
Musculoskeletal Differentiation of Cells Derived from Human Embryonic Germ Cells
11
Stem Cells 2005;23:113–123 www.StemCells.com O riginal A rticle Musculoskeletal Differentiation of Cells Derived from Human Embryonic Germ Cells Myoung Sook Kim, a Nathaniel S. Hwang, a Janice Lee, a Tae-Kyun Kim, b Kam Leong, a Michael J. Shamblott, c John Gearhart, c Jennifer Elisseeff a a Department of Biomedical Engineering, Johns Hopkins University, Baltimore, Maryland, USA; b Seoul National University School of Medicine, Seoul, Korea; c Department of Gynecology and Obstetrics, Johns Hopkins University, School of Medicine, Baltimore, Maryland, USA Key Words. Pluripotent stem cells • Embryoid bodies • EG cells • Differentiation Correspondence: Jennifer Elisseeff, Ph.D., Department of Biomedical Engineering, Johns Hopkins University, 3400 N. Charles Street, Clark 106, Baltimore, Maryland 21218,USA. Telephone: 410-516-4915; Fax: 410-516-8152; e-mail: [email protected] Received May 13, 2004; accepted for publication August 9, 2004. ©AlphaMed Press 1066–5099/2005/$12.00/0 doi: 10.1634/stemcells.2004–0110 Abstract Stem cells have the potential to significantly improve cell and tissue regeneration therapies, but little is understood about how to control their behavior. We investigated the potential differentiation capability of cells derived from human embryonic germ (EG) cells into musculoskeletal lineages by providing a three-dimensional environment with increased cell–cell contact and growth factors. Cells were clustered into pellets to mimic the mesenchyme condensation process during limb development. LVEC cells, an embryoid body–derived (EBD) cell culture generated from EG cells, were cultured in micromass pellets for 21 days in the presence of bone morphoge- netic protein 2 (BMP2) and/or transforming growth factor beta-3 (TGFβ3). Gene expression for cartilage-, bone-, and muscle-specific matrix proteins—including collagen types I, II, III, IX, X; aggrecan; cartilage pro- teoglycan link protein; cartilage oligomeric protein; chondroitin sulfate-4-S; and myf5—was upregulated in the pellets treated with TGFβ3, while mRNAs for neu- rofilament heavy (NFH), a neuron marker, and flk-1, a hematopoietic marker, decreased. Total collagen and proteoglycan production exhibited a time-dependent increase in the pellets treated with TGFβ3, further con- firming the expression of characteristic musculosk- eletal markers. Furthermore, our results indicate the ability to select or differentiate stem cells toward a mus- culoskeletal lineage from a heterogenous EBD cell line. Stem Cells 2005;23:113–123 Introduction Stem cells have significant potential to positively affect the biomedical sciences, particularly in regen- erative medicine. Human pluripotent embryonic stem (ES) cells have been derived from the inner cell mass of blastocysts and from primordial germ cells coloniz- ing the developing gonadal ridge; they are referred to as ES cells and embryonic germ (EG) cells, respectively [1, 2]. These cells are being studied to understand early development and to formulate cell-and tissue-regeneration therapies. Currently, we have limited knowledge about how to control stem cell behavior to develop therapies, particularly in the area of the musculoskeletal system. When ES and EG cells differentiate in vitro, they are often in the form of complex three-dimensional spheres termed embryoid bodies (EBs) [3]. Early developmental processes occur within the EBs, resulting in collections of precursor and differentiated cells from a wide variety of
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Musculoskeletal Differentiation of Cells Derived from Human Embryonic Germ Cells

Stem Cells 2005;23:113–123 www.StemCells.com
Original Article
Musculoskeletal Differentiation of Cells Derived from
Human Embryonic Germ Cells
Myoung Sook Kim,a Nathaniel S. Hwang,a Janice Lee,a Tae-Kyun Kim,b Kam Leong,a Michael J. Shamblott,c John Gearhart,c Jennifer Elisseeffa
aDepartment of Biomedical Engineering, Johns Hopkins University, Baltimore, Maryland, USA; bSeoul National
University School of Medicine, Seoul, Korea; cDepartment of Gynecology and Obstetrics,
Johns Hopkins University, School of Medicine, Baltimore, Maryland, USA
Key Words. Pluripotent stem cells • Embryoid bodies • EG cells • Differentiation
Correspondence: Jennifer Elisseeff, Ph.D., Department of Biomedical Engineering, Johns Hopkins University, 3400 N. Charles Street, Clark 106, Baltimore, Maryland 21218,USA. Telephone: 410-516-4915; Fax: 410-516-8152; e-mail: [email protected] Received May 13, 2004; accepted for publication August 9, 2004. ©AlphaMed Press 1066–5099/2005/$12.00/0 doi: 10.1634/stemcells.2004–0110
AbstractStem cells have the potential to significantly improve cell and tissue regeneration therapies, but little is understood about how to control their behavior. We investigated the potential differentiation capability of cells derived from human embryonic germ (EG) cells into musculoskeletal lineages by providing a three-dimensional environment with increased cell–cell contact and growth factors. Cells were clustered into pellets to mimic the mesenchyme condensation process during limb development. LVEC cells, an embryoid body–derived (EBD) cell culture generated from EG cells, were cultured in micromass pellets for 21 days in the presence of bone morphoge-netic protein 2 (BMP2) and/or transforming growth factor beta-3 (TGFβ3). Gene expression for cartilage-,
bone-, and muscle-specific matrix proteins—including collagen types I, II, III, IX, X; aggrecan; cartilage pro-teoglycan link protein; cartilage oligomeric protein; chondroitin sulfate-4-S; and myf5—was upregulated in the pellets treated with TGFβ3, while mRNAs for neu-rofilament heavy (NFH), a neuron marker, and flk-1, a hematopoietic marker, decreased. Total collagen and proteoglycan production exhibited a time-dependent increase in the pellets treated with TGFβ3, further con-firming the expression of characteristic musculosk-eletal markers. Furthermore, our results indicate the ability to select or differentiate stem cells toward a mus-culoskeletal lineage from a heterogenous EBD cell line. Stem Cells 2005;23:113–123
IntroductionStem cells have significant potential to positively
affect the biomedical sciences, particularly in regen-
erative medicine. Human pluripotent embryonic stem
(ES) cells have been derived from the inner cell mass
of blastocysts and from primordial germ cells coloniz-
ing the developing gonadal ridge; they are referred to as
ES cells and embryonic germ (EG) cells, respectively
[1, 2]. These cells are being studied to understand early
development and to formulate cell-and tissue-regeneration
therapies. Currently, we have limited knowledge about how to
control stem cell behavior to develop therapies, particularly in
the area of the musculoskeletal system.
When ES and EG cells differentiate in vitro, they are
often in the form of complex three-dimensional spheres
termed embryoid bodies (EBs) [3]. Early developmental
processes occur within the EBs, resulting in collections of
precursor and differentiated cells from a wide variety of

114 Musculoskeletal Cell Differentiation
lineages, including cells of the hematopoietic lineage, neu-
rons, and skeletal muscle. Differentiation of cells in EBs is
generally random and uncontrolled, making the resulting
cells difficult to apply to regenerative strategies.
One of the most important challenges in stem cell
research is to understand and control cell-differentiation
processes. EB–derived (EBD) cells are generated from the
disaggregation of EBs and by selection of cells that prolif-
erate well in six basic growth conditions [4]. One example,
the LVEC culture, was established on type I collagen-coated
tissue-culture dishes in the presence of 5% serum, basic
fibroblast growth factor (bFGF), insulin-like growth factor-
I (IGF-I), vascular endothelial cell growth factor (VEGF),
and epidermal growth factor (EGF). EBD cells are a mixed
cell culture, containing precursors and progenitor cells of
various lineages, and a majority of clonal EBD cell lines
retain the expression of multiple lineages. Unlike their ES
and EG precursors, EBD cells are not immortal and do not
form teratomas when injected into mice, making them a
potentially useful therapeutic tool [4]. The first studies to
evaluate chondrogenic differentiation of ES cells used a
mouse line in monolayer culture. Kramer et al. [5, 6] dem-
onstrated expression of cartilage matrix proteins after expo-
sure to bone morphogenetic protein (BMP). More recent,
Levenberg et al. [7] seeded human ES cells on polyglycolic
acid scaffolds and differentiated the cells toward a cartilage
phenotype using transforming growth factor beta (TGFβ).
The potential of EBD cells to differentiate into musculoskel-
etal tissues is unknown.
The aim of the current study is to evaluate the differen-
tiation capability of human EBD cells (LVECs) using the
micromass pellet culture system. The three-dimensional
culture condition resembles more closely the in vitro sur-
roundings during EB differentiation [8]. Increased cell–cell
interaction of cell aggregates is designed to approximate the
cell environment of condensing mesenchymes in the devel-
oping limb in vivo, which is supported by the report that
mouse ES cells derived from EBs differentiated into chon-
drocytes, progressively developing into hypertrophic, calci-
fying cells [9]. The approach of micromass cultures has been
established as a conventional method for chondroinduction
of mesynchemal stem cells (MSCs) by disassociation of
cells that are pelleted for culture [10]. In addition, the pel-
let culture system has also been used to create cartilaginous
modules from primary cultured chondrocytes and embry-
onic mesenchymal cells.
Differentiation of LVECs was investigated in a chemi-
cally defined chondrogenic differentiation medium with
addition of two growth factors that promote chondrogenesis:
BMP2 and TGFβ3 [11]. Under these conditions, LVECs
lost expression of differentiated neural and vascular mark-
ers. Protein production of markers for mesenchymal tissues
was upregulated. Gene expression of cartilage-, bone-, and
muscle-specific matrix proteins—including collagen types
I, II, III, IX, X; aggrecan; cartilage proteoglycan link pro-
tein; cartilage oligomeric protein; chondroitin sulfate-4-S;
and myf5—was upregulated, demonstrating the ability of
LVEC selection and transdifferentiation to cells of mesen-
chymal lineages.
Materials and Methods
LVEC CultureLVEC human EBD cells were cultured as previously
described [8]. EGM2MV medium (Clonetics, San Diego,
http://www.informagen.com) included 5% fetal calf serum,
hydrocortisone, human bFGF, human VEGF, R3 IGF-I,
ascorbic acid, human EGF, heparin, gentamycin, and ampho-
tericin. Tissue culture plastic was coated with bovine type
I collagen (10 μg/cm2; Collaborative Biomedical Products,
Bedford, MA, http://www.bioscience.org/company/becton.
htm). Cells were cultured at 37°C, 5% CO2, and 95% humid-
ity and then routinely passaged 1:10 to 1:40 using 0.025%
trypsin, 0.01% EDTA (Clonetics) for 5 minutes at 37°C.
Micromass CultureSubconfluent (70%–80%) undifferentiated LVECs (pas-
sage 12–15) were resuspended at a density of 2 × 105 cells/ml
in growth medium; then 1-ml aliquots of the cell suspension
were dispensed into 15-ml sterile, conical polypropylene
culture tubes (Sarstedt, Nümbrecht, Germany, http://www.
sarstedt.com) and centrifuged at 1,000 rpm for 5 minutes
to form spherical pellets [10]. The pellets were cultured at
37°C with 5% CO2 in 1 ml of chondrogenic differentiation
medium that contained 1% serum in addition to high-glu-
cose Dulbecco’s modified Eagle’s medium supplemented
with 10–7 M dexamethasone, 50 μg/ml ascorbate-2-phos-
phate, 40 μg/ml proline, 100 μg/ml sodium pyruvate, and
50 mg/ml ITS+Premix (Collaborative Biomedical: 6.25
µg/ml insulin, 6.25 µg/ml transferrin, 6.25 ng/ml selenious
acid, 1.25 mg/ml bovine serum albumin, and 5.35 mg/ml
linoleic acid). After a pellet was formed, the medium in each
tube was replaced with 0.5 ml defined medium containing
either 20 ng/ml of recombinant human BMP2 or 10 ng/ml
of TGFβ3 (Research Diagnostics, Inc., Flanders, NJ, http://
www.researchd.com), or both. Control cultures were main-
tained without adding BMP2 or TGFβ3. Culture medium
was changed three times per week. The pellets were har-
vested periodically over 3 weeks.

Kim, Hwang, Lee et al. 115
Biochemical AnalysisProtein synthesis was determined by [35S]-methionine incor-
poration. DNA content, collagen, and proteoglycan deposi-
tion were characterized using 33258 Hoechst dye, hydroxy-
proline content, and dimethylmethylene blue (DMMB)
spectrophotometric assay, respectively [12]. To investigate
whether the increase of pellet size and weight might be a pos-
sible effect of TGFβ3 on cell proliferation, tetrazolium-based
cell growth analysis was performed in LVECs cultured in
monolayer with or without BMP2 or TGFβ3 for 3 weeks.
Histology and Immunostaining PreparationPellets were fixed overnight in 4% paraformaldehyde in
phosphate-buffered solution (PBS; pH 7.5) at 40°C and trans-
ferred to 70% ethanol until they were embedded in paraffin,
according to standard histological technique. Pellets were
processed, embedded in paraffin, and cut to 5 μm in thick-
ness. Serial sections were stained with hematoxylin and
eosin (H&E) for morphology, Toluidine blue for pericellular
proteoglycan, and Masson’s trichrome for collagen detec-
tion. For immunostaining, sectioned pellets were deparaf-
finized and incubated for 30 minutes in goat serum followed
by incubation with rabbit anti-type I and anti-type X collagen
(Research Diagnostics). After washing in PBS, the sections
were incubated for 1 hour with fluorescein isothiocyanate–
conjugated or Texas Red dye–conjugated anti-rabbit second-
ary antibodies (Jackson ImmunoResearch Laboratories, West
Grove, Pennsylvania; http://www.jacksonimmuno.com).
Specimens were washed in PBS, and coverslips were
mounted in glycerol-vinyl alcohol–mount (Zymed Labora-
tories, San Francisco, http://www.zymed.com).
Reverse Transcription Polymerase Chain Reaction (RT-PCR)Total RNA was extracted from 5 million of undifferentiated
LVECs at day 0 or from 50 pellets at given time points with
TRIzol (Invitrogen, Carlsbad, CA, http://www.invitrogen.
com), following the manufacturer’s instructions. RNA was
treated with DNase I using the DNA-free Kit (Invitrogen).
Two micrograms of total RNA per 20 μ1 of reaction volume
was reverse transcribed into cDNA using the SuperScript
First-Strand Synthesis System (Invitrogen). The PCR prim-
ers are provided in online supplementary Table 1. The condi-
tions for all PCRs in this study were 35 cycles of 95°C for 30
seconds, 57°C to 60°C for 45 seconds, and 72°C for 1 minute,
with one exception of 18s rRNA (12 cycles of 95°C for 30 sec-
onds, 55°C for 30 seconds, and 72°C for 1 minute). PCR
products were separated by electrophoresis at 100 V on a
1.4% agarose gel in Tris-acetate-EDTA buffer, then visu-
alized with ethidium bromide staining.
Statistical AnalysisStatistical significance was determined within groups using
Student’s t-test. Data are presented as the mean � standard
deviations. The level of significance was set at p < .05. Each
measurement reported here was from at least two indepen-
dent pellet group preparations.
Results
Gene Expression Profiles for Differentiating Human EBD CellsIncreasing cell–cell contact by culturing the LVECs in three-
dimensional micromass configuration significantly altered
their gene-expression profiles. Previous reports show that
LVECs strongly express the neural progenitor marker nestin
and, to a lesser extent, neurofilament heavy chain (NFH) [4].
The cells also express the vascular-hematopoietic stem cell
marker flk-1 and show strong alkaline phosphatase (ALP)
activity that can be a marker of pluripotent stem cells [4].
In comparison with undifferentiated LVECs in monolayer
culture (Fig. 1A), LVECs in pellets significantly reduced the
expression of these neural and vascular endothelial markers.
NFH, flk-1, and ALP expressions decreased in control cul-
tures of pellets, whereas nestin expression showed little dif-
ference, indicating that LVECs might have lost, in part, their
original characteristics in monolayer culture when cultured
in micromass conditions.
LVEC pellets cultured in the presence of selected growth
factors increased expression of markers for mesenchymal tis-
sues, including muscle, bone, cartilage, ligament, and tendon.
When LVEC pellets were incubated with potent osteochon-
drogenic growth factors of mesenchymal precursors, BMP2
(20 ng/ml) and/or TGFβ3 (10 ng/ml), NFH, flk-1, and nestin
expression remained similar to that in control pellets without
exposure to growth factor (Fig. 1A). The skeletal muscle cell-
specific marker myf5, which was hardly detected in mono-
layer or in control pellets, was clearly expressed in BMP2
and slightly expressed in TGFβ3 cultures to suggest differ-
entiation toward this musculoskeletal tissue (Fig. 1A). Also,
the basic helix-loop-helix transcription factors, Twist-a and
Twist-b, which are required for somatic myogenesis [13] and
bone formation [14], were upregulated by all pellet cultures
compared with undifferentiated cells (online supplementary,
Table 1; not shown). While ALP is a marker for undifferen-
tiated stem cells, it is also a characteristic marker found in
bone. ALP expression, quantified by real time PCR, mark-
edly increased in pellets exposed to growth factors (Fig. 1A,
1D), and this is consistent with the activity measured in the
pellets biochemically (Fig. 1D).
One important feature of MSCs, chondrocytes, and
fibrochondrocytes is their high expression level of genes for

116 Musculoskeletal Cell Differentiation
specific types of collagen, which forms the major structural
components of bone, cartilage, and other connective tissues.
Therefore, we investigated the expression of genes involved
in the chondrogenic differentiation of LVEC pellets. Type I
collagen (Col I), which is found in bone, tendon, ligament,
and meniscal cartilage, was expressed by undifferentiated
cells, but was significantly upregulated by BMP2 and/or
TGFβ3 treatments (Fig. 1B, 1D). Both immature (Col α1(I))
and mature collagen type I are expressed in developing car-
tilage. In the study reported here, it was revealed that expres-
sions of Col α1(I) and Col I increased with pellet formation
and TGFβ3 treatment. Type III collagen (Col III) resides
at the insertion sites of the major ligaments and tendons
and within the perichondrium and periosteum and is also
involved in early chondrogenesis. Its expression was read-
ily detected in undifferentiated monolayer LVECs and was
upregulated by the pellet culture of the cells (Fig. 1B). The
expression, however, was not elevated with further treat-
ment of BMP2 or TGFβ3. Detectable levels of type II colla-
gen mRNA (Col II), the primary collagen specific to hyaline
cartilage, was observed upon differentiation and strongly
increased in TGFβ3-treated pellets (Fig. 1B). Furthermore,
the juvenile splice variant of type II collagen (Col IIA), was
not observed in cells in monolayer but was expressed after
Figure 1. Regulation of mRNA expression for extracellular matrix components determined by RT-PCR during chondrogenic dif-ferentiation of LVECs. Undifferentiated LVECs were cultured in a pellet format for 21 days in the absence or in the presence of either BMP2 (20 ng/ml) or TGFβ3 (10 ng/ml), or both. Five million undifferentiated LVECs in monolayer culture were harvested on day 0, and 50 pellets (2 × 105 cells/pellet) were harvested on day 21. (A): mRNA expression of genes involved in distinct developmental lineages of human EG cells. (B): mRNA expression of cartilage-specific matrix proteins upon differentiation. Lane 1: undifferenti-ated cells (UD); lane 2: control pellets without growth factor treatment (C); lane 3: BMP2-treated pellets (B); lane 4: TGFβ3-treated pellets (T); lane 5, BMP2 and TGFβ3 co-treated pellets (BT). Actin was used as a loading control for RT-PCR. (C): Time course for musculoskeletal differentiation of LVECs. Lane 1: undifferentiated cells (UD); lane 2: control pellet 7d (C7); lane 3: TGFβ3 pellet 7d (T7); lane 4: control pellet 14d (C14); lane 5: TGFβ3 pellet 14d (T14). (D): Real-time PCR, using SYBR green fluorescence, was used to quantify the mRNA of ALP and Col I. Abbreviations: AGN, aggrecan; ALP, alkaline phosphatase; BMP2, bone morphogenetic protein; Col I, type I collagen; COMP, cartilage oligomeric protein; LP, proteoglycan link protein; LVEC, large-vessel endothelial cell; NFH, neurofilament heavy chain; flk-1, fetal liver kinase–1; RT-PCR, reverse transcription polymerase chain reaction; TGFβ3, transforming growth factor beta-3.

Kim, Hwang, Lee et al. 117
treatment with BMP2 and was more clearly upregulated by
TGFβ3 (Fig. 1B). The adult cartilage splice variant of col-
lagen (IIB), which is activated in mature chondrocytes, was
not found in any of the cultures.
Cartilage-Specific Gene Expression in the Presence of TGFβ3Specific markers for cartilage, a tissue that cannot repair and
is a frequent target for regenerative therapies, were upregu-
lated in the micromass culture system. Col IX is cartilage
specific, and Col X is expressed in hypertrophic chondro-
cytes and bone [15]. These markers were not expressed in
undifferentiated cells but underwent a rapid increase in the
pellet cultures (Fig. 1B, 1C). The expression of cartilage
oligomeric protein (COMP) that is characteristic for devel-
oping chondrocytes was dramatically induced by pellet
culture of the cells, whereas LVECs in monolayer culture
showed no detectable expression of COMP (Fig. 1B). These
markers increased the most in TGFβ3-treated pellets. In
contrast, cartilage proteoglycan link protein (LP) and one
of the chondroitin sulfate proteoglycans, aggrecan (AGN)
were not markedly increased by TGFβ3 treatment in pel-
lets compared with control pellets (Fig. 1B). LP was readily
observed, while a trace of AGN was seen in undifferentiated
LVECs. The RT-PCR results are summarized in Table 1.
For further validation of mesenchymal tissue lineage
progression, the mRNA level of genes involved in car-
tilage development was analyzed over time. Col IX and
AGN expressions were temporally increased by TGFβ3
treatment in comparison with control in 14 days of culture.
Chondromodulin (ChM), a protein abundant in cartilage
tissue, was minimally expressed in undifferentiated LVEC
culture but substantially increased expression in all pellet
cultures (Fig. 1C).
These results suggest that LVEC pellet exposure to
BMP2 or TGFβ3 promoted the loss of propensity to neural
and hematopoietic lineages and significantly enhanced the
expression of musculoskeletal tissue markers. Since LVEC is
Table 1. Summary of reverse transcription polymerase chain reaction results in Figure 1
Expression levels
Gene UDa Controlb BMP2c TGFβ3d BMP2 + TGFβ3e
NFH +++ + + + +
flk-1 +++ + + + +
ALP +++ + +++ ++ ++
myf5 – – ++++ ++ ++
nestin +++ +++ +++ +++ +++
Col I + ++ +++ +++ ++
Col II – + ++ ++ ++
Col IIA – – + ++ nd
Col III ++ +++ +++ +++ +++
Col IX – ++ ++ +++ nd
Col X – ++ ++ +++ ++
AGN + ++ ++ ++ nd
COMP – +++ +++ +++ +++
LP +++ +++ +++ +++ nd
aUndifferentiated cells.bControl pellets without growth factor treatment. cBMP2-treated pellets. dTGFβ3-treated pellets. eBMP2 and TGFβ3 co-treated pellets. –, hardly detectable; +, weak; __, moderate; +++, strong; nd, not determined.Abbreviations: BMP2, bone morphogenetic protein; TGFβ3, transforming growth factor beta-3.

118 Musculoskeletal Cell Differentiation
a mixed cell culture, this could mean that the precursor cells
to other lineages died off or that the entire population moved
toward the musculoskeletal lineages. The experimental
design in the pellet culture system could not distinguish
between these two possibilities. However, the homogenous
immunostaining of types I and X collagen in the pellets sug-
gests that most of the cells are expressing musculoskeletal
differentiation markers. This observation, in addition to the
Figure 2. Biochemical analyses for chondrogenic differentiation of LVECs. (A): A pellet (2 × 105 cells/pellet) was papain-digested on day 21 after pulse-chase of [35S]-methionine into pellets, and new protein synthesis was determined. Data are expressed as mean � SD. *p < .05, compared with a control pellet, as determined by Student’s t-test, and triplicate analyses were performed on pellets (n = 3). (B): Duplicates of 2 million undifferentiated LVECs in monolayer culture; 20 pellets were papain-digested on day 0 and on day 21, respectively, and total collagen content was determined by measuring the hydroxyproline content of the papain-digests. (C): Determi-nation of proteoglycan in pellet-papain digests by the DMMB spectrophotometric assay. (D): Ten pellets were crushed homogenously on ice 2-amino-2-methyl-propanol solution (pH 10.3), and the supernatant was used for the detection of alkaline phosphatase activity. Each measurement reported in (B), (C), and (D) was averaged from at least two independent experiments, and data are expressed as mean � SD. *p < .05 compared with control pellets. Time course for proteoglycan production by LVECs in (E) digested control and TGFβ3-treated pellets or in (F) medium collected from the pellet cultures, determined by the DMMB spectrophotometric assay. Abbreviations: B, BMP2-treated pellets; T, TGFβ3-treated pellets; BMP2, bone morphogenetic protein; BT, BMP2 and TGFβ3 co-treated pellets; C, control pellets without growth factors; DMMB, dimethylmethylene blue; LVEC, large-vessel endothelial cell; SD, standard deviation; TGFβ3, transforming growth factor beta-3; UD, undifferentiated cells.
Kim, Hwang, Lee et al. 119
homogenous cell morphology, leads us to hypothesize that the
majority of the cells are differentiating in a similar manner.
Matrix Synthesis during the Pellet Culture of LVECsTo further examine the potential musculoskeletal differen-
tiation of LVEC pellets, protein analysis and matrix produc-
tion were evaluated. The general metabolic state of LVECs in
pellets was assessed by the measurement of total protein syn-
thesis after pellets were treated with or without BMP2 and/or
TGFβ3. Pellets were pulse-chased with [35S]-methionine for 3
hours on day 21 of culture and digested with papain, and then
the counts per minute (cpm) of each pellet was determined.
As presented in Figure 2A, cpm of the pellets treated with
TGFβ3 alone or TGFβ3 in combination with BMP2 showed
a significant increase in new protein synthesis (15,232.2 �
4,250.7 and 15,204.0 � 3,836.9, respectively) compared
with the control (6,346.6 � 1,965) or BMP2-treated pellet
(9,405.8 � 1,263.6). A time-dependent increase of the [35S]-
methionine incorporation was seen in TGFβ3-treated pellets
supplementary online Table 1; not shown).
Collagen and proteoglycan (PG) are the predominant
extracellular matrix (ECM) components in cartilage and
other musculoskeletal connective tissues and were measured
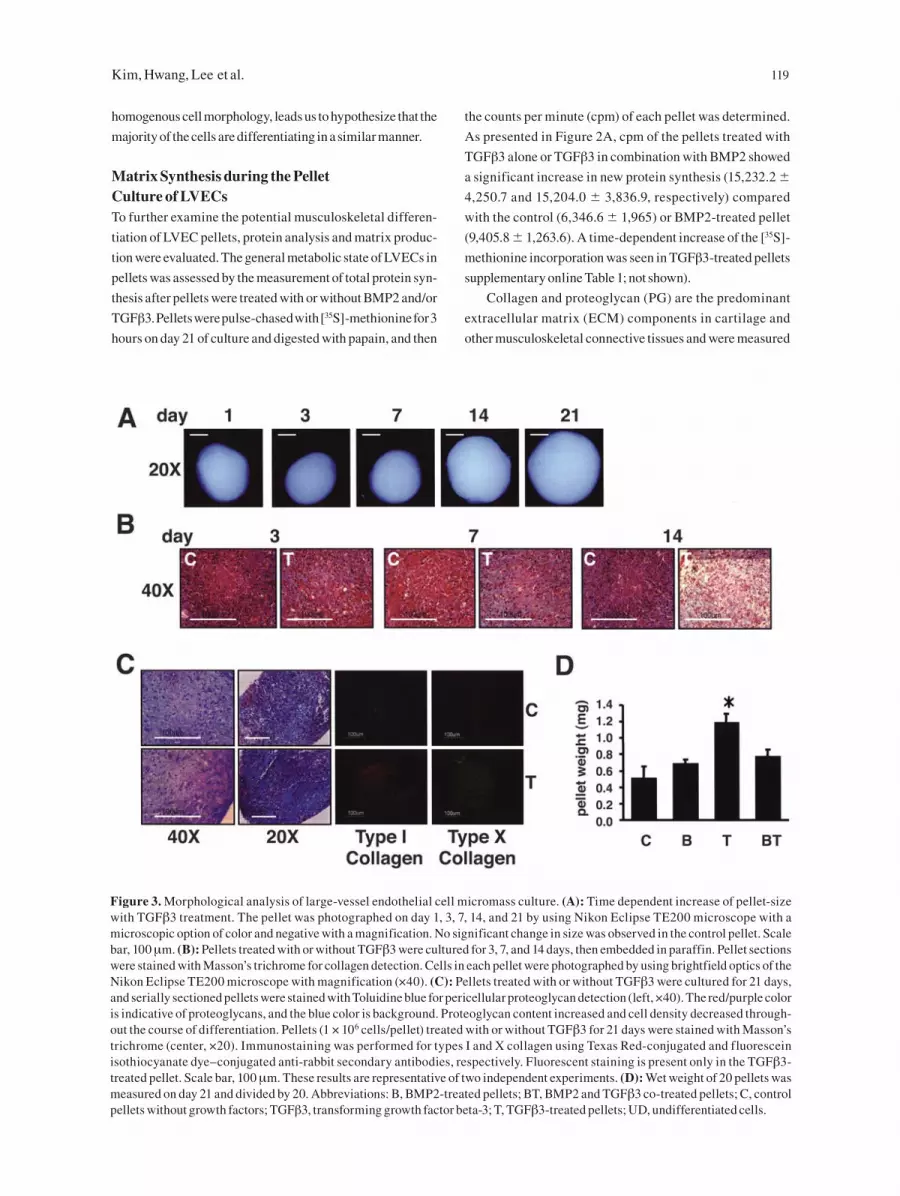
Figure 3. Morphological analysis of large-vessel endothelial cell micromass culture. (A): Time dependent increase of pellet-size with TGFβ3 treatment. The pellet was photographed on day 1, 3, 7, 14, and 21 by using Nikon Eclipse TE200 microscope with a microscopic option of color and negative with a magnification. No significant change in size was observed in the control pellet. Scale bar, 100 μm. (B): Pellets treated with or without TGFβ3 were cultured for 3, 7, and 14 days, then embedded in paraffin. Pellet sections were stained with Masson’s trichrome for collagen detection. Cells in each pellet were photographed by using brightfield optics of the Nikon Eclipse TE200 microscope with magnification (×40). (C): Pellets treated with or without TGFβ3 were cultured for 21 days, and serially sectioned pellets were stained with Toluidine blue for pericellular proteoglycan detection (left, ×40). The red/purple color is indicative of proteoglycans, and the blue color is background. Proteoglycan content increased and cell density decreased through-out the course of differentiation. Pellets (1 × 106 cells/pellet) treated with or without TGFβ3 for 21 days were stained with Masson’s trichrome (center, ×20). Immunostaining was performed for types I and X collagen using Texas Red-conjugated and fluorescein isothiocyanate dye–conjugated anti-rabbit secondary antibodies, respectively. Fluorescent staining is present only in the TGFβ3-treated pellet. Scale bar, 100 μm. These results are representative of two independent experiments. (D): Wet weight of 20 pellets was measured on day 21 and divided by 20. Abbreviations: B, BMP2-treated pellets; BT, BMP2 and TGFβ3 co-treated pellets; C, control pellets without growth factors; TGFβ3, transforming growth factor beta-3; T, TGFβ3-treated pellets; UD, undifferentiated cells.

120 Musculoskeletal Cell Differentiation
in pellets treated with or without growth factor. Collagen
contents were significantly increased in TGFβ3-treated pel-
lets (0.451 � 0.171) or TGFβ3 + BMP2–treated pellets (0.364
� 0.047) compared with undifferentiated cells (0.067 �
0.005), whereas only slight increase was observed in BMP2-
treated pellets (0.159 � 0.022) (Fig. 2B). PG content in pel-
lets treated with growth factors was much higher than in cells
from monolayer culture (Fig. 2C). Consistent with the results
of total collagen synthesis, the most dramatic increase of PG
content was shown in TGFβ3-treated pellets with approxi-
mately a fivefold increase over that of undifferentiated cells
(7.367 � 0.905). A time-dependent increase of the PG release
into the culture medium and in the pellets was observed (Fig.
2E, 2F). Control pellets that were not treated with any growth
factor also showed a slight increase of PG. Both total colla-
gen and PG content were significantly higher in control pel-
lets (0.181 � 0.020 of total collagen and 3.973 � 0.337 of PG)
than in undifferentiated cells (1.456 � 0.201), suggesting that
the pellet culture system itself could provide important envi-
ronmental cues for cell differentiation, which could be fur-
ther enhanced with the addition of BMP2 or TGFβ3.
Pellet morphology, as monitored by phase contrast
microscopy, changed over the culture period and depended
on growth factor exposure. Immediately after LVECs were
centrifuged to form pellets, the cells were aggregated and
appeared flattened at the bottom of the tube. After 24 hours,
all pellets thickened and developed into round, dense aggre-
gates. Between days 2 and 7, the pellets became spherical
without any increase in size (Fig. 3A, day 3 and 7). During
further cultivation, they changed from white and opaque
aggregates to a compact structure matrix border, suggesting
production of a dense ECM [16]. The size of pellets incubated
with TGFβ3 for 21 days showed a considerable increase
in size. Consistent with pellet size, the wet weight per pel-
let was greatest for TGFβ3 pellets (about threefold) over 3
weeks-culture (Fig. 3D). LVECs in monolayer, treated with
BMP2, showed no significant difference in proliferation
rate compared with the control without treatment. The DNA
quantity of pellets on day 21 was about 40%–60% of the ini-
tial DNA contents on day 0 (supplementary online Table 1;
not shown), similar to results from MSC pellet culture [16].
The largest decrease in DNA quantity (40%) occurred with
TGFβ3 treatment compared with control pellets, whereas
no significance was observed in BMP2 treatment alone or
BMP2 in combination with TGFβ3. Therefore, the increase
in size and weight might be at least in part due to ECM pro-
tein synthesis such as collagen and PG in pellets. Further-
more, because initial cell loss is frequently observed in pellet
cultures, the decrease in LVEC number may also suggest the
loss of precursor cells from non-musculoskeletal lineages.
TGFβ-Dependent Chondrogenic ActivityTGFβ3 demonstrated the most significant influence on
LVEC differentiation toward musculoskeletal lineages
in pellet culture. For this reason, the time-dependence of
TGFβ3 effects on LVEC pellets was more closely inves-
tigated. After 14 days in culture, the size of pellets with
TGFβ3 treatment increased significantly (Fig. 3A) com-
pared with the control pellets, which became smaller over
time in the absence of growth factor treatment. Histological
examination of the ECM was assessed in paraffin-embed-
ded, serially sectioned pellets. Masson’s trichrome staining
was selected to stain for collagen (blue or green) because it
is useful to distinguish collagen from muscle (red) or other
elements (black). As shown in Fig. 3B, a gradual increase
in blue color was observed over time in TGFβ3-treated pel-
lets after 14 days of culture. A significant increase in matrix
staining was observed on day 21, indicating a time-depen-
dent accumulation of collagen matrix in TGFβ3-treated
pellets. The metachromatic staining by Toluidine blue was
used for morphological assessment of ECM proteoglycan
synthesis. The light- to dark-red/purple staining, positive for
proteoglycans, was noticeable within the cell pellets treated
with TGFβ3 for 21 days (Fig. 3C). However, homogeneous
staining over the whole area of pellet was rarely seen. In the
absence of added growth factors, pellets had little evidence
of matrix accumulation. Evaluation of the nuclear staining
with H&E in pellet sections after 21 days revealed that sepa-
ration between cells increased with time in TGFβ3 pellets as
more ECM was produced (Fig. 3B, 3C).
DiscussionEBD cells are a potentially useful stem cell population
for cell and tissue engineering therapies because of their
multipotent differentiation capabilities, ease in expansion
such that a feeder cell layer is not required, and lack of tera-
toma formation when injected in vivo. Multipotent MSCs
in the bone marrow have the capability to differentiate into
mesodermal lineages, such as bone, fat, muscle, cartilage,
ligament, tendon, and stroma [17], and serve as a potential
model system to understand and compare the musculosk-
eletal differentiation of embryonic cells. Formation of the
vertebrate skeleton begins with the migration of undif-
ferentiated MSCs from bone marrow to sites destined to
become bone. The cells undergo a condensation step and
then form a cartilaginous scaffold, at the center of which
cells synthesize ECMs that are abundant in type II collagen
and PGs [15]. As differentiation progresses, hypertrophic
chondrocytes produce a mineralized matrix after under-
going apoptosis, and the matrices are gradually replaced
by an invasion of blood vessels, followed by synthesis of

Kim, Hwang, Lee et al. 121
bone matrix of osteoblasts. In vitro systems used to study
chondrogenesis use cells from primary cultures of adult
or embryonic cartilage tissue, cell lines of mesenchymal
origin, or MSCs derived from bone marrow. Although we
continue to learn more about the differentiation capabili-
ties of adult stem cells, they represent already committed,
differentiated cells.
In the study reported here, we used a micromass pellet
culture system to investigate the differentiation capacity of
EBD cells. This system provides a unique three-dimensional
culture condition for undifferentiated cells that resembles
more closely in vitro surroundings for EB formation and in
vivo development [8]. First, to elucidate whether the expres-
sion of markers involved in distinct developmental lineages
of human EBD cells could be differentially regulated by
micromass culture of the cells, RT-PCR analysis was per-
formed in RNA extracts from undifferentiated cells cul-
tured both as a monolayer and as pellets. Decrease of neu-
ronal (NFH) and hematopoietic (flk-1) marker expressions,
in addition to downregulation of ALP by pellet culture of
undifferentiated cells, suggests that the human EBD cells
have lost their neural and vascular progenitor cell character-
istics in pellet culture. We hypothesize that both cell selec-
tion and differentiation processes are occurring. The culture
conditions may be selecting for the growth of a subpopula-
tion of EBD stem cells with capability for mesenchymal
differentiation, as evidenced by the decreasing cell number
and increase in characteristic mesenchymal markers such
as AGN that are present in the undifferentiated cells. While
neural and vascular expression significantly decreased,
there is still low expression level, suggesting that cells with
the capability to differentiate to different lineages still exist.
Differentiation processes are also occurring, as demon-
strated by the “activation” of mesenchymal tissue-related
markers not originally present in the EBD cells, including
COMP, Col IX, and Col X.
During limb development, cartilage serves as anla-
gen for bone, in addition to forming fibrocartilage, hyaline
cartilage, tendon, and ligaments. There are ECM markers
characteristic for each of these tissues. The ECM of cartilage
contains large amounts of AGN, Col II, and other key matrix
components, including COMP. In this study, the marked
induction of Col II, Col IX, AGN, COMP, and ChM in the
presence of TGFβ3 was revealed in LVEC pellets (Fig. 1),
supporting the conclusion that the cells were differentiating
into musculoskeletal lineages. Prior to induction of chon-
drogenesis in undifferentiated cells at day 0, transcripts for
Col II, Col IIA, Col IX, Col X, COMP, and ChM were hardly
detected, whereas those for Col I, Col III, LP, AGN, CS-4S,
and CS-6S were readily or weakly detectable.
The genes that showed strong expression in our
results—Col 1α(I), Col I, and CS (Fig. 1)—are involved
in osteochondrogenesis of MSCs. ChM is not only a major
regulator in cartilage development but also a bone-remod-
eling factor [18]. Our results of expression of hypertrophic
chondrocyte markers, as assessed by the increase of ALP
activity and upregulation of Col I, Col II, and X expression
upon exposure to BMP2 or TGFβ3 (Fig. 1), also indicate that
LVECs were capable of differentiating into osteogenic lin-
eages, even though there was no detectable mineral deposi-
tion assessed by von Kossa’s stain in the pellet section (data
not shown). Protein expression of types I and X collagen was
confirmed by immunostaining (Fig. 3C). Col III and myf5
expressions further suggest the differentiation of LVECs
into ligament/tendon and myogenic lineages, respectively.
Although this speculation is based on the results of PCR
assay without definitive proof of human EBD cell differen-
tiation into specific lineages, all these results support that
LVECs in micromass have a potential to differentiate from
EG cells into musculoskeletal lineages.
The maintenance of nestin expression is compatible with
MSCs. Nestin is found in neural precursors, in the develop-
ing mouse limb buds in vivo, and in differentiating muscle
and myocardial cells [19, 20]. In addition, nestin is expressed
in immature MSC precursors [21], indicating that its expres-
sion is not restricted to developing neural cells but can be
a marker found in MSCs. In the current study, nestin was
strongly expressed in undifferentiated LVECs as reported
[4], but little change was detected in pelleted LVECs with
or without treatments of BMP2 or TGFβ3 (Fig. 1A). These
results suggest that undifferentiated LVECs might have cells
producing a complex pattern of neural and mesenchymal
cell differentiation, or the LVECs might maintain a capacity
similar to that of MSCs to differentiate into mesenchymal
cell lineages after responding to mesenchymal environ-
ments, including osteochondrogenic stimuli. This notion is
supported by the report that stem cells, mesenchymal cells,
and neural progenitors were present together during mouse
ES cell differentiation after exposure of the cells to chordin,
a BMP inhibitor [22].
PGs are a major component of cartilage ECM. The eval-
uation of PG release into the culture medium is as important
as evaluation of PG content in the cartilage matrix. In our
studies, the enhancement of TGFβ3 in osteochondrogenic
differentiation of LVEC pellets was confirmed by measur-
ing both PG content of the pellets and PG release into the cul-
ture medium. PG accumulation in pellets and in the medium
was increased by TGFβ3 during all incubation periods, and
the release was maintained higher in TGFβ3 pellets than in
the control, suggesting differentiation (Fig. 2E, 2F). How-
ever, control pellets or pellets treated with BMP2 alone or
co-treated with BMP2 and TGFβ3 released approximately

122 Musculoskeletal Cell Differentiation
the same amount of PG into the medium on day 21. Inter-
estingly, a detectable level of PG was observed in undiffer-
entiated LVECs (Figs. 2, 5). That could be explained by the
observation of PGs (AGN, LP, CS-4S, and CS-6S) in mono-
layer cultured LVECs by RT-PCR analysis (Fig. 1). In addi-
tion, it is possible that a pericellular matrix, TRA-1-60, was
responsible for DMMB-reactive PG detectable in undiffer-
entiated cells because it is one of the important markers of
human pluripotent EG cells [23].
TGFβs are potent antiproliferative agents of many dif-
ferent cell types [24]. Activities of TGFβs include induction
of cell-cycle arrest, induction of apoptosis, and induction of
expression of cell adhesion molecules and components of the
ECM, including collagen, fibronectin, and integrins. When
MSCs differentiate into chondrocytes under TGFβ3 and
BMP6 treatment, there is a continued loss of cells through
apoptosis [16]. More interestingly, MSCs exposed to TGFβ1
show a significant decline in cell number in three-dimen-
sional culture disks (REF-worster), indicating that TGFβs
can suppress cell growth in a three-dimensional system. The
TGFβRII expression remained constant in all experimental
pellet groups.
The expression of cartilage and other musculoskeletal-
specific genes, in addition to the results of biochemical anal-
ysis in this study, strongly suggests that LVEC pellets dif-
ferentiated toward chondrocytic and MSC lineages. Small
quantities of serum (1%) instead of the strictly serum-free
medium were used in this study, based on reports that meso-
dermal differentiation of ES cells can be inhibited under
serum-free conditions [25]. We could not identify morpho-
logically hypertrophic or mature chondrocytes in pellets
treated with TGFβ3 (Fig. 3C). There was a positive staining
with Toluidine blue in regions of the pellets that were incu-
bated in TGFβ3, but not throughout the pellet. Thus, it seems
that LVECs could not differentiate further down a specific
MSC lineage such as cartilage or bone. One explanation may
be that prolonged treatment of TGFβ3 in this study might
have prevented or retarded phenotype development of chon-
drocyte maturation at a later stage of cartilage formation, as
reported for cartilage nodules generated in the absence of
TGFβ3 at a later phase of chondrogenic culture of ES cells
from an E14 embryo [26]. However, the detection of Col X
transcript by PCR (Fig. 1) and immunostaining (Fig. 3C)
suggests that hypertrophic differentiation might occur in the
presence of TGFβ3. We also hypothesize that the cells may
need a more complex array of soluble and matrix-related sig-
nals to further differentiate. More complex signals, includ-
ing the enhanced cell–cell contacts provided in the pellet
culture system, may be incorporated into three-dimensional
tissue engineering biomaterial scaffold systems with a cock-
tail of growth factors to promote specific differentiation.
In summary, human LVECs showed features of chon-
drogenic and mesenchymal differentiation when they were
grown in micromass culture, as evidenced by tissue-spe-
cific gene expression and protein synthesis. Metachromatic
staining with Toluidine blue of pellet sections confirmed
that differentiated LVECs accumulated ECM. A significant
increase in PG and collagen protein synthesis was observed
in the presence of TGFβ3, indicating that the osteochondro-
inductive agent TGFβ3 further enhanced LVEC-derived
chondrogenic differentiation. This study also shows the
flexibility of partially differentiated stem cells to shift their
differentiation profile to other developmental lineages, fur-
thering our understanding of stem cell capabilities and their
potential use in musculoskeletal regeneration therapies.
AcknowledgmentsThis study was supported by the Johns Hopkins University
(JHU)–Technion, Whitaker Foundation and Institute of Cell
Engineering. The authors are grateful to Dr. Jin Hyen Baek
(JHU), and Dr. Christopher G. Williams (JHU) for their crit-
ical review and technical assistance.
Disclosure
Under a licensing agreement between Geron Corporation
and JHU, Drs. Gearhart and Shamblott are entitled to a share
of royalty received by the university on sales of products
described in this presentation. Drs. Gearhart and Shamblott
and the University own Geron stock, which is subject to cer-
tain restrictions under university policy. The terms of this
arrangement are being managed by JHU in accordance with
its conflict of interest policies.

Kim, Hwang, Lee et al. 123
References
1 Thomson JA et al. Embryonic stem cell lines derived from human blastocysts. Science 1998;282:1145–1147.
2 Shamblott MJ et al. Derivation of pluripotent stem cells from cultured human primordial germ cells. Proc Natl Acad Sci U S A 1998;95:13726–13731.
3 Dang SM et al. Efficiency of embryoid body formation and hematopoietic development from embryonic stem cells in different culture systems. Biotechnol Bioeng 2002;78:442–453.
4 Shamblott MJ et al. Human embryonic germ cell deriva-tives express a broad range of developmentally distinct markers and proliferate extensively in vitro. Proc Natl Acad Sci U S A 2001;98:113–118.
5 Kramer J et al. In vitro differentiation of mouse ES cells: bone and cartilage. Methods Enzymol 2003;365:251–268.
6 Kramer J et al. Embryonic stem cell-derived chondro-genic differentiation in vitro: activation by BMP-2 and BMP-4. Mech Dev 2000;9:193–205.
7 Levenberg S et al. Differentiation of human embryonic stem cells on three-dimensional polymer scaffolds. Proc Natl Acad Sci U S A 2003;100:12741–12746.
8 Stewart MC et al. Phenotypic stability of articular chon-drocytes in vitro: the effects of culture models, bone mor-phogenetic protein 2, and serum supplementation. J Bone Miner Res 2000;15:166–174.
9 Hegert C et al. Differentiation plasticity of chondrocytes derived from mouse embryonic stem cells. J Cell Sci 2002;115:4617–4628.
10 Johnstone B et al. In vitro chondrogenesis of bone mar-row-derived mesenchymal progenitor cells. Exp Cell Res 1998;238:265–272.
11 Wang EA et al. Purification and characterization of other distinct bone-inducing factors. Proc Natl Acad Sci U S A 1988;85:9484–9488.
12 Elisseeff J et al. Transdermal photopolymerization for minimally invasive implantation. Proc Natl Acad Sci U S A 1999;96:3104–3107.
13 Baylies MK, Bate M. Twist: a myogenic switch in Dro-sophila. Science 1996;272:1481–1484.
14 Kronenberg HM. Twist genes regulate Runx2 and bone formation. Dev Cell 2004;6:317–318.
15 Denker AE et al. Chondrogenic differentiation of murine C3H10T1/2 multipotential mesenchymal cells. I: Stimu-lation by bone morphogenetic protein-2 in high-density micromass cultures. Differentiation 1999;64:67–76.
16 Sekiya I et al. In vitro cartilage formation by human adult stem cells from bone marrow stroma defines the sequence of cellular and molecular events during chondrogenesis. Proc Natl Acad Sci U S A 2002;99:4397–4402.
17 Pittenger MF et al. Multilineage potential of adult human mesenchymal stem cells. Science 1999;284:143–147.
18 Nakamichi Y et al. Chondromodulin I is a bone remodel-ing factor. Mol Cell Biol 2003;23:636–644.
19 McKay R. Stem cells in the central nervous system. Sci-ence 1997;276:66–71.
20 Lendahl U et al. CNS stem cells express a new class of intermediate filament protein. Cell 1990;60:585–595.
21 Vogel W et al. Heterogeneity among human bone mar-row-derived mesenchymal stem cells and neural progeni-tor cells. Haematologica 2003;88:126–133.
22 Gratsch TE, O’Shea KS. Noggin and chordin have distinct activities in promoting lineage commitment of mouse embryonic stem (ES) cells. Dev Biol 2002;245:83–94.
23 Pera MF et al. Human embryonic stem cells. J Cell Sci 2000;113:5–10.
24 Filmus J et al. Role of transforming growth factor alpha (TGF-alpha) in the transformation of ras-transfected rat intestinal epithelial cells. Oncogene 1993;8:1017–1022.
25 Wiles MV, Johansson BM. Embryonic stem cell devel-opment in a chemically defined medium. Exp Cell Res 1999;247:241–248.
26 Nakayama N et al. Macroscopic cartilage formation with embryonic stem-cell-derived mesodermal progenitor cells. J Cell Sci 2003;116:2015–2028.




















