by flow cytometry. Representative results are shomatched control staining is shown in gray in each histogram. (B) Upper panel: FrozeCD112 and CD155 or isotype control antibodies, revealing expression of DNAM-1staining, magnification 200×. Frozen tissue sections were used since the CD112 ancontrol). Lower panel: Tumor biopsy bone sections obtained at diagnosis were decalMICA or goat IgG control antibodies, revealing expression of the NKG2D ligand in vi200×. All sections were counterstained with hematoxylin. All biopsies were positiv
Fig. 5C, expression levels of NKG2D on resting NK cells were para-doxically higher in EWS patients compared to age matched controls
on both CD56bright and CD56dim NK cell subsets (p = 0.004 and0.021, respectively). The expression (MFI ± S.D.) of the NCRs NKp30(patients: 7.9 ± 2.9 vs. controls: 6.2 ± 3.2; p = 0.10) and NKp46(patients: 10.2 ± 2.4 vs. controls: 6.8 ± 3.2; p = 0.08) on CD56 dimNK cells was not significantly different between patients and con-trols. The percentage of DNAM-1, NKp44, KIR and NKG2A positiveNK cells in EWS patients were comparable to age matched controls(data not shown).Since NKG2D and DNAM-1 pathways were shown to be essen-tial for the recognition of EWS cell lines, the functionality of thesereceptor systems was evaluated in NK cells from EWS patients usingblocking antibodies (n = 3 patients). As illustrated in Fig. 5D, cyto-toxic activity of EWS patient PBMC to the EWS cell line TC32 wasinhibited by blocking antibodies to either NKG2D or DNAM-1, indi-cating that both pathways contribute to the (residual) cytotoxicactivity of NK cells from EWS patients at diagnosis.
4. Discussion and conclusions
In this preclinical study, we report that Ewing sarcoma (EWS)cells were recognized and lysed by resting and, with higher effi-
of inhibitory (HLA class I) and activating NK cell receptor ligands (MICA/B, ULBP1-3,EWS cell line A673 (upper panel) and EWS cell line EW3 (lower panel). An isotype
ions of n = 4 tumor biopsies taken at diagnosis were stained with antibodies againsts CD112 and CD155 on primary EWS tissue. Representative slide with HRP-DAB
mAbs did not stain decalcified paraffin sections, also not on tonsil tissue (positiveand paraffin embedded. Sections of n = 6 biopsies were stained with goat polyclonalprimary EWS tumors. Representative slides with HRP-DAB staining, magnification:he EWS marker CD99 (data not shown).
cacy, by IL-15-activated NK cells from healthy donors. The lysis ofEWS by NK cells was dependent on the activating NK cell receptors
NKG2D and DNAM-1, whose ligands are commonly expressed byEWS cell lines and primary EWS tumor cells. At diagnosis and priorto chemotherapy, patients suffering from Ewing sarcoma displayreduced NK cell cytotoxicity despite normal numbers of NK cellsand adequate NKG2D expression.Only few reports deal with NK cell reactivity to Ewing sarcomacell lines (Chin et al., 1989; Atzpodien et al., 1988; Staege et al.,2004). We confirm the observation that cytokine-activated NK cellsare more efficient in recognizing EWS cell lines compared to rest-ing NK cells. In this report, we identify NKG2D and DNAM-1 as twomajor pathways in the recognition of EWS cell lines. Due to thenature of this (bone) tumor, it is not possible to evaluate NK cellcytotoxicity towards unmanipulated EWS cells. Attempts to gener-ate short-term cultures of EWS from diagnostic biopsies have thusfar not resulted in stable cultures.
The ligands for NKG2D are expressed on some healthy tis-sues and hematopoietic precursor cells, but they are most notablyexpressed on stressed, infected or transformed cells (Raulet, 2003;Coudert and Held, 2006). Since NKG2D ligands are expressed inresponse to stress stimuli, they can function as danger signalsto NK cells and T cells. In contrast, the physiological relevance

3922 D.H.J. Verhoeven et al. / Molecular Immunology 45 (2008) 3917–3925
Fig. 3. HLA class I expression is not a predominant factor in lysis of EWS cell lines. (A) To evaluate whether a low levels of expression of HLA class I (loss of inhibition)contributed to NK cell mediated lysis of EWS cell lines, HLA class I was induced on the HLA class I negative EWS cell line SKES-1 by incubating with IFN�. Histograms forHLA class I staining (G46-2.6 mAb, black line) and isotype controls (gray line) are shown for SKES-1 and the EBV B-LCL 107 cell line, demonstrating comparable levels ofexpression. (B) Cytotoxic activity of purified resting NK cells (left panel) or IL-15-activated NK cells (right panel) towards K562 and SKES-1 with (�) or without (�) IFN�treatment. Error bars indicate standard error of the mean (S.E.M.) of triplicates. A representative experiment is shown of five experiments using different NK cell donors. (C)Cytotoxic activity of purified resting NK cells (left panel) or IL-15-activated NK cells (right panel) to EWS cell line SKES-1 treated with IFN� (�) and with additional blockingof HLA class I using the w6/32 mAb (�). Error bars indicate S.E.M. of triplicates. A representative experiment is shown of three experiments using different NK cell donors.Additionally, blocking HLA class I on the HLA class I positive cell line A673 did not decrease lysis by NK cells (data not shown).
Fig. 4. NK cell cytotoxicity to Ewing sarcoma is dependent on the activating receptors NKG2D and DNAM-1. The dependency of either (A) resting purified NK cells or (B)IL-15-stimulated NK cells on NKG2D and DNAM-1 receptors in the recognition and lysis of EWS cell lines was investigated by blocking these receptors with monoclonalantibodies prior to chromium release assays. (A) Blocking either NKG2D (�) or DNAM-1 (�) on purified resting NK cells (>90% NK cells) abrogated NK cell cytotoxicity to EWScell line EW3 (left panel). An NK cell binding, non-blocking, anti-CD56 mAb was used as an isotype control (♦), which resulted in the same degree of cytotoxicity comparedto NK cells without antibodies (�). In the middle and right panels, dependency on NKG2D and DNAM-1 pathways is shown using three additional EWS cell lines as targetsfor non-activated NK cells. Error bars indicate S.E.M. (B) Single blocking of NKG2D (�) and DNAM-1 (�) receptors on IL-15-activated NK cells (>99% NK cells) resulted in aless pronounced reduction of NK cell cytotoxicity to EW3, whereas combined blocking of both receptor pathways (�) efficiently reduced NK cell cytotoxicity (left panel). Thisobservation was confirmed using three additional EWS cell lines as targets for IL-15-activated NK cells (middle and right panels). Representative results are shown from datausing at least three different NK cell donors for each NK cell–EWS combination.

D.H.J. Verhoeven et al. / Molecular Immunology 45 (2008) 3917–3925 3923
osis. (iagnoltane
ey U-t(p = 0.class
toxici
Fig. 5. NK cell cytotoxic activity is reduced in Ewing sarcoma patients at time of diagnby performing chromium release assays using non-stimulated PBMC obtained at dpanel, �). PBMC from age matched healthy controls (left panel, �) were tested simuthe percentage of NK cells present in the PBMC. Statistical analysis (Mann–Whitn(LU-25)/106 NK cells), demonstrating a reduced cytotoxic activity in EWS patientsusing the HLA class I negative NK cell target K562 (p = 0.005, right panel) and the HLAfrom four EWS patients with 10 ng/ml IL-15 for 3 days restored reduced NK cell cyto
age matched controls. (C) Expression of NKG2D is increased, rather than reduced on NKSince all NK cells were positive for NKG2D, surface expression was measured as mean flu(left panel) and CD56dim NK cell (right panel) subpopulations were analyzed separately. SU-test. (D) To evaluate whether NKG2D and DNAM-1 pathways were completely impairedblocked with either anti-CD56 isotype control (♦), anti-NKG2D (�) or anti-DNAM-1 (�)Results for two individual patients are shown in the left and right panels, indicating thatactivity of NK cells in EWS patients. Error bars indicate S.E.M.of the DNAM-1 derived signal in NK cell biology is less clear.Both PVR (CD155) and Nectin-2 (CD112) are expressed in manyhealthy tissues, including neuronal, endothelial and epithelialtissues (Moretta et al., 2006). They play a role as homophilic inter-cellular adhesion molecules and are known to be the entry receptorfor the alpha-herpesviridae family (reviewed in Campadelli-Fiumeet al., 2000). Furthermore, expression of DNAM-1 ligands at vascu-lar endothelial junctions was shown to be critical for the monocytetransmigration process through the endothelium (Reymond et al.,2004). In agreement with previous observations in neuroblastoma,primary ovarian carcinoma and leukemia (Castriconi et al., 2004;Pende et al., 2005; Carlsten et al., 2007), we demonstrate expressionof CD112 and CD155 on primary Ewing sarcoma and that DNAM-1is a critical receptor for NK cell cytotoxicity to tumor cells.
A) The capacity of NK cells from EWS patients to lyse tumor targets was investigatedsis and prior to chemotherapy as effector cells against the EWS cell line A673 (leftously with PBMC from EWS patients. Effector:target (E:T) ratios were corrected forest) was performed on log10 values of cytotoxic activity (expressed as Lytic Units001, middle panel). The observed reduction in NK cell cytotoxicity was confirmedI negative EWS cell line SKES-1 (Fig. 5B, left panel, p = 0.039). (B) Incubation of PBMCty to the targets SKES-1 (left panel) and K562 (right panel) compared to PBMC from
cells from EWS patients compared to NK cells from age matched healthy controls.orescence intensity (MFI) determined by flow cytometric analysis. Both CD56brighttatistical analysis was performed on log10 values of MFI using the Mann–Whitneyin NK cells from EWS patients, unstimulated PBMC from three EWS patients were
mAbs prior to chromium release assays using the EWS cell line TC32 as a target.the NKG2D as well as the DNAM-1 pathway contributed to the residual cytotoxic
The role of DNAM-1 in activation of NK cells has been inves-tigated in studies defining the signals for activation of restingNK cells. These studies conclude that DNAM-1 has a synergis-tic effect on other activating receptor systems during activationof resting NK cells, but that the DNAM-1 derived signal alone isinsufficient to trigger a calcium influx in NK cells (reviewed inBryceson et al., 2006a). The observation, that blockade of eitherNKG2D or DNAM-1 abrogates cytotoxicity of resting NK cells to EWScells, suggests that these receptors transduce activating signalsthrough synergistic signaling pathways. In contrast, the cytotoxicityof cytokine-activated NK cells to EWS cells could only be reduced bysimultaneously blocking NKG2D and DNAM-1. This could suggestthat the threshold to trigger cytotoxic activity is decreased in thesecells, for example, by modifying intracellular signaling pathways.

ar Imm
3924 D.H.J. Verhoeven et al. / MoleculAn alternative explanation could be that signaling through each ofthese receptors has increased, which is supported by the observa-tion that the expression of both receptors is upregulated upon IL-15stimulation (data not shown).
It has been demonstrated that simultaneous crosslinking ofNKG2D and DNAM-1 on resting NK cells was not sufficient to resultin a major calcium influx. DNAM-1 or NKG2D crosslinking syn-ergized with other receptors (e.g., NKp46 or 2B4) in inducing anefficient calcium influx in resting NK cells (Bryceson et al., 2006b).This suggests that a third activating signal, delivered by Ewing sar-coma tumor cells, may be involved in triggering NK cells next toNKG2D and DNAM-1.
Loss of expression of HLA class I on EWS cells does not seem toplay a predominant role in NK cell recognition, since EWS cell linesdisplayed comparable sensitivity to lysis by NK cells despite het-erogeneous expression of HLA class I. Furthermore, upregulation ofHLA class I on the EWS cell line SKES-1 (which has a genotype com-prising all known HLA class I ligands for inhibitory KIR receptors)with interferon-� did not result in a significant decrease in NK cellcytotoxicity. Furthermore, blocking of (induced) HLA class I on theEWS cell line SKES-1 (Fig. 3) as well as A673 (data not shown) bymasking with the w6/32 mAb did not significantly increase NK cellcytotoxicity. This observation might be explained by the expressionof (a combination of) activating NK cell receptor ligands on EWS celllines, which provides a strong activating signal, not easily counter-balanced by inhibitory signals. However, it cannot be excluded thatan excess expression of HLA class I on EWS tumors in individualcases results in decreased sensitivity to lysis by resting NK cells.Nevertheless, the observation that primary EWS cells also expressactivating ligands indicates that activating signals are also presentin vivo on EWS tumor cells. The fact that they have the same activat-ing receptor ligand expression pattern compared to EWS cell lines,might suggest an equal sensitivity to lysis by NK cells.
Impaired NK cell function has not been previously described inpatients suffering from Ewing sarcoma. The mechanism of NK cellimpairment in the present study does not involve downregulationof NKG2D, in contrast to what has been reported in several othertumor models (Hayakawa and Smyth, 2006; Coudert and Held,2006). The expression of NKG2D is upregulated rather than down-regulated on NK cells from EWS patients at diagnosis and NKG2D aswell as DNAM-1 function is not completely impaired since it con-tributes to the (residual) EWS patient NK cell cytotoxicity to EWScell lines. However, it cannot be excluded that signals downstreamof the NKG2D or DNAM-1 receptor are impaired in NK cells from
EWS patients.Other mechanisms, such as direct downregulation of effectormolecules and the contribution of tumor produced factors (such asTFG�) and/or regulatory T cells, could contribute to the observeddecreased NK cell activity and will be the subject of future investi-gation.
Since Ewing sarcoma cell lines are susceptible to allogeneicNK cell mediated lysis, this tumor may be a target for NK cellimmunotherapy. A role of alloreactive NK cells in anti-tumorresponses has been suggested in particular in a haploidenticalstem cell transplantation setting for the treatment of hematologictumors (Ruggeri et al., 2002). Since then, several phase I/II tri-als have been initiated, in which alloreactive allogeneic NK cellsare infused in cancer patients (Miller et al., 2005; Passweg et al.,2006). These trials demonstrated the feasibility and safety of theseinfusions, but therapeutic efficacy has to be evaluated in largephase II/III trials. An advantage of this therapeutic approach is thatit allows ex vivo activation of NK cells by cytokines, which mayimprove efficacy of NK cell reactivity in EWS patients. Future stud-ies are required to evaluate the anti-tumor potential and migratoryproperties of adoptively transferred NK cells. NK cells may serve as
unology 45 (2008) 3917–3925
direct effector cells, but they could also enhance cellular immuneresponses by their interaction with dendritic cells. The expandingknowledge of tumor and NK cell biology may provide importantclues how to best target Ewing sarcoma, especially in patients inwhom the tumor has relapsed or metastasized.
Acknowledgements
The authors wish to thank Salvatore Romeo and Ekatherina Jor-danova (Department of Pathology, LUMC, Leiden) for assistancewith immunohistochemistry.
This research is funded by grants from the Dutch PediatricCancer Foundation KiKa (to RME, MJDvT), The Netherlands Orga-nization for Health Research and Development (ZonMw, grant920-03-267; to DHJV), the Leiden University Medical Center (toDHJV) and the Dutch Cancer Society NKB/KWF (grant no. RUL 2003-2800; to ACL). The Departments of Pediatrics and Pathology (LUMC,Leiden, Netherlands) are partners of the EuroBoNeT consortium, aEuropean Commission granted Network of Excellence for studyingthe pathology and genetics of bone tumors.
References
Atzpodien, J., Gulati, S.C., Shimazaki, C., Buhrer, C., Oz, S., Kwon, J.H., Kolitz, J.E.,Clarkson, B.D., 1988. Ewing’s sarcoma: ex vivo sensitivity towards natural andlymphokine-activated killing. Oncology 45, 437–443.
Bernstein, M., Kovar, H., Paulussen, M., Randall, R.L., Schuck, A., Teot, L.A., Juergens,H., 2006. Ewing’s sarcoma family of tumors: current management. Oncologist11, 503–519.
Bottino, C., Castriconi, R., Pende, D., Rivera, P., Nanni, M., Carnemolla, B., Cantoni, C.,Grassi, J., Marcenaro, S., Reymond, N., Vitale, M., Moretta, L., Lopez, M., Moretta,A., 2003. Identification of PVR (CD155) and Nectin-2 (CD112) as cell surfaceligands for the human DNAM-1 (CD226) activating molecule. J. Exp. Med. 198,557–567.
Bryceson, Y.T., March, M.E., Ljunggren, H.G., Long, E.O., 2006a. Activation, coactiva-tion, and costimulation of resting human natural killer cells. Immunol. Rev. 214,73–91.
Bryceson, Y.T., March, M.E., Ljunggren, H.G., Long, E.O., 2006b. Synergy among recep-tors on resting NK cells for the activation of natural cytotoxicity and cytokinesecretion. Blood 107, 159–166.
Burchill, S.A., 2003. Ewing’s sarcoma: diagnostic, prognostic, and therapeutic impli-cations of molecular abnormalities. J. Clin. Pathol. 56, 96–102.
Campadelli-Fiume, G., Cocchi, F., Menotti, L., Lopez, M., 2000. The novel receptorsthat mediate the entry of herpes simplex viruses and animal alphaherpesvirusesinto cells. Rev. Med. Virol. 10, 305–319.
Carayannopoulos, L.N., Yokoyama, W.M., 2004. Recognition of infected cells by nat-ural killer cells. Curr. Opin. Immunol. 16, 26–33.
Carlsten, M., Bjorkstrom, N.K., Norell, H., Bryceson, Y., van Hall, T., Baumann, B.C.,Hanson, M., Schedvins, K., Kiessling, R., Ljunggren, H.G., Malmberg, K.J., 2007.DNAX accessory molecule-1 mediated recognition of freshly isolated ovarian
carcinoma by resting natural killer cells. Cancer Res. 67, 1317–1325.Castriconi, R., Dondero, A., Corrias, M.V., Lanino, E., Pende, D., Moretta, L., Bottino,C., Moretta, A., 2004. Natural killer cell-mediated killing of freshly isolated neu-roblastoma cells: critical role of DNAX accessory molecule-1-poliovirus receptorinteraction. Cancer Res. 64, 9180–9184.
Chin, T., Toy, C., Vandeven, C., Cairo, M.S., 1989. Lymphokine-activated killer cytotox-icity in neonatal mononuclear cells: in vitro responses to tumor cell lines frompediatric solid tumors. Pediatr. Res. 25, 156–160.
Costello, R.T., Sivori, S., Marcenaro, E., Lafage-Pochitaloff, M., Mozziconacci, M.J., Rev-iron, D., Gastaut, J.A., Pende, D., Olive, D., Moretta, A., 2002. Defective expressionand function of natural killer cell-triggering receptors in patients with acutemyeloid leukemia. Blood 99, 3661–3667.
Cotterill, S.J., Ahrens, S., Paulussen, M., Jurgens, H.F., Voute, P.A., Gadner, H., Craft,A.W., 2000. Prognostic factors in Ewing’s tumor of bone: analysis of 975 patientsfrom the European Intergroup Cooperative Ewing’s Sarcoma Study Group. J. Clin.Oncol. 18, 3108–3114.
Coudert, J.D., Held, W., 2006. The role of the NKG2D receptor for tumor immunity.Semin. Cancer Biol. 16, 333–343.
Delattre, O., Zucman, J., Melot, T., Garau, X.S., Zucker, J.M., Lenoir, G.M., Ambros,P.F., Sheer, D., Turc-Carel, C., Triche, T.J., 1994. The Ewing family of tumors—asubgroup of small-round-cell tumors defined by specific chimeric transcripts.N. Engl. J. Med. 331, 294–299.
Gasser, S., Raulet, D., 2006. The DNA damage response, immunity and cancer. Semin.Cancer Biol. 16, 344–347.
Hayakawa, Y., Smyth, M.J., 2006. NKG2D and cytotoxic effector function in tumorimmune surveillance. Semin. Immunol. 18, 176–185.
Miller, J.S., Soignier, Y., Panoskaltsis-Mortari, A., McNearney, S.A., Yun, G.H., Fautsch,S.K., McKenna, D., Le, C., Defor, T.E., Burns, L.J., Orchard, P.J., Blazar, B.R., Wagner,

ar Imm
D.H.J. Verhoeven et al. / MoleculJ.E., Slungaard, A., Weisdorf, D.J., Okazaki, I.J., McGlave, P.B., 2005. Successfuladoptive transfer and in vivo expansion of human haploidentical NK cells inpatients with cancer. Blood 105, 3051–3057.
Moretta, L., Bottino, C., Pende, D., Castriconi, R., Mingari, M.C., Moretta, A., 2006.Surface NK receptors and their ligands on tumor cells. Semin. Immunol. 18,151–158.
Parham, P., 2005. MHC class I molecules and KIRs in human history, health andsurvival. Nat. Rev. Immunol. 5, 201–214.
Passweg, J.R., Koehl, U., Uharek, L., Meyer-Monard, S., Tichelli, A., 2006. Natural-killer-cell-based treatment in haematopoietic stem-cell transplantation. Best.Pract. Res. Clin. Haematol. 19, 811–824.
Pende, D., Spaggiari, G.M., Marcenaro, S., Martini, S., Rivera, P., Capobianco, A., Falco,M., Lanino, E., Pierri, I., Zambello, R., Bacigalupo, A., Mingari, M.C., Moretta, A.,Moretta, L., 2005. Analysis of the receptor–ligand interactions in the naturalkiller-mediated lysis of freshly isolated myeloid or lymphoblastic leukemias:evidence for the involvement of the poliovirus receptor (CD155) and Nectin-2(CD112). Blood 105, 2066–2073.
Pogge von Strandmann, E., Simhadri, V.R., von Tresckow, B., Sasse, S., Reiners, K.S.,Hansen, H.P., Rothe, A., Boll, B., Simhadri, V.L., Borchmann, P., McKinnon, P.J.,Hallek, M., Engert, A., 2007. Human leukocyte antigen-B-associated transcript 3is released from tumor cells and engages the NKp30 receptor on natural killercells. Immunity 27, 965–974.
Raulet, D.H., 2003. Roles of the NKG2D immunoreceptor and its ligands. Nat. Rev.Immunol. 3, 781–790.
Reymond, N., Imbert, A.M., Devilard, E., Fabre, S., Chabannon, C., Xerri, L., Farnarier, C.,Cantoni, C., Bottino, C., Moretta, A., Dubreuil, P., Lopez, M., 2004. DNAM-1 andPVR regulate monocyte migration through endothelial junctions. J. Exp. Med.199, 1331–1341.
unology 45 (2008) 3917–3925 3925
Ruggeri, L., Capanni, M., Urbani, E., Perruccio, K., Shlomchik, W.D., Tosti, A., Posati, S.,
Rogaia, D., Frassoni, F., Aversa, F., Martelli, M.F., Velardi, A., 2002. Effectiveness ofdonor natural killer cell alloreactivity in mismatched hematopoietic transplants.Science 295, 2097–2100.Shibuya, A., Campbell, D., Hannum, C., Yssel, H., Franz-Bacon, K., McClanahan, T.,Kitamura, T., Nicholl, J., Sutherland, G.R., Lanier, L.L., Phillips, J.H., 1996. DNAM-1,a novel adhesion molecule involved in the cytolytic function of T lymphocytes.Immunity 4, 573–581.
Shibuya, A., Lanier, L.L., Phillips, J.H., 1998. Protein kinase C is involved in the regu-lation of both signaling and adhesion mediated by DNAX accessory molecule-1receptor. J. Immunol. 161, 1671–1676.
Shibuya, K., Lanier, L.L., Phillips, J.H., Ochs, H.D., Shimizu, K., Nakayama, E., Nakauchi,H., Shibuya, A., 1999. Physical and functional association of LFA-1 with DNAM-1adhesion molecule. Immunity 11, 615–623.
Staege, M.S., Hansen, G., Baersch, G., Burdach, S., 2004. Functional and molecu-lar characterization of interleukin-2 transgenic Ewing tumor cells for in vivoimmunotherapy. Pediatr. Blood Cancer 43, 23–34.
Szuhai, K., IJszenga, M., Tanke, H.J., Rosenberg, C., Hogendoorn, P.C., 2006. Molecu-lar cytogenetic characterization of four previously established and two newlyestablished Ewing sarcoma cell lines. Cancer Genet. Cytogenet. 166, 173–179.
Szuhai, K., IJszenga, M., Tanke, H.J., Taminiau, A.H., de Schepper, A., van Duinen,S.G., Rosenberg, C., Hogendoorn, P.C., 2007. Detection and molecular cytogeneticcharacterization of a novel ring chromosome in a histological variant of Ewingsarcoma. Cancer Genet. Cytogenet. 172, 12–22.
Urano, F., Umezawa, A., Yabe, H., Hong, W., Yoshida, K., Fujinaga, K., Hata, J., 1998.Molecular analysis of Ewing’s sarcoma: another fusion gene, EWS-E1AF, avail-able for diagnosis. Jpn. J. Cancer Res. 89, 703–711.




















