
Potential Role of NKG2D/MHC Class I-Related Chain A Interaction in Intrathymic Maturation of...
10
of June 13, 2013. This information is current as Maturation of Single-Positive CD8 T Cells I-Related Chain A Interaction in Intrathymic Potential Role of NKG2D/MHC Class Sophie Caillat-Zucman Sophie Hüe, Renato C. Monteiro, Sonia Berrih-Aknin and http://www.jimmunol.org/content/171/4/1909 2003; 171:1909-1917; ; J Immunol References http://www.jimmunol.org/content/171/4/1909.full#ref-list-1 , 23 of which you can access for free at: cites 55 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2003 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on June 13, 2013 http://www.jimmunol.org/ Downloaded from
Transcript of Potential Role of NKG2D/MHC Class I-Related Chain A Interaction in Intrathymic Maturation of...

of June 13, 2013.This information is current as
Maturation of Single-Positive CD8 T CellsI-Related Chain A Interaction in Intrathymic Potential Role of NKG2D/MHC Class
Sophie Caillat-ZucmanSophie Hüe, Renato C. Monteiro, Sonia Berrih-Aknin and
http://www.jimmunol.org/content/171/4/19092003; 171:1909-1917; ;J Immunol
Referenceshttp://www.jimmunol.org/content/171/4/1909.full#ref-list-1
, 23 of which you can access for free at: cites 55 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2003 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

Potential Role of NKG2D/MHC Class I-Related Chain AInteraction in Intrathymic Maturation of Single-Positive CD8T Cells1
Sophie Hue,* Renato C. Monteiro,† Sonia Berrih-Aknin, ‡ and Sophie Caillat-Zucman2*
The nonclassical MHC class I molecule MHC class I-related chain A (MICA) interacts with the NKG2D receptor expressed at thesurface of most peripheral CD8 T cells,�� T cells, and NK cells. We investigated the role of MICA-NKG2D interactions in theselection or maturation of the T cell repertoire within the thymus using MICA tetramers and anti-MICA mAbs. MICA tetramersidentified a small population of late stage CD8 single-positive, CD45RA� CD62L� CCR7� CD69� thymocytes, a phenotypecompatible with that of fully mature CD8 � cells ready to emigrate to the periphery as naive cells. MICA molecules were expressedin the outer layer of Hassal’s corpuscles within the medulla of normal thymus. In thymomas, an overexpression of MICA incortical and medullar epithelial cells was observed. This was associated with a decreased percentage of NKG2D-positive thymo-cytes, which expressed a less mature phenotype than in normal thymus. These results indicate that CD8� thymocytes up-regulateNKG2D as they complete their developmental program before leaving the thymic medulla to seed the periphery, and identifyNKG2D as a potential regulator of the developmental processes in T cells that are essential for immune homeostasis.The Journalof Immunology, 2003, 171: 1909–1917.
T lymphocyte development in the thymus is a complex pro-cess essential for the formation of the immune system.Different developmental stages of T cells within the thy-
mus can be defined by expression of the coreceptor molecules CD4and CD8. Immature T cell precursors are found within theCD4�CD8� double-negative compartment. Cells that successfullyrearrange their TCR � locus and express a functional pre-TCRproliferate rapidly and differentiate into CD4�CD8� double-pos-itive cells. These cells undergo TCR �-chain gene rearrangements,and upon expression of a functional �� TCR complex are sub-jected to the processes of positive and negative selection that gen-erate CD4� or CD8� single-positive (SP)3 thymocytes (1, 2). Inaddition to these changes in expression of surface markers, thy-mocytes traverse through the thymus as they develop. Immaturethymocytes reside in the cortex, and as they begin the selectionprocess they traffic from the cortex to the medulla.
A key role of thymic epithelial cells in positive selection is toprovide HLA/peptide ligands for the �� TCR. In addition, thethymic medulla is likely to be involved in regulating postselectiondifferentiation events at the level of SP thymocytes (3). Newlygenerated SP thymocytes reside in the medullar areas for up to 2wk, during which they undergo changes in expression of a variety
of cell surface molecules such as CD24, CD62L, and CD69, beforethey are exported to the periphery (4, 5). At the same time, posi-tively selected thymocytes undergo an expansion phase that mayact to increase numbers of T cells to aid the establishment of theperipheral T cell pool (6). However, the nature of signals requiredfor emigration of newly matured T cells out of the thymus is stillelusive.
The NKG2D-activating receptor is expressed at the surface ofmost circulating �� CD8 T cells, �� T cells, and NK cells inhumans. NKG2D forms homodimers that pair with the transmem-brane adaptor protein DAP10. Like the CD28 molecule, DAP10contains an SH2-binding YxxM motif in the cytoplasmic domainthat, upon tyrosine phosphorylation, recruits the p85 catalytic sub-unit of the phosphatidylinositol 3 kinase (7). Various families ofNKG2D ligands have been identified, including the human MHCclass I-related chains (MIC) A and B proteins (8, 9). These arenonclassical MHC class I molecules that have no role in Ag pre-sentation, but function as signals of cellular distress, and trigger arange of effector mechanisms, including cellular cytotoxicity, cy-tokine secretion, and cellular proliferation (for review, see Refs. 10and 11). In NK cells, MICA can transduce through NKG2D, anactivating signal that can override a negative signal generated byengagement of inhibitory receptors for MHC class I (12–14). InCD8 �� T cells, NKG2D/MIC engagement delivers a costimula-tory signal that complements TCR-mediated Ag recognition ontarget cells (15). Recent data indicate that two splice variants ofNKG2D may associate differently with the adaptor moleculesDAP10 and DAP12 (16, 17). NK express both DAP10 andDAP12, and can therefore transmit both direct stimulatory andcostimulatory signals. In CD8 T cells, due to the absence ofDAP12, NKG2D interacts with DAP10 and thus delivers only acostimulatory signal to T cells. In addition, it has been shown thatMIC is recognized by intraepithelial or tumor-infiltrating �� Tcells (18, 19) and is able to deliver both a TCR-dependent signal1 and a NKG2D-dependent costimulatory signal 2 for a subset ofV�1 �� T cells (20).
*Laboratory of Immunology, Hopital Necker, Paris, France; †Institut National de laSante et de la Recherche Medicale E-0225, Faculte Bichat, Paris, France; and ‡CentreNational de la Recherche Scientifique ESA 8078, Center Chirurgical Marie-Lan-nelongue, Le Plessis-Robinson, France
Received for publication February 14, 2003. Accepted for publication June 3, 2003.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 S.H. was supported by Assistance Publique Hopitaux de Paris and by a Posted’Accueil Institut National de la Sante et de la Recherche Medicale.2 Address correspondence and reprint requests to Dr. Sophie Caillat-Zucman, Labo-ratory of Immunology, Hopital Necker, 161 rue de Sevres, 75015 Paris, France. E-mail address: [email protected] Abbreviations used in this paper: SP, single positive; IEL, intraepithelial lympho-cyte; MIC, MHC class I-related chain; TREC, TCR� rearrangement excision circles;ULBP, UL-16-binding protein.
The Journal of Immunology
Copyright © 2003 by The American Association of Immunologists, Inc. 0022-1767/03/$02.00
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

At variance with the wide constitutive expression of conven-tional MHC class I molecules, expression of MICA molecules islow in adult tissues, is mostly restricted to the surface of intestinalepithelium (9), but is stress inducible in various epithelial cells. Itis also considerably up-regulated in different types of tumors (19,21) and in the context of intracellular viral (15) or bacterial infec-tion (22, 23). In addition to gut epithelium, MICA expression hasbeen suggested in a population of stellate cells located in the sub-capsular thymic cortex (9). This remarkably restricted expressionof MICA in two known sites of T lymphopoiesis suggests a pos-sible role of MICA/NKG2D interaction in the selection or matu-ration of the T cell repertoire.
In this study, we show that MICA expression is observed inepithelial cells and Hassal’s corpuscles of the thymic medulla. Fur-thermore, NKG2D is expressed on a subpopulation of CD4�CD8�
medullar thymocytes that express a phenotype of CD45RA�,CD69�, CD62L�, CCR7� naive mature T cells. These data sug-gest that NKG2D might be a signal allowing newly matured CD8T cells to emigrate to the periphery. Such hypothesis is sustainedby observation that in thymoma, an overexpression of MICA mol-ecules on tumor epithelial cells is accompanied with an abnormalmaturation phenotype of NKG2D-positive thymocytes.
Materials and MethodsCells
HT-29 (ATCC HTB-38, colonic adenocarcinoma), HeLa (ATCC CCL-2,carcinoma of cervix), CaCO2 (ATCC HTB-37, colorectal adenocarcino-ma), and Raji (ATCC CCL-86, lymphoma) cell lines were obtained fromthe American Type Culture Collection (Manassas, VA). M8 (melanomacell line) was kindly provided by P. Paul (Marseille-Luminy, France). Celllines were grown in RPMI 1640 supplemented with 10% FCS, glutamine,and antibiotics. Fresh PBMC from healthy adult volunteers were purifiedfrom whole blood by Ficoll density-gradient centrifugation (AmershamPharmacia Biotech, Piscataway, NJ). Human umbilical cord blood cellswere obtained from healthy delivered specimens and placed in heparinizedcollection tubes. Cord blood mononuclear cells were isolated, as describedabove, for whole blood specimens.
Human postnatal thymus specimens were obtained from 13 childrenduring corrective cardiac surgery. The children were 2 days to 4 years old.All the procedures followed in the study were performed in accordancewith the Declaration of Helsinki.
Thymic samples were stored in saline and processed within 1 h of ex-cision. Mononuclear cells were obtained by gentle teasing of thymus overwire mesh. Thymic cells were washed twice in RPMI before immunoflu-orescence staining. Recovered viable thymocytes (hereafter referred to astotal thymocytes) were depleted in CD4-positive cells by CD4 magneticbead depletion using the MACS system, according to the manufacturer’sprotocol (Miltenyi Biotec, Auburn, CA). The CD4�CD8� cells (up to8–10% of total thymocytes) were �98% pure.
Construction, expression, and purification of soluble MICA andMICB molecules
Recombinant MICA and MICB were produced as secreted proteins in Sf9insect cells using the pBAC4 Baculovirus expression system (Novagen,Madison, WI). The MICA and MICB expression vectors were constructedby PCR using the cDNA coding for the human MICA*001 and MICB*002alleles (kindly provided by S. Bahram, Strasbourg, France). The codingregion for the signal peptide and extracellular domains of the proteins(residues 1–276) fused to a DNA tag encoding a glycine-serine linker andthe recognition sequence for enzymatic biotinylation were amplified usingthe following primers: MICA forward, 5�-TCT GGATCC ACC ATG GGGCTG GGC CCG GTC-3�; MICB forward, 5�-TCT GGATCC ACC ATGGGG CTG GGC CGG GTC-3�; MICA/B reverse, 5�-A CTCGAG GTCCCG CAG CTC CAT CTT CAT TGC CTC AAA GAT TCC TCC AAGTCC ACC GCC GCT TTT CCC AGA GGG CAC AGG GTG AG-3�.
BamHI and XhoI restriction enzyme sites used for cloning are in italic.The antisense primer annealed to codons corresponding to the first 2 aa ofthe transmembrane domain (bold), a glycine-serine linker, and the biotinprotein ligase BirA recognition sequence 45 of Schatz (24) (underlined).
The resulting fragments were cut with BamHI and XhoI and cloned intopBAC4X-11 upstream to a six-His coding sequence and a stop codon. The
recombinant transfer vector was cotransfected with linearized mutant Ac-NPV genomes (Baculogold; BD PharMingen, San Diego, CA) into Sf9 cellsusing Insectin-Plus liposomes (Invitrogen, San Diego, CA), according to themanufacturers’ instructions. Viruses in supernatants from transfected cellswere harvested after 4 days and plaque purified using standard techniques.Plaque-pure virus clones were tested for protein expression by protein immu-noblotting of supernatants from cultures of infected cells using mouse anti-(His)4 mAb (Qiagen, Valencia, CA). Appropriate viruses were expanded incomplete TNM-FH medium (BioWhittaker, Walkersville, MD).
Large-scale preparations were carried in serum-free Sf 900-II medium(Invitrogen). Supernatants were harvested by centrifugation at day 3 afterinfection and concentrated from 80 to 10 ml in Amicon concentrator stircell on BioMax membranes (Millipore, Bedford, MA). MICA and MICBmolecules were affinity purified by Ni-NTA agarose (Qiagen) or HIS-Se-lect HC Nickel affinity gel (Sigma-Aldrich, St. Louis, MO) chromatogra-phy, respectively, and concentrated to 2 mg/ml using Centricon ultrafilters(Amicon, Beverly, MA). MICB protein was further purified by gel filtra-tion chromatography on a Superdex 200 10/30 column (Amersham Phar-macia Biotech) in 50 mM Tris, 150 mM NaCl, pH 8.0. MICA and MIBproteins were 98% pure, as determined by SDS-PAGE and Coomassiestaining.
Generation of fluorescent MICA and MICB tetramers
Biotinylation of soluble recombinant MICA or MICB proteins was donefor 16–24 h at room temperature using BirA-Ligase (Avidity, Denver,CO), according to the manufacturer’s instructions, and was estimated byimmunoprecipitation with streptavidin-agarose beads (Pierce, Rockford,IL). Efficiency was always �90%. Protein concentration was measured bymicro-bicinchoninic acid (Pierce). Tetramers were generated by mixingMICA or MICB monomers with fluorochrome-labeled streptavidin in a 4:1ratio (streptavidin PE or streptavidin FITC; BD PharMingen) without fur-ther separation by gel filtration. For staining, tetramers were used at aconcentration of 10 �g/ml of monomers.
Production of anti-MICA mAbs
Mice (BALB/c, female, 7 wk old) were primed by injecting 50 �g ofpurified rMICA*001 emulsified in CFA (Sigma-Aldrich) in the footpad.Mice were then boosted five times at 3-day interval by footpad injectionwith 50 �g of rMICA emulsified in IFA or PBS. Eighteen hours after thesixth injection, lymphocytes were isolated from inguinal lymph nodes, andwere fused with Ag8.653 myeloma cells (25) using 50% polyethylene gly-col (Sigma-Aldrich). Hybridomas were grown in RPMI-10% FCS in pres-ence of peritoneal macrophages. Supernatants were screened for specificreactivity with rMICA by ELISA and Western blot, and by flow cytometryon HT-29 cells. A total of 38 of 180 hybridomas was reactive with MICAAgs by ELISA. Three hybridomas (SR99, SR104, SR116) were furtherselected on the basis of both reactivity against rMICA by ELISA andstrong staining of HT-29 cells by flow cytometry. After subcloning, ascitefluid was harvested from mice that were pristane primed and inoculated i.p.with mAb-producing hybridoma cells. The isolated SR99 and SR104 mAbswere of the IgG1 subclass, and SR116 mAb was of IgG2a subclass.
Immunocytometry analysis
Freshly isolated cells (1–2 � 106) were stained with combinations of PE-labeled MICA or MICB tetramers and directly conjugated mAbs. Cellswere first incubated at 22°C for 90 min in the dark with MICA or MICBtetramers at concentration equivalent to 10 �g/ml of MICA protein. Thefollowing Abs were added directly for 20 min at 4°C: CD4 FITC, CD16FITC, �� TCR FITC, CD69 Cy5, and CCR7 FITC were from BD PharM-ingen; CD8 APC, CD45RA Tricolor, CD45RO FITC, CD19 Tricolor, andCD62L FITC were from Caltag (Burlingame, CA). Cells were fixed in PBSwith 1% paraformaldehyde, and immediately analyzed on a FACSCaliburflow cytometer.
For cell surface staining of MICA molecules, cells were incubated firstwith 10 �l of aggregated human IgG (10 mg/ml) for 15 min at 4°C to maskFc�R, and then with 10 �g/ml SR104 mAb, or with Ig isotype-matchedcontrol Ab, at 4°C for 30 min, washed, and stained with FITC-labeled goatanti-mouse IgG. Cells were then analyzed on a BD Biosciences FACS-Calibur flow cytometer (Mountain View, CA), and data were obtainedusing CellQuest software. The fine specificity of the mAbs was determinedby blocking the staining of Hela cells by preincubation with a 10-foldexcess of recombinant soluble MICA or MICB protein for 1 h at 4°C.
1910 NKG2D/MICA INTERACTION AND MATURATION OF CD8 THYMOCYTES
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
Thymocyte cultures
CD8�-purified thymocytes were cultured for 1–5 days in 24-well plates(Falcon; BD Biosciences) at 1 � 106 cells/well in 1 ml RMPI 1640 sup-plemented with 10% human pooled AB serum and the following cytokines,either alone or in combination: IFN-� (100 U), rIL-2 (100 U/ml), IL-7 (10ng/ml), IL-15 (50 ng/ml) (Sigma-Aldrich). Thymocytes were then har-vested and assayed for cell surface markers by flow cytometry, as describedabove.
Quantitative PCR of TCR� rearrangement excision circles(TREC)
CD8-purified thymocytes were stained with PE-labeled MICA tetramers,and positively selected by anti-PE microbeads (Miltenyi Biotec). The num-ber of TRECs was determined by quantitative real-time PCR using the5�-nuclease (TaqMan) assay in an ABI7700 system (Applied Biosystems,Foster City, CA), as previously described (26). In brief, the sortedNKG2D-positive and NKG2D-negative CD8 thymocytes and total PBLswere lysed in 100 �g/ml proteinase K for 3 h at 56°C and then 20 min at95°C. Each 25-�l reaction contained 5 �l cell lysate, and the final con-centration of each component was as follows: 1.0� reaction buffer, 20U/ml platinum Taq polymerase (Life Technologies), 3.5 mM MgCl2, 0.2mM dNTPs, 500 nM each primer, and 150 nM probe. PCR conditions were95°C for 5 min, followed by 40 cycles of 95°C for 30 s and 60°C for 1 min.A standard curve was plotted, and TREC values for samples were calcu-lated using ABI7700 software. Samples were analyzed in duplicate.
Immunohistochemical analysis
Normal human thymuses were obtained as discard tissues from healthychildren during corrective cardiac surgery (see above). In addition, thymusspecimens were obtained from patients with thymoma who underwent cur-ative thymectomy. Thymus samples were immersed in OCT compound(TissueTek; Bayer, Pittsburgh, PA) and snap frozen in liquid nitrogen.Cryosections were cut 5 �m thin onto slides. Sections were air dried for1 h, fixed in ice-cold methanol for 1 h, and stored at �20°C. Doubleimmunostaining was then performed using FITC-labeled MICA tetramers(100 �l at 50 �g/ml overnight at 4°C), followed by anti-CD8 mAb, 30 minat 22°C (DAKO, Carpenteria, CA), plus rhodamine-labeled rabbit anti-ratIgG. In parallel, staining was performed with the anti-MICA SR99 mAbplus FITC-labeled goat anti-mouse IgG, and rhodamine-labeled human anti-pan-cytokeratin Ab. Isotype-matched Ig was used as control.
Alternatively, normal or pathologic thymic tissues were fixed in 10%Formalin for 4 h, washed twice with PBS, and immersed in 70% ethanol.Serial sections of the Formalin-fixed paraffin-embedded thymic tissueblocks were prepared. After deparaffinization and rehydration, sectionswere heated in citrate buffer for 5 min in a microwave, then treated with40% normal human serum for 20 min at room temperature, and incubatedwith a titrated solution of anti-MICA mAb for 1 h, or isotype-matchedcontrol Ig. Ab binding was visualized by using biotinylated goat anti-mouse Ig (DAKO). The peroxidase EnVision System (DAKO) was used tocomplete staining, followed by counterstain with hematoxylin.
Detection of soluble MICA by ELISA
To detect soluble MICA in the serum, two different anti-MIC mAbs wereused in a sandwich ELISA. High-binding polystyrene plates (Greiner;Sigma-Aldrich) were coated with the capture SR99 Ab (5 �g/ml in PBS,100 �l per well) for 12 h at 4°C, washed five times with PBS plus 0.05%Tween 20, blocked by addition of 100 �l of 5% BSA for 1 h at 22°C, andwashed in PBS-0.05% Tween 20. The standard (serial dilutions of solublerMICA from 100 �g/ml to 0.1 ng/ml in PBS-0.05% Tween 20) and theserum samples (100 �l per well) were then added for 2 h at room temper-ature. After five washes, the biotinylated SR104 detection Ab (150 ng/mlin PBS, 100 �l per well) was added for 1 h at room temperature. Plateswere washed and incubated for 20 min at room temperature with strepta-vidin-conjugated HRP (1/30,000; Amersham Pharmacia Biotech), washed,and reacted with tetramethylbenzidine substrate (Sigma-Aldrich) for 15min at 37°C. Reaction was stopped with 0.5 M H2SO4 (50 �l per well).The absorbance was measured at 450 nm. Sera from 20 healthy individualsand 48 patients with thymoma were analyzed.
ResultsBinding characteristics of MICA and MICB tetramers on PBLs
To study the interaction of MICA and MICB molecules with theirreceptor NKG2D, we generated fluorescently labeled tetramericcomplexes of the extracellular domain of MICA and MICB. Sol-
uble forms of the MICA and MICB protein were produced in in-sect cells and purified. MIC proteins migrated on SDS-PAGE inposition consistent with the expected molecular size for the gly-cosylated form of the soluble protein (43 kDa).
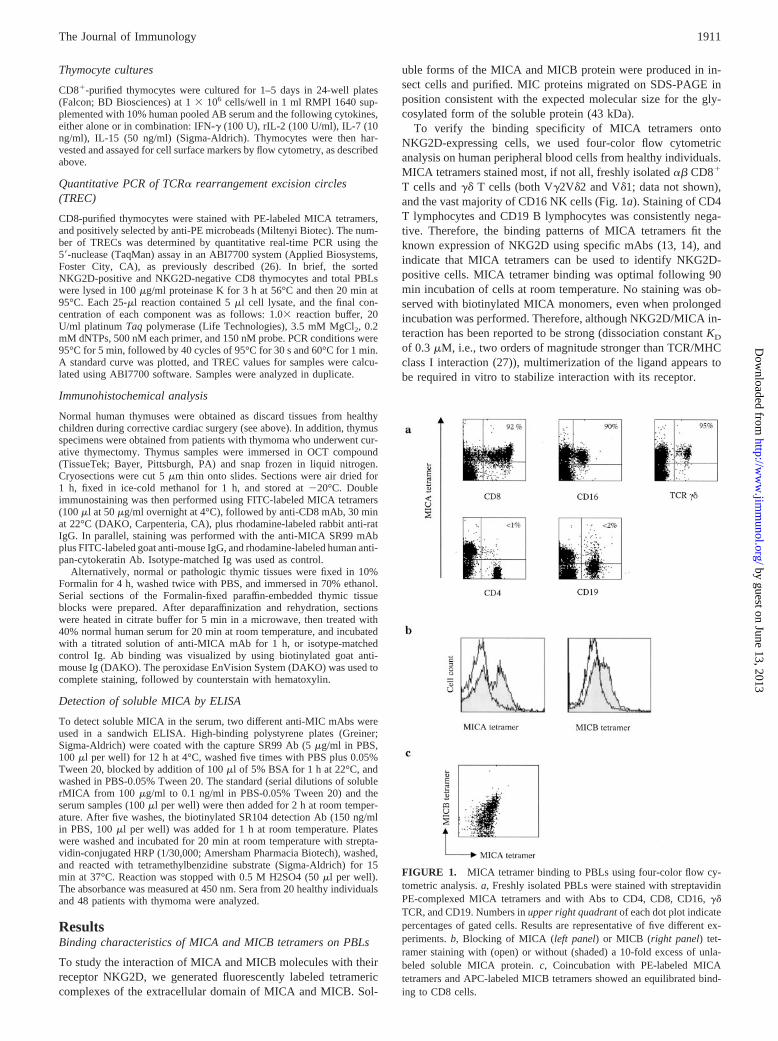
To verify the binding specificity of MICA tetramers ontoNKG2D-expressing cells, we used four-color flow cytometricanalysis on human peripheral blood cells from healthy individuals.MICA tetramers stained most, if not all, freshly isolated �� CD8�
T cells and �� T cells (both V�2V�2 and V�1; data not shown),and the vast majority of CD16 NK cells (Fig. 1a). Staining of CD4T lymphocytes and CD19 B lymphocytes was consistently nega-tive. Therefore, the binding patterns of MICA tetramers fit theknown expression of NKG2D using specific mAbs (13, 14), andindicate that MICA tetramers can be used to identify NKG2D-positive cells. MICA tetramer binding was optimal following 90min incubation of cells at room temperature. No staining was ob-served with biotinylated MICA monomers, even when prolongedincubation was performed. Therefore, although NKG2D/MICA in-teraction has been reported to be strong (dissociation constant KD
of 0.3 �M, i.e., two orders of magnitude stronger than TCR/MHCclass I interaction (27)), multimerization of the ligand appears tobe required in vitro to stabilize interaction with its receptor.
FIGURE 1. MICA tetramer binding to PBLs using four-color flow cy-tometric analysis. a, Freshly isolated PBLs were stained with streptavidinPE-complexed MICA tetramers and with Abs to CD4, CD8, CD16, ��TCR, and CD19. Numbers in upper right quadrant of each dot plot indicatepercentages of gated cells. Results are representative of five different ex-periments. b, Blocking of MICA (left panel) or MICB (right panel) tet-ramer staining with (open) or without (shaded) a 10-fold excess of unla-beled soluble MICA protein. c, Coincubation with PE-labeled MICAtetramers and APC-labeled MICB tetramers showed an equilibrated bind-ing to CD8 cells.
1911The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
Similar results were observed when staining peripheral bloodcells with MICB instead of MICA tetramers. MICA or MICB tet-ramer binding was completely inhibited by previous incubation ofcells with a 10-fold excess of soluble MICA protein (Fig. 1b). Inaddition, when cells were coincubated with PE-labeled MICA tet-ramers and APC-labeled MICB tetramers, equilibrated bindingwas observed, suggesting that MICA and MICB bind to the same(or overlapping) epitopes on their NKG2D receptor (Fig. 1c). Al-together, these results demonstrate that soluble MICA and MICBbind to NKG2D in a specific and apparently high-affinity interac-tion, and this binding of MIC molecules to NKG2D does not re-quire additional components for stability. This is in agreement withcrystallographic data showing that the MICA groove is not occu-pied by anything other than water (27).
Murine and human NKG2D ectodomains are 69% identical.Therefore, we tested whether MICA or MICB tetramers were ableto bind NKG2D on murine splenocytes from 6- to 8-wk-old
BALB/c mice. MIC tetramers did not stain CD8 T cells, while asignificant staining was observed following 3-day incubation ofsplenocytes with 100 U/ml rIL-2 (data not shown). These resultsconfirm that NKG2D is not expressed on resting, but only acti-vated T lymphocytes in mice (28), and also indicate that MICmolecules are able to bind to the murine NKG2D receptor, sug-gesting that NKG2D may use different epitopes to recognize itsvarious human or mouse ligands.
Expression of NKG2D, the receptor of MICA and MICB, onhuman thymocytes
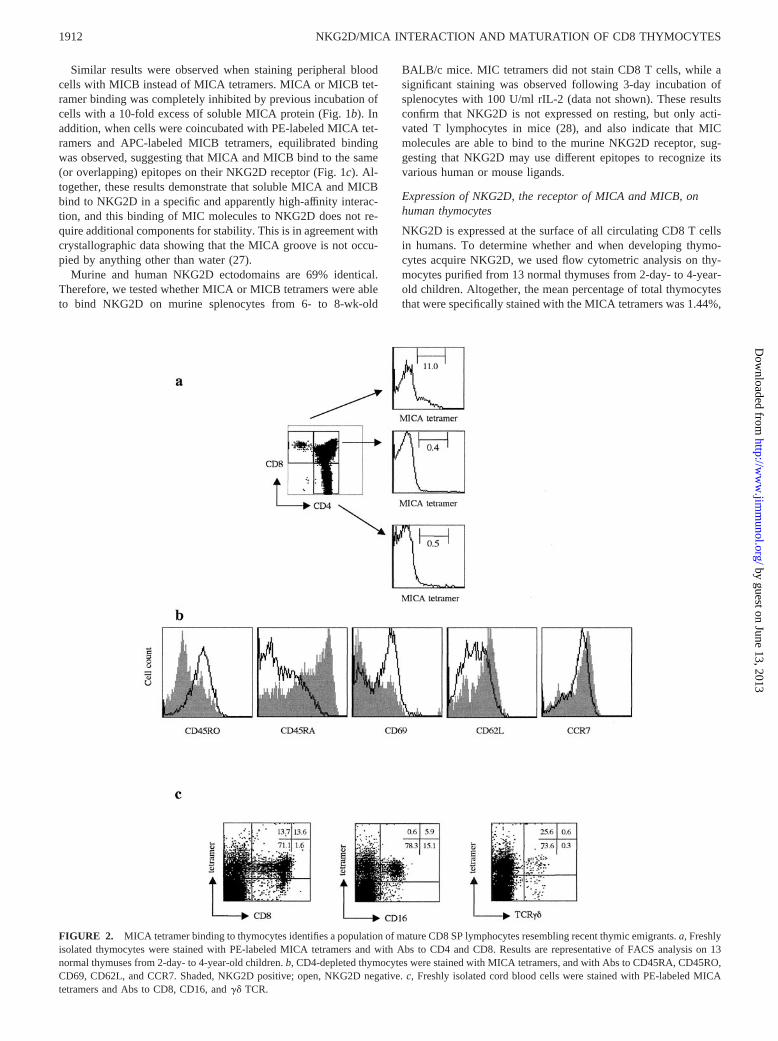
NKG2D is expressed at the surface of all circulating CD8 T cellsin humans. To determine whether and when developing thymo-cytes acquire NKG2D, we used flow cytometric analysis on thy-mocytes purified from 13 normal thymuses from 2-day- to 4-year-old children. Altogether, the mean percentage of total thymocytesthat were specifically stained with the MICA tetramers was 1.44%,
FIGURE 2. MICA tetramer binding to thymocytes identifies a population of mature CD8 SP lymphocytes resembling recent thymic emigrants. a, Freshlyisolated thymocytes were stained with PE-labeled MICA tetramers and with Abs to CD4 and CD8. Results are representative of FACS analysis on 13normal thymuses from 2-day- to 4-year-old children. b, CD4-depleted thymocytes were stained with MICA tetramers, and with Abs to CD45RA, CD45RO,CD69, CD62L, and CCR7. Shaded, NKG2D positive; open, NKG2D negative. c, Freshly isolated cord blood cells were stained with PE-labeled MICAtetramers and Abs to CD8, CD16, and �� TCR.
1912 NKG2D/MICA INTERACTION AND MATURATION OF CD8 THYMOCYTES
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

ranging from 0.85 to 2.65%. Phenotypical analysis of NKG2Dexpression was conducted on different thymocyte stages accordingto expression of the CD4 and CD8 markers. Representative resultsare depicted in Fig. 2a. Following the intrathymic lineage relation-ships, we did not detect NKG2D� thymocytes in CD4�CD8�
double-negative cells, or CD4�CD8� double-positive thymocytesubpopulation. Interestingly, MICA tetramer staining was clearlyrestricted to a small subset of thymocytes that were almost entirelyCD3 bright, CD4�, and CD8�. When quantified within this subsetof SP CD8 thymocytes, MICA tetramer-positive cells representeda mean of 11% of these cells. This percentage slightly varied ac-cording to the thymus analyzed (range 4.2–14.8%, median 11.9%),but did not appear to be developmentally regulated, because nocorrelation was observed with the age thymus was removed (be-tween day 2 to year 4 after birth). Results were strictly similarwhen using MICB instead of MICA tetramers. Among cells com-mitted to the CD8� lineage, NKG2D expression was not detectedin CD4intCD8� cells or in CD4lowCD8� cells, but was only ob-served in CD4�CD8� thymocytes. Thus, it appears that NKG2Dexpression is not stochastic, but is acquired as a postpositive se-lection event.
CD4�CD8� thymocytes comprise developmentally heteroge-neous subsets of cells. Some of them have not yet reached the stageat which functional competence is acquired and need to undergopostpositive selection differentiation before their migration to theperiphery (5). We investigated whether the NKG2D-expressingCD8� thymocytes were cells ready to be released from the thymusafter finishing their maturation. CD4-depleted thymocytes wereanalyzed for NKG2D expression together with expression of dif-ferent thymocyte maturation markers. The results (Fig. 2b) showedthat NKG2D� cells were all CD28� (data not shown), CD45RA�,CD45RO�, and CCR7�, and predominantly CD62Lhigh, CD69low,a phenotype compatible with that of fully mature CD8� cells readyto leave the thymus as naive cells. Altogether, 70% of NKG2D-positive CD8 thymocytes were CD62L� and CD69�. The sameresults were obtained when similar staining assays were done withtotal thymocytes. To confirm that this subset of NKG2D-positiveCD8 SP thymocytes represents a late stage of thymopoiesis andnot a subset of peripheral memory T cells that recirculate to thymicmedulla, we quantified TRECs in NKG2D-negative and NKG2D-positive thymocytes compared with PBLs. TRECs represent epi-somal DNA by-products of the TCR rearrangement process thatoccurs in �70% of thymocytes destined to become mature ��TCR T cells. TRECs are stable, do not replicate during mitosis,and are therefore diluted out during cell divisions. High levels ofTRECs were detected in the same proportion in NKG2D-positiveand NKG2D-negative CD8 thymocytes (�5,000/10,000 cells, i.e.,the same order of magnitude as previously reported in CD8 SPthymocytes (29)). This TREC content was 10 times higher thanthat of peripheral blood T cells (�500/10,000 cells), confirmingthat the CD8�NKG2D� subset of thymocytes does represent a latestage of thymopoiesis and not a subset of mature peripheral T cellsthat recirculate to the thymic medulla.
Because human cord blood T cells are composed almost exclu-sively of naive T cells and are representative of recent thymicemigrants (30), we also analyzed NKG2D expression on cordblood cells. Flow cytometric analysis confirmed that �95% ofCD8� T cells in unstimulated cord blood carried the naiveCD45RA� CD62L� phenotype, and showed that 90% of CD8 Tcells (as well as 90% of NK and 67% of �� TCR cells) werestained with MICA tetramer (Fig. 2c).
Altogether, these results indicate that acquisition of NKG2Dcharacterizes a percentage of mature CD8 SP thymocytes ready toemigrate to the periphery.
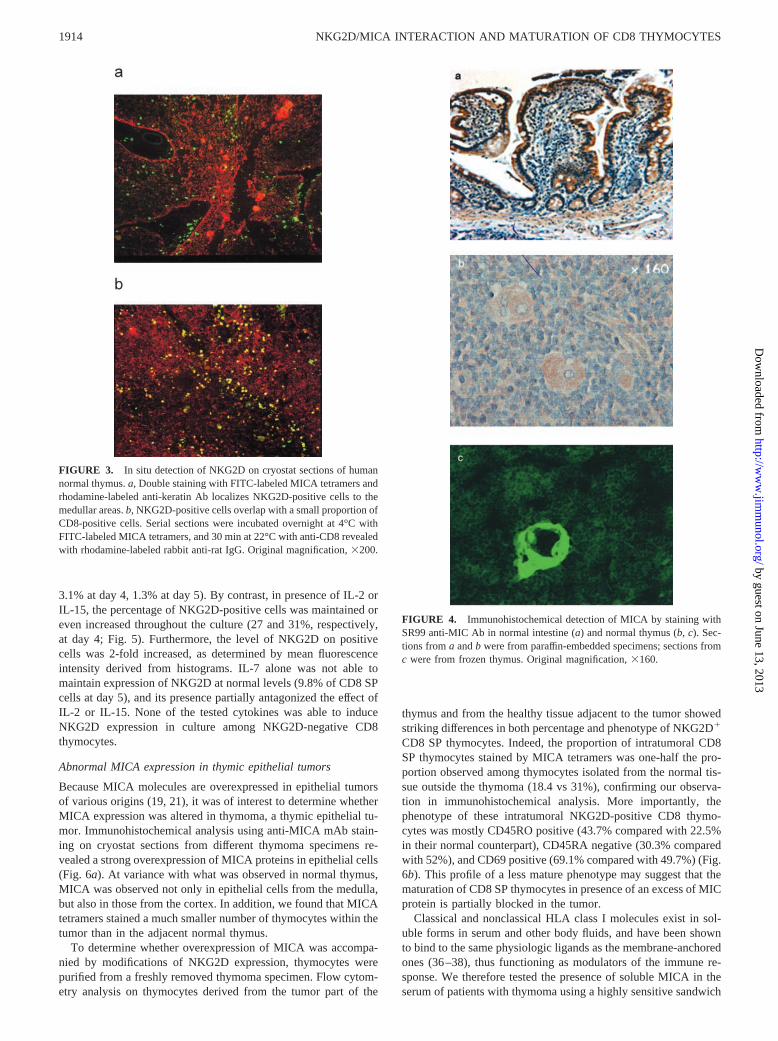
Immunolocalization of NKG2D in human thymus to medullar SPCD8� thymocytes
Mature thymocytes are localized in the thymic medulla, wherethey are supposed to spend up to 2 wk before emigration to theperiphery. To provide additional information on the nature andlocalization of NKG2D� cells within the thymus, immunohisto-chemical analyses using FITC-labeled MICA tetramers were per-formed on thymic cryostat sections from normal thymuses.NKG2D-expressing cells were all scattered throughout the me-dulla region, in the vicinity of blood vessels, whereas cortical areasshowed no staining (Fig. 3a). Double immunostaining with anti-CD8 mAb confirmed that NKG2D-expressing cells overlappedwith a small percentage of CD8� cells in the thymic medulla (Fig.3b). These results demonstrate that NKG2D-expressing thymo-cytes represent a fraction of mature CD4�CD8� SP present in situin the thymic medulla only.
Expression of MICA and MICB in the human thymus
To establish whether the acquisition of the NKG2D molecules onlate stage thymocytes is related to the presence of their ligand inthe thymus, we analyzed intrathymic MICA expression in the thy-mus specimens. To this aim, anti-MICA mAbs were produced byimmunizing mice with soluble rMICA. Three mAbs (SR99,SR104, and SR116) were selected for their ability to react withMICA molecules on the surface of HT-29, HeLa, and CaCo2 ep-ithelial cell lines by flow cytometry and with denatured rMICAmolecules in ELISA. SR104 recognized MICA molecules only,while SR99 and SR116 mAbs recognized both MICA and MICB(data not shown). MIC molecules are highly polymorphic, with�60 alleles described to date (31–33). Therefore, there is a pos-sibility that mAbs could be raised against allele-specific determi-nants, because the MICA*001 variant was used for immunizingmice. However, because of their reactivity with several lines ex-pressing different alleles, such as HT-29 (MICA*004/016), HeLa(MICA*008), and CaCo2 (MICA*033) (34), our mAbs were likelyto recognize a nonpolymorphic determinant on the MICA protein.In peripheral tissues, MICA molecules have been observed at thesurface of gut epithelium (9). Indeed, as shown in Fig. 4a, theSR99 mAb significantly stained epithelial cells in normal gutsections.
To decipher the potential role of MICA in the acquisition ofNKG2D maturation on SP CD8� thymocytes, we examined thelocalization of MICA molecules on paraffin-embedded or frozenthymus sections (Fig. 4, b and c). MICA proteins were exclusivelydetected in the thymic medulla, and were located in cells formingthe outer walls of Hassal’s corpuscles and in cells morphologicallyresembling mesenchymal cells (35). A very faint staining was ob-served in epithelial cells of the medulla, but not in cortical epithe-lial cells, endothelial cells, dendritic cells, or thymocytes. The ep-ithelial nature of MICA-expressing cells in the thymus wasconfirmed by double immunostaining for MICA and cytokeratin(data not shown). These results indicate that MICA molecules inthe thymic medulla are located within close proximity to matureCD8� SP thymocytes.
Acquisition of NKG2D on human thymocytes
We wanted to determine whether cytokines affect the acquisitionof NKG2D on late stage CD8 thymocytes. Total or purified SPCD8 thymocytes were cultured for 1–5 days in presence of IFN-�,IL-2, IL-15, or IL-7 either alone or in conjunction, and were an-alyzed for expression of NKG2D by flow cytometry. Surprisingly,in absence of any cytokine, NKG2D-positive cells progressivelydisappeared within 5 days (17% of CD8 SP thymocytes at day 0,
1913The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
3.1% at day 4, 1.3% at day 5). By contrast, in presence of IL-2 orIL-15, the percentage of NKG2D-positive cells was maintained oreven increased throughout the culture (27 and 31%, respectively,at day 4; Fig. 5). Furthermore, the level of NKG2D on positivecells was 2-fold increased, as determined by mean fluorescenceintensity derived from histograms. IL-7 alone was not able tomaintain expression of NKG2D at normal levels (9.8% of CD8 SPcells at day 5), and its presence partially antagonized the effect ofIL-2 or IL-15. None of the tested cytokines was able to induceNKG2D expression in culture among NKG2D-negative CD8thymocytes.
Abnormal MICA expression in thymic epithelial tumors
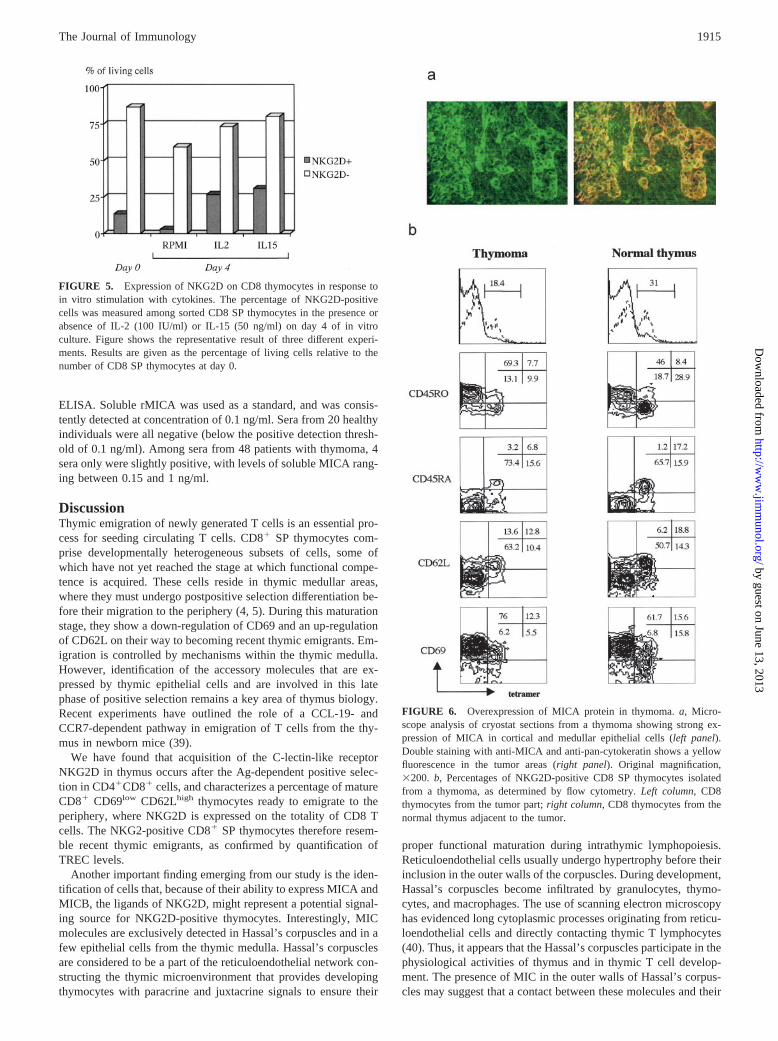
Because MICA molecules are overexpressed in epithelial tumorsof various origins (19, 21), it was of interest to determine whetherMICA expression was altered in thymoma, a thymic epithelial tu-mor. Immunohistochemical analysis using anti-MICA mAb stain-ing on cryostat sections from different thymoma specimens re-vealed a strong overexpression of MICA proteins in epithelial cells(Fig. 6a). At variance with what was observed in normal thymus,MICA was observed not only in epithelial cells from the medulla,but also in those from the cortex. In addition, we found that MICAtetramers stained a much smaller number of thymocytes within thetumor than in the adjacent normal thymus.
To determine whether overexpression of MICA was accompa-nied by modifications of NKG2D expression, thymocytes werepurified from a freshly removed thymoma specimen. Flow cytom-etry analysis on thymocytes derived from the tumor part of the
thymus and from the healthy tissue adjacent to the tumor showedstriking differences in both percentage and phenotype of NKG2D�
CD8 SP thymocytes. Indeed, the proportion of intratumoral CD8SP thymocytes stained by MICA tetramers was one-half the pro-portion observed among thymocytes isolated from the normal tis-sue outside the thymoma (18.4 vs 31%), confirming our observa-tion in immunohistochemical analysis. More importantly, thephenotype of these intratumoral NKG2D-positive CD8 thymo-cytes was mostly CD45RO positive (43.7% compared with 22.5%in their normal counterpart), CD45RA negative (30.3% comparedwith 52%), and CD69 positive (69.1% compared with 49.7%) (Fig.6b). This profile of a less mature phenotype may suggest that thematuration of CD8 SP thymocytes in presence of an excess of MICprotein is partially blocked in the tumor.
Classical and nonclassical HLA class I molecules exist in sol-uble forms in serum and other body fluids, and have been shownto bind to the same physiologic ligands as the membrane-anchoredones (36–38), thus functioning as modulators of the immune re-sponse. We therefore tested the presence of soluble MICA in theserum of patients with thymoma using a highly sensitive sandwich
FIGURE 3. In situ detection of NKG2D on cryostat sections of humannormal thymus. a, Double staining with FITC-labeled MICA tetramers andrhodamine-labeled anti-keratin Ab localizes NKG2D-positive cells to themedullar areas. b, NKG2D-positive cells overlap with a small proportion ofCD8-positive cells. Serial sections were incubated overnight at 4°C withFITC-labeled MICA tetramers, and 30 min at 22°C with anti-CD8 revealedwith rhodamine-labeled rabbit anti-rat IgG. Original magnification, �200.
FIGURE 4. Immunohistochemical detection of MICA by staining withSR99 anti-MIC Ab in normal intestine (a) and normal thymus (b, c). Sec-tions from a and b were from paraffin-embedded specimens; sections fromc were from frozen thymus. Original magnification, �160.
1914 NKG2D/MICA INTERACTION AND MATURATION OF CD8 THYMOCYTES
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
ELISA. Soluble rMICA was used as a standard, and was consis-tently detected at concentration of 0.1 ng/ml. Sera from 20 healthyindividuals were all negative (below the positive detection thresh-old of 0.1 ng/ml). Among sera from 48 patients with thymoma, 4sera only were slightly positive, with levels of soluble MICA rang-ing between 0.15 and 1 ng/ml.
DiscussionThymic emigration of newly generated T cells is an essential pro-cess for seeding circulating T cells. CD8� SP thymocytes com-prise developmentally heterogeneous subsets of cells, some ofwhich have not yet reached the stage at which functional compe-tence is acquired. These cells reside in thymic medullar areas,where they must undergo postpositive selection differentiation be-fore their migration to the periphery (4, 5). During this maturationstage, they show a down-regulation of CD69 and an up-regulationof CD62L on their way to becoming recent thymic emigrants. Em-igration is controlled by mechanisms within the thymic medulla.However, identification of the accessory molecules that are ex-pressed by thymic epithelial cells and are involved in this latephase of positive selection remains a key area of thymus biology.Recent experiments have outlined the role of a CCL-19- andCCR7-dependent pathway in emigration of T cells from the thy-mus in newborn mice (39).
We have found that acquisition of the C-lectin-like receptorNKG2D in thymus occurs after the Ag-dependent positive selec-tion in CD4�CD8� cells, and characterizes a percentage of matureCD8� CD69low CD62Lhigh thymocytes ready to emigrate to theperiphery, where NKG2D is expressed on the totality of CD8 Tcells. The NKG2-positive CD8� SP thymocytes therefore resem-ble recent thymic emigrants, as confirmed by quantification ofTREC levels.
Another important finding emerging from our study is the iden-tification of cells that, because of their ability to express MICA andMICB, the ligands of NKG2D, might represent a potential signal-ing source for NKG2D-positive thymocytes. Interestingly, MICmolecules are exclusively detected in Hassal’s corpuscles and in afew epithelial cells from the thymic medulla. Hassal’s corpusclesare considered to be a part of the reticuloendothelial network con-structing the thymic microenvironment that provides developingthymocytes with paracrine and juxtacrine signals to ensure their
proper functional maturation during intrathymic lymphopoiesis.Reticuloendothelial cells usually undergo hypertrophy before theirinclusion in the outer walls of the corpuscles. During development,Hassal’s corpuscles become infiltrated by granulocytes, thymo-cytes, and macrophages. The use of scanning electron microscopyhas evidenced long cytoplasmic processes originating from reticu-loendothelial cells and directly contacting thymic T lymphocytes(40). Thus, it appears that the Hassal’s corpuscles participate in thephysiological activities of thymus and in thymic T cell develop-ment. The presence of MIC in the outer walls of Hassal’s corpus-cles may suggest that a contact between these molecules and their
FIGURE 5. Expression of NKG2D on CD8 thymocytes in response toin vitro stimulation with cytokines. The percentage of NKG2D-positivecells was measured among sorted CD8 SP thymocytes in the presence orabsence of IL-2 (100 IU/ml) or IL-15 (50 ng/ml) on day 4 of in vitroculture. Figure shows the representative result of three different experi-ments. Results are given as the percentage of living cells relative to thenumber of CD8 SP thymocytes at day 0.
FIGURE 6. Overexpression of MICA protein in thymoma. a, Micro-scope analysis of cryostat sections from a thymoma showing strong ex-pression of MICA in cortical and medullar epithelial cells (left panel).Double staining with anti-MICA and anti-pan-cytokeratin shows a yellowfluorescence in the tumor areas (right panel). Original magnification,�200. b, Percentages of NKG2D-positive CD8 SP thymocytes isolatedfrom a thymoma, as determined by flow cytometry. Left column, CD8thymocytes from the tumor part; right column, CD8 thymocytes from thenormal thymus adjacent to the tumor.
1915The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

receptor NKG2D is of functional importance for the final matura-tion and export of SP CD8 thymocytes. In addition, it has beenpostulated that Hassal’s corpuscles play an important role in re-moval and disintegration of dying cells within the thymus. In thiscontext, MIC-expressing Hassal’s corpuscles might also reflect thehigh turnover of medullar thymic epithelial cells because of sig-naling provided by the interaction with NKG2D-positive thymo-cytes. Other thymus-expressed receptor/ligand pairs are implicatedin the control of thymocyte migration. The CCR9/CCL25 interac-tion is involved in the intrathymic trafficking of both �� TCR�
and �� TCR� T cells (41), while CCR7-CCL19 interaction playsa role in the emigration of newly matured T lymphocytes (39).
In addition to MIC proteins, several other NKG2D ligands havebeen identified. These include in the human the UL16-binding pro-teins (ULBP) (12, 42), and in the mouse the retinoic acid-inducibleearly genes protein family and the H-60 minor histocompatibilityAg (28, 43). Rodents lack any recognizable homologues of theMIC proteins, but murine NKG2D ligands RAE and H60, likeULBPs, are homologous to the platform domains of the MHC classI proteins, and are anchored in the membrane by GPI linkages.Although it is unclear which NKG2D ligands are more physiolog-ically important, it is likely that different members of the ULBPand MIC families have distinct biological roles (44). ULBPmRNAs have been detected in thymus (12), but protein expressionhas not been analyzed. Crystallographic analyses of NKG2D haverevealed a surprising degree of ligand plasticity, as it can accom-modate the platform domains of the distantly related MIC andULBP class Ib molecules. In addition, ULBP1 and 2 are capable ofbinding to mouse NK cells (42), but conversely, human NKG2Ddoes not bind to retinoic acid-inducible early genes (45). It hasbeen predicted that murine NKG2D will bind MICB (46), andindeed we could observe a binding of MIC tetramers to murineIL-2-activated splenocytes.
How CD8 SP thymocytes acquire NKG2D remains unclear.CD8� TCR ��� jejunal intraepithelial lymphocytes (IEL) expressNKG2D, but at levels consistently lower than those found on pe-ripheral blood T cells (47). Because MICA is mostly expressed ingut epithelium, many IELs are continuously exposed to MICA andthus, conceptually, are in state of alert. Therefore, it is not surpris-ing that IELs express low levels of NKG2D, which is probablydown-modulated to prevent chronic T cell stimulation and to limitautoreactive bystander T cell activation (48). Expression ofNKG2D on IELs is, however, restored to levels similar to those inPBMCs by exposure to IL-15, a cytokine secreted by intestinalepithelial cells and induced upon inflammation and viral infec-tions. IL-15 exerts stimulatory effects similar to those of IL-2 andshares the same receptor. In NK cells, stimulation by IL-15 hasalso been shown to up-regulate NKG2D expression (42, 44). In-deed, we observed that both IL-15 and IL-2 not only allowedNKG2D-positive thymocytes to survive in vitro, but also increasedthe levels of NKG2D molecules on CD8 SP thymocytes.
Human thymoma is a thymic epithelial cell tumor that oftencontains a large number of lymphocytes. Although thymomas havethe capacity to generate mature T cells, intratumoral T cell matu-ration is abnormal in that the mature CD45RA� T cell subset isdecreased compared with normal thymus (49). Furthermore, ma-ture intratumoral T cells often have an increased autoantigen-spe-cific potential toward fragments of the � subunit of acetylcholinereceptor, compared with T cells from normal thymus (50). Thisexplains why thymoma is occasionally associated with autoim-mune myasthenia gravis. It has been proposed that thymomas maygenerate autoantigen-specific T cells by a process of abnormal pos-itive or negative T cell selection. Such T cells would leave thethymoma and contribute to the peripheral autoimmune cascade
that leads to myasthenic symptoms of muscle weakness, followinginteraction between thymoma-derived autoantigen-specific T cellsand autoantibody-producing B cells that reside entirely outside thetumor (51, 52). Because various epithelial tumors are known toconsiderably up-regulate MICA, it was particularly informative tohave access to thymomas to analyze MICA expression and theconsequences on NKG2D acquisition. We observed a strikingoverexpression of MICA in thymic epithelial cells in thymoma.Surprisingly, this overexpression was accompanied by a decreasedproportion of NKG2D-positive cells among SP CD8 thymocytescompared with that in the healthy thymus. This observation mir-rors that in normal gut, where strong expression of MICA on gutepithelium is associated with down-modulation of NKG2D onIELs, and may suggest a defect in the maturation events associatedwith the tumoral process. Indeed, intratumoral NKG2D-positiveCD8� thymocytes did not express a fully mature phenotype, asshown by the persistence of CD69 and the presence of CD45RO.An increased expression of MICA on epithelial cells might beaccompanied by the release of soluble MICA shedded from tumorcell death. At the time the present work was completed, Groh et al.(53) showed that soluble MIC is released by shedding from MIC-positive tumors, and causes down-regulation of NKG2D at thesurface of tumor-specific effector T cells. Salih et al. (54) simul-taneouly reported the presence of soluble MIC in the serum ofpatients with gastrointestinal tumors. Although we observed thepresence of very low levels of soluble MIC in only a minority ofour patients, it is conceivable that soluble MIC is locally releasedfrom thymoma in the vicinity of thymocytes, leading to a down-regulation of NKG2D and a blockade of CD8 thymocytes in animmature phenotype. Such blockade might prevent T cell export ofmature CD8 SP thymocytes. Together with events triggering de-regulation of cytokine networks, MICA might therefore representa pathogenicity factor, which could exacerbate autoimmune con-ditions. Our results are in the same line of thinking as recent datathat suggest that the maturation of the CD8� lineage might beaffected in a subset of thymomas, leading to some thymopoieticincompetence (55). Alternatively, we cannot exclude the possibil-ity that part of these NKG2D-positive cells represent effectormemory T cells infiltrating the tumoral tissue.
In summary, we have identified the NKG2D� CD8 SP thymo-cyte population as a thymus-dependent late stage in CD8 T celldevelopment. Mature CD8 SP T cells are able to seed the peripheryonce they have acquired the NKG2D� phenotype. Inability to up-regulate NKG2D expression on thymocytes might cause them toremain in the thymus, as suggested by results in thymoma. Ourdata bring new lights in the nature of the thymic cross talk betweenthe thymic medulla and SP thymocytes.
AcknowledgmentsWe thank Dr. Sophie Ezine for critical reading of the manuscript, and Dr.Salima Hacein-Bey for determination of TREC levels.
References1. Von Boehmer, H., H. S. Teh, and P. Kisielow. 1989. The thymus selects the
useful, neglects the useless and destroys the harmful. Immunol. Today 10:57.2. Robey, E., and B. J. Fowlkes. 1994. Selective events in T cell development.
Annu. Rev. Immunol. 12:675.3. Anderson, G., and E. J. Jenkinson. 2001. Lymphostromal interactions in thymic
development and function. Nat. Rev. Immunol. 1:31.4. Anderson, G., K. J. Hare, and E. J. Jenkinson. 1999. Positive selection of thy-
mocytes: the long and winding road. Immunol. Today 20:463.5. Hare, K. J., E. J. Jenkinson, and G. Anderson. 1999. CD69 expression discrim-
inates MHC-dependent and -independent stages of thymocyte positive selection.J. Immunol. 162:3978.
6. Penit, C., and F. Vasseur. 1997. Expansion of mature thymocyte subsets beforeemigration to the periphery. J. Immunol. 159:4848.
1916 NKG2D/MICA INTERACTION AND MATURATION OF CD8 THYMOCYTES
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

7. Wu, J., Y. Song, A. B. Bakker, S. Bauer, T. Spies, L. L. Lanier, and J. H. Phillips.1999. An activating immunoreceptor complex formed by NKG2D and DAP10.Science 285:730.
8. Bahram, S., M. Bresnahan, D. E. Geraghty, and T. Spies. 1994. A second lineageof mammalian major histocompatibility complex class I genes. Proc. Natl. Acad.Sci. USA 91:6259.
9. Groh, V., S. Bahram, S. Bauer, A. Herman, M. Beauchamp, and T. Spies. 1996.Cell stress-regulated human major histocompatibility complex class I gene ex-pressed in gastrointestinal epithelium. Proc. Natl. Acad. Sci. USA 93:12445.
10. Bahram, S. 2000. MIC genes: from genetics to biology. Adv. Immunol. 76:1.11. Vivier, E., E. Tomasello, and P. Paul. 2002. Lymphocyte activation via NKG2D:
towards a new paradigm in immune recognition? Curr. Opin. Immunol. 14:306.12. Cosman, D., J. Mullberg, C. L. Sutherland, W. Chin, R. Armitage, W. Fanslow,
M. Kubin, and N. J. Chalupny. 2001. ULBPs, novel MHC class I-related mole-cules, bind to CMV glycoprotein UL16 and stimulate NK cytotoxicity throughthe NKG2D receptor. Immunity 14:123.
13. Bauer, S., V. Groh, J. Wu, A. Steinle, J. H. Phillips, L. L. Lanier, and T. Spies.1999. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science 285:727.
14. Pende, D., C. Cantoni, P. Rivera, M. Vitale, R. Castriconi, S. Marcenaro,M. Nanni, R. Biassoni, C. Bottino, A. Moretta, and L. Moretta. 2001. Role ofNKG2D in tumor cell lysis mediated by human NK cells: cooperation with nat-ural cytotoxicity receptors and capability of recognizing tumors of nonepithelialorigin. Eur. J. Immunol. 31:1076.
15. Groh, V., R. Rhinehart, J. Randolph-Habecker, M. S. Topp, S. R. Riddell, andT. Spies. 2001. Costimulation of CD8�� T cells by NKG2D via engagement byMIC induced on virus-infected cells. Nat. Immun. 2:255.
16. Diefenbach, A., E. Tomasello, M. Lucas, A. M. Jamieson, J. K. Hsia, E. Vivier,and D. H. Raulet. 2002. Selective associations with signaling proteins determinestimulatory versus costimulatory activity of NKG2D. Nat. Immun. 3:1142.
17. Gilfillan, S., E. L. Ho, M. Cella, W. M. Yokoyama, and M. Colonna. 2002.NKG2D recruits two distinct adapters to trigger NK cell activation and costimu-lation. Nat. Immun. 3:1150.
18. Groh, V., A. Steinle, S. Bauer, and T. Spies. 1998. Recognition of stress-inducedMHC molecules by intestinal epithelial �� T cells. Science 279:1737.
19. Groh, V., R. Rhinehart, H. Secrist, S. Bauer, K. H. Grabstein, and T. Spies. 1999.Broad tumor-associated expression and recognition by tumor-derived �� T cellsof MICA and MICB. Proc. Natl. Acad. Sci. USA 96:6879.
20. Wu, J., V. Groh, and T. Spies. 2002. T cell antigen receptor engagement andspecificity in the recognition of stress-inducible MHC class I-related chains byhuman epithelial �� T cells. J. Immunol. 169:1236.
21. Vetter, C. S., V. Groh, P. thor Straten, T. Spies, E. B. Brocker, and J. C. Becker.2002. Expression of stress-induced MHC class I related chain molecules on hu-man melanoma. J. Invest. Dermatol. 118:600.
22. Das, H., V. Groh, C. Kuijl, M. Sugita, C. T. Morita, T. Spies, and J. F. Bukowski.2001. MICA engagement by human V�2V�2 T cells enhances their antigen-dependent effector function. Immunity 15:83.
23. Tieng, V., C. Le Bouguenec, L. du Merle, P. Bertheau, P. Desreumaux, A. Janin,D. Charron, and A. Toubert. 2002. Binding of Escherichia coli adhesin AfaE toCD55 triggers cell-surface expression of the MHC class I-related moleculeMICA. Proc. Natl. Acad. Sci. USA 99:2977.
24. Schatz, P. J. 1993. Use of peptide libraries to map the substrate specificity of apeptide-modifying enzyme: a 13 residue consensus peptide specifies biotinylationin Escherichia coli. Bio/Technology 11:1138.
25. Kearney, J. F., A. Radbruch, B. Liesegang, and K. Rajewsky. 1979. A new mousemyeloma cell line that has lost immunoglobulin expression but permits the con-struction of antibody-secreting hybrid cell lines. J. Immunol. 123:1548.
26. Douek, D. C., R. A. Vescio, M. R. Betts, J. M. Brenchley, B. J. Hill, L. Zhang,J. R. Berenson, R. H. Collins, and R. A. Koup. 2000. Assessment of thymicoutput in adults after hematopoietic stem-cell transplantation and prediction ofT-cell reconstitution. Lancet 355:1875.
27. Li, P., D. L. Morris, B. E. Willcox, A. Steinle, T. Spies, and R. K. Strong. 2001.Complex structure of the activating immunoreceptor NKG2D and its MHC classI-like ligand MICA. Nat. Immun. 2:443.
28. Diefenbach, A., A. M. Jamieson, S. D. Liu, N. Shastri, and D. H. Raulet. 2000.Ligands for the murine NKG2D receptor: expression by tumor cells and activa-tion of NK cells and macrophages. Nat. Immun. 1:119.
29. Okamoto, Y., D. C. Douek, R. D. McFarland, and R. A. Koup. 2002. Effects ofexogenous interleukin-7 on human thymus function. Blood 99:2851.
30. Hassan, J., and D. J. Reen. 2001. Human recent thymic emigrants: identification,expansion, and survival characteristics. J. Immunol. 167:1970.
31. Fodil, N., P. Pellet, L. Laloux, G. Hauptmann, I. Theodorou, and S. Bahram.1999. MICA haplotypic diversity. Immunogenetics 49:557.
32. Pellet, P., M. Renaud, N. Fodil, L. Laloux, H. Inoko, G. Hauptmann, P. Debre,S. Bahram, and I. Theodorou. 1997. Allelic repertoire of the human MICB gene.Immunogenetics 46:434.
33. Robinson, J., M. Perez-Rodriguez, M. J. Waller, B. Cuillerier, S. Bahram, Z. Yao,E. D. Albert, J. A. Madrigal, and S. G. Marsh. 2001. MICA sequences 2000.Immunogenetics 53:150.
34. Zhang, Y., A. M. Lazaro, B. Lavingia, and P. Stastny. 2001. Typing for all knownMICA alleles by group-specific PCR and SSOP. Hum. Immunol. 62:620.
35. Suniara, R. K., E. J. Jenkinson, and J. J. Owen. 2000. An essential role for thymicmesenchyme in early T cell development. J. Exp. Med. 191:1051.
36. Puppo, F., P. Contini, M. Ghio, S. Brenci, M. Scudeletti, G. Filaci, S. Ferrone,and F. Indiveri. 2000. Soluble human MHC class I molecules induce soluble Fasligand secretion and trigger apoptosis in activated CD8� Fas (CD95)� T lym-phocytes. Int. Immunol. 12:195.
37. Spaggiari, G. M., P. Contini, R. Carosio, M. Arvigo, M. Ghio, D. Oddone,A. Dondero, M. R. Zocchi, F. Puppo, F. Indiveri, and A. Poggi. 2002. SolubleHLA class I molecules induce natural killer cell apoptosis through the engage-ment of CD8: evidence for a negative regulation exerted by members of theinhibitory receptor superfamily. Blood 99:1706.
38. Fournel, S., M. Aguerre-Girr, X. Huc, F. Lenfant, A. Alam, A. Toubert,A. Bensussan, and P. Le Bouteiller. 2000. Cutting edge: soluble HLA-G1 triggersCD95/CD95 ligand-mediated apoptosis in activated CD8� cells by interactingwith CD8. J. Immunol. 164:6100.
39. Ueno, T., K. Hara, M. S. Willis, M. A. Malin, U. E. Hopken, D. H. Gray,K. Matsushima, M. Lipp, T. A. Springer, R. L. Boyd, et al. 2002. Role for CCR7ligands in the emigration of newly generated T lymphocytes from the neonatalthymus. Immunity 16:205.
40. Bodey, B., B. Bodey, Jr., S. E. Siegel, and H. E. Kaiser. 2000. Novel insights intothe function of the thymic Hassall’s bodies. In Vivo 14:407.
41. Uehara, S., K. Song, J. M. Farber, and P. E. Love. 2002. Characterization ofCCR9 expression and CCL25/thymus-expressed chemokine responsiveness dur-ing T cell development: CD3highCD69� thymocytes and ��TCR� thymocytespreferentially respond to CCL25. J. Immunol. 168:134.
42. Kubin, M., L. Cassiano, J. Chalupny, W. Chin, D. Cosman, W. Fanslow,J. Mullberg, A. M. Rousseau, D. Ulrich, and R. Armitage. 2001. ULBP1, 2, 3:novel MHC class I-related molecules that bind to human cytomegalovirus gly-coprotein UL16, activate NK cells. Eur. J. Immunol. 31:1428.
43. Cerwenka, A., A. B. Bakker, T. McClanahan, J. Wagner, J. Wu, J. H. Phillips,and L. L. Lanier. 2000. Retinoic acid early inducible genes define a ligand familyfor the activating NKG2D receptor in mice. Immunity 12:721.
44. Sutherland, C. L., N. J. Chalupny, and D. Cosman. 2001. The UL16-bindingproteins, a novel family of MHC class I-related ligands for NKG2D, activatenatural killer cell functions. Immunol. Rev. 181:185.
45. Strong, R. K. 2002. Asymmetric ligand recognition by the activating natural killercell receptor NKG2D, a symmetric homodimer. Mol. Immunol. 38:1029.
46. Li, P., G. McDermott, and R. K. Strong. 2002. Crystal structures of RAE-1� andits complex with the activating immunoreceptor NKG2D. Immunity 16:77.
47. Roberts, A. I., L. Lee, E. Schwarz, V. Groh, T. Spies, E. C. Ebert, and B. Jabri.2001. NKG2D receptors induced by IL-15 costimulate CD28-negative effectorCTL in the tissue microenvironment. J. Immunol. 167:5527.
48. Spies, T. 2002. Induction of T cell alertness by bacterial colonization of intestinalepithelium. Proc. Natl. Acad. Sci. USA 99:2584.
49. Nenninger, R., A. Schultz, V. Hoffacker, M. Helmreich, A. Wilisch,B. Vandekerckhove, T. Hunig, B. Schalke, C. Schneider, S. Tzartos, et al. 1998.Abnormal thymocyte development and generation of autoreactive T cells inmixed and cortical thymomas. Lab. Invest. 78:743.
50. Zhang, G. X., B. G. Xiao, M. Bakhiet, P. van der Meide, H. Wigzell, H. Link, andT. Olsson. 1996. Both CD4� and CD8� T cells are essential to induce experi-mental autoimmune myasthenia gravis. J. Exp. Med. 184:349.
51. Muller-Hermelink, H. K., and A. Marx. 2000. Thymoma. Curr. Opin. Oncol.12:426.
52. Drachman, D. B. 1994. Myasthenia gravis. N. Engl. J. Med. 330:1797.53. Groh, V., J. Wu, C. Yee, and T. Spies. 2002. Tumor-derived soluble MIC ligands
impair expression of NKG2D and T-cell activation. Nature 419:734.54. Salih, H. R., H. G. Rammensee, and A. Steinle. 2002. Cutting edge: down-reg-
ulation of MICA on human tumors by proteolytic shedding. J. Immunol. 169:4098.
55. Strobel, P., M. Helmreich, G. Menioudakis, S. R. Lewin, T. Rudiger, A. Bauer,V. Hoffacker, R. Gold, W. Nix, B. Schalke, et al. 2002. Paraneoplastic myasthe-nia gravis correlates with generation of mature naive CD4� T cells in thymomas.Blood 100:159.
1917The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from




















