
Primers for a PCR-Based Approach to Mitochondrial Genome Sequencing in Birds and Other Vertebrates
Upload
independentCategory
view
5download
0
Human DNA Polymerase l Possesses TerminalDeoxyribonucleotidyl Transferase Activity And CanElongate RNA Primers: Implications forNovel Functions
Kristijan Ramadan1†, Giovanni Maga2†, Igor V. Shevelev1,3†Giuseppe Villani4, Luis Blanco5 and Ulrich Hubscher1*
1Institute of VeterinaryBiochemistry and MolecularBiology, University ofZurich-IrchelWinterthurerstrasse 190CH-8057 Zurich, Switzerland
2Istituto di Genetica MolecolareIGM-CNR, Via Abbiategrasso207, I-27100 Pavia, Italy
3Department of Molecular andRadiation BiophysicsPetersburg Nuclear PhysicsInstitute, Leningrad districtGatchina 188300, RussianFederation
4Institut de Pharmacologie et deBiologie Structurale, CentreNational de la RechercheScientifique, 205 route deNarbonne, 31077 ToulouseCedex, France
5Centro de Biologia MolecularSevero Ochoa, (CSIC-UAM)Universidad AutonomaMadrid 28049, Spain
DNA polymerase l is a novel enzyme of the family X of DNA poly-merases. The recent demonstration of an intrinsic 50-deoxyribose-50-phos-phate lyase activity, a template/primer dependent polymerase activity, adistributive manner of DNA synthesis and sequence similarity to DNApolymerase b suggested a novel b-like enzyme. All these properties sup-port a role of DNA polymerase l in base excision repair. On the otherhand, the biochemical properties of the polymerisation activity of DNApolymerase l are still largely unknown. Here we give evidence thathuman DNA polymerase l has an intrinsic terminal deoxyribonucleotidyltransferase activity that preferentially adds pyrimidines onto 30OH endsof DNA oligonucleotides. Furthermore, human DNA polymerase l effi-ciently elongates an RNA primer hybridized to a DNA template. Thesetwo novel properties of human DNA polymerase l might suggestadditional roles for this enzyme in DNA replication and repair processes.
q 2003 Elsevier Science Ltd. All rights reserved
Keywords: DNA polymerase l; terminal deoxyribonucleotidyl transferaseactivity; RNA primer elongation; human; DNA replication and repair*Corresponding author
Introduction
DNA polymerases (pols) play an essential role inthe maintenance of genome integrity during DNA
replication, DNA repair, DNA recombination, andalso in checkpoint functions in response to DNAdamage. Recently, various novel pols have beenidentified.1 All known pols share both functionaland structural similarities. However, each of theseenzymes possesses unique features that enable itto cope with different DNA transactions where itis involved in vivo. Noteworthy, a large and evengrowing number of enzymes has been recentlyfound to be specialized in different types of DNAsynthesis such as translesion synthesis and lowfidelity synthesis on normal undamaged DNA.
0022-2836/03/$ - see front matter q 2003 Elsevier Science Ltd. All rights reserved
† These authors contributed equally to this work.
E-mail address of the corresponding author:[email protected]
Abbreviations used: pol, DNA polymerase; TdT,terminal deoxyribonucleotidyl transferase; dRP-lyase, 50-deoxyribose-50-phosphate lyase; BER, base excisionrepair; NER, nucleotide excision repair; NHEJ, non-homologous end-joining.
doi:10.1016/S0022-2836(03)00265-1 J. Mol. Biol. (2003) 328, 63–72

One of these novel enzymes is pol l, a memberof the family X of pols 2,3 which shares significantsimilarity with other members of this family suchas pol b, terminal deoxyribonucleotidyl transferase(TdT), and pol m.4 Pol l is a nuclear enzyme thathas been identified in mammals as well as indifferent higher eukaryotes including vertebratesand plants such as Arabidopsis thaliana5. In additionto its 32% amino acid identity with pol b, sequencecomparison and three-dimensional structuremodelling predict that pol l contains in all thefour pol b such as structural subdomains, calledfingers, palm, thumb, and 8 kDa 50-deoxyribose-50-phosphate lyase (dRP-lyase) domain. However,unlike pol b, pol l has a BRCT motif and aproline/serine rich region at its N terminus. Asimilar BRCT domain is also present in TdT andpol m and is believed to take part in protein–protein and/or protein–DNA interactions,whereas the proline/serine rich part is unique topol l. Northern blot analysis shows that murineand human pol l mRNAs are highly abundant intestis. Moreover, predominant expression ofmurine pol l in pachytene spermatocytes has ledto the hypothesis that it might play a role in DNArepair synthesis coupled to meiotic recombination.2
Since its discovery, pol l has been suggested toplay a role in DNA synthesis associated withDNA repair processes in the nucleus of mam-malian cells. In particular, its polymerase features,combined with the intrinsic dRP-lyase activity,make pol l an important enzyme for base excisionrepair (BER).6,7 Moreover, deletion of exons 5 to 7in the pol l-targeted allele, which resulted in theremoval of the polymerase and dRP-lyase catalyticdomains, has no influence on viability and fertilityof mice over several generations, thus arguingthat pol l is dispensable for mouse development.8
The sensitivity of pol l 2 /2 , 2/þ and þ/þembryonic stem cells to different DNA damagingagents such as UV-irradiation (for nucleotideexcision repair (NER)), X-ray irradiation (for non-homologous end-joining (NHEJ)), and methyl-methansulphonate for BER was not affected,suggesting that pol l is not essential for theserepair systems.9 Probably, some other pol(s) (forexample pol b, pol d and pol 1) compensate forthe deficiency of pol l in these repair processes.Recent data suggested a role of human pol l (hpoll) in translesion DNA synthesis over an abasicsite, in cooperation with the replication accessoryprotein PCNA.10 Thus, in vivo functions of pol lare still not understood. In addition, its biochemi-cal properties are still largely unknown. Recently,it was shown that hpol l is a template dependentpol, distributive on a template/primer but pro-cessive on a gapped DNA, and it does not containa 30 ! 50 proofreading exonuclease activity.7
Here, we describe two novel activities of hpol l:first, a terminal deoxyribonucleotidyl transferaseactivity with sequence preference for the pyrimi-dine nucleotides, and second, the capacity toelongate RNA primers hybridised to DNA tem-
plates. We discuss these two enzymatic propertiesof hpol l in view of its possible role in DNA repli-cation and double strand break (DSB) repair.
Results
Human DNA polymerase l has terminaldeoxyribonucleotidyl transferase activity
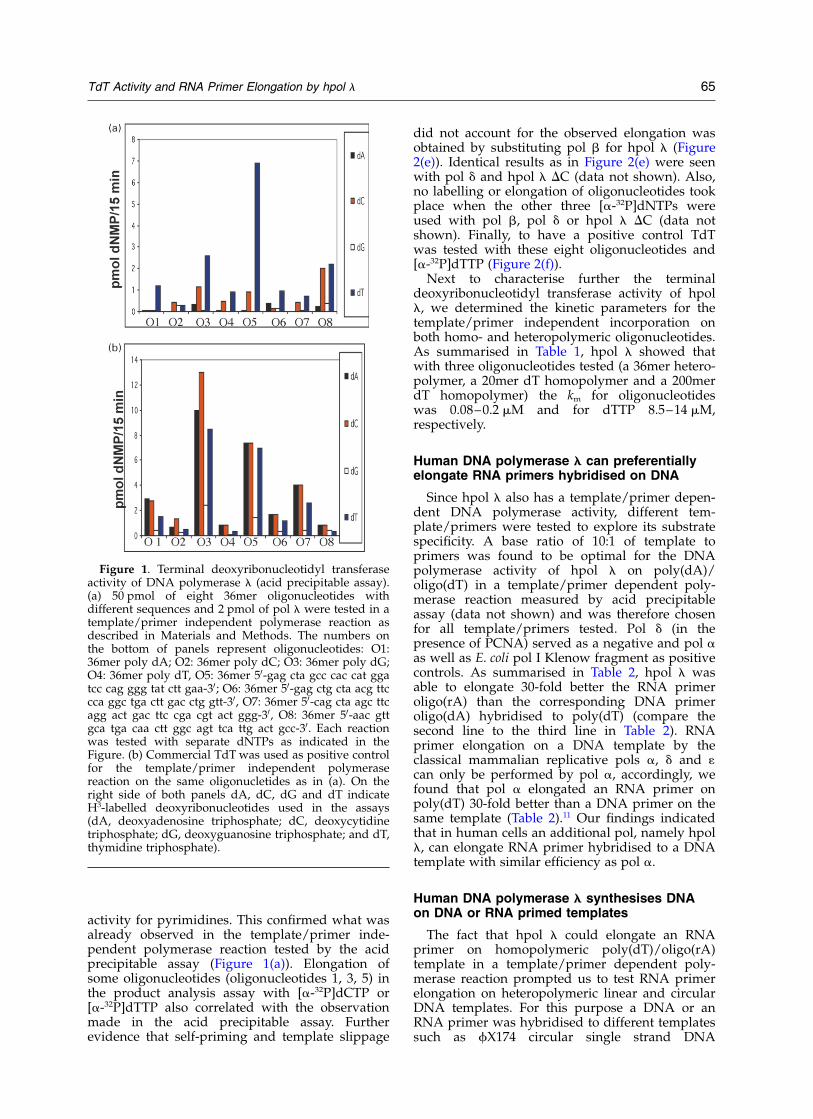
Terminal deoxyribonucleotidyl transferaseactivity of hpol l was tested using two methods:first, by precipitation with 10% trichloroacetic acidfollowed by radioactivity determination (acid pre-cipitable assay) and second, by product analysisby electrophoresis and visualisation by auto-radiography (for details see Materials andMethods). Figure 1 gives an example of a tem-plate/primer independent polymerase activitymeasured by acid precipitable assay with[3H]dNTPs and different DNA oligonucleotides(oligonucleotides). To avoid the possibility that theoligonucleotides might have been self-hybridisedeight different oligonucleotides of 36 bases weretested. As outlined in the legend to Figure 1, fourof these oligonucleotides are homopolymeric, andfour contain heteropolymeric sequences. All four[3H]dNTPs were tested separately. These experi-ments showed that hpol l was able to catalysepolymerisation with dTTP or dCTP to single-strand oligonucleotides even in the absence of aprimer (Figure 1(a)). The extent of incorporationwas different for different oligonucleotides (com-pare e.g. Figures 1 and 2). In contrast to TdT, usedas a positive control (Figure 1(b)) hpol l couldonly efficiently incorporate dCTP and dTTP. Noincorporation or very weak incorporation of dATPand dGTP was seen by this acid precipitableassay. With the negative controls, pol b, pol d andhpol l DC (truncated form of hpol l-; for detailssee Materials and Methods) no incorporation ofnucleotides on the oligonucleotides could beobserved (data not shown). From these experi-ments we can conclude that hpol l has a terminaldeoxyribonucleotidyl transferase activity with apreference for pyrimidines.
To investigate further the template/primer inde-pendent polymerase activity of hpol l, oligo-nucleotides mentioned above were tested in atemplate/primer independent polymerase reactionby product analysis assay where the incorporatednucleotides were visualised by autoradiographyafter gel electrophoresis (Figure 2). It can benoticed that on some of the oligonucleotides hpoll could add only one nucleotide to the 30 OH,thus only labelling it (Figure 2(a), lanes 2, 4, 6, 7, 8and Figure 2(b)), where on others hpol l couldincorporate additional nucleotides (Figure 2(a),lanes 1, 3 and 5), thus elongating the oligonucleo-tide. As shown in Figure 2(a)–(d), the level ofincorporation on the 30 OH with dCTP or dTTPrespect to dATP or dGTP, indicated preference ofhpol l terminal deoxyribonucleotidyl transferase
64 TdT Activity and RNA Primer Elongation by hpol l
activity for pyrimidines. This confirmed what wasalready observed in the template/primer inde-pendent polymerase reaction tested by the acidprecipitable assay (Figure 1(a)). Elongation ofsome oligonucleotides (oligonucleotides 1, 3, 5) inthe product analysis assay with [a-32P]dCTP or[a-32P]dTTP also correlated with the observationmade in the acid precipitable assay. Furtherevidence that self-priming and template slippage
did not account for the observed elongation wasobtained by substituting pol b for hpol l (Figure2(e)). Identical results as in Figure 2(e) were seenwith pol d and hpol l DC (data not shown). Also,no labelling or elongation of oligonucleotides tookplace when the other three [a-32P]dNTPs wereused with pol b, pol d or hpol l DC (data notshown). Finally, to have a positive control TdTwas tested with these eight oligonucleotides and[a-32P]dTTP (Figure 2(f)).
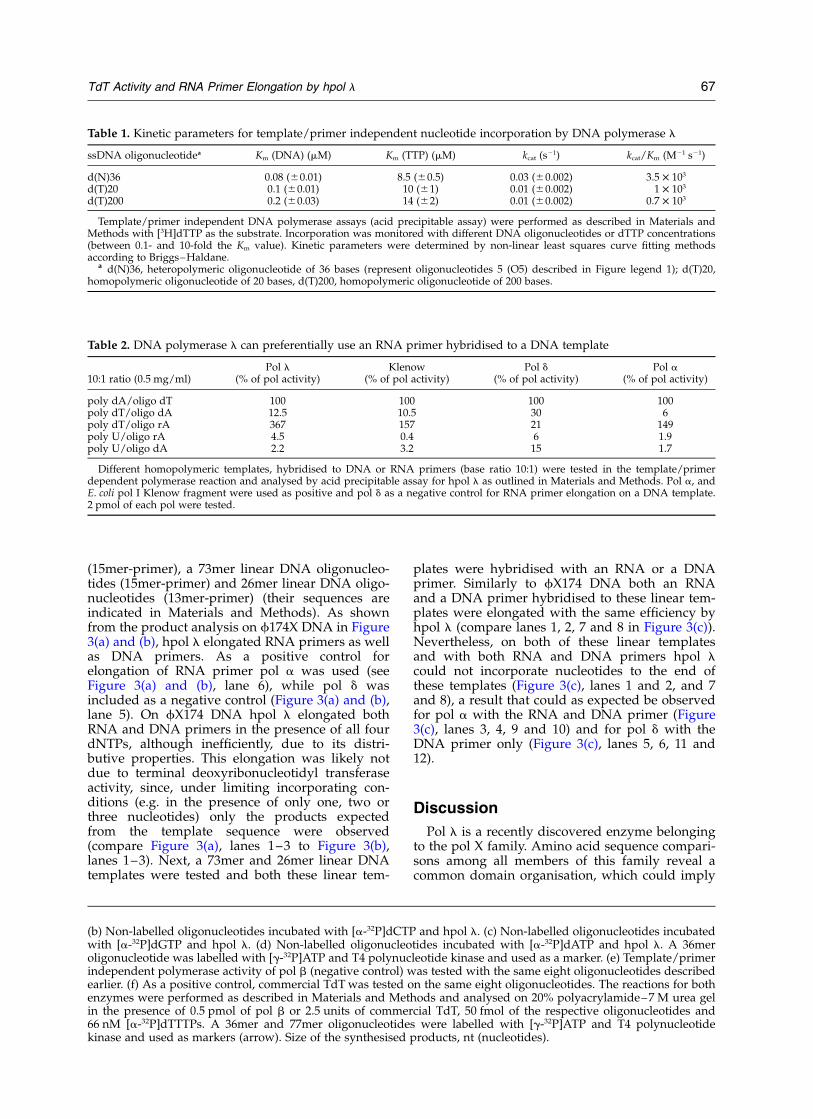
Next to characterise further the terminaldeoxyribonucleotidyl transferase activity of hpoll, we determined the kinetic parameters for thetemplate/primer independent incorporation onboth homo- and heteropolymeric oligonucleotides.As summarised in Table 1, hpol l showed thatwith three oligonucleotides tested (a 36mer hetero-polymer, a 20mer dT homopolymer and a 200merdT homopolymer) the km for oligonucleotideswas 0.08–0.2 mM and for dTTP 8.5–14 mM,respectively.
Human DNA polymerase l can preferentiallyelongate RNA primers hybridised on DNA
Since hpol l also has a template/primer depen-dent DNA polymerase activity, different tem-plate/primers were tested to explore its substratespecificity. A base ratio of 10:1 of template toprimers was found to be optimal for the DNApolymerase activity of hpol l on poly(dA)/oligo(dT) in a template/primer dependent poly-merase reaction measured by acid precipitableassay (data not shown) and was therefore chosenfor all template/primers tested. Pol d (in thepresence of PCNA) served as a negative and pol aas well as E. coli pol I Klenow fragment as positivecontrols. As summarised in Table 2, hpol l wasable to elongate 30-fold better the RNA primeroligo(rA) than the corresponding DNA primeroligo(dA) hybridised to poly(dT) (compare thesecond line to the third line in Table 2). RNAprimer elongation on a DNA template by theclassical mammalian replicative pols a, d and 1can only be performed by pol a, accordingly, wefound that pol a elongated an RNA primer onpoly(dT) 30-fold better than a DNA primer on thesame template (Table 2).11 Our findings indicatedthat in human cells an additional pol, namely hpoll, can elongate RNA primer hybridised to a DNAtemplate with similar efficiency as pol a.
Human DNA polymerase l synthesises DNAon DNA or RNA primed templates
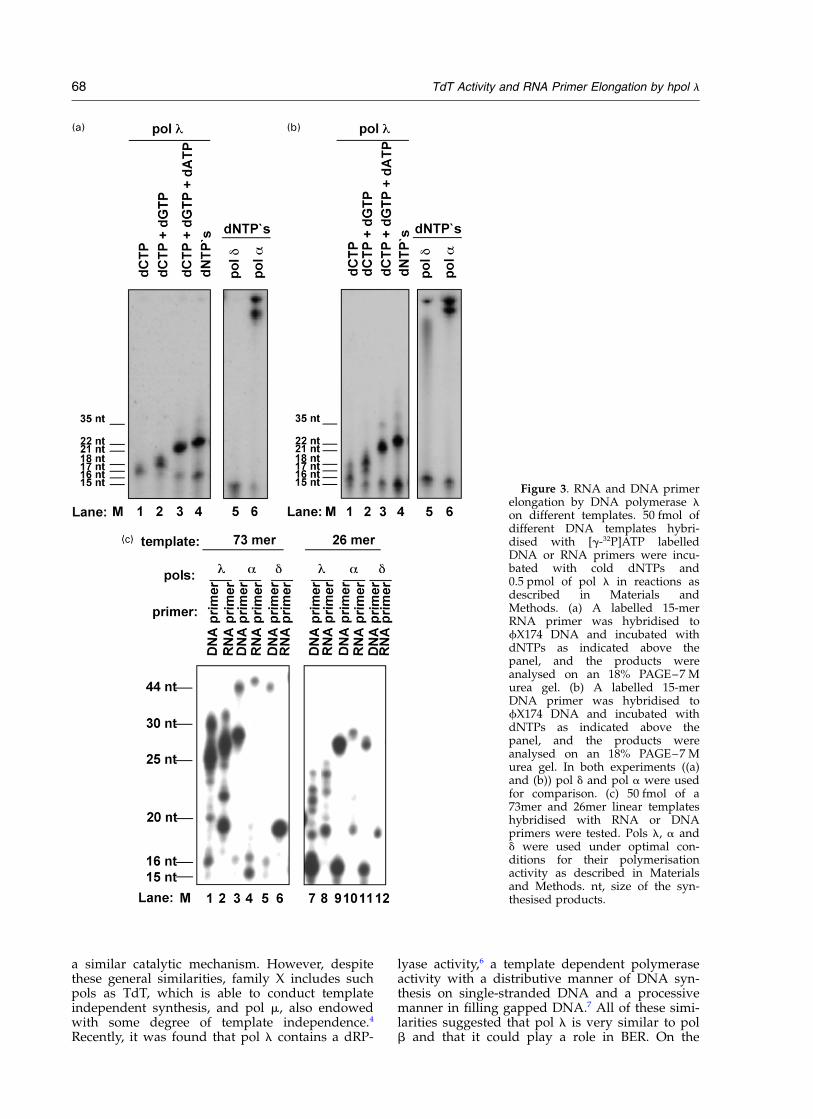
The fact that hpol l could elongate an RNAprimer on homopolymeric poly(dT)/oligo(rA)template in a template/primer dependent poly-merase reaction prompted us to test RNA primerelongation on heteropolymeric linear and circularDNA templates. For this purpose a DNA or anRNA primer was hybridised to different templatessuch as fX174 circular single strand DNA
Figure 1. Terminal deoxyribonucleotidyl transferaseactivity of DNA polymerase l (acid precipitable assay).(a) 50 pmol of eight 36mer oligonucleotides withdifferent sequences and 2 pmol of pol l were tested in atemplate/primer independent polymerase reaction asdescribed in Materials and Methods. The numbers onthe bottom of panels represent oligonucleotides: O1:36mer poly dA; O2: 36mer poly dC; O3: 36mer poly dG;O4: 36mer poly dT, O5: 36mer 50-gag cta gcc cac cat ggatcc cag ggg tat ctt gaa-30; O6: 36mer 50-gag ctg cta acg ttccca ggc tga ctt gac ctg gtt-30, O7: 36mer 50-cag cta agc ttcagg act gac ttc cga cgt act ggg-30, O8: 36mer 50-aac gttgca tga caa ctt ggc agt tca ttg act gcc-30. Each reactionwas tested with separate dNTPs as indicated in theFigure. (b) Commercial TdT was used as positive controlfor the template/primer independent polymerasereaction on the same oligonucletides as in (a). On theright side of both panels dA, dC, dG and dT indicateH3-labelled deoxyribonucleotides used in the assays(dA, deoxyadenosine triphosphate; dC, deoxycytidinetriphosphate; dG, deoxyguanosine triphosphate; and dT,thymidine triphosphate).
TdT Activity and RNA Primer Elongation by hpol l 65
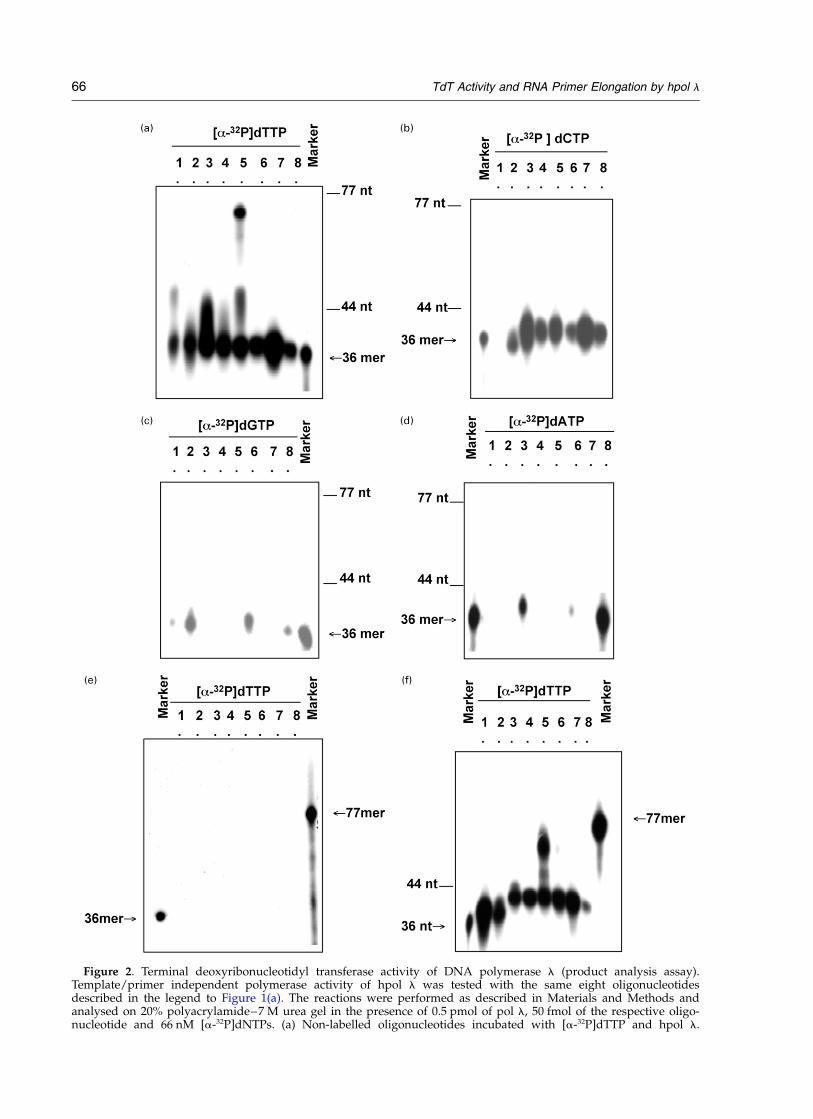
Figure 2. Terminal deoxyribonucleotidyl transferase activity of DNA polymerase l (product analysis assay).Template/primer independent polymerase activity of hpol l was tested with the same eight oligonucleotidesdescribed in the legend to Figure 1(a). The reactions were performed as described in Materials and Methods andanalysed on 20% polyacrylamide–7 M urea gel in the presence of 0.5 pmol of pol l, 50 fmol of the respective oligo-nucleotide and 66 nM [a-32P]dNTPs. (a) Non-labelled oligonucleotides incubated with [a-32P]dTTP and hpol l.
66 TdT Activity and RNA Primer Elongation by hpol l
(15mer-primer), a 73mer linear DNA oligonucleo-tides (15mer-primer) and 26mer linear DNA oligo-nucleotides (13mer-primer) (their sequences areindicated in Materials and Methods). As shownfrom the product analysis on f174X DNA in Figure3(a) and (b), hpol l elongated RNA primers as wellas DNA primers. As a positive control forelongation of RNA primer pol a was used (seeFigure 3(a) and (b), lane 6), while pol d wasincluded as a negative control (Figure 3(a) and (b),lane 5). On fX174 DNA hpol l elongated bothRNA and DNA primers in the presence of all fourdNTPs, although inefficiently, due to its distri-butive properties. This elongation was likely notdue to terminal deoxyribonucleotidyl transferaseactivity, since, under limiting incorporating con-ditions (e.g. in the presence of only one, two orthree nucleotides) only the products expectedfrom the template sequence were observed(compare Figure 3(a), lanes 1–3 to Figure 3(b),lanes 1–3). Next, a 73mer and 26mer linear DNAtemplates were tested and both these linear tem-
plates were hybridised with an RNA or a DNAprimer. Similarly to fX174 DNA both an RNAand a DNA primer hybridised to these linear tem-plates were elongated with the same efficiency byhpol l (compare lanes 1, 2, 7 and 8 in Figure 3(c)).Nevertheless, on both of these linear templatesand with both RNA and DNA primers hpol lcould not incorporate nucleotides to the end ofthese templates (Figure 3(c), lanes 1 and 2, and 7and 8), a result that could as expected be observedfor pol a with the RNA and DNA primer (Figure3(c), lanes 3, 4, 9 and 10) and for pol d with theDNA primer only (Figure 3(c), lanes 5, 6, 11 and12).
Discussion
Pol l is a recently discovered enzyme belongingto the pol X family. Amino acid sequence compari-sons among all members of this family reveal acommon domain organisation, which could imply
(b) Non-labelled oligonucleotides incubated with [a-32P]dCTP and hpol l. (c) Non-labelled oligonucleotides incubatedwith [a-32P]dGTP and hpol l. (d) Non-labelled oligonucleotides incubated with [a-32P]dATP and hpol l. A 36meroligonucleotide was labelled with [g-32P]ATP and T4 polynucleotide kinase and used as a marker. (e) Template/primerindependent polymerase activity of pol b (negative control) was tested with the same eight oligonucleotides describedearlier. (f) As a positive control, commercial TdT was tested on the same eight oligonucleotides. The reactions for bothenzymes were performed as described in Materials and Methods and analysed on 20% polyacrylamide–7 M urea gelin the presence of 0.5 pmol of pol b or 2.5 units of commercial TdT, 50 fmol of the respective oligonucleotides and66 nM [a-32P]dTTTPs. A 36mer and 77mer oligonucleotides were labelled with [g-32P]ATP and T4 polynucleotidekinase and used as markers (arrow). Size of the synthesised products, nt (nucleotides).
Table 1. Kinetic parameters for template/primer independent nucleotide incorporation by DNA polymerase l
ssDNA oligonucleotidea Km (DNA) (mM) Km (TTP) (mM) kcat (s21) kcat/Km (M21 s21)
d(N)36 0.08 (^0.01) 8.5 (^0.5) 0.03 (^0.002) 3.5 £ 103
d(T)20 0.1 (^0.01) 10 (^1) 0.01 (^0.002) 1 £ 103
d(T)200 0.2 (^0.03) 14 (^2) 0.01 (^0.002) 0.7 £ 103
Template/primer independent DNA polymerase assays (acid precipitable assay) were performed as described in Materials andMethods with [3H]dTTP as the substrate. Incorporation was monitored with different DNA oligonucleotides or dTTP concentrations(between 0.1- and 10-fold the Km value). Kinetic parameters were determined by non-linear least squares curve fitting methodsaccording to Briggs–Haldane.
a d(N)36, heteropolymeric oligonucleotide of 36 bases (represent oligonucleotides 5 (O5) described in Figure legend 1); d(T)20,homopolymeric oligonucleotide of 20 bases, d(T)200, homopolymeric oligonucleotide of 200 bases.
Table 2. DNA polymerase l can preferentially use an RNA primer hybridised to a DNA template
10:1 ratio (0.5 mg/ml)Pol l
(% of pol activity)Klenow
(% of pol activity)Pol d
(% of pol activity)Pol a
(% of pol activity)
poly dA/oligo dT 100 100 100 100poly dT/oligo dA 12.5 10.5 30 6poly dT/oligo rA 367 157 21 149poly U/oligo rA 4.5 0.4 6 1.9poly U/oligo dA 2.2 3.2 15 1.7
Different homopolymeric templates, hybridised to DNA or RNA primers (base ratio 10:1) were tested in the template/primerdependent polymerase reaction and analysed by acid precipitable assay for hpol l as outlined in Materials and Methods. Pol a, andE. coli pol I Klenow fragment were used as positive and pol d as a negative control for RNA primer elongation on a DNA template.2 pmol of each pol were tested.
TdT Activity and RNA Primer Elongation by hpol l 67
a similar catalytic mechanism. However, despitethese general similarities, family X includes suchpols as TdT, which is able to conduct templateindependent synthesis, and pol m, also endowedwith some degree of template independence.4
Recently, it was found that pol l contains a dRP-
lyase activity,6 a template dependent polymeraseactivity with a distributive manner of DNA syn-thesis on single-stranded DNA and a processivemanner in filling gapped DNA.7 All of these simi-larities suggested that pol l is very similar to polb and that it could play a role in BER. On the
Figure 3. RNA and DNA primerelongation by DNA polymerase lon different templates. 50 fmol ofdifferent DNA templates hybri-dised with [g-32P]ATP labelledDNA or RNA primers were incu-bated with cold dNTPs and0.5 pmol of pol l in reactions asdescribed in Materials andMethods. (a) A labelled 15-merRNA primer was hybridised tofX174 DNA and incubated withdNTPs as indicated above thepanel, and the products wereanalysed on an 18% PAGE–7 Murea gel. (b) A labelled 15-merDNA primer was hybridised tofX174 DNA and incubated withdNTPs as indicated above thepanel, and the products wereanalysed on an 18% PAGE–7 Murea gel. In both experiments ((a)and (b)) pol d and pol a were usedfor comparison. (c) 50 fmol of a73mer and 26mer linear templateshybridised with RNA or DNAprimers were tested. Pols l, a andd were used under optimal con-ditions for their polymerisationactivity as described in Materialsand Methods. nt, size of the syn-thesised products.
68 TdT Activity and RNA Primer Elongation by hpol l

other hand, differently from pol b, the viability ofpol l deficient mice over several generationsargues that this enzyme is dispensable for mousedevelopment.8 Taken together, these data indicatedthat the cellular function(s) of pol l are largelyunknown. Since pol l shares high homology withboth TdT and pol m, those contain intrinsic tem-plate/primer independent polymerase activity,4
we have tested different sizes and sequences ofoligonucleotides and found that hpol l can extendoligonucleotides in the absence or primers. Thisactivity was detected with a variety of oligonucleo-tide sizes (see Table 1), suggesting that the oligo-nucleotide length is not important for the terminaldeoxyribonucleotidyl transferase activity of hpoll. By testing such oligonucleotides with otherpols, such as hpol b (Figure 2(e)), hpol d and hpoll DC (data not shown) we could demonstrate thatlabelling and elongation of the oligonucleotides byhpol l were not due to self-priming since no label-ling or elongation was observed with these pols.When template/primer independent polymeraseactivity of hpol l was tested without Mn2þ, whichis the metal-ion used here, no incorporation ofnucleotides was detected (data not shown), indi-cating that it was a metal ion-dependent mechan-ism. Nevertheless, since it is known that Mn ionsincrease pol b activity and lower the fidelity ofDNA synthesis7,12 it was conceivable that Mn2þ
could allow the template/primer independentreaction of hpol l. However, we were not able tocheck the influence of Mg2þ on template/primerindependent polymerase activity with recombinanthpol l, since recombinant hpol l has a very lowactivity with Mg2þ even in a template/primerdependent reaction, a finding that correlates withprevious results.7 For this reason we tookadvantage of the natural calf thymus pol lenzyme5 since our previous results showed thatthe natural calf thymus pol l was equally activewith both Mg2þ and Mn2þ in a template/primerdependent polymerase reaction. When the naturalcalf thymus pol l was tested in the template/primer independent polymerase reaction withboth metal ions and [a-32P]dNTP0s, labelling andelongation occurred (data not shown). This resultshowed that template/primer independent poly-merase activity of pol l was not due to the muta-genic effect of Mn2þ on pol l by promotinggreater reactivity than Mg2þ at the catalytic site.12
A possible explanation for the different behaviourof the natural calf thymus pol l and the recombi-nant hpol l towards the use of divalent cationscould stem from different subunit compositions orposttranslational modifications.
In addition, our results suggested that the tem-plate/primer independent polymerase activity ofhpol l is sensitive to oligonucleotide sequences.The sequence specificity was also seen for TdTused as a positive control (see Figures 1(b) and2(f)), thus indicating a similarity between thesetwo enzymes. From both acid precipitable andproduct analysis template/primer independent
polymerase reactions, it is obvious that hpol lpreferentially incorporates pyrimidines. Thispreference for pyrimidine incorporation was alsoobserved for pol m.4
Our data are in controversy with a very recentpublished work.13 In this paper the terminal deoxy-ribonucleotidyl transferase activity of pol l wasonly measured with two oligonucleotides (polydA and oligo dT and only with labelled dTTP),while here, as discussed above, eight differentoligonucleotides with the four labelled dNTPreflected a much broader approach. Moreover, theproducts were also tested by gel analysis. Finallytheir pol l was only active on homopolymericDNA template/primer but not as in our case on anatural circular DNA template/primer, suggestingthat these authors might have inhibited both DNAtemplate dependent and independent polymeraseactivity of pol l with huge amount of dNTPs(80 mM) in the enzymatic reactions. This fact isknown from published result7 and from our data(K.R., G.M., I.V.S. & U.H., unpublished).
We also designed two truncated hpol l proteins,which were overexpressed and purified followingthe procedures employed for the hpol l wt (seeMaterials and Methods). Hpol l DN is a mutantwithout the first 275 aa from the N terminus, butstill contains the polymerase catalytic site andhpol l DC is a mutant lacking 300 aa from the Cterminus and does not contain the polymerasecatalytic active site. Using both of these mutantsin the template/primer dependent and indepen-dent polymerase reactions with both acid precipi-table and product analysis assays we observedthat only hpol l DN displays template/primerdependent polymerase activity as well as terminaldeoxyribonucleotidyl transferase activity whilehpol l DC did not (data not shown). These resultssuggested that the catalytic site of terminal doxy-ribonucleotidyl transferase activity resides also inthe polymerase domain and renders highlyunlikely the possibility that this activity is due tosome contamination during hpol l purification.
The 31-kDa domain of pol b has sequence hom-ology with the catalytic core domain of the p49pol a/primase subunit, and the 8-kDa dRP-lyasedomain of pol b has sequence homology with thep58 pol a/primase subunit.14 It is well known thatthe primase activity in the eukaryotic cell is associ-ated with two of the pol a subunits.15,16 Based onthese similarities between pol b and the primasesubunits and on the fact that pol l has 32%amino acid identity with pol b in the polymerase(31-kDa domain) and the dRP-lyase domain(8-kDa domain),2 we tested the possibility thathpol l could have the ability to elongate RNAprimers, a property previously thought to be con-fined in mammalian cells to pol a only.11 Testing avariety of DNA templates hybridised with RNAor DNA primers we observed that hpol l couldelongate both RNA and DNA primers. BetterDNA synthesis with RNA primer than with aDNA primer was only observed with the linear
TdT Activity and RNA Primer Elongation by hpol l 69

homopolymeric poly(dT)/oligo(rA) template in atemplate/primer dependent polymerase reactionmeasured by acid precipitable assay, an obser-vation which was also made for pol a. Theproducts of DNA synthesis analysed by auto-radiography with both RNA and DNA primerswere similar. The size and pattern of the productsindicated that hpol l polymerises DNA in a distri-butive way and cannot elongate primer to the endof template.
The two novel activities of hpol l described herecould be viewed in the light of possible cellularroles of the enzymes. Calf thymus and human poll can replicate over an abasic site.5,10 AP sites arethose of the most frequently formed lesions inDNA.17–19 If the AP sites are not removed by excisionrepair processes, they present a strong block to thereplication DNA machinery. During replication, APsites can be bypassed either by specialised muta-genic pols, or by error-free mechanisms such asrecombination or a copy-choice type of DNAsynthesis.20 The fact that hpol l can efficientlyelongate RNA primers on DNA templates suggestsa similarity or a supplementarity in function withpol a. These findings, namely the capacity of pol lto extend RNA primers and to efficiently bypass APsites, suggest that pol l might participate in Okazakifragment maturation, allowing to bypass AP sitesfrequently occurring in mammalian cells which canbe also bypassed by pol a.21
The second physiological role of these two novelhpol l activities might be an involvement in end-joining of DSB repair. Pol l closely related pols,TdT and pol m have already been shown to partici-pate in end-joining DSB repair. For instance, TdT isexpressed only in lymphatic tissue, and duringV(D)J recombination randomly adds nucleotidesto the end of coding segments. In addition it wasshown that pol m functions in DSB repair in non-lymphatic tissue after ionizing radiation treatmentof the cells.22 Moreover, the budding yeast Pol4also belonging to the pol X family, efficiently fillsin short gaps formed by the alignment of comple-mentary single strands at the ends of duplexDNA.23 On the other side, genetic studies indicatethat Pol4 participates in end joining events thatrequire nucleolytic processing.24 The terminaldeoxyribonucleotidyl transferase activity of pol lcould play a role in DSB repair. It is also interestingto note that end joining occurs in the G1 stage ofthe cell cycle when low concentrations of dNTPsare present in the cell. The fact that hpol l has avery high affinity for dNTPs (Km ¼ 0.5 mM) in thetemplate/primer dependent polymerase reaction7
may facilitate its participation in this process.
Materials and Methods
Nucleotides and enzymes
Nucleotides were purchased from Pharmacia P-L Bio-chemicals. [g-32P]ATP (3000 Ci/mmol), [a-32P]dATP
(3000 Ci/mmol) and [a-32P]dCTP (3000 Ci/mmol),[a-32P]dGTP (3000 Ci/mmol) and [a-32P]dTTP (3000 Ci/mmol), [3H]dATP (24 Ci/mmol), [3H]dCTP (16 Ci/mmol),[3H]dGTP (14.1 Ci/mmol), [3H]dTTP (49 Ci/mmol), wereobtained from Amersham International Plc. T4 poly-nucleotide kinase and TdT were from BoehringerMannheim. E. coli pol I Klenow fragment was from NewEngland Biolabs. Recombinant human pol b and d werepurified as described in Refs. 25,26 and natural pol a frommammalian cells according to Ref. 27.
Construction of bacterial expression plasmids
The human pol l cDNA was obtained from humanplacenta by PCR amplification, cloned in the bacterialexpression vector pRSET-B which allows expression ofrecombinant protein as fusion with a peptide containinga hexahistidyl sequence for purification on Ni2þ affinityresins as described earlier.2 Truncated forms of hpol lcDNA were amplified by PCR using pRSET-B-pol l as atemplate. A deletion mutant of cDNA for hpol l whichlacks the N-terminal 1–217 aa (hpol l DN), was ampli-fied with primers DN-forward (50gctcggatccgcccctgctgtcctggat03) and DN-reverse (50ggtgaattcaccagtcccgctcagcagg30). A cDNA, which lacks the C-terminal217–575 aa (hpol l DC) was amplified with primersDC-forward ð50cgccggatcccaggggtatcttgaag30Þ; and DC-reverse ð50ccggaattcatgggctaggctcacaatc30Þ: The primersDN-forward and reverse, and DC-forward and reversecontain BamHI and EcoRI sites (underlined), respectively.Both truncated hpol l cDNAs were cloned in thebacterial expression vector pRSET-B, the same onewhich has been used for the full length hpol l (hpol lwt) cDNA and described earlier.2
Overproduction and purification of wt, DN and DCforms of human DNA polymerase l proteins
Expression of full length and both truncated forms ofhpol l were carried out in the E. coli strain BL-21-Codonplus(DE-3)-RIL (Stratagene), with extra copies of theargU, ileY, and leuW tRNA genes. Expression and purifi-cation of all three hpol l proteins were made asdescribed earlier.2 The final fractions of hpol l wt, DNand DC proteins were obtained as single polypeptidesrunning at the expected position when analysed byCoomassie blue stained 10% SDS-polyacrylamide gel.Hpol l wt run at position 68 kDa, hpol l DN at positionabout 40 kDa and hpol l DC at position about 28 kDa.Purified proteins were analysed by Western blotanalysis; hpol l wt, DN and DC polypeptides cross-reacted with polyclonal antibody against hpol l, andonly two of them, hpol l wt and DN polypeptides, cross-reacted with monoclonal antibody against human pol b.
DNA templates and substrates
Pharmacia was the supplier of poly(dA), poly(A),poly(dT), oligo(dT12 – 18), oligo(dA12 – 18) and oligo(A12 – 18).fX174 DNA was purified as described28 and differentDNA primers were from Microsynth (Switzerland). TheRNA primers were from Genset (France). The linearDNA templates (73mer, 26mer) were chemically syn-thesised and purified on a denaturing polyacrylamidegel. The sequences are: 73 mer: 50-GATCGGGAGGGTAGGAATATTGAGGATGAAGGGTTGAGTTGAGT GGAGATAGTGGAGGGTAGTATGGTGGATA-30 26 mer: 50-GTCAGCATCTGTGTGAATTCGGCACT-30.
70 TdT Activity and RNA Primer Elongation by hpol l

The sequences complementary to the correspondingprimers are underlined.
Template/primer dependent DNA polymerase assay
Acid precipitable assay
Final volume of 25 ml contained the following com-ponents: 50 mM Tris–HCl (pH 7.5), 1 mM dithiothreitol,250 mg/ml BSA, 1 mM MnCl2 (for pol l, pol b andE. coli pol I Klenow fragment), 6 mM MgCl2 (for pol aand d), 0.5 mg/ml of different homopolymeric templatesin ratio template:primer as bases of 10:1, and enzymes tobe tested. The concentration of [3H]dNTPs was used inoptimal condition for each of the pols, as describedearlier.11,29 Reactions containing pol d were supple-mented with 50 ng of PCNA. All reactions wereincubated for 15 minutes, 37 8C, precipitated with 10%trichloroacetic acid, and the insoluble radioactivematerial was determined by scintilation counting asdescribed.30
Product analysis assay
Final volume of 5 ml contained the same compositionof buffer as described for the acid precipitable assay,50 fmol of different DNA templates hybridised withDNA or RNA [g-32P]ATP labelled primers (15mer or13mer) in ratio (mole:mole) 1:1, unlabelled dNTPs(different concentrations of nucleotides were used asindicated in the respective Figure legend) and enzymesto be tested. All reactions were incubated at 37 8C for15 minutes, the samples were mixed with denaturinggel loading buffer (95% (v/v) formamide, 10 mM EDTA,0.05% bromophenol blue, 0.05% xylene cyanol), heatedfor five minutes at 95 8C, and then subjected to electro-phoresis on a denaturing gel (7 M urea and different %of polyacrylamide, as indicated in the respective Figurelegend). The reaction products were analysed by auto-radiography.
Template/primer independent DNA polymeraseassays (terminal deoxyribonucleotidyltransferase activity)
Acid precipitable assay
The reaction volume and composition of buffers forpols l, a, b and d were identical to the acid precipitabletemplate/primer dependent DNA polymerase assay.Commercial TdT was used according to the manu-facturer’s protocol. The reactions contained 50 pmolsingle-strand oligonucleotides and 2 mM [3H]dNTPs.The samples were incubated 30 minutes at 37 8C, precipi-tated with 10% trichloroacetic acid, and the insolubleradioactive material was determined by scintilationcounting as described.30
Product analysis assay
The volume of the reaction and the buffer compo-sitions were identical as in the product analysistemplate/primer dependent DNA polymerase assay,but contained 50 fmol of oligonucleotides and 66 nM of[a-32P]dNTPs. Size and sequence of oligonucleotides,concentration of nucleotides and enzymes are indicatedin the respective Figure legends. All reactions were incu-bated at 37 8C for 15 minutes, samples were mixed with
denaturing gel loading buffer and subjected to electro-phoresis on a denaturing gel (7 M urea and different %of polyacrylamide gel, as indicated in the respectiveFigure legends). The reaction products were finallyanalysed by autoradiography.
Acknowledgements
This work was supported by the Swiss NationalScience Foundation (grant 3100. 061361. 00) toK.R., by the “UBS AG im Auftrag eines Kunden”to I.S. and the Kanton of Zurich to U.H., I.S. andG.M.
References
1. Hubscher, U., Maga, G. & Spadari, S. (2002).Eukaryotic DNA polymerases. Annu. Rev. Biochem.71, 133–163.
2. Garcia-Diaz, M., Dominguez, O., Lopez-Fernandez,L. A., de Lera, L. T., Saniger, M. L., Ruiz, J. F. et al.(2000). DNA polymerase lambda (Pol lambda), anovel eukaryotic DNA polymerase with a potentialrole in meiosis. J. Mol. Biol. 301, 851–867.
3. Aoufouchi, S., Flatter, E., Dahan, A., Faili, A.,Bertocci, B., Storck, S. et al. (2000). Two novel humanand mouse DNA polymerases of the polX family.Nucl. Acids Res. 28, 3684–3693.
4. Dominguez, O., Ruiz, J. F., Lain de Lera, T., Garcia-Diaz, M., Gonzalez, M. A., Kirchhoff, T. et al. (2000).DNA polymerase mu (Pol mu), homologous to TdT,could act as a DNA mutator in eukaryotic cells.EMBO J. 19, 1731–1742.
5. Ramadan, K., Shevelev, I. V., Maga, G. & Hubscher,U. (2002). DNA polymerase lambda from calfthymus preferentially replicates damaged DNA.J. Biol. Chem. 277, 18454–18458.
6. Garcia-Diaz, M., Bebenek, K., Kunkel, T. A. & Blanco,L. (2001). Identification of an intrinsic 50-deoxyribose-5-phosphate lyase activity in human DNA polymer-ase lambda: a possible role in base excision repair.J. Biol. Chem. 276, 34659–34663.
7. Garcia-Diaz, M., Bebenek, K., Sabariegos, R.,Dominguez, O., Rodriguez, J., Kirchhoff, T. et al.(2002). DNA polymerase lambda, a novel DNArepair enzyme in human cells. J. Biol. Chem. 277,13184–13191.
8. Bertocci, B., De Smet, A., Flatter, E., Dahan, A.,Bories, J. C., Landreau, C. et al. (2002). Cutting edge:DNA polymerases mu and lambda are dispensablefor Ig gene hypermutation. J. Immunol. 168,3702–3706.
9. Kobayashi, Y., Watanabe, M., Okada, Y., Sawa, H.,Takai, H., Nakanishi, M. et al. (2002). Hydrocephalus,situs inversus, chronic sinusitis, and male infertilityin DNA polymerase lambda-deficient mice: possibleimplication for the pathogenesis of immotile ciliasyndrome. Mol. Cell. Biol. 22, 2769–2776.
10. Maga, G., Villani, G., Ramadan, K., Shevelev, I.,Tanguy Le Gac, N., Blanco, L. et al. (2002). HumanDNA polymerase lambda functionally and physi-cally interacts with proliferating cell nuclear antigenin normal and translesion DNA synthesis. J. Biol.Chem. 277, 48434–48440.
TdT Activity and RNA Primer Elongation by hpol l 71

11. Weiser, T., Gassmann, M., Thommes, P., Ferrari, E.,Hafkemeyer, P. & Hubscher, U. (1991). Biochemicaland functional comparison of DNA polymerasesalpha, delta, and epsilon from calf thymus. J. Biol.Chem. 266, 10420–10428.
12. Pelletier, H., Sawaya, M. R., Wolfle, W., Wilson, S. H.& Kraut, J. (1996). A structural basis for metal ionmutagenicity and nucleotide selectivity in humanDNA polymerase beta. Biochemistry, 35, 12762–12777.
13. Shimazaki, N., Yoshida, K., Kobayashi, T., Toji, S.,Tamai, K. & Koiwai, O. (2002). Over-expression ofhuman DNA polymerase lambda in E. coli andcharacterization of the recombinant enzyme. GeneCells, 7, 639–651.
14. Zerbe, L. K. & Kuchta, R. D. (2002). The p58 subunitof human DNA primase is important for primerinitiation, elongation, and counting. Biochemistry, 41,4891–4900.
15. Takada-Takayama, R., Suzuki, M., Enomoto, T.,Hanaoka, F. & Ui, M. (1990). Purification and charac-terization of mouse DNA polymerase alpha devoidof primase activity. FEBS Letters, 273, 27–30.
16. Arezi, B. & Kuchta, R. D. (2000). Eukaryotic DNAprimase. Trends Biochem. Sci. 25, 572–576.
17. Fortini, P., Bignami, M. & Dogliotti, E. (1990). Evi-dence for AP site formation related to DNA–oxygenalkylation in CHO cells treated with ethylatingagents. Mutat. Res. 236, 129–137.
18. Hang, B., Chenna, A., Fraenkel-Conrat, H. & Singer,B. (1996). An unusual mechanism for the majorhuman apurinic/apyrimidinic (AP) endonucleaseinvolving 50 cleavage of DNA containing a benzene-derived exocyclic adduct in the absence of an APsite. Proc. Natl Acad. Sci. USA, 93, 13737–13741.
19. Podlutsky, A. J., Dianova, I. I., Podust, V. N., Bohr,V. A. & Dianov, G. L. (2001). Human DNA polymer-ase beta initiates DNA synthesis during long-patchrepair of reduced AP sites in DNA. EMBO J. 20,1477–1482.
20. Haracska, L., Kondratick, C. M., Unk, I., Prakash, S.& Prakash, L. (2001). Interaction with PCNA is essen-tial for yeast DNA polymerase eta function. Mol. Cell,8, 407–415.
21. Zerbe, L. K., Goodman, M. F., Efrati, E. & Kuchta,R. D. (1999). Abasic template lesions are strongchain terminators for DNA primase but not forDNA polymerase alpha during the synthesis of newDNA strands. Biochemistry, 38, 12908–12914.
22. Mahajan, K. N., Nick McElhinny, S. A., Mitchell, B. S.& Ramsden, D. A. (2002). Association of DNA poly-merase mu (pol mu) with Ku and ligase IV: role forpol mu in end-joining double-strand break repair.Mol. Cell. Biol. 22, 5194–5202.
23. Tseng, H. M. & Tomkinson, A. E. (2002). A physicaland functional interaction between yeast Pol4 andDnl4–Lif1 Links DNA synthesis and ligation in non-homologous end joining. J. Biol. Chem. 277,45630–45637.
24. Wilson, T. E. & Lieber, M. R. (1999). Efficient proces-sing of DNA ends during yeast nonhomologous endjoining. Evidence for a DNA polymerase beta(Pol4)-dependent pathway. J. Biol. Chem. 274,23599–23609.
25. Beard, W. A. & Wilson, S. H. (1995). Purification anddomain-mapping of mammalian DNA polymerasebeta. Methods Enzymol. 262, 98–107.
26. Podust, V. N., Chang, L. S., Ott, R., Dianov, G. L. &Fanning, E. (2002). Reconstitution of human DNApolymerase delta using recombinant baculoviruses:the p12 subunit potentiates DNA polymerizingactivity of the four-subunit enzyme. J. Biol. Chem.277, 3894–3901.
27. Hubscher, U. (1983). The mammalian primase is partof a high molecular weight DNA polymerase alphapolypeptide. EMBO J. 2, 133–136.
28. Kunkel, T. A., Schaaper, R. M., James, E. & Loeb, L. A.(1983). Infidelity of DNA replication as a basis ofmutagenesis. Basic Life Sci. 23, 63–82.
29. Freemont, P. S., Ollis, D. L., Steitz, T. A. & Joyce, C. M.(1986). A domain of the Klenow fragment ofEscherichia coli DNA polymerase I has polymerasebut no exonuclease activity. Proteins: Struct. Funct.Genet. 1, 66–73.
30. Hubscher, U. & Kornberg, A. (1979). The delta sub-unit of Escherichia coli DNA polymerase III holo-enzyme is the dnaX gene product. Proc. Natl Acad.Sci. USA, 76, 6284–6288.
Edited by J. Karn
(Received 8 January 2003; received in revised form 19 February 2003; accepted 20 February 2003)
72 TdT Activity and RNA Primer Elongation by hpol l
Copyright © 2022 FDOKUMEN




















