
An 18 kDa Acid Phosphatase from Chicken Heart Possesses Phosphotransferase Activity
12
An 18 kDa Acid Phosphatase from Chicken Heart Possesses Phosphotransferase Activity Rubina Naz, 1 Asma Saeed, 2 and Ahmad Saeed 1,3 A low molecular weight acid phosphatase was purified to homogeneity from chicken heart with a specific activity of 42 U/mg and a recovery of about 1%. Nearly 800 fold purification was achieved. The molecular weight was estimated to be 18 kDa by SDS-polyacrylamide gel electrophoresis. Para-nitrophenyl phosphate, phenyl phosphate and flavin mononucleotide were efficiently hydrolysed by the enzyme and found to be good substrates. Fluoride and tartrate had no inhibitory effect while phosphate, vanadate and molybdate strongly inhibited the enzyme. The acid phosphatase was stimulated in the presence of glycerol, ethylene glycol, methanol, ethanol and acetone, which reflected the phosphotransferase activity. When phosphate acceptors such as ethylene glycol concentrations were increased, the ratio of phosphate transfer to hydrolysis was also increased, demonstrating the presence of a transphosphorylation reaction where an acceptor can compete with water in the rate limiting step involving hydrolysis of a covalent phospho enzyme intermediate. Partition experiments carried out with two substrates, para-nitrophenyl phosphate and phenyl phosphate, revealed a constant product ratio of 1.7 for phosphotransfer to ethylene glycol versus hydrolysis, strongly supporting the existence of common covalent phospho enzyme intermediate. A constant ratio of K cat /K m , 4.310 4 , found at different ethylene glycol concentrations, also supported the idea that the rate limiting step was the hydrolysis of the phospho enzyme intermediate. KEY WORDS: 18 kDa Acid phosphatase isoenzymes; acid phosphatase; phosphotransferase; Phosp- hotyrosine protein phosphatase. 1. INTRODUCTION Acid phosphatases (EC 3.1.3.2) are a heterogeneous enzyme family which catalyse the hydrolysis of syn- thetic and natural phosphomonoesters with libera- tion of inorganic phosphate and the respective base alcohol or phenol (Bachwald et al., 1984; Vincent et al., 1992). Although the biological role of acid phosphatases still remains largely unknown, their involvement in the metabolism of phosphorylated compounds and their ubiquitous nature indicate their importance in cell physiology (Panara et al., 1998). Acid phosphatases are present in all mamma- lian tissues but differ in protein structure and enzyme properties (Ostrowsky and Kuciel, 1994; Panara et al., 1993; Vincent et al., 1992). Several mammalian tissues have been shown to contain at least three types of acid phosphatases, based on differences in molecular size; a high molec- ular weight (M r 80–200 kDa) acid phosphatase, an intermediate molecular weight (M r 35–50 kDa) form and a low molecular weight (M r 10–30 kDa) enzyme. These types are clearly distinguishable 1 Department of Chemistry Gomal University, Dera Ismail Khan, Pakistan. 2 Department of Biological Sciences Gomal University, Dera Ismail Khan, Pakistan. 3 To whom correspondence should be addressed. E-mail: [email protected] Abbreviations: DTT, dithiothreitol; EDTA, ethylenediamine tet- raacetate; FMN, flavin mononucleotide; M r , molecular weight; PAGE, polyacrylamide gel electrophoresis; PMSF, phenyl-meth- ylsulfonyl fluoride; PTPase, phosphotyrosine protein phosphatase; SDS, sodium dodecyl sulphate. 135 1572-3887/06/0200-0135/0 Ó 2006 Springer Science+Business Media, Inc. The Protein Journal, Vol. 25, No. 2, February 2006 (Ó 2006) DOI: 10.1007/s10930-006-0005-5
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of An 18 kDa Acid Phosphatase from Chicken Heart Possesses Phosphotransferase Activity

An 18 kDa Acid Phosphatase from Chicken HeartPossesses Phosphotransferase Activity
Rubina Naz,1 Asma Saeed,2 and Ahmad Saeed1,3
A low molecular weight acid phosphatase was purified to homogeneity from chicken heartwith a specific activity of 42 U/mg and a recovery of about 1%. Nearly 800 fold purification
was achieved. The molecular weight was estimated to be 18 kDa by SDS-polyacrylamide gelelectrophoresis. Para-nitrophenyl phosphate, phenyl phosphate and flavin mononucleotidewere efficiently hydrolysed by the enzyme and found to be good substrates. Fluoride and
tartrate had no inhibitory effect while phosphate, vanadate and molybdate strongly inhibitedthe enzyme. The acid phosphatase was stimulated in the presence of glycerol, ethylene glycol,methanol, ethanol and acetone, which reflected the phosphotransferase activity. Whenphosphate acceptors such as ethylene glycol concentrations were increased, the ratio of
phosphate transfer to hydrolysis was also increased, demonstrating the presence of atransphosphorylation reaction where an acceptor can compete with water in the rate limitingstep involving hydrolysis of a covalent phospho enzyme intermediate. Partition experiments
carried out with two substrates, para-nitrophenyl phosphate and phenyl phosphate, revealed aconstant product ratio of 1.7 for phosphotransfer to ethylene glycol versus hydrolysis, stronglysupporting the existence of common covalent phospho enzyme intermediate. A constant ratio
of Kcat/Km, 4.3�104, found at different ethylene glycol concentrations, also supported the ideathat the rate limiting step was the hydrolysis of the phospho enzyme intermediate.
KEY WORDS: 18 kDa Acid phosphatase isoenzymes; acid phosphatase; phosphotransferase; Phosp-hotyrosine protein phosphatase.
1. INTRODUCTION
Acid phosphatases (EC 3.1.3.2) are a heterogeneousenzyme family which catalyse the hydrolysis of syn-thetic and natural phosphomonoesters with libera-tion of inorganic phosphate and the respective basealcohol or phenol (Bachwald et al., 1984; Vincentet al., 1992). Although the biological role of acidphosphatases still remains largely unknown, theirinvolvement in the metabolism of phosphorylatedcompounds and their ubiquitous nature indicate
their importance in cell physiology (Panara et al.,1998). Acid phosphatases are present in all mamma-lian tissues but differ in protein structure and enzymeproperties (Ostrowsky and Kuciel, 1994; Panaraet al., 1993; Vincent et al., 1992).
Several mammalian tissues have been shown tocontain at least three types of acid phosphatases,based on differences in molecular size; a high molec-ular weight (Mr 80–200 kDa) acid phosphatase,an intermediate molecular weight (Mr 35–50 kDa)form and a low molecular weight (Mr 10–30 kDa)enzyme. These types are clearly distinguishable
1 Department of Chemistry Gomal University, Dera Ismail Khan,
Pakistan.2 Department of Biological Sciences Gomal University, Dera
Ismail Khan, Pakistan.3 To whom correspondence should be addressed. E-mail:
Abbreviations: DTT, dithiothreitol; EDTA, ethylenediamine tet-
raacetate; FMN, flavin mononucleotide; Mr, molecular weight;
PAGE, polyacrylamide gel electrophoresis; PMSF, phenyl-meth-
ylsulfonyl fluoride; PTPase, phosphotyrosine protein phosphatase;
SDS, sodium dodecyl sulphate.
1351572-3887/06/0200-0135/0 � 2006 Springer Science+Business Media, Inc.
The Protein Journal, Vol. 25, No. 2, February 2006 (� 2006)DOI: 10.1007/s10930-006-0005-5

from each other by their location within cells, andin their substrate specificity and sensitivity to activa-tors or inhibitors. None of these enzymes needsa metal ion for catalytic activity (Fujimoto et al.,1993).
High molecular weight acid phosphatases arepresent as two molecular species: a microsomal asso-ciated enzyme of about 200 kDa and a 80–120 kDaacid phosphatase of lysosomal origin (Panara et al.,1992). Both acid phosphatases are inhibited by fluo-ride and tartrate, are resistant to formaldehyde andreagents with sulphydryl groups and have an essen-tial histidyl residue at the active site (Hollander,1971; Van Etten and McTigue, 1977).
The low molecular weight acid phosphatases(10–30 kDa) are not particle associated enzymes butseem to be of cytosolic origin (Baxter and Suelter,1984; De Araujo et al., 1976). These low molecularweight enzymes are found to be inhibited by SH-blocking agents and formaldehyde but unaffectedby either fluoride or tartrate (Igarashi andHollander, 1968). The substrate specificity of lowmolecular weight acid phosphatases is morerestricted than that of high molecular weight acidphosphatases in that the former efficiently hydrolyseonly arylphosphates and flavin mononucleotide(FMN), while high molecular weight acid phospha-tases catalyse the hydrolysis of most phosphateesters (De Araujo et al., 1976). Recently a phosp-hotyrosine protein phosphatase activity for the lowmolecular weight acid phosphatase enzyme wasreported (Chernoff and Li, 1985).
The low molecular weight acid phosphatasesisolated from human liver (Taga and Van Etten,1982) human placenta (Waheed et al., 1988) bovineliver (Lawrence and Van Etten, 1981; Ramponiet al., 1989) and bovine heart (Zhang and VanEtten, 1990) showed hydrolytic activity againstphosphotyrosine IgG/phosphotyrosyl casein or tyro-sine autophosphorylated epidermal growth factorreceptors. These low molecular weight acid phos-phatases are now recognized to be active phosp-hotyrosine protein phosphatases (PTPases).
Acid phosphatases with intermediate molecularweights (Mr 35–50 kDa) are reported in rat liver(Murakami et al., 1986), sea urchin egg and embryo(Yokota and Nakano, 1984), Xenopus laevis (Fil-burn, 1973) and frog liver (Panara et al., 1989).These intermediate molecular weight enzymes showdifferent properties. A study of the relationshipbetween these enzymes and the other acid phospha-tase classes has not been reported.
The present work describe purification of an18 kDa acid phosphatase enzyme from chickenheart possessing phosphotyrosine protein phospha-tase and phosphotransferase activities.
2. EXPERIMENTAL PROCEDURES
2.1. Materials
Chicken hearts were obtained from a local poul-try shop. Para-nitrophenyl phosphate, phenyl phos-phate, bovine serum albumin, SDS molecular weightmarkers, SP-Sephadex C-50 and Sephadex G-75were purchased from Sigma Chemical Co.; Thematerials for SDS-polyacrylamide gel electrophoresiswere from Bio-Rad and Acros Chemical Co.; p-ami-no benzylphosphonic acid-agarose gel was obtainedfrom Pierce Chemical Co.; All other reagents were ofthe highest purity commercially available.
2.2. Enzyme Assays
Acid phosphatase activity was determined at37�C in 0.1 M acetate buffer pH 5.5 containing1 mM EDTA in the presence of 4 mM para-nitro-phenyl phosphate as previously reported by Ramp-oni et al. (1989). One unit of activity is defined asthe amount of enzyme that is required to produce 1lmol of para-nitrophenol product from its substrateper min at 37�C ( e405 ¼ 1:8� 104M)1 cm)1). Spe-cific activity is defined as number of enzyme unitsper milligram protein.
The activity over pH range of 2.9–9.0, the tem-perature optima measured at 5�C intervals from 25to 80�C and the effect of metal ions and modifierswere determined (Panara, 1985, 1986).
Km values were determined graphically(Lineweaver-Burk plot) with 6 substrate concentra-tions ranging from 0.25 to 16 km in the absence andpresence of ethylene glycol or inhibitors. The inhibi-tion constants were calculated at fixed concentra-tions of orthophosphate (0.5, 1.0 and 2.5 mM),molybdate (0.02, 0.05 mM) and vanadate (0.05 mM)and varied concentrations of para-nitrophenyl phos-phate (Taga and Van Etten, 1982).
2.3. Phosphotransferase Activity
The transphosphorylation reaction mixture(500 ll) containing 4 mM para-nitrophenyl
136 Naz, Saeed, and Saeed

phosphate, alcohol (1 M or 2 M), 0.1 M acetatebuffer pH 5.5 and catalytic amount of enzyme (25ll) was incubated at 37�C for 5 min. For determi-nation of para-nitrophenol, the reaction was stop-ped by addition of 1.5 ml of 0.1 N KOH and theabsorbance at 405 nm was measured. The amount ofpara-nitrophenol was determined from the absor-bance using para-nitrophenol standard curve. Foractivities with a phenylphosphate substrate, theamount of phenol produced was measured spectro-photometrically at 287 nm using e ¼ 2560M)1 cm)1
(Cirri et al., 1993). For inorganic phosphate determi-nation, the reaction was quenched by addition of0.2 ml of 10% trichloroacetic acid (TCA) and themolybdate colour reaction was performed accordingto method of Black and Jones (1983). The amountof inorganic phosphate produced was calculatedfrom the standard curve using KH2PO4 as a stan-dard. The amount of phosphate transferred to thealcohol was the difference between the amount ofpara-nitrophenol and inorganic phosphate produced.
2.4. Substrate Specificity
Substrate specificity studies were carried out bydetermining the liberated inorganic phosphate asdescribed above. All substrates were used at 4 mMconcentrations.
2.5. Protein Determination
Protein concentration was determined by theBiuret method. For column effluents, the relativeprotein concentration was estimated from the absor-bance at 280 nm.
2.6. Electrophoresis
SDS-polyacrylamide gel electrophoresis wascarried out by the method of Laemmli (1970).
2.7. Enzyme Purification
All procedures were carried out at 4�C. Acidphosphatase from chicken heart was purified as fol-lows according to a procedure described by Manaoet al., 1992.
Step-1. Fresh chicken hearts were washed withcold 10 mM Tris–HCl buffer pH 7.4 containing0.25 M sucrose and 1 mM EDTA and were cut into
1–2 cm pieces. Then, they were homogenized in aWaring blender with 3 volumes of 0.3 M sodiumacetate buffer pH 5.0 containing 1 mM EDTA,1 mM b-mercaptoethanol and 0.1 mM PMSF untila smooth suspension was obtained. The homoge-nate was stirred for 2–3 h at 4�C to achieve com-plete extraction. The homogenate was centrifuged at2740�g in Beckman Centrifuge J2-21 (using rotorJA-14). The supernatant was collected and the pel-let was discarded.
Step-2. Solid ammonium sulphate (176 g/l) wasadded to the supernatant to give 30% saturation.After stirring gently to dissolve all the ammoniumsulphate, it was centrifuged at 5000 rpm (2740�g)for 30 min. The supernatant was brought to 60%saturation with solid ammonium sulphate (198 g/l)with gradual additions and constant stirring. Theresulting mixture was allowed to stir for 1–2 h andthen centrifuged at 2740�g for 1 h. The precipitatewas dissolved in about 300 ml of 0.01 M acetatebuffer pH 5.1 containing 1 mM EDTA, 1 mMb-mercaptoethanol and 0.1 mM PMSF. The suspen-sion was stirred for 1–2 h to extract the enzyme,and again centrifuged at 10,000�g for 1 h. Theclear supernatant was dialysed against 10 l of samebuffer overnight with 2–3 replacements of fresh buf-fer. The dialysate was centrifuged at 10,000�g toobtain a clear supernatant.
Step-3. The clear supernatant solution was ap-plied to a SP-Sephadex C-50 column (2.8�36 cm),previously equilibrated with the dialysing buffer.The column was washed with the same buffer untilthe absorbance of the eluate at 280 nm was lessthan 0.08. The bound acid phosphatase activity waseluted by 0.3 M phosphate buffer pH 5.5 containing1 mM EDTA, 0.1 mM PMSF and 1 mM b-mercap-toethanol (single step). The fractions containingactivity were pooled together and the enzyme wasprecipitated by adding ammonium sulphate to 70%saturation. The precipitate was collected by centri-fugation at 10,000�g for 1 h and dissolved in15 m1 of 0.01 M acetate buffer pH 5.1 containing1 mM EDTA, 0.1 mM PMSF and 1 mM b-mercap-toethanol.
Step-4. A 15 ml sample (in two portions) wasapplied to a Sephadex G-75 column (2.6�70 cm)previously equilibrated with 0.01 M acetate bufferpH 5.0 containing the same additives and 0.1 MNaCl. The sample was eluted with same buffer andthe active fractions were pooled (60 ml).
Step-5. The enzyme after gel filtration was dial-ysed overnight against 1 l of 0.1 M citrate buffer
137Chicken Heart Possesses Phosphotransferase Activity

pH 6.5 containing 1 mM EDTA and 1 mM DTT.The sample was applied to a column (1�8 cm) ofpara-aminobenzyl phosphonic acid-agarose affinitygel previously equilibrated with same buffer. Thecolumn was washed extensively with the buffer toeliminate unbound proteins. During the washing,contaminant proteins were eluted together withsome of the acid phosphatase activity (PeakI, prob-ably isoform-1 or IF-1). The pure enzyme (PeakII)corresponding to IF-2 (Ramponi and Stefani, 1997)was eluted by a 0–0.1 M sodium phosphate lineargradient in the same buffer (total volume of 60 ml).The active fractions were pooled separately andconcentrated by Amicon ultrafiltration with YM3membrane.
3. RESULTS AND DISCUSSION
The major purification steps for purificationof low molecular weight acid phosphatase fromchicken heart to homogeneity are summarized inTable 1. Elution profiles for ion-exchange chroma-tography, gel filtration and affinity chromatographyare shown in Fig. 1(a–c). Para-nitrophenyl phos-phate was used as substrate to monitor the enzymeactivity throughout the purification procedures.About 800 fold purification was achieved with anoverall recovery of 1% and a specific activity of 42 U/mg/min. at pH 5.5 and 37�C. The SDS-polyacryl-amide gel-electrophoresis (12%) showed single bandat 18 kDa with Coomassive blue staining (Fig. 2).
Chicken heart low molecular weight acid phos-phatase activity showed maximum activity over a pHof 4.2–6.2. The optimal temperature for activity was55–60�C. The Km for para-nitrophenyl phosphate
was 0.25 mM at pH 5.5 with a Vmax of 36.35 lmol ofsubstrate hydrolysed/min/mg at 37�C while the Km
for phenyl phosphate was 0.3 mM and Vmax was 38lmol/min/mg. Table 2 shows the hydrolysis rates ofvarious phosphate monoesters by the chicken heartenzyme. Only para-nitrophenyl phosphate, phenyl-phosphate and flavin mononucleotide were efficientlyhydrolysed. Other substrates, including a and bglycerophosphate, glucose-1-phosphate, glucose-6-phosphate, AMP, UMP, phospho-serine and phos-pho-threonine were not hydrolysed at appreciablerates. Thus, it showed a limited substrate specificitycompared with high molecular weight acid phospha-tases (Hollander, 1971; Rehkop and Van Etten,1975; Taga and Van Etten, 1982). This enzyme issimilar to low molecular weight PTPases isolatedfrom bovine liver (Ramponi et al., 1989), bovineheart (Zhang and Van Etten, 1990) bovine brain(Saeed et al., 1990) human placenta (Waheed et al.,1988) rat liver (Manao et al., 1992) and chicken liver(Saeed, 1999) that efficiently catalyse the hydrolysisof arylphosphates (Heinrikson, 1969; Lawrence andVan Etten, 1981) and acyl phosphates such as ben-zoyl phosphate (Ramponi et al., 1980; Ramponi,1994). On the other hand, aliphatic phosphates werepoor substrates with the exception of FMN. Tagaand Van Etten (1982) also noted that FMN is hydro-lysed by human liver enzyme with a Vmax three timesgreater than that of para-nitrophenyl phosphate.
As reported for mammalian tissues (Sainiand Van Etten, 1978; Taga and Van Etten, 1982);fluoride and tartrate are potent inhibitors of highmolecular weight acid phosphatases while theseenzymes are insensitive to formaldehyde. Chickenheart enzyme was found to be insensitive to fluo-ride and tartrate, but was strongly deactivated by
Table 1. Purification Scheme for Acid Phosphatase from Chicken Heart
Volume
(ml)
Total
activity (U)
Total
proteinb (mg)
Specific
activity (U/mg)
Purification
fold
Recovery
%
Extracta 2250 742.5 13,950 0.054 1 100
30%(NH4)2SO4 2350 681.5 9870 0.069 1.27 91.7
60%(NH4)2SO4 300 243 4080 0.059 1.0 32.7
Dialysis & centrifugation 500 210 3400 0.062 1.15 28.2
SP-Sephadex C-50 15 43.5 145.5 0.298 5.52 5.8
Sephadex G-75 60 40.38 102.0 0.398 7.33 5.4
Affinity chromatography PeakI
24 7.32 0.72 10.16 188 0.98
PeakII
9 6.12 0.144 42.05 787 0.82
a Starting from 1 kg of chicken heart.b Protein concentration was determined by Biuret method.
138 Naz, Saeed, and Saeed
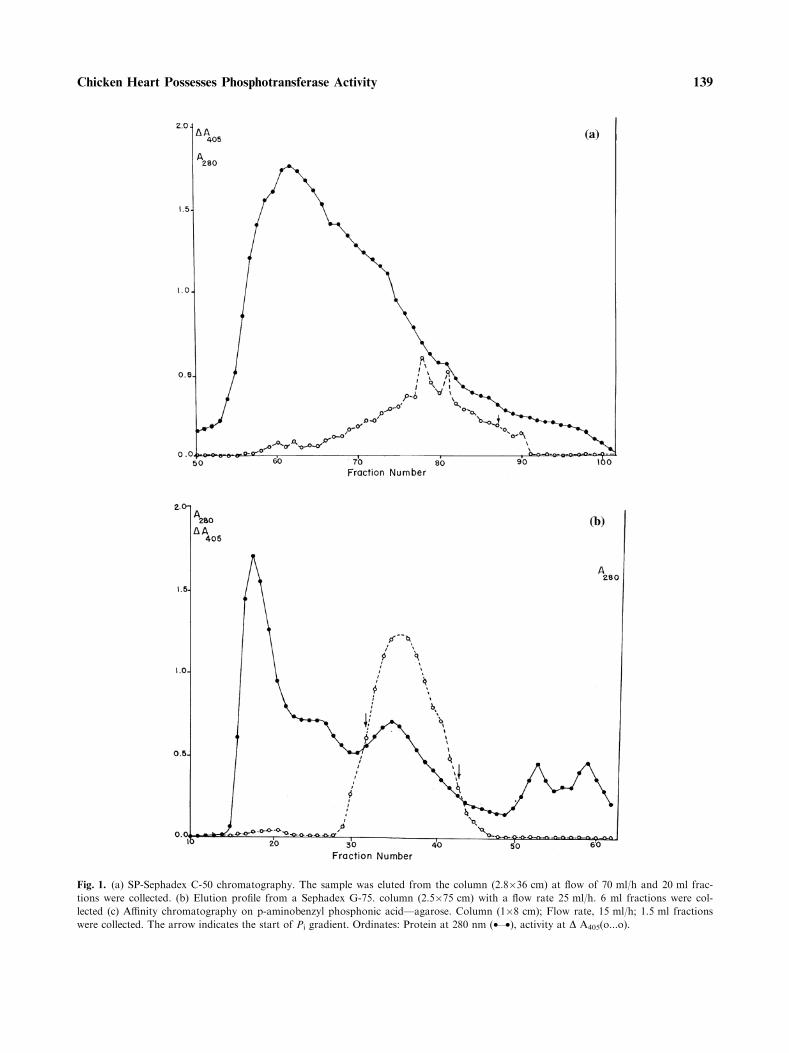
Fig. 1. (a) SP-Sephadex C-50 chromatography. The sample was eluted from the column (2.8�36 cm) at flow of 70 ml/h and 20 ml frac-
tions were collected. (b) Elution profile from a Sephadex G-75. column (2.5�75 cm) with a flow rate 25 ml/h. 6 ml fractions were col-
lected (c) Affinity chromatography on p-aminobenzyl phosphonic acid—agarose. Column (1�8 cm); Flow rate, 15 ml/h; 1.5 ml fractions
were collected. The arrow indicates the start of Pi gradient. Ordinates: Protein at 280 nm (•—•), activity at D A405(o...o).
139Chicken Heart Possesses Phosphotransferase Activity

Fig. 1. Continued
Fig. 2. SDS-Polyacrylamide gel electrophoresis of acid phospha-
tase isoform 2 from chicken heart. Lane 1—The standard pro-
teins used from top to bottom were bovine plasma albumin
(66,000 Da), egg albumin (45,000 Da), pepsin (34,700 Da), b-lac-toglobulin (18,400 Da) lysozyme (14,300 Da). Lane 2—Acid
phosphatase from chicken heart. Lane 3—Standard proteins as
above.
Table 2. Substrate Specificity of Chicken heart Acid Phosphatase
Substrate Activity %
p-Nitrophenyl phosphate 100
Phenyl phosphate 75
a-Glycero phosphate 4.3
b-Glycero phosphate 6.5
Flavin mononucleotide 108
Glucose-1-phosphate 7.8
Glucose-6-phosphate 1.5
Phosphoserine 3
Phosphothreonine 6.4
AMP 2.8
Diphenyl phosphate 1
Phosphoenol phosphate 8.2
UMP 3
Assays were carried out at 37�C and pH 5.5. The assay mixture
contained 0.1 M acetate buffer, 4 mM of substrate and the enzyme
in a total volume of 0.5 m1. After incubation for an appropriate
period, the reaction was quenched by addition of 0.2 m1 of 10%
TCA and the amount of inorganic phosphate released was deter-
mined by Black & Jones as described earlier. The enzyme activity
was expressed as a percent of that of the same enzyme towards
p-nitrophenyl phosphate as 100.
140 Naz, Saeed, and Saeed

formaldehyde, an activity that differentiates lowand high molecular weight acid phosphatases(Table 3). Phosphate, orthovanadate and molyb-date strongly inhibited the enzyme and theseinhibitors were purely competitive (Fig. 3). Phos-phate was a less powerful inhibitor than vanadateor molybdate. Their Ki values were estimated tobe 1.28 mM, 14 lM and 19 lM, respectively.Millimolar concentrations of phosphate were nee-ded to effectively inhibit the enzyme, whereas onlymicromolar concentrations of vanadate or molyb-date were required to abolish the activity of theenzyme. The results are similar to those reportedfor other acid phosphatases (Chernoff and Li,1985; Waheed et al., 1988; Zhang and Van Etten,1990). Phosphotyrosin protein phosphatases werestrongly inhibited by orthovanadate and molyb-date in the micromolar range. Moreover theseinhibitors were more effective for high molecularweight acid phosphatases than for low molecularweight enzymes (Khan et al., 1997; Panara, 1985).
Triton X-100 and EDTA had little or no effecton the activity. The enzyme did not require divalentmetal ions for activity. Assays carried out with 5 mMion concentrations indicated that Ag+, Hg++,Ni++, Co++ and Zn++ were potent inhibitors ofthe enzyme whereas other cations such as Mg++,Ca++, Mn++ and Cd++ showed little effect.
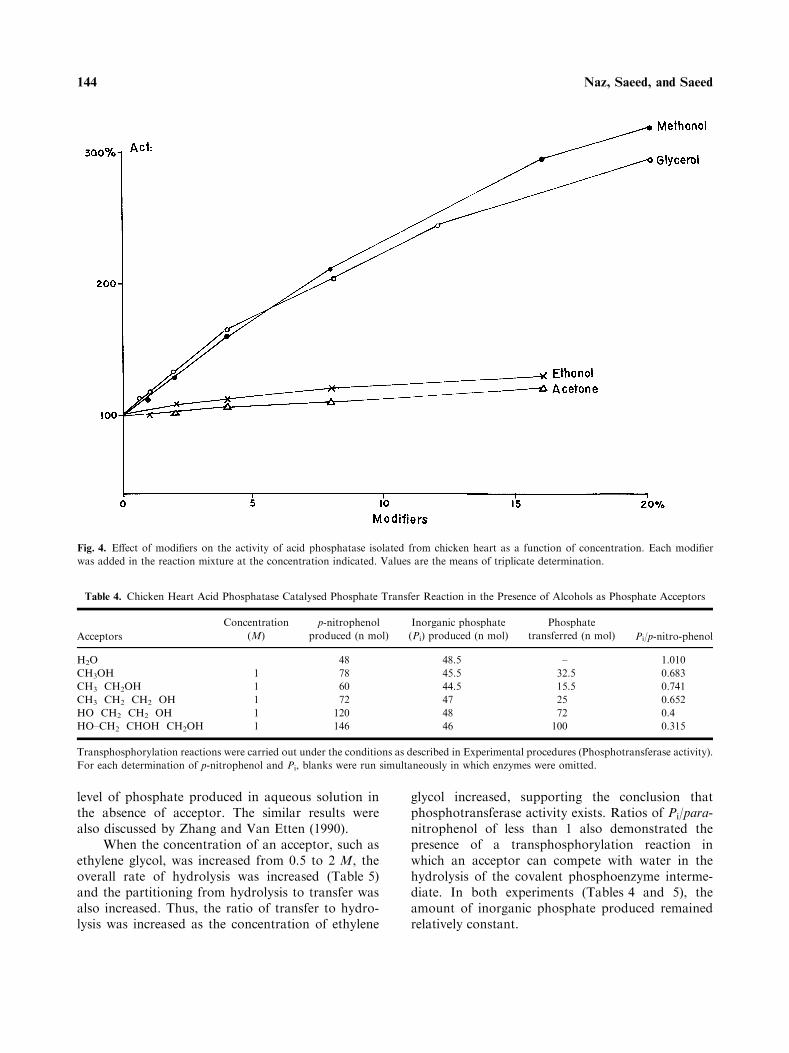
The enzyme activity was stimulated in the pres-ence of acetone, ethanol, methanol and glycerol(Fig. 4). Glycerol and methanol, at 8% concentra-tions, caused two-fold increases in the enzyme activ-ity, while 20% concentrations caused three-foldincreases. This behaviour is typical of phospho-transferase activity.
Several authors have demonstrated that lowmolecular weight PTPases possess phosphotransfer-ase activity in addition to hydrolytic activity (Baldi-jao et al., 1975; Hollander, 1971; Ostrowski andBarnard, 1973; Tanizaki et al., 1977; Zhang and VanEtten, 1990, 1991). The ability to catalyse phosphatetransfer and partitioning of an intermediate betweencompeting acceptors has been useful method fordemonstrating the formation of phospho-enzymeintermediate and for identifying the rate limiting stepin the reaction mechanism specifically whether it isthe phosphorylation of the enzyme or the dephos-phorylation of a phosphoenzyme intermediate.
According to Zhang and Van Etten (1991) if therate limiting step is the formation of a phosphoen-zyme intermediate, then the acceptor reacts with thephosphoenzyme, it will not increase the overall rateof substrate disappearance. Thus, the addition of aphosphate acceptor would not change the rate offormation of para-nitrophenol but would decreasethe rate of formation of inorganic phosphate becausethe new pathway competes to lower the level of E–P.In other words, the presence of the acceptor onlychanges the product distribution but not the overallrate of destruction of the substrate (see reactionscheme 1) where E–P is a phosphoenzyme intermedi-ate, A is an acceptor and pNP is para-nitrophenol.
On the other hand, if the rate limiting step ishydrolysis of the phosphoenzyme intermediate, thenthe presence of the acceptor will increase the rate of
Scheme 1.
Table 3. Effect of Inhibitors on the Activity of Chicken Heart
Acid Phosphatase
Inhibitor Concentration Activity %
None – 100
NaF 10 mM 102
Tartrate 10 mM 98
Na3PO4 5 mM 40
Molybdate 0.2 mM 92.5
0.4 mM 68
Vanadate 0.1 mM 20
ZnCl2 10 mM 1.6
Formaldehyde 0.5% 14
EDTA 10 mM 86
Triton X-100 1% 83.6
The activity was determined by incubating the enzyme in a final
mixture volume of 0.5 m1 containing 4 mM p-nitrophenyl phos-
phate, 0.1 M acetate buffer pH 5.5 in the presence and absence of
the indicated additions. After a 5 min incubation at 37�C, the
reaction was stopped by the addition of 1.5 m1 of 0.1 N KOH and
the absorbance at 405 nm was measured. The control activity
without inhibitor was taken as 100% and the other activities were
expressed as a percentage of this activity. Values are the means of
triplicate determination.
141Chicken Heart Possesses Phosphotransferase Activity

the break down of the intermediate and henceincrease the overall reaction rate. Thus, addition of aphosphate acceptor would increase the rate of forma-tion of para-nitrophenol in response to the new path-way, the level of E–P would not fall, and the rate offormation of inorganic phosphate would remainunchanged. Zhang and Van Etten (1991) demon-strated that the latter is the case for hydrolysis ofpara-nitrophenyl phosphate catalysed by bovine
heart low molecular weight PTPase in the presenceof ethylene glycol. Similarly, Cirri et al. (1993) alsofound the same results with benzoyl phosphate assubstrate of MBP-PTPase (maltose-binding protein-PTPase fusion protein) and glycerol as the acceptormolecule.
In our case, phosphotransferase activitywas shown by the phosphotransfer experimentsusing a variety of alcohols as phosphate acceptors
Fig. 3. Competitive inhibition of chicken heart acid phosphatase. Lineweaver-Burk plots of 1/v versus 1/S. (A) In the absence (a) or pres-
ence of 0.5 mM (b), 1 mM (c), and 2.5 mM (d) Na3PO4. (B) In the absence (a) or presence of 0.05 mM (b) Na3VO4. (C) In the absence
(a) or presence of 0.02 mM (b) and 0.05 mM (c) ammonium molybdate.
142 Naz, Saeed, and Saeed

(Table 4). The polyfunctional alcohols (ethylene gly-col and glycerol), were better phosphate acceptorsthan monofunctional alcohols. The presence of aphosphate acceptor in the incubation mixture
increased the overall rate of substrate breakdown asdetermined by the amount of para-nitrophenol pro-duced. The level of inorganic phosphate that wasproduced remained unchanged compared with the
Fig. 3. Continued
143Chicken Heart Possesses Phosphotransferase Activity
level of phosphate produced in aqueous solution inthe absence of acceptor. The similar results werealso discussed by Zhang and Van Etten (1990).
When the concentration of an acceptor, such asethylene glycol, was increased from 0.5 to 2 M, theoverall rate of hydrolysis was increased (Table 5)and the partitioning from hydrolysis to transfer wasalso increased. Thus, the ratio of transfer to hydro-lysis was increased as the concentration of ethylene
glycol increased, supporting the conclusion thatphosphotransferase activity exists. Ratios of Pi/para-nitrophenol of less than 1 also demonstrated thepresence of a transphosphorylation reaction inwhich an acceptor can compete with water in thehydrolysis of the covalent phosphoenzyme interme-diate. In both experiments (Tables 4 and 5), theamount of inorganic phosphate produced remainedrelatively constant.
Fig. 4. Effect of modifiers on the activity of acid phosphatase isolated from chicken heart as a function of concentration. Each modifier
was added in the reaction mixture at the concentration indicated. Values are the means of triplicate determination.
Table 4. Chicken Heart Acid Phosphatase Catalysed Phosphate Transfer Reaction in the Presence of Alcohols as Phosphate Acceptors
Acceptors
Concentration
(M)
p-nitrophenol
produced (n mol)
Inorganic phosphate
(Pi) produced (n mol)
Phosphate
transferred (n mol) Pi/p-nitro-phenol
H2O 48 48.5 – 1.010
CH3OH 1 78 45.5 32.5 0.683
CH3 Æ CH2OH 1 60 44.5 15.5 0.741
CH3 Æ CH2 Æ CH2 Æ OH 1 72 47 25 0.652
HO Æ CH2 Æ CH2 Æ OH 1 120 48 72 0.4
HO–CH2 Æ CHOH Æ CH2OH 1 146 46 100 0.315
Transphosphorylation reactions were carried out under the conditions as described in Experimental procedures (Phosphotransferase activity).
For each determination of p-nitrophenol and Pi, blanks were run simultaneously in which enzymes were omitted.
144 Naz, Saeed, and Saeed

These data suggest that the breakdown of thephosphoenzyme intermediate is the rate determiningstep since there is a constant level of phosphoen-zyme intermediate, E–P under the conditions usedin these experiments. The presence of a nucleophilesuch as a low molecular weight alcohol, which is abetter nucleophile than water, increases the break-down of E–P and thus the overall production ofpara-nitrophenol. Since alcohol concentrations werelow (0.5 or 2 M), the rate of hydrolysis was nearlythe same as that in the absence of an acceptor,resulting in a constant level of phosphate producedin each case.
Zhang and Van Etten (1991) determined Kcat
values for 21 phosphate monoesters, both arylphosphates and alkyl phosphates. The Kcat valuesfor aryl phosphate were almost identical (26.1 s)1 to34.3 s)1) whereas those for alkyl phosphates werevariable and generally lower (0.73 s)1 to 3.54 s)1).These results were consistent with the existence of acommon phosphoenzyme intermediate, with itsbreakdown being rate limiting. This conclusion wasconfirmed by a variety of experiments, includingpre-steady-state kinetics, 18O exchange and phos-phoenzyme trapping (Wo et al., 1992; Zhang andVan Etten, 1991). Our results for chicken heart acidphosphatase show Kcat values for para-nitrophenylphosphate and phenyl phosphate, 10.9 s)1 and11.0 s)1, respectively that are almost the same.
Similar results were also reported for both naturaland mutated MBP-PTPase fusion protein (Cirriet al., 1993), suggesting that these phosphate mono-esters are hydrolysed through the formation of acommon phosphoenzyme intermediate and that itsbreakdown is the common rate-determining step.
Furthermore, this conclusion is supported bythe partition experiments between differentsubstrate hydrolysis and transphosphorylation to anacceptor molecule that completes with water in thephosphoenzyme breakdown reaction. A commonintermediate that is formed from different substratesmust give the same ratio of products at a given con-centration of acceptor. In the presence of 1 M eth-ylene glycol, partitioning of the intermediatebetween competing acceptors, ethylene glycol andwater, was determined using two substrates, para-nitrophenyl phosphate and phenyl phosphate(Table 6). The constant ratio of 1.7 between phos-phate transferred to ethylene glycol and the phos-phate produced strongly supports the conclusionthat a common covalent intermediate exists.
Addition of an acceptor changed both the cata-lytic constant (Kcat) and Michaelis Menten constant(Km) (Table 7). These results are similar to resultsfor bovine heart enzyme and natural PTPase andrecombinant MBP-PTPase (Cirri et al., 1993; Zhangand Van Etten, 1991). The constant ratio of KCat/Km i.e. 4.3�104 found at different ethylene glycol
Table 5. Chicken Heart Acid Phosphatase Catalysed Phosphate Transfer Reaction in the Presence of Various Concentrations of Ethylene
Glycol
Concentration of
ethylene glycol (M)
p-nitrophenol
produced (n mol)
Phosphate (Pi) produced
(hydrolysis) (n mol)
Phosphate
transferred (n mol)
Ratio of transfer
to hydrolysis Pi/p-nitro phenol
0 48 47.5 0.5 0.010 0.989
0.5 86 42.5 43.5 1.023 0.494
1.0 110 44.0 66.0 1.500 0.400
2.0 136 43.75 92.25 2.108 0.321
As mentioned in Table 4.
Table 6. Partition Experiment: Relative Rates of Phenols and Pi Production by Chicken Heart Acid Phosphatase from Para-Nitrophenyl
Phosphate and Phenyl Phosphate in the Presence of 1 M Ethylene Glycol
Substrate
Phenols
produced/min (n mol)
Phosophate
produced/min (n mol)
Phosphate
transferred/min (n mol)
Ratio of transfer
to hydrolysis
Para-nitrophenyl phosphate 18.8(7.12) 6.9(7.08) 11.9 1.724
Phenyl phosphate 3.24(1.19) 1.22(1.15) 2.02 1.655
Assays were carried out at 37�C and pH 5.5. The reaction mixture contained 0.1 M acetate buffer, 4 mM substrate, 1 M ethylene glycol and
25 ll of enzyme in a final volume of 0.5 m1. After incubation for 5 min., the reactions were terminated by the addition of either KOH or
TCA (see Experimental procedures). The amount of p-nitrophenol and phenol produced/min. were calculated using molar extinction coef-
ficient of 1.8�104 and 2560 M)1 Cm)1, respectively. The amount of Pi produced/min. were estimated from standard curve of KH2PO4.
Readings within parenthesis indicate the production of phenols and Pi in the absence of 1 Methylene glycol.
145Chicken Heart Possesses Phosphotransferase Activity

concentrations excluded a mechanism in which eth-ylene glycol directly attacks the E-pNPP, Michaelis-complex (Fersht, 1985; Zhang and Van Etten,1991). From this it can also be concluded that ratedetermining step is the hydrolysis of the phosphoen-zyme intermediate.
Thus, the phosphotransferase activity, overallincrease in the rate of hydrolysis in the presence ofacceptors, and the constant product ratio in thepartition experiments all suggest the existence of acovalent phosphoenzyme intermediate and that therate limiting step in the hydrolytic pathway is thedephosphorylation of this intermediate.
ACKNOWLEDGMENT
This work was supported in part by grant fromGomal University Research Project No.12/GU.
REFERENCES
Buchwald, S. L., Saini, M. S., Knowles, J. R., and Van Etten, R. L.(1984). J. Biol. Chem. 259: 2208–2213.
Baxter, J. H., and Suelter, C. H. (1984). Arch. Biochem. Biophys.228: 397–406.
Black, M. J., and Jones, M. E. (1983). Anal. Biochem. 135: 233–238.Baldijao, C. E. M., Guija, E., Bittencourt, H. M. S., and
Chaimovich, H. (1975). Biochem. Biophys. Acta 391: 316–325.Chernoff, J., and Li, H. C. (1985). Arch. Biochem. Biophys. 240:
135–145.Cirri, P., Chiarugi, P., Camici, G., Manao, G., Raugei, G.,
Cappugi, G., and Ramponi, G. (1993). Eur. J. Biochem. 214:647–657.
De Araujo, P. S., Mies, V., and Miranda, O. (1976). Biochim.Biophys. Acta 456: 121–130.
Fujimoto, O., Gotoh, H., Ohbayashi, T., Yazaki, H., and Ohara,A. (1993). Biol. Pharm. Bull. 16: 745–750.
Filburn, C. R. (1973) Arch. Biochem. Biophys. 159: 683–693.Fersht, A. (1985) Enzyme Structure and Mechanism New York:
W. H. Freeman and Company.Hollander, P. V. (1971). Acid phosphatases, The Enzymes. In:
Boyer, P. D. (ed.), pp. 449–498. Vol. 4, New York: AcademicPress.
Heinrikson, R. L. (1969) J. Biol. Chem. 244: 299–307.Igarashi, M., and Hollander, P. V. (1968). J. Biol. Chem. 243:
6084–6089.Khan, A. R., Naz, R., Hassan, M., and Saeed, A. (1997). J. Chem.
Soc. Pak. 19: 60–69.
Lawrence, G. L., and Van Etten, R. L. (1981). Arch. Biochem.Biophys. 206: 122–131.
Laemmli, U. K. (1970) Nature 227: 680–685.Murakami, K., Fujimoto, S., and Ohara, A. (1986). Chem. Pharm.
Bull. 34: 3320–3327.Manao, G., Pazzagli, L., Cirri, P., Casselli, A., Camici, G.,
Cappugi, G., Saeed, A., and Ramponi, G. (1992). J. Prot.Chem. 11: 333–345.
Ostrowsky, W. S., and Kuciel, R. (1994). Clin. Chim. Acta 226:
121–129.Ostrowski, W., and Barnard, E. A. (1973). Biochemistry 12:
3893–3898.Panara, F. (1985) Int. J. Biochem. 17: 1213–1217.Panara, F. (1986) Biochem. J. 235: 265–268.Panara, F., Angiolillo, A., and Pascolini, R. (1989). Comp.
Biochem. Physiol. 93B: 877–882.Panara, F., Angiolillo, A., Fagotti, A., Di Rosa, I., Simoncelli, F.,
and Pascolini, R. (1992). Int. J. Biochem. 24: 1619–1623.Panara, F., Angiolillo, A., Fagotti, A., Di Rosa, I., and Pascolini,
R. (1993). Trends Comp. Biochem. Physiol. 1: 675–692.Panara, F., Guiderdone, S., Pellegrini, M., Zuccaccia, C., and
Contenti, S. (1998). Cytobios 94: 111–119.Ramponi, G., Cappugi, G., Manao, G., and Camici, G. (1980).
26th Natl. Meet. Ital. Biochem. Soc. p. 167.Ramponi, G., Manao, G., Camici, G., Cappugi, G., Ruggiero, M.,
and Bottaro, B. P. (1989). FEBS Lett. 250: 469–473.Ramponi, G. (1994) Adv. Prot. Phosphatases. 8: 1–25.Ramponi, G., and Stefani, M. (1997). Int. J. Biochem. Cell Biol. 29:
279–292.Rehkop, D. M., and Van Etten, R. L. (1975). Hoppe Seyler’s Z.
physiol. Chem. 356: 1775–1782.Saeed, A., Tremori, E., Manao, G., Camici, G., Cappugi, G., and
Ramponi, G. (1990). Physiol. Chem. Phys. Med. NMR 22: 81–94.
Saeed, A. (1999) J. Chem. Soc. Pak. 21: 311–320.Saini, M. S., and Van Etten, R. L. (1978). Arch. Biochem. Biophys.
191: 613–618.Taga, E. M., and Van Etten, R. L. (1982). Arch. Biochem. Biophys.
214: 505–515.Tanizaki, M. M., Bittencourt, H. M. S., and Chaimovich, H.
(1977). Biochim. Biophys. Acta 485: 116–123.Vincent, J. B., Crowder, M. W., and Averill, B. A. (1992). Trends
Biochem. Sci. 17: 105–110.Van Etten, R. L., and Mc Tigue, J. J. (1977). Biochim. Biophys.
Acta. 484: 386–397.Waheed, A., Laidler, P. M., Wo, Y. Y. P., and Van Etten, R. L.
(1988). Biochemistry 27: 4265–4273.Wo, Y. Y. P., Zhou, M. M., Stevis, P., Davis, J. P., Zhang, Z.
Y., and Van Etten, R. L. (1992). Biochemistry 31:
1712–1721.Yokota, Y., and Nakano, E. (1984). Comp. Biochem. Physiol. 79B:
17–22.Zhang, Z. Y., and Van Etten, R. L. (1990). Arch. Biochem.
Biophys. 282: 39–49.Zhang, Z. Y., and Van Etten, R. L. (1991). J. Biol. Chem. 266:
1516–1525.
Table 7. Kcat and Km Values for Para-Nitrophenyl Phosphate (pNPP) in the Presence of Ethylene Glycol
Ethylene glycol Km mM Vmaxlmol min)1 mg)1 Kcat s)1 Kcat/Km M)1 s)1
0 0.25 36.35 10.9 4.3�1040.5 0.4 54.65 16.4 4.1�1041.0 0.55 76.51 22.97 4.1�1042.0 0.9 127.27 38.28 4.2�104
Vmax expressed as lmol of p-nitrophenol liberated/min/mg of protein.
146 Naz, Saeed, and Saeed




















