
SNP Detection in mRNA in Living Cells Using Allele Specific FRET Probes
Upload
khangminh22Category
view
0download
0
Overview of the Cellular Basis of Life (pp. 62–63)
The Plasma Membrane: Structure (pp. 63–67)
The Fluid Mosaic Model (pp. 63–66)
Membrane Junctions (pp. 66–67)
The Plasma Membrane: MembraneTransport (pp. 68–77)
Passive Processes (pp. 68–72)
Active Processes (pp. 72–77)
The Plasma Membrane: Generation of aResting Membrane Potential (pp. 79–80)
The Plasma Membrane: Cell-Environment Interactions (pp. 80–81)
Roles of Cell Adhesion Molecules (pp. 80–81)
Roles of Membrane Receptors (p. 81)
Role of Voltage-Sensitive Membrane ChannelProteins (p. 81)
The Cytoplasm (pp. 81–91)
Cytoplasmic Organelles (pp. 83–89)
Cellular Extensions (pp. 90–91)
The Nucleus (pp. 91–95)
The Nuclear Envelope (pp. 91–93)
Nucleoli (p. 93)
Chromatin (pp. 93–95)
Cell Growth and Reproduction (pp. 95–107)
The Cell Life Cycle (pp. 95–100)
Protein Synthesis (pp. 100–105)
Other Roles of DNA (pp. 105–106)
Cytosolic Protein Degradation (pp. 106–107)
Extracellular Materials (p. 107)
Developmental Aspects of Cells (pp. 108–109)
Just as bricks and timbers are the structural units of a house, cells arethe structural units of all living things, from one-celled “generalists”like amoebas to complex multicellular organisms such as humans,
dogs, and trees. The human body has 50 to 100 trillion of these tinybuilding blocks.
This chapter focuses on structures and functions shared by all cells.We address specialized cells and their unique functions in later chapters.
Cells: The Living
Units
61
3000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 61

Overview of the Cellular Basis of Life� Define cell.
� List the three major regions of a generalized cell and indi-cate the function of each.
The English scientist Robert Hooke first observed plant cellswith a crude microscope in the late 1600s. However, it was notuntil the 1830s that two German scientists, Matthias Schleidenand Theodor Schwann, were bold enough to insist that all livingthings are composed of cells. The German pathologist RudolfVirchow extended this idea by contending that cells arise onlyfrom other cells. Virchow’s proclamation was revolutionary be-cause it openly challenged the widely accepted theory of sponta-neous generation, which held that organisms arise spontaneouslyfrom garbage or other nonliving matter.
Since the late 1800s, cell research has been exceptionallyfruitful and provided us with four concepts collectively knownas the cell theory:
1. A cell is the basic structural and functional unit of livingorganisms. So when you define cell properties you are infact defining the properties of life.
2. The activity of an organism depends on both the individ-ual and the collective activities of its cells.
3. According to the principle of complementarity of structureand function, the biochemical activities of cells are dic-tated by the relative number of their specific subcellularstructures.
4. Continuity of life from one generation to another has acellular basis.
We will expand on all of these concepts as we progress. Let usbegin with the idea that the cell is the smallest living unit. What-ever its form, however it behaves, the cell is the microscopicpackage that contains all the parts necessary to survive in anever-changing world. It follows then that loss of cellular homeo-stasis underlies virtually every disease.
The trillions of cells in the human body include over 200 dif-ferent cell types that vary greatly in shape, size, and function(Figure 3.1). The spherical fat cells, disc-shaped red blood cells,branching nerve cells, and cubelike cells of kidney tubules are justa few examples of the shapes cells take. Depending on type, cellsalso vary greatly in length—ranging from 2 micrometers(1/12,000 of an inch) in the smallest cells to over a meter in thenerve cells that cause you to wiggle your toes. A cell’s shape re-flects its function. For example, the flat, tilelike epithelial cells thatline the inside of your cheek fit closely together, forming a livingbarrier that protects underlying tissues from bacterial invasion.
Regardless of type, all cells are composed chiefly of carbon,hydrogen, nitrogen, oxygen, and trace amounts of several otherelements. In addition, all cells have the same basic parts andsome common functions. For this reason, it is possible to speakof a generalized, or composite, cell (Figure 3.2).
Human cells have three main parts: the plasma membrane,the cytoplasm, and the nucleus. The plasma membrane, a fragile
62 UNIT 1 Organization of the Body
3
Fibroblasts
Erythrocytes
Epithelial cells
(a) Cells that connect body parts, form linings, or transport gases
(c) Cell that stores nutrients
(b) Cells that move organs and body parts
(d) Cell that fights disease
Macrophage
Nerve cell
Fat cell
Sperm
(e) Cell that gathers information and controls body functions
(f) Cell of reproduction
Skeletalmusclecell
Smoothmuscle cells
Figure 3.1 Cell diversity. (Note that cells are not drawn to thesame scale.)
barrier, is the outer boundary of the cell. Internal to this mem-brane is the cytoplasm (si�to-plazm), the intracellular fluid thatis packed with organelles, small structures that perform specificcell functions. The nucleus (nu�kle-us) controls cellular activi-ties and typically it lies near the cell’s center. We use these threemain parts of the cell to organize the summary in Table 3.3(pp. 94–95), and we describe them in greater detail next.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 62

C H E C K Y O U R U N D E R S TA N D I N G
1. Name the three basic parts of a cell and describe the func-tion of each.
2. How would you explain the meaning of a “generalized cell”to a classmate?
For answers, see Appendix G.
The Plasma Membrane: Structure� Describe the chemical composition of the plasma mem-
brane and relate it to membrane functions.
� Compare the structure and function of tight junctions,desmosomes, and gap junctions.
The flexible plasma membrane defines the extent of a cell,thereby separating two of the body’s major fluid compartments—the intracellular fluid within cells and the extracellular fluid out-side cells. The term cell membrane is commonly used as a syn-onym for plasma membrane, but because nearly all cellularorganelles are enclosed in a membrane, in this book we will al-ways refer to the cell’s surface, or outer limiting membrane, asthe plasma membrane. The plasma membrane is much morethan a passive envelope. As you will see, its unique structureallows it to play a dynamic role in many cellular activities.
The Fluid Mosaic ModelThe fluid mosaic model of membrane structure depicts theplasma membrane as an exceedingly thin (7–10 nm) structure
Chapter 3 Cells: The Living Units 63
3
Secretion being releasedfrom cell by exocytosis
Peroxisome
Ribosomes
Roughendoplasmicreticulum
Nucleus
Nuclear envelopeChromatin
Golgi apparatus
Nucleolus
Smooth endoplasmicreticulum
Cytosol
Lysosome
Mitochondrion
Centrioles
Centrosomematrix
• Microtubule
Cytoskeletalelements
• Intermediate filaments
Plasmamembrane
Figure 3.2 Structure of the generalized cell. No cell is exactly like this one, but this compositeillustrates features common to many human cells. Note that not all of the organelles are drawnto the same scale in this illustration.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 63

composed of a double layer, or bilayer, of lipid molecules withprotein molecules “plugged into” or dispersed in it (Figure 3.3).The proteins, many of which float in the fluid lipid bilayer, forma constantly changing mosaic pattern. The model is named forthis characteristic.
Membrane Lipids
The lipid bilayer forms the basic “fabric” of the membrane. It isconstructed largely of phospholipids, with smaller amounts ofcholesterol and glycolipids. Each lollipop-shaped phospholipidmolecule has a polar “head” that is charged and is hydrophilic(hydro � water, philic � loving), and an uncharged, nonpolar“tail” that is made of two fatty acid chains and is hydrophobic(phobia � fear). The polar heads are attracted to water—themain constituent of both the intracellular and extracellularfluids—and so they lie on both the inner and outer surfaces ofthe membrane. The nonpolar tails, being hydrophobic, avoidwater and line up in the center of the membrane.
The result is that all biological membranes share a com-mon sandwich-like structure: They are composed of two par-allel sheets of phospholipid molecules lying tail to tail, withtheir polar heads exposed to water both inside and outsidethe cell. This self-orienting property of phospholipids en-courages biological membranes to self-assemble into closed,generally spherical, structures and to reseal themselvesquickly when torn.
The plasma membrane is a dynamic fluid structure that is inconstant flux. Its consistency is like that of olive oil. The lipidmolecules of the bilayer move freely from side to side, parallelto the membrane surface, but their polar-nonpolar interac-tions prevent them from flip-flopping or moving from onephospholipid layer (half of the bilayer) to the other. Theinward-facing and outward-facing surfaces of the plasmamembrane differ in the kinds and amounts of lipids they con-tain, and these variations are important in determining localmembrane structure and function. The majority of membranephospholipids are unsaturated, a condition which kinks theirtails (increasing the space between them) and increases mem-brane fluidity. (See the illustration of phosphatidylcholine inFigure 2.16b, p. 46.)
Glycolipids (gli�ko-lip�idz) are lipids with attached sugargroups. They are found only on the outer plasma membranesurface and account for about 5% of the total membrane lipid.Their sugar groups, like the phosphate-containing groups ofphospholipids, make that end of the glycolipid molecule polar,whereas the fatty acid tails are nonpolar.
Some 20% of membrane lipid is cholesterol. Like phospho-lipids, cholesterol has a polar region (its hydroxyl group) and anonpolar region (its fused ring system). It wedges its platelikehydrocarbon rings between the phospholipid tails, stabilizingthe membrane, while increasing the mobility of the phospho-lipids and the fluidity of the membrane.
About 20% of the outer membrane surface contains lipidrafts, dynamic assemblies of saturated phospholipids (whichpack together tightly) associated with unique lipids calledsphingolipids and lots of cholesterol. These quiltlike patches are
more stable and orderly and less fluid than the rest of the mem-brane, and they can include or exclude specific proteins to variousextents. Because of these qualities, lipid rafts are assumed to beconcentrating platforms for certain receptor molecules or formolecules needed for cell signaling. (Cell signaling will be dis-cussed on pp. 81–82.)
Membrane Proteins
Proteins make up about half of the plasma membrane by massand are responsible for most of the specialized membrane func-tions. There are two distinct populations of membrane pro-teins, integral and peripheral (Figure 3.3). Integral proteins arefirmly inserted into the lipid bilayer. Some protrude from onemembrane face only, but most are transmembrane proteins thatspan the entire width of the membrane and protrude on bothsides. Whether transmembrane or not, all integral proteinshave both hydrophobic and hydrophilic regions. This struc-tural feature allows them to interact both with the nonpolarlipid tails buried in the membrane and with water inside andoutside the cell.
Although some are enzymes, most transmembrane pro-teins are involved in transport. Some cluster together to formchannels, or pores, through which small, water-soluble mole-cules or ions can move, thus bypassing the lipid part of themembrane. Others act as carriers that bind to a substance andthen move it through the membrane. Still others are receptorsfor hormones or other chemical messengers and relay mes-sages to the cell interior (a process called signal transduction)(Figure 3.4a, b).
Peripheral proteins (Figure 3.3), in contrast, are not em-bedded in the lipid. Instead, they attach rather loosely only tointegral proteins and are easily removed without disrupting themembrane. Peripheral proteins include a network of filamentsthat helps support the membrane from its cytoplasmic side(Figure 3.4c). Some peripheral proteins are enzymes. Others aremotor proteins involved in mechanical functions, such aschanging cell shape during cell division and muscle cell con-traction. Still others link cells together.
Some of the proteins float freely. Others, particularly theperipheral proteins, are restricted in their movements be-cause they are “tethered” to intracellular structures that makeup the cytoskeleton. Many of the proteins that abut the extra-cellular fluid are glycoproteins with branching sugar groups.The term glycocalyx (gli�ko-kal�iks; “sugar covering”) is usedto describe the fuzzy, sticky, carbohydrate-rich area at the cellsurface. You can think of your cells as sugar-coated. The gly-cocalyx that clings to each cell’s surface is enriched both byglycolipids and by glycoproteins secreted by the cell.
Because every cell type has a different pattern of sugars in itsglycocalyx, the glycocalyx provides highly specific biologicalmarkers by which approaching cells recognize each other(Figure 3.4f). For example, a sperm recognizes an ovum (eggcell) by the ovum’s unique glycocalyx. Cells of the immune sys-tem identify a bacterium by binding to certain membraneglycoproteins in the bacterial glycocalyx.
64 UNIT 1 Organization of the Body
3
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 64

Chapter 3 Cells: The Living Units 65
3
Integral proteins
Extracellular fluid(watery environment)
Cytoplasm (watery environment)
Polar head of phospholipid molecule
GlycolipidCholesterol
Peripheral proteins
Bimolecular lipid layercontaining proteins
Inward-facing layer of phospholipids
Outward-facing layer of phospholipids
Carbohydrate of glycocalyx
Glycoprotein
Nonpolar tail of phospholipid molecule
Filament of cytoskeleton
Figure 3.3 Structure of the plasma membrane according to the fluid mosaic model. Thelipid bilayer forms the basic structure of the membrane. The associated proteins are involved inmembrane functions such as membrane transport, catalysis, and cell-to-cell recognition.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 65

H O M E O S TAT I C I M B A L A N C E
Definite changes in the glycocalyx occur in a cell that is becom-ing cancerous. In fact, a cancer cell’s glycocalyx may change al-most continuously, allowing it to keep ahead of immune systemrecognition mechanisms and avoid destruction. (Cancer is dis-cussed on pp. 142–143.) ■
C H E C K Y O U R U N D E R S TA N D I N G
3. What basic structure do all cellular membranes share?4. Why do phospholipids, which form the greater part of cell
membranes, organize into a bilayer—tail-to-tail—in a wateryenvironment?
5. What is the importance of the glycocalyx in cell interactions?
For answers, see Appendix G.
Membrane JunctionsAlthough certain cell types—blood cells, sperm cells, and somephagocytic cells (which ingest and destroy bacteria and othersubstances)—are “footloose” in the body, many other types,particularly epithelial cells, are knit into tight communities.Typically, three factors act to bind cells together:
1. Glycoproteins in the glycocalyx act as an adhesive.2. Wavy contours of the membranes of adjacent cells fit to-
gether in a tongue-and-groove fashion.3. Special membrane junctions are formed (Figure 3.5).
Because junctions are the most important factor securing cellstogether, let us look more closely at the various types.
Tight Junctions In a tight junction, a series of integral proteinmolecules (including occludins and claudins) in the plasmamembranes of adjacent cells fuse together, forming animpermeable junction that encircles the cell (Figure 3.5a). Tightjunctions help prevent molecules from passing through the ex-tracellular space between adjacent cells. For example, tight junc-tions between epithelial cells lining the digestive tract keepdigestive enzymes and microorganisms in the intestine fromseeping into the bloodstream. (Although called “impermeable”junctions, some tight junctions are somewhat leaky and may al-low certain types of ions to pass.)
Desmosomes Desmosomes (des�mo-somz; “binding bod-ies”) are anchoring junctions—mechanical couplings scatteredlike rivets along the sides of abutting cells that prevent their sep-aration (Figure 3.5b). On the cytoplasmic face of each plasmamembrane is a buttonlike thickening called a plaque. Adjacentcells are held together by thin linker protein filaments (cad-herins) that extend from the plaques and fit together like the teethof a zipper in the intercellular space. Thicker keratin filaments(intermediate filaments, which form part of the cytoskeleton)extend from the cytoplasmic side of the plaque across the widthof the cell to anchor to the plaque on the cell’s opposite side. Inthis way, desmosomes not only bind neighboring cells together,
66 UNIT 1 Organization of the Body
3
ATP
A protein (left) that spans the membrane may provide a hydrophilic channel across the membrane that is selective for a particular solute. Some transport proteins (right) hydrolyze ATP as an energy source to actively pump substances across the membrane.
A membrane protein exposed to the outside of the cell may have a binding site with a specific shape that fits the shape of a chemical messenger, such as a hormone. The external signal may cause a change in shape in the protein that initiates a chain ofchemical reactions in the cell.
Elements of the cytoskeleton (cell’s internal supports) and the extracellular matrix (fibers and other substances outside the cell) may be anchored to membrane proteins, which help maintain cell shape and fix the location of certain membrane proteins. Others play a role in cell movement or bind adjacent cells together.
A protein built into the membrane may be an enzyme with its active site exposed to substances in the adjacent solution. In some cases, several enzymes in a membrane act as a team that catalyzes sequential steps of ametabolic pathway as indicated (left to right) here.
Membrane proteins of adjacent cells may be hooked together in various kinds of intercellular junctions. Some membrane proteins (CAMs) of this group provide temporary binding sites that guide cell migration and other cell-to-cell interactions.
Some glycoproteins (proteins bonded to short chains of sugars) serve as identification tags that are specifically recognized by other cells.
CAMs
(f) Cell-cell recognition
(e) Intercellular joining
Glycoprotein
(d) Enzymatic activity
(c) Attachment to the cytoskeleton and extracellular matrix (ECM)
(b) Receptors for signal transduction
(a) Transport
Signal
Enzymes
Receptor
Figure 3.4 Membrane proteins performmany tasks. A single protein may performsome combination of these tasks.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 66

but they also contribute to a continuous internal network ofstrong “guy-wires.”
This arrangement distributes tension throughout a cellularsheet and reduces the chance of tearing when it is subjected topulling forces. Desmosomes are abundant in tissues subjectedto great mechanical stress, such as skin and heart muscle.
Gap Junctions A gap junction, or nexus (nek�sus; “bond”), isa communicating junction between adjacent cells. At gap junc-tions the adjacent plasma membranes are very close, and thecells are connected by hollow cylinders called connexons (ko-nek�sonz), composed of transmembrane proteins. The manydifferent types of connexon proteins vary the selectivity of the
gap junction channels. Ions, simple sugars, and other smallmolecules pass through these water-filled channels from onecell to the next (Figure 3.5c).
Gap junctions are present in electrically excitable tissues, suchas the heart and smooth muscle, where ion passage from cell tocell helps synchronize their electrical activity and contraction.
C H E C K Y O U R U N D E R S TA N D I N G
6. What two types of membrane junctions would you expect tofind between muscle cells of the heart?
For answer, see Appendix G.
Chapter 3 Cells: The Living Units 67
3
Interlockingjunctionalproteins
Intercellularspace
Intercellularspace
Plasma membranesof adjacent cells
Microvilli
Intercellularspace
Basement membrane
Plaque
Linkerglycoproteins(cadherins)Intermediate
filament(keratin)
Intercellularspace
Channelbetween cells(connexon)
(a) Tight junctions: Impermeable junctions prevent molecules from passing through the intercellular space.
(b) Desmosomes: Anchoring junctions bind adjacent cells together and help form an internal tension-reducing network of fibers.
(c) Gap junctions: Communicating junctions allow ions and small molecules to pass from one cell to the next for intercellular communication.
Figure 3.5 Cell junctions. An epithelial cell is shown joined to adjacent cells by three commontypes of cell junctions. (Note: Except for epithelia, it is unlikely that a single cell will have allthree junction types.)
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 67

The Plasma Membrane: Membrane Transport� Relate plasma membrane structure to active and passive
transport processes.
� Compare and contrast simple diffusion, facilitated diffu-sion, and osmosis relative to substances transported, direc-tion, and mechanism.
Our cells are bathed in an extracellular fluid called interstitialfluid (in�ter-stish�al) that is derived from the blood. Interstitialfluid is like a rich, nutritious “soup.” It contains thousands of in-gredients, including amino acids, sugars, fatty acids, vitamins,regulatory substances such as hormones and neurotransmitters,salts, and waste products. To remain healthy, each cell must ex-tract from this mix the exact amounts of the substances it needsat specific times.
Although there is continuous traffic across the plasma mem-brane, it is a selectively, or differentially, permeable barrier,meaning that it allows some substances to pass while excludingothers. It allows nutrients to enter the cell, but keeps many un-desirable substances out. At the same time, it keeps valuablecell proteins and other substances in the cell, but allows wastesto exit.
Substances move through the plasma membrane in essen-tially two ways—passively or actively. In passive processes, sub-stances cross the membrane without any energy input from thecell. In active processes, the cell provides the metabolic energy(ATP) needed to move substances across the membrane. Thevarious transport processes that occur in cells are summarizedin Table 3.1 on p. 72 and Table 3.2 on p. 80. Let’s examine eachof these types of membrane transport.
H O M E O S TAT I C I M B A L A N C E
Selective permeability is a characteristic of healthy, intact cells.When a cell (or its plasma membrane) is severely damaged, themembrane becomes permeable to virtually everything, andsubstances flow into and out of the cell freely. This phenome-non is evident when someone has been severely burned. Pre-cious fluids, proteins, and ions “weep” from the dead anddamaged cells. ■
Passive ProcessesThe two main types of passive transport are diffusion (di-fu�zhun) and filtration. Diffusion is an important means of pas-sive membrane transport for every cell of the body. Becausefiltration generally occurs only across capillary walls, that topicis more properly covered in conjunction with capillary trans-port processes later in the book.
Diffusion
Diffusion is the tendency of molecules or ions to move from anarea where they are in higher concentration to an area wherethey are in lower concentration, that is, down or along theirconcentration gradient. The constant random and high-speedmotion of molecules and ions (a result of their intrinsic kineticenergy) results in collisions. With each collision, the particlesricochet off one another and change direction. The overall effectof this erratic movement is the scattering or dispersion of theparticles throughout the environment (Figure 3.6). The greaterthe difference in concentration of the diffusing molecules andions between the two areas, the more collisions occur and thefaster the net diffusion of the particles.
Because the driving force for diffusion is the kinetic energy ofthe molecules themselves, the speed of diffusion is influenced bymolecular size (the smaller, the faster) and by temperature (thewarmer, the faster). In a closed container, diffusion eventuallyproduces a uniform mixture of molecules. In other words, thesystem reaches equilibrium, with molecules moving equally inall directions (no net movement).
Diffusion is occurring all around us, but obvious examples ofpure diffusion are almost impossible to see. The reason is thatany diffusion process that occurs over an easily observable dis-tance takes a long time, and is often accompanied by otherprocesses (convection, for example) that affect the movement ofmolecules and ions. In fact, one should suspect any readily ob-servable “example” of diffusion. Nonetheless, diffusion is im-mensely important in physiological systems and it occursrapidly because the distances molecules are moving are veryshort, perhaps 1/1000 (or less) the thickness of this page! Exam-ples include the movement of ions across cell membranes andthe movement of neurotransmitters between two nerve cells.
The plasma membrane is a physical barrier to free diffusionbecause of its hydrophobic core. However, a molecule will diffuse
68 UNIT 1 Organization of the Body
3 Figure 3.6 Diffusion. Molecules in solution move continuously and collide constantly withother molecules, causing them to move away from areas of their highest concentration andbecome evenly distributed. From left to right, molecules from a dye pellet diffuse into thesurrounding water down their concentration gradient.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 68

through the membrane if the molecule is (1) lipid soluble,(2) small enough to pass through membrane channels, or(3) assisted by a carrier molecule. The unassisted diffusion oflipid-soluble or very small particles is called simple diffusion. Aspecial name, osmosis, is given to the unassisted diffusion of asolvent (usually water) through a membrane. Assisted diffusionis known as facilitated diffusion.
Simple Diffusion In simple diffusion, nonpolar and lipid-soluble substances diffuse directly through the lipid bilayer(Figure 3.7a). Such substances include oxygen, carbon dioxide,and fat-soluble vitamins. Because oxygen concentration is al-ways higher in the blood than in tissue cells, oxygen continu-ously diffuses from the blood into the cells. Carbon dioxide, onthe other hand, is in higher concentration within the cells, so itdiffuses from tissue cells into the blood.
Facilitated Diffusion Certain molecules, notably glucose andother sugars, some amino acids, and ions are transported pas-sively even though they are unable to pass through the lipid bi-layer. Instead they move through the membrane by a passivetransport process called facilitated diffusion in which thetransported substance either (1) binds to protein carriers in themembrane and is ferried across or (2) moves through water-filled protein channels.
■ Carriers are transmembrane integral proteins that showspecificity for molecules of a certain polar substance or classof substances that are too large to pass through membranechannels, such as sugars and amino acids. The most popularmodel for the action of carriers indicates that changes in theshape of the carrier allow it to first envelop and then releasethe transported substance, shielding it en route from the
nonpolar regions of the membrane. Essentially, the bindingsite is moved from one face of the membrane to the other bychanges in the conformation of the carrier protein (Figure 3.7band Table 3.1).
Note that a substance transported by carrier-mediated fa-cilitated diffusion, such as glucose, moves down its concentra-tion gradient, just as in simple diffusion. Glucose is normallyin higher concentrations in the blood than in the cells, whereit is rapidly used for ATP synthesis. So, glucose transportwithin the body is typically unidirectional—into the cells.However, carrier-mediated transport is limited by the num-ber of protein carriers present. For example, when all the glu-cose carriers are “engaged,” they are said to be saturated, andglucose transport is occurring at its maximum rate.
■ Channels are transmembrane proteins that serve to trans-port substances, usually ions or water, through aqueouschannels from one side of the membrane to the other(Figure 3.7c and d). Binding or association sites exist withinthe channels, and the channels are selective due to pore sizeand the charges of the amino acids lining the channel. Somechannels, the so-called leakage channels, are always open andsimply allow ion or water fluxes according to concentrationgradients. Others are gated and are controlled (opened orclosed) by various chemical or electrical signals.
Like carriers, many channels can be inhibited by certainmolecules, show saturation, and tend to be specific. Sub-stances moving through them also follow the concentrationgradient (always moving down the gradient). When a sub-stance crosses the membrane by simple diffusion, the rate ofdiffusion is not controllable because the lipid solubility ofthe membrane is not immediately changeable. By contrast,the rate of facilitated diffusion is controllable because the
Chapter 3 Cells: The Living Units 69
3
Extracellular fluid
Lipid-solublesolutes
Cytoplasm
Lipid-insoluble solutes (such as sugars or amino acids)
Small lipid-insoluble solutes
Watermolecules
Lipidbillayer
Aquaporin
(a) Simple diffusion of fat-soluble molecules directly through the phospholipid bilayer
(b) Carrier-mediated facilitated diffusion via a protein carrier specific for one chemical; binding of substrate causes shape change in transport protein
(c) Channel-mediated facilitated diffusion through a channel protein; mostly ions selected on basis of size and charge
(d) Osmosis, diffusion of a solvent such as water through a specific channel protein (aquaporin) or through the lipid bilayer
Figure 3.7 Diffusion through the plasma membrane.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 69

permeability of the membrane can be altered by regulatingthe activity or number of individual carriers or channels.
Oxygen, water, glucose, and various ions are vitally impor-tant to cellular homeostasis. Their passive transport by diffusion(either simple or facilitated) represents a tremendous saving ofcellular energy. Indeed, if these substances had to be transportedactively, cell expenditures of ATP would increase exponentially!
Osmosis The diffusion of a solvent, such as water, through aselectively permeable membrane is osmosis (oz-mo�sis; osmos �pushing). Even though water is highly polar, it passes via os-mosis through the lipid bilayer (Figure 3.7d). This is surprisingbecause you’d expect water to be repelled by the hydrophobiclipid tails. Although still hypothetical, one explanation is that
random movements of the membrane lipids open small gapsbetween their wiggling tails, allowing water to slip and slide itsway through the membrane by moving from gap to gap.
Water also moves freely and reversibly through water-specificchannels constructed by transmembrane proteins calledaquaporins (AQPs). Although aquaporins are believed to bepresent in all cell types, they are particularly abundant in redblood cells and in cells involved in water balance such as kidneytubule cells.
Osmosis occurs whenever the water concentration differs onthe two sides of a membrane. If distilled water is present on bothsides of a selectively permeable membrane,no net osmosis occurs,even though water molecules move in both directions throughthe membrane. If the solute concentration on the two sides of
70 UNIT 1 Organization of the Body
3
(a) Membrane permeable to both solutes and water
(b) Membrane permeable to water, impermeable to solutes
Solute and water molecules move down their concentration gradientsin opposite directions. Fluid volume remains the same in both compartments.
Leftcompartment:Solution withlower osmolarity
Rightcompartment:Solution with greater osmolarity
Membrane
H2O
Solute
Solutemolecules(sugar)
Both solutions have thesame osmolarity: volumeunchanged
Both solutions have identicalosmolarity, but volume of thesolution on the right is greaterbecause only water is free to move
Solute molecules are prevented from moving but water moves by osmosis.Volume increases in the compartment with the higher osmolarity.
Leftcompartment
Rightcompartment
Membrane
Solutemolecules(sugar)
H2O
Figure 3.8 Influence of membrane permeability on diffusion and osmosis.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 70

Chapter 3 Cells: The Living Units 71
3
the membrane differs, water concentration differs as well (assolute concentration increases, water concentration decreases).
The extent to which water’s concentration is decreased bysolutes depends on the number, not the type, of solute particles,because one molecule or one ion of solute (theoretically) dis-places one water molecule. The total concentration of all soluteparticles in a solution is referred to as the solution’s osmolarity(oz�mo-lar�ı-te). When equal volumes of aqueous solutions ofdifferent osmolarity are separated by a membrane that ispermeable to all molecules in the system, net diffusion of bothsolute and water occurs, each moving down its own concentra-tion gradient. Eventually, equilibrium is reached when the waterconcentration on the left equals that on the right, and the soluteconcentration on both sides is the same (Figure 3.8a).
If we consider the same system, but make the membraneimpermeable to solute molecules, we see quite a different result(Figure 3.8b). Water quickly diffuses from the left to the rightcompartment and continues to do so until its concentration isthe same on the two sides of the membrane. Notice that in thiscase equilibrium results from the movement of water alone (thesolutes are prevented from moving). Notice also that the move-ment of water leads to dramatic changes in the volumes of thetwo compartments.
The last example mimics osmosis across plasma membranesof living cells, with one major difference. In our examples, thevolumes of the compartments are infinitely expandable and theeffect of pressure exerted by the added weight of the higher fluidcolumn is not considered. In living plant cells, which have rigidcell walls external to their plasma membranes, this is not thecase. As water diffuses into the cell, the point is finally reached
where the hydrostatic pressure (the back pressure exerted bywater against the membrane) within the cell is equal to itsosmotic pressure (the tendency of water to move into the cellby osmosis). At this point, there is no further (net) waterentry. As a rule, the higher the amount of nondiffusible, ornonpenetrating, solutes in a cell, the higher the osmotic pressureand the greater the hydrostatic pressure that must be developedto resist further net water entry.
However, such major changes in hydrostatic (and osmotic)pressures do not occur in living animal cells, which lack rigidcell walls. Osmotic imbalances cause animal cells to swell orshrink (due to net water gain or loss) until either the solute con-centration is the same on both sides of the plasma membrane,or the membrane is stretched to its breaking point.
Such changes in animal cells lead us to the important con-cept of tonicity (to-nis�ı-te). As noted, many molecules, particu-larly intracellular proteins and selected ions, are prevented fromdiffusing through the plasma membrane. Consequently, anychange in their concentration alters the water concentration onthe two sides of the membrane and results in a net loss or gainof water by the cell.
The ability of a solution to change the shape or tone of cellsby altering their internal water volume is called tonicity (tono �tension). Solutions with the same concentrations of nonpene-trating solutes as those found in cells (0.9% saline or 5% glu-cose) are isotonic (“the same tonicity”). Cells exposed to suchsolutions retain their normal shape, and exhibit no net loss orgain of water (Figure 3.9a). As you might expect, the body’sextracellular fluids and most intravenous solutions (solutionsinfused into the body via a vein) are isotonic.
(a) Isotonic solutions
Cells retain their normal size andshape in isotonic solutions (same
solute/water concentration as insidecells; water moves in and out).
(b) Hypertonic solutions
Cells lose water by osmosis and shrink in a hypertonic solution (contains a
higher concentration of solutes than are present inside the cells).
(c) Hypotonic solutions
Cells take on water by osmosis until they become bloated and burst (lyse) in a hypotonic solution (contains a lower
concentration of solutes than are present in cells).
Figure 3.9 The effect of solutions of varying tonicities on living red blood cells.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 71

Solutions with a higher concentration of nonpenetratingsolutes than seen in the cell (for example, a strong saline solu-tion) are hypertonic. Cells immersed in hypertonic solutionslose water and shrink, or crenate (kre�nat) (Figure 3.9b).
Solutions that are more dilute (contain a lower concentrationof nonpenetrating solutes) than cells are called hypotonic. Cellsplaced in a hypotonic solution plump up rapidly as water rushesinto them (Figure 3.9c). Distilled water represents the most ex-treme example of hypotonicity. Because it contains no solutes,water continues to enter cells until they finally burst or lyse.
Notice that osmolarity and tonicity are not the same thing. Asolution’s osmolarity is based solely on its total solute concentra-tion. In contrast, its tonicity is based on how the solution affectscell volume, which depends on (1) solute concentration and(2) solute permeability of the plasma membrane. Osmolarity isexpressed as osmoles per liter (osmol/L) where 1 osmol is equalto 1 mole of nonionizing molecules.* A 0.3-osmol/L solution ofNaCl is isotonic because sodium ions are usually prevented fromdiffusing through the plasma membrane. But if the cell is im-mersed in a 0.3-osmol/L solution of a penetrating solute, thesolute will enter the cell and water will follow. The cell will swelland burst, just as if it had been placed in pure water.
Osmosis is extremely important in determining distributionof water in the various fluid-containing compartments of thebody (in cells, in blood, and so on). In general, osmosis contin-ues until osmotic and hydrostatic pressures acting at the mem-brane are equal. For example, water is forced out of capillaryblood by the hydrostatic pressure of the blood against the capil-lary wall, but the presence in blood of solutes that are too large tocross the capillary membrane draws water back into the blood-stream. As a result, very little net loss of plasma fluid occurs.
Simple diffusion and osmosis occurring directly throughthe plasma membrane are not selective processes. In thoseprocesses, whether a molecule can pass through the membrane
depends chiefly on its size or its solubility in lipid, not on itsunique structure. Facilitated diffusion, on the other hand, isoften highly selective. The carrier for glucose, for example,combines specifically with glucose, in much the same way anenzyme binds to its specific substrate and ion channels allowonly selected ions to pass.
H O M E O S TAT I C I M B A L A N C E
Hypertonic solutions are sometimes infused intravenously intothe bloodstream of edematous patients (those swollen becausewater is retained in their tissues) to draw excess water out of theextracellular space and move it into the bloodstream so that itcan be eliminated by the kidneys. Hypotonic solutions may beused (with care) to rehydrate the tissues of extremely dehy-drated patients. In less extreme cases of dehydration, drinkinghypotonic fluids (colas, apple juice, and sports drinks) usuallydoes the trick. ■
Table 3.1 summarizes passive membrane transport processes.
C H E C K Y O U R U N D E R S TA N D I N G
7. What is the energy source for all types of diffusion?8. What determines the direction of any diffusion process?9. What are the two types of facilitated diffusion and how do
they differ?
For answers, see Appendix G.
Active Processes� Differentiate between primary and secondary active
transport.
� Compare and contrast endocytosis and exocytosis in termsof function and direction.
� Compare and contrast pinocytosis, phagocytosis, andreceptor-mediated endocytosis.
72 UNIT 1 Organization of the Body
3
*Osmolarity (Osm) is determined by multiplying molarity (moles per liter, or M)by the number of particles resulting from ionization. For example, since NaCl ion-izes to Na� � Cl�, a 1M solution of NaCl is a 2-Osm solution. For substances thatdo not ionize (e.g., glucose), molarity and osmolarity are the same.
Passive Membrane Transport Processes
PROCESS ENERGY SOURCE DESCRIPTION EXAMPLES
Diffusion
Simple diffusion Kinetic energy Net movement of molecules from an areaof their higher concentration to an areaof their lower concentration, that is, alongtheir concentration gradient
Movement of fats, oxygen, carbon dioxidethrough the lipid portion of the membrane
Facilitated diffusion Kinetic energy Same as simple diffusion, but the diffusingsubstance is attached to a lipid-solublemembrane carrier protein or movesthrough a membrane channel
Movement of glucose and some ions into cells
Osmosis Kinetic energy Simple diffusion of water through a selec-tively permeable membrane
Movement of water into and out of cellsdirectly through the lipid phase of the membrane or via membrane channels(aquaporins)
TABLE 3.1
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 72

Whenever a cell uses the bond energy of ATP to move solutesacross the membrane, the process is referred to as active. Sub-stances moved actively across the plasma membrane are usuallyunable to pass in the necessary direction by passive transportprocesses. The substance may be too large to pass through thechannels, incapable of dissolving in the lipid bilayer, or unableto move down its concentration gradient. There are two majormeans of active membrane transport: active transport andvesicular transport.
Active Transport
Active transport, like carrier-mediated facilitated diffusion, re-quires carrier proteins that combine specifically and reversiblywith the transported substances. However, facilitated diffusionalways follows concentration gradients because its driving forceis kinetic energy. In contrast, the active transporters or solutepumps move solutes, most importantly ions (such as Na�, K�,and Ca2�), “uphill” against a concentration gradient. To do thiswork, cells must expend the energy of ATP.
Active transport processes are distinguished according totheir source of energy. In primary active transport, the energy todo work comes directly from hydrolysis of ATP. In secondary ac-tive transport, transport is driven indirectly by energy stored inionic gradients created by operation of primary active transportpumps. Secondary active transport systems are all coupledsystems; that is, they move more than one substance at a time. Ifthe two transported substances are moved in the same direc-tion, the system is a symport system (sym � same). If the trans-ported substances “wave to each other” as they cross themembrane in opposite directions, the system is an antiportsystem (anti � opposite, against). Let’s examine these processesmore carefully.
Primary Active Transport In primary active transport, hy-drolysis of ATP results in the phosphorylation of the transportprotein. This step causes the protein to change its shape insuch a manner that it “pumps” the bound solute across themembrane.
Primary active transport systems include calcium and hy-drogen pumps, but the most investigated example of a primaryactive transport system is the operation of the sodium-potassium pump, for which the carrier, or “pump,” is an en-zyme called Na�-K� ATPase. In the body, the concentration ofK� inside the cell is some 10 times higher than that outside,and the reverse is true of Na�. These ionic concentration dif-ferences are essential for excitable cells like muscle and nervecells to function normally and for all body cells to maintaintheir normal fluid volume. Because Na� and K� leak slowlybut continuously through leakage channels in the plasmamembrane along their concentration gradient (and cross morerapidly in stimulated muscle and nerve cells), the Na�-K�
pump operates more or less continuously as an antiporter. It si-multaneously drives Na� out of the cell against a steep concen-tration gradient and pumps K� back in.
The electrochemical gradients maintained by the Na�-K�
pump underlie most primary and secondary active transport ofnutrients and ions, and are crucial for cardiac and skeletal mus-cle and neuron function.
The step-by-step operation of the Na�-K� pump is de-scribed in Focus on Primary Active Transport: The Na+-K+ Pump(Figure 3.10) on p. 74. Make sure you understand this processthoroughly before moving on to the topic of secondary activetransport.
Secondary Active Transport A single ATP-powered pump,such as the Na�-K� pump, can indirectly drive the secondaryactive transport of several other solutes. By moving sodiumacross the plasma membrane against its concentration gradient,the pump stores energy (in the ion gradient). Then, just as wa-ter pumped uphill can do work as it flows back down (to turn aturbine or water wheel), a substance pumped across a mem-brane can do work as it leaks back, propelled “downhill” alongits concentration gradient. In this way, as sodium moves backinto the cell with the help of a carrier protein (facilitated diffu-sion), other substances are “dragged along,” or cotransported,by a common carrier protein (Figure 3.11). A carrier movingtwo substances in the same direction is a symport system.
For example, some sugars, amino acids, and many ions arecotransported in this way into cells lining the small intestine.Both cotransported substances move passively because the en-ergy for this type of transport is the concentration gradient ofthe ion (in this case Na�). Na� has to be pumped back out intothe lumen (cavity) of the intestine to maintain its diffusion gra-dient. Ion gradients can also be used to drive antiport systemssuch as those that help to regulate intracellular pH by using thesodium gradient to expel hydrogen ions.
Regardless of whether the energy is provided directly(primary active transport) or indirectly (secondary activetransport), each membrane pump or cotransporter trans-ports only specific substances. For this reason, active transportsystems provide a way for the cell to be very selective in caseswhere substances cannot pass by diffusion. (No pump—notransport.)
Vesicular Transport
In vesicular transport, fluids containing large particles andmacromolecules are transported across cellular membranes in-side membranous sacs called vesicles. Vesicular transportprocesses that eject substances from the cell interior into the ex-tracellular fluid are called exocytosis (ek�so-si-to�sis;“out of thecell”). Those in which the cell ingests small patches of the plasma
Chapter 3 Cells: The Living Units 73
3
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 73

Extracellular fluid
6 K+ is released from the pump protein and Na+ sites are ready to bind Na+ again. The cycle repeats.
Primary active transport is the process in which ions are moved across cell membranes against electrochemical gradients using energy supplied directly by ATP. The action of the Na+-K+ pump is an important example of primary active transport.
2 Binding of Na+ promotes phosphorylation of the protein by ATP.
1 Cytoplasmic Na+ binds to pump protein.
Na+
Na+-K+ pump
K+ released
ATP-binding site Na+ bound
Cytoplasm
ATPADP
P
K+
5 K+ binding triggers release of the phosphate. Pump protein returns to its original conformation.
3 Phosphorylation causes the protein to change shape, expelling Na+ to the outside.
4 Extracellular K+ binds to pump protein.
Na+ released
K+ bound
P
K+
PPi
Figure 3.10 FOCUS Primary Active Transport: The Na+-K+ Pump
74
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 74

membrane and moves substances from the cell exterior to the cellinterior are called endocytosis (en�do-si-to�sis;“within the cell”).
Vesicular transport is also used for combination processessuch as transcytosis, moving substances into, across, and thenout of the cell, and substance, or vesicular, trafficking, movingsubstances from one area (or organelle) in the cell to another.
Like solute pumping, vesicular transport processes are ener-gized by ATP (or in some cases another energy-rich compound,GTP—guanosine triphosphate).
Endocytosis, Transcytosis, and Vesicular Trafficking Virtuallyall forms of vesicular transport involve an assortment ofprotein-coated vesicles of three types and, with some excep-tions, all are mediated by membrane receptors. Before we getspecific about each type of coated vesicular transport, let’s lookat the general scheme of endocytosis.
Protein-coated vesicles provide the main route for endocyto-sis and transcytosis of bulk solids, most macromolecules, andfluids. On occasion, these vesicles are also hijacked by pathogensseeking entry into a cell.
Figure 3.12 shows the basic steps in endocytosis and transcy-tosis. The substance to be taken into the cell by endocytosisis progressively enclosed by an infolding portion of the plasmamembrane called a coated pit. The coating is most often thebristlelike clathrin (kla�thrin; “lattice clad”) protein coatingfound on the cytoplasmic face of the pit. The clathrin coat(clathrin and some accessory proteins) acts both in cargo selec-tion and in deforming the membrane to produce the vesicle.
The vesicle detaches, and the coat proteins are recycledback to the plasma membrane.
The uncoated vesicle then typically fuses with a process-ing and sorting vesicle called an endosome. Some membranecomponents and receptors of the fused vesicle may be recycledback to the plasma membrane in a transport vesicle. The6
54
32
1
remaining contents of the vesicle may (a) combine with alysosome (li�so--so-m), a specialized cell structure containingdigestive enzymes, where the ingested substance is degraded orreleased (if iron or cholesterol), or (b) be transported com-pletely across the cell and released by exocytosis on the oppositeside (transcytosis). Transcytosis is common in the endothelialcells lining blood vessels because it provides a quick means toget substances from the blood to the interstitial fluid.
Based on the nature and quantity of material taken up andthe means of uptake, three types of endocytosis that useclathrin-coated vesicles are recognized: phagocytosis, pinocyto-sis, and receptor-mediated endocytosis.
Phagocytosis (fag�o-si-to�sis; “cell eating”) is the type of en-docytosis in which the cell engulfs some relatively large or solidmaterial, such as a clump of bacteria, cell debris, or inanimateparticles (asbestos fibers or glass, for example) (Figure 3.13a).When a particle binds to receptors on the cell’s surface, cyto-plasmic extensions called pseudopods (soo�do-pahdz; pseudo �false, pod � foot) form and flow around the particle and engulfit. The endocytotic vesicle formed in this way is called aphagosome (fag�o-som;“eaten body”). In most cases, the phago-some then fuses with a lysosome and its contents are digested.Any indigestible contents are ejected from the cell by exocytosis.
In the human body, only macrophages and certain whiteblood cells are “experts” at phagocytosis. Commonly referred toas phagocytes, these cells help police and protect the body by in-gesting and disposing of bacteria, other foreign substances, anddead tissue cells. The disposal of dying cells is crucial, becausedead cell remnants trigger inflammation in the surroundingarea or may stimulate an undesirable immune response. Mostphagocytes move about by amoeboid motion (ah-me�boyd;“changing shape”); that is, the flowing of their cytoplasm intotemporary pseudopods allows them to creep along.
Chapter 3 Cells: The Living Units 75
3
1 2The ATP-driven Na+-K+ pump stores energy by creating a steep concentration gradient for Na+ entry into the cell.
As Na+ diffuses back across the membranethrough a membrane cotransporter protein, itdrives glucose against its concentration gradientinto the cell. (ECF = extracellular fluid)
K+
Na+
Na+
Na+
Na+
Na+
Na+-glucosesymporttransporterloadingglucose fromECF
Na+-glucosesymport transporterreleasing glucoseinto the cytoplasm
Glucose
Na+-K+
pump
Cytoplasm
Extracellular fluid
ATP
Figure 3.11 Secondary active transport.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 75

that is present only in very small amounts in the extracellularfluid. The receptors for this process are plasma membrane pro-teins that bind only certain substances. Both the receptors andattached molecules are internalized in a clathrin-coated pit andthen dealt with in one of the ways discussed above. Substancestaken up by receptor-mediated endocytosis include enzymes,insulin (and some other hormones), low-density lipoproteins(such as cholesterol attached to a transport protein), and iron.Unfortunately, flu viruses, diphtheria, and cholera toxins usethis route to enter and attack our cells.
Other coat proteins are also used for certain types of vesiculartransport. Caveolae (ka�ve-o�le; “little caves”), tubular or flask-shaped inpocketings of the plasma membrane seen in many celltypes, are involved in a unique kind of receptor-mediated endo-cytosis called potosis. Like clathrin-coated pits, caveolae capturespecific molecules (folic acid, tetanus toxin) from the extracellularfluid in coated vesicles and participate in some forms of trans-cytosis. However, caveolae are smaller than clathrin-coated
76 UNIT 1 Organization of the Body
3
Coated pit ingests substance.
Protein-coated vesicle detaches.
Coat proteins detach and are recycled to plasma membrane.
Uncoated vesicle fuses with a sorting vesicle called an endosome.
Transport vesicle containing membrane components moves to the plasma membrane for recycling.
Fused vesicle may (a) fuse with lysosome for digestion of its contents, or (b) deliver its contents to the plasma membrane on the opposite side of the cell (transcytosis).
Protein coat(typically clathrin)
Extracellular fluidPlasma membrane
Endosome
Lysosome
Transportvesicle
(b)(a)
Uncoatedendocyticvesicle
Cytoplasm
1
2
3
4 5
6
Figure 3.12 Events of endocytosis mediated by protein-coated pits. Note the threepossible fates for a vesicle and its contents, shown in and .65
In pinocytosis (“cell drinking”), also called fluid-phaseendocytosis, a bit of infolding plasma membrane (which beginsas a clathrin-coated pit) surrounds a very small volume of extra-cellular fluid containing dissolved molecules (Figure 3.13b).This droplet enters the cell and fuses with an endosome. Unlikephagocytosis, pinocytosis is a routine activity of most cells, af-fording them a nonselective way of sampling the extracellularfluid. It is particularly important in cells that absorb nutrients,such as cells that line the intestines.
As mentioned, bits of the plasma membrane are removedwhen the membranous sacs are internalized. However, thesemembranes are recycled back to the plasma membrane by exo-cytosis as described shortly, so the surface area of the plasmamembrane remains remarkably constant.
Receptor-mediated endocytosis is the main mechanism forthe specific endocytosis and transcytosis of most macromole-cules by body cells, and it is exquisitely selective (Figure 3.13c).It is also the mechanism that allows cells to concentrate material
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 76

Chapter 3 Cells: The Living Units 77
3
vesicles. Additionally, their cage-like protein coat is thinner andcomposed of a different protein called caveolin.
Caveolae are closely associated with lipid rafts that are plat-forms for G proteins, receptors for hormones (for example, in-sulin), and enzymes involved in cell regulation. These vesiclesappear to provide sites for cell signaling and cross talk between
signaling pathways. Their precise role in the cell is still beingworked out.
Vesicles coated with coatomer (COP1 and COP2) proteinsare used in most types of intracellular vesicular trafficking, inwhich vesicles transport substances between organelles.
Active Membrane Transport Processes
PROCESS ENERGY SOURCE DESCRIPTION EXAMPLES
Active Transport
Primary active transport
ATP Transport of substances against a concentration(or electrochemical) gradient. Performed acrossthe plasma membrane by a solute pump, directlyusing energy of ATP hydrolysis.
Ions (Na�, K�, H�, Ca2�, and others)
Secondary activetransport
Ion concentrationgradient main-tained with ATP
Cotransport (coupled transport) of two solutesacross the membrane. Energy is supplied indi-rectly by the ion gradient created by primary ac-tive transport. Symporters move the transportedsubstances in the same direction; antiportersmove transported substances in opposite direc-tions across the membrane.
Movement of polar or charged sol-utes, e.g., amino acids (into cell bysymporters); Ca2+, H+ (out of cells viaantiporters)
Vesicular Transport
Exocytosis ATP Secretion or ejection of substances from a cell.The substance is enclosed in a membranous vesi-cle, which fuses with the plasma membrane andruptures, releasing the substance to the exterior.
Secretion of neurotransmitters, hormones, mucus, etc.; ejection of cell wastes
Endocytosis■ Via clathrin-coated
vesicles
ATP
Phagocytosis ATP “Cell eating”: A large external particle (proteins,bacteria, dead cell debris) is surrounded by a “seizing foot” and becomes enclosed in a vesicle(phagosome).
In the human body, occurs pri-marily in protective phagocytes (some white blood cells and mac-rophages)
Pinocytosis (fluid-phase endocytosis)
ATP Plasma membrane sinks beneath an external fluiddroplet containing small solutes. Membrane edgesfuse, forming a fluid-filled vesicle.
Occurs in most cells; important fortaking in dissolved solutes by absorp-tive cells of the kidney and intestine
Receptor-mediatedendocytosis
ATP Selective endocytosis and transcytosis. Externalsubstance binds to membrane receptors.
Means of intake of some hormones,cholesterol, iron, and most macromol-ecules
■ Via caveolin-coatedvesicles (caveolae)
ATP Selective endocytosis (and transcytosis). Externalsubstance binds to membrane receptors (oftenassociated with lipid rafts).
Roles not fully known; proposed rolesinclude cholesterol regulation andtrafficking, and platforms for signaltransduction
Intracellular vesiculartrafficking
■ Via coatomer-coated vesicles
ATP Vesicles pinch off from organelles and travel toother organelles to deliver their cargo.
Accounts for nearly all intracellulartrafficking between certain organ-elles (endoplasmic reticulum andGolgi apparatus). Exceptions includevesicles budding from the trans faceof the Golgi apparatus, which areclathrin-coated.
TABLE 3.2
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 77

78 UNIT 1 Organization of the Body
3
1 The membrane-bound vesicle migrates to the plasma membrane.
2 There, proteins at the vesicle surface (v-SNAREs) bind with t-SNAREs (plasma membrane proteins).
3 The vesicle and plasma membrane fuse and a pore opens up.
4 Vesicle contents are released to the cell exterior.
(a) The process of exocytosis
(b) Photomicrograph of a secretory vesicle releasing its contents by exocytosis (100,000�)
Extracellularfluid
Plasma membraneSNARE (t-SNARE)
Secretoryvesicle Vesicle SNARE
(v-SNARE)
Molecule tobe secreted
Cytoplasm
Fusedv- andt-SNAREs
Fusion pore formed
Figure 3.14Exocytosis.
Phagosome
Vesicle
Vesicle
Receptor recycledto plasma membrane
(a) PhagocytosisThe cell engulfs a large particle by forming pro-jecting pseudopods (”false feet”) around it and en-closing it within a membrane sac called a phagosome. The phagosome is combined with a lysosome. Undigested contents remain in the vesicle (now called a residual body) or are ejected by exocytosis. Vesicle may or may not be protein-coated but has receptors capable of binding to microorganisms or solid particles.
(b) PinocytosisThe cell “gulps” drops of extracellular fluid containing solutes into tiny vesicles. No receptors are used, so the process is nonspecific. Most vesicles are protein-coated.
(c) Receptor-mediated endocytosisExtracellular substances bind to specific receptor proteins in regions of coated pits, enabling the cell to ingest and concentrate specific substances (ligands) in protein-coated vesicles. Ligands may simply be released inside the cell, or combined with a lysosome to digest contents. Receptors are recycled to the plasma membrane in vesicles.
Figure 3.13 Comparison of three types of endocytosis.
Exocytosis The process of exocytosis, typically stimulated by acell-surface signal such as binding of a hormone to a membranereceptor or a change in membrane voltage, accounts for hor-mone secretion, neurotransmitter release, mucus secretion, andin some cases, ejection of wastes. The substance to be removedfrom the cell is first enclosed in a protein-coated membranoussac called a vesicle. In most cases, the vesicle migrates to theplasma membrane, fuses with it, and then ruptures, spilling thesac contents out of the cell (Figure 3.14).
Exocytosis, like other cases in which vesicles are targeted totheir destinations, involves a “docking” process in which trans-membrane proteins on the vesicles, fancifully called v-SNAREs(v for vesicle), recognize certain plasma membrane proteins,
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 78

called t-SNAREs (t for target), and bind with them. This bindingcauses the membranes to “corkscrew”together and fuse, rearrang-ing the lipid monolayers without mixing them (Figure 3.14a). Asdescribed, membrane material added by exocytosis is removedby endocytosis—the reverse process.
Table 3.2 summarizes active membrane transport processes.
C H E C K Y O U R U N D E R S TA N D I N G
10. What happens when the Na+-K+ pump is phosphorylated?When K+ binds to the pump protein?
11. As a cell grows, its plasma membrane expands. Does thismembrane expansion involve endocytosis or exocytosis?
12. Phagocytic cells gather in the lungs, particularly in the lungsof smokers. What is the connection?
13. What vesicular transport process allows a cell to take in cho-lesterol from the extracellular fluid?
For answers, see Appendix G.
The Plasma Membrane: Generationof a Resting Membrane Potential� Define membrane potential and explain how the resting
membrane potential is established and maintained.
As you’re now aware, the selective permeability of the plasmamembrane can lead to dramatic osmotic flows, but that is not itsonly consequence. An equally important result is the generationof a membrane potential, or voltage, across the membrane. A
voltage is electrical potential energy resulting from the separa-tion of oppositely charged particles. In cells, the oppositelycharged particles are ions, and the barrier that keeps them apartis the plasma membrane.
In their resting state, all body cells exhibit a resting membranepotential that typically ranges from –50 to –100 millivolts (mV),depending on cell type. For this reason, all cells are said to bepolarized. The minus sign before the voltage indicates that theinside of the cell is negative compared to its outside. This voltage(or charge separation) exists only at the membrane. If we were toadd up all the negative and positive charges in the cytoplasm, wewould find that the cell interior is electrically neutral. Likewise,the positive and negative charges in the extracellular fluid bal-ance each other exactly.
So how does the resting membrane potential come about,and how is it maintained? The short answer is that diffusioncauses ionic imbalances that polarize the membrane, and activetransport processes maintain that membrane potential. First,let’s look at how diffusion polarizes the membrane.
Many kinds of ions are found both inside cells and in the ex-tracellular fluid, but the resting membrane potential is deter-mined mainly by the concentration gradient of potassium (K�)and by the differential permeability of the plasma membraneto K� and other ions (Figure 3.15). Recall that K� and proteinanions predominate inside body cells, and the extracellular fluidcontains relatively more Na�, which is largely balanced by Cl–.The unstimulated plasma membrane is somewhat permeable toK� because of leakage channels, but impermeable to the proteinanions. Consequently, K� diffuses out of the cell along its con-centration gradient but the protein anions are unable to follow,
Chapter 3 Cells: The Living Units 79
3
1
2
3
K+ diffuse down their steep concentration gradient (out of the cell) via leakage channels. Loss of K+ results in a negative charge on the inner plasma membrane face.
K+ also move into the cell because they are attracted to the negative charge established on the inner plasma membrane face.
A negative membrane potential (–90 mV) is established when the movement of K+ out of the cell equals K+ movement into the cell. At this point, the concentration gradient promoting K+ exit exactly opposes the electrical gradient for K+ entry.
Na+Na+
Na+
K+ K+K+
K+K+
K+
K+ K+
K+
K+
K+ K+ K+
K+
Cl–Cl–
Potassiumleakagechannels
Protein anion (unable tofollow K+ through themembrane)Cytoplasm
Extracellular fluid
+ + + ++ +
++
– – – ––
–––
A–
A–
Figure 3.15 The key role of K+ in generating the resting membrane potential. Theresting membrane potential is largely determined by K+ because the membrane is much morepermeable to K+ than Na+ at rest. The active transport of sodium and potassium ions (in a ratioof 3:2) by the Na+-K+ pump maintains these conditions.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 79

and this loss of positive charges makes the membrane interiormore negative (Figure 3.15). As more and more K� leaves thecell, the negativity of the inner membrane face becomes greatenough to attract K� back toward and even into the cell. At thispoint (–90 mV), potassium’s concentration gradient is exactlybalanced by the electrical gradient (membrane potential), andone K� enters the cell as one leaves.
In many cells, sodium (Na�) also contributes to the restingmembrane potential. Sodium is strongly attracted to the cell in-terior by its concentration gradient, bringing the resting mem-brane potential to –70 mV. However, potassium still largelydetermines the resting membrane potential because the mem-brane is much more permeable to K� than to Na�. Even thoughthe membrane is permeable to Cl–, in most cells Cl– does notcontribute to the resting membrane potential, because its entryis resisted by the negative charge of the interior. We may betempted to believe that massive flows of K� ions are needed togenerate the resting potential, but this is not the case. Surpris-ingly, the number of ions producing the membrane potential isso small that it does not change ion concentrations in any signif-icant way.
In a cell at rest, very few ions cross its plasma membrane.However, Na� and K� are not at equilibrium and there is somenet movement of K� out of the cell and of Na� into the cell. Na�
is strongly pulled into the cell by both its concentration gradientand the interior negative charge. If only passive forces were atwork, these ion concentrations would eventually become equalinside and outside the cell.
Now let’s look at how active transport processes maintainthe membrane potential that diffusion has established, with theresult that the cell exhibits a steady state. The rate of activetransport is equal to, and depends on, the rate of Na� diffusioninto the cell. If more Na� enters, more is pumped out. (This islike being in a leaky boat. The more water that comes in, thefaster you bail!) The Na�-K� pump couples sodium andpotassium transport and, on average, each “turn” of the pumpejects 3Na� out of the cell and carries 2K� back in (see Figure3.10). Because the membrane is always 50 to 100 times morepermeable to K�, the ATP-dependent Na�-K� pump main-tains both the membrane potential (the charge separation) andthe osmotic balance. Indeed, if Na� was not continuouslyremoved from cells, in time so much would accumulate intra-cellularly that the osmotic gradient would draw water into thecells, causing them to burst.
Now that we’ve introduced the membrane potential, we canadd a detail or two to our discussion of diffusion. Earlier we saidthat solutes diffuse down their concentration gradient. This istrue for uncharged solutes, but only partially true for ions. Thenegatively and positively charged faces of the plasma membranecan help or hinder diffusion of ions driven by a concentrationgradient. It is more correct to say that ions diffuse according toelectrochemical gradients, thereby recognizing the effect ofboth electrical and concentration (chemical) forces.
Consequently, although diffusion of K� across the plasmamembrane is aided by the membrane’s greater permeability to itand by the ion’s concentration gradient, its diffusion is resistedsomewhat by the positive charge on the cell exterior. In contrast,Na� is drawn into the cell by a steep electrochemical gradient,
and the limiting factor is the membrane’s relative impermeabil-ity to it. As we will describe in detail in later chapters, “upset-ting” the resting membrane potential by transient opening ofNa� and K� channels in the plasma membrane is a normalmeans of activating neurons and muscle cells.
C H E C K Y O U R U N D E R S TA N D I N G
14. What event or process establishes the resting membranepotential?
15. Is the inside of the plasma membrane negative or positiverelative to its outside in a polarized membrane?
For answers, see Appendix G.
The Plasma Membrane: Cell-Environment Interactions� Describe the role of the glycocalyx when cells interact with
their environment.
� List several roles of membrane receptors and that ofvoltage-sensitive membrane channel proteins.
Cells are biological minifactories and, like other factories, theyreceive and send orders from and to the outside community.But how does a cell interact with its environment, and what ac-tivates it to carry out its homeostatic functions?
Although cells sometimes interact directly with other cells,this is not always the case. In many cases cells respond to extra-cellular chemicals, such as hormones and neurotransmittersdistributed in body fluids. Cells also interact with extracellularmolecules that act as signposts to guide cell migration duringdevelopment and repair.
Whether cells interact directly or indirectly,however, the glyco-calyx is always involved. The best understood of the participatingglycocalyx molecules fall into two large families—cell adhesionmolecules and plasma membrane receptors (see Figure 3.4). An-other group of membrane proteins, the voltage-sensitive channelproteins, are important in cells that respond to electrical signals.
Roles of Cell Adhesion MoleculesThousands of cell adhesion molecules (CAMs) are found onalmost every cell in the body. They play key roles in embryonicdevelopment and wound repair (situations where cell mobilityis important) and in immunity. These sticky glycoproteins(cadherins and integrins) act as
1. The molecular “Velcro” that cells use to anchor themselvesto molecules in the extracellular space and to each other(see desmosome discussion on pp. 66–67)
2. The “arms” that migrating cells use to haul themselves pastone another
3. SOS signals sticking out from the blood vessel lining thatrally protective white blood cells to a nearby infected or in-jured area
80 UNIT 1 Organization of the Body
3
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 80

Chapter 3 Cells: The Living Units 81
3
4. Mechanical sensors that respond to local tension at the cellsurface by stimulating synthesis or degradation of adhe-sive membrane junctions
5. Transmitters of intracellular signals that direct cell migra-tion, proliferation, and specialization
Roles of Membrane ReceptorsA huge and diverse group of integral proteins and glycoproteinsthat serve as binding sites are collectively known as membranereceptors. Some function in contact signaling, and others inchemical signaling. Let’s take a look.
Contact Signaling Contact signaling is the actual coming to-gether and touching of cells, and it is the means by which cellsrecognize one another. It is particularly important for normaldevelopment and immunity. Some bacteria and other infec-tious agents use contact signaling to identify their “preferred”target tissues or organs.
Chemical Signaling Most plasma membrane receptors are in-volved in chemical signaling, and this group will receive the bulkof our attention. Signaling chemicals that bind specifically toplasma membrane receptors are called ligands. Among theseligands are most neurotransmitters (nervous system signals),hormones (endocrine system signals), and paracrines (chemicalsthat act locally and are rapidly destroyed).
Different cells respond in different ways to the same ligand.Acetylcholine, for instance, stimulates skeletal muscle cells tocontract, but inhibits heart muscle. Why do different cells havesuch different responses? The reason is that a target cell’s re-sponse depends on the internal machinery that the receptor islinked to, not the specific ligand that binds to it.
Though cell responses to receptor binding vary widely, there isa fundamental similarity.When a ligand binds to a membrane re-ceptor, the receptor’s structure changes, and cell proteins are al-tered in some way. For example, muscle proteins change shape togenerate force. Some membrane receptor proteins are catalyticproteins that function as enzymes. Others, such as the chemicallygated channel-linked receptors common in muscle and nerve cells,respond to ligands by transiently opening or closing ion gates,which in turn changes the excitability of the cell.
Still other receptors are coupled to enzymes or ion channelsby a regulatory molecule called a G protein. Because nearlyevery cell in the body displays at least some of these receptors,we will spend a bit more time with them. The lipid rafts men-tioned earlier group together many receptor-mediated ele-ments, thus facilitating cell signaling.
G protein–linked receptors exert their effect indirectlythrough a G protein, which acts as a middleman or relay to ac-tivate (or inactivate) a membrane-bound enzyme or ion chan-nel. As a result, one or more intracellular chemical signals,commonly called second messengers, are generated and con-nect plasma membrane events to the internal metabolic ma-chinery of the cell. Two very important second messengers arecyclic AMP and ionic calcium, both of which typically activateprotein kinase enzymes, which transfer phosphate groups from
ATP to other proteins. In this way, the protein kinases can acti-vate a whole series of enzymes that bring about the desired cel-lular activity. Because a single enzyme can catalyze hundreds ofreactions, the amplification effect of such a chain of events istremendous. Focus on G Proteins (Figure 3.16) on p. 82 describeshow a G protein signaling system works. Take a moment tostudy this carefully because this key signaling pathway is in-volved in neurotransmission, smell, vision, and hormone action(Chapters 11, 15, and 16).
One important signaling molecule must be mentioned eventhough it doesn’t act in any of the ways we have already de-scribed. Nitric oxide (NO), one of nature’s simplest molecules, ismade of a single atom of nitrogen and one of oxygen. It is alsoan environmental pollutant and the first gas known to act as abiological messenger. Because of its tiny size, it slips into and outof cells easily. Its unpaired electron makes it highly reactive andit reacts with head-spinning speed with other key molecules tospur cells into a broad array of activities. You will be hearingmore about NO later (in the neural, cardiovascular, and im-mune system chapters).
Role of Voltage-Sensitive Membrane Channel ProteinsElectrical Signaling In the process known as electrical signal-ing, certain plasma membrane proteins are channel proteinsthat respond to changes in membrane potential by opening orclosing the channel. Such voltage-gated channels are commonin excitable tissues like neural and muscle tissues, and are indis-pensable to their normal functioning.
C H E C K Y O U R U N D E R S TA N D I N G
16. What term is used to indicate signaling chemicals that bindto membrane receptors? Which type of membrane receptoris most important in directing intracellular events by promot-ing formation of second messengers?
For answer, see Appendix G.
The Cytoplasm� Describe the composition of the cytosol. Define inclusions
and list several types.
� Discuss the structure and function of mitochondria.
� Discuss the structure and function of ribosomes, the endo-plasmic reticulum, and the Golgi apparatus, including func-tional interrelationships among these organelles.
� Compare the functions of lysosomes and peroxisomes.
Cytoplasm (“cell-forming material”) is the cellular material be-tween the plasma membrane and the nucleus. It is the site wheremost cellular activities are accomplished. Although early micro-scopists thought that the cytoplasm was a structureless gel, theelectron microscope reveals that it consists of three major ele-ments: the cytosol, organelles, and inclusions.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 81

1 2 3Ligand (1st messenger) binds to the receptor. The receptor is activated and changes shape.
The activated receptor binds to a G protein and activates it. During activation, a G protein changes shape (turns “on”) causing it to release GDP and bind GTP.
Activated G protein activates (or inactivates) effector protein (e.g., an enzyme) by causing its shape to change.
G proteins act as middlemen or relays between extracellular first messengers and intracellular second messengers that cause responses within the cell.
The sequence described here is like a molecular relay race. Instead of a baton passed from runner to runner, the message is passed from molecule to molecule as it makes its way across the cell membrane from outside to inside the cell.
Extracellular fluid
Intracellular fluid
GTP
GTPGDP
Inactive 2nd messenger
Active 2ndmessenger
Activatedkinaseenzymes
Ligand (1st messenger)
Receptor
Effector protein(e.g., an enzyme)
Receptor
G protein
Ligand
G protein Enzyme 2ndmessenger
GTP
Cascade of cellular responses (metabolic and structural changes)
4 Activated effector enzymes catalyze reactions that produce 2nd messengers in the cell.(Common 2nd messengers include cyclic AMP and Ca2�.)
5 Second messengers activate other enzymes or ion channels. Cyclic AMP typically activates protein kinase enzymes.
6 Kinase enzymes transfer phosphate groups from ATP to specific proteins and activate a series of other enzymes that trigger various cell responses.
Figure 3.16 FOCUS G Proteins
82
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 82

The cytosol (si�to-sol) is the viscous, semitransparent fluidin which the other cytoplasmic elements are suspended. It is acomplex mixture with properties of both a colloid and a truesolution. Dissolved in the cytosol, which is largely water, areproteins, salts, sugars, and a variety of other solutes.
The cytoplasmic organelles are the metabolic machinery ofthe cell. Each type of organelle carries out a specific function forthe cell—some synthesize proteins, others package those pro-teins, and so on.
Inclusions are chemical substances that may or may not bepresent, depending on cell type. Examples include stored nutri-ents, such as the glycogen granules abundant in liver and mus-cle cells; lipid droplets common in fat cells; pigment (melanin)granules seen in certain cells of skin and hair; water-containingvacuoles; and crystals of various types.
Cytoplasmic OrganellesThe cytoplasmic organelles (“little organs”) are specialized cel-lular compartments, each performing its own job to maintainthe life of the cell. Some organelles, the nonmembranous or-ganelles, lack membranes. Examples are the cytoskeleton, centri-oles, and ribosomes.
Most organelles, however, are bounded by a membrane simi-lar in composition to the plasma membrane. This membraneenables such membranous organelles to maintain an internal en-vironment different from that of the surrounding cytosol. Thiscompartmentalization is crucial to cell functioning. Without it,thousands of enzymes would be randomly mixed and biochem-ical activity would be chaotic. The cell’s membranous organellesinclude the mitochondria, peroxisomes, lysosomes, endoplasmicreticulum, and Golgi apparatus.
Besides providing “splendid isolation,” an organelle’s mem-brane often unites it with the rest of an interactive intracellularsystem called the endomembrane system (see p. 87) and the lipidmakeup of its membrane allows it to recognize and interactwith other organelles. Now, let us consider what goes on in eachof the workshops of our cellular factory.
Mitochondria
Mitochondria (mi�to-kon�dre-ah) are threadlike (mitos �thread) or lozenge-shaped membranous organelles. In livingcells they squirm, elongate, and change shape almost continu-ously. They are the power plants of a cell, providing most of itsATP supply. The density of mitochondria in a particular cell re-flects that cell’s energy requirements, and mitochondria are gen-erally clustered where the action is. Busy cells like kidney andliver cells have hundreds of mitochondria, whereas relatively in-active cells (such as unchallenged lymphocytes) have just a few.
Mitochondria are enclosed by two membranes, each with thegeneral structure of the plasma membrane (Figure 3.17). Theouter membrane is smooth and featureless, but the inner mem-brane folds inward, forming shelflike cristae (krı�ste; “crests”)that protrude into the matrix, the gel-like substance within themitochondrion. Intermediate products of food fuels (glucoseand others) are broken down to water and carbon dioxide by
teams of enzymes, some dissolved in the mitochondrial matrixand others forming part of the crista membrane.
As the metabolites are broken down and oxidized, some ofthe energy released is captured and used to attach phosphategroups to ADP molecules to form ATP. This multistep mito-chondrial process (described in Chapter 24) is generally re-ferred to as aerobic cellular respiration (a-er-o�bik) because itrequires oxygen.
Mitochondria are complex organelles: They contain theirown DNA, RNA, and ribosomes and are able to reproducethemselves. Mitochondrial genes (some 37 of them) direct thesynthesis of 1% of the proteins required for mitochondrialfunction, and the DNA of the cell’s nucleus encodes the remain-ing proteins needed to carry out cellular respiration. When cel-lular requirements for ATP increase, the mitochondriasynthesize more cristae or simply pinch in half (a process calledfission) to increase their number, then grow to their former size.
Intriguingly, mitochondria are similar to a specific group ofbacteria (the purple bacteria phylum), and mitochondrial DNA
Chapter 3 Cells: The Living Units 83
3
Enzymes
Matrix
Cristae
Mitochondrial DNA
Ribosome
Outer mitochondrial membrane
Innermitochondrial membrane
(b)
(a)
(c)
Figure 3.17 Mitochondrion. (a) Diagrammatic view of a longitudi-nally sectioned mitochondrion. (b) Close-up view of a crista showingenzymes (stalked particles). (c) Electron micrograph of a mitochon-drion (50,000�).
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 83

is bacteria-like. It is widely believed that mitochondria arosefrom bacteria that invaded the ancient ancestors of plant andanimal cells, and that this unique merger gave rise to all com-plex cells.
Ribosomes
Ribosomes (ri�bo-somz) are small, dark-staining granulescomposed of proteins and a variety of RNAs called ribosomalRNAs. Each ribosome has two globular subunits that fit togetherlike the body and cap of an acorn. Ribosomes are sites of proteinsynthesis, a function we discuss in detail later in this chapter.
Some ribosomes float freely in the cytoplasm. Others are at-tached to membranes, forming a complex called the rough en-doplasmic reticulum (Figure 3.18). These two ribosomalpopulations appear to divide the chore of protein synthesis. Freeribosomes make soluble proteins that function in the cytosol, aswell as those imported into mitochondria and some other or-ganelles. Membrane-bound ribosomes synthesize proteins des-tined either for incorporation into cell membranes or for exportfrom the cell. Ribosomes can switch back and forth betweenthese two functions, attaching to and detaching from the mem-branes of the endoplasmic reticulum, according to the type ofprotein they are making at a given time.
Endoplasmic Reticulum
The endoplasmic reticulum (ER) (en�do-plaz�mik re-tik�u-lum;“network within the cytoplasm”) is an extensive system of inter-connected tubes and parallel membranes enclosing fluid-filled
cavities, or cisternae (sis-ter�ne). Coiling and twisting throughthe cytosol, the ER is continuous with the nuclear membraneand accounts for about half of the cell’s membranes. There aretwo distinct varieties of ER: rough ER and smooth ER.
Rough Endoplasmic Reticulum The external surface of therough ER is studded with ribosomes (Figure 3.18a, b). Pro-teins assembled on these ribosomes thread their way into thefluid-filled interior of the ER cisternae, where various fatesawait them (as described on p. 106). When complete, the newlymade proteins are enclosed in coatomer-coated vesicles fortheir journey to the Golgi apparatus where they undergo fur-ther processing.
The rough ER has several functions. Its ribosomes manufac-ture all proteins secreted from cells. For this reason, the roughER is particularly abundant and well developed in most secre-tory cells, antibody-producing plasma cells, and liver cells,which produce most blood proteins. It is also the cell’s “mem-brane factory” because integral proteins and phospholipids thatform part of all cellular membranes are manufactured there.The enzymes needed to catalyze lipid synthesis have their activesites on the external (cytosolic) face of the ER membrane, wherethe needed substrates are readily available.
Smooth Endoplasmic Reticulum The smooth ER (see Figures3.2 and 3.18a, b) is continuous with the rough ER and consistsof tubules arranged in a looping network. Its enzymes (all inte-gral proteins forming part of its membranes) play no role in
84 UNIT 1 Organization of the Body
3
Nuclearenvelope
Ribosomes
Rough ER
(a) Diagrammatic view of smooth and rough ER
Smooth ER
(b) Electron micrograph of smooth and rough ER (10,000�)
Figure 3.18 The endoplasmic reticulum.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 84

protein synthesis. Instead, they catalyze reactions involved withthe following processes:
1. Lipid metabolism, cholesterol synthesis, and synthesis ofthe lipid components of lipoproteins (in liver cells)
2. Synthesis of steroid-based hormones such as sex hor-mones (testosterone-synthesizing cells of the testes are fullof smooth ER)
3. Absorption, synthesis, and transport of fats (in intesti-nal cells)
4. Detoxification of drugs, certain pesticides, and carcino-gens (in liver and kidneys)
5. Breakdown of stored glycogen to form free glucose (inliver cells especially)
Additionally, skeletal and cardiac muscle cells have an elabo-rate smooth ER (called the sarcoplasmic reticulum) that playsan important role in calcium ion storage and release duringmuscle contraction. Except for the examples given above, mostbody cells contain little, if any, true smooth ER.
Golgi Apparatus
The Golgi apparatus (gol�je) consists of stacked and flattenedmembranous sacs, shaped like hollow dinner plates, associated
with swarms of tiny membranous vesicles (Figure 3.19). TheGolgi apparatus is the principal “traffic director” for cellularproteins. Its major function is to modify, concentrate, and pack-age the proteins and lipids made at the rough ER.
The transport vesicles that bud off from the rough ER moveto and fuse with the membranes at the convex cis face, the“receiving” side, of the Golgi apparatus. Inside the apparatus,the proteins are modified: Some sugar groups are trimmedwhile others are added, and in some cases, phosphate groups areadded. The various proteins are “tagged” for delivery to a spe-cific address, sorted, and packaged in at least three types of vesi-cles that bud from the concave trans face (the “shipping” side) ofthe Golgi stack.
Vesicles containing proteins destined for export pinch offfrom the trans face as secretory vesicles, or granules, which mi-grate to the plasma membrane and discharge their contentsfrom the cell by exocytosis (pathway A, Figure 3.20). Specializedsecretory cells, such as the enzyme-producing cells of thepancreas, have a very prominent Golgi apparatus. Besides itspackaging-for-release function, the Golgi apparatus pinches offvesicles containing lipids and transmembrane proteins destinedfor a “home” in the plasma membrane (pathway B, Figure 3.20)or other membranous organelles. It also packages digestive
Chapter 3 Cells: The Living Units 85
3
Figure 3.19 Golgi apparatus. Note: In (a), the various and abundant vesicles shown in theprocess of pinching off from the membranous Golgi apparatus would have a protein coating ontheir external surfaces. These proteins are omitted from the diagram for simplicity.
Cis face—“receiving” side of Golgi apparatus
Secretory vesicle
(a) Many vesicles in the process of pinching off from the membranous Golgi apparatus.
(b) Electron micrograph of the Golgi apparatus (90,000�)
Transport vesicle from the Golgi apparatus
Transportvesiclefromtrans face
Trans face—“shipping” side ofGolgi apparatus
New vesiclesforming
New vesicles forming
Cisternae
Transport vesiclefrom rough ER
Golgi apparatus
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 85

86 UNIT 1 Organization of the Body
3
Protein-containing vesicles pinch off rough ER and migrate to fuse with membranes of Golgi apparatus.
Proteins are modified within the Golgi compartments.
Proteins are then packaged within different vesicle types, depending on their ultimate destination.
Plasmamembrane
Secretion by exocytosis
Vesicle becomeslysosome
Golgi apparatus
Rough ER ER membrane Phagosome
Proteins in cisterna
Pathway B:Vesicle membraneto be incorporatedinto plasmamembranePathway A:
Vesicle contentsdestined forexocytosis
Extracellular fluid
Secretoryvesicle
Pathway C:Lysosomecontaining acidhydrolaseenzymes
1
3
2
Figure 3.20 The sequence of events from protein synthesis on the rough ER to the finaldistribution of those proteins. The protein coats on the transport vesicles are not illustrated.
Lysosomes
Light areas are regions where materials are being digested.
Figure 3.21 Electron micrograph of a cell containing lysosomes (12,000�).
enzymes into membranous lysosomes that remain in the cell(pathway C in Figure 3.20, and discussed next).
Lysosomes
Born as endosomes which contain inactive enzymes, lysosomes(“disintegrator bodies”) are spherical membranous organellescontaining activated digestive enzymes (Figure 3.21).As you mightguess, lysosomes are large and abundant within phagocytes, thecells that dispose of invading bacteria and cell debris. Lysosomalenzymes can digest almost all kinds of biological molecules. Theywork best in acidic conditions and so are called acid hydrolases.
The lysosomal membrane is adapted to serve lysosomalfunctions in two important ways. First, it contains H� (proton)“pumps,” which are ATPases that gather hydrogen ions from thesurrounding cytosol to maintain the organelle’s acidic pH. Sec-ond, it retains the dangerous acid hydrolases while permittingthe final products of digestion to escape so that they can be usedby the cell or excreted. In this way, lysosomes provide sites wheredigestion can proceed safely within a cell.
Lysosomes function as a cell’s “demolition crew” by
1. Digesting particles taken in by endocytosis, particularlyingested bacteria, viruses, and toxins
2. Degrading worn-out or nonfunctional organelles3. Performing metabolic functions, such as glycogen break-
down and release4. Breaking down nonuseful tissues, such as the webs be-
tween the fingers and toes of a developing fetus and theuterine lining during menstruation
5. Breaking down bone to release calcium ions into the blood
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 86
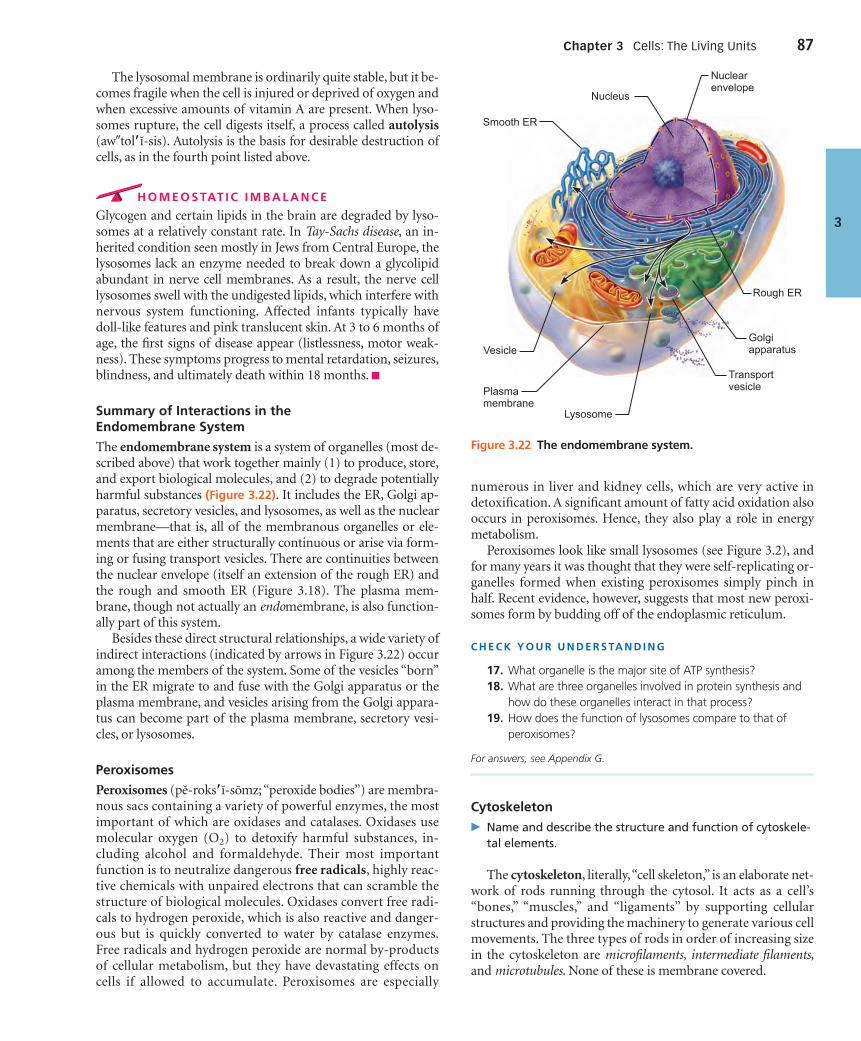
The lysosomal membrane is ordinarily quite stable, but it be-comes fragile when the cell is injured or deprived of oxygen andwhen excessive amounts of vitamin A are present. When lyso-somes rupture, the cell digests itself, a process called autolysis(aw�tol�ı-sis). Autolysis is the basis for desirable destruction ofcells, as in the fourth point listed above.
H O M E O S TAT I C I M B A L A N C E
Glycogen and certain lipids in the brain are degraded by lyso-somes at a relatively constant rate. In Tay-Sachs disease, an in-herited condition seen mostly in Jews from Central Europe, thelysosomes lack an enzyme needed to break down a glycolipidabundant in nerve cell membranes. As a result, the nerve celllysosomes swell with the undigested lipids, which interfere withnervous system functioning. Affected infants typically havedoll-like features and pink translucent skin. At 3 to 6 months ofage, the first signs of disease appear (listlessness, motor weak-ness). These symptoms progress to mental retardation, seizures,blindness, and ultimately death within 18 months. ■
Summary of Interactions in the Endomembrane System
The endomembrane system is a system of organelles (most de-scribed above) that work together mainly (1) to produce, store,and export biological molecules, and (2) to degrade potentiallyharmful substances (Figure 3.22). It includes the ER, Golgi ap-paratus, secretory vesicles, and lysosomes, as well as the nuclearmembrane—that is, all of the membranous organelles or ele-ments that are either structurally continuous or arise via form-ing or fusing transport vesicles. There are continuities betweenthe nuclear envelope (itself an extension of the rough ER) andthe rough and smooth ER (Figure 3.18). The plasma mem-brane, though not actually an endomembrane, is also function-ally part of this system.
Besides these direct structural relationships, a wide variety ofindirect interactions (indicated by arrows in Figure 3.22) occuramong the members of the system. Some of the vesicles “born”in the ER migrate to and fuse with the Golgi apparatus or theplasma membrane, and vesicles arising from the Golgi appara-tus can become part of the plasma membrane, secretory vesi-cles, or lysosomes.
Peroxisomes
Peroxisomes (pe-roks�ı-somz; “peroxide bodies”) are membra-nous sacs containing a variety of powerful enzymes, the mostimportant of which are oxidases and catalases. Oxidases usemolecular oxygen (O2) to detoxify harmful substances, in-cluding alcohol and formaldehyde. Their most importantfunction is to neutralize dangerous free radicals, highly reac-tive chemicals with unpaired electrons that can scramble thestructure of biological molecules. Oxidases convert free radi-cals to hydrogen peroxide, which is also reactive and danger-ous but is quickly converted to water by catalase enzymes.Free radicals and hydrogen peroxide are normal by-productsof cellular metabolism, but they have devastating effects oncells if allowed to accumulate. Peroxisomes are especially
numerous in liver and kidney cells, which are very active indetoxification. A significant amount of fatty acid oxidation alsooccurs in peroxisomes. Hence, they also play a role in energymetabolism.
Peroxisomes look like small lysosomes (see Figure 3.2), andfor many years it was thought that they were self-replicating or-ganelles formed when existing peroxisomes simply pinch inhalf. Recent evidence, however, suggests that most new peroxi-somes form by budding off of the endoplasmic reticulum.
C H E C K Y O U R U N D E R S TA N D I N G
17. What organelle is the major site of ATP synthesis?18. What are three organelles involved in protein synthesis and
how do these organelles interact in that process?19. How does the function of lysosomes compare to that of
peroxisomes?
For answers, see Appendix G.
Cytoskeleton
� Name and describe the structure and function of cytoskele-tal elements.
The cytoskeleton, literally,“cell skeleton,” is an elaborate net-work of rods running through the cytosol. It acts as a cell’s“bones,” “muscles,” and “ligaments” by supporting cellularstructures and providing the machinery to generate various cellmovements. The three types of rods in order of increasing sizein the cytoskeleton are microfilaments, intermediate filaments,and microtubules. None of these is membrane covered.
3
Figure 3.22 The endomembrane system.
Golgiapparatus
TransportvesiclePlasma
membrane
Vesicle
Smooth ER
Rough ER
Nuclearenvelope
Lysosome
Nucleus
Chapter 3 Cells: The Living Units 87
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 87

Microtubules (mi�kro-tu�bulz), the elements with thelargest diameter, are hollow tubes made of spherical proteinsubunits called tubulins (Figure 3.23c). Most microtubules radi-ate from a small region of cytoplasm near the nucleus called thecentrosome or cell center (see Figures 3.2, 3.25). Microtubules areremarkably dynamic organelles, constantly growing out fromthe centrosome, disassembling, and then reassembling at thesame or different sites. The stiff but bendable microtubules de-termine the overall shape of the cell, as well as the distributionof cellular organelles.
Mitochondria, lysosomes, and secretory vesicles attach tothe microtubules like ornaments hanging from tree branches.They are continually moved along the microtubules and repo-sitioned by tiny protein machines called motor proteins(kinesins, dyneins, and others) or motor molecules. The vari-ous motor proteins work by changing their shapes. Powered byATP, some motor proteins appear to act like train engines mov-ing substances along on the microtubular “railroad tracks.” Noone analogy completely captures their action, but the mostcommon analogy has these motor molecules moving “handover hand” somewhat like an orangutan—gripping, releasing,and then gripping again at a new site further along the micro-tubule (Figure 3.24a).
Microfilaments (mi�kro-fil�ah-ments), the thinnest elementsof the cytoskeleton, are strands of the protein actin (“ray”)(Figure 3.23a). Each cell has its own unique arrangement of mi-
crofilaments, so no two cells are alike. However, nearly all cellshave a fairly dense cross-linked network of microfilaments,called the terminal web, attached to the cytoplasmic side of theirplasma membrane. The web strengthens the cell surface and re-sists compression.
Most microfilaments are involved in cell motility or changesin cell shape. You could say that cells move “when they get theiract(in) together.” For example, actin filaments interact with an-other protein, unconventional myosin (mi�o-sin), to generatecontractile forces in a cell (Figure 3.24b). This interaction alsoforms the cleavage furrow that pinches one cell into two duringcell division. Microfilaments attached to CAMs (see Figure 3.4e)are responsible for the crawling movements of amoeboid mo-tion, and for the membrane changes that accompany endocyto-sis and exocytosis. Except in muscle cells, where they are highlydeveloped, stable, and long-lived, actin filaments are constantlybreaking down and re-forming from smaller subunits wheneverand wherever their services are needed.
Intermediate filaments are tough, insoluble protein fibersthat have a diameter between those of microfilaments and mi-crotubules (Figure 3.23b). Constructed like woven ropes, inter-mediate filaments are the most stable and permanent of thecytoskeletal elements and have high tensile strength. Unlike mi-crotubules and microfilaments, intermediate filaments do notbind ATP or serve as “tracks” for molecular motors to transportintracellular substances. Instead, they attach to desmosomes,
88 UNIT 1 Organization of the Body
3
(a) Microfilaments
Strands made of spherical proteinsubunits called actins
(b) Intermediate filaments
Tough, insoluble protein fibersconstructed like woven ropes
(c) Microtubules
Hollow tubes of spherical proteinsubunits called tubulins
Actin subunit
7 nm 10 nm 25 nm
Fibrous subunits
Tubulin subunits
Microfilaments form the blue networksurrounding the pink nucleus in this photo.
Intermediate filaments form the purplebatlike network in this photo.
Microtubules appear as gold networkssurrounding the cells’ pink nuclei in this photo.
Figure 3.23 Cytoskeletal elements support the cell and help to generate movement.Diagrammatic views (above) and photos (below). The photos are of fibroblasts treated tofluorescently tag the structure of interest.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 88

Chapter 3 Cells: The Living Units 89
3
Receptor formotor molecule
Microtubuleof cytoskeleton
Motor molecule(ATP powered)
Vesicle
Cytoskeletal elements(microtubules or microfilaments)
Motor molecule(ATP powered)
ATP
ATP
(a) Motor molecules can attach to receptors on vesicles or organelles, and “walk” the organelles along the microtubules of the cytoskeleton.
(b) In some types of cell motility, motor molecules attached to one element of the cytoskeleton can cause it to slide over another element, as in muscle contraction and cilia movement.
Figure 3.24 Microtubules and microfilaments function in cellmotility by interacting with motor molecules. The variousmotor molecules, all powered by ATP, work by changing their shapes,moving back or forth on something like microscopic legs. With eachcycle of shape changes, the motor molecule releases its free end and grips at a site farther along the microtubule or microfilament.
and their main job is to act as internal guy-wires to resist pullingforces exerted on the cell. Because the protein composition ofintermediate filaments varies in different cell types, there arenumerous names for these cytoskeletal elements—for example,they are called neurofilaments in nerve cells and keratin fila-ments in epithelial cells.
Centrosome and Centrioles
� Describe the roles of centrioles in mitosis and in formationof cilia and flagella.
As mentioned, microtubules are anchored at one end in an in-conspicuous region near the nucleus called the centrosome orcell center. The centrosome acts as a microtubule organizing center.It has few distinguishing marks other than a granular-looking
matrix that contains paired centrioles, small, barrel-shaped or-ganelles oriented at right angles to each other (Figure 3.25). Thecentrosome matrix is best known for generating microtubulesand organizing the mitotic spindle in cell division (seeFigure 3.33). Each centriole consists of a pinwheel array of ninetriplets of microtubules, each connected to the next by non-tubulin proteins and arranged to form a hollow tube. Centriolesalso form the bases of cilia and flagella, our next topics.
Centrosome matrix
(b)
(a)
Centrioles
Microtubules
Figure 3.25 Centrioles. (a) Three-dimensional view of a centriolepair oriented at right angles, as they are usually seen in the cell. Thecentrioles are located in an inconspicuous region to one side of thenucleus called the centrosome, or cell center. (b) An electronmicrograph showing a cross section of a centriole (120,000�). Noticethat it is composed of nine microtubule triplets.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 89

Cellular Extensions� Describe how the two main types of cell extensions, cilia
and microvilli, differ in structure and function.
Cilia and Flagella
Cilia (sil�e-ah; “eyelashes”) are whiplike, motile cellular exten-sions that occur, typically in large numbers, on the exposedsurfaces of certain cells (Figure 3.26). Ciliary action moves sub-stances in one direction across cell surfaces. For example, cili-ated cells that line the respiratory tract propel mucus laden withdust particles and bacteria upward away from the lungs.
When a cell is about to form cilia, the centrioles multiply andline up beneath the plasma membrane at the free cell surface. Mi-crotubules then begin to “sprout”from each centriole, forming theciliary projections by exerting pressure on the plasma membrane.
When the projections formed by centrioles are substantiallylonger, they are called flagella (flah-jel�ah). The single exampleof a flagellated cell in the human body is a sperm, which has onepropulsive flagellum, commonly called a tail. Notice that ciliapropel other substances across a cell’s surface, whereas a flagellumpropels the cell itself.
Centrioles forming the bases of cilia and flagella are com-monly referred to as basal bodies (ba�sal) (Figure 3.26). Theywere given a separate name because they were thought to be
different from the structures seen in the centrosome. We nowknow that centrioles and basal bodies are modifications of thesame structure. The “9 � 2” pattern of microtubules in thecilium or flagellum itself (nine doublets, or pairs, of microtubulesencircling one central pair) differs slightly from that of a centriole(nine microtubule triplets). Additionally, the cilium has flexible“wagon wheels”of cross-linking proteins (purple in Figure 3.26),and motor proteins (green dynein arms in Figure 3.26) thatpromote movement of the cilium or flagellum.
Just how ciliary activity is coordinated is not fully under-stood, but microtubules are definitely involved. Extending fromthe microtubule doublets are arms composed of the motor pro-tein dynein (Figure 3.26). The dynein side arms of one doubletgrip the adjacent doublet, and powered by ATP, push it up, re-lease, and then grip again. Because the doublets are physicallyrestricted by other proteins, they cannot slide far and instead areforced to bend. The collective bending action of all the doubletscauses the cilium to bend.
As a cilium moves, it alternates rhythmically between apropulsive power stroke, when it is nearly straight and moves inan arc, and a recovery stroke, when it bends and returns to its ini-tial position (Figure 3.27a). With these two strokes, the ciliumproduces a pushing motion in a single direction that is repeatedsome 10 to 20 times per second. The activity of cilia in a partic-ular region is coordinated so that the bending of one cilium is
90 UNIT 1 Organization of the Body
3
Plasmamembrane
Outer microtubuledoublet
Dynein arms
Centralmicrotubule
Radial spoke
Radial spoke
TEM
TEM
Triplet
Basal body(centriole)
Cilium
Microtubules
Plasmamembrane
Basal body
Cross-linkingproteins insideouter doublets
Cross-linkingproteins insideouter doublets
A longitudinal section of acilium shows microtubules running the length of thestructure.
The doublets also have attached motor proteins, the dynein arms.
The outer microtubule doublets and the two central microtubules are held together by cross-linking proteins and radial spokes.
A cross section through the basal body. The nine outer doublets of a cilium extend into a basal body where each doublet joins another microtubule to form a ring of nine triplets.
A cross section through the cilium shows the ”9 + 2” arrangement of microtubules.
TEM
Figure 3.26 Structure of a cilium. (TEM � transmission electron micrograph.)
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 90

quickly followed by the bending of the next and then the next,creating a current at the cell surface that brings to mind thetraveling waves that pass across a field of grass on a windy day(Figure 3.27b).
Microvilli
Microvilli (mi�kro-vil�i; “little shaggy hairs”) are minute, fin-gerlike extensions of the plasma membrane that project from afree, or exposed, cell surface (Figure 3.5 top and Figure 3.28).They increase the plasma membrane surface area tremendouslyand are most often found on the surface of absorptive cells suchas intestinal and kidney tubule cells. Microvilli have a core ofbundled actin filaments that extend into the so-called terminalweb of the cytoskeleton of the cell. Actin is sometimes a contrac-tile protein, but in microvilli it appears to function as a mechan-ical “stiffener.”
C H E C K Y O U R U N D E R S TA N D I N G
20. How are microtubules and microfilaments related func-tionally?
21. Of microfilaments, microtubules, or intermediate filaments,which is most important in maintaining cell shape?
22. The major function of cilia is to move substances across thefree cell surface. What is the major role of microvilli?
For answers, see Appendix G.
The Nucleus� Outline the structure and function of the nuclear envelope,
nucleolus, and chromatin.
Anything that works, works best when it is controlled. For cells,the control center is the gene-containing nucleus (nucle � pit,kernel). The nucleus can be compared to a computer, design de-partment, construction boss, and board of directors—all rolledinto one. As the genetic library, it contains the instructionsneeded to build nearly all the body’s proteins. Additionally, itdictates the kinds and amounts of proteins to be synthesized atany one time in response to signals acting on the cell.
Most cells have only one nucleus, but some, including skele-tal muscle cells, bone destruction cells, and some liver cells, aremultinucleate (mul�tı-nu�kle-at), that is, they have many nu-clei. The presence of more than one nucleus usually signifiesthat a larger-than-usual cytoplasmic mass must be regulated.
With one exception, all of our body cells are nucleated. Theexception is mature red blood cells, whose nuclei are ejected be-fore the cells enter the bloodstream. These anucleate (a-nu�kle-at;a � without) cells cannot reproduce and therefore live in thebloodstream for only three to four months before they begin todeteriorate. Without a nucleus, a cell cannot produce mRNA tomake proteins, and when its enzymes and cell structures startto break down (as all eventually do), they cannot be replaced.
The nucleus, averaging 5 µm in diameter, is larger than any ofthe cytoplasmic organelles. Although most often spherical oroval, its shape usually conforms to the shape of the cell. Thenucleus has three recognizable regions or structures: the nuclearenvelope (membrane), nucleoli, and chromatin (Figure 3.29a).Besides these structures, there are several distinct compart-ments (a splicing factor compartment and others) rich in spe-cific sets of proteins. These are not limited by membranes and
Chapter 3 Cells: The Living Units 91
3
(a) Phases of ciliary motion.
(b)
Power, orpropulsive, stroke
Layer of mucus
Cell surface
Recovery stroke, when ciliumis returning to its initial position
1 2 3 4 5 6 7
Traveling wave created by the activity ofmany cilia acting together propels mucusacross cell surfaces.
Figure 3.27 Ciliary function.
Microvillus
Actinfilaments
Terminalweb
Figure 3.28 Microvilli.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 91

are in a constant state of dynamic flux. Much remains to belearned about them.
The Nuclear EnvelopeThe nucleus is bounded by the nuclear envelope, a doublemembrane barrier separated by a fluid-filled space (similar tothe mitochondrial membrane). The outer nuclear membrane iscontinuous with the rough ER of the cytoplasm and is studdedwith ribosomes on its external face. The inner nuclear mem-brane is lined by the nuclear lamina, a network of lamins (rod-
shaped proteins that assemble to form intermediate filaments)that maintains the shape of the nucleus and acts as a scaffold toorganize DNA in the nucleus (Figure 3.29b, bottom).
At various points, the two layers of the nuclear envelope inter-connect to form the edges of nuclear pores. An intricate complexof proteins, called a nuclear pore complex (NPC), lines each pore,forming an aqueous transport channel and regulating entry andexit of molecules (e.g., mRNAs) and large particles into and outof the nucleus (see Figure 3.29b, middle).
Like other cell membranes, the nuclear envelope is selectivelypermeable, but here passage of substances is much freer than
92 UNIT 1 Organization of the Body
3
Chromatin (condensed)
Nuclear envelope Nucleus
Nuclearpores
Fractureline of outermembrane
Nuclear pore complexes. Each pore is ringed by protein particles.
Surface of nuclear envelope.
Nuclear lamina. The netlike laminacomposed of intermediate filaments formed bylamins lines the inner surface of the nuclearenvelope.
Nucleolus
Cisternae of rough ER
(a)
(b)
Figure 3.29 The nucleus. (a) Three-dimensional diagrammaticview of the nucleus, showing the continuity of its double membranewith the ER. (b) Freeze-fracture transmission electron micrographs(TEMs).
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 92
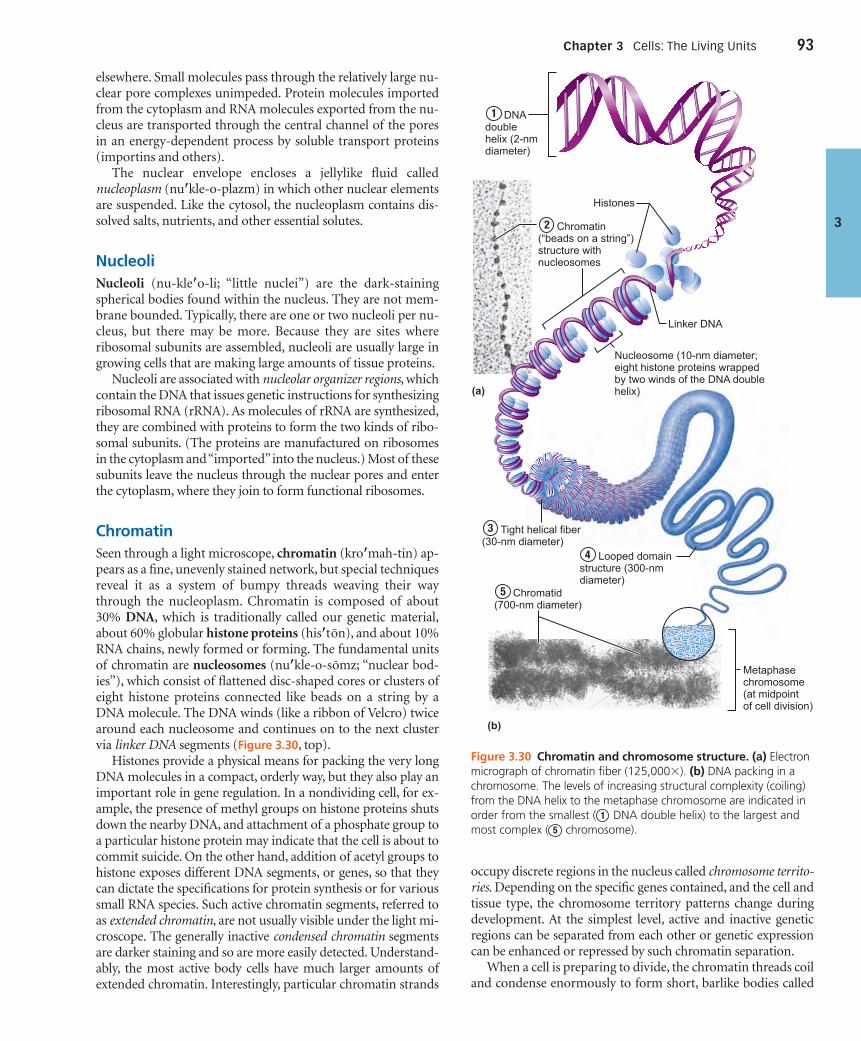
Chapter 3 Cells: The Living Units 93
3
elsewhere. Small molecules pass through the relatively large nu-clear pore complexes unimpeded. Protein molecules importedfrom the cytoplasm and RNA molecules exported from the nu-cleus are transported through the central channel of the poresin an energy-dependent process by soluble transport proteins(importins and others).
The nuclear envelope encloses a jellylike fluid callednucleoplasm (nu�kle-o-plazm) in which other nuclear elementsare suspended. Like the cytosol, the nucleoplasm contains dis-solved salts, nutrients, and other essential solutes.
NucleoliNucleoli (nu-kle�o-li; “little nuclei”) are the dark-stainingspherical bodies found within the nucleus. They are not mem-brane bounded. Typically, there are one or two nucleoli per nu-cleus, but there may be more. Because they are sites whereribosomal subunits are assembled, nucleoli are usually large ingrowing cells that are making large amounts of tissue proteins.
Nucleoli are associated with nucleolar organizer regions, whichcontain the DNA that issues genetic instructions for synthesizingribosomal RNA (rRNA). As molecules of rRNA are synthesized,they are combined with proteins to form the two kinds of ribo-somal subunits. (The proteins are manufactured on ribosomesin the cytoplasm and “imported”into the nucleus.) Most of thesesubunits leave the nucleus through the nuclear pores and enterthe cytoplasm, where they join to form functional ribosomes.
ChromatinSeen through a light microscope, chromatin (kro�mah-tin) ap-pears as a fine, unevenly stained network, but special techniquesreveal it as a system of bumpy threads weaving their waythrough the nucleoplasm. Chromatin is composed of about30% DNA, which is traditionally called our genetic material,about 60% globular histone proteins (his�ton), and about 10%RNA chains, newly formed or forming. The fundamental unitsof chromatin are nucleosomes (nu�kle-o-somz; “nuclear bod-ies”), which consist of flattened disc-shaped cores or clusters ofeight histone proteins connected like beads on a string by aDNA molecule. The DNA winds (like a ribbon of Velcro) twicearound each nucleosome and continues on to the next clustervia linker DNA segments (Figure 3.30, top).
Histones provide a physical means for packing the very longDNA molecules in a compact, orderly way, but they also play animportant role in gene regulation. In a nondividing cell, for ex-ample, the presence of methyl groups on histone proteins shutsdown the nearby DNA, and attachment of a phosphate group toa particular histone protein may indicate that the cell is about tocommit suicide. On the other hand, addition of acetyl groups tohistone exposes different DNA segments, or genes, so that theycan dictate the specifications for protein synthesis or for varioussmall RNA species. Such active chromatin segments, referred toas extended chromatin, are not usually visible under the light mi-croscope. The generally inactive condensed chromatin segmentsare darker staining and so are more easily detected. Understand-ably, the most active body cells have much larger amounts ofextended chromatin. Interestingly, particular chromatin strands
Metaphasechromosome(at midpointof cell division)
Nucleosome (10-nm diameter; eight histone proteins wrappedby two winds of the DNA double helix)
Linker DNA
Histones
(a)
(b)
1 DNA doublehelix (2-nm diameter)
2 Chromatin (“beads on a string”) structure with nucleosomes
3 Tight helical fiber(30-nm diameter)
5 Chromatid(700-nm diameter)
4 Looped domain structure (300-nm diameter)
Figure 3.30 Chromatin and chromosome structure. (a) Electronmicrograph of chromatin fiber (125,000�). (b) DNA packing in achromosome. The levels of increasing structural complexity (coiling)from the DNA helix to the metaphase chromosome are indicated inorder from the smallest ( DNA double helix) to the largest andmost complex ( chromosome).5
1
occupy discrete regions in the nucleus called chromosome territo-ries. Depending on the specific genes contained, and the cell andtissue type, the chromosome territory patterns change duringdevelopment. At the simplest level, active and inactive geneticregions can be separated from each other or genetic expressioncan be enhanced or repressed by such chromatin separation.
When a cell is preparing to divide, the chromatin threads coiland condense enormously to form short, barlike bodies called
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 93

94 UNIT 1 Organization of the Body
3
Parts of the Cell: Structure and Function
CELL PART STRUCTURE FUNCTIONS
Plasma Membrane (Figure 3.3)
Membrane made of a double layer of lip-ids (phospholipids, cholesterol, and so on)within which proteins are embedded. Pro-teins may extend entirely through the lipidbilayer or protrude on only one face. Exter-nally facing proteins and some lipids haveattached sugar groups.
Serves as an external cell barrier, and acts intransport of substances into or out of the cell. Maintains a resting potential that is es-sential for functioning of excitable cells.Externally facing proteins act as recep-tors (for hormones, neurotransmitters, andso on) and in cell-to-cell recognition.
Cytoplasm
Cellular region between the nuclear and plasma membranes. Consists of fluid cytosol contain-ing dissolved solutes, organelles (the metabolic machinery of the cytoplasm), and inclusions(stored nutrients, secretory products, pigment granules).
Cytoplasmic Organelles
■ Mitochondria (Figure 3.17) Rodlike, double-membrane structures; innermembrane folded into projections calledcristae.
Site of ATP synthesis; powerhouse of the cell.
■ Ribosomes (Figures 3.18, 3.37–3.39) Dense particles consisting of two subunits,each composed of ribosomal RNA and pro-tein. Free or attached to rough endoplasmicreticulum.
The sites of protein synthesis.
■ Rough endoplasmic reticulum (Figures 3.18, 3.39)
Membrane system enclosing a cavity, the cis-terna, and coiling through the cytoplasm.Externally studded with ribosomes.
Sugar groups are attached to proteins withinthe cisternae. Proteins are bound in vesiclesfor transport to the Golgi apparatus andother sites. External face synthesizes phos-pholipids.
■ Smooth endoplasmic reticulum (Figure 3.18)
Membranous system of sacs and tubules; freeof ribosomes.
Site of lipid and steroid (cholesterol) synthe-sis, lipid metabolism, and drug detoxifi-cation.
■ Golgi apparatus (Figures 3.19, 3.20) A stack of smooth membrane sacs and asso-ciated vesicles close to the nucleus.
Packages, modifies, and segregates pro-teins for secretion from the cell, inclusion inlysosomes, and incorporation into the plasmamembrane.
■ Lysosomes (Figure 3.21) Membranous sacs containing acid hy-drolases.
Sites of intracellular digestion.
■ Peroxisomes (Figure 3.2) Membranous sacs of oxidase enzymes. The enzymes detoxify a number of toxic sub-stances. The most important enzyme, cata-lase, breaks down hydrogen peroxide.
■ Microtubules (Figures 3.23–3.25) Cylindrical structures made of tubulinproteins.
Support the cell and give it shape. Involved inintracellular and cellular movements. Formcentrioles and cilia and flagella, if present.
■ Microfilaments (Figures 3.23, 3.24) Fine filaments composed of the pro-tein actin.
Involved in muscle contraction and othertypes of intracellular movement, help formthe cell’s cytoskeleton.
■ Intermediate filaments (Figure 3.23) Protein fibers; composition varies. The stable cytoskeletal elements; resist me-chanical forces acting on the cell.
■ Centrioles (Figure 3.25) Paired cylindrical bodies, each composed ofnine triplets of microtubules.
Organize a microtubule network during mi-tosis to form the spindle and asters. Form thebases of cilia and flagella.
Inclusions Varied; includes stored nutrients such as lipiddroplets and glycogen granules, protein crys-tals, pigment granules.
Storage for nutrients, wastes, and cell products.
TABLE 3.3
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 94

Chapter 3 Cells: The Living Units 95
3
(continued)
CELL PART STRUCTURE FUNCTIONS
Cellular Extensions
■ Cilia (Figure 3.26, 3.27) Short cell-surface projections; each ciliumcomposed of nine pairs of microtubules sur-rounding a central pair.
Coordinated movement creates a unidi-rectional current that propels substancesacross cell surfaces.
■ Flagella Like a cilium, but longer; only example in hu-mans is the sperm tail.
Propel the cell.
■ Microvilli (Figure 3.28) Tubular extensions of the plasma membrane;contain a bundle of actin filaments.
Increase surface area for absorption.
Nucleus (Figure 3.2, 3.29)
Largest organelle. Surrounded by the nuclearenvelope; contains fluid nucleoplasm, nucle-oli, and chromatin.
Control center of the cell; responsible fortransmitting genetic information and provid-ing the instructions for protein synthesis.
■ Nuclear envelope (Figure 3.29) Double-membrane structure pierced bypores. Outer membrane continuous with theendoplasmic reticulum.
Separates the nucleoplasm from the cyto-plasm and regulates passage of substances toand from the nucleus.
■ Nucleoli (Figure 3.29) Dense spherical (non-membrane-bounded)bodies, composed of ribosomal RNA andproteins.
Site of ribosome subunit manufacture.
■ Chromatin (Figure 3.30) Granular, threadlike material composed ofDNA and histone proteins.
DNA constitutes the genes.
TABLE 3.3
chromosomes (“colored bodies”) (Figure 3.30, bottom). Chro-mosome compactness prevents the delicate chromatin strandsfrom tangling and breaking during the movements that occurduring cell division. We describe the functions of DNA and theevents of cell division in the next section.
Table 3.3 summarizes the parts of the cell.
C H E C K Y O U R U N D E R S TA N D I N G
23. If a cell ejects or loses its nucleus, what is its fate and why?24. What is the role of nucleoli?25. What is the importance of the histone proteins present in the
nucleus?
For answers, see Appendix G.
Cell Growth and ReproductionThe Cell Life Cycle� List the phases of the cell life cycle and describe the key
events of each phase.
� Describe the process of DNA replication.
The cell life cycle is the series of changes a cell goes throughfrom the time it is formed until it reproduces. The outer ring ofFigure 3.31 shows the two major periods of the cell cycle:
interphase (in green), in which the cell grows and carries on itsusual activities, and cell division or the mitotic phase (in yellow),during which it divides into two cells.
Interphase
Interphase is the period from cell formation to cell division.Early cytologists, unaware of the constant molecular activity incells and impressed by the obvious movements of cell division,called interphase the resting phase of the cell cycle. (The terminterphase reflects this idea of a stage between cell divisions.)However, this image is grossly misleading because during inter-phase a cell is carrying out all its routine activities and is “resting”only from dividing. Perhaps a more accurate name for thisphase would be metabolic phase or growth phase.
Subphases In addition to carrying on its life-sustaining reac-tions, an interphase cell prepares for the next cell division.Interphase is divided into G1, S, and G2 subphases (the Gs standfor gaps before and after the S phase, and S is for synthetic).In all three subphases, the cell grows by producing proteinsand organelles, but chromatin is reproduced only during theS subphase.
During G1 (gap 1), the cell is metabolically active, synthesiz-ing proteins rapidly and growing vigorously (Figure 3.31, lightgreen area). This is the most variable phase in terms of length.In cells that divide rapidly, G1 typically lasts several minutes tohours, but in those that divide slowly, it may last for days or even
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 95

96 UNIT 1 Organization of the Body
3
years. Cells that permanently cease dividing are said to be in theG0 phase. For most of G1, virtually no activities directly relatedto cell division occur. However, as G1 ends, the centrioles start toreplicate in preparation for cell division.
During the S phase, DNA is replicated, ensuring that the twofuture cells being created will receive identical copies of the ge-netic material (Figure 3.31, blue area). New histones are madeand assembled into chromatin. One thing is sure: Without aproper S phase, there can be no correct mitotic phase. (We willdescribe DNA replication next.)
The final phase of interphase, called G2, is brief (Figure 3.31,dark green area). Enzymes and other proteins needed for divi-sion are synthesized and moved to their proper sites. By the endof G2, centriole replication (begun in G1) is complete. The cell isnow ready to divide. Throughout S and G2, the cell continues togrow and carries on with business as usual.
DNA Replication Before a cell can divide, its DNA must bereplicated exactly, so that identical copies of the cell’s genes canbe passed on to each of its offspring. During the S phase, repli-cation begins simultaneously on several chromatin threads andcontinues until all the DNA has been replicated.
Replication is still being studied but appears to involve thefollowing events:
1. The DNA helices begin unwinding from the histones ofthe nucleosomes.
2. In an ATP-requiring process, a helicase enzyme untwiststhe double helix and gradually separates the DNA mole-cule into two complementary nucleotide chains, exposingthe nitrogenous bases. The Y-shaped site of separation,where active DNA replication will soon begin, is called thereplication fork (Figure 3.32).
3. Each nucleotide strand then serves as a template, or set ofinstructions, for building a new complementary nu-cleotide strand from free DNA precursors dissolved in thenucleoplasm.
4. At sites where DNA synthesis is to occur, the needed “ma-chinery” gradually accumulates until several different pro-teins (mostly enzymes) are present in a large complexcalled a replisome. Primase enzymes, which are part of thereplisome, catalyze the formation of short (about ten baseslong) RNA primers, which actually initiate DNA synthesis.
5. Once the primer is in place, the enzyme DNA polymerasecomes into the picture. Continuing from the primer, itpositions complementary nucleotides along the templatestrand and then covalently links them together. So, if theDNA polymerase encounters the sequence of bases GCTon the template strand, it assembles the bases CGA to bindto it. DNA polymerase works only in one direction. Con-sequently, one strand, the leading strand, is synthesizedcontinuously (once primed by the RNA primer) followingthe movement of the replication fork. The other strand,called the lagging strand, is constructed in segments in theopposite direction and requires that a primer initiate repli-cation of each segment. These primers are eventually re-placed with DNA nucleotides by DNA polymerases.
6. The short segments of DNA are then spliced together byDNA ligase. The end result is that two DNA molecules areformed from the original DNA helix and are identical to it.Because each new molecule consists of one old and one newnucleotide strand, this mechanism of DNA replication iscalled semiconservative replication (Figure 3.32). Replica-tion also involves the generation of two new telomeres (tel �end; mer � piece), snugly fitting nucleoprotein caps thatprevent degradation of the ends of the chromatin strands.
As soon as replication ends, histones (synthesized in the cyto-plasm and imported into the nucleus) associate with the DNA,completing the formation of two new chromatin strands. Thechromatin strands, united by a buttonlike centromere (a stretchof repetitive DNA), remain attached, held together by thecentromere and a protein complex called cohesin, until the cellenters the anaphase stage of mitotic cell division (see p. 99).They are then distributed to the daughter cells as described next,ensuring that each cell has identical genetic information.
The progression from DNA synthesis into the events of celldivision presumes that the newly synthesized DNA is not dam-aged or broken in any way. If damage occurs, progress through
Interphase
G1
Growth
SGrowth and DNA
synthesis G2
Growth and finalpreparations for
division
AnaphaseMetaphase
Prophase
TelophaseCytokinesis
M
Mitotic phase (M)
G2 checkpoint
G1 checkpoint(restriction point)
Mitosis
Figure 3.31 The cell cycle. During G1, cells grow rapidly and carryout their routine functions. The S phase is the period of DNA syn-thesis. In G2, materials needed for cell division are synthesized andgrowth continues. During the M phase (cell division), mitosis andcytokinesis occur, producing two daughter cells. Important check-points at which mitosis may be prevented from occurring are foundthroughout interphase; two are shown on the diagram.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 96

the cell cycle is arrested until the DNA repair mechanism hasfixed the problem.
Cell Division
Cell division is essential for body growth and tissue repair. Cellsthat continually wear away, such as cells of the skin and intes-tinal lining, reproduce themselves almost continuously. Oth-ers, such as liver cells, divide more slowly (to maintain theparticular size of the organ they compose) but retain the abilityto reproduce quickly if the organ is damaged. Most cells ofnervous tissue, skeletal muscle, and heart muscle lose their abil-ity to divide when they are fully mature, and repairs are madewith scar tissue (a fibrous type of connective tissue).
Events of Cell Division In most body cells, cell division, whichis called the M (mitotic) phase of the cell life cycle, involves twodistinct events: mitosis (mi-to�sis; mit � thread; osis �process), or division of the nucleus, and cytokinesis (si-to-kı-ne�sis; kines � movement), or division of the cytoplasm (seeFigure 3.31, yellow area). A somewhat different process of nu-clear division called meiosis (mi-o�sis) produces sex cells (ova
and sperm) with only half the number of genes found in otherbody cells. We discuss the details of meiosis in Chapter 27. Herewe concentrate on mitotic cell division.
Mitosis Mitosis is the series of events that parcel out the repli-cated DNA of the mother cell to two daughter cells. It is describedin terms of four phases: prophase, metaphase, anaphase, andtelophase, but it is actually a continuous process, with one phasemerging smoothly into the next. Its duration varies accordingto cell type, but in human cells it typically lasts about an houror less from start to finish. Focus on Mitosis (Figure 3.33),pp. 98–99, describes the phases of mitosis in detail.
Cytokinesis Cytokinesis, or the division of the cytoplasm, be-gins during late anaphase and is completed after mitosis ends.The plasma membrane over the center of the cell (the spindleequator) is drawn inward to form a cleavage furrow by the ac-tivity of a contractile ring (Figure 3.33) made of actin filaments.The furrow deepens until the cytoplasmic mass is pinched intotwo parts, so that at the end of cytokinesis there are two daughtercells. Each is smaller and has less cytoplasm than the mother
Chapter 3 Cells: The Living Units 97
3
Adenine
Thymine
Cytosine
GuanineOld (template) strand
Two new strands (leading and lagging)synthesized in opposite directions
DNA polymerase
DNA polymerase
Laggingstrand
Leadingstrand
Free nucleotides Old strand acts as a template forsynthesis of new strandChromosome
Helicase unwindsthe double helix andexposes the bases
Old DNA
Replicationfork
Figure 3.32 Replication of DNA. The DNA helix uncoils, and the hydrogen bonds betweenits base pairs are broken. Then, each nucleotide strand of the DNA acts as a template for con-structing a complementary strand, as illustrated on the right-hand side of the diagram. DNApolymerases work in one direction only, so the two new strands (leading and lagging) aresynthesized in opposite directions. (The DNA ligase enzymes that join the DNA fragments onthe lagging strand are not illustrated.) Each DNA molecule formed consists of one old (tem-plate) strand and one newly assembled strand and constitutes a chromatid of a chromosome.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 97

The light micrographs show dividing lung cells from a newt. The chromosomes appear blue and the microtubules green. (The red fibers are intermediate filaments.) The schematic drawings show details not visible in the micrographs. For simplicity, only four chromosomes are drawn.
Centrosomes (eachhas 2 centrioles)
Early mitoticspindle
Spindle pole
Kinetochore Kinetochoremicrotubule
Polar microtubule
Nucleolus Centromere
Plasmamembrane Fragments
of nuclearenvelopeAster
Nuclearenvelope
Chromosomeconsisting of twosister chromatids
Chromatin
InterphaseInterphase is the period of a cell’s life when it carries out its normal metabolic activities and grows.
• During interphase, the DNA-containing material is in the form of chromatin. The nuclear envelope and one or more nucleoli are intact and visible.• There are three distinct periods of this phase: G1: The centrioles begin replicating. S: DNA is replicated. G2: Final preparations for mitosis are completed and centrioles finish replicating.
Early Prophase• The chromatin condenses, forming barlike chromosomes that are visible with a light microscope. • Each duplicated chromosome appears as two identical threads, now called sister chromatids, held together at a small, constricted region called a centromere. (After the chromatids separate, each is considered a new chromo-some.) • As the chromosomes appear, the nucleoli disappear, and the two centrosomes separate from one another. • The centrosomes act as focal points for growth of a microtubule assembly called the mitotic spindle. As these microtubules lengthen, they propel the centrosomes toward opposite ends (poles) of the cell.• Microtubule arrays called asters (”stars”) are seen extending from the matrix around the centrosomes.
Late Prophase• While the centrosomes are still moving apart, the nuclear envelope fragments, allowing the spindle to interact with the chromosomes.• Some of the growing spindle microtubules attach to kinetochores (ki-ne -to-korz), special protein structures at each chromosome’s centromere. Such microtubules are called kinetochore microtubules.• The remaining spindle microtubules (not attached to any chromosomes) are called polar microtubules. The microtubules slide past each other, forcing the poles apart.• The kinetochore microtubules pull on each chromosome from both poles in a kind of tug-of-war that ultimately draws the chromosomes to the exact center or equator of the cell.
Interphase Early Prophase Late Prophase
Mitosis is the process of nuclear division in which the chromosomes are distributed to two daughter nuclei. Together with cytokinesis, it produces two identical daughter cells.
Figure 3.33 FOCUS Mitosis
98
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 98

Contractilering atcleavagefurrow
Nuclearenvelopeforming
Nucleolus formingSpindle
Metaphaseplate
Daughterchromosomes
Metaphase Anaphase Telophase and Cytokinesis
Telophase Telophase begins as soon as chromosomal movement stops. This final phase is like prophase in reverse.
• The identical sets of chromosomes at the opposite poles of the cell uncoil and resume their threadlike chromatin form.• A new nuclear envelope forms around each chromatin mass, nucleoli reappear within the nuclei, and the spindle breaks down and disappears. • Mitosis is now ended. The cell, for just a brief period, is binucleate (has two nuclei) and each new nucleus is identical to the original mother nucleus.
Cytokinesis• As a rule, as mitosis draws to a close, cytokinesis completes the division of the cell into two identical daughter cells. Cytokinesis occurs as a contractile ring of actin microfila-ments forms the cleavage furrow and pinches the cell apart. It begins during late anaphase and continues through and beyond telophase.
AnaphaseAnaphase is the third and shortest phase of mitosis. Anaphase begins abruptly as the centromeres of the chromosomes split simultaneously. Each chromatid now becomes a chromosome in its own right.
• The kinetochore microtubules, moved along by motor proteins in the kinetochores, gradually pull each chromosome toward the pole it faces. • At the same time, the polar microtubules slide past each other, lengthen, and push the two poles of the cell apart. • Anaphase is easy to recognize because the moving chromosomes look V shaped. The centromeres lead the way, and the chromosomal “arms” dangle behind them.• This process of moving and separating the chromosomes is helped by the fact that the chromosomes are short, compact bodies. Diffuse threads of chromatin would tangle, trail, and break, resulting in imprecise “parceling out” to the daughter cells.
MetaphaseMetaphase is the second phase of mitosis.
• The two centrosomes are at opposite poles of the cell.• The chromosomes cluster at the middle of the cell, with their centromeres precisely aligned at the equator of the spindle. This imaginary plane midway between the poles is called the metaphase plate.
99
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 99

cell, but is genetically identical to it. The daughter cells then en-ter the interphase portion of the life cycle until it is their turn todivide.
Control of Cell Division The signals that prod cells to divideare incompletely understood, but we know that the ratio of cellsurface area to cell volume is important. The amount of nutri-ents a growing cell requires is directly related to its volume. Vol-ume increases with the cube of cell radius, whereas surface areaincreases more slowly with the square of the radius. For exam-ple, a 64-fold (43) increase in cell volume is accompanied byonly a 16-fold (42) increase in surface area. Consequently, thesurface area of the plasma membrane becomes inadequate fornutrient and waste exchange when a cell reaches a certain criti-cal size. Cell division solves this problem because the smallerdaughter cells have a favorable ratio of surface area to volume.These surface-volume relationships help explain why most cellsare microscopic in size.
Two other factors that influence when cells divide are chem-ical signals (growth factors, hormones, and others) released byother cells and the availability of space. Normal cells stop prolif-erating when they begin touching, a phenomenon called contactinhibition. The cell life cycle is controlled by a system that hasbeen compared to an automatic washer’s timer control. Likethat timer, the control system for the cell cycle is driven by abuilt-in clock. However, just as the washer’s cycle is subject toadjustments (by regulating the flow from the faucet, say, or byan internal water-level sensor), the cell cycle is regulated by bothinternal and external factors.
Two groups of proteins are crucial to the ability of a cell toaccomplish the S phase and enter mitosis. They are cyclins (reg-ulatory proteins whose levels rise and fall during each life cycle)and Cdks (cyclin-dependent kinases), which are present in aconstant concentration in the cell and are activated by bindingto particular cyclins. In response to specific signals, a new batchof cyclins accumulates during each interphase. Subsequent join-ing of specific Cdk and cyclin proteins initiates enzymatic cas-cades that phosphorylate histones and other proteins neededfor the various stages of cell division. At the end of mitosis, thecyclins are abruptly destroyed by enzymes.
A number of “switches” and crucial checkpoints for cell di-vision occur throughout interphase. These built-in stop sig-nals halt the cell cycle until overridden by internal or externalgo-ahead signals. In many cells, a G1 checkpoint, called therestriction point, seems to be most important (see Figure 3.31).If the cell is diverted from dividing at this checkpoint, it entersthe nondividing state (G0). Another important checkpoint,and the first to be understood, occurs late in G2, when athreshold amount of a protein complex called MPF (M-phasepromoting factor) is required to give the okay signal to passthe G2 checkpoint and enter the M phase. Later in M phase,MPF is inactivated.
Besides these “go” signals, there are a number of so-calledrepressor genes that inhibit cell division. One example is thep53 gene that initiates a series of enzymatic events that producegrowth-inhibiting factors. Roughly half of all cancers have ab-normal p53 genes. These cancers are not inhibited by contact
with other cells and divide wildly, making them dangerous totheir host.
C H E C K Y O U R U N D E R S TA N D I N G
26. If one of the DNA strands being replicated “reads” CGAATG,what will be the base sequence of the corresponding DNAstrand?
27. During what phase of the cell cycle is DNA synthesized?28. What are three events occurring in prophase that are undone
in telophase?
For answers, see Appendix G.
Protein Synthesis� Define gene and genetic code and explain the function
of genes.
� Name the two phases of protein synthesis and describe theroles of DNA, mRNA, tRNA, and rRNA in each phase.
� Contrast triplets, codons, and anticodons.
In addition to directing its own replication, DNA serves as themaster blueprint for protein synthesis. Although cells also makelipids and carbohydrates, DNA does not dictate their structure.Historically, DNA is said to specify only the structure of proteinmolecules, including the enzymes that catalyze the synthesis ofall classes of biological molecules. Most of the metabolic ma-chinery of the cell is concerned in some way with protein syn-thesis. This is not surprising, seeing as structural proteinsconstitute most of the dry cell material, and functional proteinsdirect almost all cellular activities. Essentially, cells are miniatureprotein factories that synthesize the huge variety of proteinsthat determine the chemical and physical nature of cells—andtherefore of the whole body.
Proteins, as you will recall from Chapter 2, are composed ofpolypeptide chains, which in turn are made up of amino acids.For purposes of this discussion, we can define a gene as a seg-ment of a DNA molecule that carries instructions for creatingone polypeptide chain. (Note, however, that some genes specifythe structure of certain varieties of RNA as their final product.)
The four nucleotide bases (A, G, T, and C) are the “letters”used in the genetic dictionary, and the information of DNA isfound in the sequence of these bases. Each sequence of threebases, called a triplet, can be thought of as a “word” that speci-fies a particular amino acid. For example, the triplet AAA callsfor the amino acid phenylalanine, and CCT calls for glycine.The sequence of triplets in each gene forms a “sentence” thattells exactly how a particular polypeptide is to be made: It spec-ifies the number, kinds, and order of amino acids needed tobuild a particular polypetide.
Variations in the arrangement of A, T, C, and G allow ourcells to make all the different kinds of proteins needed. Even anunusually “small” gene has an estimated 210 base pairs in se-quence. The ratio between DNA bases in the gene and aminoacids in the polypeptide is 3:1 (because each triplet stands for
100 UNIT 1 Organization of the Body
3
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 100

one amino acid), so we would expect the polypeptide specifiedby such a gene to contain 70 amino acids.
Most genes of higher organisms contain exons, which areamino acid–specifying informational sequences. These exonsare often separated by introns, which are noncoding, oftenrepetitive, segments that range from 60 to 100,000 nucleotides.Once considered a type of “junk DNA,” intron DNA is now be-lieved to represent a genome scrapyard that provides a reservoirof ready-to-use DNA segments for genome evolution, as well asa rich source of a large variety of small RNA molecules. Therest of the DNA (the great majority of it) is essentially “darkmatter” whose function is a mystery. It is in these regionsthat pseudogenes (false genes) are found. Pseudogenes “looklike” real genes but have deficits that render them apparentlyfunctionless.
The Role of RNA
By itself, DNA is like a CD recording: The information it con-tains cannot be used without a decoding mechanism (a CDplayer). Furthermore, most polypeptides are manufactured atribosomes in the cytoplasm, but in interphase cells, DNA neverleaves the nucleus. So, DNA requires not only a decoder, but amessenger as well. The decoding and messenger functions arecarried out by RNA, the second type of nucleic acid.
As you learned in Chapter 2, RNA differs from DNA in beingsingle stranded and in having the sugar ribose instead of de-oxyribose and the base uracil (U) in place of thymine (T). Threeforms of RNA typically act together to carry out DNA’s instruc-tions for polypeptide synthesis:
1. Messenger RNA (mRNA), relatively long nucleotidestrands resembling “half-DNA” molecules (one of the twostrands of a DNA molecule coding for protein structure)
2. Ribosomal RNA (rRNA), part of the ribosomes3. Transfer RNA (tRNA), small, roughly L-shaped molecules
All types of RNA are formed on the DNA in the nucleus inmuch the same way as DNA replicates itself: The DNA helix sep-arates and one of its strands serves as a template for synthesiz-ing a complementary RNA strand. Once formed, the RNAmolecule is released from the DNA template and migrates intothe cytoplasm. Its job done, the DNA simply recoils into its hel-ical, inactive form.
Approximately 2% of the nuclear DNA codes for the synthe-sis of short-lived mRNA, so named because it carries, from geneto ribosome, the “message” containing the instructions forbuilding a polypeptide. (Kind of a genetic memo for proteinstructure.) DNA in the nucleolar organizer regions (mentionedpreviously) codes for the synthesis of rRNA, which is long-livedand stable, as is tRNA coded by other DNA sequences. BecauserRNA and tRNA do not transport codes for synthesizing othermolecules, they are the final products of the genes that code forthem. Ribosomal RNA and tRNA act together to “translate” themessage carried by mRNA.
Essentially, polypeptide synthesis involves two majorsteps: (1) transcription, in which DNA’s information is encoded
in mRNA, and (2) translation, in which the informationcarried by mRNA is decoded and used to assemblepolypeptides. Figure 3.34 shows an overview of the infor-mation flow in these two major steps. The figure also indi-cates the “RNA processing” that removes introns frommRNA before this molecule leaves the nucleus and movesinto the cytoplasm.
Transcription
A transcriptionist converts a message from a recording or short-hand notes into a word-processed or electronic copy. In otherwords, information is transformed, or transferred, from oneform or format to another. In cells, transcription involves thetransfer of information from a DNA’s base sequence to thecomplementary base sequence of an mRNA molecule. Theform is different, but the same information is being conveyed.Once the mRNA molecule is made, it detaches and leaves thenucleus via a nuclear pore.
Essentially, three basic phases are involved in transcription:(1) initiation, (2) elongation, and (3) termination. In additionto looking at the synthesis of mRNA, we will also examine theediting and further processing needed to clean up the mRNAtranscript.
How does transcription get started? Transcription cannotbegin until gene-activating chemicals called transcription factorsstimulate loosening of the histones at the site-to-be of gene
Chapter 3 Cells: The Living Units 101
3
Nuclearpores
mRNA
Pre-mRNARNA Processing
Transcription
Translation
DNA
Nuclearenvelope
Ribosome
Polypeptide
Figure 3.34 Simplified scheme of information flow from theDNA gene to mRNA to protein structure during transcriptionand translation. (Note that mRNA is first synthesized as pre-mRNA,which is processed by enzymes before leaving the nucleus.)
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 101

transcription and then bind to the promoter. The promoter is aspecial DNA sequence that contains the start point (beginningof the structural gene to be transcribed). It specifies wheremRNA synthesis starts and which DNA strand is going to serveas the template strand (Figure 3.35, top). The uncoiled DNAstrand not used as a template is called the coding strand becauseit has the same (coded) sequence as the mRNA to be built (ex-cept for the use of U in mRNA in place of T in DNA). The tran-scription factors also help position RNA polymerase, theenzyme that oversees the synthesis of mRNA, correctly at thepromoter. Once these preparations are made, RNA polymerasecan initiate transcription.
Figure 3.35 illustrates the following steps in transcription:
Initiation. Once bound with the help of the transcriptionfactors, RNA polymerase pulls apart the strands of the
1
DNA double helix so that transcription can begin at thestart point in the promoter.
Elongation. Using incoming RNA nucleotides as sub-strates, the RNA polymerase aligns them with complemen-tary DNA bases on the template strand and then links themtogether. As RNA polymerase elongates the mRNA strandone base at a time, it unwinds the DNA helix in front of it,and rewinds the helix behind it. At any given moment, 16 to18 base pairs of DNA are unwound and the most recentlymade mRNA is still hydrogen-bonded (H-bonded) to thetemplate DNA. This small region—called the DNA-RNAhybrid—is up to 12 base pairs long.
Termination. When the polymerase reaches a special basesequence called a termination signal, transcription endsand the newly formed mRNA pulls off the DNA template.
3
2
102 UNIT 1 Organization of the Body
3
RNA polymerase
RNA polymerase
RNApolymerase
DNACoding strand
Template strandPromoterregion
Terminationsignal
mRNA
mRNA
Template strand
mRNA transcript
Completed mRNA transcript
Rewinding of DNA
Coding strand of DNA
DNA-RNA hybrid region
The DNA-RNA hybrid: At any given moment, 16–18 base pairs ofDNA are unwound and the most recently made RNA is still bound to DNA. This small region is called the DNA-RNA hybrid.
Templatestrand
Unwindingof DNA
RNA nucleotidesDirection oftranscription
1 Initiation: With the help of transcription factors, RNA polymerase binds to the promoter, pries apart the two DNA strands, and initiates mRNA synthesis at the start point on the template strand.
3 Termination: mRNA synthesis ends when the termination signal is reached. RNA polymerase and the completed mRNA transcript are released.
2 Elongation: As the RNA polymerase moves along the template strand, elongating the mRNA transcript one base at a time, it unwinds the DNA double helix before it and rewinds the double helix behind it.
Figure 3.35 Overview of stages of transcription.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 102

Processing of mRNA Although it would appear that transla-tion can begin as soon as the mRNA is made, that is not the case.The process is a bit more complex. Recall that mammalian DNAlike ours has coding regions (exons) separated by non-protein-coding regions (introns). Because the DNA is transcribed se-quentially, the mRNA initially made, called pre-mRNA orprimary transcript, is littered with intron “junk” or “nonsense.”Before the newly formed RNA can be used as a messenger, itmust be processed, or edited—that is, sections corresponding tointrons must be removed. This job is done by spliceosomes.These large RNA-protein complexes snip out the introns andsplice together the remaining exon-coded sections in the orderin which they occurred in the DNA, producing the functionalmRNA that directs translation at the ribosome.
Although many introns naturally degrade, some contain ac-tive segments (such as microRNAs) that can function to con-trol, interfere with, or silence other genes. Additionally, beforethe edited mRNA can function in protein synthesis, a numberof specific RNA-binding proteins, called mRNA complex pro-teins, must become associated with it. These mRNA complexproteins guide its export from the nucleus, determine its local-ization, translation, and stability, and check it for premature ter-mination codons.
Translation
A translator takes a message in one language and restates it inanother. In the translation step of protein synthesis, the lan-guage of nucleic acids (base sequence) is translated into the lan-guage of proteins (amino acid sequence).
The rules by which the base sequence of a gene is translatedinto an amino acid sequence are called the genetic code. Foreach triplet, or three-base sequence on DNA, the correspondingthree-base sequence on mRNA is called a codon. Since there arefour kinds of RNA (or DNA) nucleotides, there are 43, or 64,possible codons. Three of these 64 codons are “stop signs” thatcall for termination of polypeptide synthesis. All the rest codefor amino acids. Because there are only about 20 amino acids,some are specified by more than one codon. This redundancy inthe genetic code helps protect against problems due to tran-scription (and translation) errors. The genetic code and a com-plete codon list are provided in Figure 3.36.
The process of translation occurs in the cytoplasm and in-volves the mRNAs, tRNAs, and rRNAs mentioned above, andoccurs in the sequence of events described next and illustratedin Figure 3.37.
When it reaches the cytoplasm after processing in the nu-cleus, the mRNA molecule carrying instructions for a particularprotein binds to a small ribosomal subunit (Figure 3.37, ).Then tRNA comes into the picture, its job being to transferamino acids, dissolved in the cytosol, to the ribosome.
Shaped like a handheld drill, tRNA is well suited to its dualfunction of binding to both an amino acid and an mRNAcodon. The amino acid is bound to one end of tRNA, at a regioncalled the stem. At the other end, the head, is its anticodon(an�ti-ko�don), a three-base sequence complementary to themRNA codon calling for the amino acid carried by that partic-
1
Chapter 3 Cells: The Living Units 103
3
SECOND BASE
UUG
UUA
UUC
UUUPhe
Leu
FIR
ST B
ASE
THIR
D B
ASE
CUG
CUA
CUC
CUU
Leu
AUA
AUC
AUU
Ile
GUG
GUA
GUC
GUU
Val
UCG
UCA
UCC
UCU
Ser
CCG
CCA
CCC
CCU
Pro
ACG
ACA
ACC
ACU
Thr
GCG
GCA
GCC
GCU
Ala
UAC
UAUTyr
CAG
CAA
CAC
CAUHis
Gln
AAG
AAA
AAC
AAUAsn
Lys
GAG
GAA
GAC
GAUAsp
Glu
UGC
UGUCys
Trp
CGG
CGA
CGC
CGU
Arg
AGG
AGA
AGC
AGUSer
Arg
GGG
GGA
GGC
GGU
Gly
UAA Stop UGA Stop
AUGMet orStart
UAG Stop UGG
U C A G
G
A
C
U
G
A
C
U
G
A
C
U
G
A
C
U
U
C
A
G
Figure 3.36 The genetic code. The three bases in an mRNA codonare designated as the first, second, and third. Each set of threespecifies a particular amino acid, represented here by an abbreviation(see list below). The codon AUG (which specifies the amino acidmethionine) is the usual start signal for protein synthesis. The wordstop indicates the codons that serve as signals to terminate proteinsynthesis.
Abb.* Amino acidAla alanineArg arginineAsn asparagineAsp aspartic acidCys cysteineGlu glutamic acidGln glutamineGly glycineHis histidineIle isoleucine
Abb.* Amino acidLeu leucineLys lysineMet methioninePhe phenylalaninePro prolineSer serineThr threonineTrp tryptophanTyr tyrosineVal valine
*Abbreviation for the amino acid
ular tRNA. Because anticodons form hydrogen bonds withcomplementary codons, tRNA is the link between the languageof nucleic acids and the language of proteins. For example, if themRNA codon is AUA, which specifies isoleucine, the tRNAs car-rying isoleucine will have the anticodon UAU, which can bind tothe AUA codons.
There are approximately 45 different types of tRNA, eachcapable of binding with a specific amino acid. The attachment
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 103

process is controlled by an aminoacyl-tRNA synthetase enzymeand is activated by ATP (see the top right side of Figure 3.37).Once its amino acid is loaded, the tRNA (now called anaminoacyl-tRNA because of its amino acid cargo) migrates to theribosome, where it maneuvers the amino acid into the properposition, as specified by the mRNA codons (Figure 3.37, ).2
The ribosome is more than just a passive attachment site formRNA and tRNA. Like a vise, the ribosome holds the tRNAand mRNA close together to coordinate the coupling of codonsand anticodons, and positions the next (incoming) amino acidfor addition to the growing polypeptide chain. To do its job, theribosome has a binding site for mRNA and three binding sites
104 UNIT 1 Organization of the Body
3 1 After mRNA synthesis in the nucleus, mRNA leaves the nucleus and attaches to a ribosome.
2 Translation begins as incoming aminoacyl-tRNA recognizes the complementary codon calling for it at the A site on the ribosome. It hydrogen-bonds to the codon via its anticodon.
3 As the ribosome moves along the mRNA, and each codon is read in sequence, a new amino acid is added to the growing protein chain and the tRNA in the A site is translocated to theP site.
4 Once its amino acid is released from the P site, tRNA is ratcheted to the E site and then released to reenter the cytoplasmic pool, ready to be recharged with a new amino acid. The polypeptide is released when the stop codon is read.
Leu
Leu
Energized by ATP, the correct aminoacid is attached to each species oftRNA by aminoacyl-tRNA synthetaseenzyme.
Amino acid
tRNA
Aminoacyl-tRNAsynthetase
G A A
G A AtRNA “head”bearinganticodon
Psite A
siteE
site
Ile
Pro
A AU
UU
A
U UC C C
CG G
G
Largeribosomalsubunit
Smallribosomalsubunit
Direction ofribosome advancePortion of mRNA
already translated
Codon15
Codon16
Codon17
Nucleus
mRNA
Released mRNA
Nuclearmembrane
Nuclear pore
RNA polymerase
Templatestrand ofDNA
Figure 3.37 The basic steps of translation. The diagram presumes that initiation wasaccomplished and the 17th amino acid as dictated by the mRNA codons is being brought tothe ribosome.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 104

for tRNA: an A (aminoacyl) site for an incoming aminoacyl-tRNA, a P (peptidyl) site for the tRNA holding the growingpolypeptide chain, and an E (exit) site for an outgoing tRNA(Figure 3.37).
How does translation get started? When a special methionine-charged initiator tRNA binds to the middle (P) site on the smallribosomal subunit, the translation process officially begins. Thenewly formed mRNA has an initial base sequence, called aleader sequence, that allows it to attach to its binding site on thesmall ribosomal subunit. With the initiator tRNA still in tow,the small ribosomal subunit scans along the mRNA until it en-counters the start codon (the first AUG triplet it meets). Whenthe initiator tRNA’s UAC anticodon “recognizes” and binds tothe start codon, a large ribosomal subunit unites with the smallone, forming a functional ribosome. With the mRNA firmly po-sitioned in the “groove” between the two ribosomal subunits,translation begins in earnest. This initiation process requires thehelp of a number of initiation factors and is energized by GTP.
Now the ribosome slides the mRNA strand along, bringingthe next codon into position to be “read” by an aminoacyl-tRNAcoming into the A site (Figure 3.37, ). It is at this point that theribosome does a little “proofreading” to make sure of the codon-anticodon match. That accomplished, an enzymatic componentin the large ribosomal particle peptide-bonds the amino acid ofthe initiator tRNA in the P site to that of the tRNA at the A site.The ribosome then translocates the tRNA that is now carryingtwo amino acids to the P site (as in Figure 3.37, ). At the sametime, it ratchets the initiator tRNA to the E site, from which itleaves the ribosome (as in Figure 3.37, ).
This orderly “musical chairs” process continues: the peptidyl-tRNAs transferring their polypeptide cargo to the aminoacyl-tRNAs, and then the P-site-to-E-site and A-site-to-P-sitemovements of the tRNAs (see again Figure 3.37, and ). Asthe ribosome “chugs” along the mRNA track and the mRNA isprogressively read, its initial portion passes through the ribo-some and may ultimately become attached successively to severalother ribosomes, all reading the same message simultaneouslyand sequentially. Such a multiple ribosome–mRNA complex iscalled a polyribosome, and it provides an efficient system for pro-ducing many copies of the same protein (Figure 3.38).
The mRNA strand continues to be read sequentially until itslast codon, the stop codon (one of UGA, UAA, or UAG) enters theribosomal groove. The stop codon is the “period” at the end ofthe mRNA sentence that tells the ribosome that translation ofthat mRNA is finished. The completed polypeptide chain is thenreleased from the ribosome, and the ribosome separates into itstwo subunits (Figure 3.38a). The released protein folds into acomplex 3-D structure and floats off, ready to work. When themessage of the mRNA that directed its formation becomes out-dated, it is degraded in structures called processing (P) bodies.
As noted earlier in the chapter, ribosomes attach to and de-tach from the rough ER. When a short “leader”peptide called anER signal sequence is present in a protein being synthesized, theassociated ribosome attaches to the membrane of the rough ER.This signal sequence, with its attached cargo of a ribosomeand mRNA, is guided to appropriate receptor sites on the ERmembrane by a signal-recognition particle (SRP), a molecular
43
4
3
2
chaperone that cycles between the ER and the cytosol. The sub-sequent events occurring at the ER are detailed in Figure 3.39.
In summary, the genetic information of a cell is translatedinto the production of proteins via a sequence of informationtransfer that is completely directed by complementary basepairing. The transfer of information goes from DNA base se-quence (triplets) to the complementary base sequence ofmRNA (codons) and then to the tRNA base sequence (anti-codons), which is identical to the DNA sequence except for thesubstitution of uracil (U) for thymine (T) (Figure 3.40).
Other Roles of DNAThe story of DNA doesn’t end with the production of proteinsencoded by exons. Scientists are finding that other intron or“junk” DNA actually codes for a surprising variety of activeRNA species, including the following:
■ Antisense RNAs, made on the DNA strand complementaryto the template strand for mRNA, can intercept and bind tothe protein-coding mRNA strand and prevent it from beingtranslated into protein.
■ MicroRNAs are small RNAs that can use RNA interferencemachinery to interfere with and suppress mRNAs made bycertain exons, thus effectively silencing them.
■ Riboswitches are folded RNAs that code, like mRNA, for aparticular protein. What sets them apart from other mRNAs is
Chapter 3 Cells: The Living Units 105
3Start of mRNA
Incomingribosomalsubunits
Growingpolypeptides
Completedpolypeptide
End of mRNA
Polyribosome
Ribosomes
mRNA
(b) This transmission electron micrograph showsa large polyribosome (400,000�).
(a) Each polyribosome consists of one strand of mRNA being read by several ribosomes simultaneously. In this diagram, the mRNA is moving to the left and the “oldest” functional ribosome is farthest to the right.
Figure 3.38 Polyribosome arrays allow a single strand ofmRNA to be translated into hundreds of the same polypeptidemolecules in a short time.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 105

a region that acts as a switch to turn protein synthesis on or offin response to specific metabolic changes in the environment,such as changes in the concentration of vitamins, amino acids,nucleotides, or other small molecules in the cell. When itsenses these changes, the riboswitch changes shape, therebystopping or starting production of the protein it specifies.
We still have much to learn about these versatile RNA speciesthat arise from intron and “junk” DNA and appear to play a rolein heredity. Other areas of research focus on the consequences ofthe triplet code being degenerate (that is, having several codonscode for a single amino acid), and on the multitasking of DNAsegments. For example, a sequence can code for structure of aprotein and also manage to guide positioning of a nucleosome.
C H E C K Y O U R U N D E R S TA N D I N G
29. Codons and anticodons are both three-base sequences. Howdo they differ?
30. How do the A, P, and E ribosomal sites differ functionallyduring protein synthesis?
31. What is the role of DNA in transcription?
For answers, see Appendix G.
Cytosolic Protein Degradation� Describe the importance of ubiquitin-dependent degradation
of soluble proteins.
What happens to proteins that are no longer useful? To carryout their physiological roles, proteins must be in the right placeat the right time and in the right amounts. Like everything else,proteins outlive their usefulness and are eventually degraded.Organelle proteins are digested by lysosomes, but lysosomal en-zymes do not have access to misfolded, damaged, or unneededsoluble proteins in the cytosol that need to be disposed of. Forexample, these proteins might include some used only in cell di-vision, or short-lived transcription factors.
So, how does the cell prevent such proteins from accumulat-ing while avoiding wholesale destruction of virtually all solubleproteins by the cytosolic enzymes? Doomed proteins aremarked for attack (proteolysis) by attachment of proteins calledubiquitins (u-bı�kwı-tinz) in an ATP-dependent reaction. Thetagged proteins are then hydrolyzed to small peptides by solubleenzymes or by proteasomes, giant “waste disposal” complexescomposed of protein-digesting enzymes and the ubiquitin is
106 UNIT 1 Organization of the Body
3
1 The mRNA-ribosome complex is directed to the rough ER by the SRP. There the SRP binds to a receptor site.
2 Once attached to the ER, the SRP is released and the growing polypeptide snakes through the ER membrane pore into the cisterna.
3 The signal sequence is clipped off by an enzyme. As protein synthesis continues, sugar groups may be added to the protein.
4 In this example, the completed protein is released from the ribosome and folds into its 3-D conformation, a process aided by molecular chaperones.
5 The protein is enclosed within a protein (coatomer)-coated transport vesicle.The transport vesicles make their way to theGolgi apparatus, where further processingof the proteins occurs (see Figure 3.19).
RibosomeER signalsequence
Signalrecognitionparticle(SRP)
Receptor site
mRNA
Growingpolypeptide
Signalsequenceremoved
Sugargroup
Releasedprotein
Transport vesiclepinching off
Coatomer-coatedtransport vesicle
Rough ER cisterna
Cytoplasm
Figure 3.39 Presence of an endoplasmic reticulum (ER) signal sequence in a newlyforming protein causes the signal recognition particle (SRP) to direct the mRNA-ribosomecomplex to the rough ER.
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 106

recycled. Proteasome activity is critical during starvation whenthese complexes degrade preexisting proteins to provide aminoacids for synthesis of new and needed proteins.
Extracellular Materials� Name and describe the composition of extracellular ma-
terials.
Extracellular materials are any substances contributing tobody mass that are found outside the cells. One class of extracel-lular materials is body fluids, mainly interstitial fluid, bloodplasma, and cerebrospinal fluid. These fluids are importanttransport and dissolving media. Cellular secretions are also ex-tracellular materials and include substances that aid in digestion(intestinal and gastric fluids) and some that act as lubricants(saliva, mucus, and serous fluids).
By far the most abundant extracellular material is theextracellular matrix. Most body cells are in contact with a jelly-like substance composed of proteins and polysaccharides. Thesemolecules are secreted by the cells and self-assemble into an or-ganized mesh in the extracellular space, where they serve as auniversal “cell glue” that helps to hold body cells together. As de-scribed in Chapter 4, the extracellular matrix is particularlyabundant in connective tissues—in some cases so abundantthat it (rather than living cells) accounts for the bulk of that tis-sue type. Depending on the structure to be formed, the extracel-lular matrix in connective tissue ranges from soft to rock-hard.
C H E C K Y O U R U N D E R S TA N D I N G
32. What is the importance of ubiquitin in the life of a cell?33. What are two body fluids that inhabit the extracellular space
and what role does each play in the body?
For answers, see Appendix G.
Chapter 3 Cells: The Living Units 107
3
DNA molecule
Gene 1
Gene 2
Gene 4
DNA: DNA base sequence(triplets) of the gene codesfor synthesis of a particularpolypeptide chain
T A
1
C G T A G C G A T T T C C C T G C G A A A A C T
2 3 4 5 6 7 8 9
Codons
Triplets
tRNA
A U
1
G C C A U C G C U A A A G G G A C G C U U U
U A C G G U A G C G A U U U C C C U G C G A A A
U G A
2 3 4 5 6 7 8 9
Met Pro Ser Leu Lys Gly Arg Phe
Start
Stop;detach
G
Anticodon
mRNA: Base sequence(codons) of the transcribedmRNA
tRNA: Consecutive basesequences of tRNA anticodons able to recognize the mRNA codons calling for the amino acids they transport
Polypeptide: Amino acid sequence of the polypeptide chain
Figure 3.40 Information transfer from DNA to RNA to polypeptide. Information istransferred from the DNA of the gene to the complementary messenger RNA molecule, whosecodons are then “read” by transfer RNA anticodons. Notice that the “reading” of the mRNA bytRNA anticodons reestablishes the base (triplet) sequence of the DNA genetic code (except thatT is replaced by U).
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 107

Developmental Aspects of Cells� Discuss some theories of cell differentiation and aging.
� Indicate the value of apoptosis to the body.
We all begin life as a single cell, the fertilized egg, and all the cellsof our body arise from it. Very early in development, cells beginto specialize. Some become liver cells, some nerve cells, and soon. All our cells carry the same genes, so how can one cell be-come so different from another? This is a fascinating question.
Apparently, cells in various regions of the embryo are ex-posed to different chemical signals that channel them into spe-cific pathways of development. When the embryo consists ofjust a few cells (these are the embryonic stem cells you keep hear-ing about), the major signals may be nothing more than slightdifferences in oxygen and carbon dioxide concentrations be-tween the more superficial and the deeper cells. But as devel-opment continues, cells release chemicals that influencedevelopment of neighboring cells by triggering processes thatswitch some genes “off.”Some genes are active in all cells. For ex-ample, genes for rRNA and ATP synthesis are “on” in all cells,but genes for synthesizing the enzymes needed to produce thy-roxine are “on” only in cells that are going to be part of the thy-roid gland. Hence, the secret of cell specialization lies in thekinds of proteins made and reflects the activation of differentgenes in different cell types.
Cell specialization leads to structural variation—different or-ganelles come to predominate in different cells. For example,muscle cells make large amounts of actin and myosin, and theircytoplasm fills with microfilaments. Liver and phagocytic cellsproduce more lysosomes. The development of specific and dis-tinctive features in cells is called cell differentiation.
During early development, cell death and destruction arenormal events. Nature takes few chances. More cells thanneeded are produced, and excesses are eliminated later in a typeof programmed cell death called apoptosis (ap�o-to�sis; “fallingaway”). This process of controlled cellular suicide also elimi-nates cells that are stressed, no longer needed, injured, or aged.
How does apoptosis work? In response to damaged macro-molecules within the cell or to some extracellular signal, mito-chondrial membranes become permeable, allowing cytochromec and other factors to leak into the cytosol. These chemicals, inturn, trigger apoptosis by causing activation of a series of intra-cellular enzymes called caspases. These enzymes are normallydormant, but when they are activated, they unleash a torrent ofprotein digestion activity within the cell, destroying the cell’sDNA, cytoskeleton, and so on, producing a quick, neat death.The apoptotic cell shrinks without leaking its contents into thesurrounding tissue, detaches from other cells, and rounds up.Because the dying cell releases a chemical (lysophosphatidyl-choline) that attracts macrophages, and sprouts “eat me” signals,it is immediately phagocytized.
Cancer cells do not undergo apoptosis, but oxygen-starvedcells do so excessively (heart-muscle and brain cells during heartattacks and strokes, for example). Apoptosis is particularly com-mon in the developing nervous system. It is also responsible for“carving out” fingers and toes from their embryonic precursors.
Most organs are well formed and functional long beforebirth, but the body continues to grow and enlarge by formingnew cells throughout childhood and adolescence. Once wereach adult size, cell division is important mainly to replaceshort-lived cells and repair wounds.
During young adulthood, cell numbers remain fairly con-stant. However, local changes in the rate of cell division arecommon. For example, when a person is anemic, his or herbone marrow undergoes hyperplasia (hi�per-pla�ze-ah), or ac-celerated growth (hyper � over; plas � grow), so that red bloodcells are produced at a faster rate. If the anemia is remedied, theexcessive marrow activity ceases. Atrophy (at�ro-fe), a decreasein size of an organ or body tissue, can result from loss of normalstimulation. Muscles that lose their nerve supply atrophy andwaste away, and lack of exercise leads to thinned, brittle bones.
Cell aging occurs and it accounts for most problems associ-ated with old age. Cell aging is a complicated process withmany causes. The wear-and-tear theory attributes aging to lit-tle chemical insults and formation of free radicals, both ofwhich have cumulative effects. For example, environmentaltoxins such as pesticides, alcohol, and bacterial toxins maydamage cell membranes, poison enzyme systems, or cause“mistakes” in DNA replication. Temporary lack of oxygen,which occurs increasingly with age as our blood vessels clogwith fatty materials, leads to accelerated rates of cell deaththroughout the body.
Most free radicals are produced in the mitochondria, becausethese organelles have the highest metabolic rate. This findingimplies that diminished energy production by radical-damagedmitochondria, due perhaps to an increasing burden of muta-tions in mitochondrial DNA, weakens (and ages) cells. X raysand other types of radiation, and some chemicals, also generatehuge numbers of free radicals, which can overwhelm the perox-isomal enzymes. Vitamins C and E act as antioxidants in thebody and may help to prevent excessive free radical formation.With age, glucose (blood sugar) becomes party to cross-linkingproteins together, a condition that severely disrupts proteinfunction and accelerates the course of arteriosclerosis.
Another theory attributes cell aging to progressive disordersin the immune system. According to this theory, cell damage re-sults from (1) autoimmune responses, which means the im-mune system turns against one’s own tissues, and (2) aprogressive weakening of the immune response, so that thebody is less and less able to get rid of cell-damaging pathogens.
Perhaps the most widely accepted theory of cell aging is thegenetic theory, which suggests that cessation of mitosis and cellaging are “programmed into our genes.” One interesting notionhere is that a telomere clock determines the number of times acell can divide. Telomeres (telo � end; mer � piece) are strings ofnucleotides that cap the ends of chromosomes, protecting themfrom fraying or fusing with other chromosomes, much likeplastic caps preserve the ends of shoelaces. In human telomeres,the base sequence TTAGGG is repeated a thousand times ormore (like a DNA stutter). Though telomeres carry no genes,they appear to be vital for chromosomal survival, because eachtime DNA is replicated, 50 to 100 of the end nucleotides are lost
108 UNIT 1 Organization of the Body
3
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 108

and the telomeres get a bit shorter. When telomeres reach a cer-tain minimum length, the stop-division signal is given.
The idea that cell longevity depends on telomere integritywas supported by the discovery of telomerase, an enzyme thatprotects telomeres from degrading. Pegged as the “immortalityenzyme,” telomerase is found in germ line cells (cells that giverise to sperm and ova), but it is barely detectable or absent inother adult cell types.
C H E C K Y O U R U N D E R S TA N D I N G
34. What is apoptosis and what is its importance in the body?35. What is the wear-and-tear theory of aging?
For answers, see Appendix G.
In this chapter we have described the structure and functionof the generalized cell. One of the wonders of the cell is the dis-parity between its minute size and its extraordinary activity,which reflects the diversity of its organelles. The evidence for di-vision of labor and functional specialization among organellesis inescapable. Only ribosomes synthesize proteins, while pro-tein packaging is the bailiwick of the Golgi apparatus. Mem-branes compartmentalize most organelles, allowing them towork without hindering or being hindered by other cell activi-ties, and the plasma membrane regulates molecular trafficacross the cell’s boundary. Now that you know what cells have incommon, you are ready to explore how they differ in the variousbody tissues, the topic of Chapter 4.
Chapter 3 Cells: The Living Units 109
3
Anaplasia (an�ah-pla�ze-ah; an � without, not; plas � to grow) Ab-normalities in cell structure and loss of differentiation; for exam-ple, cancer cells typically lose the appearance of the parent cellsand come to resemble undifferentiated or embryonic cells.
Dysplasia (dis-pla�ze-ah; dys � abnormal) A change in cell size,shape, or arrangement due to chronic irritation or inflammation(infections, etc.).
Hypertrophy (hi-per�tro-fe) Growth of an organ or tissue due to anincrease in the size of its cells. Hypertrophy is a normal responseof skeletal muscle cells when they are challenged to lift excessiveweight; differs from hyperplasia, which is an increase in size dueto an increase in cell number.
Liposomes (lip�o-somz) Hollow microscopic sacs formed of phos-pholipids that can be filled with a variety of drugs. Serve as mul-tipurpose vehicles for drugs, genetic material, and cosmetics.
Mutation A change in DNA base sequence that may lead to incorpo-ration of incorrect amino acids in particular positions in the re-sulting protein; the affected protein may remain unimpaired ormay function abnormally or not at all, leading to disease.
Necrosis (ne-kro�sis; necros � death; osis � process) Death of a cellor group of cells due to injury or disease. Acute injury causes thecells to swell and burst, and induces the inflammatory response.(This is uncontrolled cell death, in contrast to apoptosis de-scribed in the text.)
RELATED CLINICAL TERMS
Media study tools that could provide you additional help in review-ing specific key topics of Chapter 3 are referenced below.
= Interactive Physiology
Overview of the Cellular Basis of Life (pp. 62–63)
1. All living organisms are composed of cells—the basic structuraland functional units of life. Cells vary widely in both shape andsize.
2. The principle of complementarity states that the biochemicalactivity of cells reflects the operation of their organelles.
3. The generalized cell is a concept that typifies all cells. The gener-alized cell has three major regions—the nucleus, cytoplasm, andplasma membrane.
The Plasma Membrane: Structure (pp. 63–67)
1. The plasma membrane encloses cell contents, mediates exchangeswith the extracellular environment, and plays a role in cellularcommunication.
The Fluid Mosaic Model (pp. 63–66)
2. The fluid mosaic model depicts the plasma membrane as a fluidbilayer of lipids (phospholipids, cholesterol, and glycolipids)within which proteins are inserted.
3. The lipids have both hydrophilic and hydrophobic regions thatorganize their aggregation and self-repair. The lipids form thestructural part of the plasma membrane.
4. Most proteins are integral transmembrane proteins that extendentirely through the membrane. Some, appended to the integralproteins, are peripheral proteins.
5. Proteins are responsible for most specialized membrane func-tions: Some are enzymes, some are receptors, and others mediatemembrane transport functions. Externally facing glycoproteinscontribute to the glycocalyx.
Membrane Junctions (p. 66–67)
6. Membrane junctions join cells together and may aid or inhibitmovement of molecules between or past cells.
7. Tight junctions are impermeable junctions. Desmosomes me-chanically couple cells into a functional community. Gap junc-tions allow joined cells to communicate.
The Plasma Membrane: Membrane Transport (pp. 68–77)
1. The plasma membrane acts as a selectively permeable barrier.Substances move across the plasma membrane by passiveprocesses, which depend on the kinetic energy of molecules,and by active processes, which depend on the use of cellular en-ergy (ATP).
CHAPTER SUMMARY
000200010270575674_R1_CH03_p0061-0112.qxd 12/2/2011 11:30 AM Page 109
Copyright © 2022 FDOKUMEN




















