
Protein tyrosine phosphatase PTPRJ is negatively regulated by microRNA-328
12
Protein tyrosine phosphatase PTPRJ is negatively regulated by microRNA-328 Francesco Paduano 1 , Vincenzo Dattilo 1 , Domenico Narciso 1 , Anna Bilotta 1 , Eugenio Gaudio 1 , Miranda Menniti 2 , Valter Agosti 1 , Camillo Palmieri 1 , Nicola Perrotti 2 , Alfredo Fusco 3 , Francesco Trapasso 1 and Rodolfo Iuliano 1 1 Dipartimento di Medicina Sperimentale e Clinica, Universita ` ‘Magna Graecia’, Catanzaro, Italy 2 Dipartimento di Scienze della Salute, Universita ` ‘Magna Graecia’, Catanzaro, Italy 3 Dipartimento di Biologia e Patologia Cellulare e Molecolare, Universita ` ‘Federico II’, Napoli, Italy Keywords microRNA; miR-328; protein tyrosine phosphatase; PTPRJ; tumour suppressor gene Correspondence R. Iuliano, Dipartimento di Medicina Sperimentale e Clinica, Universita ` ‘Magna Graecia’, viale Europa, 88100 Catanzaro, Italy Fax: +39 0961 3694090 Tel: +39 0961 3695182 E-mail: [email protected] Website: http://www.unicz.it F. Paduano, Dipartimento di Medicina Sperimentale e Clinica, Universita ` ‘Magna Graecia’, viale Europa, 88100 Catanzaro, Italy Fax: +39 0961 3694090 Tel: +39 0961 3694106 E-mail: [email protected] Website: http://www.unicz.it (Received 22 December 2011, revised 31 March 2012, accepted 3 May 2012) doi:10.1111/j.1742-4658.2012.08624.x Expression of PTPRJ, which is a ubiquitous receptor-type protein tyrosine phosphatase, is significantly reduced in a vast majority of human epithelial cancers and cancer cell lines (i.e. colon, lung, thyroid, mammary and pan- creatic tumours). A possible role for microRNAs (miRNAs) in the negative regulation of PTPRJ expression has never been investigated. In this study, we show that overexpression of microRNA-328 (miR-328) decreases PTPRJ expression in HeLa and SKBr3 cells. Further investigations demon- strate that miR-328 acts directly on the 3¢UTR of PTPRJ, resulting in reduced mRNA levels. Luciferase assay and site-specific mutagenesis were used to identify a functional miRNA response element in the 3¢UTR of PTPRJ. Expression of miR-328 significantly enhances cell proliferation in HeLa and SKBr3 cells, similar to the effects of downregulation of PTPRJ with small interfering RNA. Additionally, in HeLa cells, the proliferative effect of miR-328 was not observed when PTPRJ was silenced with small interfering RNA; conversely, restoration of PTPRJ expression in miR-328- overexpressing cells abolished the proliferative activity of miR-328. In con- clusion, we report the identification of miR-328 as an important player in the regulation of PTPRJ expression, and we propose that the interaction of miR-328 with PTPRJ is responsible for miR-328-dependent increase of epi- thelial cell proliferation. Introduction The tyrosine phosphatase PTPRJ (also called DEP-1, HPTPg or CD148) is a widely expressed transmem- brane protein that plays a role in multiple signalling pathways [1]. Substrates of PTPRJ include proteins that belong to the family of tyrosine kinase receptors (EGFR, HGFR, PDGFR, RET and VEGFR-2) [2–6] as well as those involved in cell adhesion (c-Src, p120- catenin and ZO-1) [7–9]. Consequently, in several epi- thelium-derived cell lines, PTPRJ inhibits cell prolifera- tion and promotes cell adhesion [10–13]. PTPRJ has been proposed to be a tumour suppressor owing to downregulation of its expression in cancer cell lines [10,12–14] and inhibition of tumorous growth upon restoration of normal expression levels [12–14]. Abbreviations CFSE, carboxyfluorescein succinimidyl ester; hsa-miR, Homo sapiens microRNA; LOH, loss of heterozygosity; miR-328, microRNA-328; miRNA, microRNA; MRE, microRNA response element; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; siRNA, small interfering RNA. FEBS Journal (2012) ª 2012 The Authors Journal compilation ª 2012 FEBS 1
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Protein tyrosine phosphatase PTPRJ is negatively regulated by microRNA-328

Protein tyrosine phosphatase PTPRJ is negativelyregulated by microRNA-328Francesco Paduano1, Vincenzo Dattilo1, Domenico Narciso1, Anna Bilotta1, Eugenio Gaudio1,Miranda Menniti2, Valter Agosti1, Camillo Palmieri1, Nicola Perrotti2, Alfredo Fusco3, FrancescoTrapasso1 and Rodolfo Iuliano1
1 Dipartimento di Medicina Sperimentale e Clinica, Universita ‘Magna Graecia’, Catanzaro, Italy
2 Dipartimento di Scienze della Salute, Universita ‘Magna Graecia’, Catanzaro, Italy
3 Dipartimento di Biologia e Patologia Cellulare e Molecolare, Universita ‘Federico II’, Napoli, Italy
Keywords
microRNA; miR-328; protein tyrosine
phosphatase; PTPRJ; tumour suppressor
gene
Correspondence
R. Iuliano, Dipartimento di Medicina
Sperimentale e Clinica, Universita ‘Magna
Graecia’, viale Europa, 88100 Catanzaro,
Italy
Fax: +39 0961 3694090
Tel: +39 0961 3695182
E-mail: [email protected]
Website: http://www.unicz.it
F. Paduano, Dipartimento di Medicina
Sperimentale e Clinica, Universita ‘Magna
Graecia’, viale Europa, 88100 Catanzaro,
Italy
Fax: +39 0961 3694090
Tel: +39 0961 3694106
E-mail: [email protected]
Website: http://www.unicz.it
(Received 22 December 2011, revised 31
March 2012, accepted 3 May 2012)
doi:10.1111/j.1742-4658.2012.08624.x
Expression of PTPRJ, which is a ubiquitous receptor-type protein tyrosine
phosphatase, is significantly reduced in a vast majority of human epithelial
cancers and cancer cell lines (i.e. colon, lung, thyroid, mammary and pan-
creatic tumours). A possible role for microRNAs (miRNAs) in the negative
regulation of PTPRJ expression has never been investigated. In this study,
we show that overexpression of microRNA-328 (miR-328) decreases
PTPRJ expression in HeLa and SKBr3 cells. Further investigations demon-
strate that miR-328 acts directly on the 3¢UTR of PTPRJ, resulting in
reduced mRNA levels. Luciferase assay and site-specific mutagenesis were
used to identify a functional miRNA response element in the 3¢UTR of
PTPRJ. Expression of miR-328 significantly enhances cell proliferation in
HeLa and SKBr3 cells, similar to the effects of downregulation of PTPRJ
with small interfering RNA. Additionally, in HeLa cells, the proliferative
effect of miR-328 was not observed when PTPRJ was silenced with small
interfering RNA; conversely, restoration of PTPRJ expression in miR-328-
overexpressing cells abolished the proliferative activity of miR-328. In con-
clusion, we report the identification of miR-328 as an important player in
the regulation of PTPRJ expression, and we propose that the interaction of
miR-328 with PTPRJ is responsible for miR-328-dependent increase of epi-
thelial cell proliferation.
Introduction
The tyrosine phosphatase PTPRJ (also called DEP-1,
HPTPg or CD148) is a widely expressed transmem-
brane protein that plays a role in multiple signalling
pathways [1]. Substrates of PTPRJ include proteins
that belong to the family of tyrosine kinase receptors
(EGFR, HGFR, PDGFR, RET and VEGFR-2) [2–6]
as well as those involved in cell adhesion (c-Src, p120-
catenin and ZO-1) [7–9]. Consequently, in several epi-
thelium-derived cell lines, PTPRJ inhibits cell prolifera-
tion and promotes cell adhesion [10–13]. PTPRJ has
been proposed to be a tumour suppressor owing to
downregulation of its expression in cancer cell lines
[10,12–14] and inhibition of tumorous growth upon
restoration of normal expression levels [12–14].
Abbreviations
CFSE, carboxyfluorescein succinimidyl ester; hsa-miR, Homo sapiens microRNA; LOH, loss of heterozygosity; miR-328, microRNA-328;
miRNA, microRNA; MRE, microRNA response element; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; siRNA, small
interfering RNA.
FEBS Journal (2012) ª 2012 The Authors Journal compilation ª 2012 FEBS 1

Although expression of PTPRJ decreases significantly
during the progression of human tumours [15,16], little
is known about the mechanisms involved in the pro-
cess. Loss of heterozygosity (LOH) was detected at the
PTPRJ locus (11p11.2) in lung, breast, colon and thy-
roid carcinomas [17,18]; however, LOH and promoter
methylation of PTPRJ [19,20] can only partly explain
the association between tumour progression and
reduced PTPRJ expression.
MicroRNAs (miRNAs) are tiny non-coding RNAs
that downregulate gene expression by binding to the
3¢UTRs of their targets resulting in the inhibition of
protein translation or mRNA degradation [21]. Given
their varied effects on gene regulation, miRNAs have
been implicated in several diseases, including cancer.
MiRNA signatures have been used in the classification
of human tumours and in cancer prognosis predictions
[22–24]. MiRNA expression levels are not only useful as
biomarkers of disease progression, but it is now clear
that they also play a causative role in cancer progres-
sion. Genetic and epigenetic variations of miRNAs have
been detected in tumours [25,26], and polymorphisms in
miRNA coding regions have been associated with can-
cer susceptibility [27]. Probing their mechanistic role in
cancer, it appears that miRNAs can target either onco-
genes or tumour suppressor genes, and act by blocking
or favouring tumour progression as a consequence. The
balance in targets between oncogenes and tumour sup-
pressors by a certain miRNA defines its role in cancer
development [24]. Therefore, it is crucial to identify can-
cer-related genes targeted by miRNAs.
In this study, we report that microRNA-328 (miR-328)
targets the 3¢UTR of PTPRJ resulting in its decreased
expression. Furthermore, PTPRJ downregulation could
Table 1. Computational analysis with MIRTAR, FINDTAR and RNAHYBRID of microRNA recognition elements (MREs) for miR-328 in the 3¢UTR of
PTPRJ mRNA. MREs identified by RNAHYBRID are indicated as MRE_1 to MRE_5 in the text.
PTPRJ expression is negatively regulated by microRNA-328 F. Paduano et al.
2 FEBS Journal (2012) ª 2012 The Authors Journal compilation ª 2012 FEBS

explain the proliferative effects observed in HeLa cells
withmiR-328 overexpression. Therefore, we propose that
miR-328 is a negative regulator of PTPRJ expression,
with a possible role in the development of human
tumours.
Results
The 3¢UTR of PTPRJ was analysed by the bioinfor-
matics algorithm mirtar [28] to identify possible inter-
acting miRNAs. Several miRNAs were selected as
likely interactors and were considered for further bio-
informatic analyses with other computational algo-
rithms (findtar and rnahybrid) [29,30]. Among the
selected miRNAs, for miR-328, two miRNA response
elements (MREs) were found with mirtar; four with
findtar; and five with rnahybrid in the 3¢UTR of
PTPRJ (Table 1). For further analyses on MREs, we
considered the sites picked with rnahybrid.
In order to evaluate the reliability of computer pre-
diction, we performed the transfection of synthetic
miR-328 in two different cell lines (HeLa and SKBr3),
and we then detected PTPRJ by western blot. We
found a significant decrease of PTPRJ protein levels in
miR-328 transfected cells compared with the control
(Fig. 1A). As a positive control, a PTPRJ-specific
small interfering RNA (siRNA) was used. To evaluate
whether miR-328 affects PTPRJ expression by decreas-
ing mRNA levels, we quantified, by qRT-PCR,
mRNA levels of PTPRJ and its alternatively spliced
shorter isoform, encoding an mRNA with a different
3¢UTR (Fig. 1B). We found that miR-328 decreased
mRNA levels of PTPRJ but not of the shorter iso-
form, indicating that miR-328 probably acts on the
3¢UTR of PTPRJ mRNA. In addition, the PTPRJ
siRNA oligonucleotide that binds a region on exon 6
shared by the two PTPRJ isoforms decreased mRNA
levels of both isoforms (Fig. 1C). Thus, we demon-
strated that miR-328 downregulates endogenous
PTPRJ at both the mRNA and the protein levels.
We hypothesized that miR-328 interacts directly with
the 3¢UTR of PTPRJ mRNA to suppress PTPRJ
expression. To test this hypothesis, the ability of miR-
328 to regulate the 3¢UTR of PTPRJ was evaluated by
luciferase reporter assays. The region starting from
nucleotide +6259 to nucleotide +7761 of the PTPRJ
sequence (NM_002843.3) was cloned downstream of a
reporter luciferase gene (pmirGLO 3¢-WT) (Fig. 2A), as
described in Materials and methods. Co-transfection of
synthetic miR-328 and pmirGLO 3¢-WT reduced the
luciferase activity significantly in comparison with Lipo-
fectamine and miR-342 (Fig. 2B), an miRNA that did
not influence endogenous PTPRJ expression in HeLa
cells (data not shown). However, co-transfection of
miR-328 in cells transfected with pmirGLO empty vec-
tor did not influence luciferase activity significantly.
These results prompted us to study recognition elements
of miR-328 in the PTPRJ 3¢UTR. Five possible MREs
(MRE_1 to MRE_5, Fig. 2A) containing an miR-328
seeding region were predicted by computational
*
*
*
Rel
- m
RN
A le
vel
1.2
0.8
0.6
0.4
0.2
0
1.4
1
1.2
0.8
0.6
0.4
0.2
0
1.4
1
* *
*
HeLa SKBr3
PTPRJ long form
PTPRJ short form
SKBr3 HeLa
PTPRJ
γ-tubulin
220 kDa
50 kDa
PTPRJ transcript long form
PTPRJ transcript short form miR-328
siRNA-PTPRJ
3 UTR exon 6
A
C
BFig. 1. PTPRJ expression is downregulated
by miR-328. (A) Western blot analysis for
PTPRJ detection in protein extracts from
cells transfected transiently for 24 h with
siRNA-PTPRJ, Lipofectamine or miR-328.
Expression is normalized on c-tubulin. (B)
Schematic representation of the PTPRJ
pre-mRNA (long and short forms) and loca-
tion of miRNA and siRNA target sites on
PTPRJ mRNA (Genbank accession no.
NM_002843.3). (C) qRT-PCR with specific
primers amplifying the two isoforms of the
PTPRJ gene from cell lines transfected
transiently for 24 h with Lipofectamine,
siRNA-PTPRJ or miR-328. Values were
normalized to HPRT RNA levels. *P < 0.01.
F. Paduano et al. PTPRJ expression is negatively regulated by microRNA-328
FEBS Journal (2012) ª 2012 The Authors Journal compilation ª 2012 FEBS 3

analyses, as reported before (Table 1). To map interac-
tion sites of miR-328 in the 3¢UTR of PTPRJ, we gener-
ated a deletion mutant that includes the 3¢UTR
(pmirGLO 3¢D) downstream of the SacI site and con-
tains just two possible miR-328 binding sites (MRE_4
and MRE_5). Luciferase activity of that mutant was sig-
nificantly affected by the co-transfection with miR-328,
and the amount of luciferase reduction was not lower
than that detected by the pmirGLO 3¢UTR. Therefore,
deletion of nucleotides +6259 to +7082 of the human
PTPRJ 3¢UTR containing three MREs (MRE_1 to
MRE_3) did not decrease susceptibility to miR-328
inhibition (Fig. 2C), indicating that MRE_1 to MRE_3
were not functional. To better characterize the interac-
tion, we generated two site-specific mutations in
MRE_4 and MRE_5 (3¢D-4 mut and 3¢D-5 mut)
affecting the possible remaining miR-328 binding sites
in the 3¢UTR of PTPRJ mRNA (Fig. 2D). When the
MRE_5 site is mutated (pmirGLO 3¢D-5 mut), only a
slight decrease in the luciferase activity was observed.
Mutation of the MRE_4 site (pmirGLO 3¢D-4 mut) sig-
nificantly abolished the repression of luciferase activity
operated by miR-328. The same results were obtained
with the double mutant 3¢UTR (pmirGLO 3¢D-4 ⁄ 5 mut),
indicating that MRE_4 in position from +7174 to
+7203 plays a key role in the miR-328 mediated repres-
sion (Fig. 2E). Taken together, these data indicate that
miR-328 directly regulates PTPRJ expression mainly
through interaction with the predicted MRE_4 in the
3¢UTR of PTPRJ.
Proliferation of HeLa and SKBr3 cells is sensitive to
PTPRJ levels modulated by vector transfection [10,31].
Fig. 2. PTPRJ is a direct target of miR-328. (A) Reporter plasmid containing five potential miR-328 binding sites within the 3¢UTR of human
PTPRJ gene (MRE_1 to MRE_5). (B) Luciferase assay of HeLa cells co-transfected with PTPRJ WT 3¢UTR reporter plasmid (pmirGLO 3¢-WT)
or empty vector (pmirGLO) and synthetic miR-328 or miR-342. (C) Luciferase assay of HeLa cells co-transfected with 3¢UTR reporter plasmid
(pmirGLO 3¢-WT) or deleted 3¢UTR (pmirGLO 3¢D) and synthetic miR-328. (D) Base pairing for comparison between mature miR-328 and
wild-type (PTPRJ WT MRE) or mutant (PTPRJ mut MRE) putative target sites (MRE_4 and MRE_5) in the 3¢UTR of PTPRJ mRNA. Mutations
were generated on the potential target sequence of PTPRJ (bold letters with underlining) by site-directed mutagenesis. (E) Reporter repres-
sion of miR-328 in transfected HeLa cells with vector is indicated. Luciferase values were normalized to the double mutant (pmirGLO 3¢D-4 ⁄ 5 mut). Significant differences are indicated by an asterisk, P < 0.01. All the experiments were performed in quadruplicate and repeated
twice.
PTPRJ expression is negatively regulated by microRNA-328 F. Paduano et al.
4 FEBS Journal (2012) ª 2012 The Authors Journal compilation ª 2012 FEBS
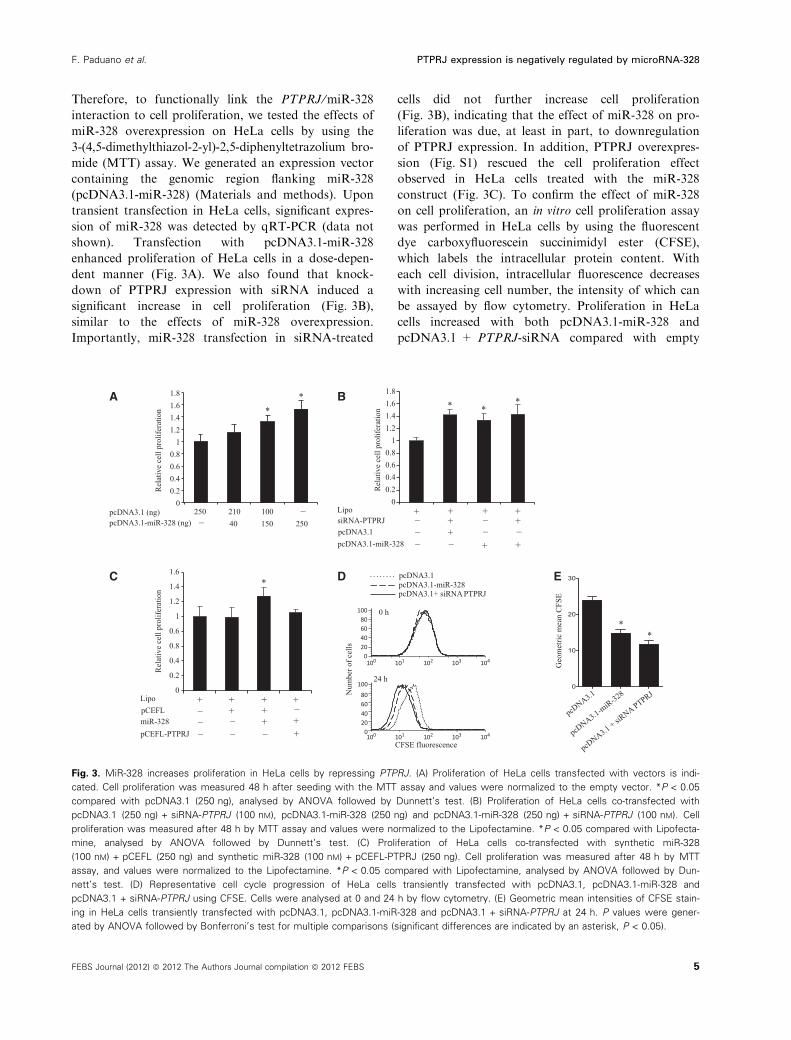
Therefore, to functionally link the PTPRJ ⁄miR-328
interaction to cell proliferation, we tested the effects of
miR-328 overexpression on HeLa cells by using the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bro-
mide (MTT) assay. We generated an expression vector
containing the genomic region flanking miR-328
(pcDNA3.1-miR-328) (Materials and methods). Upon
transient transfection in HeLa cells, significant expres-
sion of miR-328 was detected by qRT-PCR (data not
shown). Transfection with pcDNA3.1-miR-328
enhanced proliferation of HeLa cells in a dose-depen-
dent manner (Fig. 3A). We also found that knock-
down of PTPRJ expression with siRNA induced a
significant increase in cell proliferation (Fig. 3B),
similar to the effects of miR-328 overexpression.
Importantly, miR-328 transfection in siRNA-treated
cells did not further increase cell proliferation
(Fig. 3B), indicating that the effect of miR-328 on pro-
liferation was due, at least in part, to downregulation
of PTPRJ expression. In addition, PTPRJ overexpres-
sion (Fig. S1) rescued the cell proliferation effect
observed in HeLa cells treated with the miR-328
construct (Fig. 3C). To confirm the effect of miR-328
on cell proliferation, an in vitro cell proliferation assay
was performed in HeLa cells by using the fluorescent
dye carboxyfluorescein succinimidyl ester (CFSE),
which labels the intracellular protein content. With
each cell division, intracellular fluorescence decreases
with increasing cell number, the intensity of which can
be assayed by flow cytometry. Proliferation in HeLa
cells increased with both pcDNA3.1-miR-328 and
pcDNA3.1 + PTPRJ-siRNA compared with empty
00.20.40.60.8
11.21.41.61.8
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
00.20.40.60.8
11.21.41.61.8
Rel
ativ
e ce
ll pr
olife
ratio
n
pcDNA3.1 (ng) pcDNA3.1-miR-328 (ng)
250 210 40
100 150 250 –
–
**
*
pCEFL miR-328 pCEFL-PTPRJ
+ ––
+ + –
–+ +
–––
Rel
ativ
e ce
ll pr
olife
ratio
n
* **
siRNA-PTPRJ pcDNA3.1 pcDNA3.1-miR-328
+ + –
+ –+
–– –
–– +
Rel
ativ
e ce
ll pr
olife
ratio
n
Lipo + + + +
Lipo + + + +
CFSE fluorescence
Num
ber o
f cel
ls
0 h
24 h
100 101 102 103 104
100 101 102 103 104
020406080100
020406080
100
pcDNA3.1 pcDNA3.1-miR-328 pcDNA3.1+ siRNA PTPRJ
0
10
20
30
Geo
met
ric m
ean
CFS
E
**
A B
C D E
Fig. 3. MiR-328 increases proliferation in HeLa cells by repressing PTPRJ. (A) Proliferation of HeLa cells transfected with vectors is indi-
cated. Cell proliferation was measured 48 h after seeding with the MTT assay and values were normalized to the empty vector. *P < 0.05
compared with pcDNA3.1 (250 ng), analysed by ANOVA followed by Dunnett’s test. (B) Proliferation of HeLa cells co-transfected with
pcDNA3.1 (250 ng) + siRNA-PTPRJ (100 nM), pcDNA3.1-miR-328 (250 ng) and pcDNA3.1-miR-328 (250 ng) + siRNA-PTPRJ (100 nM). Cell
proliferation was measured after 48 h by MTT assay and values were normalized to the Lipofectamine. *P < 0.05 compared with Lipofecta-
mine, analysed by ANOVA followed by Dunnett’s test. (C) Proliferation of HeLa cells co-transfected with synthetic miR-328
(100 nM) + pCEFL (250 ng) and synthetic miR-328 (100 nM) + pCEFL-PTPRJ (250 ng). Cell proliferation was measured after 48 h by MTT
assay, and values were normalized to the Lipofectamine. *P < 0.05 compared with Lipofectamine, analysed by ANOVA followed by Dun-
nett’s test. (D) Representative cell cycle progression of HeLa cells transiently transfected with pcDNA3.1, pcDNA3.1-miR-328 and
pcDNA3.1 + siRNA-PTPRJ using CFSE. Cells were analysed at 0 and 24 h by flow cytometry. (E) Geometric mean intensities of CFSE stain-
ing in HeLa cells transiently transfected with pcDNA3.1, pcDNA3.1-miR-328 and pcDNA3.1 + siRNA-PTPRJ at 24 h. P values were gener-
ated by ANOVA followed by Bonferroni’s test for multiple comparisons (significant differences are indicated by an asterisk, P < 0.05).
F. Paduano et al. PTPRJ expression is negatively regulated by microRNA-328
FEBS Journal (2012) ª 2012 The Authors Journal compilation ª 2012 FEBS 5

vector, confirming that miR-328-dependent induction
of cell proliferation can also be achieved by repressing
PTPRJ levels (Fig. 3D,E). The proliferative effect of
miR-328 was also evaluated in SKBr3 cells. MiR-328
increased cell proliferation in transiently transfected
SKBr3 cells as demonstrated by MTT and CFSE
assays (Fig. S2).
To further investigate the link between PTPRJ and
miR-328, we generated stable HeLa cell lines overex-
pressing miR-328. Expression of miR-328 was quanti-
fied by qRT-PCR and two clones, miR-328 clone-1
and miR-328 clone-2, showed 50- and 80-fold
increases, respectively, in miR-328 expression com-
pared with empty vector control (Fig. 4A). Western
blot analysis of the HeLa miR-328 clones confirmed
PTPRJ downregulation (Fig. 4B). Additionally, flow
cytometry showed a reduction in membrane PTPRJ
expression in the miR-328 stable clones compared with
empty vector control (Fig. 4C). Finally, we aimed to
elucidate the effects of miR-328 overexpression on pro-
liferation rates in HeLa cells. This was performed by
using the MTT assay, and the miR-328 stable clones
showed an increase in cell proliferation compared with
empty vector control (Fig. 4D). This was also con-
firmed by the CFSE assay: clearly, the two miR-328
clones displayed an increase in cell proliferation com-
pared with empty vector control (Fig. 4E,F). PTPRJ is
a negative regulator of cell migration [11]. To investi-
gate whether miR-328 affects HeLa cell migration, we
used the wound-healing assay. We found that cell
migration was increased in both miR-328 stable clones
compared with empty vector control (Fig. S3).
Fig. 4. HeLa clones stably expressing miR-328 decrease PTPRJ expression and induce cell proliferation. (A) Relative miR-328 expression in
two HeLa stable clones (miR-328 clone-1 and miR-328 clone-2) quantified by qRT-PCR. Values were normalized to pcDNA3.1 vector.
*P < 0.01. (B) Western blot analysis of PTPRJ expression in HeLa stable clones. Expression is normalized to c-tubulin. Relative levels of
PTPRJ were quantified by QUANTITY ONE software (BioRad Laboratories). (C) Fluorescence-activated cell sorting analysis of membrane-bound
PTPRJ expression in HeLa clones stably expressing miR-328 or empty vector. Grey peaks represent unlabelled cells; open peaks represent
cells labelled with PTPRJ antibody. (D) Proliferation of HeLa stable clones measured after 24 and 48 h with the MTT assay. Values were nor-
malized to the empty vector. *P < 0.05 compared with pcDNA3.1 at 24 h, analysed by ANOVA followed by Dunnett’s test. (E) Representa-
tive cell cycle progression of HeLa stable clones using CFSE. Cells were analysed at 0, 24 and 48 h by flow cytometry. (F) Geometric mean
intensities of CFSE staining in HeLa stable clones at 24 and 48 h. *P < 0.05 compared with pcDNA3.1, analysed by ANOVA followed by
Bonferroni’s test.
PTPRJ expression is negatively regulated by microRNA-328 F. Paduano et al.
6 FEBS Journal (2012) ª 2012 The Authors Journal compilation ª 2012 FEBS

Discussion
Several studies have addressed the function of PTPRJ in
the regulation of cellular pathways and a number of
PTPRJ-interacting proteins or substrates have been iden-
tified [1], underlining the role ofPTPRJ as a negative reg-
ulator of cell proliferation in epithelial [10,12–14] and
endothelial [32] cells. PTPRJ acts by downregulating
receptor-type tyrosine kinase activity [2–6] or by inhibit-
ing the pro-proliferative PI3K and Ras-dependent path-
ways [33,34]. To clarify the role of PTPRJ in vivo,
knockout mice were generated resulting in viable mice
that do not develop tumours spontaneously [35]. How-
ever, significant downregulation of PTPRJ expression
has been observed in tumours [15,16]. Thus, it is impor-
tant to study the mechanisms involved in the regulation
of PTPRJ expression that is currently not verywell under-
stood. In general, regulation of the expression of protein
tyrosine phosphatases is poorly characterized [36].
Post-transcriptional regulation of PTPRJ has been
studied by Karagyozov et al. [37], who found a novel
mechanism of translational regulation that was based
on the use of different translational start sites. Our
study provides an independent mechanism of control of
PTPRJ expression which involves the 3¢UTR. We used
three independent bioinformatics algorithms to identify
MREs in the 3¢UTR of PTPRJ. Interestingly, MRE_4,
which was the only MRE to be detected by all three
algorithms, appeared to interact with miR-328.
Recently, a novel mechanism of action has been
attributed to miR-328. In fact, miR-328 acts through a
decoy activity that interferes with the function of regula-
tory proteins [38]. However, miR-328 is also able to
repress gene expression through the typical miRNA
mechanism of acting on the 3¢UTR of target genes [38],
as shown here.
In our study, miR-328 increased proliferation of
HeLa and SKBr3 cells by means of a mechanism
dependent, at least in part, on the downregulation of
PTPRJ expression. The number of possible miR-328
gene targets in the cell lines analysed probably influ-
enced the extent of PTPRJ downregulation. However,
effects on cell proliferation would not necessarily have
to correlate with the extent of PTPRJ downregulation
as different cell lines could have varied sensitivities to
expression of PTPRJ, or other targets of miR-328.
The list of validated targets of miR-328 includes
CD44, ABCG2 and PIM-1 [38–40]. CD44 is involved
in cell adhesion and could act synergistically with
PTPRJ. Given that CD44 and PIM-1 are considered
pro-oncogenic proteins, this could appear to contrast
with the tumour suppressor role of PTPRJ. However,
the oncogenic activity of PIM-1 is mainly restricted to
haematological malignancies [41] in which, it should be
emphasized, the role of PTPRJ is still unclear. Indeed,
CD148 ⁄PTPRJ has been proposed to be a valuable
marker of mantle cell lymphomas in an unbiased pro-
teomic study [42].
Loss of expression of miR-328 is strongly correlated
with blast crisis in patients afflicted with chronic mye-
logenous leukaemia [38]. In glioblastoma multiforme,
miR-328 levels are lower compared with normal brain
tissues [43]. Conversely, miR-328 levels are upregulated
in patients with brain metastasis of lung cancer [44].
Expression levels of miR-328 are significantly higher in
blood samples of melanoma patients compared with
healthy individuals [45].
Studies on miR-328 discussed thus far reflect the
well understood feature of miRNAs: they can act as
onco-miRs (favouring tumour progression) or onco-
suppressor miRs (blocking tumour progression)
depending on the cellular context, as exemplified by
miR-221 and miR-222 that behave as tumour suppres-
sors in haematological malignancies and as oncogenes
in other tumours [46]. In fact, the role of an miRNA
in a pathological context is also dependent on the rela-
tive expression levels of its targets in the tissue under
examination [47]. However, in epithelium-derived cells,
miR-328 increases cell proliferation (our results) and
cell migration [44], and this is in line with the opposing
role played by PTPRJ in these processes [10–15,48,49].
In conclusion, we have identified a novel mechanism
of regulation of PTPRJ expression in which the
3¢UTR of PTPRJ mRNA is targeted by miR-328. This
interaction is important for proliferation of epithelium-
derived cells. It would be of interest to investigate in
future studies the correlation between expression of
PTPRJ and miR-328 in human tumours.
Materials and methods
Bioinformatic analysis
The algorithms used were mirtar, rnahybrid and find-
tar. We first screened the possible miRNAs interacting with
the 3¢UTR of PTPRJ by using mirtar [28], a bioinformat-
ics algorithm that allows a selection of potential interacting
miRNAs with a given 3¢UTR sequence. Every miRNA can-
didate resulting from that analysis was further investigated
with findtar [29] and rnahybrid [30] to determine the best
possible miRNA candidate for the interaction with the
3¢UTR of PTPRJ. The threshold value of minimum free
energy for the identification of MREs located on PTPRJ
3¢UTR was set at the default value ()16 kcalÆmol)1) in the
screening with mirtar and at )25 kcalÆmol)1 in the analyses
with findtar and rnahybrid.
F. Paduano et al. PTPRJ expression is negatively regulated by microRNA-328
FEBS Journal (2012) ª 2012 The Authors Journal compilation ª 2012 FEBS 7

Cells and transfections
HeLa (human cervix adenocarcinoma) and SKBr3 (human
breast adenocarcinoma) cell lines were maintained in culture
at 37 �C and 5% CO2 in DMEM supplemented with 10%
fetal bovine serum, 100 U penicillin and 100 lgÆmL)1 strepto-
mycin sulfate. Transfections were made with Lipofectamine�
2000, following the manufacturer’s instructions (Invitrogen,
Carlsbad, CA, USA). For western blot and qRT-PCR experi-
ments, 4 · 105 cells were seeded in six-well plates and trans-
fected with 50 pmol Pre-miRTM (Ambion, Life Technologies,
Paisley, UK) or 100 pmol siRNA. For luciferase assays, 2 ·05 cells were seeded in 24-well plates and co-transfected with
10 pmolÆmL)1 Pre-miRTM (Ambion) and 25 ngÆmL)1 repor-
ter plasmid (pmirGLO vector; Promega,Madison,WI, USA).
Western blot analysis
Cells were harvested after 48 h, washed with ice-cold
NaCl ⁄Pi and lysed in a buffer containing 50 mm Tris ⁄HCl
pH 7.5, 1% Nonidet P40, 150 mm NaCl, 25 mm NaF, 1 mm
Na3VO4 and a complete mixture of protease inhibitors
(Roche, Basel, Switzerland). Lysates were clarified by centri-
fugation at 10 000 g for 15 min at 4 �C. Protein concentra-
tion was determined by a modified Bradford assay (BioRad,
Hercules, CA, USA). Fifty micrograms of proteins were sub-
jected to electrophoresis carried out in an 8%–12% gradient
gel (Invitrogen) and then transferred to nitrocellulose mem-
branes (Hybond-C; Amersham Biosciences, Piscataway, NJ,
USA). The membranes were blocked with 5% non-fat dry
milk and then probed for 2 h with the appropriate primary
antibodies. After incubation with specific horseradish peroxi-
dase conjugated secondary antibodies, protein bands were
revealed with an enhanced chemiluminescence system (Santa
Cruz Biotechnology, Santa Cruz, CA, USA). Primary anti-
bodies used were anti-human PTPRJ and anti-tubulin pur-
chased from R&D Systems (Minneapolis, MN, USA) and
Santa Cruz Biotechnology, respectively.
Quantitative real-time PCR
RNA extraction was performed with miRNeasy Mini Kit�(Qiagen, Valencia, CA, USA), following the manufacturer’s
instructions, and total RNA was quantified with a spectro-
photometer. Total RNA samples (250 ng) were subjected to
the reaction of reverse-transcription using the High Capa-
city RNA-to-cDNA Kit (Applied Biosystems, Foster City,
CA, USA), following the manufacturer’s instructions. Five
hundred nanolitres of cDNAs were amplified by real-time
PCR with Promega SYBR green kit and 5 pmol of primers
in a total volume of 25 lL.The primers used were as follows: 5¢-GTATTAT
CATTGGTGGCTTGTTC-3¢ (forward for long and short
form of PTPRJ), 5¢-CATCTCCGTGGTGGTGAC-3¢ (reversefor long form of PTPRJ) and 5¢-AGGCAGGTGTTCAA
ATCATCC-3¢ (reverse for short form ofPTPRJ). Specific oligo-
nucleotides used for hypoxanthine phosphoribosyltransferase
(normalization control) amplification were reported by Vande-
sompele et al. [50].
Real-time PCR reactions were carried out in a BioRad
iQ�5 apparatus with the following conditions: initial dena-
turation step at 95 �C for 3 min, followed by 40 cycles of
10 s at 95 �C and 1 min at 57 �C. Specificity of PCR prod-
ucts was checked by melting curve analysis and gel electro-
phoresis. Efficiencies of real-time PCRs were calculated by
constructing curves with cycle threshold (Ct) obtained from
amplifications of serial tenfold dilutions of a cDNA sam-
ple, resulting in higher than 90%. For miR-328 quantifica-
tion, 50 ng of total RNA was subjected to reverse-
transcription with miRCURY LNA� Universal cDNA
synthesis kit (Exiqon, Vedbaek, Denmark), following the
manufacturer’s instructions. One hundredth of the cDNA
was amplified by real-time PCR with SYBR� Green master
mix 2X (Exiqon) and miRCURY LNA� hsa-miR-328 spe-
cific PCR primer set (Exiqon) or miRCURY LNA� U6
snRNA specific PCR primer set (Exiqon), following the
manufacturer’s instructions. Reactions were performed in a
total volume of 20 lL in a BioRad IQ�5 apparatus.
Plasmids and mutagenesis
The 3¢UTR of PTPRJ containing the predicted sites for
miR-328 ⁄PTPRJ interaction (the region from +6259 to
+7761 of PTPRJ) was amplified by PCR using genomic
DNA as the template, Pfu polymerase and the following
primers: forward 5¢-CATGTTTAAACCTTCTCAAATG
GAAATTGCA-3¢ and reverse 5¢- GCCCTCGAGCAACC
AACAGCAATACTCTGTA-3¢. The amplified product was
purified and then cloned in pmirGLO vector (Promega) at
PmeI and XhoI restriction sites, downstream of the firefly
luciferase reporter gene.
A deletion mutant (Delta) was generated by exciding from
pmirGLO-PTPRJ the PmeI-SacI fragment and re-ligating
the plasmid after blunting of the SacI digested end. Site-
specific mutants of Delta plasmid were generated by PCR
mutagenesis. Primers used were forward PmeI, 5¢-CATGTTTAAACCTCAAATTGGAAGAGTCC-3¢; reverseXhoI, 5¢- GCCCTCGAGCAACCAACAGCAATACTCT-
GTA-3¢; forward mut7440, 5¢-GAGGGGCGGAAATAAG-
CTATGCAGC-3¢; reverse mut7440, 5¢-GCTGCATAGCT-
TATTTCCGCCCCTC-3¢; forward mut7174: 5¢-GGTGT
CAATGAAAAAACAAAAGACTGGTG-3¢; reverse mut71
74, 5¢-CACCAGTCTTTTGTTTTTTCATTGACACC-3¢.PCR amplification was performed using Pfu polymerase
(Promega) and pmirGLO-Delta mutant as the template.
Mutants were all cloned at PmeI and XhoI sites of pmirGLO.
The 75 nucleotide precursor hairpin sequences of miR-328
and 100 nucleotides of genomic sequences flanking each side
of the hairpin sequence were amplified by PCR using the
following primers: forward, 5¢-CACGAATTCCCTTGTC
PTPRJ expression is negatively regulated by microRNA-328 F. Paduano et al.
8 FEBS Journal (2012) ª 2012 The Authors Journal compilation ª 2012 FEBS

GAAGTCCTCCTGTGTAG-3¢; reverse, 5¢- CACCTCGAG
GACTCCTTGCCCCATTCTCTGCG-3¢.To generate the miR-328 expression construct, the 467-
nucleotide DNA fragment obtained was digested with
EcoRI and XhoI and inserted into the EcoRI-XhoI sites of
the pCDNA3.1 expression vector (Invitrogen). All plasmids
were checked by sequence analysis.
Luciferase assay
HeLa transfected cells were lysed in a passive lysis buffer
(Promega). One hundred microlitres of luciferase assay
reagent 2 were added to 20 lL of lysate, and samples were
analysed in a luminometer (firefly reading). After the addi-
tion of Stop and Glo reagent (Promega), the samples were
again analysed in the luminometer (Renilla reading). Values
were expressed as the ratio of firefly ⁄Renilla readings.
Cell proliferation assay
HeLa cells were plated in 96-well plates at a density of
2 · 103 cells per well in 100 lL of medium and were trans-
fected with siRNA and ⁄or plasmid DNA. Each condition
was plated in octuplicate. Cell proliferation rates were deter-
mined 48 h post-transfection with the MTT assay (Sigma-
Aldrich, St Louis, MO, USA). Absorbance of the formazan
dye that is formed was measured at a wavelength of 570 nm,
using a Microplate Reader (BioRad); absorbance is propor-
tional to the number of metabolically active cells in culture.
For CFSE staining, cells were harvested and washed twice
with NaCl ⁄Pi, followed by labelling with 5 mm CFSE for
5 min at room temperature. Residual CFSE was removed by
washing twice with NaCl ⁄Pi, and cells were seeded in six-well
plates containing growth medium. CFSE fluorescence was
measured by fluorescence-activated cell sorting analysis, and
data were analysed with flowjo software (Tree Star,
Ashland, OR, USA).
Flow cytometry analysis
Cells were centrifuged at 400 g for 3 min, the supernatant
was discarded and the cells were re-suspended in 500 lL of
NaCl ⁄Pi. Briefly, cells were blocked with 1% BSA in
NaCl ⁄Pi, washed once and incubated for 20 min with
PTPRJ primary antibody (R&D), followed by incubation
with phycoerythrin-conjugated secondary antibody. Samples
were analysed with FACSCan instruments (Becton Dickinson,
Franklin Lakes, NJ, USA). Data were analysed with flow-
jo software (Tree Star).
Generation of stable cell clones
HeLa cells were transfected using Lipofectamine 2000 (Invi-
trogen). Cells were plated into selection medium containing
0.4 mgÆmL)1 G418 (Sigma Aldrich) 1 day post-transfection.
After 25 days of selection, individual G418-resistant colo-
nies were isolated, expanded and analysed for miR-328
expression levels by qRT-PCR.
Cell migration assay
Cell migration assay was performed using the scratch
wound-healing method on HeLa cells that were 80–90%
confluent in a six-well plate format. Cell migration was
monitored by microscopy. Images acquired for each sample
were then quantitatively analysed. For each image, dis-
tances between the points of the scratch were measured. By
comparing the images from t = 0 to t = 12 h, which was
the last time point, the distance of each scratch closure was
obtained by measuring the distances using imagej software
(NIH Image, Bethesda, MD, USA).
Statistical analyses
Results are expressed as the mean ± standard deviation of
three separate experiments. The significance of differences
between groups was evaluated by using Student’s t test or one-
way analysis of variance (ANOVA), followed by Dunnett’s
test ⁄Bonferroni’s test for multiple comparisons. Analysis was
conducted using the graphpad prism software (San Diego, CA,
USA), and differences were considered significant if P < 0.05.
Acknowledgements
The present study was supported by a grant from Min-
istero dell’Istruzione, dell’Universita e della Ricerca
(MIUR) to CP and RI (No. 2007F7T537_003) and a
grant from the Associazione Italiana Ricerca Cancro
(AIRC) to FT. FP was a recipient of an FIRC (Fond-
azione Italiana Ricerca Cancro) fellowship.
References
1 Ostman A, Hellberg C & Bohmer FD (2006)
Protein-tyrosine phosphatases and cancer. Nat Rev
Cancer 6, 307–320.
2 Tarcic G, Boguslavsky SK, Wakim J, Kiuchi T, Liu A,
Reinitz F, Nathanson D, Takahashi T, Mischel PS, Ng
T et al. (2009) An unbiased screen identifies DEP-1
tumor suppressor as a phosphatase controlling EGFR
endocytosis. Curr Biol 19, 1788–1798.
3 Palka HL, Park M & Tonks NK (2003) Hepatocyte
growth factor receptor tyrosine kinase met is a substrate
of the receptor protein-tyrosine phosphatase DEP-1.
J Biol Chem 278, 5728–5735.
4 Kovalenko M, Denner K, Sandstrom J, Persson C,
Gross S, Jandt E, Vilella R, Bohmer F & Ostman A
(2000) Site selective dephosphorylation of the platelet-
F. Paduano et al. PTPRJ expression is negatively regulated by microRNA-328
FEBS Journal (2012) ª 2012 The Authors Journal compilation ª 2012 FEBS 9

derived growth factor beta-receptor by the receptor-like
protein tyrosine phosphatase DEP-1. J Biol Chem 275,
16219–16226.
5 Iervolino A, Iuliano R, Trapasso F, Viglietto G, Melillo
RM, Carlomagno F, Santoro M & Fusco A (2006) The
receptor-type protein tyrosine phosphatase J
antagonizes the biochemical and biological effects of
RET-derived oncoproteins. Cancer Res 66, 6280–6287.
6 Lampugnani MG, Zanetti A, Corada M, Takahashi T,
Balconi G, Breviario F, Orsenigo F, Cattelino A,
Kemler R, Daniel TO et al. (2003) Contact inhibition
of VEGF-induced proliferation requires vascular
endothelial cadherin, beta-catenin, and the phosphatase
DEP-1 ⁄CD148. J Cell Biol 161, 793–804.
7 Le Pera I, Iuliano R, Florio T, Susini C, Trapasso F,
Santoro M, Chiariotti L, Schettini G, Viglietto G &
Fusco A (2005) The rat tyrosine phosphatase eta
increases cell adhesion by activating c-Src through
dephosphorylation of its inhibitory phosphotyrosine
residue. Oncogene 24, 3187–3195.
8 Holsinger LJ, Ward K, Duffield B, Zachwieja J & Jallal
B (2002) The transmembrane receptor protein tyrosine
phosphatase DEP1 interacts with p120(ctn). Oncogene
21, 7067–7076.
9 Sallee JL & Burridge K (2009) Density-enhanced phos-
phatase 1 regulates phosphorylation of tight junction
proteins and enhances barrier function of epithelial
cells. J Biol Chem 284, 14997–15006.
10 Keane MM, Lowrey GA, Ettenberg SA, Dayton MA &
Lipkowitz S (1996) The protein tyrosine phosphatase
DEP-1 is induced during differentiation and inhibits
growth of breast cancer cells. Cancer Res 56,
4236–4243.
11 Jandt E, Denner K, Kovalenko M, Ostman A &
Bohmer FD (2003) The protein-tyrosine phosphatase
DEP-1 modulates growth factor-stimulated cell migra-
tion and cell–matrix adhesion. Oncogene 22, 4175–4185.
12 Trapasso F, Yendamuri S, Dumon KR, Iuliano R,
Cesari R, Feig B, Seto R, Infante L, Ishii H, Vecchione
A et al. (2004) Restoration of receptor-type protein
tyrosine phosphatase eta function inhibits human
pancreatic carcinoma cell growth in vitro and in vivo.
Carcinogenesis 25, 2107–2114.
13 Balavenkatraman KK, Jandt E, Friedrich K, Kauten-
burger T, Pool-Zobel BL, Ostman A & Bohmer FD
(2006) DEP-1 protein tyrosine phosphatase inhibits pro-
liferation and migration of colon carcinoma cells and is
upregulated by protective nutrients. Oncogene 25,
6319–6324.
14 Iuliano R, Trapasso F, Le Pera I, Schepis F, Sama I,
Clodomiro A, Dumon KR, Santoro M, Chiariotti L,
Viglietto G et al. (2003) An adenovirus carrying the rat
protein tyrosine phosphatase eta suppresses the growth
of human thyroid carcinoma cell lines in vitro and
in vivo. Cancer Res 63, 882–886.
15 Trapasso F, Iuliano R, Boccia A, Stella A, Visconti R,
Bruni P, Baldassarre G, Santoro M, Viglietto G &
Fusco A (2000) Rat protein tyrosine phosphatase eta
suppresses the neoplastic phenotype of retrovirally
transformed thyroid cells through the stabilization of
p27(Kip1). Mol Cell Biol 20, 9236–9246.
16 Massa A, Barbieri F, Aiello C, Arena S, Pattarozzi A,
Pirani P, Corsaro A, Iuliano R, Fusco A, Zona G et al.
(2004) The expression of the phosphotyrosine phospha-
tase DEP-1 ⁄PTPeta dictates the responsivity of glioma
cells to somatostatin inhibition of cell proliferation.
J Biol Chem 279, 29004–29012.
17 Ruivenkamp CA, van Wezel T, Zanon C, Stassen AP,
Vlcek C, Csikos T, Klous AM, Tripodis N, Perrakis A,
Boerrigter L et al. (2002) Ptprj is a candidate for the
mouse colon-cancer susceptibility locus Scc1 and is
frequently deleted in human cancers. Nat Genet 31,
295–300.
18 Iuliano R, Le Pera I, Cristofaro C, Baudi F, Arturi F, Pal-
lante P, Martelli ML, Trapasso F, Chiariotti L & Fusco A
(2004) The tyrosine phosphatase PTPRJ ⁄DEP-1 genotype
affects thyroid carcinogenesis.Oncogene 23, 8432–8438.
19 Cuozzo C, Porcellini A, Angrisano T, Morano A, Lee
B, Di Pardo A, Messina S, Iuliano R, Fusco A, Santillo
MR et al. (2007) DNA damage, homology-directed
repair, and DNA methylation. PLoS Genet 3, e110.
20 Venkatachalam R, Ligtenberg MJ, Hoogerbrugge N,
Schackert HK, Gorgens H, Hahn MM, Kamping EJ,
Vreede L, Hoenselaar E, van der Looij E et al. (2010)
Germline epigenetic silencing of the tumor suppressor
gene PTPRJ in early-onset familial colorectal cancer.
Gastroenterology 139, 2221–2224.
21 Bartel DP (2009) MicroRNAs: target recognition and
regulatory functions. Cell 136, 215–233.
22 Calin GA, Ferracin M, Cimmino A, Di Leva G, Shi-
mizu M, Wojcik SE, Iorio MV, Visone R, Sever NI,
Fabbri M et al. (2005) A microRNA signature associ-
ated with prognosis and progression in chronic lym-
phocytic leukemia. N Engl J Med 353, 1793–1801.
23 Visone R, Pallante P, Vecchione A, Cirombella R,
Ferracin M, Ferraro A, Volinia S, Coluzzi S, Leone V,
Borbone E et al. (2007) Specific microRNAs are down-
regulated in human thyroid anaplastic carcinomas.
Oncogene 26, 7590–7595.
24 Garzon R, Calin GA & Croce CM (2009) MicroRNAs
in cancer. Annu Rev Med 60, 167–179.
25 Calin GA & Croce CM (2006) MicroRNAs and chro-
mosomal abnormalities in cancer cells. Oncogene 25,
6202–6210.
26 Bueno MJ, Perez de Castro I, Gomez de Cedron M,
Santos J, Calin GA, Cigudosa JC, Croce CM, Fern-
andez-Piqueras J & Malumbres M (2008) Genetic and
epigenetic silencing of microRNA-203 enhances ABL1
and BCR-ABL1 oncogene expression. Cancer Cell 13,
496–506.
PTPRJ expression is negatively regulated by microRNA-328 F. Paduano et al.
10 FEBS Journal (2012) ª 2012 The Authors Journal compilation ª 2012 FEBS

27 Jazdzewski K, Murray EL, Franssila K, Jarzab B,
Schoenberg DR & de la Chapelle A (2008) Common
SNP in pre-miR-146a decreases mature miR expression
and predisposes to papillary thyroid carcinoma. Proc
Natl Acad Sci USA 105, 7269–7274.
28 Hsu JB, Chiu CM, Hsu SD, Huang WY, Chien CH,
Lee TY & Huang HD (2011) miRTar: an integrated
system for identifying miRNA-target interactions in
human. BMC Bioinformatics 12, 300.
29 Ye W, Lv Q, Wong CK, Wu S, Fu C, Hua Z, Cai G,
Li G, Yang BB & Zhang Y (2008) The effect of central
loops in miRNA:MRE duplexes on the efficiency of
miRNA-mediated gene regulation. PLoS One 3, e1719.
30 Rehmsmeier M, Steffen P, Hochsmann M & Giegerich
R (2004) Fast and effective prediction of microR-
NA ⁄ target duplexes. RNA 10, 1507–1517.
31 Iuliano R, Raso C, Quintiero A, Pera IL, Pichiorri F,
Palumbo T, Palmieri D, Pattarozzi A, Florio T, Vigli-
etto G et al. (2009) The eighth fibronectin type III
domain of protein tyrosine phosphatase receptor
J influences the formation of protein complexes and cell
localization. J Biochem 145, 377–385.
32 Takahashi T, Takahashi K, Mernaugh RL, Tsuboi N,
Liu H & Daniel TO (2006) A monoclonal antibody
against CD148, a receptor-like tyrosine phosphatase,
inhibits endothelial-cell growth and angiogenesis. Blood
108, 1234–1242.
33 Tsuboi N, Utsunomiya T, Roberts RL, Ito H, Takah-
ashi K, Noda M & Takahashi T (2008) The tyrosine
phosphatase CD148 interacts with p85 regulatory
subunit of PI 3-kinase. Biochem J 413, 193–200.
34 Sacco F, Tinti M, Palma A, Ferrari E, Nardozza AP,
Hooft van Huijsduijnen R, Takahashi T, Castagnoli L
& Cesareni G (2009) Tumor suppressor density-
enhanced phosphatase-1 (DEP-1) inhibits the RAS
pathway by direct dephosphorylation of ERK1 ⁄ 2kinases. J Biol Chem 284, 22048–22058.
35 Trapasso F, Drusco A, Costinean S, Alder H, Aqeilan
RI, Iuliano R, Gaudio E, Raso C, Zanesi N, Croce CM
et al. (2006) Genetic ablation of Ptprj, a mouse cancer
susceptibility gene, results in normal growth and devel-
opment and does not predispose to spontaneous tumor-
igenesis. DNA Cell Biol 25, 376–382.
36 Den Hertog J, Ostman A & Bohmer FD (2008) Protein
tyrosine phosphatises: regulatory mechanisms. FEBS J
275, 831–847.
37 Karagyozov L, Godfrey R, Bohmer SA, Petermann A,
Holters S, Ostman A & Bohmer FD (2008) The struc-
ture of the 5¢-end of the protein-tyrosine phosphatase
PTPRJ mRNA reveals a novel mechanism for
translation attenuation. Nucleic Acids Res 36, 4443–
4453.
38 Eiring AM, Harb JG, Neviani P, Garton C, Oaks JJ,
Spizzo R, Liu S, Schwind S, Santhanam R, Hickey CJ
et al. (2010) miR-328 functions as an RNA decoy to
modulate hnRNP E2 regulation of mRNA translation
in leukemic blasts. Cell 140, 652–665.
39 Wang CH, Lee DY, Deng Z, Jeyapalan Z, Lee SC,
Kahai S, Lu WY, Zhang Y & Yang BB (2008)
MicroRNA miR-328 regulates zonation morphogenesis
by targeting CD44 expression. PLoS One 3, e2420.
40 Pan YZ, Morris ME & Yu AM (2009) MicroRNA-328
negatively regulates the expression of breast cancer
resistance protein (BCRP ⁄ABCG2) in human cancer
cells. Mol Pharmacol 75, 1374–1379.
41 Brault L, Gasser C, Bracher F, Huber K, Knapp S
& Schwaller J (2010) PIM serine ⁄ threonine kinases in
the pathogenesis and therapy of hematologic
malignancies and solid cancers. Haematologica 95,
1004–1015.
42 Miguet L, Bechade G, Fornecker L, Zink E, Felden
C, Gervais C, Herbrecht R, Van Dorsselaer A, Mau-
vieux L & Sanglier-Cianferani S (2009) Proteomic
analysis of malignant B-cell derived microparticles
reveals CD148 as a potentially useful antigenic biomar-
ker for mantle cell lymphoma diagnosis. J Proteome
Res 8, 3346–3354.
43 Malzkorn B, Wolter M, Liesenberg F, Grzendowski M,
Stuhler K, Meyer HE & Reifenberger G (2010) Identifi-
cation and functional characterization of microRNAs
involved in the malignant progression of gliomas. Brain
Pathol 20, 539–550.
44 Arora S, Ranade AR, Tran NL, Nasser S, Sridhar S,
Korn RL, Ross JT, Dhruv H, Foss KM, Sibenaller Z
et al. (2011) MicroRNA-328 is associated with (non-
small) cell lung cancer (NSCLC) brain metastasis and
mediates NSCLCmigration. Int J Cancer 129, 2621–2631.
45 Leidinger P, Keller A, Borries A, Reichrath J, Rass K,
Jager SU, Lenhof HP & Meese E (2010) High-through-
put miRNA profiling of human melanoma blood
samples. BMC Cancer 10, 262.
46 Di Leva G & Croce CM (2010) Roles of small RNAs
in tumor formation. Trends Mol Med 16, 257–267.
47 Leung AK & Sharp PA (2010) MicroRNA functions in
stress responses. Mol Cell 40, 205–215.
48 Brunner PM, Heier PC, Mihaly-Bison J, Priglinger U,
Binder BR & Prager GW (2011) Density enhanced
phosphatase-1 down-regulates urokinase receptor
surface expression in confluent endothelial cells. Blood
117, 4154–4161.
49 Petermann A, Haase D, Wetzel A, Balavenkatraman
KK, Tenev T, Guhrs KH, Friedrich S, Nakamura M,
Mawrin C & Bohmer FD (2011) Loss of the protein-
tyrosine phosphatase DEP-1 ⁄PTPRJ drives meningioma
cell motility. Brain Pathol 21, 405–418.
50 Vandesompele J, De Preter K, Pattyn F, Poppe B, Van
Roy N, De Paepe A & Speleman F (2002) Accurate
normalization of real-time quantitative RT-PCR data
by geometric averaging of multiple internal control
genes. Genome Biol 3, 0034.
F. Paduano et al. PTPRJ expression is negatively regulated by microRNA-328
FEBS Journal (2012) ª 2012 The Authors Journal compilation ª 2012 FEBS 11

Supporting information
The following supplementary material is available:
Fig. S1. PTPRJ protein expression in HeLa transfected
cells.
Fig. S2. miR-328 increases cell proliferation of SKBr3
cells.
Fig. S3. Stable transfection of miR-328 in HeLa cells
increased the cell migration.
This supplementary material can be found in the
online version of this article.
Please note: As a service to our authors and readers,
this journal provides supporting information supplied
by the authors. Such materials are peer-reviewed and
may be reorganized for online delivery, but are not
copy-edited or typeset. Technical support issues arising
from supporting information (other than missing files)
should be addressed to the authors.
PTPRJ expression is negatively regulated by microRNA-328 F. Paduano et al.
12 FEBS Journal (2012) ª 2012 The Authors Journal compilation ª 2012 FEBS




















