
SAP1 is a microvillus-specific protein tyrosine phosphatase that modulates intestinal tumorigenesis
14
© 2009 The Authors Genes to Cells (2009) 14, 295–308 Journal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd. 295 DOI: 10.1111/j.1365-2443.2008.01270.x Blackwell Publishing Inc SAP-1 is an intestinal MV-specific PTP H Sadakata et al. SAP-1 is a microvillus-specific protein tyrosine phosphatase that modulates intestinal tumorigenesis Hisanobu Sadakata 1,2,† , Hideki Okazawa 1,† , Takashi Sato 3,† , Yana Supriatna 1 , Hiroshi Ohnishi 1 , Shinya Kusakari 1 , Yoji Murata 1 , Tomokazu Ito 1 , Uichi Nishiyama 4 , Takashi Minegishi 2 , Akihiro Harada 3 and Takashi Matozaki 1, * 1 Laboratory of Biosignal Sciences, Institute for Molecular and Cellular Regulation, Gunma University, 3-39-15 Showa-Machi, Maebashi, Gunma 371-8512, Japan 2 Department of Obstetrics and Gynecology, Graduate School of Medicine, Gunma University, 3-39-22 Showa-Machi, Maebashi, Gunma 371-8511, Japan 3 Laboratory of Molecular Traffic, Institute for Molecular and Cellular Regulation, Gunma University, 3-39-15 Showa-Machi, Maebashi, Gunma 371-8512, Japan 4 Pharmaceutical Development Laboratories, Kirin Brewery Co. Ltd., Takasaki, Gunma 370-1295, Japan SAP-1 (PTPRH) is a receptor-type protein tyrosine phosphatase (RPTP) with a single catalytic domain in its cytoplasmic region and fibronectin type III-like domains in its extracellular region. The cellular localization and biological functions of this RPTP have remained unknown, however. We now show that mouse SAP-1 mRNA is largely restricted to the gastrointestinal tract and that SAP-1 protein localizes to the microvilli of the brush border in gastrointestinal epithelial cells. The expression of SAP-1 in mouse intestine is minimal during embryonic development but increases markedly after birth. SAP-1-deficient mice manifested no marked changes in morphology of the intestinal epithelium. In contrast, SAP-1 ablation inhibited tumorigenesis in mice with a heterozygous mutation of the adenomatous polyposis coli gene. These results thus suggest that SAP-1 is a microvillus-specific RPTP that regulates intestinal tumorigenesis. Introduction Intestinal epithelial cells of mammals are highly polar- ized. The apical surface of these cells consists of the brush border, which is formed by a large number of microvilli (Mooseker 1985; Holmes & Lobley 1989; Ross & Pawlina 2006), whereas the lateral membrane adjacent to the apical surface contains tight junctions, adherens junc- tions and desmosomes (in this order from the apical to the basal side), resulting in a relatively straight and tight interface with neighboring cells (Takeichi 1995; Gumbiner 2005; Miyoshi & Takai 2005; Ross & Pawlina 2006). This cell–cell adhesion is mediated by homophilic binding in trans of various adhesion molecules, including claudin at tight junctions as well as E-cadherin and nectin at adherens junctions (Takeichi 1995; Tsukita & Furuse 2002; Gumbiner 2005; Miyoshi & Takai 2005). Protein tyrosine phosphorylation plays a key role in regulation of cell–cell adhesion, with catenins, which are associated with the intracellular region of E-cadherin (Gumbiner 2005; Lilien & Balsamo 2005), being important targets for such phosphorylation and dephosphorylation. Indeed, receptor tyrosine kinases, such as the receptors for epidermal growth factor or hepatocyte growth factor, as well as Src family kinases and protein tyrosine phos- phatases (PTPs) such as PTPμ are localized at sites of cell–cell adhesion in epithelial cells (Maratos-Flier et al. 1987; Takeda et al. 1995; Kamei et al. 1999; Sallee et al. 2006). Moreover, these enzymes contribute to the dynamic plasticity of cell–cell adhesion in epithelial cells (Takeda et al. 1995; Lilien & Balsamo 2005; Sallee et al. 2006). In contrast, it remains unknown whether tyrosine phosphorylation–dephosphorylation participates in regulation of brush border function or which tyrosine kinases or PTPs localize to and function at the brush border of epithelial cells. Communicated by: Yoshimi Takai *Correspondence: [email protected] † These authors have contributed equally to this work.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of SAP1 is a microvillus-specific protein tyrosine phosphatase that modulates intestinal tumorigenesis

© 2009 The Authors Genes to Cells (2009) 14, 295–308Journal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
295DOI: 10.1111/j.1365-2443.2008.01270.x
Blackwell Publishing IncSAP-1 is an intestinal MV-specific PTPH Sadakata et al.SAP-1 is a microvillus-specific protein tyrosine phosphatase that modulates intestinal tumorigenesis
Hisanobu Sadakata1,2,†, Hideki Okazawa1,†, Takashi Sato3,†, Yana Supriatna1, Hiroshi Ohnishi1, Shinya Kusakari1, Yoji Murata1, Tomokazu Ito1, Uichi Nishiyama4, Takashi Minegishi2, Akihiro Harada3 and Takashi Matozaki1,*1Laboratory of Biosignal Sciences, Institute for Molecular and Cellular Regulation, Gunma University, 3-39-15 Showa-Machi, Maebashi, Gunma 371-8512, Japan 2Department of Obstetrics and Gynecology, Graduate School of Medicine, Gunma University, 3-39-22 Showa-Machi, Maebashi, Gunma 371-8511, Japan 3Laboratory of Molecular Traffic, Institute for Molecular and Cellular Regulation, Gunma University, 3-39-15 Showa-Machi, Maebashi, Gunma 371-8512, Japan 4Pharmaceutical Development Laboratories, Kirin Brewery Co. Ltd., Takasaki, Gunma 370-1295, Japan
SAP-1 (PTPRH) is a receptor-type protein tyrosine phosphatase (RPTP) with a single catalyticdomain in its cytoplasmic region and fibronectin type III-like domains in its extracellular region.The cellular localization and biological functions of this RPTP have remained unknown, however.We now show that mouse SAP-1 mRNA is largely restricted to the gastrointestinal tract and thatSAP-1 protein localizes to the microvilli of the brush border in gastrointestinal epithelial cells.The expression of SAP-1 in mouse intestine is minimal during embryonic development butincreases markedly after birth. SAP-1-deficient mice manifested no marked changes in morphologyof the intestinal epithelium. In contrast, SAP-1 ablation inhibited tumorigenesis in mice witha heterozygous mutation of the adenomatous polyposis coli gene. These results thus suggest thatSAP-1 is a microvillus-specific RPTP that regulates intestinal tumorigenesis.
Introduction
Intestinal epithelial cells of mammals are highly polar-ized. The apical surface of these cells consists of the brushborder, which is formed by a large number of microvilli(Mooseker 1985; Holmes & Lobley 1989; Ross & Pawlina2006), whereas the lateral membrane adjacent to theapical surface contains tight junctions, adherens junc-tions and desmosomes (in this order from the apicalto the basal side), resulting in a relatively straight andtight interface with neighboring cells (Takeichi 1995;Gumbiner 2005; Miyoshi & Takai 2005; Ross & Pawlina2006). This cell–cell adhesion is mediated by homophilicbinding in trans of various adhesion molecules, includingclaudin at tight junctions as well as E-cadherin and nectinat adherens junctions (Takeichi 1995; Tsukita & Furuse
2002; Gumbiner 2005; Miyoshi & Takai 2005). Proteintyrosine phosphorylation plays a key role in regulation ofcell–cell adhesion, with catenins, which are associatedwith the intracellular region of E-cadherin (Gumbiner2005; Lilien & Balsamo 2005), being important targetsfor such phosphorylation and dephosphorylation. Indeed,receptor tyrosine kinases, such as the receptors forepidermal growth factor or hepatocyte growth factor, aswell as Src family kinases and protein tyrosine phos-phatases (PTPs) such as PTPμ are localized at sites ofcell–cell adhesion in epithelial cells (Maratos-Flier et al.1987; Takeda et al. 1995; Kamei et al. 1999; Sallee et al.2006). Moreover, these enzymes contribute to thedynamic plasticity of cell–cell adhesion in epithelial cells(Takeda et al. 1995; Lilien & Balsamo 2005; Sallee et al.2006). In contrast, it remains unknown whether tyrosinephosphorylation–dephosphorylation participates inregulation of brush border function or which tyrosinekinases or PTPs localize to and function at the brushborder of epithelial cells.
Communicated by: Yoshimi Takai*Correspondence: [email protected]†These authors have contributed equally to this work.

H Sadakata et al.
Genes to Cells (2009) 14, 295–308 © 2009 The AuthorsJournal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
296
Stomach cancer-associated protein tyrosine phos-phatase-1 (SAP-1) (also known as PTPRH) was originallyidentified as a PTP expressed in a human stomach cancercell line (Matozaki et al. 1994). It is a receptor-type PTP(RPTP, R3 subtype) (Andersen et al. 2001), with a singlecatalytic domain in its cytoplasmic region and eightfibronectin type III-like domains in its extracellularregion (Matozaki et al. 1994). Although its expressionwas found to be markedly increased in human colon andpancreatic cancers (Matozaki et al. 1994; Seo et al. 1997),the expression and localization of SAP-1 in normaltissues have not been well defined. A “substrate-trapping”approach identified p130Cas, a prominent focal adhesion-associated component of the integrin signaling pathway,as a likely physiological substrate of SAP-1 (Noguchiet al. 2001). Forced expression of SAP-1 in cultured cellsalso resulted in the dephosphorylation of several addi-tional focal adhesion-associated proteins, including focaladhesion kinase and paxillin, as well as in impairment ofreorganization of the actin-based cytoskeleton (Noguchiet al. 2001), suggesting that SAP-1 regulates the latterprocess. Forced expression of this PTP also inhibited cellproliferation, an effect that was mediated in part either byattenuation of growth factor-induced mitogen-activatedprotein kinase activation or by induction of caspase-dependent apoptosis (Noguchi et al. 2001; Takada et al.2002). The cytoplasmic region of SAP-1 binds directlyto Lck, and over-expression of SAP-1 resulted indown-regulation of the kinase activity of Lck andconsequent inhibition of T cell receptor-mediated T cellfunction (Ito et al. 2003). However, given that most ofthese observations were made with cultured cells, thephysiological or pathological functions of SAP-1 in vivoremain unknown.
We have now determined the sequence of mouseSAP-1 cDNA and generated a mAb to the mouseprotein. With the use of these materials, we have shownthat SAP-1 is expressed predominantly in the gastro-intestinal tract, most prominently in the small intestine,and that it is specifically localized at the microvilli ofgastrointestinal epithelial cells. Moreover, our observa-tions suggest that SAP-1 plays an important role in theregulation of intestinal tumorigenesis.
ResultsPredicted structure of mouse SAP-1 and its predominant expression in the intestine
To examine the expression of SAP-1 in various tissues,we first cloned a cDNA encoding mouse SAP-1.Searching of mouse Expressed Sequence Tag (EST) and
other DNA data bases revealed several mouse DNA orEST sequences with similarity to those of humanSAP-1 cDNA. With the use of these sequences, weobtained partial cDNA fragments for mouse SAP-1 byreverse transcription-polymerase chain reaction (RT-PCR) as described in Experimental procedures. Wefurther searched the mouse cDNA data base provided bythe Functional Annotation of the mouse (FANTOM)project (Carninci et al. 2005) for sequences similar tothose of the partial mouse SAP-1 cDNA fragmentsand thereby obtained a full-length mouse SAP-1 cDNA(approximately 3.5 kb) (DDBJ Accession No. AB217856).The predicted structure of the mouse SAP-1 protein,which comprises 971 amino acids, was found to besimilar to that of human SAP-1 (Fig. 1A), with theexception that the mouse protein contains six fibronectintype III-like domains in its extracellular region, whereasthe human protein contains eight such domains (Mato-zaki et al. 1994). The amino acid sequences of mouseand human SAP-1 proteins share an overall identity of56%, with the corresponding values for the extracellularand intracellular regions being 43% and 77%, respec-tively (Fig. 1A). A search of the Novartis GeneAtlas<http://symatlas.gnf.org/symatlas> with the mousecDNA sequence revealed that SAP-1 mRNA is abun-dant in the small intestine and colon. Indeed, Northernblot analysis showed that mouse SAP-1 mRNA waspreferentially distributed to the small intestine, stomachand colon (Fig. 1B).
To investigate the expression and localization ofSAP-1 protein, we generated a monoclonal antibody(mAb) (clone 123) to the extracellular region of mouseSAP-1. Immunoblot analysis with this mAb also revealedprominent expression of the approximately 250-kDaSAP-1 protein in the intestine, and a low level of theexpression was observed in testis (Fig. 1C). Further-more, the amount of SAP-1 in the duodenum or jejunumwas markedly greater than that in the stomach or colon(Fig. 1D).
Specific localization of mouse SAP-1 to microvilli of gastrointestinal epithelial cells
We next examined the localization of SAP-1 in the smallintestine. Immunohistofluorescence analysis with themAb to SAP-1 revealed that SAP-1 was localized at theapical surface of intestinal epithelial cells (Fig. 2A),whereas β-catenin was present at sites of cell–celladhesion as well as in the cytoplasm of epithelial cells, asdescribed previously (Sena et al. 2006). The localizationof SAP-1 was similar to that of ezrin or alkaline phos-phatase (Fig. 2B), both of which are known to be

SAP-1 is an intestinal MV-specific PTP
© 2009 The Authors Genes to Cells (2009) 14, 295–308Journal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
297
localized at the microvilli of the intestinal epithelium(Narisawa et al. 2003; Saotome et al. 2004). Further-more, the localization of SAP-1 overlapped with thatof ezrin/radixin/moesin binding phosphoprotein of50 kDa (EBP-50) at the apical surface of the intestinalepithelium (Fig. 2C). EBP-50 is a protein that linksCD44 at the cell surface to ezrin and the actin cyto-skeleton and is thus concentrated at the microvilli of theintestinal epithelium (Tsukita & Yonemura 1999). Eachmicrovillus at the brush border of intestinal epithelialcells contains a bundle of actin filaments that extends theentire length of the microvillus as well as into the actinfilament meshwork below it known as the terminal web(Mooseker 1985; Tyska et al. 2005; Ross & Pawlina2006). SAP-1 immunoreactivity was detected immedi-ately above the prominent staining of F-actin revealed byphalloidin, which may correspond to the terminal web,
at the brush border of intestinal epithelial cells (Fig. 2C).Immuno-electron microscopy (EM) with the mAb toSAP-1 also revealed prominent staining at the microvilliof intestinal epithelial cells from wild-type mice (Fig. 2D),whereas such staining was virtually undetectable insections from SAP-1-deficient mice, the generation ofwhich is described below. These results thus indicatedthat SAP-1 is specifically expressed in intestinal epithelialcells and localizes to the microvilli of these cells.
SAP-1 was also localized at the apical surface ofepithelial cells of the stomach, although the level ofexpression was markedly lower than that in the smallintestine (Fig. 3A). Microvilli are more prominent inthe small intestine than in the stomach (Ross & Pawlina2006). Moreover, the expression of SAP-1 was absent ingoblet cells (Fig. 3B), which were stained with Ulexeuropaeus agglutinin (UEA)-I and lack microvilli (Fischer
Figure 1 Structure of mouse SAP-1 andits predominant expression in the gastro-intestinal tract. (A) Schematic representationof the structures of human and mouseSAP-1. Numbers indicate amino acidresidues. FN III, fibronectin type III; TM,transmembrane region; PTP, phosphataseregion. The sequence identity between theextracellular or cytoplasmic regions ofthe human and mouse SAP-1 proteins isindicated. (B) Distribution of SAP-1 mRNAamong mouse tissues. PolyadenylatedRNA (2 μg) isolated from the indicatedtissues was subjected to Northern hybridi-zation with approximately 3.5-kb mouseSAP-1 (upper panel) or approximately0.5-kb mouse glyceraldehyde-3-phosphatedehydrogenase (GAPDH) (loading control,lower panel) cRNA probes. (C) Expressionof SAP-1 protein in mouse tissues. Lysatesof various tissues (20 μg of protein) weresubjected to immunoblot analysis withmAbs to SAP-1 (upper panel) or to β-tubulin(loading control, lower panel). Thepositions of molecular size standards areindicated on the left. (D) Distribution ofSAP-1 in the mouse gastrointestinal tract.Lysates of the stomach, duodenum,jejunum and colon were subjected toimmunoblot analysis as in (C).

H Sadakata et al.
Genes to Cells (2009) 14, 295–308 © 2009 The AuthorsJournal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
298
et al. 1984). These observations were thus also consistentwith the notion that SAP-1 is localized specifically atmicrovilli of the gastrointestinal epithelium.
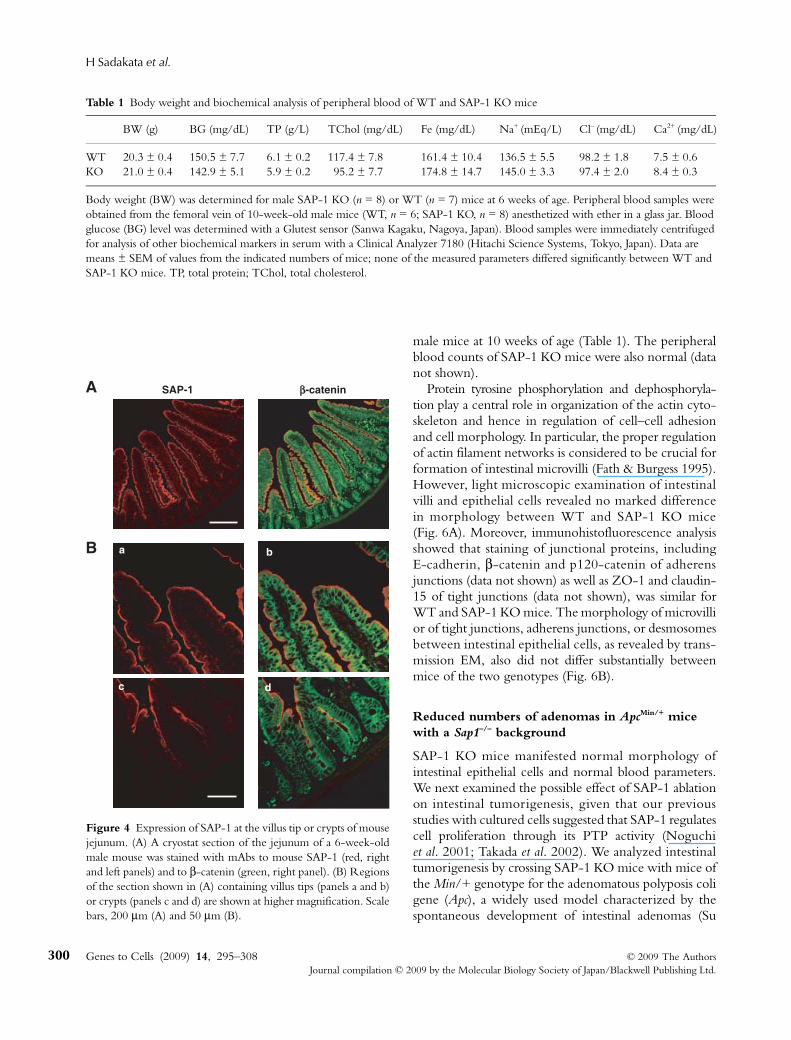
We next examined the expression of SAP-1 duringdevelopment of the intestine in mouse embryos. Immuno-blot and immunohistofluorescence analyses revealedthat SAP-1 was virtually undetectable in the intestineat embryonic day (E) 14.5, whereas the expression ofE-cadherin was apparent at this time (Fig. 3C,D).Expression of SAP-1 was detected at a low level at E16.5and increased markedly after birth (Fig. 3C,D). Theseresults suggested that expression of SAP-1 increases inparallel with the development of intestinal epithelialcells. We further examined the expression of SAP-1during differentiation of intestinal epithelial cells. Inadult mice, staining for SAP-1 was absent or present atonly a low level in intestinal crypts (Fig. 4), where newlygenerated intestinal epithelial cells and their progenitorsreside (Ross & Pawlina 2006; Blanpain et al. 2007). Theseresults suggested that expression of SAP-1 increases inparallel with differentiation of intestinal epithelial cells.
Normal morphology of intestinal epithelial cells in SAP-1-deficient mice
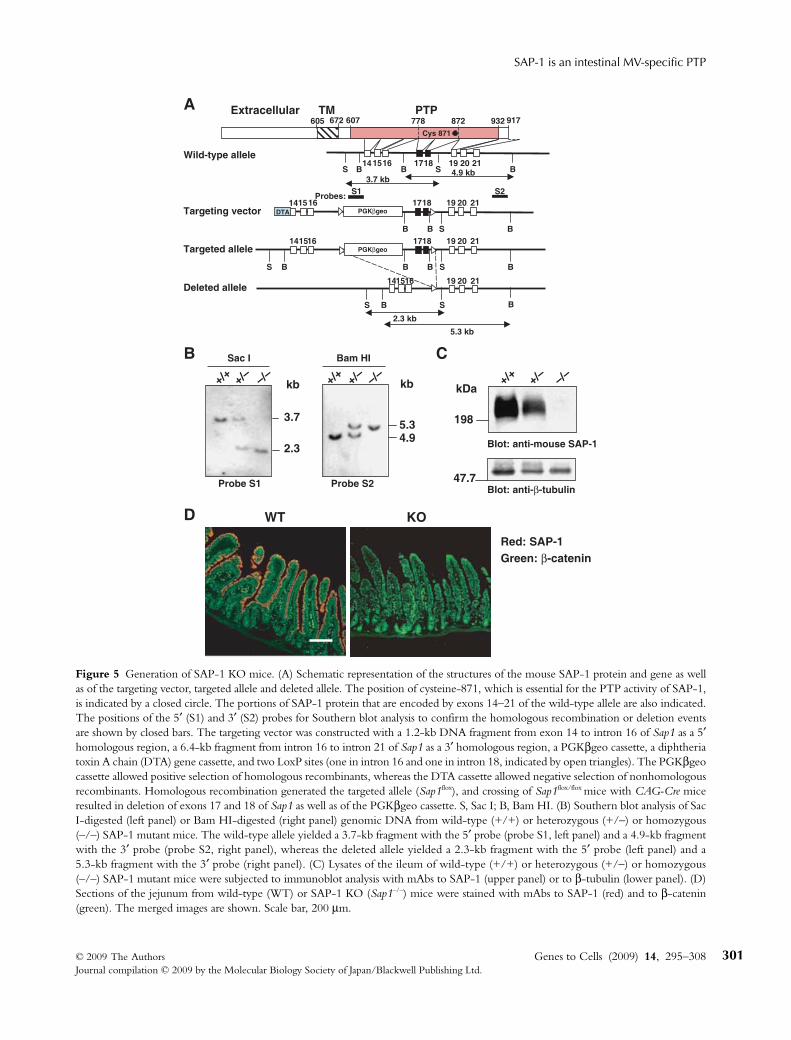
To investigate the physiological or pathological roles ofSAP-1 in vivo, we generated SAP-1-deficient mice bytargeted deletion of a fragment of genomic DNAcontaining exons 17 and 18 of the SAP-1 gene, whichencode a region of the protein (amino acids 778–872)required for enzymatic activity. We generated a Sap1mutant allele (Sap1flox) by introducing LoxP sites into theintrons flanking exons 17 and 18 (Fig. 5A). Deletion ofthese two exons would be expected to introduce aframeshift mutation, resulting in loss of the COOH-terminal half of the catalytic domain as well as thedownstream region of the protein. Crossing of Sap1flox/flox
mice with CAG-Cre transgenic animals (Sakai &Miyazaki 1997) resulted in the generation of SAP-1-deficient (SAP-1 KO) mice. Southern blot analysisconfirmed the presence of the deleted allele in SAP-1KO mice (Fig. 5B). Immunoblot analysis with the mAbto SAP-1 also demonstrated the complete loss of SAP-1protein or a reduction in its abundance of approximately50% in the intestine of SAP-1 KO (Sap1–/–) mice andSap1+/– heterozygotes, respectively (Fig. 5C). Immuno-histofluorescence staining with this mAb also revealedthe complete loss of SAP-1 in the apical brush border ofthe small intestine of SAP-1 KO mice (Fig. 5D). Giventhat the mAb to SAP-1 reacts with the extracellularregion of the protein, we also performed immunoblotanalysis with polyclonal antibodies to the cytoplasmic
Figure 2 Localization of SAP-1 to the epithelium of the mousesmall intestine. (A) A cryostat section of the jejunum of a 6-week-oldmale mouse was subjected to immunofluorescence stainingwith mAbs to mouse SAP-1 (left panels) and to β-catenin (middlepanels). Merged images are also shown in the right panels. Boxedregions in the upper panels are shown at higher magnification inthe lower panels. Scale bars, 200 μm (upper panels) and 50 μm(lower panels). (B) Sections of mouse jejunum were stained withmAbs to SAP-1 (left panel) and to ezrin (middle panel) as well asfor alkaline phosphatase activity (right panel). Scale bar, 50 μm.(C) Sections of mouse jejunum were stained with a mAb toSAP-1 and either polyclonal antibodies to EBP-50 (upper panels)or rhodamine-conjugated phalloidin (lower panels), as indicated.Merged images are shown in the right panels. Scale bars, 10 μm.(D) Immuno-EM of the jejunum of wild-type (WT) or SAP-1knockout (KO) mice with a mAb to mouse SAP-1. Scale bar, 200 nm.

SAP-1 is an intestinal MV-specific PTP
© 2009 The Authors Genes to Cells (2009) 14, 295–308Journal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
299
region of mouse SAP-1. This analysis also revealed thecomplete ablation of SAP-1 in the intestine of SAP-1KO mice (data not shown). These results thus indicatedthat the entire SAP-1 protein is absent or undetectable inthe intestine of SAP-1 KO mice, likely as a result ofinstability of the mutant SAP-1 mRNA or its proteinproduct. We attempted to generate mice with microvillus-specific deficiency of SAP-1, but we found that theexpression level of SAP-1 in Sap1flox/flox mice wasreduced by > 50% relative to that in wild-type animals(data not shown). The SAP-1 KO mice were thereforebackcrossed onto the C57BL/6 background for morethan four generations, and offspring homozygous for
the wild-type (WT) or mutant Sap1 alleles were studiedin the following experiments.
The SAP-1 KO mice were born apparently healthy,survived to adulthood, and fertile. Although intestinalepithelial cells play a central role in food digestion and inabsorption of nutrients and electrolytes through theintestine, and SAP-1 localizes specifically to microvilli ofthe gastrointestinal epithelium, the body weight of maleSAP-1 KO mice at 6 weeks of age was similar to thatof WT controls (Table 1). Moreover, the blood glucoseconcentration as well as the serum concentrations oftotal protein, total cholesterol, iron, Na+, Cl– and Ca2+
did not differ substantially between WT and SAP-1 KO
Figure 3 Expression of SAP-1 in variousregions of the epithelium of the gastrointes-tinal tract and during development of thesmall intestine. (A) Sections of the jejunum(left panels) or stomach (right panels) of6-week-old male mice were stained withboth a mAb to mouse SAP-1 (upper panels)and rhodamine-conjugated phalloidin(middle panels). Merged images are shownin the lower panels. Scale bar, 50 μm. (B) Asection of mouse ileum was stained withboth a mAb to SAP-1 (upper panel) andFITC-conjugated UEA-I (middle panel).A merged image is shown in the lowerpanel. The arrowhead indicates a goblet cellin the ileal epithelium. Scale bar, 10 μm.(C) Lysates (20 μg of protein) of intestinaltissue from mice at the indicated stages ofdevelopment were subjected to immunoblotanalysis with mAbs to mouse SAP-1 (upperpanel) or to actin (loading control, lowerpanel). (D) Sections of mouse intestinaltissue at the indicated developmental stageswere stained with mAbs to mouse SAP-1(upper panels) or to E-cadherin (lowerpanels). Scale bar, 200 μm. P1 and P8,postnatal days 1 and 8, respectively.
H Sadakata et al.
Genes to Cells (2009) 14, 295–308 © 2009 The AuthorsJournal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
300
male mice at 10 weeks of age (Table 1). The peripheralblood counts of SAP-1 KO mice were also normal (datanot shown).
Protein tyrosine phosphorylation and dephosphoryla-tion play a central role in organization of the actin cyto-skeleton and hence in regulation of cell–cell adhesionand cell morphology. In particular, the proper regulationof actin filament networks is considered to be crucial forformation of intestinal microvilli (Fath & Burgess 1995).However, light microscopic examination of intestinalvilli and epithelial cells revealed no marked differencein morphology between WT and SAP-1 KO mice(Fig. 6A). Moreover, immunohistofluorescence analysisshowed that staining of junctional proteins, includingE-cadherin, β-catenin and p120-catenin of adherensjunctions (data not shown) as well as ZO-1 and claudin-15 of tight junctions (data not shown), was similar forWT and SAP-1 KO mice. The morphology of microvillior of tight junctions, adherens junctions, or desmosomesbetween intestinal epithelial cells, as revealed by trans-mission EM, also did not differ substantially betweenmice of the two genotypes (Fig. 6B).
Reduced numbers of adenomas in ApcMin/+ mice with a Sap1–/– background
SAP-1 KO mice manifested normal morphology ofintestinal epithelial cells and normal blood parameters.We next examined the possible effect of SAP-1 ablationon intestinal tumorigenesis, given that our previousstudies with cultured cells suggested that SAP-1 regulatescell proliferation through its PTP activity (Noguchiet al. 2001; Takada et al. 2002). We analyzed intestinaltumorigenesis by crossing SAP-1 KO mice with mice ofthe Min/+ genotype for the adenomatous polyposis coligene (Apc), a widely used model characterized by thespontaneous development of intestinal adenomas (Su
Table 1 Body weight and biochemical analysis of peripheral blood of WT and SAP-1 KO mice
BW (g) BG (mg/dL) TP (g/L) TChol (mg/dL) Fe (mg/dL) Na+ (mEq/L) Cl– (mg/dL) Ca2+ (mg/dL)
WT 20.3 ± 0.4 150.5 ± 7.7 6.1 ± 0.2 117.4 ± 7.8 161.4 ± 10.4 136.5 ± 5.5 98.2 ± 1.8 7.5 ± 0.6KO 21.0 ± 0.4 142.9 ± 5.1 5.9 ± 0.2 95.2 ± 7.7 174.8 ± 14.7 145.0 ± 3.3 97.4 ± 2.0 8.4 ± 0.3
Body weight (BW) was determined for male SAP-1 KO (n = 8) or WT (n = 7) mice at 6 weeks of age. Peripheral blood samples were obtained from the femoral vein of 10-week-old male mice (WT, n = 6; SAP-1 KO, n = 8) anesthetized with ether in a glass jar. Blood glucose (BG) level was determined with a Glutest sensor (Sanwa Kagaku, Nagoya, Japan). Blood samples were immediately centrifuged for analysis of other biochemical markers in serum with a Clinical Analyzer 7180 (Hitachi Science Systems, Tokyo, Japan). Data are means ± SEM of values from the indicated numbers of mice; none of the measured parameters differed significantly between WT and SAP-1 KO mice. TP, total protein; TChol, total cholesterol.
Figure 4 Expression of SAP-1 at the villus tip or crypts of mousejejunum. (A) A cryostat section of the jejunum of a 6-week-oldmale mouse was stained with mAbs to mouse SAP-1 (red, rightand left panels) and to β-catenin (green, right panel). (B) Regionsof the section shown in (A) containing villus tips (panels a and b)or crypts (panels c and d) are shown at higher magnification. Scalebars, 200 μm (A) and 50 μm (B).
SAP-1 is an intestinal MV-specific PTP
© 2009 The Authors Genes to Cells (2009) 14, 295–308Journal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
301
Figure 5 Generation of SAP-1 KO mice. (A) Schematic representation of the structures of the mouse SAP-1 protein and gene as wellas of the targeting vector, targeted allele and deleted allele. The position of cysteine-871, which is essential for the PTP activity of SAP-1,is indicated by a closed circle. The portions of SAP-1 protein that are encoded by exons 14–21 of the wild-type allele are also indicated.The positions of the 5′ (S1) and 3′ (S2) probes for Southern blot analysis to confirm the homologous recombination or deletion eventsare shown by closed bars. The targeting vector was constructed with a 1.2-kb DNA fragment from exon 14 to intron 16 of Sap1 as a 5′homologous region, a 6.4-kb fragment from intron 16 to intron 21 of Sap1 as a 3′ homologous region, a PGKβgeo cassette, a diphtheriatoxin A chain (DTA) gene cassette, and two LoxP sites (one in intron 16 and one in intron 18, indicated by open triangles). The PGKβgeocassette allowed positive selection of homologous recombinants, whereas the DTA cassette allowed negative selection of nonhomologousrecombinants. Homologous recombination generated the targeted allele (Sap1flox), and crossing of Sap1flox/flox mice with CAG-Cre miceresulted in deletion of exons 17 and 18 of Sap1 as well as of the PGKβgeo cassette. S, Sac I; B, Bam HI. (B) Southern blot analysis of SacI-digested (left panel) or Bam HI-digested (right panel) genomic DNA from wild-type (+/+) or heterozygous (+/–) or homozygous(–/–) SAP-1 mutant mice. The wild-type allele yielded a 3.7-kb fragment with the 5′ probe (probe S1, left panel) and a 4.9-kb fragmentwith the 3′ probe (probe S2, right panel), whereas the deleted allele yielded a 2.3-kb fragment with the 5′ probe (left panel) and a5.3-kb fragment with the 3′ probe (right panel). (C) Lysates of the ileum of wild-type (+/+) or heterozygous (+/–) or homozygous(–/–) SAP-1 mutant mice were subjected to immunoblot analysis with mAbs to SAP-1 (upper panel) or to β-tubulin (lower panel). (D)Sections of the jejunum from wild-type (WT) or SAP-1 KO (Sap1–/–) mice were stained with mAbs to SAP-1 (red) and to β-catenin(green). The merged images are shown. Scale bar, 200 μm.

H Sadakata et al.
Genes to Cells (2009) 14, 295–308 © 2009 The AuthorsJournal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
302
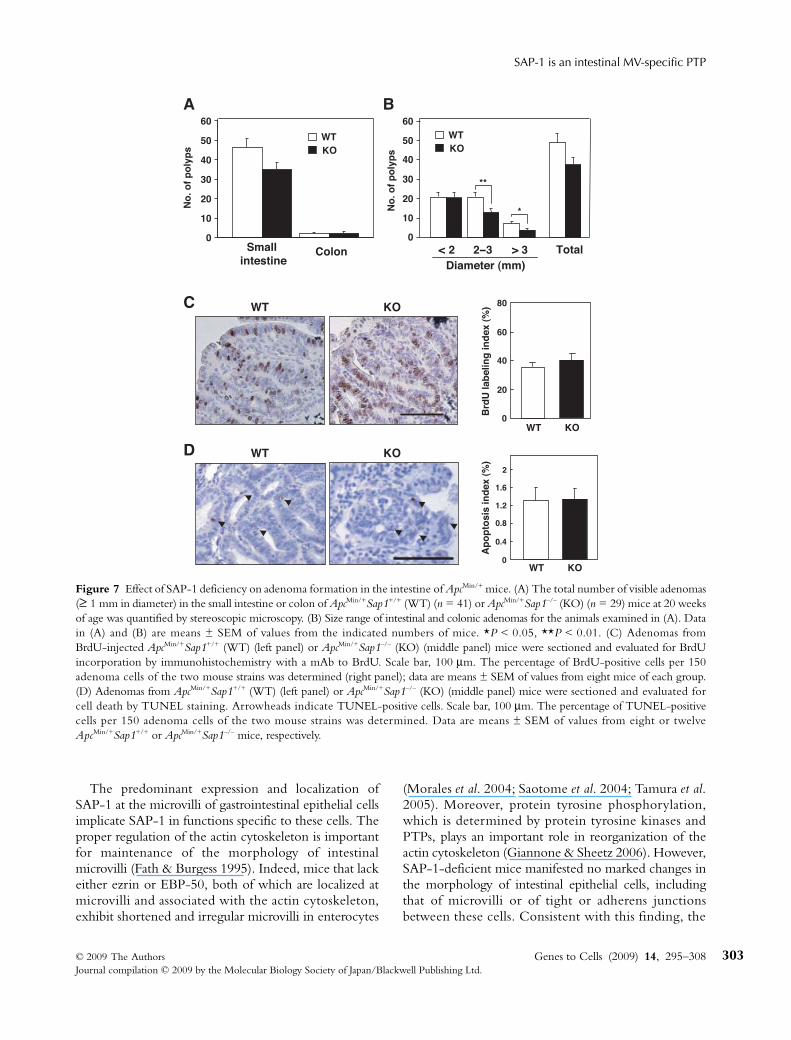
et al. 1992; Rakoff-Nahoum & Medzhitov 2007). Thenumbers of visible adenomatous polyps (≥ 1 mm indiameter) that formed in the small intestine and colonwere quantified by stereoscopic microscopy. At 20 weeks
of age, the total number of adenomas in the smallintestine, but not that in the colon, appeared to bereduced in ApcMin/+Sap1–/– mice compared with that inApcMin/+Sap1+/+ mice (Fig. 7A), although such reductionwas not statistically significant. However, the number ofmacroadenomas (≥ 2 mm in diameter) in the intestinewas markedly reduced in ApcMin/+Sap1–/– mice comparedwith that in ApcMin/+Sap1+/+ mice (Fig. 7B). These resultsthus suggested that SAP-1 regulates intestinal tumorgrowth in ApcMin/+ mice.
Given that the size of tumors is influenced by thebalance between cell proliferation and cell death withinthe tumor mass, we investigated the rates of cell pro-liferation and apoptosis in size-matched tumors fromApcMin/+Sap1+/+ and ApcMin/+Sap1–/– mice. Neither thepercentage of 5-bromo-2′-deoxyuridine (BrdU)-positive(proliferating) cells (Fig. 7C) nor the percentage ofTdT-mediated dUTP-biotin nick end labeling (TUNEL)-positive (apoptotic) cells (Fig. 7D) differed significantlybetween mice of the two genotypes. The reduced extentof intestinal tumorigenesis in ApcMin/+Sap1–/– mice thusdid not appear to be attributable to a reduced rate of cellproliferation or an increased rate of apoptosis.
DiscussionWe have here shown that mouse SAP-1 is localized atthe microvilli of the brush border in gastrointestinalepithelial cells. SAP-1 is classified as an RPTP of the R3subtype, as are PTP-RO, Vascular endothelial cell-specific phosphotyrosine phosphatase (VE-PTP) andDEP-1 (Andersen et al. 2001). All of these enzymesshare similar structures, with a single catalytic domainin the cytoplasmic region and fibronectin type III-likedomains in the extracellular region. PTP-RO (alsoknown as GLEPP1) is specifically expressed in podocytesof the renal glomerulus (Thomas et al. 1994; Yang et al.1996); in particular, it is localized at the apical surface offoot processes in the highly polarized visceral glomerularepithelial cells (Yang et al. 1996; Wharram et al. 2000).Although the morphology of foot processes is distinctfrom that of microvilli, both of these structures are richin actin (Fath & Burgess 1995; Benzing 2004). Inaddition, the expression of VE-PTP is restricted toendothelial cells (Bäumer et al. 2006; Dominguez et al.2007), with VE-PTP also being localized at the apicalsurface of these cells (Bäumer et al. 2006; Dominguezet al. 2007). Together, these observations suggest thatthe expression of RPTPs of the R3 subtype may berestricted to a single or limited number of cell types. Inaddition, these RPTPs may be expressed specifically atthe apical surface of polarized cells.
Figure 6 Morphology of intestinal epithelial cells of SAP-1 KOmice. (A) Sections of the jejunum of male SAP-1 KO or WTlittermates at 10 weeks of age were stained with hematoxylin–eosin. The images show villus-crypt units (upper panels) or theepithelium (lower panels). Scale bars, 200 μm (upper panels) or20 μm (lower panels). (B) Transmission EM of the jejunumsamples shown in (A). Images of the microvilli (upper panels) andof the upper region of cell–cell adhesion of the epithelium (lowerpanels) are shown. TJ, tight junction; AJ, adherens junction; DS,desmosome. Scale bars, 500 nm.
SAP-1 is an intestinal MV-specific PTP
© 2009 The Authors Genes to Cells (2009) 14, 295–308Journal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
303
The predominant expression and localization ofSAP-1 at the microvilli of gastrointestinal epithelial cellsimplicate SAP-1 in functions specific to these cells. Theproper regulation of the actin cytoskeleton is importantfor maintenance of the morphology of intestinalmicrovilli (Fath & Burgess 1995). Indeed, mice that lackeither ezrin or EBP-50, both of which are localized atmicrovilli and associated with the actin cytoskeleton,exhibit shortened and irregular microvilli in enterocytes
(Morales et al. 2004; Saotome et al. 2004; Tamura et al.2005). Moreover, protein tyrosine phosphorylation,which is determined by protein tyrosine kinases andPTPs, plays an important role in reorganization of theactin cytoskeleton (Giannone & Sheetz 2006). However,SAP-1-deficient mice manifested no marked changes inthe morphology of intestinal epithelial cells, includingthat of microvilli or of tight or adherens junctionsbetween these cells. Consistent with this finding, the
Figure 7 Effect of SAP-1 deficiency on adenoma formation in the intestine of ApcMin/+ mice. (A) The total number of visible adenomas(≥ 1 mm in diameter) in the small intestine or colon of ApcMin/+Sap1+/+ (WT) (n = 41) or ApcMin/+Sap1–/– (KO) (n = 29) mice at 20 weeksof age was quantified by stereoscopic microscopy. (B) Size range of intestinal and colonic adenomas for the animals examined in (A). Datain (A) and (B) are means ± SEM of values from the indicated numbers of mice. *P < 0.05, **P < 0.01. (C) Adenomas fromBrdU-injected ApcMin/+Sap1+/+ (WT) (left panel) or ApcMin/+Sap1–/– (KO) (middle panel) mice were sectioned and evaluated for BrdUincorporation by immunohistochemistry with a mAb to BrdU. Scale bar, 100 μm. The percentage of BrdU-positive cells per 150adenoma cells of the two mouse strains was determined (right panel); data are means ± SEM of values from eight mice of each group.(D) Adenomas from ApcMin/+Sap1+/+ (WT) (left panel) or ApcMin/+Sap1–/– (KO) (middle panel) mice were sectioned and evaluated forcell death by TUNEL staining. Arrowheads indicate TUNEL-positive cells. Scale bar, 100 μm. The percentage of TUNEL-positivecells per 150 adenoma cells of the two mouse strains was determined. Data are means ± SEM of values from eight or twelveApcMin/+Sap1+/+ or ApcMin/+Sap1–/– mice, respectively.

H Sadakata et al.
Genes to Cells (2009) 14, 295–308 © 2009 The AuthorsJournal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
304
expression of SAP-1 in the intestine was shown to beminimal during embryonic development and to increasemarkedly after birth, suggesting that SAP-1 is not importantfor determination of cell architecture in the intestinalepithelium.
Intestinal epithelial cells play an essential role infood digestion and in the absorption of nutrients andelectrolytes through the intestine. Indeed, severaldigestive enzymes as well as channels and transportersfor electrolytes or nutrients, including those for glucose,amino acids and fatty acids, are associated with thegastrointestinal microvilli (Holmes & Lobley 1989; Ross& Pawlina 2006). However, general nutrition statusappeared unaffected in SAP-1 KO mice, as were peripheralblood cell counts. SAP-1 may thus be dispensable forregulation of food digestion and absorption of nutrientsand electrolytes in the intestine. We are not able toexclude the possibility, however, that SAP-1 indeedcontributes to such functions and that other PTPs, suchas other RPTPs of the R3 subtype, are able to compensatefor the loss of SAP-1 activity in SAP-1 KO mice.
We also found that SAP-1 deficiency inhibitedtumorigenesis in mice heterozygous for mutation of Apc.The number of large adenomas (≥ 2 mm in diameter) inSAP-1 KO mice was markedly reduced whereas thatof small adenomas (< 2 mm) was similar to that in WTmice, suggesting that SAP-1 contributes to tumorexpansion but not to the initial transformation of normalepithelial cells into dysplastic cells. Functional impair-ment of the APC protein in ApcMin/+ mice is consideredto result in stabilization and marked accumulation ofβ-catenin, which initiates transformation of normalepithelial cells and promotes tumorigenesis throughconstitutive activation of the β-catenin-transcriptionfactor 4 (TCF-4) transcriptional pathway (Clarke 2006).Our results thus suggest that SAP-1 regulates intes-tinal tumorigenesis through cooperation with thispathway, although the precise mechanism of suchregulation remains to be determined. Indeed, the ratesof cell proliferation and apoptosis in adenomas ofApcMin/+Sap1–/– mice did not differ significantly fromthose apparent in ApcMin/+Sap1+/+ mice. In addition, thecytoplasmic and nuclear accumulation of β-cateninin adenomas were similar in mice of these two genotypes(unpublished data), suggesting that SAP-1 likely does notregulate the β-catenin-TCF-4 pathway itself. Instead,SAP-1 may promote intestinal cell proliferation throughactivation of protein tyrosine kinases such as Src familykinases. Indeed, PTPα, another RPTP, was shown toinduce transformation of Rat-1 fibroblasts as a result ofactivation of Src mediated by dephosphorylation of itsCOOH-terminal tyrosine residue (Zheng et al. 1992).
However, forced expression of SAP-1 failed to increasethe activity of Src in cultured fibroblasts (unpublisheddata). Moreover, the levels of c-Src activity (in eitheradenomas or normal tissues) of ApcMin/+Sap1–/– mice didnot differ from those of ApcMin/+Sap1+/+ mice (unpub-lished data). We previously showed that the expression ofSAP-1 is markedly increased in human colon cancers(Matozaki et al. 1994; Seo et al. 1997). Our presentresults thus support the notion that SAP-1 promotes thetumorigenic potential of intestinal epithelial cells. Bycontrast, Src kinase may not be responsible for promotionby SAP-1 of intestinal tumorigenesis. Recent studiesalso suggests the transforming properties of PTPs. Gainof function mutations in SHP-2 are implicated tocause juvenile myelomonocytic leukemia (Tartaglia et al.2003). In addition, deletion of PTP1B activity delaysmammary tumorigenesis and protects lung metastasis intransgenic mice with an active mutant of ErbB2 (Julienet al. 2007). Further studies are warranted to characterizethe physiological and pathological roles of SAP-1 as anintestinal epithelium-specific PTP.
Experimental proceduresAntibodies and reagents
Mouse mAbs to β-tubulin (TUB 2.1) or to actin (AC-40) wereobtained from Sigma (St. Louis, MO), those to β-catenin or top120-catenin were from BD Biosciences (San Diego, CA), andthat to E-cadherin (ECCD-2) was from Takara BIO (Otsu, Shiga,Japan). Rabbit polyclonal antibodies to EBP-50 were from Abcam(Cambridge, UK) and those to claudin-15 were kindly providedby M. Furuse (Kobe University, Kobe, Japan). A rat mAb to CD31(MEC13.3) was from BD Biosciences and that to ezrin (M11) waskindly provided by S. Tsukita (Osaka University, Suita, Japan).Rabbit polyclonal antibodies to ZO-1, rhodamine-conjugatedphalloidin, and secondary antibodies labeled with fluoresceinisothiocyanate (FITC), Cy3 or Alexa488 for immunofluorescenceanalysis were obtained from Invitrogen (Carlsbad, CA). Horse radishperoxidase (HRP)-conjugated goat polyclonal antibodies to rat,mouse or rabbit IgG for immunoblot analysis were from JacksonImmunoResearch (West Grove, PA). FITC-conjugated UEA-Iwas from Vector (Burlingame, CA).
Isolation of a full-length mouse SAP-1 cDNA
To obtain a full-length mouse SAP-1 cDNA, we searched mousedata bases for sequences similar to that of human SAP-1 cDNAwith the use of the Basic Local Alignment Search Tool (BLAST)program (NCBI). Several EST fragments as well as genomic DNAfragments potentially corresponding to the mouse SAP-1 genewere identified and were used to obtain partial cDNAs for mouseSAP-1 by RT-PCR. The sequences of two RT-PCR products(226 and 431 bp) were then used to search the mouse DNA data

SAP-1 is an intestinal MV-specific PTP
© 2009 The Authors Genes to Cells (2009) 14, 295–308Journal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
305
base provided by the FANTOM project (Carninci et al. 2005),resulting in the identification of a cDNA clone (#9030616J04)that contained the full-length SAP-1 cDNA.
Generation of monoclonal and polyclonal antibodies to mouse SAP-1
A rat mAb to mouse SAP-1 (clone 123) was generated with theuse of a recombinant fusion protein comprising the extracellularregion of mouse SAP-1 (amino acids 1–604) and the Fc region ofhuman IgG as the antigen. Rabbit polyclonal antibodies to mouseSAP-1 were also generated with the use of a glutathione-S-transferase fusion protein containing the cytoplasmic region ofmouse SAP-1 (amino acids 648–971) as antigen, as previouslydescribed (Matozaki et al. 1994).
Northern blot analysis
A digoxigenin-labeled cRNA probe corresponding to nt –31 to3454 of mouse SAP-1 cDNA was synthesized with the use of aDIG RNA Labeling Kit (Roche, Basel, Switzerland) and T7RNA polymerase. Polyadenylated RNA (2 μg) fixed on a nylonmembrane was subjected to hybridization overnight at 50 °Cwith 400 ng of the cRNA probe in DIG Easy Hyb (Roche). Themembrane was then exposed to alkaline phosphatase–conjugatedsheep polyclonal Fab fragments to digoxigenin (1 : 10 000 dilution;Roche), and hybridization complexes were visualized with theCDP-Star reagent (Roche).
Immunoblot analysis
Mouse tissues were homogenized on ice in lysis buffer (20 mmTris–HCl (pH 7.6), 140 mm NaCl, 1 mm EDTA, 1% NonidetP-40) containing 1 mm phenylmethylsulfonyl fluoride, aprotinin(10 μg/mL), and 1 mm sodium vanadate. The lysates were centri-fuged at 10 000 g for 15 min at 4 °C, and the resulting supernatantswere subjected to immunoblot analysis.
Histological and immunofluorescence analyses
For analysis of mouse tissues, mice were anesthetized by intraperi-toneal (i.p.) injection of sodium pentobarbital at 25 mg/kg ofweight and were then perfused transcardially with 4% paraformal-dehyde (PFA) in phosphate buffered saline (PBS). Tissues werethen dissected, fixed again with 4% PFA in PBS for 1 h at roomtemperature, transferred to a series of sucrose solutions [7%, 20%and 30% (w/v), sequentially] in PBS, embedded in optimalcutting temperature (OCT) compound (Sakura, Tokyo, Japan),and rapidly frozen in liquid nitrogen. Frozen sections of 5 μmthickness were then prepared and subjected to immunofluor-escence analysis with primary antibodies and secondary antibodieslabeled with fluorescent dyes, as described previously (Ishikawa-Sekigami et al. 2006). Fluorescence images were obtained with aconfocal laser-scanning microscope (LSM Pascal; Carl Zeiss, Jena,Germany). To examine the expression of alkaline phosphatase, we
used a Vector Red Alkaline Phosphatase Kit I (Vector Laboratories,Burlingame, CA). Fixed tissue samples were also embedded inparaffin, sectioned, and stained with Mayer’s hematoxylin–eosin.
Transmission EM
Mice were anesthetized as described above and were then perfusedtranscardially with 2% PFA and 2.5% glutaraldehyde in 0.1 msodium cacodylate buffer (pH 7.4). Tissues were removed,immersed in the same fixative for 1 h at 4 °C, and then incubatedfor 1 h at 4 °C with 1% OsO4 in the same buffer. They were thendehydrated and embedded in Epon. For immuno-EM, tissuesamples were fixed by immersion for 2 h at room temperature inPBS containing 4% PFA and 1% glutaraldehyde. Frozen sectionsof 10 μm thickness were then prepared and processed for immuno-staining with a mAb to mouse SAP-1, and with Fab fragmentsof goat antibodies to rat IgG that were labeled with 1.4-nm goldparticles (Nanoprobes, Yaphank, NY); signals were enhanced withthe use of an HQ Silver Enhancement kit (Nanoprobes). Thesections were fixed again with 1% OsO4 and 0.1% potassiumferrocyanide and were finally embedded in Epon. Ultrathin sections(90 nm) were prepared, stained with uranyl acetate and lead citrate,and examined with a JEM 1010 electron microscope ( JEOL,Tokyo, Japan).
Generation of SAP-1-deficient mice
To generate SAP-1-deficient mice, we first obtained two bacterialartificial chromosome clones containing 129/Sv mouse Sap1genomic DNA from BACPAC Resources Center (Children’sHospital Oakland Research Institute, Oakland, CA). The targetingvector (Fig. 5A) was constructed with a 1.2-kb DNA fragmentspanning exon 14 to intron 16 as a 5′ homologous region, with a6.4-kb fragment spanning intron 16 to intron 21 as a 3′ homologousregion, with a construct (PGKβgeo) comprising the promoterof the mouse phosphoglycerate kinase 1 gene linked to a β-galactosidase–neomycin phosphotransferase fusion gene (Frie-drich & Soriano 1991), and with a diphtheria toxin A chain(DTA) gene cassette (Takara BIO). A LoxP site was also insertedinto intron 16 as well as into intron 18 of the targeting vector.The targeting vector was introduced into the J1 line of mouseembryonic stem cells by electroporation, and homologous recom-bination was confirmed by Southern blot analysis, as describedpreviously (Harada et al. 1994), with two DNA probes (S1 and S2)(Fig. 5A). Chromosomal DNA was thus purified from each cellclone and was digested with Hind III (for the S1 probe) or BamHI (for the S2 probe). We crossed transgenic mice that express Crerecombinase under the control of CAG promoter (CAG-Cremice) (Sakai & Miyazaki 1997) with mice harboring a targetedSap1 allele. The genotype of the resulting offspring wasdetermined by either Southern blot (Fig. 5B) or PCR analysis. InPCR analysis, 396- and 496-bp DNA fragments correspondingto the wild-type and targeted alleles, respectively, were generatedwith the sense primer MSAP-MS (5′-ATGGAGAGACTGGCTCTTGC-3′) and the antisense primer MSAP LOX-AS(5′-CCATGCAGCTAGTAACATGG-3′) and a 300-bp DNA

H Sadakata et al.
Genes to Cells (2009) 14, 295–308 © 2009 The AuthorsJournal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
306
fragment corresponding to the deleted allele was generated withthe primers Lox-g (5′-ACGGTATCGATAAGCTTGGC-3′) andMSAP LOX-AS. SAP-1-KO mice were then backcrossed ontothe C57BL/6 background for four generations. Mice were bredand maintained at the Institute of Experimental Animal Researchof Gunma University under specific pathogen-free conditions.All animal experiments were performed with the guidelines ofthe Animal Care and Experimentation Committee of GunmaUniversity.
Adenoma formation in SAP-1-deficient mice with the ApcMin allele
ApcMin/+ mice (C57BL/6J-ApcMin/J) were obtained from the JacksonLaboratory (Bar Harbor, ME) and were crossed with SAP-1 KOmice. The resulting ApcMin/+Sap1+/– offspring were crossed togenerate ApcMin/+Sap1–/– mice and their ApcMin/+Sap1+/+ littermatesfor comparison. The small intestine and colon were removedfrom mice at 20 weeks of age and were fixed in methacarn(methanol : chloroform : acetic acid, 4 : 2 : 1, v/v) for 3 min(Prokhortchouk et al. 2006). The number and diameter of adenomaswere determined with the use of a stereoscopic microscope(SZX7; Olympus, Tokyo, Japan) at a magnification of 10×. Tissuewas also embedded in paraffin for histological analysis.
BrdU incorporation assay
Mice were injected i.p. with BrdU (50 μg/g of body weight inPBS) and killed 1 h later. Intestinal tissue was fixed in methacarn,embedded in paraffin, sectioned, and stained with a mAb to BrdUwith the use of a BrdU In-Situ Detection Kit (BD Pharmingen,Franklin Lakes, NJ). The BrdU labeling index was defined as thepercentage of BrdU-positive cells per 150 adenoma cells.
Apoptosis assay
Intestinal tissue was fixed in methacarn, transferred to a series ofsucrose solutions [7%, 20% and 30% (w/v), sequentially], embeddedin OCT compound, and evaluated for apoptosis with an apoptosisdetection kit (Takara BIO). The number of apoptotic cells per 150adenoma cells was determined.
Statistical analysis
Data are presented as means ± SEM and were analyzed by Student’st test. A P value of < 0.05 was considered statistically significant.
AcknowledgementsWe thank S. Tsukita and M. Furuse for antibodies, Y. Hayashizakifor a RIKEN full-length cDNA clone (#9030616J04), J. Miyazakifor CAG-Cre mice, T. Fujimoto for his suggestion and instructionfor EM analysis, as well as K. Tomizawa, H. Kobayashi, Y. Hayashi,Y. Niwayama and T. Horie for technical assistance. This work wassupported by a Grant-in-Aid for Scientific Research on Priority
Areas Cancer, a Grant-in-Aid for Scientific Research (B), and agrant of the Global Center of Excellence Program from theMinistry of Education, Culture, Sports, Science and Technologyof Japan.
ReferencesAndersen, J.N., Mortensen, O.H., Peters, G.H., Drake, P.G.,
Iversen, L.F., Olsen, O.H., Jansen, P.G., Andersen, H.S., Tonks,N.K. & Møller, N.P. (2001) Structural and evolutionaryrelationships among protein tyrosine phosphatase domains.Mol. Cell. Biol. 21, 7117–7136.
Bäumer, S., Keller, L., Holtmann, A., Funke, R., August, B.,Gamp, A. Wolburg, H., Wolburg-Buchholz, K., Deutsch, U. &Vestweber, D. (2006) Vascular endothelial cell-specific phos-photyrosine phosphatase (VE-PTP) activity is required forblood vessel development. Blood 107, 4754–4762.
Benzing, T. (2004) Signaling at the slit diaphragm. J. Am. Soc.Nephrol. 15, 1382–1391.
Blanpain, C., Horsley, V. & Fuchs, E. (2007) Epithelial stem cells:turning over new leaves. Cell 128, 445–458.
Carninci, P., Kasukawa, T., Katayama, S., et al. (2005) Thetranscriptional landscape of the mammalian genome. Science309, 1559–1563.
Clarke, A.R. (2006) Wnt signalling in the mouse intestine.Oncogene 25, 7512–7521.
Dominguez, M.G., Hughes, V.C., Pan, L., Simmons, M., Daly,C., Anderson, K., Noguera-Troise, I., Murphy, A.J.,Valenzuela, D.M., Davis, S., Thurston, G., Yancopoulos, G.D.& Gale, N.W. (2007) Vascular endothelial tyrosine phosphatase(VE-PTP)-null mice undergo vasculogenesis but die embryon-ically because of defects in angiogenesis. Proc. Natl Acad. Sci.USA 104, 3243–3248.
Fath, K.R. & Burgess, D.R. (1995) Microvillus assembly. Notactin alone. Curr. Biol. 5, 591–593.
Fischer, J., Klein, P.J., Vierbuchen, M., Skutta, B., Uhlenbruck, G.& Fischer, R. (1984) Characterization of glycoconjugates ofhuman gastrointestinal mucosa by lectins. I. Histochemicaldistribution of lectin binding sites in normal alimentary tract aswell as in benign and malignant gastric neoplasms. Histochem.Cytochem. 32, 681–689.
Friedrich, G. & Soriano, P. (1991) Promoter traps in embryonicstem cells: a genetic screen to identify and mutate developmentalgenes in mice. Genes Dev. 5, 1513–1523.
Giannone, G. & Sheetz, M.P. (2006) Substrate rigidity and forcedefine form through tyrosine phosphatase and kinase pathways.Trends Cell Biol. 16, 213–223.
Gumbiner, B.M. (2005) Regulation of cadherin-mediatedadhesion in morphogenesis. Nat. Rev. Mol. Cell Biol. 6, 622–634.
Harada, A., Oguchi, K., Okabe, S., Kuno, J., Terada, S., Ohshima,T., Sato-Yoshitake, R., Takei, Y., Noda, T. & Hirokawa, N.(1994) Altered microtubule organization in small-calibre axonsof mice lacking tau protein. Nature 369, 488–491.
Holmes, R. & Lobley, R.W. (1989) Intestinal brush borderrevisited. Gut 30, 1667–1678.

SAP-1 is an intestinal MV-specific PTP
© 2009 The Authors Genes to Cells (2009) 14, 295–308Journal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
307
Ishikawa-Sekigami, T., Kaneko, Y., Okazawa, H., Tomizawa, T.,Okajo, J., Saito, Y., Okuzawa, C., Sugawara-Yokoo, M.,Nishiyama, U., Ohnishi, H., Matozaki, T. & Nojima, Y. (2006)SHPS-1 promotes the survival of circulating erythrocytesthrough inhibition of phagocytosis by splenic macrophages.Blood 107, 341–348.
Ito, T., Okazawa, H., Maruyama, K., Tomizawa, K., Motegi, S.,Ohnishi, H., Kuwano, H., Kosugi, A. & Matozaki, T. (2003)Interaction of SAP-1, a transmembrane-type protein-tyrosinephosphatase, with the tyrosine kinase Lck. Roles in regulationof T cell function. J. Biol. Chem. 278, 34854–34863.
Julien, S.G., Dubé, N., Read, M., Penney, J., Paquet, M., Han, Y.,Kennedy, B.P., Muller, W.J. & Tremblay, M.L. (2007) Proteintyrosine phosphatase 1B deficiency or inhibition delaysErbB2-induced mammary tumorigenesis and protects fromlung metastasis. Nat. Genet. 39, 338–346.
Kamei, T., Matozaki, T., Sakisaka, T., Kodama, A., Yokoyama, S.,Peng, Y.F., Nakano, K., Takaishi, K. & Takai, Y. (1999)Coendocytosis of cadherin and c-Met coupled to disruption ofcell–cell adhesion in MDCK cells—regulation by Rho, Racand Rab small G proteins. Oncogene 18, 6776–6784.
Lilien, J. & Balsamo, J. (2005) The regulation of cadherin-mediated adhesion by tyrosine phosphorylation/dephosphor-ylation of β-catenin. Curr. Opin. Cell Biol. 17, 459–465.
Maratos-Flier, E., Kao, C.Y., Verdin, E.M. & King, G.L. (1987)Receptor-mediated vectorial transcytosis of epidermal growthfactor by Madin–Darby canine kidney cells. J. Cell Biol. 105,1595–1601.
Matozaki, T., Suzuki, T., Uchida, T., Inazawa, J., Ariyama, T.,Matsuda, K., Hirita, K., Noguchi, H., Mizuno, H., Sakamoto,C. & Kasuga, M. (1994) Molecular cloning of a humantransmembrane-type protein tyrosine phosphatase and its expres-sion in gastrointestinal cancers. J. Biol. Chem. 269, 2075–2081.
Miyoshi, J. & Takai, Y. (2005) Molecular perspective on tight-junction assembly and epithelial polarity. Adv. Drug Deliv. Rev.57, 815–855.
Mooseker, M.S. (1985) Organization, chemistry, and assembly ofthe cytoskeletal apparatus of the intestinal brush border. Annu.Rev. Cell Biol. 1, 209–241.
Morales, F.C., Takahashi, Y., Kreimann, E.L. & Georgescu, M.M.(2004) Ezrin–radixin–moesin (ERM)-binding phosphoprotein50 organizes ERM proteins at the apical membrane of polarizedepithelia. Proc. Natl. Acad. Sci. USA 101, 17705–17710.
Narisawa, S., Huang, L., Iwasaki, A., Hasegawa, H., Alpers, D.H.& Millan, J.L. (2003) Accelerated fat absorption in intestinalalkaline phosphatase knockout mice. Mol. Cell. Biol. 23, 7525–7530.
Noguchi, T., Tsuda, M., Takeda, H., Takada, T., Inagaki, K.,Yamao, T., Fukunaga, K., Matozaki, T. & Kasuga, M. (2001)Inhibition of cell growth and spreading by stomach cancer-associated protein-tyrosine phosphatase-1 (SAP-1) throughdephosphorylation of p130cas. J. Biol. Chem. 276, 15216–15224.
Prokhortchouk, A., Sansom, O., Selfridge, J., et al. (2006)Kaiso-deficient mice show resistance to intestinal cancer. Mol.Cell. Biol. 26, 199–208.
Rakoff-Nahoum, S. & Medzhitov, R. (2007) Regulation ofspontaneous intestinal tumorigenesis through the adaptorprotein MyD88. Science 317, 124–127.
Ross, M.H. & Pawlina, W. (2006) Histology. In: A Text and Atlaswith Correlated Cell and Molecular Biology, 5th edn, pp. 102–121,518–575. Baltimore: Lippincott Williams & Wilkins.
Sakai, K. & Miyazaki, J. (1997) A transgenic mouse line thatretains Cre recombinase activity in mature oocytes irrespectiveof the cre transgene transmission. Biochem. Biophys. Res. Commun.237, 318–324.
Sallee, J.L., Wittchen, E.S. & Burridge, K. (2006) Regulation ofcell adhesion by protein–tyrosine phosphatases: II. Cell–celladhesion. J. Biol. Chem. 281, 16189–16192.
Saotome, I., Curto, M. & McClatchey, A.I. (2004) Ezrin isessential for epithelial organization and villus morphogenesis inthe developing intestine. Dev. Cell 6, 855–864.
Sena, P., Saviano, M., Monni, S., Losi, L., Roncucci, L., Marzona,L. & De Pol, A. (2006) Subcellular localization of β-catenin andAPC proteins in colorectal preneoplastic and neoplastic lesions.Cancer Lett. 241, 203–212.
Seo, Y., Matozaki, T., Tsuda, M., Hayashi, Y., Itoh, H. & Kasuga,M. (1997) Overexpression of SAP-1, a transmembrane-typeprotein tyrosine phosphatase, in human colorectal cancers.Biochem. Biophys. Res. Commun. 231, 705–711.
Su, L.K., Kinzler, K.W., Vogelstein, B., Preisinger, A.C., Moser,A.R., Luongo, C., Gould, K.A. & Dove, W.F. (1992) Multipleintestinal neoplasia caused by a mutation in the murinehomolog of the APC gene. Science 256, 668–670.
Takada, T., Noguchi, T., Inagaki, K., Hosooka, T., Fukunaga, K.,Yamao, T., Ogawa, W., Matozaki, T. & Kasuga, M. (2002)Induction of apoptosis by stomach cancer-associated protein–tyrosine phosphatase-1. J. Biol. Chem. 277, 34359–34366.
Takeda, H., Nagafuchi, A., Yonemura, S., Tsukita, S., Behrens, J.,Birchmeier, W. & Tsukita, S. (1995) V-src kinase shifts thecadherin-based cell adhesion from the strong to the weak stateand β-catenin is not required for the shift. J. Cell Biol. 131,1839–1847.
Takeichi, M. (1995) Morphogenetic roles of classic cadherins.Curr. Opin. Cell Biol. 7, 619–627.
Tamura, A., Kikuchi, S., Hata, M., Katsuno, T., Matsui, T., Hayashi,H., Suzuki, Y., Noda, T., Tsukita, S. & Tsukita, S. (2005)Achlorhydria by ezrin knockdown: defects in the formation/expansion of apical canaliculi in gastric parietal cells. J. Cell Biol.169, 21–28.
Tartaglia, M., Niemeyer, C.M., Fragale, A., Song, X., Buechner,J., Jung, A., Hählen, K., Hasle, H., Licht, J.D. & Gelb, B.D.(2003) Somatic mutations in PTPN11 in juvenile myelomono-cytic leukemia, myelodysplastic syndromes and acute myeloidleukemia. Nat. Genet. 34, 148–150.
Thomas, P.E., Wharram, B.L., Goyal, M., Wiggins, J.E., Holzman,L.B. & Wiggins, R.C. (1994) GLEPP1, a renal glomerularepithelial cell (podocyte) membrane protein–tyrosine phos-phatase. Identification, molecular cloning, and characterizationin rabbit. J. Biol. Chem. 269, 19953–19962.
Tsukita, S. & Furuse, M. (2002) Claudin-based barrier in simpleand stratified cellular sheets. Curr. Opin. Cell Biol. 14, 531–536.

H Sadakata et al.
Genes to Cells (2009) 14, 295–308 © 2009 The AuthorsJournal compilation © 2009 by the Molecular Biology Society of Japan/Blackwell Publishing Ltd.
308
Tsukita, S. & Yonemura, S. (1999) Cortical actin organization:lessons from ERM (ezrin/radixin/moesin) proteins. J. Biol.Chem. 274, 34507–34510.
Tyska, M.J., Mackey, A.T., Huang, J.D., Copeland, N.G., Jenkins,N.A. & Mooseker, M.S. (2005) Myosin-1a is critical for normalbrush border structure and composition. Mol. Biol. Cell 16:2443–2457.
Wharram, B.L., Goyal, M., Gillespie, P.J., Wiggins, J.E., Kershaw,D.B., Holzman, L.B., Dysko, R.C., Sanders, T.L., Samuelson,L.C. & Wiggins, R.C. (2000) Altered podocyte structure inGLEPP1 (Ptpro)-deficient mice associated with hypertensionand low glomerular filtration rate. J. Clin. Invest. 106, 1281–1290.
Yang, D.H., Goyal, M., Sharif, K., Kershaw, D., Thomas, P.,Dysko, R. & Wiggins, R. (1996) Glomerular epithelial protein1 and podocalyxin-like protein 1 in inflammatory glomerulardisease (crescentic nephritis) in rabbit and man. Lab. Invest. 74,571–584.
Zheng, X.M., Wang, Y. & Pallen, C.J. (1992) Cell transformationand activation of pp60c-src by overexpression of a proteintyrosine phosphatase. Nature 359, 336–339.
Received: 5 November 2008Accepted: 21 November 2008




















