
Sphingosine Forms Channels in Membranes That Differ Greatly from Those Formed by Ceramide
Upload
independentCategory
view
2download
0
0014-2980/99/0909-2923$17.50+.50/0© WILEY-VCH Verlag GmbH, D-69451 Weinheim, 1999
Greatly reduced efficiency of both positive andnegative selection of thymocytes in CD45 tyrosinephosphatase-deficient mice
P. Joseph Mee, Martin Turner, M. Albert Basson, Patrick S. Costello, Rose Zamoyskaand Victor L. J. Tybulewicz
National Institute for Medical Research, The Ridgeway, Mill Hill, London, NW7 1AA, UK
The T cell repertoire is shaped by positive and negative selection of thymocytes. TCR-mediated signals that determine these selection processes are only partly understood. TheCD45 tyrosine phosphatase has been shown to be important for signal transduction throughthe TCR, but there has been disagreement about whether CD45 is a positive or negative reg-ulator of TCR signaling. Using CD45-deficient mice expressing transgenic TCR, we showthat in the absence of CD45 there is a large increase in the thresholds of TCR stimulationrequired for both positive and negative selection. Our results conclusively demonstrate thatin double-positive thymocytes CD45 is a positive regulator of the TCR signals that drive thy-mic selection events.
Key words: CD45 / T cell selection / Tyrosine phosphatase / Thymus / Gene targeting
Received 22/4/99Revised 22/6/99Accepted 24/6/99
[I 19566]
Abbreviations: DN: Double negative DP: Double positiveES: Embryonic stem NTOC: Neonatal thymic organ cultureSEB: Staphylococcal enterotoxin B SP: Single positive
1 Introduction
The T cell repertoire is shaped by the positive and nega-tive selection of immature CD4+CD8+ double-positive(DP) thymocytes. The outcome of these selection eventsis determined by interactions between the T cell receptor(TCR) on DP thymocytes and peptides presented by theclass I and class II molecules of the major histocompati-bility complex (MHC) expressed on thymic stroma [1];however, the TCR-mediated signals that determine theseprocesses are only partially understood.
CD45 is a transmembrane tyrosine phosphatase impli-cated in the control of TCR signaling. Most CD45-deficient T cell lines show defective TCR signaling [2–4].Analysis in some cell lines has shown that in the absenceof CD45 the Src-family kinases Lck and Fyn are muchless active and hyperphosphorylated on a C-terminal“negative regulatory” tyrosine [5–8]. Thus, it has beenproposed that the function of CD45 is to dephosphory-late this tyrosine, thereby priming the Src-family kinases,ready for their activation following TCR stimulation.However, other studies have come to opposite conclu-
sions, showing that in other CD45-deficient T cell linesLck and Fyn have increased kinase activity due toincreased phosphorylation of a “positive regulatory”tyrosine within the kinase domain, suggesting that thefunction of CD45 is to dephosphorylate this residue andinactivate the Src-family kinases [9–11]. Recent experi-ments showing that CD45 can antagonize the ability ofLck to transform fibroblasts also support the notion thatCD45 is a negative regulator of Src-family kinases [12].Thus, while CD45 is acknowledged to be important forTCR signaling, there is disagreement about whether itacts as a positive or negative regulator of this process.
A role for CD45 in T cell selection in vivo was suggestedby a partial block in thymic development between DPand CD4+ or CD8+ single-positive (SP) thymocytes inmice with a targeted mutation in exon 6 of the Cd45 gene(Cd45ex6/ex6) [13]. However, interpretation of phenotype inthese mice was complicated by the observation that asmany as 40 % of mature T cells still expressed surface-bound CD45 [14]. Subsequently, analysis of a mousestrain with a disruption of exon 9 of the Cd45 gene(Cd45ex9/ex9), which expresses no detectable CD45, con-firmed that the transition from DP to SP thymocytes wasseverely disrupted by the absence of CD45 [15]. Such adevelopmental block could result from defective positiveor negative selection or both. In this study we haveextended the analysis of the role of CD45 in T cell devel-opment in a novel CD45-deficient mouse strain. Usingmice carrying transgenic TCR we show that both posi-
Eur. J. Immunol. 1999. 29: 2923–2933 Positive and negative thymocyte selection in Cd45–/– mice 2923
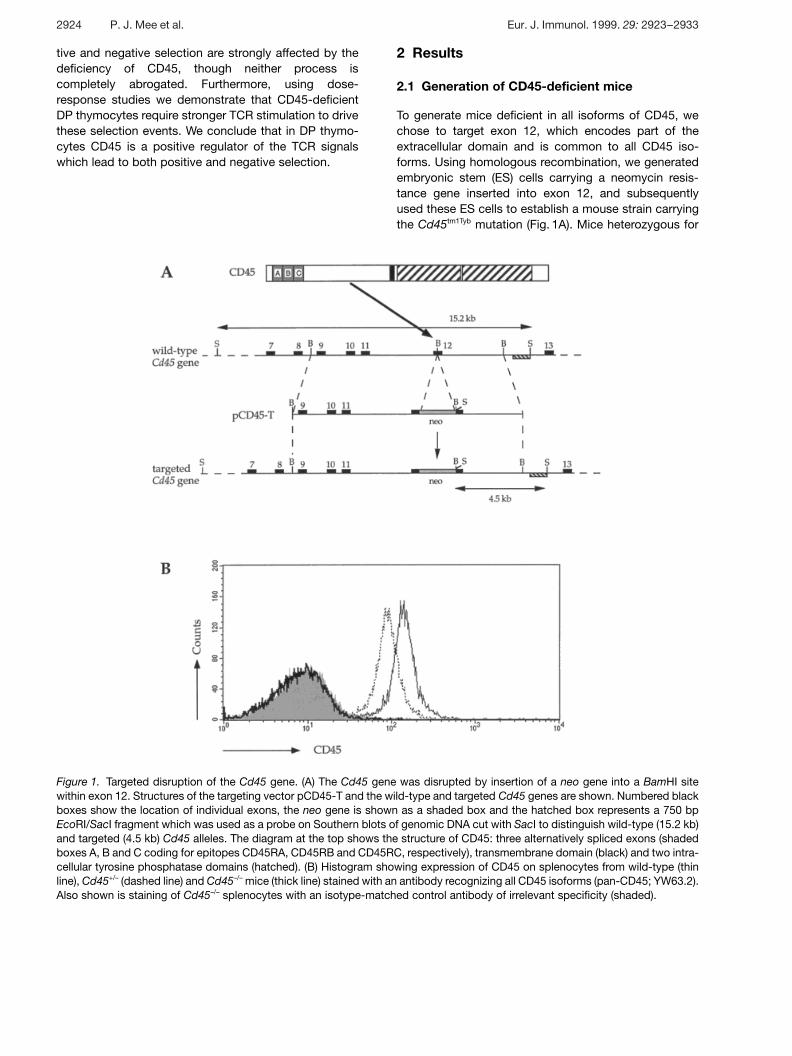
Figure 1. Targeted disruption of the Cd45 gene. (A) The Cd45 gene was disrupted by insertion of a neo gene into a BamHI sitewithin exon 12. Structures of the targeting vector pCD45-T and the wild-type and targeted Cd45 genes are shown. Numbered blackboxes show the location of individual exons, the neo gene is shown as a shaded box and the hatched box represents a 750 bpEcoRI/SacI fragment which was used as a probe on Southern blots of genomic DNA cut with SacI to distinguish wild-type (15.2 kb)and targeted (4.5 kb) Cd45 alleles. The diagram at the top shows the structure of CD45: three alternatively spliced exons (shadedboxes A, B and C coding for epitopes CD45RA, CD45RB and CD45RC, respectively), transmembrane domain (black) and two intra-cellular tyrosine phosphatase domains (hatched). (B) Histogram showing expression of CD45 on splenocytes from wild-type (thinline), Cd45+/– (dashed line) and Cd45–/– mice (thick line) stained with an antibody recognizing all CD45 isoforms (pan-CD45; YW63.2).Also shown is staining of Cd45–/– splenocytes with an isotype-matched control antibody of irrelevant specificity (shaded).
tive and negative selection are strongly affected by thedeficiency of CD45, though neither process iscompletely abrogated. Furthermore, using dose-response studies we demonstrate that CD45-deficientDP thymocytes require stronger TCR stimulation to drivethese selection events. We conclude that in DP thymo-cytes CD45 is a positive regulator of the TCR signalswhich lead to both positive and negative selection.
2 Results
2.1 Generation of CD45-deficient mice
To generate mice deficient in all isoforms of CD45, wechose to target exon 12, which encodes part of theextracellular domain and is common to all CD45 iso-forms. Using homologous recombination, we generatedembryonic stem (ES) cells carrying a neomycin resis-tance gene inserted into exon 12, and subsequentlyused these ES cells to establish a mouse strain carryingthe Cd45tm1Tyb mutation (Fig. 1A). Mice heterozygous for
2924 P. J. Mee et al. Eur. J. Immunol. 1999. 29: 2923–2933

this mutation (Cd45+/–) were intercrossed to generatehomozygous mutant mice (Cd45–/–). On a segregatinggenetic background of 129/Sv, C57BL/6 and DBA/2strains, the homozygous mutant mice were grossly nor-mal and fertile. In contrast, when inbred on the 129/Svbackground, the mutants exhibited severe diarrhoea anddecreased viability beginning around the time of wean-ing, at 3 weeks of age, such that typically less than 25 %of the mutants survived to adulthood. This phenotype isstrain specific, since Cd45–/– mice inbred on the BALB/cbackground showed no decreased viability, althoughthey often presented with splenomegaly.
Flow cytometric analysis of splenocytes, thymocytes,lymph node and bone marrow cells from Cd45–/– micewith an antibody recognizing all CD45 isoforms (pan-CD45) showed no detectable CD45 on any cells (Fig. 1Band data not shown). Furthermore, analysis of mutantcells from the same organs with antibodies specific forthe CD45RA, CD45RB and CD45RC isoforms, againshowed no detectable protein expression (not shown);note that the exon 12 mutation does not disrupt any ofthe exons coding for the epitopes recognized by theseisoform-specific antibodies. To extend this analysis fur-ther, we immunoblotted total cytoplasmic protein fromCd45–/– splenocytes, thymocytes and primary mast cellcultures with an antiserum specific for the cytoplasmicdomain of CD45 (R02.3, gift of P. Johnson); this showedthe presence of a truncated ˚ 150-kDa form of CD45 inall mutant cell types analyzed, at approximately 5 % ofthe level of wild-type CD45 (not shown). In conclusion,the exon 12 mutation has eliminated the production offull-length membrane-associated CD45. A low level of atruncated form of the protein is still made; however, sinceno CD45 could be detected on the outside of the cell byflow cytometry, it is likely that this protein is not mem-brane associated. Reinitiation of translation after exon12, or a fused translation product of the neo gene andexons 13–33 of Cd45, would result in a protein whichwould not be membrane-localized since it would notposses a signal sequence.
2.2 Altered thymic development in Cd45–/– mice
Thymic cellularity of Cd45–/– mice was typically 40 % thatof control mice. This can be almost entirely accountedfor by a 2.6-fold reduction in the number of CD4+CD8+
DP thymocytes (Fig. 2A, note logarithmic scale). Theabsolute number of CD4–CD8– double-negative (DN)cells was similar between mutant and control mice. TheDN compartment was further analyzed for expression ofCD44 and CD25, which can be used to define four sub-sets of cells (DN1–DN4) [16]. CD45-deficient thymusescontained normal numbers of the earliest populations
(DN1, CD44+CD25– and DN2, CD44+CD25+) but showeda fivefold increase in the numbers of DN3, CD44–CD25+
cells (Fig. 2B). A developmental block between the DN3,CD44–CD25+ and DN4, CD44–CD25– subsets of DN thy-mocytes has been reported in mice mutant for Rag-1,TCR g and preT § , suggesting that this transition isdependent on signaling through the pre-TCR and is usedto monitor for successful rearrangement of TCR g [17].The existence of a similar, although not complete, blockin Cd45–/– mice suggests that CD45 may play a role inpre-TCR signaling. Alternatively, it may be involved in thetransition of DN to DP cells or their subsequent survivalor expansion.
2.3 Positive selection of both class I and class II-restricted thymocytes is severelycompromised in Cd45–/– mice
In addition to the reduction in numbers of DP thymo-cytes, CD45-deficient mice showed a more dramaticreduction (7–13-fold) in the number of CD4+ and CD8+
SP cells, suggestive of a block in positive selection(Fig. 2A, C). A similar decrease was also seen in the num-bers of CD4+ and CD8+ peripheral T cells in the spleenand lymph nodes of the mutant mice (not shown). A hall-mark of DP thymocytes undergoing positive selection isan increase in TCR expression from intermediate to highlevels and the induction of expression of CD69. Flowcytometric analysis of CD45-deficient DP thymocytesdemonstrated that they contained many fewer cells thatwere TCRhi or CD69+, consistent with a failure of positiveselection (Fig. 2C). CD45-deficient DP thymocytes alsoshowed other anomalies; they expressed lower levels ofCD5 and higher levels of CD4 and CD8. In addition, theintermediate level of TCR expression found on most DPcells was slightly higher in the Cd45–/– mice than in con-trols (Fig. 2C).
Since CD45 is expressed in multiple hemopoietic line-ages, it was possible that this altered thymic develop-ment was a consequence of CD45 deficiency in a non-Tcell. To address this issue we reconstituted irradiatedmice with a mixture of wild-type and CD45-deficientbone marrow. Flow cytometric analysis demonstratedthat in these mixed chimeras, CD45-deficient thymo-cytes still did not undergo normal development, butrecapitulated the large decrease in numbers of SP thy-mocytes and peripheral T cells (not shown). In contrast,in the same mice, thymic development from the wild-type marrow was normal. Thus, the effect of the CD45mutation on T cell development is cell autonomous, sug-gesting that CD45 plays a role in selection-induced sig-nal transduction events within the thymocytes them-selves.
Eur. J. Immunol. 1999. 29: 2923–2933 Positive and negative thymocyte selection in Cd45–/– mice 2925
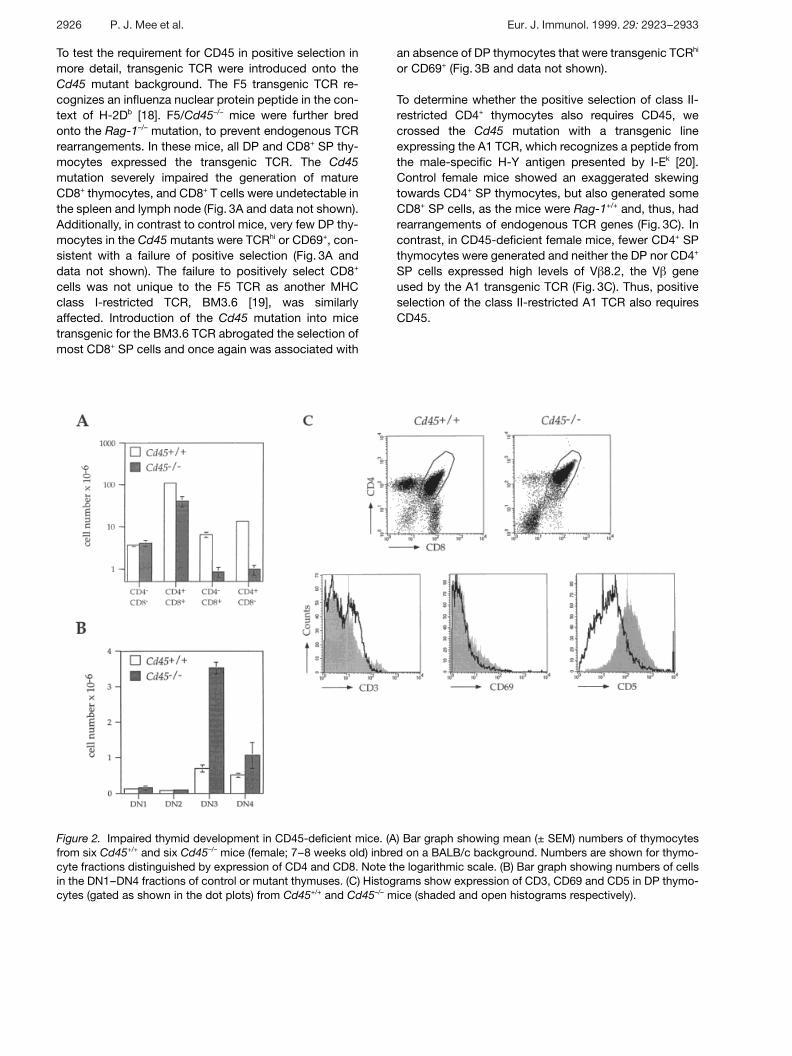
Figure 2. Impaired thymid development in CD45-deficient mice. (A) Bar graph showing mean (± SEM) numbers of thymocytesfrom six Cd45+/+ and six Cd45–/– mice (female; 7–8 weeks old) inbred on a BALB/c background. Numbers are shown for thymo-cyte fractions distinguished by expression of CD4 and CD8. Note the logarithmic scale. (B) Bar graph showing numbers of cellsin the DN1–DN4 fractions of control or mutant thymuses. (C) Histograms show expression of CD3, CD69 and CD5 in DP thymo-cytes (gated as shown in the dot plots) from Cd45+/+ and Cd45–/– mice (shaded and open histograms respectively).
To test the requirement for CD45 in positive selection inmore detail, transgenic TCR were introduced onto theCd45 mutant background. The F5 transgenic TCR re-cognizes an influenza nuclear protein peptide in the con-text of H-2Db [18]. F5/Cd45–/– mice were further bredonto the Rag-1–/– mutation, to prevent endogenous TCRrearrangements. In these mice, all DP and CD8+ SP thy-mocytes expressed the transgenic TCR. The Cd45mutation severely impaired the generation of matureCD8+ thymocytes, and CD8+ T cells were undetectable inthe spleen and lymph node (Fig. 3A and data not shown).Additionally, in contrast to control mice, very few DP thy-mocytes in the Cd45 mutants were TCRhi or CD69+, con-sistent with a failure of positive selection (Fig. 3A anddata not shown). The failure to positively select CD8+
cells was not unique to the F5 TCR as another MHCclass I-restricted TCR, BM3.6 [19], was similarlyaffected. Introduction of the Cd45 mutation into micetransgenic for the BM3.6 TCR abrogated the selection ofmost CD8+ SP cells and once again was associated with
an absence of DP thymocytes that were transgenic TCRhi
or CD69+ (Fig. 3B and data not shown).
To determine whether the positive selection of class II-restricted CD4+ thymocytes also requires CD45, wecrossed the Cd45 mutation with a transgenic lineexpressing the A1 TCR, which recognizes a peptide fromthe male-specific H-Y antigen presented by I-Ek [20].Control female mice showed an exaggerated skewingtowards CD4+ SP thymocytes, but also generated someCD8+ SP cells, as the mice were Rag-1+/+ and, thus, hadrearrangements of endogenous TCR genes (Fig. 3C). Incontrast, in CD45-deficient female mice, fewer CD4+ SPthymocytes were generated and neither the DP nor CD4+
SP cells expressed high levels of V g 8.2, the V g geneused by the A1 transgenic TCR (Fig. 3C). Thus, positiveselection of the class II-restricted A1 TCR also requiresCD45.
2926 P. J. Mee et al. Eur. J. Immunol. 1999. 29: 2923–2933
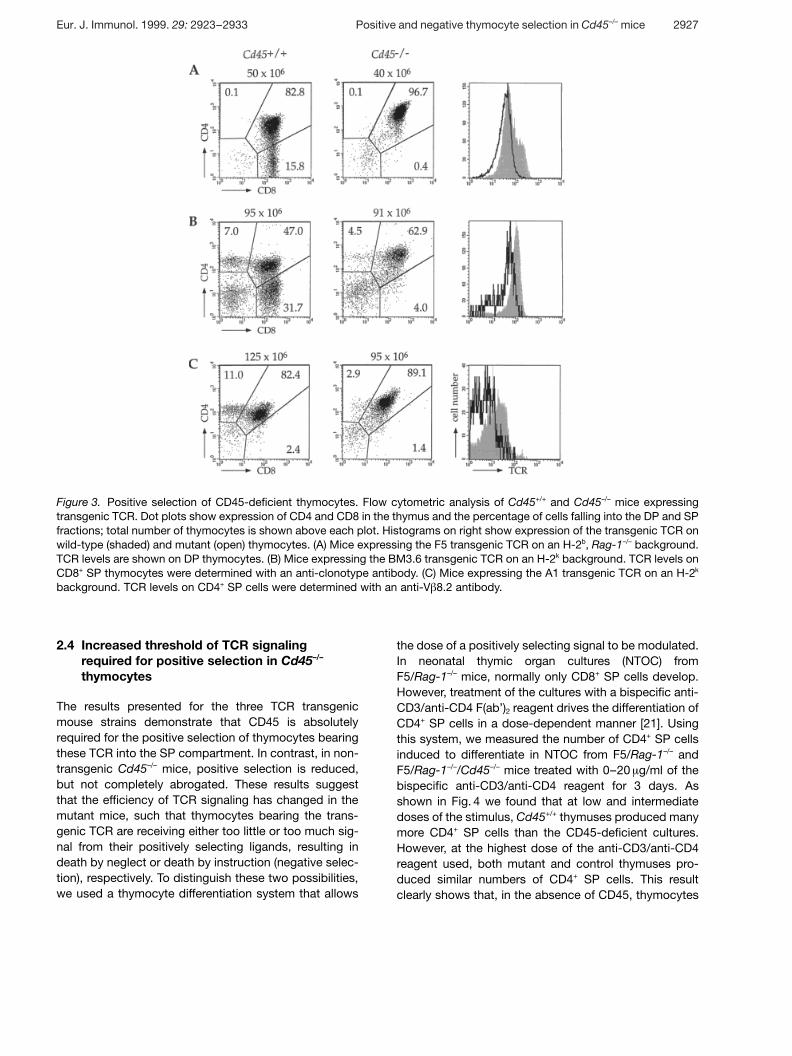
Figure 3. Positive selection of CD45-deficient thymocytes. Flow cytometric analysis of Cd45+/+ and Cd45–/– mice expressingtransgenic TCR. Dot plots show expression of CD4 and CD8 in the thymus and the percentage of cells falling into the DP and SPfractions; total number of thymocytes is shown above each plot. Histograms on right show expression of the transgenic TCR onwild-type (shaded) and mutant (open) thymocytes. (A) Mice expressing the F5 transgenic TCR on an H-2b, Rag-1–/– background.TCR levels are shown on DP thymocytes. (B) Mice expressing the BM3.6 transgenic TCR on an H-2k background. TCR levels onCD8+ SP thymocytes were determined with an anti-clonotype antibody. (C) Mice expressing the A1 transgenic TCR on an H-2k
background. TCR levels on CD4+ SP cells were determined with an anti-V g 8.2 antibody.
2.4 Increased threshold of TCR signalingrequired for positive selection in Cd45–/–
thymocytes
The results presented for the three TCR transgenicmouse strains demonstrate that CD45 is absolutelyrequired for the positive selection of thymocytes bearingthese TCR into the SP compartment. In contrast, in non-transgenic Cd45–/– mice, positive selection is reduced,but not completely abrogated. These results suggestthat the efficiency of TCR signaling has changed in themutant mice, such that thymocytes bearing the trans-genic TCR are receiving either too little or too much sig-nal from their positively selecting ligands, resulting indeath by neglect or death by instruction (negative selec-tion), respectively. To distinguish these two possibilities,we used a thymocyte differentiation system that allows
the dose of a positively selecting signal to be modulated.In neonatal thymic organ cultures (NTOC) fromF5/Rag-1–/– mice, normally only CD8+ SP cells develop.However, treatment of the cultures with a bispecific anti-CD3/anti-CD4 F(ab’)2 reagent drives the differentiation ofCD4+ SP cells in a dose-dependent manner [21]. Usingthis system, we measured the number of CD4+ SP cellsinduced to differentiate in NTOC from F5/Rag-1–/– andF5/Rag-1–/–/Cd45–/– mice treated with 0–20 ? g/ml of thebispecific anti-CD3/anti-CD4 reagent for 3 days. Asshown in Fig. 4 we found that at low and intermediatedoses of the stimulus, Cd45+/+ thymuses produced manymore CD4+ SP cells than the CD45-deficient cultures.However, at the highest dose of the anti-CD3/anti-CD4reagent used, both mutant and control thymuses pro-duced similar numbers of CD4+ SP cells. This resultclearly shows that, in the absence of CD45, thymocytes
Eur. J. Immunol. 1999. 29: 2923–2933 Positive and negative thymocyte selection in Cd45–/– mice 2927

Figure 4. Reduced efficiency of positive selection of CD45-deficient thymocytes. Graph showing mean (± SEM) num-bers of CD4+CD8– V g 11highPNAlowSP thymocytes generatedin NTOC from Cd45+/+ or Cd45–/– F5/Rag-1–/– mice, inresponse to different doses of a bispecific F(ab’)2 reagentrecognizing both CD3 4 and CD4 (anti-CD3/CD4).
transduce positively selecting signals much less effi-ciently, thus resulting in an increased threshold of TCR/CD4 signaling required to drive positive selection.
2.5 CD45-deficient thymocytes can undergonegative selection
To examine the requirement for CD45 in negative selec-tion of thymocytes we bred the Cd45 mutation onto abackground expressing the BM3.6 TCR and H-2b. SinceBM3.6 is an alloreactive anti-H-2Kb TCR, in Cd45+/+ micealmost all the DP thymocytes were deleted and thymiccellularity was greatly reduced (Fig. 5A). Since thesemice were Rag-1+/+, they contained a small number ofCD8+ SP cells which expressed TCR generated fromendogenous TCR genes; however, these were negativefor the BM3.6 clonotype (Fig. 5A). The thymuses ofCd45–/– mice on this background were similarly reducedin size, contained no DP cells, and the few remainingCD8+ SP cells were negative for the BM3.6 clonotype.Therefore, CD45 is not required for negative selection ofthis class I-restricted TCR.
2.6 Cd45–/– mice have an altered sensitivity tonegative selection
We also studied negative selection by endogenoussuperantigens. In BALB/c mice, proteins encoded bymouse mammary tumor proviruses (Mtv loci) are pre-sented in association with the class II molecule I-Ed andresult in the deletion of CD4+ and CD8+ SP cells express-
ing V g 3, 5 and 11 but not V g 8 [22]. Flow cytometric anal-ysis of Cd45–/– mice which had been backcrossed onto aBALB/c background demonstrated that, in comparisonto C57BL/6 mice which do not undergo Mtv-mediateddeletion, the Cd45 mutants had specifically deletedCD4+ SP thymocytes expressing V g 3,5 and 11 (Fig. 5B).However, comparison of Cd45–/– (BALB/c) with wild-typeBALB/c mice demonstrated that the extent of deletionwas not as great in the mutant mice. Thus, although neg-ative selection by Mtv superantigens can occur inCd45–/– mice, it appears to be less efficient.
To examine the altered sensitivity of CD45-deficient thy-mocytes to negative selection in more detail, we ana-lyzed their deletion by staphylococcal enterotoxin B(SEB), which binds to class II molecules and to V g 3, 7, 8,17 [23]. This binding leads to the selective elimination ofSP thymocytes expressing these particular V g . SEB-mediated negative selection is a “late” acting event anddoes not cause V g -specific deletion of DP thymocytes(not shown). CD45-deficient and control mice wereinjected with different amounts of SEB and were moni-tored for the selective disappearance of SP cellsexpressing V g 8. This allowed us to determine the doseresponse of SEB-driven negative selection in both geno-types. CD45-deficient mice were clearly much less sus-ceptible to deletion by SEB, since injections of 0.3–1 ? gSEB induced half-maximal deletion in control mice,whereas even 10 ? g caused little or no deletion of V g 8+
CD8+ SP thymocytes in Cd45–/– mice, although somedeletion of CD4+ SP cells was seen (Fig. 5C and data notshown). Taken together these results show that CD45 isnot absolutely required for negative selection, but in theabsence of CD45, the efficiency of the process is greatlyreduced and the threshold of TCR signaling required fornegative selection is increased.
2.7 Altered calcium flux in Cd45–/– thymocytes
One of the hallmarks of TCR-induced signaling is themobilization of intracellular calcium (Ca2+) and this canbe demonstrated in mature T cells as well as both SPand DP thymocytes. Furthermore, such calcium fluxeshave been implicated in both positive and negativeselection in the thymus [24, 25]. To examine TCR-induced calcium flux in Cd45–/– thymocytes, the cellswere preloaded with the calcium-sensitive dye Indo-1and flow cytometry was used to monitor intracellular cal-cium concentration following cross-linking of the TCR/CD3 complex. These studies clearly showed that cal-cium fluxes were greatly reduced in the CD45-deficientthymocytes (Fig. 6). In control DP and CD4+ SP thymo-cytes, calcium fluxes were readily detectable across arange of doses of cross-linking antibody (10–100 ? g/ml)
2928 P. J. Mee et al. Eur. J. Immunol. 1999. 29: 2923–2933
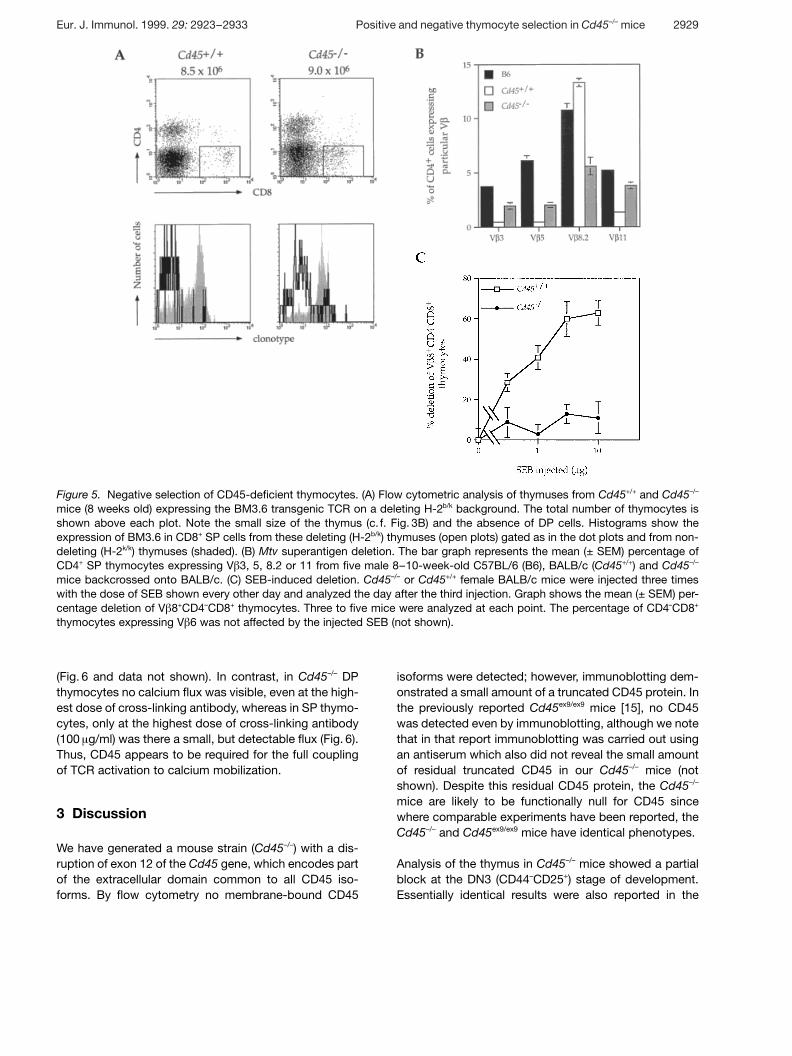
Figure 5. Negative selection of CD45-deficient thymocytes. (A) Flow cytometric analysis of thymuses from Cd45+/+ and Cd45–/–
mice (8 weeks old) expressing the BM3.6 transgenic TCR on a deleting H-2b/k background. The total number of thymocytes isshown above each plot. Note the small size of the thymus (c. f. Fig. 3B) and the absence of DP cells. Histograms show theexpression of BM3.6 in CD8+ SP cells from these deleting (H-2b/k) thymuses (open plots) gated as in the dot plots and from non-deleting (H-2k/k) thymuses (shaded). (B) Mtv superantigen deletion. The bar graph represents the mean (± SEM) percentage ofCD4+ SP thymocytes expressing V g 3, 5, 8.2 or 11 from five male 8–10-week-old C57BL/6 (B6), BALB/c (Cd45+/+) and Cd45–/–
mice backcrossed onto BALB/c. (C) SEB-induced deletion. Cd45–/– or Cd45+/+ female BALB/c mice were injected three timeswith the dose of SEB shown every other day and analyzed the day after the third injection. Graph shows the mean (± SEM) per-centage deletion of V g 8+CD4–CD8+ thymocytes. Three to five mice were analyzed at each point. The percentage of CD4–CD8+
thymocytes expressing V g 6 was not affected by the injected SEB (not shown).
(Fig. 6 and data not shown). In contrast, in Cd45–/– DPthymocytes no calcium flux was visible, even at the high-est dose of cross-linking antibody, whereas in SP thymo-cytes, only at the highest dose of cross-linking antibody(100 ? g/ml) was there a small, but detectable flux (Fig. 6).Thus, CD45 appears to be required for the full couplingof TCR activation to calcium mobilization.
3 Discussion
We have generated a mouse strain (Cd45–/–) with a dis-ruption of exon 12 of the Cd45 gene, which encodes partof the extracellular domain common to all CD45 iso-forms. By flow cytometry no membrane-bound CD45
isoforms were detected; however, immunoblotting dem-onstrated a small amount of a truncated CD45 protein. Inthe previously reported Cd45ex9/ex9 mice [15], no CD45was detected even by immunoblotting, although we notethat in that report immunoblotting was carried out usingan antiserum which also did not reveal the small amountof residual truncated CD45 in our Cd45–/– mice (notshown). Despite this residual CD45 protein, the Cd45–/–
mice are likely to be functionally null for CD45 sincewhere comparable experiments have been reported, theCd45–/– and Cd45ex9/ex9 mice have identical phenotypes.
Analysis of the thymus in Cd45–/– mice showed a partialblock at the DN3 (CD44–CD25+) stage of development.Essentially identical results were also reported in the
Eur. J. Immunol. 1999. 29: 2923–2933 Positive and negative thymocyte selection in Cd45–/– mice 2929
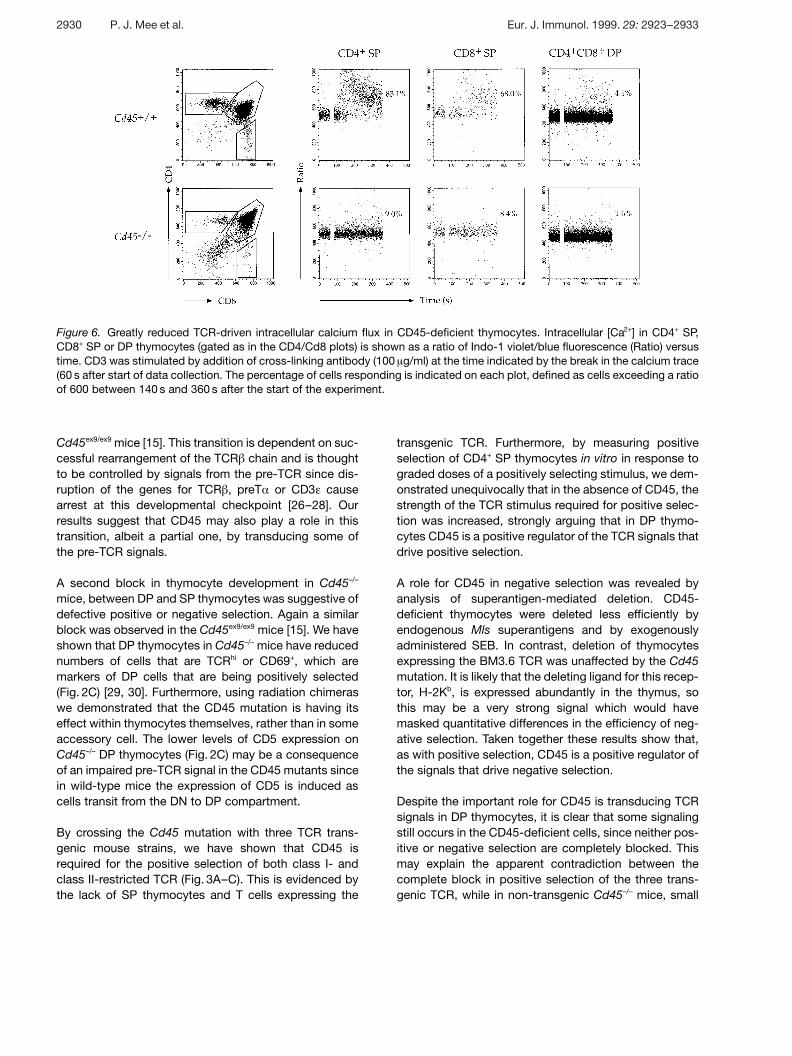
Figure 6. Greatly reduced TCR-driven intracellular calcium flux in CD45-deficient thymocytes. Intracellular [Ca2+] in CD4+ SP,CD8+ SP or DP thymocytes (gated as in the CD4/Cd8 plots) is shown as a ratio of Indo-1 violet/blue fluorescence (Ratio) versustime. CD3 was stimulated by addition of cross-linking antibody (100 ? g/ml) at the time indicated by the break in the calcium trace(60 s after start of data collection. The percentage of cells responding is indicated on each plot, defined as cells exceeding a ratioof 600 between 140 s and 360 s after the start of the experiment.
Cd45ex9/ex9 mice [15]. This transition is dependent on suc-cessful rearrangement of the TCR g chain and is thoughtto be controlled by signals from the pre-TCR since dis-ruption of the genes for TCR g , preT § or CD3 4 causearrest at this developmental checkpoint [26–28]. Ourresults suggest that CD45 may also play a role in thistransition, albeit a partial one, by transducing some ofthe pre-TCR signals.
A second block in thymocyte development in Cd45–/–
mice, between DP and SP thymocytes was suggestive ofdefective positive or negative selection. Again a similarblock was observed in the Cd45ex9/ex9 mice [15]. We haveshown that DP thymocytes in Cd45–/– mice have reducednumbers of cells that are TCRhi or CD69+, which aremarkers of DP cells that are being positively selected(Fig. 2C) [29, 30]. Furthermore, using radiation chimeraswe demonstrated that the CD45 mutation is having itseffect within thymocytes themselves, rather than in someaccessory cell. The lower levels of CD5 expression onCd45–/– DP thymocytes (Fig. 2C) may be a consequenceof an impaired pre-TCR signal in the CD45 mutants sincein wild-type mice the expression of CD5 is induced ascells transit from the DN to DP compartment.
By crossing the Cd45 mutation with three TCR trans-genic mouse strains, we have shown that CD45 isrequired for the positive selection of both class I- andclass II-restricted TCR (Fig. 3A–C). This is evidenced bythe lack of SP thymocytes and T cells expressing the
transgenic TCR. Furthermore, by measuring positiveselection of CD4+ SP thymocytes in vitro in response tograded doses of a positively selecting stimulus, we dem-onstrated unequivocally that in the absence of CD45, thestrength of the TCR stimulus required for positive selec-tion was increased, strongly arguing that in DP thymo-cytes CD45 is a positive regulator of the TCR signals thatdrive positive selection.
A role for CD45 in negative selection was revealed byanalysis of superantigen-mediated deletion. CD45-deficient thymocytes were deleted less efficiently byendogenous Mls superantigens and by exogenouslyadministered SEB. In contrast, deletion of thymocytesexpressing the BM3.6 TCR was unaffected by the Cd45mutation. It is likely that the deleting ligand for this recep-tor, H-2Kb, is expressed abundantly in the thymus, sothis may be a very strong signal which would havemasked quantitative differences in the efficiency of neg-ative selection. Taken together these results show that,as with positive selection, CD45 is a positive regulator ofthe signals that drive negative selection.
Despite the important role for CD45 is transducing TCRsignals in DP thymocytes, it is clear that some signalingstill occurs in the CD45-deficient cells, since neither pos-itive or negative selection are completely blocked. Thismay explain the apparent contradiction between thecomplete block in positive selection of the three trans-genic TCR, while in non-transgenic Cd45–/– mice, small
2930 P. J. Mee et al. Eur. J. Immunol. 1999. 29: 2923–2933

numbers of SP thymocytes and peripheral T cells aregenerated. The transgenic receptors are blocked in theirdevelopment as they do not receive a large enough sig-nal from binding to their positively selecting ligand,whereas in non-transgenic mutant mice those thymo-cytes which are selected may be expressing TCR with ahigher affinity/avidity for positively selecting MHCligands, resulting in a stimulus strong enough to over-come the Cd45 mutation.
In conclusion, we have shown that the absence of CD45results in a large increase in the thresholds of TCR stimu-lation required for both positive and negative selection.Thus, in DP thymocytes, CD45 is a critical positive regu-lator of the TCR signals that drive these selection events.
4 Materials and methods
4.1 Targeting of the Cd45 gene
Q phage containing about 32 kb of the Cd45 gene were iso-lated from a genomic library of 129/Sv mouse DNA in Q FIXII(Stratagene) using an 806-bp EcoRV/HindIII fragment of themouse Cd45 cDNA spanning exons 7 to 13 of the extracellu-lar domain, taken from p70ZLC (gift of M. Thomas). The tar-geting construct pCD45-T was built by subcloning a 6.7-kbBamHI fragment extending from intron 8 to exon 12 and a3.1-kb BamHI fragment extending from exon 12 to intron 12into the XhoI and EcoRI sites, respectively, of pPNT [31].pCD45-T DNA linearized with NotI was electroporated intoD3 ES cells and G418- and gancyclovir-resistant clonesselected using 250 ? g/ml G418 and 2 ? M gancyclovir as pre-viously described [31]. Lines carrying the correct targetingevent were identified by Southern blot analysis as describedin Fig. 1A. We identified 11 targeted clones out of 480 clonesscreened. The correct structure of the targeted Cd45 genewas confirmed by further Southern blots (data not shown).
4.2 Mice
Chimeric mice were generated by injecting targeted ES cellsinto blastocysts from C57BL/6 mice using standard proce-dures. The Cd45 mutation was established in the germ lineby breeding male chimeras to (B6D2)F1 mice. To generatemice inbred on 129/Sv, chimeras were bred directly to 129/Sv female mice; for mice on the BALB/c background,Cd45+/– mice were backcrossed to wild-type BALB/c micefor 10 generations and then intercrossed. TCR transgenicmice have been described previously: F5 [18], BM3.6 [19],A1(m-2) [20]. Rag-1–/– mice were kindly provided by D. Balti-more [32].
4.3 Staining of cells with fluorescent antibodies andcalcium analysis
Staining with the indicated combinations of monoclonalantibodies was performed as previously described [33]. Therise in intracellular calcium in thymocytes stimulated throughcross-linking of CD3 was monitored using Indo-1 acetoxy-methyl ester exactly as previously described [33].
4.4 Dose-response studies of positive and negativeselection
Neonatal (day of birth) thymus lobes were obtained frommatings of Cd45+/+ or Cd45–/– mice that were also Rag-1–/–
and carried the F5 TCR transgene. Mature CD4+ SP thymo-cytes were generated in NTOC in the presence of a bispe-cific F(ab’)2 reagent recognizing both CD3 4 and CD4 as pre-viously described [21]. Mature (V g 11hi/PNAlo), CD4+CD8–
cells that differentiated were calculated from three individuallobes for each concentration of anti-CD3/CD4. Thymocytedeletion induced by SEB was analyzed exactly as previouslydescribed, except that CD8+ rather than CD4+ SP cells wereevaluated [33].
Acknowledgements: We thank C. Atkins for flow cyto-metric analysis; M. Thomas for a CD45 cDNA clone; D.Kioussis and A. Mellor for provision of TCR transgenic mice,advice and discussions; E. Schweighoffer, L. Duddy, M.Quinn and A. Walters for technical assistance; E. Schweig-hoffer for critical reading of the manuscript; G. Klaus, P.Johnson and M. Thomas for antibodies. The work was sup-ported by a grant from the Leukemia Research Fund (toV.L.J.T.) and by the Medical Research Council, U.K.
5 References
1 Jameson, S. C., Hogquist, K. A. and Bevan, M. J.,Positive selection of thymocytes. Annu. Rev. Immunol.1995. 13: 93–126.
2 Pingel, J. T. and Thomas, M. L., Evidence that the leu-kocyte common antigen is required for antigen-inducedT lymphocyte proliferation. Cell 1989. 58: 1055–1065.
3 Koretzky, G. A., Picus, J., Thomas, M. L. and Weiss,A., Tyrosine phosphatase CD45 is essential for couplingT-cell antigen receptor to the phosphatidyl inositol path-way. Nature 1990. 346: 66–68.
4 Shiroo, M. L. G., Biffen, E., Shivnan, E. and Alexander,D., CD45-tyrosine phosphatase-activated p59fyn cou-ples the T cell antigen receptor to pathways of diacylgly-cerol production, protein kinase C activation and cal-cium influx. EMBO J. 1992. 11: 4887.
Eur. J. Immunol. 1999. 29: 2923–2933 Positive and negative thymocyte selection in Cd45–/– mice 2931

5 Ostergaard, H. L., Shackelford, D. A., Hurley, T. R.,Johnson, P., Hyman, R., Sefton, B. M. and Trow-bridge, I. S., Expression of CD45 alters phosphorylationof the lack-encoded tyrosine protein kinase in murinelymphoma T-cell lines. Proc. Natl. Acad. Sci. USA 1989.86: 8959–8963.
6 Sieh, M., Bolen, J. B. and Weiss, A., CD45 specificallymodulates binding of Lck to a phosphopeptide encom-passing the negative regulatory tyrosine of Lck. EMBO J.1993. 12: 315–321.
7 McFarland, E. D. C., Hurley, T. R., Pingel, J. T., Sefton,B. M., Shaw, A. and Thomas, M. L., Correlationbetween Src family member regulation by the protein-tyrosine-phosphatase CD45 and transmembrane signal-ing through the T-cell receptor. Proc. Natl. Acad. Sci.USA 1993. 90: 1402–1406.
8 Hurley, T. R., Hyman, R. and Sefton, B. M., Differentialeffects of expression of the CD45 tyrosine protein phos-phatase on the tyrosine phosphorylation of the lck, fyn,and c-src tyrosine protein kinases. Mol. Cell. Biol. 1993.13: 1651–1656.
9 Deans, J. P., Kanner, S. B., Torres, R. M. and Ledbet-ter, J. A., Interaction of CD4:lck with the T cell receptor/CD3 complex induces early signaling events in theabsence of CD45 tyrosine phosphatase. Eur. J. Immu-nol. 1992. 22: 661–668.
10 Burns, C. M., Sakaguchi, K., Appella, E. and Ashwell,J. D., CD45 regulation of tyrosine phosphorylation andenzyme-activity of Src-family kinases. J. Biol. Chem.1994. 269: 13594.
11 D’Oro, U., Sakaguchi, K., Appella, E. and Ashwell, J.D., Mutational analysis of Lck in CD45-negative T cells:dominant role of tyrosine 394 phosphorylation in kinaseactivity. Mol. Cell. Biol. 1996. 16: 4996–5003.
12 Gervais, F. G. and Veillette, A., Reconstitution of inter-actions between protein-tyrosine phosphatase CD45and tyrosine-protein kinase p56(lck) in nonlymphoidcells. J. Biol. Chem. 1997. 272: 12754–12761.
13 Kishihara, K., Penninger, J., Wallace, V. A., Kundig, T.M., Kawai, K., Wakeham, A., Timms, E., Pfeffer, K.,Ohashi, P. S., Thomas, M. L., Furlonger, C., Paige, C.J. and Mak, T. W., Normal B lymphocyte developmentbut impaired T cell maturation in CD45-exon 6 proteintyrosine phosphatase-deficient mice. Cell 1993. 74:143–156.
14 Kong, Y. Y., Kishihara, K., Yoshida, H., Mak, T. W. andNomoto, K., Generation of T cells with differentialresponses to alloantigens in CD45 exon 6-deficientmice. J. Immunol. 1995. 154: 5725–5735.
15 Byth, K. F., Conroy, L. A., Howlett, S., Smith, A. J. H.,May, J., Alexander, D. R. and Holmes, N., CD45-nulltransgenic mice reveal a positive regulatory role for
CD45 in early thymocyte development, in the selectionof CD4+CD8+ lymphocytes, and in B cell maturation. J.Exp. Med. 1996. 183: 1707–1718.
16 Godfrey, D. I., Kennedy, J., Suda, T. and Zlotnik, A., Adevelopmental pathway involving four phenotypicallyand functionally distinct subsets of CD3-CD4-CD8-triple-negative adult mouse thymocytes defined byCD44 and CD25 expression. J. Immunol. 1993. 150:4244–4252.
17 Haks, M. C., Oosterwegel, M. A., Blom, B., Spits, H.M. and Kruisbeek, A. M., Cell-fate decisions in early Tcell development regulation by cytokine receptors andthe pre-TCR. Semin Immunol. 1999. 11: 23–37.
18 Mamalaki, C., Elliot, J., Norton, T., Yannoutsos, N.,Townsend, A. R., Chandler, P., Simpson, E. andKioussis, D., Positive and negative selection in trans-genic mice expressing a T-cell receptor specific for influ-enza nucleoprotein and endogenous superantigen. Dev.Immunol. 1993. 3: 159–174.
19 Sponaas, A. M., Tomlinson, P. D., Antoniou, J.,Auphan, N., Langlet, C., Malissen, B., Schmitt Ver-hulst, A. M. and Mellor, A. L., Induction of tolerance toself MHC class I molecules expressed under the controlof milk protein or beta-globin gene promoters. Int.Immunol. 1994. 6: 277–287.
20 Douek, D. C., Corley, K. T. T., Zal, T., Mellor, A., Dyson,P. J. and Altmann, D. M., Negative selection by endoge-nous antigen and superantigen occurs at multiple thymicsites. Int. Immunol. 1996. 8: 1413–1420.
21 Basson, M. A., Bommhardt, U., Cole, M. S., Tso, J. Y.and Zamoyska, R., CD3 ligation on immature thymo-cytes generates antagonist-like signals appropriate forCD8 lineage commitment, independently of T cell recep-tor specificity. J. Exp. Med. 1998. 187: 1249–1260.
22 Simpson, E., Dyson, P. J., Knight, A. M., Robinson, P.J., Elliott, J. I. and Altmann, D. M., T-cell receptor rep-ertoire selection by mouse mammary tumor viruses andMHC molecules. Immunol. Rev. 1993. 131: 93–115.
23 Marrack, P. and Kappler, J., The staphylococcalenterotoxins and their relatives. Science 1990. 248:705–711.
24 Vasquez, N. J., Kane, L. P. and Hedrick, S. M., Intracel-lular signals that mediate thymic negative selection.Immunity 1994. 1: 45–56.
25 Takahama, Y. and Nakauchi, H., Phorbol ester and cal-cium ionophore can replace TCR signals that inducepositive selection of CD4 T cells. J. Immunol. 1996. 157:1508–1513.
26 Mombaerts, P., Clarke, A. R., Rudnicki, M. A., Iaco-mini, J., Itohara, S., Lafaille, J. J., Wang, L., Ichikawa,Y., Jaenisch, R., Hooper, M. L. and Tonegawa, S.,Mutations in T-cell antigen receptor genes alpha and
2932 P. J. Mee et al. Eur. J. Immunol. 1999. 29: 2923–2933

beta block thymocyte development at different stages.Nature 1992. 360: 225–231.
27 Fehling, H. J., Krotkova, A., Saint Ruf, C. and vonBoehmer, H., Crucial role of the pre-T-cell receptoralpha gene in development of alpha beta but not gammadelta T cells. Nature 1995. 375: 795–798.
28 Malissen, M., Gillet, A., Ardouin, L., Bouvier, G., Trucy,J., Ferrier, P., Vivier, E. and Malissen, B., Altered T celldevelopment in mice with a targeted mutation of theCD3-epsilon gene. EMBO J. 1995. 14: 4641–4653.
29 Swat, W., Dessing, M., von Boehmer, H. and Kisielow,P., CD69 expression during selection and maturation ofCD4+8+ thymocytes. Eur. J. Immunol. 1993. 23:739–746.
30 Yamashita, I., Nagata, T., Tada, T. and Nakayama, T.,CD69 cell surface expression identifies developing thy-mocytes which audition for T cell antigen receptor-mediated positive selection. Int. Immunol. 1993. 5:1139–1150.
31 Tybulewicz, V. L., Crawford, C. E., Jackson, P. K.,Bronson, R. T. and Mulligan, R. C., Neonatal lethalityand lymphopenia in mice with a homozygous disruptionof the c-abl proto-oncogene. Cell 1991. 65: 1153–1163.
32 Spanopoulou, E., Roman, C. A., Corcoran, L. M.,Schlissel, M. S., Silver, D. P., Nemazee, D., Nussen-zweig, M. C., Shinton, S. A., Hardy, R. R. and Balti-more, D., Functional immunoglobulin transgenes guideordered B-cell differentiation in Rag-1-deficient mice.Genes Dev. 1994. 8: 1030–1042.
33 Turner, M., Mee, P. J., Walters, A., Quinn, M. E., Mel-lor, A. L., Zamoyska, R. and Tybulewicz, V. L. J., Arequirement for the Rho-family GTP exchange factor Vavin positive and negative selection of thymocytes. Immu-nity 1997. 7: 451–460.
Correspondence: Victor L. J. Tybulewicz, National Institutefor Medical Research, The Ridgeway, Mill Hill, London, NW71AA, UKFax: +44-181-906-4477e-mail: v-tybule — nimr.mrc.ac.uk
Present addresses: P. Joseph Mee, Centre for GenomeResearch, West Main Road, Edingburgh EH9 3JQ, U.K.;Martin Turner, The Babraham Institute, Babraham, Cam-bridge CB2 4AT, U.K.; and Patrick S. Costello, Imperial Can-cer Research Fund, Lincoln’s Inn Fields, London WC2A 3PX,U.K.
Eur. J. Immunol. 1999. 29: 2923–2933 Positive and negative thymocyte selection in Cd45–/– mice 2933
Copyright © 2022 FDOKUMEN




















