
Scavenger receptor class-A has a central role in cerebral ischemia–reperfusion injury
Upload
independentCategory
view
2download
0
Ž .Matrix Biology 19 2000 61]71
Selective adhesion of macrophages to denatured forms oftype I collagen is mediated by scavenger receptors
Brian B. Gowena, Thomas K. Borgb, Abdul Ghaffar a, Eugene P. Mayer a,U
aDepartment of Microbiology and Immunology, Uni ersity of South Carolina, School of Medicine, Columbia, SC 29208, USAbDepartment of De¨elopmental Biology and Anatomy, Uni ersity of South Carolina, School of Medicine, Columbia, USA
Accepted 13 October 1999
Abstract
Ž .Macrophages Mfs are multifunctional immune cells which are involved in the regulation of immune and inflammatoryresponses, as well as in tissue repair and remodeling. In tissues, Mfs reside in areas which are rich in extracellular matrixŽ .ECM , the structural component which also plays an essential role in regulating a variety of cellular functions. A major ECMprotein encountered by Mfs is type I collagen, the most abundant of the fibril-forming collagens. In this study, the adhesionof RAW 264.7 murine Mfs to native fibrillar, monomeric, and denatured type I collagen was investigated. Using atomic forcemicroscopy, structural differences between fibrillar and monomeric type I collagen were clearly resolved. When cultured onfibrillar type I collagen, Mfs adhered poorly. In contrast, they adhered significantly to monomeric, heat-denatured, orcollagenase-modified type I collagen. Studies utilizing anti-b1 and -b2 integrin adhesion-blocking antibodies, RGD-containingpeptides, or divalent cation-free conditions did not inhibit Mf adhesion to monomeric or denatured type I collagen. However,
Ž .macrophage scavenger receptor MSR ligands and anti-MSR antibodies significantly blocked Mf adhesion to denatured andmonomeric type I collagen strongly suggesting the involvement of the MSR as an adhesion molecule for denatured type Icollagen. Further analysis by Western blot identified the MSR as the primary receptor for denatured type I collagen amongMf proteins purified from a heat-denatured type I collagen affinity column. These findings indicate that Mfs adhereselectively to denatured forms of type I collagen, but not the native fibrillar conformation, via their scavenger receptors.Q 2000 Elsevier Science B.V.rInternational Society of Matrix Biology. All rights reserved.
Keywords: Cell adhesion; Collagen; Macrophage scavenger receptor
1. Introduction
Monocytes, recruited from peripheral blood to sitesof infection and inflammation, migrate through thevascular endothelium and into various tissues via acomplex series of interactions with the extracellular
Ž .matrix ECM . Once in the underlying tissues, mono-cytes differentiate and develop into macrophages
U Tel.: q1-803-733-3281; fax: q1-803-733-3192.Ž .E-mail address: [email protected] E.P. Mayer
Ž .Mfs , which are essential to proper immune func-Žtion and wound resolution Duerksen-Hughes and
Gooding, 1993; Barbul and Regan, 1995; DiPietro,.1995 . However, the infiltration and accumulation of
Mfs at inflammatory sites can also be detrimental incertain cases. In atherosclerosis, Mfs are believed toplay a central role in the development and progres-sion of the disease, which is characterized as a chronicinflammatory disorder where the persistent presence
Žof Mfs is evident at all stages Gown et al., 1986;.Ross, 1993 .
Resident Mfs, which reside in areas rich in ECM
0945-053Xr00r$ - see front matter Q 2000 Elsevier Science B.V.rInternational Society of Matrix Biology. All rights reserved.Ž .PII: S 0 9 4 5 - 0 5 3 X 9 9 0 0 0 5 2 - 9

( )B.B. Gowen et al. r Matrix Biology 19 2000 61]7162
constituents, are constantly exposed to collagens, fi-bronectin, and a variety of proteoglycans. In mam-malian tissues, in excess of 20 distinct collagens have
Ž .been described Kadler et al., 1996 . Type I collagencomprises the large majority of the total body colla-gen including )60% of the total dry weight of der-
Ž .mal tissue Thomas et al., 1995 . It is a fibril-formingcollagen which polymerizes with other collagenmolecules to generate fibrils and fibers under physio-
Ž .logical conditions Kadler et al., 1996 . Type I, as wellŽ .as type III collagen also a fibril-forming collagen ,
Žare major components of atherosclerotic lesions Mc-.Cullagh et al., 1980; Rekhter et al., 1993 .
Extracellular matrix damage and denaturation canresult from a variety of insults including thermalinjury, trauma, tumor invasion, and chronic inflam-
Ž .mation. During acute wound and chronicŽ .atherosclerosis inflammation, local ECM proteinsare exposed to an environment laden with proteolyticenzymes which can degrade and alter their native
Žconformation Weiss, 1989; Jasin and Taurog, 1991;.Sukhova et al., 1999 . Previous studies have shown
that certain recognition sequences become accessibleto cell-surface receptors, the integrins, only when
Žpresented in a denatured form Davis, 1992a,b; Davis.et al., 1997; Messent et al., 1998 . Other modifications
to ECM proteins such as oxidation and non-enzymaticglycation occur in arterial basement membranes as a
Žresult of aging andror vascular injury or disease El.Khoury et al., 1994; Mattana et al., 1995 . Moreover,
Ž .the resulting advanced glycation end products AGEaccumulate at an accelerated rate in diabetics, whichare at high risk for developing atherosclerosisŽ .Brownlee et al., 1988 . Macrophage scavenger recep-
Ž .tors MSR , have been shown to mediate adhesion toŽ .AGE-modified matrix El Khoury et al., 1994 , sug-
gesting that these receptors may also function toaugment the accumulation of Mfs in the arterialintima where these modifications occur. These studiessuggest that modified matrix proteins can influencecell localization and function. It is unclear as towhether Mfs can recognize and respond to differ-ences in damaged or denatured collagens which resultfrom thermal injury or increased proteolytic activity atsites of inflammation.
MSRs are trimeric integral membrane proteinswhich exhibit a remarkably broad ligand specificitywhich includes various polyanions, bacterial cell wallconstituents, crocidolite asbestos, b-amyloid fibrils,
Ž . Žand modified low density lipoproteins LDL Hamp-ton et al., 1991; Resnick et al., 1993; Krieger andHerz, 1994; Araki et al., 1995; Christie et al., 1996;
.Horiuchi et al., 1996; Paresce et al., 1996 . Interest-ingly, these receptors have also been shown to medi-
Žate cell adhesion to apoptotic thymocytes Platt et al.,. Ž1996; Terpstra et al., 1997 , B lymphocytes Yokota et
. Žal., 1998 , serum-coated tissue culture plastic Fraser. Žet al., 1993 , and AGE-modified type IV collagen El
.Khoury et al., 1994 . However, MSRs are best knownfor their role in the uptake of modified LDLs, impli-cating them in foam cell formation and atherogenesis.The ability of these receptors to recognize other ma-trix proteins has not been investigated.
The majority of studies which have investigatedmonocyterMf]ECM interactions have focused onthe role of matrix proteins in modulating cell func-
Žtions Bohnsack et al., 1985; Eierman et al., 1989;Newman and Tucci, 1990; McKay et al., 1992; Mc-
.Cutcheon et al., 1998 . Adhesion is a factor in thelocalization and retention of cells at specific sitesŽ .Ruoslahti and Obrink, 1996 . With the exception ofb integrin-mediated adhesion to laminin and fi-
Žbronectin Shaw et al., 1990; Pucillo et al., 1993;.Bauvois et al., 1996; Weston and Weeks, 1996 , very
little is known about how Mfs attach to other ECMproteins. Previous studies which have investigatedmonocyterMf and leukocyte adhesion to type I col-
Žlagen have had contrasting results Newman andTucci, 1990; Borel et al., 1993; Lin et al., 1995; Frieser
.et al., 1996 . The results are difficult to interpret sincevarious cell types and methods of coating collagenwere used and not well detailed. Moreover, the mech-anism of adhesion to type I collagen was not ad-dressed nor was the binding capacity of Mfs todenatured vs. native type I fibrillar collagen examined.
In this study, the adhesion of Mfs to native fibril-lar, monomeric, and denatured type I collagens wasinvestigated. Furthermore, the mechanism by whichMfs bind to denatured forms of type I collagen wasalso addressed. The data presented strongly suggestthat MSRs, not the integrins, are the primary recep-tors for denatured type I collagen present on Mfs.
2. Materials and methods
2.1. Antibodies and reagents
Ž . Ž .The anti-b1 Ha2r5 and anti-b2 GAME-46 inte-Ž .grin monoclonal antibodies mAb s, as well as the
isotype-matched control mAbs, were obtained fromŽ .Pharmingen San Diego, CA, USA . Monoclonal anti-
Ž .bodies directed against murine MSR clone 2F8 ,Žwere purchased from Chemicon Temecula, CA,
.USA . Secondary HRP-conjugated anti-rat antibodieswere obtained from Santa Cruz Biotechnology, IncŽ .Santa Cruz, CA, USA . Streptavidin-horseradish per-
Ž .oxidase conjugate SA-HRP was obtained fromŽ .Amersham Arlington Heights, IL, USA . Hexapep-
Ž .tides Gly]Arg]Gly]Asp]Thr]Pro GRGDTP ,Ž .Gly]Arg]Gly]Asp]Ser]Pro GRGDSP , and inac-
tive control peptide Gly]Arg]Gly]Glu]Ser]Pro

( )B.B. Gowen et al. r Matrix Biology 19 2000 61]71 63
Ž .GRGESP were obtained from American PeptideŽ .Company Sunnyvale, CA, USA . For the divalent
cation-free studies, HBSS devoid of Ca2q and Mg2q
Žand standard HBSS were purchased from Sigma St..Louis, MO, USA . Dextran sulfate, chondroitin sul-
Ž w x.fate, polyguanylic acid poly G , polycytidylic acidŽ w x. Ž w x.poly C , polyuridylic acid poly U , and polyinosinic
Ž w x.acid poly I were also obtained from Sigma.
2.2. Cells
The RAW 264.7 and J774.1 murine Mf cell lines,as well as the B16 murine melanoma cell line, wereobtained from the American Type Tissue CollectionŽ .Rockville, MD, USA and maintained in RPMI 1640medium supplemented with 10% fetal bovine serum,
Žpenicillin]streptomycin 100 IU penicillin and 100 mg.of streptomycin per ml , and 2 mM L-glutamine, all
Ž .obtained from Gibco BRL Grand Island, NY, USA .Male C3HrHeN mice were obtained from Harlan
Ž .Sprague Dawley Indianapolis, ID, USA at 4]6 weeksŽ .of age. Thioglycollate TG -elicited Mfs were ac-
quired from mice which had been injected i.p. with 1Ž . Žml of 4% wrv TG broth Difco Laboratories, De-
.troit, MI, USA 4 days before cells were harvested byperitoneal lavage. Typically )90% of the TG-elicitedperitoneal exudate cells were Mfs as determined byWright]Giemsa staining. All experiments were con-ducted in serum-free RPMI 1640 medium, at 378C,and 5% CO .2
2.3. Coating of collagen and laminin
Concentrated stocks of type I bovine skin collagenand Engelbreth-Holm-Swarm mouse tumor lamininŽ .Becton Dickinson, Bedford, MA, USA were diluted
Ž .in phosphate-buffered saline PBS to 50 mgrml and15 mgrml, respectively. The diluted proteins weredispensed, 100 mlrwell, into 96-well microtiter tissueculture plates and incubated overnight at 48C. Incertain experiments, type I collagen was also dilutedin 0.02 N glacial acetic acid to produce monomericcollagen matrices. Coated wells were routinely washedthree times with sterile PBS to remove any residualunbound ECM protein prior to the addition of cells.
( )2.4. Atomic Force Microscopy AFM
Fibrillar or monomeric type I collagen was coatedŽonto a mica surface Ted Pella, Inc., Redding, CA,
.USA and the unbound protein was removed by wash-ing three times with sterile distilled water. A Nanos-
Žcope Multimode Scanning Probe Microscope Digital.Instruments, Santa Barbara, CA, USA was utilized to
analyze coated type I collagen. Scanning of the colla-gen was done in tapping mode, in air. In AFM the
surface topography of a sample is imaged by moving afine stylus, mounted on a cantilever spring, over thesurface and recording the deflection of the stylus.This technique is capable of resolving a sample at the
Žatomic level Schabert et al., 1995; Muller et al.,.1997 .
2.5. Denaturation and detection of type I collagen
For heat-denaturation experiments, type I collagenand laminin were incubated at 4, 37, 60, 80, and1008C, for 30 min, prior to being coated onto 96-wellmicrotiter plates. Preliminary experiments with wellscoated with various doses of heat-denatured collagenand laminin indicated that complete coating was
Žachieved with as little as 10 mgrml of protein data.not shown . For collagenase digestions, wells were
first coated with fibrillar type I collagen prior totreatment with varying amounts of Clostridium hysto-
Ž .lytica collagenase Sigma, St. Louis, MO, USA . Theplates were allowed to incubate for 30 min at 378Cand washed four times with PBS prior to the additionof Mfs.
Detection of type I collagen after collagenase di-gestion was performed by a standard ELISA on aduplicate plate. Collagenase-treated plates werewashed four times with sterile distilled water and thenon-specific binding sites were blocked with 1% BSAin PBS prior to the addition of rabbit anti-bovine type
ŽI collagen antibodies Chemicon, Temecula, CA,.USA . After washing away unbound antibodies, alka-
line phosphatase-conjugated, goat anti-rabbit sec-Žondary antibodies Santa Cruz Biotechnology, Santa
.Cruz, CA, USA were added to each well. Afterremoving the unbound secondary antibodies, 75 ml of
Ž3 mM r-nitrophenyl phosphate Sigma, St. Louis,.MO, USA in 0.05 M NaCO and 0.05 mM MgCl3 2
was added to the wells. Color development was moni-tored visually and the reaction stopped by the addi-tion of 25 ml of 0.5 M NaOH. The plates were read at405 nm on a Titertek MultiSkan MCCr340 plate
Ž .reader Titertek Instruments, Huntsville, AL, USA .
2.6. Adhesion assay
Adhesion was assessed by measuring the level ofthe endogenous lysosomal enzyme, hexosaminidase,
Ž .as originally described Landegren, 1984 . Briefly, 7.5=104 cells were cultured on coated or uncoatedŽ .controls 96-well microtiter plate wells and incubatedfor 1 h. Unbound cells were removed by washing 3times with PBS prior to the addition of 50 mlrwell of
Žhexosaminidase substrate 3.75 mM r-nitrophenyl-N-acetyl-b-D-glucosaminide, 0.25% Triton X-100, 0.05
.M citrate buffer, pH 5 . After a 1]2-h incubationperiod, hexosaminidase activity was stopped and color

( )B.B. Gowen et al. r Matrix Biology 19 2000 61]7164
developed by the addition of 75 mlrwell of stoprde-Žvelopment buffer 5 mM EDTA, 50 mM glycine, pH
.10.4 . Plates were read at 405 nm on a TitertekMultiSkan microtiter plate reader. The number ofadherent cells was determined from a standard curveof Mfs cultured on uncoated tissue culture plasticŽ .TCP . Preliminary experiments showed that )90%of Mfs cultured on TCP adhered within 1 h. Allassays were done in replicates of four.
Inhibition of adhesion to denatured forms of type Icollagen with anti-b integrin adhesion-blocking anti-bodies, RGD-containing hexapeptides, MSR ligands,and anti-MSR mAbs was performed as follows. Cellswere preincubated with the specified antibodies,hexapeptides, or MSR ligands for 30 min, at 48C, andthen cultured on the coated collagens. Adhesion wasdetermined after a 1-h incubation period, also in thepresence of antibodies, hexapeptides, or MSR ligands.
2.7. Cell surface protein biotinylation and analysis
RAW 264.7 Mf cell surface membrane proteinswere biotinylated using EZ-Link Sulfo-NHS-LC-Bio-
Ž .tin Pierce, Rockford, IL, USA following the manu-facturers specifications. Heat-denatured and fibrillartype I collagens were immobilized onto cyanogen
Žbromide activated-sepharose 4B Sigma, St. Louis,.MO, USA following the manufacturers specifications
and utilized for affinity chromatography. All stepswere performed at 48C. Biotinylated cells were lysedwith 50 mM Tris]HCl, pH 7.5, containing 1 mMCaCl , 1 mM MgCl , 40 mM octylthioglucoside2 2Ž .OTG , 0.5 mM PMSF, 10 mgrml aprotonin, and 1mgrml pepstatin A. Insoluble material was removedby centrifugation at 12 000=g for 15 min. The super-natant was slowly allowed to enter the heat-denatured
type I collagen-sepharose 4B affinity column. After a15-min incubation period, the column was washedwith 5 column volumes of equilibration buffer while
Ž .monitoring the absorbancy at 280 A280 nm with aŽBeckman DU-50 Series spectrophotometer Irvine,
.CA, USA . The column was eluted with 0.1 M glycine,0.5 M NaCl, pH 3, 1 mM CaCl , 1 mM MgCl , 1 mM2 2MnCl , and 20 mM OTG, and the fractions were2immediately neutralized with 1 M Tris, pH 8. TheA280 of fractions was monitored until baseline wasreached and the fractions containing eluted proteinswere pooled and concentrated. The eluted proteinswere separated under reducing and non-reducing con-
Žditions on 4]15% precast polyacrylamide gels Bio-.Rad, Hercules, CA, USA and analyzed by the pico
Western blot chemiluminescence detection systemŽ .Pierce, Rockford, IL, USA following the manufac-turers specifications. In control experiments, the su-pernatants were initially passed over fibrillar type Icollagen-sepharose 4B affinity columns and elutedwith 50 mM Tris-HCl, pH 7.5, containing 10 mMEDTA and 20 mM OTG. Fractions were collectedand analyzed as above. Detectable levels of proteincould not be eluted from the fibrillar type I collagencolumns. All affinity column experiments were per-formed at least twice and similar results were ob-tained.
3. Results
3.1. Adhesion of Mf s to fibrillar and monomeric formsof type I collagen
Investigations of cell]matrix interactions which in-clude type I collagen are generally conducted using
Ž . Ž .Fig. 1. Structure of coated fibrillar and monomeric type I collagen. A Fibrillar and B monomeric type I collagen were coated onto mica andŽ .analyzed by atomic force microscopy. Note the extensive network of type I collagen fibrils native conformation in panel A.
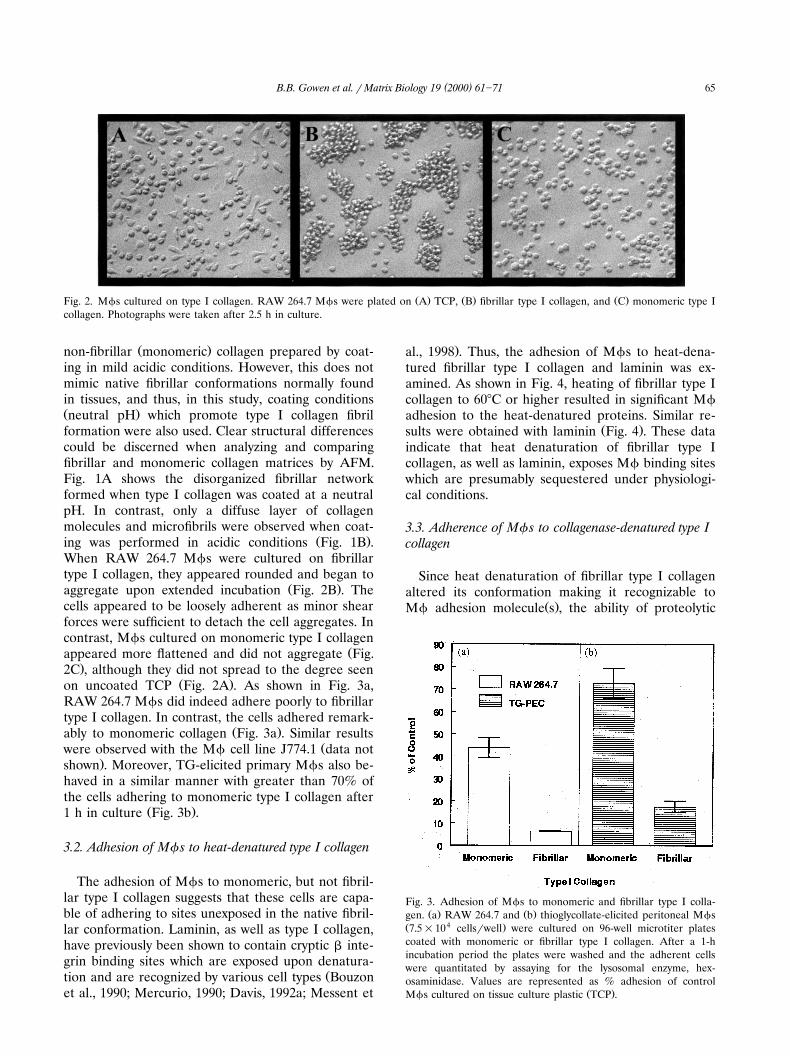
( )B.B. Gowen et al. r Matrix Biology 19 2000 61]71 65
Ž . Ž . Ž .Fig. 2. Mfs cultured on type I collagen. RAW 264.7 Mfs were plated on A TCP, B fibrillar type I collagen, and C monomeric type Icollagen. Photographs were taken after 2.5 h in culture.
Ž .non-fibrillar monomeric collagen prepared by coat-ing in mild acidic conditions. However, this does notmimic native fibrillar conformations normally foundin tissues, and thus, in this study, coating conditionsŽ .neutral pH which promote type I collagen fibrilformation were also used. Clear structural differencescould be discerned when analyzing and comparingfibrillar and monomeric collagen matrices by AFM.Fig. 1A shows the disorganized fibrillar networkformed when type I collagen was coated at a neutralpH. In contrast, only a diffuse layer of collagenmolecules and microfibrils were observed when coat-
Ž .ing was performed in acidic conditions Fig. 1B .When RAW 264.7 Mfs were cultured on fibrillartype I collagen, they appeared rounded and began to
Ž .aggregate upon extended incubation Fig. 2B . Thecells appeared to be loosely adherent as minor shearforces were sufficient to detach the cell aggregates. Incontrast, Mfs cultured on monomeric type I collagen
Žappeared more flattened and did not aggregate Fig..2C , although they did not spread to the degree seen
Ž .on uncoated TCP Fig. 2A . As shown in Fig. 3a,RAW 264.7 Mfs did indeed adhere poorly to fibrillartype I collagen. In contrast, the cells adhered remark-
Ž .ably to monomeric collagen Fig. 3a . Similar resultsŽwere observed with the Mf cell line J774.1 data not
.shown . Moreover, TG-elicited primary Mfs also be-haved in a similar manner with greater than 70% ofthe cells adhering to monomeric type I collagen after
Ž .1 h in culture Fig. 3b .
3.2. Adhesion of Mf s to heat-denatured type I collagen
The adhesion of Mfs to monomeric, but not fibril-lar type I collagen suggests that these cells are capa-ble of adhering to sites unexposed in the native fibril-lar conformation. Laminin, as well as type I collagen,have previously been shown to contain cryptic b inte-grin binding sites which are exposed upon denatura-
Žtion and are recognized by various cell types Bouzonet al., 1990; Mercurio, 1990; Davis, 1992a; Messent et
.al., 1998 . Thus, the adhesion of Mfs to heat-dena-tured fibrillar type I collagen and laminin was ex-amined. As shown in Fig. 4, heating of fibrillar type Icollagen to 608C or higher resulted in significant Mfadhesion to the heat-denatured proteins. Similar re-
Ž .sults were obtained with laminin Fig. 4 . These dataindicate that heat denaturation of fibrillar type Icollagen, as well as laminin, exposes Mf binding siteswhich are presumably sequestered under physiologi-cal conditions.
3.3. Adherence of Mf s to collagenase-denatured type Icollagen
Since heat denaturation of fibrillar type I collagenaltered its conformation making it recognizable to
Ž .Mf adhesion molecule s , the ability of proteolytic
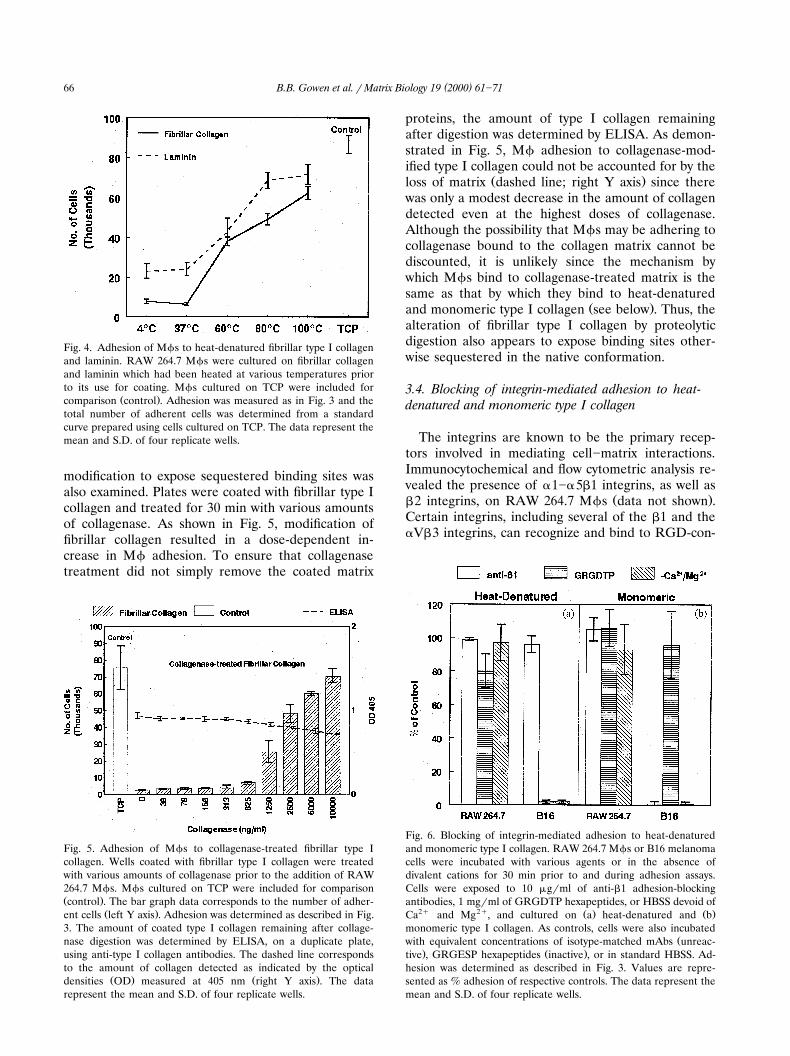
Fig. 3. Adhesion of Mfs to monomeric and fibrillar type I colla-Ž . Ž .gen. a RAW 264.7 and b thioglycollate-elicited peritoneal Mfs
Ž 4 .7.5=10 cellsrwell were cultured on 96-well microtiter platescoated with monomeric or fibrillar type I collagen. After a 1-hincubation period the plates were washed and the adherent cellswere quantitated by assaying for the lysosomal enzyme, hex-osaminidase. Values are represented as % adhesion of control
Ž .Mfs cultured on tissue culture plastic TCP .
( )B.B. Gowen et al. r Matrix Biology 19 2000 61]7166
Fig. 4. Adhesion of Mfs to heat-denatured fibrillar type I collagenand laminin. RAW 264.7 Mfs were cultured on fibrillar collagenand laminin which had been heated at various temperatures priorto its use for coating. Mfs cultured on TCP were included for
Ž .comparison control . Adhesion was measured as in Fig. 3 and thetotal number of adherent cells was determined from a standardcurve prepared using cells cultured on TCP. The data represent themean and S.D. of four replicate wells.
modification to expose sequestered binding sites wasalso examined. Plates were coated with fibrillar type Icollagen and treated for 30 min with various amountsof collagenase. As shown in Fig. 5, modification offibrillar collagen resulted in a dose-dependent in-crease in Mf adhesion. To ensure that collagenasetreatment did not simply remove the coated matrix
Fig. 5. Adhesion of Mfs to collagenase-treated fibrillar type Icollagen. Wells coated with fibrillar type I collagen were treatedwith various amounts of collagenase prior to the addition of RAW264.7 Mfs. Mfs cultured on TCP were included for comparisonŽ .control . The bar graph data corresponds to the number of adher-
Ž .ent cells left Y axis . Adhesion was determined as described in Fig.3. The amount of coated type I collagen remaining after collage-nase digestion was determined by ELISA, on a duplicate plate,using anti-type I collagen antibodies. The dashed line correspondsto the amount of collagen detected as indicated by the optical
Ž . Ž .densities OD measured at 405 nm right Y axis . The datarepresent the mean and S.D. of four replicate wells.
proteins, the amount of type I collagen remainingafter digestion was determined by ELISA. As demon-strated in Fig. 5, Mf adhesion to collagenase-mod-ified type I collagen could not be accounted for by the
Ž .loss of matrix dashed line; right Y axis since therewas only a modest decrease in the amount of collagendetected even at the highest doses of collagenase.Although the possibility that Mfs may be adhering tocollagenase bound to the collagen matrix cannot bediscounted, it is unlikely since the mechanism bywhich Mfs bind to collagenase-treated matrix is thesame as that by which they bind to heat-denatured
Ž .and monomeric type I collagen see below . Thus, thealteration of fibrillar type I collagen by proteolyticdigestion also appears to expose binding sites other-wise sequestered in the native conformation.
3.4. Blocking of integrin-mediated adhesion to heat-denatured and monomeric type I collagen
The integrins are known to be the primary recep-tors involved in mediating cell]matrix interactions.Immunocytochemical and flow cytometric analysis re-vealed the presence of a1]a5b1 integrins, as well as
Ž .b2 integrins, on RAW 264.7 Mfs data not shown .Certain integrins, including several of the b1 and theaVb3 integrins, can recognize and bind to RGD-con-
Fig. 6. Blocking of integrin-mediated adhesion to heat-denaturedand monomeric type I collagen. RAW 264.7 Mfs or B16 melanomacells were incubated with various agents or in the absence ofdivalent cations for 30 min prior to and during adhesion assays.Cells were exposed to 10 mgrml of anti-b1 adhesion-blockingantibodies, 1 mgrml of GRGDTP hexapeptides, or HBSS devoid of
2q 2q Ž . Ž .Ca and Mg , and cultured on a heat-denatured and bmonomeric type I collagen. As controls, cells were also incubated
Žwith equivalent concentrations of isotype-matched mAbs unreac-. Ž .tive , GRGESP hexapeptides inactive , or in standard HBSS. Ad-
hesion was determined as described in Fig. 3. Values are repre-sented as % adhesion of respective controls. The data represent themean and S.D. of four replicate wells.

( )B.B. Gowen et al. r Matrix Biology 19 2000 61]71 67
taining sequences found within matrix proteins suchŽas collagens and laminin Humphries, 1990; Davis,
.1992a; Ruoslahti et al., 1994; Messent et al., 1998 .Moreover, binding of integrins requires the presence
Ž .of divalent cations Ruoslahti and Obrink, 1996 . Thus,the involvement of integrins was investigated by wellestablished standard approaches which exploit inte-grin binding requirements, as well as treatment withanti-b1 and b2 integrin adhesion-blocking antibodies.As shown in Fig. 6, anti-b1 integrin blocking anti-
Ž .bodies, an RGD-containing hexapeptide GRGDTPpreviously shown to inhibit cellular adhesion to colla-
Žgens and other ECM proteins Dedhar et al., 1987;. 2qGehlsen et al., 1988 , or conditions devoid of Ca
and Mg2q failed to significantly inhibit RAW 264.7Mf adhesion to heat-denatured or monomeric type Icollagen. Furthermore, anti-b2 integrin antibodies, aswell as a different RGD-containing hexapeptideŽ .GRGDSP , were also unable to abrogate Mf adhe-
Ž .sion to the collagen matrices data not shown . ForŽ .comparison, melanoma cells B16 cell line , whose
binding to type I collagen has been shown to be bŽ .integrin-mediated Davis, 1992a , were used to verify
the effectiveness of the various treatments and condi-tions. As demonstrated in Fig. 6, only the adhesion ofthe B16 melanoma cells to monomeric type I collagenwas blocked with the anti-b1 integrin antibodies,
Žwhereas GRGDTP as well as GRGDSP, data not.shown exclusively inhibited the adhesion of cells to
heat-denatured type I collagen. These data are inaccordance with previous findings that showed thatmelanoma cell adhesion to heat-denatured type I
Ž .collagen is via the aVb3 integrin Davis, 1992a .Furthermore, the inactive control hexapeptide,GRGESP, had no effect on B16 attachment to heat-denatured collagen demonstrating the specificity of
Ž .the RGD-containing peptides data not shown . Lastly,adhesion experiments conducted in the absence ofCa2q and Mg2q completely blocked B16 melanomaadhesion to both monomeric and heat-denatured type
Ž .I collagen Fig. 6 . This finding is consistent with therequirement of divalent cations for integrin-mediatedbinding. Taken together, these data suggest that inte-grins previously shown to be involved in adhesion totype I collagen play a minor role, if any, in theadhesion of RAW 264.7 Mfs to heat-denatured ormonomeric type I collagen.
3.5 Blocking of Mf adhesion to denatured forms of typeI collagen with MSR ligands and anti-MSR antibodies
Since there was no evidence of integrin involve-ment in the observed adhesion of Mfs to denaturedforms of type I collagen and previous reports havedemonstrated the ability of the MSR to function as an
Žadhesion molecule Fraser et al., 1993; El Khoury et.al., 1994 , MSR ligands were used to try and block
adhesion to heat-denatured, collagenase-modified, andmonomeric type I collagen. The polyanion, dextransulfate, has been shown to effectively block MSR
Ž .activity Acton et al., 1993; El Khoury et al., 1994whereas chondroitin sulfate has no effect. As shownin Fig. 7a, dextran sulfate abrogated RAW 264.7 Mfadhesion to all forms of denatured type I collagenwhile chondroitin sulfate had little effect. Other MSR
w x w xligands, poly G , poly I , and fucoidan were also ableto block Mf binding to denatured collagens whereas
Ž w x w x.other control polyanions poly U and poly C did notŽ .data not shown . These findings provided indirectevidence that the MSR was involved in Mf adhesionto damaged or non-native type I collagen. To obtainmore specific evidence, anti-MSR 2F8 mAbs, origi-nally demonstrated to inhibit divalent cation indepen-dent Mf adhesion to serum coated-tissue culture
Ž .plastic Fraser et al., 1993 , were used to treat Mfs inadhesion-blocking studies. As demonstrated in Fig.7b, the 2F8 mAb specifically blocked RAW 264.7 Mfadhesion to all forms of denatured type I collagen.Isotype-matched control antibodies had very little ef-
Ž .fect on adhesion Fig. 7b . Taken together, these datademonstrate that the MSR can function as an adhe-sion molecule for heat-denatured, collagenase-di-gested, and monomeric type I collagen.
Fig. 7. Blocking of MSR-mediated adhesion to heat-denatured,collagenase-modified, and monomeric type I collagen. RAW 264.7
Ž . ŽMfs were incubated with the MSR ligand a dextran sulfate 100. Ž . Ž .mgrml and b MSR-specific mAb, 2F8 10 mgrml , for 30 min
prior to culturing on heat-denatured and monomeric type I colla-gen. Negative controls consisted of Mfs incubated with equivalent
Ž . Ž .concentrations of a chondroitin sulfate and b isotype-matchedmAbs. Adhesion was determined as described in Fig. 3. Values arerepresented as % adhesion of respective controls. The data repre-sent the mean and S.D. of four replicate wells. Chondroitin sulfateŽ . Ž . Ž .CS ; dextran sulfate DS ; heat-denatured H-D ; collagenase-
Ž . Ž .treated C-T ; monomeric Mono .

( )B.B. Gowen et al. r Matrix Biology 19 2000 61]7168
3.6 MSR affinity for immobilized heat-denatured type Icollagen
To verify the affinity of the MSR for denaturedtype I collagen, RAW 264.7 Mf biotinylated cellsurface membrane proteins were exposed to type Icollagen affinity chromatography matrices. To demon-strate specificity for heat-denatured type I collagen,proteins were initially passed over fibrillar type Icollagen columns. Detectable levels of proteins couldnot be eluted from these columns, indicating the lackof affinity of RAW 264.7 Mf membrane proteinstowards fibrillar type I collagen. This finding wasconsistent with the demonstrated poor adhesion ofMfs to the native fibrillar conformation of type I
Ž .collagen Fig. 3 . Proteins which bound to heat-dena-tured type I collagen were eluted and analyzed byWestern blot using anti-MSR mAbs, which onlyrecognize the intact trimeric receptor of reported Mrof 200]230 kDa. Fig. 8a shows the presence of a
Ž .single band at ;205 kDa lane 2 which is consistentwith the size of the MSR. In agreement with theknown reactivity of the 2F8 mAb, no bands werevisible in the lanes where proteins were separated
Ž .under reducing conditions lanes 1 and 3 . As ex-pected, the MSR was also readily detected in cell
Ž .extracts from TG-elicited peritoneal Mfs Fig. 8a .To investigate whether other proteins were also bind-ing to heat-denatured type I collagen, the elutedbiotinylated cell surface membrane proteins were alsoprobed with streptavidin-HRP to identify other candi-date receptors. Although an assortment of bands wereclearly resolved, the most prominent ones, by far,were of apparent M of ;75 kDa, under reducingrSDS-PAGE, and ;140 and ;205 kDa, under non-
Ž .reducing conditions Fig. 8b . Again, the apparentsizes of these bands are consistent with those re-ported for the murine MSR which is a trimeric inte-gral membrane protein composed of ;75 kDamonomers. Taken together, these data demonstratethe affinity of the MSR for heat-denatured type Icollagen.
4. Discussion
Adherence is an important determinant which fac-tors in the homing and retention of Mfs at sites ofinflammation. Previous studies have shown thatmonocyterMf adhesion is a gene inductive event,particularly for those genes involved in inflammatory
Žresponses Eierman et al., 1989; Sporn et al., 1990;.Haskill et al., 1992; Yurochko et al., 1992 . The aim of
this study was to assess the differential adhesion ofMfs to native fibrillar and non-native conformationsof type I collagen. We found that RAW 264.7 Mfs
Fig. 8. Western blot analysis of biotinylated cell surface membraneproteins eluted from heat-denatured type I collagen affinity column.RAW 264.7 Mf proteins with affinity for heat-denatured type Icollagen were fractionated, pooled, and concentrated prior to theirseparation by SDS-PAGE on 4]15% gradient gels. After separa-tion, proteins were transferred onto PVDF membranes for Western
Ž . Ž .blot analysis using a MSR-specific 2F8 mAbs and b HRP-con-Ž .jugated streptavidin. a Lanes 1 and 3; proteins separated in the
Ž .presence of 2-mercaptoethanol 2-ME, reducing conditions . Lanes2 and 4; proteins separated under non-reducing conditions. Lanes 1and 2; RAW 264.7 Mf proteins. Lanes 3 and 4; TG-elicited
Ž .peritoneal Mf cell extract. b RAW 264.7 Mf proteins separatedŽ .under reducing conditions Lane 1 and non-reducing conditions
Ž .Lane 2 .
adhered poorly to fibrillar type I collagen; however,when cultured on denatured conformations of type Icollagen the cells adhered remarkably well. Similarresults were observed when primary peritoneal MfsŽ . ŽFig. 3b , as well as the J774.1 Mf cell line data not
.shown , were cultured under the same conditions.These findings confirmed that the selective adhesionof Mfs to denatured or monomeric collagen was notan artifact of the RAW 264.7 cell line since normalMfs, and the J774.1 cell line, behaved similarly.
Previous reports which have studied adhesion ofleukocytes and other cells to denatured proteins andECM have implicated the b integrins as the primary
Žadhesion receptors Davis, 1992a,b; Davis et al., 1997;.Messent et al., 1998 . On melanoma cells, the aVb3
integrin is the major receptor for denatured type IŽ .collagen Davis, 1992a . The aVb3 integrin-mediated
adhesion is RGD-dependent, in contrast to b1 inte-grin-mediated adhesion to native collagen, which is
ŽRGD-independent Gullberg et al., 1989; Davis,. Ž1992a . Moreover, several b2 integrins Mac-1 and.p150,95 , as well as the a4b1 integrin, have all been
shown to mediate adhesion to denatured proteinsŽ .Davis, 1992b; Davis et al., 1997 . Despite previousfindings which suggested integrin involvement, wecould not significantly inhibit the adhesion of RAW264.7 Mfs to heat-denatured or non-fibrillarmonomeric type I collagen with anti-b1 or -b2 inte-

( )B.B. Gowen et al. r Matrix Biology 19 2000 61]71 69
grin adhesion-blocking antibodies, RGD-containingpeptides, or divalent cation-free conditions. These
Ž .results suggested that Mf receptor s , other thanthose integrins previously reported to function asreceptors for type I collagen and denatured proteins,may be responsible for mediating adhesion to type Icollagen.
Evidence that MSRs can also function as adhesionmolecules prompted us to investigate whether theywere involved in mediating RAW 264.7 Mf adhesionto denatured forms of type I collagen. Using MSRligands and specific anti-MSR mAbs, we identified theMSR as the major Mf receptor for non-native formsof type I collagen. Preliminary data suggest that theseMSRs can also recognize denatured forms of type III
Ž .collagen data not shown , another fibril-forming col-lagen commonly found in association with type I. In
Žcontrast, adhesion to type IV collagen a network-for-.ming collagen , which is a major component of base-
ment membranes, does not appear to be mediated byŽ .these receptors data not shown . Thus, MSRs may
function as receptors for denatured fibril-forming, butnot network-forming, types of collagens. The fact thatthese receptors have been shown to mediate adhesionto denatured, as well as AGE-modified matrix sug-gests that they may be involved in the homing andlocalization of Mfs to specific sites. Along with itsfunction as an adhesion molecule, the engagement ofthe MSR can also transmit signals to the cell. Indeed,studies have demonstrated the ability of MSRs toinfluence the release of growth factors and inflam-
Žmatory mediators Kirstein et al., 1990; Palkama,.1991 . Moreover, despite its broad ligand specificity,
there is evidence that the MSR can discriminatebetween different ligands and transmit ligand-specific
Žsignals to Mfs Paulnock, D., personal communica-.tion . We are currently investigating the functional
consequences of MSR binding to denatured type Icollagen.
Evidence that cell adhesion to wound sites plays animportant role in wound healing is supported by theaccelerated healing which occurs following experi-mental therapies involving the use of fibronectin orsticky synthetic substances, which may aid in the
Žlocalization of cells to these sites Clark, 1990; Ruos-.lahti, 1991 . Mfs are the predominant cell type found
in wounds during the inflammatory stage and havebeen shown to play a key role in the wound healing
Ž .process DiPietro, 1995 . Wound resolution involvesŽECM degradation, synthesis, and re-organization Bi-
rkedal-Hansen, 1995; Shingleton et al., 1996; Chin.and Werb, 1997 . Thus, ECM proteins, including type
I collagen, are digested by a variety of proteases,thereby altering their native conformation. Fragmentsof various ECM proteins have been shown to be
chemotactic for circulating leukocytes and the pres-ence of such fragments, as a result of injury, has been
Žpreviously reported Laskin et al., 1986; La Celle et.al., 1990 . In cutaneous wounds, Mfs are exposed to
large quantities of denatured type I collagen. Ourfindings that Mfs adhere to damaged or denaturedforms of type I collagen may help explain how Mfshome to and are retained at these sites until thedamage is repaired and new matrix is in place. More-over, the inability of Mfs to adhere to fibrillar colla-gen prevents the cells from adhering to native ornewly remodeled type I collagen, facilitating theirmigration through tissues and away from resolvedwounds. Thus, the ability of Mfs to differentiallyadhere to denatured but not native type I collagenmay be an important aspect in their role in woundhealing.
In addition to their role in acute inflammatoryresponses, Mfs are also linked to the development ofthe chronic inflammatory disease, atherosclerosisŽ .Gown et al., 1986; Ross, 1993 . Moreover, the MSRplays a central role in the accumulation of lipids andthe formation of foam cells which is characteristic ofthe disease. In atherosclerotic plaques, there is anabundance of type I collagen present. As lesionsprogress, the amount of type I collagen increases andthe ratio of type III to type I collagen decreasesŽMcCullagh, 1983; Rekhter et al., 1993; Rekhter,
.1999 . Increased interstitial collagenase activity hasbeen shown to occur at these sites which are continu-ously undergoing various degrees of matrix repair and
Žremodeling Galis et al., 1994, 1995; Sukhova et al.,.1999 . Our data indicate that the MSR can also
function as an adhesion molecule for damaged fibril-lar type I collagen. Therefore, the ability of Mfs to
Ž .recognize proteolytically modified denatured type Icollagen in atherosclerotic lesions via their MSRs mayfactor in the pathophysiology of the disease. Theincreased number of Mfs retained at these sites mayresult from their ability to adhere to denatured, aswell as AGE-modified type collagen via the MSR.This engagement may be involved in signaling therelease of proinflammatory cytokines and growth fac-tors, exacerbating the progression of the disease.
Acknowledgements
We would like to thank Digital Instruments for theuse of the Nanoscope Multimode Scanning ProbeMicroscope and Dr Irene Revenko for her experttechnical assistance. This study was partially sup-ported by an Interdisciplinary Grant from the SouthCarolina Cancer Center.

( )B.B. Gowen et al. r Matrix Biology 19 2000 61]7170
References
Acton, S., Resnick, D., Freeman, M., Ekkel, Y., Ashkenas, J.,Krieger, M., 1993. The collagenous domains of macrophagescavenger receptors and complement component C1q mediatetheir similar, but not identical, binding specificities for polyan-ionic ligands. J. Biol. Chem. 268, 3530]3537.
Araki, N., Higashi, T., Mori, T. et al., 1995. Macrophage scavengerreceptor mediates the endocytic uptake and degradation ofadvanced glycation end products of the Maillard reaction. Eur. J.Biochem. 230, 408]415.
Barbul, A., Regan, M.C., 1995. Immune involvement in woundhealing. Otolaryngol. Clin. North Am. 28, 955]968.
Bauvois, B., Van Weyenbergh, J., Rouillard, D., Wietzerbin, J.,1996. TGF-beta 1-stimulated adhesion of human mononuclearphagocytes to fibronectin and laminin is abolished by IFN-gamma: dependence on alpha 5 beta 1 and beta 2 integrins. Exp.Cell Res. 222, 209]217.
Birkedal-Hansen, H., 1995. Proteolytic remodeling of extracellularmatrix. Curr. Opin. Cell Biol. 7, 728]735.
Bohnsack, J.F., Kleinman, H.K., Takahashi, T., O’Shea, J.J., Brown,E.J., 1985. Connective tissue proteins and phagocytic cell func-tion. J. Exp. Med. 161, 912]923.
Borel, J.P., Bellon, G., Garnotel, R., Monboisse, J.C., 1993. Adhe-sion and activation of human neutrophils on basement mem-brane molecules. Kidney Int. 43, 26]29.
Bouzon, M., Dussert, C., Lissitzky, J.C., Martin, P.M., 1990. Spread-ing of B16 F1 cells on laminin and its proteolytic fragments P1and E8: involvement of laminin carbohydrate chains. Exp. CellRes. 190, 47]56.
Brownlee, M., Cerami, A., Vlassara, H., 1988. Advanced glycosyla-tion end products in tissue and the biochemical basis of diabeticcomplications. N. Engl. J. Med. 318, 1315]1321.
Chin, J.R., Werb, Z., 1997. Matrix metalloproteinases regulatemorphogenesis, migration and remodeling of epithelium, tongueskeletal muscle and cartilage in the mandibular arch. Develop-ment 124, 1519]1530.
Christie, R.H., Freeman, M., Hyman, B.T., 1996. Expression of themacrophage scavenger receptor, a multifunctional lipoproteinreceptor, in microglia associated with senile plaques inAlzheimer’s disease. Am. J. Pathol. 148, 399]403.
Clark, R.A., 1990. Fibronectin matrix deposition and fibronectinreceptor expression in healing and normal skin. J. Invest. Der-matol 94, 128S]134S.
Davis, G.E., 1992a. Affinity of integrins for damaged extracellularmatrix: alpha v beta 3 binds to denatured collagen type Ithrough RGD sites. Biochem. Biophys. Res. Commun. 182,1025]1031.
Davis, G.E., 1992b. The Mac-1 and p150,95 beta 2 integrins binddenatured proteins to mediate leukocyte cell-substrate adhesion.Exp. Cell Res. 200, 242]252.
Davis, G.E., Thomas, J.S., Madden, S., 1997. The alpha4beta1integrin can mediate leukocyte adhesion to casein and dena-tured protein substrates. J. Leuk. Biol. 62, 318]328.
Dedhar, S., Ruoslahti, E., Pierschbacher, M.D., 1987. A cell surfacereceptor complex for collagen type I recognizes the Arg-Gly-Aspsequence. J. Cell Biol. 104, 585]593.
DiPietro, L.A., 1995. Wound healing: the role of the macrophageand other immune cells. Shock 4, 233]240.
Duerksen-Hughes, P.J., Gooding, L.R., 1993. Macrophage-media-Ž .ted cytotoxicity. In: Sitkovsky, M.V., Henkart, P.A. Eds. , Cyto-
toxic Cells: Recognition, Effector Function, and Methods.Birkhauser, Boston, pp. 439]454.
Eierman, D.F., Johnson, C.E., Haskill, J.S., 1989. Human monocyteinflammatory mediator gene expression is selectively regulatedby adherence substrates. J. Immunol. 142, 1970]1976.
El Khoury, J., Thomas, C.A., Loike, J.D., Hickman, S.E., Cao, L.,Silverstein, S.C., 1994. Macrophages adhere to glucose-modifiedbasement membrane collagen IV via their scavenger receptors.J. Biol. Chem. 269, 10197]10200.
Fraser, I., Hughes, D., Gordon, S., 1993. Divalent cation-indepen-dent macrophage adhesion inhibited by monoclonal antibody tomurine scavenger receptor. Nature 364, 343]346.
Frieser, M., Hallmann, R., Johansson, S., Vestweber, D., Goodman,S.L., Sorokin, L., 1996. Mouse polymorphonuclear granulocytebinding to extracellular matrix molecules involves beta 1 inte-grins. Eur. J. Immunol. 26, 3127]3136.
Galis, Z.S., Muszynski, M., Sukhova, G.K., Simon-Morrissey, E.,Libby, P., 1995. Enhanced expression of vascular matrix metal-loproteinases induced in vitro by cytokines and in regions ofhuman atherosclerotic lesions. Ann. New York Acad. Sci. 748,501]507.
Galis, Z.S., Sukhova, G.K., Lark, M.W., Libby, P., 1994. Increasedexpression of matrix metalloproteinases and matrix degradingactivity in vulnerable regions of human atherosclerotic plaques.J. Clin. Invest. 94, 2493]2503.
Gehlsen, K.R., Argraves, W.S., Pierschbacher, M.D., Ruoslahti, E.,1988. Inhibition of in vitro tumor cell invasion by Arg-Gly-Asp-
Žcontaining synthetic peptides published erratum appears in JŽ . .Cell Biol 1989 Jun; 108 6 : following 2546 . J. Cell Biol. 106,
925]930.Gown, A.M., Tsukada, T., Ross, R., 1986. Human atherosclerosis II.
Immunocytochemical analysis of the cellular composition ofhuman atherosclerotic lesions. Am. J. Pathol. 125, 191]207.
Gullberg, D., Terracio, L., Borg, T.K., Rubin, K., 1989. Identifica-tion of integrin-like matrix receptors with affinity for interstitialcollagens. J. Biol. Chem. 264, 12686]12694.
Hampton, R.Y., Golenbock, D.T., Penman, M., Krieger, M., Raetz,C.R., 1991. Recognition and plasma clearance of endotoxin byscavenger receptors. Nature 352, 342]344.
Haskill, S., Yurochko, A.D., Isaacs, K.L., 1992. Regulation ofmacrophage infiltration and activation in sites of chronic in-flammation. Ann. New York Acad. Sci. 664, 93]102.
Horiuchi, S., Higashi, T., Ikeda, K. et al., 1996. Advanced glycationend products and their recognition by macrophage and
Ž .macrophage-derived cells. Diabetes 45 Suppl. 3 , S73]S76.Humphries, M.J., 1990. The molecular basis and specificity of
integrin]ligand interactions. J. Cell Sci. 97, 585]592.Jasin, H.E., Taurog, J.D., 1991. Mechanisms of disruption of the
articular cartilage surface in inflammation. Neutrophil elastaseincreases availability of collagen type II epitopes for bindingwith antibody on the surface of articular cartilage. J. Clin. Invest.87, 1531]1536.
Kadler, K.E., Holmes, D.F., Trotter, J.A., Chapman, J.A., 1996.Collagen fibril formation. Biochem. J. 316, 1]11.
Kirstein, M., Brett, J., Radoff, S., Ogawa, S., Stern, D., Vlassara, H.,1990. Advanced protein glycosylation induces transendothelialhuman monocyte chemotaxis and secretion of platelet-derivedgrowth factor: role in vascular disease of diabetes and aging.Proc. Nat. Acad. Sci. USA 87, 9010]9014.
Krieger, M., Herz, J., 1994. Structures and functions of multiligandlipoprotein receptors: macrophage scavenger receptors and LDL
Ž .receptor-related protein LRP . Annu. Rev. Biochem. 63,601]637.
La Celle, P., Blumenstock, F.A., McKinley, C., Saba, T.M., Vincent,P.A., Gray, V., 1990. Blood-borne collagenous debris complexeswith plasma fibronectin after thermal injury. Blood 75, 470]478.
Landegren, U., 1984. Measurement of cell numbers by means ofthe endogenous enzyme hexosaminidase. Applications to detec-tion of lymphokines and cell surface antigens. J. Immunol. Meth.67, 379]388.
Laskin, D.L., Kimura, T., Sakakibara, S., Riley, D.J., Berg, R.A.,

( )B.B. Gowen et al. r Matrix Biology 19 2000 61]71 71
1986. Chemotactic activity of collagen-like polypeptides for hu-man peripheral blood neutrophils. J. Leuk. Biol. 39, 255]266.
Lin, T.H., Rosales, C., Mondal, K., Bolen, J.B., Haskill, S., Juliano,R.L., 1995. Integrin-mediated tyrosine phosphorylation and cy-tokine message induction in monocytic cells. A possible signalingrole for the Syk tyrosine kinase. J. Biol. Chem. 270, 16189]16197.
Mattana, J., Margiloff, L., Singhal, P.C., 1995. Oxidation of mesan-gial matrix using a mixed function oxidase system augmentsadhesion of macrophages: possible role of macrophage scav-enger receptors. Biochem. Biophys. Res. Commun. 212, 63]69.
McCullagh, K.G., Duance, V.C., Bishop, K.A., 1980. The distribu-Ž .tion of collagen types I, III and V AB in normal and atheros-
clerotic human aorta. J. Pathol. 130, 45]55.McCullagh, K.G., 1983. Increased type I collagen in human
atherosclerotic plaque. Atherosclerosis 46, 247]248.McCutcheon, J.C., Hart, S.P., Canning, M., Ross, K., Humphries,
M.J., Dransfield, I., 1998. Regulation of macrophage phagocyto-sis of apoptotic neutrophils by adhesion to fibronectin. J. Leuk.Biol. 64, 600]607.
McKay, D.B., Vasquez, M.A., Redline, R.W., Lu, C.Y., 1992.Macrophage functions are regulated by murine decidual andtumor extracellular matrices. J. Clin. Invest. 89, 134]142.
Mercurio, A.M., 1990. Laminin: multiple forms, multiple receptors.Curr. Opin. Cell Biol. 2, 845]849.
Messent, A.J., Tuckwell, D.S., Knauper, V., Humphries, M.J., Mur-phy, G., Gavrilovic, J., 1998. Effects of collagenase-cleavage oftype I collagen on alpha2beta1 integrin-mediated cell adhesion.J. Cell Sci. 111, 1127]1135.
Muller, D.J., Schoenenberger, C.A., Schabert, F., Engel, A., 1997.Structural changes in native membrane proteins monitored atsubnanometer resolution with the atomic force microscope: areview. J. Struct. Biol. 119, 149]157.
Newman, S.L., Tucci, M.A., 1990. Regulation of monocytermacro-phage function by extracellular matrix. J. Clin. Invest. 86,703]714.
Palkama, T., 1991. Induction of interleukin-1 production by ligandsbinding to the scavenger receptor in human monocytes and theTHP-1 cell line. Immunology 74, 432]438.
Paresce, D.M., Ghosh, R.N., Maxfield, F.R., 1996. Microglial cellsinternalize aggregates of the Alzheimer’s disease amyloid beta-protein via a scavenger receptor. Neuron 17, 553]565.
Platt, N., Suzuki, H., Kurihara, Y., Kodama, T., Gordon, S., 1996.Role for the class A macrophage scavenger receptor in thephagocytosis of apoptotic thymocytes in vitro. Proc. Natl. Acad.Sci. USA 93, 12456]12460.
Pucillo, C.E., Colombatti, A., Vitale, M., Salzano, S., Rossi, G.,Formisano, S., 1993. Interactions of promonocytic U937 cellswith proteins of the extracellular matrix. Immunology 80,248]252.
Rekhter, M.D., Zhang, K., Narayanan, A.S., Phan, S., Schork, M.A.,Gordon, D., 1993. Type I collagen gene expression in humanatherosclerosis. Localization to specific plaque regions. Am. J.Pathol. 143, 1634]1648.
Rekhter, M.D., 1999. Collagen synthesis in atherosclerosis: toomuch and not enough. Cardiovasc. Res. 41, 376]384.
Resnick, D., Freedman, N.J., Xu, S., Krieger, M., 1993. Secretedextracellular domains of macrophage scavenger receptors formelongated trimers which specifically bind crocidolite asbestos. J.Biol. Chem. 268, 3538]3545.
Ross, R., 1993. The pathogenesis of atherosclerosis: a perspectivefor the 1990s. Nature 362, 801]809.
Ruoslahti, E., 1991. Integrins. J. Clin. Invest. 87, 1]5.Ruoslahti, E., Noble, N.A., Kagami, S., Border, W.A., 1994. Inte-
Ž .grins. Kidney Int. 45 Suppl. 44 , S17]S22.Ruoslahti, B.O., Obrink, B., 1996. Common principles in cell adhe-
sion. Exp. Cell Res. 227, 1]11.Schabert, F.A., Henn, C., Engel, A., 1995. Native Escherichia coli
OmpF porin surfaces probed by atomic force microscopy. Sci-ence 268, 92]94.
Shaw, L.M., Messier, J.M., Mercurio, A.M., 1990. The activationdependent adhesion of macrophages to laminin involves cy-toskeletal anchoring and phosporylation of the a6b1 integrin. J.Cell Biol. 110, 2167]2174.
Shingleton, W.D., Hodges, D.J., Brick, P., Cawston, T.E., 1996.Collagenase: a key enzyme in collagen turnover. Biochem. CellBiol. 74, 759]775.
Sporn, S.A., Eierman, D.F., Johnson, C.E. et al., 1990. Monocyteadherence results in selective induction of novel genes sharinghomology with mediators of inflammation and tissue repair. J.Immunol. 144, 4434]4441.
Sukhova, G.K., Schonbeck, U., Rabkin, E. et al., 1999. Evidence forincreased collagenolysis by interstitial collagenase-1 and -3 invulnerable human atheromatous plaques. Circulation 99,2503]2509.
Terpstra, V., Kondratenko, N., Steinberg, D., 1997. Macrophageslacking scavenger receptor A show a decrease in binding anduptake of acetylated low-density lipoprotein and of apoptoticthymocytes, but not of oxidatively damaged red blood cells. Proc.Natl. Acad. Sci. USA 94, 8127]8131.
Thomas, D.W., O’Neill, I.D., Harding, K.G., Sheperd, J.P., 1995.Cutaneous wound healing: a current perspective. J. Oral. Max-illofac. Surg. 53, 442]447.
Weiss, S.J., 1989. Tissue destruction by neutrophils. N. Engl. J.Med. 320, 365]376.
Weston, C.A., Weeks, B.S., 1996. bFGF stimulates U937 cell adhe-sion to fibronectin and secretion of gelatinase B. Biochem.Biophys. Res. Commun. 228, 318]323.
Yokota, T., Ehlin-Henriksson, B., Hansson, G.K., 1998. Scavengerreceptors mediate adhesion of activated B lymphocytes. Exp.Cell Res. 239, 16]22.
Yurochko, A.D., Liu, D.Y., Eierman, D., Haskill, S., 1992. Integrinsas a primary signal transduction molecule regulating monocyteimmediate]early gene induction. Proc. Nat. Acad. Sci. USA 89,9034]9038.
Copyright © 2022 FDOKUMEN




















