
Review of vascularised bone tissue-engineering strategies with a focus on co-culture systems
21
Review of vascularised bone tissue-engineering strategies with a focus on co-culture systems Yuchun Liu 1,2 , Jerry K Y Chan 2,3,4 and Swee-Hin Teoh 1 * 1 Division of Bioengineering, School of Chemical and Biomedical Engineering, Nanyang Technological University, Singapore 637459 2 Experimental Fetal Medicine Group, Department of Obstetrics and Gynaecology, Yong Loo Lin School of Medicine, National University of Singapore, Singapore 119228 3 Department of Reproductive Medicine, KK Women’s and Children’s Hospital, Singapore 229899 4 Cancer and Stem Cell Biology, Duke-NUS Graduate Medical School, Singapore Abstract Poor angiogenesis within tissue-engineered grafts has been identified as a main challenge limiting the clinical introduction of bone tissue-engineering (BTE) approaches for the repair of large bone defects. Thick BTE grafts often exhibit poor cellular viability particularly at the core, leading to graft failure and lack of integration with host tissues. Various BTE approaches have been explored for improving vascularisation in tissue-engineered constructs and are briefly discussed in this review. Recent investigations relating to co-culture systems of endothelial and osteoblast-like cells have shown evidence of BTE efficacy in increasing vascularization in thick constructs. This review provides an overview of key concepts related to bone formation and then focuses on the current state of engineered vascularized co-culture systems using bone repair as a model. It will also address key questions regarding the generation of clinically relevant vascularized bone constructs as well as potential directions and considerations for research with the objective of pursuing engineered co-culture systems in other disciplines of vascularized regenerative medicine. The final objective is to generate serious and functional long-lasting vessels for sustainable angiogenesis that will enable enhanced cellular survival within thick voluminous bone grafts, thereby aiding in bone formation and remodelling in the long term. However, more evidence about the quality of blood vessels formed and its associated functional improvement in bone formation as well as a mechanistic understanding of their interactions are necessary for designing better therapeutic strategies for translation to clinical settings. Copyright © 2012 John Wiley & Sons, Ltd. Received 11 April 2012; Revised 13 July 2012; Accepted 25 August 2012 Keywords co-culture; endothelial; mesenchymal stem cell; vascularization; scaffold; bioreactor; hypoxia; bone tissue engineering 1. Bone architecture 1.1. Microarchitecture of vascularized bone tissue The human skeletal system consists of 80% dense cortical bone that surrounds the entire bone and primarily occupies the marrow space at the diaphysis, while the honeycombed network of trabecular bone largely constitutes the inner region of the epiphysis and metaphysis. Due to the high density of cortical bone, haversian systems contribute to its porosity through concentric layers of lamellae surrounding haversian canals that contain bone nerve and blood supply (Deng and Liu, 2005; Eriksen et al., 1994). Examination of the anatomy of bone reveals a complex hierarchical struc- ture, beginning with the sub-nano level of molecules and minerals where collagenous proteins are arranged in fibrillar bundles that form lamellae on the sub-micron level. The microstructure of bone is comprised of haversian systems, osteons and layers of lamellae that form the macrostructure of cortical and cancellous bone (Fig. 1A) (Rho et al., 1998). In addition, bone has high vascularity, which is necessary for *Correspondence to: Prof. Teoh Swee-Hin, PhD., Division of Bioengineering, School of Chemical and Biomedical Engineering, 70 Nanyang Drive, Nanyang Technological University, Singapore 637457. Telephone: 65-65141017 E-mail: [email protected] Copyright © 2012 John Wiley & Sons, Ltd. JOURNAL OF TISSUE ENGINEERING AND REGENERATIVE MEDICINE REVIEW J Tissue Eng Regen Med (2012) Published online in Wiley Online Library (wileyonlinelibrary.com) DOI: 10.1002/term.1617
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Review of vascularised bone tissue-engineering strategies with a focus on co-culture systems

Review of vascularised bone tissue-engineeringstrategies with a focus on co-culture systemsYuchun Liu1,2, Jerry K Y Chan2,3,4 and Swee-Hin Teoh1*1Division of Bioengineering, School of Chemical and Biomedical Engineering, Nanyang Technological University, Singapore 6374592Experimental Fetal Medicine Group, Department of Obstetrics and Gynaecology, Yong Loo Lin School of Medicine, National University ofSingapore, Singapore 1192283Department of Reproductive Medicine, KK Women’s and Children’s Hospital, Singapore 2298994Cancer and Stem Cell Biology, Duke-NUS Graduate Medical School, Singapore
Abstract
Poor angiogenesis within tissue-engineered grafts has been identified as a main challenge limitingthe clinical introduction of bone tissue-engineering (BTE) approaches for the repair of large bonedefects. Thick BTE grafts often exhibit poor cellular viability particularly at the core, leading to graftfailure and lack of integration with host tissues. Various BTE approaches have been explored forimproving vascularisation in tissue-engineered constructs and are briefly discussed in this review.Recent investigations relating to co-culture systems of endothelial and osteoblast-like cells haveshown evidence of BTE efficacy in increasing vascularization in thick constructs. This review providesan overview of key concepts related to bone formation and then focuses on the current state ofengineered vascularized co-culture systems using bone repair as a model. It will also address keyquestions regarding the generation of clinically relevant vascularized bone constructs as well aspotential directions and considerations for research with the objective of pursuing engineeredco-culture systems in other disciplines of vascularized regenerative medicine. The final objective isto generate serious and functional long-lasting vessels for sustainable angiogenesis that will enableenhanced cellular survival within thick voluminous bone grafts, thereby aiding in bone formationand remodelling in the long term. However, more evidence about the quality of blood vessels formedand its associated functional improvement in bone formation as well as a mechanistic understandingof their interactions are necessary for designing better therapeutic strategies for translation toclinical settings. Copyright © 2012 John Wiley & Sons, Ltd.
Received 11 April 2012; Revised 13 July 2012; Accepted 25 August 2012
Keywords co-culture; endothelial; mesenchymal stem cell; vascularization; scaffold; bioreactor; hypoxia;bone tissue engineering
1. Bone architecture
1.1. Microarchitecture of vascularized bonetissue
The human skeletal system consists of 80% dense corticalbone that surrounds the entire bone and primarily occupiesthe marrow space at the diaphysis, while the honeycombed
network of trabecular bone largely constitutes the innerregion of the epiphysis and metaphysis. Due to the highdensity of cortical bone, haversian systems contribute to itsporosity through concentric layers of lamellae surroundinghaversian canals that contain bone nerve and blood supply(Deng and Liu, 2005; Eriksen et al., 1994). Examination ofthe anatomy of bone reveals a complex hierarchical struc-ture, beginning with the sub-nano level of molecules andminerals where collagenous proteins are arranged in fibrillarbundles that form lamellae on the sub-micron level. Themicrostructure of bone is comprised of haversian systems,osteons and layers of lamellae that form the macrostructureof cortical and cancellous bone (Fig.1A) (Rho et al., 1998). Inaddition, bone has high vascularity, which is necessary for
*Correspondence to: Prof. Teoh Swee-Hin, PhD., Division ofBioengineering, School of Chemical and Biomedical Engineering,70 Nanyang Drive, Nanyang Technological University, Singapore637457. Telephone: 65-65141017 E-mail: [email protected]
Copyright © 2012 John Wiley & Sons, Ltd.
JOURNAL OF TISSUE ENGINEERING AND REGENERATIVE MEDICINE REVIEWJ Tissue Eng Regen Med (2012)Published online in Wiley Online Library (wileyonlinelibrary.com) DOI: 10.1002/term.1617

the maintenance of cellular survival, active remodelling andskeletal integrity (Fig. 1B).
1.2. The four stages of bone formation
To achieve successful bone tissue-engineering (BTE), currentstrategies should be directed towards achieving late-stagetissue-engineered bone with an organised structural orienta-tion that is associated with at least moderate mechanicalproperties. In the current study, we summarized the fourprogressive stages of bone regeneration from precipitationof mineralized nodules to lamellar bone formation (Table 1).
1.3. Natural bone healing and formationprocesses
1.3.1. Biochemical factors and oxygen microenvir-onments required for vascularization
During natural bone reparative processes, both endochon-dral and intramembraneous ossification occurred in proxim-ity of a vascular network (Kanczler and Oreffo, 2008).Following fracture, the disrupted vascular supply resultedin a hypoxic environment that acted as a stimulus for upregu-lation of a group of angiogenic factors that formed a vascularnetwork. Endochondral ossification was first initiated by aninflammatory phase that activated a cascade of cellularevents that recruited several cell types to the fracturearea, including resident mesenchymal stem cells (MSCs).
MSCs differentiated and formed cartilaginous material tharesorbed and mineralized into bone following formationof a vascular network. Intramembranous ossificationoccurred through direct differentiation of MSCs presentin the medullar cavity without the formation of cartilageduring the reparative process. Briefly, it was activation ofthe hypoxia-inducible factor 1 (HIF-1) pathway in anischemia microenvironment that triggered upregulationof angiogenic factors such as vascular endothelial growthfactor (VEGF) (Germain et al., 2010). The generation ofgrowth factors then triggered the migration of endothelialcells with their tip cells acting as oxygen sensors guidingvascular invasion into hypoxic zones while their stalk cellsproliferated, thereby forming a vascular network (Germainet al., 2010; Coulon et al., 2010) (Fig. 2).
Early predictions from the 19th century suggested thatvascular stimulating factors present at bone damage sitesinitiate osteogenesis (Trueta and Buhr, 1963). This has sincebeen verified by studies that showed a need for an adequatevascular supply for bone regeneration to succeed. For exam-ple, endogenous VEGF has been shown to be essential forendochondral bone formation (Gerber et al., 1999; Zelzeret al., 2002; Street et al., 2002). Inhibition of VEGF by asoluble neutralising VEGF receptor during endochondraland intramembraneous ossification led to decreased angio-genesis, with reduced callus mineralisation and bone forma-tion in a femoral fracture and tibia cortical bone defect inmice. Conversely, an exogenous VEGF supply acceleratedbone bridging across a rabbit radius critical-sized defect,implicating a distinct role of VEGF as an angiogenic factorin promoting bone healing (Street et al., 2002). Optimal
Figure 1. (A) Hierarchical structure of bone from a sub-nanostructure of collagenmolecules to a nanostructure of cylindrically arrangedmicrofibrils to lamellar layers forming the macrostructure of cortical bone (adapted from Rho et al. (1998). (B) Vascularised boneanatomy and microenvironmental influences such as biomechanical, microarchitectural and low oxygen tension factors
Y. Liu et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

healing of the defect region was heavily dependant onadequate vascularisation, thereby suggesting an integral roleof blood vessels in bone regeneration.
To mediate the downstream responses of growth factors,the potent catalytic action of bioactive cofactors involved infacilitating their binding to targeted ligands, thereby concen-trating them on the cell surface and preventing extracellularprotease degradation, was examined. For example, heparinsulphate, a linear unbranched sugar, has been identified as apotent cofactor responsible for regulating cell progenitorphenotype, growth and differentiation by coordinatinggrowth factor orientation (Cool and Nurcombe, 2006).Particularly during osteogenesis, heparin sulphates and theirbinding factors have been found to be important bioactive ele-ments in the mineralised matrix by potentiating the responseof bone-inducing factors and by regulating cellular transduc-tion and downstream processes (Cool and Nurcombe, 2005).A key factor was the fibroblast growth factor family, whichplayed an integral role in osteogenesis (Boilly et al., 2000; Coolet al., 2002; Jackson et al., 2006). Conversely, a loss or defectin heparin sulphates led to disruptions in various osteogenicpathways, thereby resulting in bone anomalies (Marie et al.,2000; Song et al., 2005). Based on these findings, we assumedthat the underlying catalytic functions of bioactive cofactorssuch as heparin sulphates had a significant influence on theperformance of various bioactive growth factors towardsenhancing cellular responses. Understanding the molecularsignalling pathways during bone development is thereforeimportant for achieving successful BTE therapy.
Figure 2. Role of hypoxia in the angiogenic sprouting process whereangiogenic genes were upregulated and growth factors secreted,thereby inducing (a) sprouting of endothelial cells and (b), tip cellmigration towards the hypoxic stimulus while stalk cells followedbehind. (c) A new vascular network formed with the emergence ofvarious branch points over time (Carmeliet and Jain, 2000; Woodand Asada, 2011). Illustration of emergence of tip cells during angio-genesis adapted fromWood and Asada (2011)
Table 1. Four-stage bone forming process of structural bone, mechanical properties and vessel formation over healing duration;X indicates relative intensity
Stages ofBone Formation
Stage 1 Stage 2 Stage 3 Stage 4
Mineralisation Woven bone Trabecular bone Lamellae bone
MechanicalStrength
X XX XXX XXXX
Bone Structure Random distribution:Nucleation and rapidgrowth of mineralnodules occurred undera thermodynamic drivingforce when in 3D space
Disjointed fine plate-likenetworks of two or moreconnecting fibresrandomly oriented;Low density with largeporosity
Bone plates are joined andhighly oriented, conformingto applied stresses
Dense and compact:Multilayered bone platesarranged in a concentricmanner conforming toapplied stresses
Time interval 1-2weeks 3-6weeks ~ 3-6months Months-years
Vascularisation
(a) Extraosseous X XXX XXX XXX
(b) Medullary - X XX XXX
Cytokines Discontinuous and temporally regulated at high levels Moderate regulation for homeostasis and remodelling
Vascularised Bone Tissue Engineering using Coculture Systems
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

1.3.2. Biomechanical factors duringbone formation
Besides the role of vascularization in aiding bone formation,there are various biomechanical factors involved in bone for-mation and remodelling. During daily activities, the humanskeletal system experiences various biomechanical forcesincluding compressive, bending and torsional loads. On amicroscopic level, strain is experienced due to tissuedeformation. Fluid shear stresses are also exerted by intersti-tial fluid flow through the lacunae, driven largely by com-pression and tension under loading on the bone (McCoyand O’Brien, 2010). Bone develops by two mechanisms;endochondral and intramembraneous ossification. It isknown that mechanical stimuli have an influence on embry-onic bone formation (Lelkes, 1958; Hosseini and Hogg,1991a, 1991b) as well as during post-embryonic bone repair(Carter and Beaupré, 2001; Carter et al., 1998). Its effectson bone formation have been hypothesized to be medi-ated through mechanosensitive genes that alter mechan-ical influences, thereby modulating the mechanosensitivepathways during bone development and regeneration(Nowlan et al., 2008).
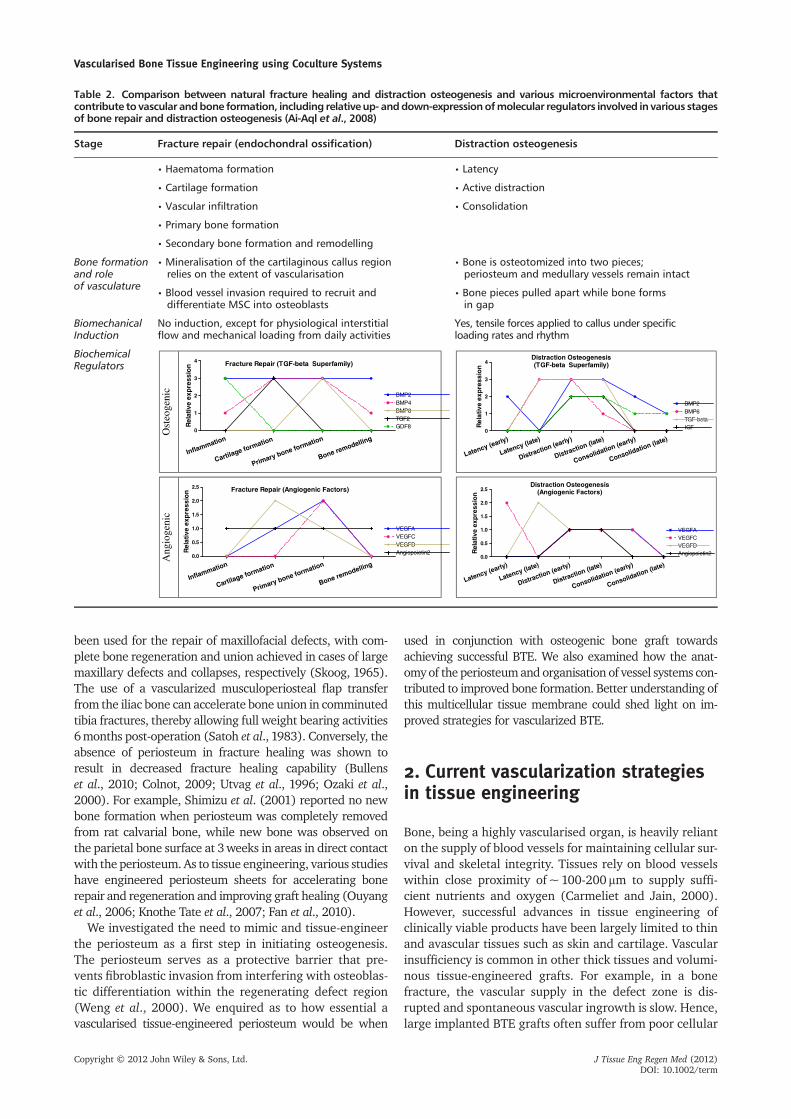
In addition, bone is a dynamic living tissue that under-goes remodelling by which tissue continuously resorbsand deposits new bone simultaneously during bonehomeostasis during an individual’s lifetime. Early observa-tions have shown that this process can be adapted bymechanical stimuli by which Wolff (1892) states thatbone adapts its structure and remodels according tomechanical loads. Although the influence of the mechan-ical transduction pathway on bone cells during thisprocess of adaptive remodelling is poorly understood, ithas been shown that mechanical stress favours the forma-tion of bone as opposed to bone loss under non-weightbearing circumstances (Leondes, 2001). In soft tissueremodelling, a similar concept has been proposed, statingthat remodelling efficacy of soft tissues is based onimposed demands (Henenfeld, 1953). Putting Davis’ lawinto perspective during bone formation, we revisited theprocess of distraction osteogenesis, a surgical processfor bone lengthening. This surgical technique relies onmechanical stimuli (tensile forces) by which twoosteotomized bone pieces are gradually pulled apartwhile new bone forms within the gap. The longitudinalmechanical strain exerted on the callus during thedistraction phase initiates biological responses thatstimulate the formation of mineralized bone andeventual remodelling (Ai-Aql et al., 2008).
There are various microenvironmental factors presentin natural bone during bone formation and remodelling,as illustrated in Figure 1 and Table 2. Both fracturehealing and distraction osteogenesis processes are regu-lated by a differential expression of a mix of biochemicalfactors including inflammatory, osteogenic and angiogenicgrowth factors (Ai-Aql et al., 2008). Biomechanical factorsare also relevant in vascular physiology in bone, whereendothelial cells in blood vessels are constantly exposedto blood flow tangential to the endothelial monolayer.
This is essential for controlling many aspects duringthe progress of an endothelial cell, including mediationof transcriptional changes, endothelial sprouting (Songand Munn, 2011), proliferation (Lin et al., 2000) and vesselhomeostasis (Hudlicka et al., 2006; Jones et al., 2006). Thesignificant effects of biomechanical forces for stimulatingvessel and bone formation and remodelling have alsobeen well demonstrated experimentally (Stolberg andMcCloskey, 2009; Grellier et al., 2009a; Kimelman-Bleichet al., 2011; Ehrlich and Lanyon, 2002; Yamamoto et al.,2005; Yamamoto et al., 2003; McCloskey et al., 2006).
1.4. Periosteum and its vasculature
Researchers have long explored the bone forming poten-tial of periosteum. The periosteum is a thin and highlyvascularized bilayered tissue membrane attached to bonesurfaces by a matrix of collagenous fibre bundles knownas Sharpeys fibres (McBride et al., 2010). The outer layeris dense and consists of fibroblasts and the inner cambiumlayer is rich in MSCs capable of directed differentiationinto bone and cartilage for bone growth and regeneration(Soltan et al., 2009; De Bari et al., 2006). The inner layerof the periosteum also plays an essential role in boneremodelling, where osteoblasts differentiate after bonematrix deposition and osteoclast activity (Jacobsen,1997; Tran Van et al., 1982).
As to the vascular system, the highly vascularised peri-osteum consists of a network of microvasculature in theouter fibrous layer of the periosteum known as the intrin-sic periosteal system and is comprised of vessels encirclingthe bone in a random, circular or longitudinally pat-tern (Simpson, 1985). Other periosteal-associated vesselsystems include musculoperiosteal and fascioperiostealvessels that connect muscles and periosteum, while corti-cal capillary anastomoses are linked to intramedullarycirculation in the bone cortex (Simpson, 1985). Thisextensive blood vessel supply, which exists within thenatural periosteum, suggests the importance of mimickingits vessel network organisation and spatial distribution whendesigning and constructing artificial vessel systems for BTE.
The periosteumplays a crucial role during fracture healing(Bullens et al., 2010). During a fracture, the outer cambiumlayer of the activated periosteum is responsible for drivingthe initiation of osteogenesis and angiogenesis for bonerepair and directs the differentiation of stem cells into bonelineage (Tran Van et al., 1982). It has been suggested thatsince the periosteum serves as a local source of osteoprogeni-tors for bone healing and remodelling (Eyre-Brook, 1984;Kostopoulos and Karring, 1995; Stevenson, 1999), itsabsence would therefore rely on mobilizing cells from othersources, which would delay the onset of bone repair. Experi-mentally, the osteogenic and angiogenic potencies of theperiosteum have been investigated in various in vitro andin vivo studies and showed revitalisation of allografts andenhancement in bone regeneration (Jacobsen, 1997; TranVan et al., 1982; Lemperle et al., 1998; Melcher and Accursi,1971; Runyan et al., 2010). Clinically, periosteal flaps have
Y. Liu et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term
been used for the repair of maxillofacial defects, with com-plete bone regeneration and union achieved in cases of largemaxillary defects and collapses, respectively (Skoog, 1965).The use of a vascularized musculoperiosteal flap transferfrom the iliac bone can accelerate bone union in comminutedtibia fractures, thereby allowing full weight bearing activities6months post-operation (Satoh et al., 1983). Conversely, theabsence of periosteum in fracture healing was shown toresult in decreased fracture healing capability (Bullenset al., 2010; Colnot, 2009; Utvag et al., 1996; Ozaki et al.,2000). For example, Shimizu et al. (2001) reported no newbone formation when periosteum was completely removedfrom rat calvarial bone, while new bone was observed onthe parietal bone surface at 3weeks in areas in direct contactwith the periosteum. As to tissue engineering, various studieshave engineered periosteum sheets for accelerating bonerepair and regeneration and improving graft healing (Ouyanget al., 2006; Knothe Tate et al., 2007; Fan et al., 2010).
We investigated the need to mimic and tissue-engineerthe periosteum as a first step in initiating osteogenesis.The periosteum serves as a protective barrier that pre-vents fibroblastic invasion from interfering with osteoblas-tic differentiation within the regenerating defect region(Weng et al., 2000). We enquired as to how essential avascularised tissue-engineered periosteum would be when
used in conjunction with osteogenic bone graft towardsachieving successful BTE. We also examined how the anat-omyof the periosteumand organisation of vessel systems con-tributed to improved bone formation. Better understanding ofthis multicellular tissue membrane could shed light on im-proved strategies for vascularized BTE.
2. Current vascularization strategiesin tissue engineering
Bone, being a highly vascularised organ, is heavily relianton the supply of blood vessels for maintaining cellular sur-vival and skeletal integrity. Tissues rely on blood vesselswithin close proximity of~100-200 mm to supply suffi-cient nutrients and oxygen (Carmeliet and Jain, 2000).However, successful advances in tissue engineering ofclinically viable products have been largely limited to thinand avascular tissues such as skin and cartilage. Vascularinsufficiency is common in other thick tissues and volumi-nous tissue-engineered grafts. For example, in a bonefracture, the vascular supply in the defect zone is dis-rupted and spontaneous vascular ingrowth is slow. Hence,large implanted BTE grafts often suffer from poor cellular
Table 2. Comparison between natural fracture healing and distraction osteogenesis and various microenvironmental factors thatcontribute to vascular andbone formation, including relative up- anddown-expressionofmolecular regulators involved in various stagesof bone repair and distraction osteogenesis (Ai-Aql et al., 2008)
Stage Fracture repair (endochondral ossification) Distraction osteogenesis
• Haematoma formation • Latency
• Cartilage formation • Active distraction
• Vascular infiltration • Consolidation
• Primary bone formation
• Secondary bone formation and remodelling
Bone formationand roleof vasculature
• Mineralisation of the cartilaginous callus regionrelies on the extent of vascularisation
• Bone is osteotomized into two pieces;periosteum and medullary vessels remain intact
• Blood vessel invasion required to recruit anddifferentiate MSC into osteoblasts
• Bone pieces pulled apart while bone formsin gap
BiomechanicalInduction
No induction, except for physiological interstitialflow and mechanical loading from daily activities
Yes, tensile forces applied to callus under specificloading rates and rhythm
BiochemicalRegulators
Ost
eoge
nic
Fracture Repair (TGF-beta Superfamily)
Inflammation
Cartilage formation
Primary bone formation
Bone remodelling0
1
2
3
4
BMP2BMP4BMP8TGF2GDF8R
elat
ive
exp
ress
ion
Distraction Osteogenesis(TGF-beta Superfamily)
Latency (early)
Latency (late)
Distraction (early)
Distraction (late)
Consolidation (early)
Consolidation (late)0
1
2
3
4
BMP2BMP6TGF-betaIGFR
elat
ive
exp
ress
ion
Ang
ioge
nic
Fracture Repair (Angiogenic Factors)
Inflammation
Cartilage formation
Primary bone formation
Bone remodelling0.0
0.5
1.0
1.5
2.0
2.5
VEGFAVEGFCVEGFDAngiopoietin2R
elat
ive
exp
ress
ion
Distraction Osteogenesis(Angiogenic Factors)
Latency (early)
Latency (late)
Distraction (early)
Distraction (late)
Consolidation (early)
Consolidation (late)0.0
0.5
1.0
1.5
2.0
2.5
VEGFAVEGFCVEGFDAngiopoietin2R
elat
ive
exp
ress
ion
Vascularised Bone Tissue Engineering using Coculture Systems
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

survival particularly in the core due to inadequate exchangeof nutrients and oxygen (Fig. 3) (Rouwkema et al., 2008).This results in non-uniform cell integration, cellular necrosisand eventual failure of the BTE graft (Kanczler and Oreffo,2008; Rouwkema et al., 2008; Ko et al., 2007), thereby limit-ing the clinical translation of tissue-engineering research. Toaddress this issue, several strategies have been undertakento introduce vasculature into tissue-engineered constructs(Rouwkema et al., 2008; Lovett et al., 2009).
2.1. Smart scaffolds and growth factors
One strategy involves the use of smart scaffold technology byfunctionalizing scaffold surfaces through physical adsorptionby intermediate proteins and biological molecules or by directcovalent immobilisation of bioactive molecules such asgrowth factors that promote vasculogenesis and angiogenesis(Fig. 4). Growth factors can also be physically encapsu-lated within the scaffolds, thereby acting as delivery systemsfor controlled release over time (Lee et al., 2011). For
example, VEGF covalent-linked collagen scaffolds resultedin rapid vascularisation in vivo, leading to improved myocar-dial repair compared to untreated control. In another study,Kaigler et al. (2006) showed increased blood vessel formationinvading the VEGF-encapsulated poly(lactic-co-glycolic acid)scaffolds accompanied by earlier and greater bone formationin an irradiated rat calvarial defect model compared toscaffolds alone. In gene therapy, stem cells such as MSCshave been used as vehicles for angiogenic growth factordelivery where cells were transduced ex vivo, thereby encod-ing growth factors. For example, the use of VEGF-transducedMSCs showed increased vascularity on VEGF expressionalone, which was in synergy with VEGF/bone morphogeneticprotein-2 (BMP-2) that transduced MSC-enhanced boneformation in a segmental tibia defect mouse model (Kumaret al., 2010). Many studies have demonstrated the significantbenefits of angiogenic growth factor delivery (Kempen et al.,2009; Street et al., 2002) and gene therapy (Ito et al., 2005;Peng et al., 2002) in aiding bone regeneration for use inBTE applications.
Although these methods allowed mobilization of thepatient’s own stem cells for self-reparative bone healingmechanisms and thereby avoided all issues pertaining toex vivo cell culture and expansion, they however relied oningrowth of host vasculature, which often takes time.Moreover, the expression of angiogenic factors followed atemporal pattern that was tightly regulated. Ai-Aql et al.(2008) reported a sequential temporal expression of differentangiogenic factors occurring at different stages of bone repair(endochondral ossification) and distraction osteogenesis.However, current growth factor delivery systems releasegrowth factors on a continuous burst release profile, makingit difficult to control the temporal and spatial release profileof growth factors. We recently published and demonstratedfor the first time the efficacy of a discontinuous release ofBMP-2 from a 3D scaffold in promoting healing in a segmentalrabbit ulnamodel (Bae et al., 2011). Furthermore, recent issuesrelating to the safety (Boraiah et al., 2009; Shields et al., 2006;Wong et al., 2008) and efficacy of the use of growth factors inclinical trials have also been raised (Simons et al., 2002), likelyattributed to poor understanding of growth factors such asappropriate dosage, mode of administration and otherclinical trial design-related issues (Lee et al., 2011).
Figure 3. Conceptual illustration of cell viability and death in athick scaffold graft and core at a critical distance of≥100-200mmfrom blood vessel supply
Figure 4. Diagrammatic representation of porous scaffold constructs (A) encapsulated with growth factors and their time-dependentcontrol release (B) surface functionalized with growth factors by physical adsorption and covalent immobilisation methods
Y. Liu et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

2.2. In vitro and in vivo prevascularization
A rapidly growing field of research is currently focusing onthe generation of prevascularization in grafts through bothin vitro and in vivo approaches. A two-step in vivo prevascu-larization strategy involves the initial implantation of tissue-engineered grafts within well-vascularised sites such asmuscle for complete vascularization prior to subsequentimplantation at the defect site (Warnke et al., 2004; Kneseret al., 2006b; Kneser et al., 2006a). This approach allowsfor instantaneous reperfusion by surgical anastomosis,thereby ensuring its therapeutic efficacy. However, it islimited by the need for multiple surgeries, hence limiting itsclinical appeal. Another approach proposes the use of endo-thelial cells in co-culture with other cell types to achievein vitro prevascularization and generate vascularizationwithin the scaffold construct (Melero-Martin et al., 2008;Fuchs et al., 2009a; Fuchs et al., 2007; Tsigkou et al., 2010;Liu et al., 2012). However, this method involves the chal-lenge of generating stable in vitro vasculature in the con-structs with a slower rate of anastomosis efficacy comparedto in vivo prevascularization. With pre-formed vessels, hostvessels only need to grow to the outer regions of the graftwhere they can anastomose the prevascularized vessels,thereby shortening the time required to achieve effectivevascularisation of the tissue-engineered graft. Such anapproach avoids the need for multiple surgeries, althoughreperfusion efficacy might not be as efficient as in vivoprevascularization (Rouwkema et al., 2008). In the currentstudy,we focused on the use of a cell-based co-culture systemfor the generation of prevascularized BTE grafts.
2.3. Trends in co-culture studies
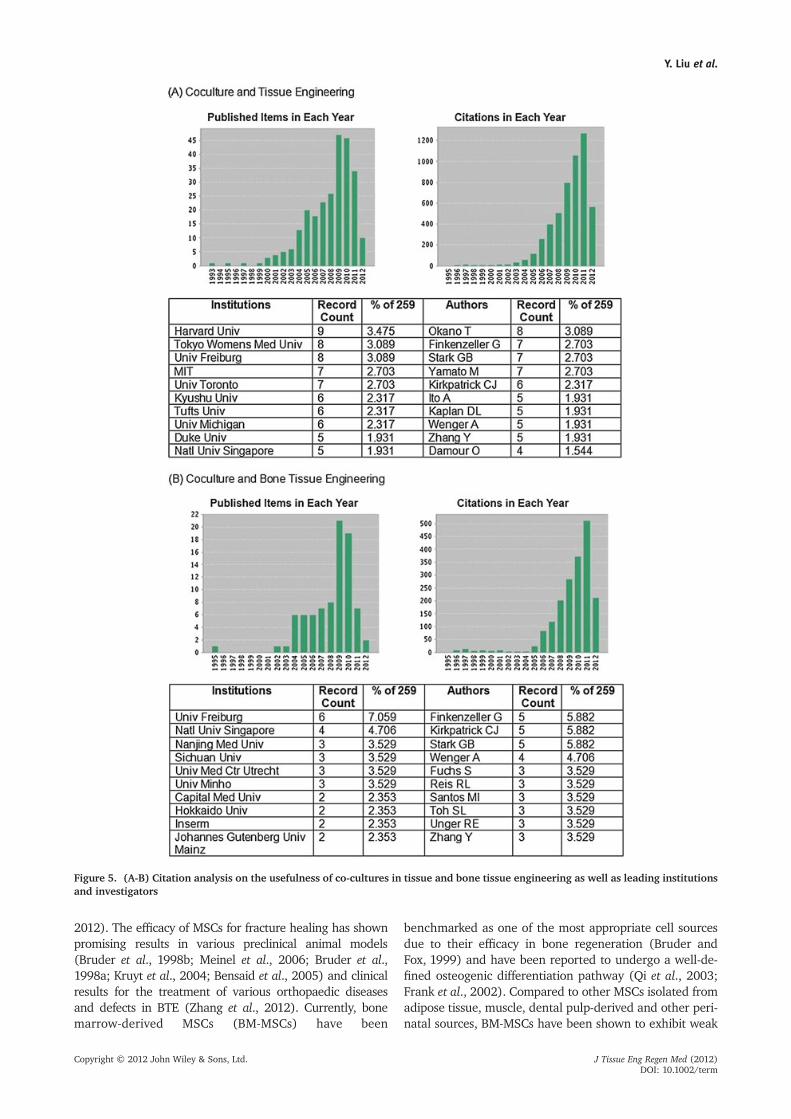
We reviewed the current state of the art and identified asignificant rise in the number of publications on co-culturesystems in tissue engineering (259 publications) over thepast two decades and found a significant proportion ofstudies focused on engineering vascularized bone (85 publi-cations) (Fig. 5, Web of Science, 2012).
3. Current status of co-culturesystems in BTE
Tissue engineering involves an integrative approach to theinteractions between stem cells, biomaterials, molecularfactors and other microenvironmental factors translated intoappropriate animal models. It would be ideal if animalstudies could be monitored through non-invasive imagingprocedures to help address animal-specific efficacy of theimplanted graft. The study of a co-culture system using morethan one cell type resulted in increased complexities. Therewere different factors that remained unexplored and poorlyunderstood that need to be examined in such co-culturesystems, thereby making experimental comparisonsbetween reported studies difficult if not impossible.
These parameters need to be optimized and standardizedin future co-culture studies. To better engineer tissues,we need to break down the approach to “basic units” tobetter understand and develop steps for a more system-atic approach. Several culture conditions need to becarefully considered for optimal performance to assesstherapeutic effectiveness in these tissue-engineered grafts.In this section, we will review the current state of co-culturesystems based on various key parameters as they relate tonatural mimicry of the bone (Fig. 1). Thorough investiga-tions are necessary before identifying the best approachfor the generation of functional and effective vascular-ized bone tissue-engineered constructs.
3.1. Parameters to consider
3.1.1. Cell type
Osteogenic cells. The choice of cell type is crucial fordetermining the efficacy of bone regeneration. Variouscell types have been used for BTE and can be broadlycategorized as bone marrow aspirates, lineage-specificosteoblasts and directed differentiation of stem cells(Zhang et al., 2012; Szpalski et al., 2012). The use of freshbone marrow aspirates was the first cell-based approachused in BTE and was shown to contribute to effective boneregeneration in various experimental animal models(Grundel et al., 1991; Werntz et al., 1996) as well as inclinical studies (Connolly et al., 1989; Quarto et al.,2001). However, the quantity of osteoprogenitors in thebone marrow reservoir was largely dependent on patientage and health. As a result, they were not necessarily pres-ent in sufficient cell numbers to induce a therapeuticeffect. Osteoblasts are osteogenic cell types found in bonethat contribute to mineralization and bone formation(Tortelli et al., 2009; Biggs et al., 2009; Ishaug et al.,1997). However, their isolation remains challenging andhas a limited proliferative potential that further hinderstheir use in the clinical setting. The periosteum alsocontains a rich source of osteoprogenitor cells thatcontribute directly to bone regeneration (Hutmacherand Sittinger, 2003) that, however, requires improvedmethods for its purification and expansion for it to be effi-cacious (Colnot, 2009).
MSCs are multipotent cells that have the ability to differ-entiate into various cell types such as bone, cartilage,muscle, fat and other tissue types (Caplan and Dennis,2006). They can be readily isolated by plastic adherencefrom various sources such as umbilical cord blood, adiposetissue, liver and amniotic fluid, with bone marrow beingthe most well-studied source (Prockop, 1997; Pittengeret al., 1999). Ease of isolation, extensive expansion withoutdifferentiation (Haynesworth et al., 1992; Bruder et al.,1997) and stable phenotype in culture (Bruder et al.,1997; Pittenger et al., 1999) make MSCs an attractivetherapeutic cell source for BTE. Furthermore, their low im-munogenicity also suggests a potential for allergenic use intherapeutic applications (Le Blanc, 2003; Zhang et al.,
Vascularised Bone Tissue Engineering using Coculture Systems
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term
2012). The efficacy of MSCs for fracture healing has shownpromising results in various preclinical animal models(Bruder et al., 1998b; Meinel et al., 2006; Bruder et al.,1998a; Kruyt et al., 2004; Bensaid et al., 2005) and clinicalresults for the treatment of various orthopaedic diseasesand defects in BTE (Zhang et al., 2012). Currently, bonemarrow-derived MSCs (BM-MSCs) have been
benchmarked as one of the most appropriate cell sourcesdue to their efficacy in bone regeneration (Bruder andFox, 1999) and have been reported to undergo a well-de-fined osteogenic differentiation pathway (Qi et al., 2003;Frank et al., 2002). Compared to other MSCs isolated fromadipose tissue, muscle, dental pulp-derived and other peri-natal sources, BM-MSCs have been shown to exhibit weak
Figure 5. (A-B) Citation analysis on the usefulness of co-cultures in tissue and bone tissue engineering as well as leading institutionsand investigators
Y. Liu et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

osteogenic differentiation in vivo (Colnot, 2009) and to notsupport the formation of hematopoietic marrow (Robey,2011). In addition, recent identification of human fetal MSCs(hfMSCs) from blood, liver and bone marrow (Campagnolietal., 2001) showed superiority over their adult sources byexpressing higher osteogenic gene expression (Gotherstromet al., 2005), greater proliferative capacity (Guillot et al.,2007; Gotherstrom et al., 2005) andmore robust mineraliza-tion in vitro and in vivo (Zhang et al., 2009a).
Other stem cell sources such as embryonic stem cells andinduced pluripotent stem cells are currently under investiga-tion for their bone-forming abilities (Kahle et al., 2010;Gruenloh et al., 2011; Illich et al., 2011; Kuznetsov et al.,2011). Embryonic stem cells are pluripotent cell types thathave the ability to differentiate into all derivatives of thethree germ layers including bone and can propagate to largenumbers indefinitely. However, these cells pose a clinicalconcern relating to tumorigenesis (Fong et al., 2010) andimmunorejection (Grinnemo et al., 2008; Wu et al., 2008;Drukker and Benvenisty, 2004; Zhao et al., 2011). Currently,there are many potential cell sources that require more rigor-ous in vitro and in vivo investigation to verify their osteogenicabilities and safety prior to use in a clinical setting (Robey,2011; Zhang et al., 2012; Szpalski et al., 2012).
Vasculogenic (endothelial) cells. In vascularized tissueengineering, the choice of endothelial cell (EC) type forprevascularization of grafts is crucial. Mature ECs have beentraditionally thought to be responsible for postnatalangiogenesis by which capillaries sprout from pre-existingstructures (D’Amore and Thompson, 1987) until recent iden-tification of peripheral blood-derived endothelial progenitorcells (PB-EPCs) by Asahara et al. (1997) were found to be10 times more proliferative than human umbilical veinendothelial cells (HUVECs) (Murohara, 2010). PB-EPCs havebeen shown to increase vascularization in diseases such ashindlimb ischemia (Tateishi-Yuyama et al., 2002) andmyocardial infarction (Dimmeler and Zeiher, 2009) bycontributing to fracture healing by neovasculogenesis(Atesok et al., 2010; Rozen et al., 2009). Transplantation ofEPC grafts into immunodeficient mice resulted in the forma-tion of chimeric blood vessels interspersed with red bloodcells compared to none observed in EC grafts (Yoder et al.,2007). Furthermore, mature ECs have been shown to bephenotypically and functionally heterogeneous cell typesdepending on the origin of isolation and the endothelium,which resulted in difficulties in experimental comparisons(Aird, 2007) compared to the well-defined circulating popu-lation of EPCs (Masuda et al., 2011).
The two most commonly explored sources of EPCs areadult PB and umbilical cord blood (UCB), and showedthat the more primitive perinatally-derived UCB-EPCshad superior expansion and differentiation abilities com-pared to their adult counterparts (Murohara, 2010).Ingram et al. (2004) reported greater proliferative abili-ties with at least 100 population doublings from UCB-EPCs, while maintaining high levels of telomerase activitycompared to 20-30 population doublings in PB-EPCs. UCB-EPCs formed larger colonies that emerged at an earlier time
point according to isolation. They also underwent fasterdifferentiation (Kalka et al., 2000) and formed capillary-likestructures faster on Matrigel™ substrate compared to theiradult counterparts (Ingram et al. 2004) (Table 3).
Apart from promoting therapeutic neovascularization,the contribution of EPCs in BTE applications has also beendemonstrated by several recent studies. Liu et al. (2012)showed the secretion of osteogenic soluble factors suchas BMP-1, 2, 3, 6, 7 and 8 and TGF-bs by EPCs, afterwhich their use as conditioned media combined withbone inductive media significantly enhanced osteogenicdifferentiation of MSCs. In a rodent tibia fracture anddistraction osteogenesis model, it was suggested thatEPCs participated in neovascularization and subsequentbone repair as a result of temporal coincidences ofincreased EPC mobilization from bone marrow followedby homing at the site of injury in fracture healing anddistraction/consolidation models, respectively (Lee et al.,2008). Matsumoto et al. (2006; 2008) and Mifune et al.(2008) also showed functional fracture healing by PBEPC-derived vasculogenesis (Mifune et al. 2008). Morerecently, Rozen et al. (2009) demonstrated the formationof dense woven bone in a critical size defect zone in sheeptibiae 12weeks after autologous EPC implantation, whileAtesok et al. (2010) showed complete union of a ratfemoral defect within 10weeks. These studies showedthe therapeutic efficacy of EPCs in fracture repair throughneovasculogenesis.
3.1.2. Media
The choice of culture media should be carefully consid-ered in the maintenance of cellular viability of both celltypes as well as tissue development. Optimal conditionsfor inducing both vessel network and bone formationwithout negative cellular differentiation of the other celltype should be identified. In addition, the state of cellulardifferentiation prior to implantation should also beconsidered. It has been reported that pre-differentiationof osteoblast-like cells in bone inductive media prior toimplantation accelerated rate of bone formation in vivocompared to undifferentiated MSCs (Breitbart et al.,1998; Bruder and Fox, 1999). In addition, a recent inves-tigation by Schantz et al. (2012) compared the efficacyof tissue formation of undifferentiated MSCs and osteo-blast-loaded scaffolds in a rabbit calvarial defect. Resultsrevealed the importance of directing MSCs towards a spe-cific lineage prior to implantation; otherwise, their cellfate could likely have been determined by their immediatehost environment. However, it is imperative to keep inmind that the differentiated state of MSCs could interferewith their role as pericytes for wrapping and stabilisingthe pre-forming vascular network.
Most studies have investigated MSC co-culture systems inone media type only, often without justification for thechoice ofmedia used, while other studies used a combinationof culture media optimal for different cell types. Table 4shows the various media types used in current co-culturesystems for BTE. However, the heterogeneity of the
Vascularised Bone Tissue Engineering using Coculture Systems
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

experimental parameters in these studies makes comparisonbetween co-culture systems challenging. In a recent head-to-head study of various media types (bone media with/with-out endothelial supplements and endothelial-based mediawith/without osteoinductive supplements) for MSC/EPCco-culture, a general trend towards enhanced vessel
formation and more robust mineralization was noted whenendothelial growth and bone media were used, respectively.Kolbe et al. (2011) showed increased microvessel formationefficiency when MSCs were cultured in EGM2 compared tobone media with and without growth factor supplements,while the use of bone media was found to demonstratehigher osteogenic differentiation capacity of MSCs comparedto other media types. Few studies have used bone inductivemedia for co-cultures due to poor endothelial survival. Thisis, however, in conflict with a recent finding by Liu et al.(2012) that showed support of an in vitro vessel networkstructure within the 3D osteogenic graft, suggesting that boneinductive media alone was able to adequately support endo-thelial survival and distribution in the short term, even in theabsence of angiogenic supplements (Fig. 6). They also showedthat bone inductive media were most effective towards induc-ing osteogenic differentiation of MSCs, while there was a lackof osteogenic modulation in EGM variants. While growth fac-tors supplemented in endothelial growthmedia can effectivelypromote endothelial differentiation, care must be taken to en-sure that osteogenic differentiation is not inhibited. As a resultand depending on the tissue, the choice of media and varioussoluble components such as angiogenic growth factors and
Table 3. Characteristics of endothelial cells and theirprogenitors
Cell type EPCs ECs
Progenitor; endotheliallineage-directed
Mature
Source • Cord blood • Umbilical vein• Peripheral blood • Skin• Bone marrow • Adipose tissue• Monocytes
Existence Circulating in blood Lining interiorsurface of bloodvessels and lymphatics
Proliferation rate " -
Vessel formingefficiency
" -
Table 4. Maintenance of co-culture systems in BTE includingmedia used, co-culture ratios and seedingmethodologywith co-culture indirect contact unless otherwise stated
Co-culture media Cell types References
IMDM-based media MSC, EC (Villars et al. 2000)MSC, HUVEC (Villars et al. 2002)OB, HUVEC (Guillotin et al. 2008)OB, HUVEC (Hofmann et al. 2008)MSC, HUVEC (Grellier et al. 2009b)
MSC, EPC (Aguirre et al. 2010)
MesenCult-based media MSC, EPC (Henrich et al. 2009)MSC, EPC (Seebach et al. 2010)
EGM-based media MG-63 or OB, HDMEC (Unger et al. 2007)MSC, EPC (Fuchs et al. 2009a)OB, EPC (Fuchs et al. 2009b)MSC, EC (Xue et al. 2009)OB, EPC (Dohle et al. 2010)
Bone media MSC, EPC in BM (Liu et al. 2012)
Separately cultured or in mixed media MSC, EC in EGM2: DMEM10 (1:1) (Kaigler et al. 2005)MSC, HUVEC in EBM2, BM, EBM2:BM (1:1) (Rouwkema et al. 2006)MG-63 or OB, EPC in EGM-2with or without osteogenic supplements
(Fuchs et al. 2007)
MSC predifferentiated in BM,EPC in EGM2 individually culturedand implanted
(Usami et al. 2009)
MSC, EPC in aMEM15:EGM20 (1:1) (Geuze et al. 2009)MSC predifferentiated in BM, thenin co-culture with EC in EGM2
(Tsigkou et al. 2010)
MSC, HUVEC in IMDM, Medium 199 (M199),DMEM and M199:DMEM (1:1)
(Bidarra et al. 2011)
MSCs predifferentiated in BM, then co-cultured with HUVEC grown in EBM with VEGF-A and bFGF
(Koob et al. 2011)
MSC, EPC in EBM2, EGM2 with or withoutosteogenic supplements, BM with or withoutendothelial supplements
(Kolbe et al. 2011)
Legend: MSCs, mesenchymal stem cells; ECs, endothelial cells; OB, osteoblast; MG-63, osteosarcoma cell line; HDMEC, human dermal microvas-cular EC; BM, bone media; EGM, endothelial growth media; IMDM, Iscove’s Modified Dulbecco’s Media; DMEM, Dulbecco’s Modified Eagle’sMedium; aMEM, Alpha Minimum Essential Medium; Figure behind media type denotes % fetal bovine serum
Y. Liu et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

bone inductive components plays an important role in deter-mining cellular survival and differentiation.
3.1.3. Seeding methodology
There are several different cell-seeding methodologies for3D scaffold constructs, although an optimal methodologyhas yet to be determined. Some factors that need to beconsidered include how cells are cultured prior to implan-tation, whether cells should be combined or culturedapart in their optimal culture media, and whether adynamic culture system should be used and for howlong. Other concerns relate to whether cells should bemixed and what seeding ratios should be used to allowfor efficient cellular interactions and crosstalk. Table 5summarizes the possible advantages and disadvantagesassociated with various seeding methodologies, keepingin mind other considerations discussed in this review.
Future studies should consider the generation of patternedco-cultures to allow for more precise spatial control of celldistribution and cell-cell inter-actions. Techniques forgenerating patterned co-cultures include soft lithogra-phy, dielectrophoresis-based patterning and switchablesurface-based patterning on 2D layering systems (Kajiet al., 2011). The use of 3D patterned co-cultures is likelyto result in a more homogenously distributed vascular net-work within the osteogenic graft, thereby maximisingheterotypic interactions and influence of soluble factors.
3.1.4. Extracellular matrix
As to seeding methodologies onto a scaffold construct, amatrix is sometimes used for retaining cells within theporous space in the scaffold for high efficiency seeding. Theuse of extracellular matrix (ECM) in combination withscaffolds could play a critical role in orientating growth
Figure 6. GFP-labelled UCB-EPCs showed high levels of survival among RFP-hfMSCs when co-cultured in bone inductive media, whichlater extended into long tube-like structures by Day 7. Light microscopy images showed extensive mineralization of the co-cultures onDay 14
Table 5. Advantages and disadvantages of various seeding methodologies of co-cultures in vascularized BTE
Seedingmethodology Advantages Disadvantages Author, year, references
Combiningco-culture directly
• Logistically simple • Need to ensure cellsurvival in the samemedia type prior toseeding without alterationto monoculture cellularproperties
1. (Unger et al. 2007) 10.(Melero-Martin et al. 2008)2. (Unger et al. 2010) 11. (Fuchs et al. 2009b)• Even cellular mix
distributed withinthe construct
12. (Rouwkema et al. 2009)3. (Hofmann et al. 2008)13. (Koob et al. 2011)4. (Fuchs et al. 2007)14. (Liu et al. 2012)5. (Henrich et al. 2009)
6. (Seebach et al. 2010)7. (Villars et al. 2002)(Guillotin et al. 2008)8. (Geuze et al. 2009)9. (Bidarra et al. 2011)
2-step protocol • Allows monoculturesto develop optimally intheir respective growthmedia, maintainingindividual phenotypiccharacteristics
• Uneven distribution ofcells, where EPCs are mostlyattached to scaffold surfacesrather than within thescaffold core
1. (Dohle et al. 2010)2. (Fuchs et al. 2009b)3. (Tsigkou et al. 2010)(A) Seeding OB firs,
then EPC on samescaffold(B) Seeding OBand EPC ondifferent scaffoldsthat can be thencombined together
• Prevascularises the EPC zonesof the scaffold only and mightbe difficult for vessels toinfiltrate if osteogenic graftis highly confluent
1. (Usami et al. 2009)
• Minimal EPC/MSC interactionsbetween co-cultures
Co-culturefollowed bydynamic stimulation
• Improves metabolicand osteogenic activity
• Logistically more challenging 1. (Grellier et al. 2009b)
• Assists in maturation of graft
Vascularised Bone Tissue Engineering using Coculture Systems
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

factors for regulating cellular signalling, adhesion andsurvival. However, we need to consider the use of an appro-priate matrix that could block cell-cell interactions betweenthe two cell types. Some studies have usedMatrigel to immo-bilise co-cultures (Melero-Martin et al., 2008; Fuchs et al.,2009a; Rouwkema et al., 2009; Koob et al., 2011). However,they did not effectively investigate the performance of theco-culture and its crosstalk effects due to the growth factorenvironment of the Matrigel that masked the effects of theco-culture. Furthermore, Matrigel is a matrix derived froma basement membrane matrix generated by a mouse tumourcell-line, thereby making its usefulness in a clinical settingless feasible (Kleinman et al., 1982). Conversely, the use offibrin has been clinically approved and widely used invarious clinical settings. It is derived from components fromnormal wound healing and has desirable material propertiesfor wound healing, biodegradability and biocompatibility(Sierra, 1993) and is considered a possible alternative as anECM substratum in future co-culture systems.
3.1.5. Static and dynamic culture
Cell-based BTE approaches have used bonemarrow aspiratesthat were either infused directly without culture or culture-expanded for harvesting of MSCs and predifferentiated priorto transplantation (Salter et al., 2011). For culture expansionof MSCs, the use of a bioreactor is beneficial for ensuringhomogenous cellular distribution and efficient mass trans-port within the scaffold prior to implantation. Furthermore,the mechanical cues involved in onset of several signallingpathways present during normal physiologically conditionssuch as bone formation, development and homeostasis, havebeen known to positively influence the osteogenic propertiesof the graft (McCoy and O’Brien, 2010). We and others havedemonstrated that the dynamic culture of MSC-based tissue-engineered grafts was associated with improved cell survivalwithin the grafts and enhanced mineralization both in vitroand in vivo (Zhang et al., 2009b; Zhang et al., 2010a; Meinelet al., 2005). The choice of different dynamic systems canplay a part in optimal seeding, proliferation and differentia-tion of cells (Zhang et al., 2010b).
Several studies have shown the successful use of co-cul-tures of osteoprogenitor-like endothelial cells for fracturerepair in an orthotopic femoral defect model in the absenceof mechanical stimulation (Seebach et al., 2010; Henrichet al., 2009; Grellier et al., 2009b). However, results onlyfulfilled short- and medium-term goals in terms of bone for-mation, thereby possibly limiting late stage bone formationand remodelling in the long term. To date, Grellier et al.(2009b) are the only authors to have reported a dynamicculture for cell-encapsulated alginate microspheres prior tosubsequent assays by comparing the co-culture of humanosteoprogenitors and HUVECs to its monocultures. Therehave been no studies on the influence of dynamic cultureon osteoblast-endothelial cell co-cultures for vascularizedBTE. Due to the mechano-sensitive nature of both osteoblas-tic and endothelial cell types and their associated physiol-ogy, we proposed biomechanical stimulation of co-culturesystems to promote both endothelial (Tzima et al., 2005)
and osteogenic differentiation (Sikavitsas et al., 2003;Holtorf et al., 2006) towards more robust vascular and bonedevelopment and remodelling. The duration of thesein vitro pre-cultivations required careful consideration.
3.1.6. Scaffolds
3D scaffolds have been used in BTE to provide a localenvironment for cells and to serve as a temporary matrixto support and direct cellular growth and differentiation(Langer and Vacanti, 1993) as well as provide mechano-induction for expression of growth factors and ultimatelybone remodeling. Particularly for bone, an ideal scaffoldshould be mechanically stable to temporarily support thedefect zone while resorbing at a rate in tandem withnew bone formation in the defect region (Hutmacher,2000). In tissue remodelling, cells are able to recognisethe architectural feature of the environment and differen-tiate accordingly. The structural characteristics of thescaffold are important in guiding cellular distributionand organization and are thus designed to closely mimicthe in vivo environment of the tissue. The central spongyregion is trabecular in structure while the osteons in theouter cortical zones are supported by an extensivenetwork of vasculature. A trabecular microarchitectureand porous network are necessary for support of vascularingrowth, new tissue formation and pathways for masstransport (Fig. 7) (Muschler et al., 2004). Most impor-tantly, the biomaterial should interact with the host tissueand exert a desirable cellular response that will assist intissue repair (Lanza et al., 2007).
Fundamental scaffold design factors to consider includematerial type, mechanical strength, pore size, ease of manu-facturability, surface chemistry and spatial properties(Table 6) (Hutmacher, 2000).
3.1.7. Microenvironment (oxygen tension)
A bone fracture results in the disruption of the vascularsupply and leads to hypoxia in the defect zone and upregula-tion of various angiogenic genes such as VEGF (Fig. 2).Experimentally, the role of HIF-1a and its activated pathwayhave been shown to aid in skeletal development by stimulat-ing osteoprogenitor differentiation and promoting robustbone formation and remodelling in combination with angio-genic mechanisms (Wan et al., 2010). Wang et al. (2007)showed robust bone formation accompanied by VEGF upre-gulation following constitutive activation of the HIF-1apathway in osteoblasts, while the inhibition of HIF-1a ledto the formation of narrow and less vascularised bones. Asimilar improvement in vascularity and bone formation wasobserved in distraction osteogenesis mice following vonHippel-Lindau deletion with constitutive HIF-1a activationcompared tomice lacking HIF-1a (Wan et al., 2008; Komatsuand Hadjiargyrou, 2004). Additionally, lentivirally-drivenoverexpression of HIF-1a in bone marrow-derived MSCs forthe repair of a rat calvarial CSD defect showed enhancedvessel formation and bony repair compared to its lacZ control(Zou et al., 2011).
Y. Liu et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

While various studies have shown enhanced efficacy of theangiogenic and osteogenic responses of endothelial (Fig. 8)and osteoprogenitor monocultures, respectively, followinghypoxic stimulus, few studies have investigated the effectsof lowoxygen tensions on a co-culture system. A recent studyinvestigated the effect of hypoxic cultures on MSC/EPCsbased on the hypothesis that a hypoxic environment contrib-uted to prevascularization. Results, however, showed animpediment of vasculogenesis following hypoxia stimulationcompared to normoxic cultures, which supported the forma-tion of prevascularized structures (Gawlitta et al., 2012).Additional studies are required to confirm the importanceof a hypoxic stimulus for prevascularizing tissue-engineeredco-culture constructs towards facilitating bone regenerationand fracture repair response in vivo.
3.1.8. Animal model
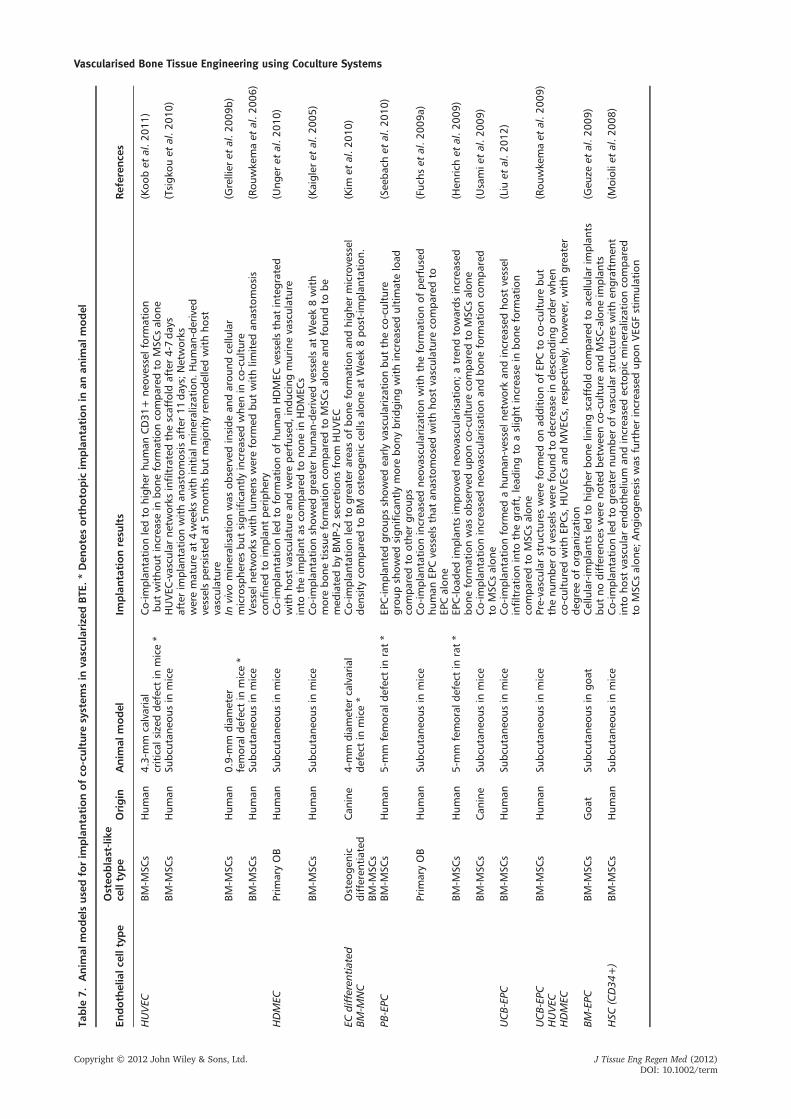
Most co-culture studies have examined co-culture graftsin subcutaneous implantation models, with a few studies
focusing on orthotopic bone repair models (Table 7).However, not all studies showed an accompanyingincrease in bone formation following increased vasculari-sation on co-culture. Usami et al. (2009) and Liu et al.(2012) showed minor increases in ectopic bone formationand higher densities in capillary growth in EPC/MSCsover MSC constructs when implanted in a nude mousemodel. Seebach et al. (2010) showed significant improve-ments in bone formation with enhanced early angiogene-sis in human-EPC/MSC groups in a rat CSD. In contrast,Koob et al. (2011) did not find any improvement in boneregeneration when HUVECs were used in co-culture withMSCs despite triggering neovascularisation, while MSCsalone supported bone formation in a calvarial CSD micemodel with higher bone content compared to its optimalco-culture ratio group implanted in an intramuscular goatmodel, albeit with no statistical significance.
It was difficult to compare the outcomes of these differentstudies because of the use of different animal models andspecies with different types of bone bearing healing potentials
Table 6. Basic criteria and considerations for designing a scaffold for use in BTE applications (Salgado et al., 2004; Hutmacher,2000)
Criteria Feature Function
Biocompatibility Good integration with host tissue Does not elicit immune response
Microarchitecture Interconnected scaffold strutsthat comply to applied stresses
Uniformly distributes stresses throughoutscaffold
Porosity Large surface area: volume, ideallywith pore size ranging between200-900mm
Allows cell in-growth, neovascularisation,sufficient mass transport and osteogenesis
Surface properties Appropriate chemical andtopographical properties
Affects cellular adhesion, proliferation,protein interactions and osteoconduction
Osteoinductivity Osteoinductive properties Recruits and differentiates osteoprogenitorsto the defect region
Mechanical properties Sufficient mechanical strengthappropriately tuned with porosity
Withstands loading forces in vivo, withstandshydrostatic pressure, and allows for cellularin-growth and matrix production
Biodegradability Biodegradable at an appropriaterate to match growth of neotissue
Temporarily supports the defect zone as scaffoldgets replaced by new bone
Figure 7. Porous trabecular-like scaffold (polycaprolactone/tricalcium phosphate) with a high porosity that enabled vascular infiltration,mass transport and new tissue formation
Vascularised Bone Tissue Engineering using Coculture Systems
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

(Horner et al., 2010). For example, it was shown that tibialperiosteum had greater reparative efficiency compared tocalvarial periosteum (Fujii et al., 2006), while the ratcalvaria has been described as a hostile environment for bonehealing due to poor blood supply and little bone marrow(Conn, 2008). Of significance, most studies examinedectopically implanted implants in rodents in which implantswere in close proximity to neighbouring vascularized subder-mal tissues. In orthotopic rodent models, the small depth ofdefects allowed an adequate exchange of oxygen and nutri-ents. Hence, these small animal studies did not accuratelyreflect the performance of implanted grafts and did not an-swer our questions regarding vascularization. Instead, they of-fered a simple model for optimizing various parameters suchas scaffold biocompatibility, osteoconductivity, extent of bloodvessel formation and osteogenic potential of the graft in vivo.
To address the limitations of mass transport and vascularinsufficiency in thick grafts, it would be of greater relevanceto consider models of large animals (sheep, goat, pigs),where diffusion distances are clinically relevant (Muschleret al., 2010). This would allow investigation of co-culturegraft efficacy and biomechanical related issues that moreclosely mimic that of the human skeletal system and its clin-ical relevance (Conn, 2008). These models would also allowlong-term monitoring to better track structural changes andhelp understand the performance of tissue-engineered graftsfrom the formation of woven to lamellar bone. As a result,the choice of suitable animal models that use long-termimplantation studies need to be carefully selected to trackthe structural changes of bone and to test the mechanicalstrength of newly formed compared to natural bone tissueto more accurately determine the efficacy and clinical feasi-bility of co-cultured bone grafts. Various factors that influ-ence the quality and quantity of bone healing include speciesand age of the animal model, type and size of defect, site ofimplantation, presence of periosteum, soft tissue and vascu-larisation, diet of the animal, appropriate animal modelsand comparisons of bone properties between different ani-mal species (Reichert et al., 2009). These factors should beconsidered as more accurate predictors of the efficacy ofBTE-grafts when choosing a preclinical animal model (Conn2008; Horner et al., 2010; O’Loughlin et al., 2008; Reichertet al., 2009). Conducting animal studies under regulatoryapproved animal facilities is also important towards translat-ing these studies to clinical applications.
4. Future directions
4.1. Quality of vessels formed
4.1.1. Current co-culture systems
Most studies have focused on observations of increasedvascularization following addition of endothelial lineagesinto osteoprogenitor-like co-cultures and showed immuno-chemical staining of endothelial markers, with some studiesshowing evidence of integration and chimeric vessel forma-tion (Melero-Martin et al., 2008; Fuchs et al., 2009a; Ungeret al., 2010; Rouwkema et al., 2009). These studies, however,showed little evidence of the integrity and functionality ofthe vessels within the grafts. A few studies reported acorresponding increase in bone formation due to increasedvascularization in co-culture following in vivo implantation(Seebach et al., 2010; Usami et al., 2009; Liu et al., 2012),while others showed functional improvements in boneformation (Koob et al., 2011; Geuze et al., 2009). Theseconflicting data raise questions about the functional efficacyof in vitro prevascularization. How significant are the contri-butions of in vitro prevascularization in enhancing boneformation and how can we efficiently improve the qualityand functionality of these vessels?
4.1.2. Need for advanced imaging methodologies
To assist in our understanding of the efficacy of theseco-culture approaches, real-time tracking of vascularbehaviour within osteogenic grafts through the use ofadvanced imaging methodologies is essential for under-standing the various features of the microvasculaturein vivo. For example, intravital microscopy (IVM) allowsfor continuous non-invasive imaging of both quantitativeand qualitative aspects of angiogenesis in live animalsvia a “window” chamber (Jain et al., 2002; Menger andLehr, 1993). Microcirculation of the host is visible follow-ing IV injection of fluorescently labelled molecules (Jainet al., 2002), while monitoring the time course dynamicsof origin-specific endothelial cells to vessel networkformation within the graft. The contributions of bothrecruited endothelial cells and pre-formed in vitro vascu-lature as well as their timing of anastomosis can be
Figure 8. In vitro study of HUVECs cultured in 2% O2 during differentiation on Matrigel showed more extensive vessel-forming abilityand exhibited 1.7 fold higher branch points compared to<21% O2 conditions (p<0.001).
Y. Liu et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term
Table
7.Anim
almodelsusedforim
plantationofco
-culture
system
sin
vascularize
dBTE
.*Den
otesorthoto
pic
implantationin
anan
imal
model
Endoth
elialc
elltyp
eOsteo
blast-like
celltype
Origin
Anim
almodel
Implantationresu
lts
Reference
s
HUVEC
BM-M
SCs
Hum
an4.3-mm
calvarial
critical
sizedde
fect
inmice*
Co-im
plan
tation
ledto
high
erhu
man
CD31
+ne
ovesselformation
butwitho
utincrea
sein
bone
form
ationco
mpa
redto
MSC
salon
e(Koo
bet
al.2
011)
BM-M
SCs
Hum
anSu
bcutan
eous
inmice
HUVEC
-vascu
larne
tworks
infiltratedthescaffold
after4-7da
ysafterim
plan
tation
withan
astomosisafter11
days;N
etworks
werematureat
4wee
kswithinitialm
ineralization.
Hum
an-derived
vesselspe
rsistedat
5mon
thsbu
tmajorityremod
elledwithho
stvasculature
(Tsigk
ouet
al.2
010)
BM-M
SCs
Hum
an0.9-mm
diam
eter
femoral
defect
inmice*
Invivo
mineralisationwas
observed
inside
andarou
ndcellu
lar
microsphe
resbu
tsign
ificantly
increa
sedwhe
nin
co-culture
(Grellier
etal.2
009b
)
BM-M
SCs
Hum
anSu
bcutan
eous
inmice
Vesseln
etworks
withlumen
swereform
edbu
twithlim
ited
anastomosis
confi
nedto
implan
tpe
riph
ery
(Rou
wke
maet
al.2
006)
HDMEC
Prim
aryOB
Hum
anSu
bcutan
eous
inmice
Co-im
plan
tation
ledto
form
ationof
human
HDMEC
vesselsthat
integrated
withho
stvasculaturean
dwerepe
rfused
,ind
ucingmurinevasculature
into
theim
plan
tas
compa
redto
none
inHDMEC
s
(Ung
eret
al.2
010)
BM-M
SCs
Hum
anSu
bcutan
eous
inmice
Co-im
plan
tation
show
edgrea
terhu
man
-derived
vesselsat
Wee
k8with
morebo
netissue
form
ationco
mpa
redto
MSC
salon
ean
dfoun
dto
bemed
iatedby
BMP-2secretions
from
HUVEC
(Kaigler
etal.2
005)
ECdifferen
tiated
BM-M
NC
Osteo
genic
differen
tiated
BM-M
SCs
Can
ine
4-mm
diam
eter
calvarial
defect
inmice*
Co-im
plan
tation
ledto
grea
terarea
sof
bone
form
ationan
dhigh
ermicrovessel
densityco
mpa
redto
BMosteog
enic
cells
alon
eat
Wee
k8po
st-im
plan
tation
.(Kim
etal.2
010)
PB-EPC
BM-M
SCs
Hum
an5-mm
femoral
defect
inrat*
EPC-im
plan
tedgrou
psshow
edea
rlyvascularizationbu
ttheco
-culture
grou
pshow
edsign
ificantly
morebo
nybridging
withincrea
sedultimateload
compa
redto
othe
rgrou
ps
(See
bach
etal.2
010)
Prim
aryOB
Hum
anSu
bcutan
eous
inmice
Co-im
plan
tation
increa
sedne
ovascu
larization
withtheform
ationof
perfused
human
EPCvesselsthat
anastomosed
withho
stvasculatureco
mpa
redto
EPCalon
e
(Fuc
hset
al.2
009a
)
BM-M
SCs
Hum
an5-mm
femoral
defect
inrat*
EPC-lo
aded
implan
tsim
prov
edne
ovascu
larisation
;atren
dtowards
increa
sed
bone
form
ationwas
observed
upon
co-culture
compa
redto
MSC
salon
e(Hen
rich
etal.2
009)
BM-M
SCs
Can
ine
Subc
utan
eous
inmice
Co-im
plan
tation
increa
sedne
ovascu
larisation
andbo
neform
ationco
mpa
red
toMSC
salon
e(U
samie
tal.2
009)
UCB-EP
CBM
-MSC
sHum
anSu
bcutan
eous
inmice
Co-im
plan
tation
form
edahu
man
-vesseln
etworkan
dincrea
sedho
stvessel
infiltration
into
thegraft,lead
ingto
aslight
increa
sein
bone
form
ation
compa
redto
MSC
salon
e
(Liu
etal.2
012)
UCB-EP
CBM
-MSC
sHum
anSu
bcutan
eous
inmice
Pre-vascular
structures
wereform
edon
addition
ofEP
Cto
co-culture
but
thenu
mbe
rof
vesselswerefoun
dto
decrea
sein
descen
ding
orde
rwhe
nco
-culturedwithEP
Cs,HUVEC
san
dMVEC
s,resp
ectively,h
owever,w
ithgrea
ter
degree
oforga
nization
(Rou
wke
maet
al.2
009)
HUVEC
HDMEC
BM-EPC
BM-M
SCs
Goa
tSu
bcutan
eous
ingo
atCellular-im
plan
tsledto
high
erbo
nelin
ingscaffold
compa
redto
acellularim
plan
tsbu
tno
differen
ceswereno
tedbe
twee
nco
-culture
andMSC
-alone
implan
ts(G
euze
etal.2
009)
HSC
(CD34
+)
BM-M
SCs
Hum
anSu
bcutan
eous
inmice
Co-im
plan
tation
ledto
grea
ternu
mbe
rof
vascular
structures
withen
graftm
ent
into
host
vascular
endo
thelium
andincrea
sedectopicmineralizationco
mpa
red
toMSC
salon
e;Ang
ioge
nesiswas
furthe
rincrea
sedup
onVEG
Fstim
ulation
(Moiolie
tal.2
008)
Vascularised Bone Tissue Engineering using Coculture Systems
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

tracked. High-resolution imaging also allows assessmentof functionality through observation of vessel morphology(diameter, length, inter-capillary distance, branchingpatterns and tortuosity) and microhemodynamic vari-ables (vascular permeability, blood cells velocity andother blood flow related parameters) as well as cell-celland cell-biomaterial interactions, which often requiretissue removal. An experimental model for using IVM forimaging provides molecular, cellular, phenotypical andfunctional perspectives of the vasculature within thepremineralizing bone graft of the co-culture system isessential for improving strategies in vascularized regener-ative medicine and tissue engineering. A recent studyreported functional anastomosis of human EPC/MSCvessel networks with the host mouse following infusionof species-specific lectin. After disconnecting the pre-formed vessel network and transplanting it into a second-ary mouse, reperfusion was restored within 3 days (Kanget al., 2011), suggesting the possibility of generatingfunctional prevascularized networks through co-cultures.
4.1.3. Formation of stabilized vessels
Timely maturation of vessels is necessary for preventingregression of leaky and disorganized vessels and subsequentfailure, while establishing anastomosis with the host circula-tion (Jain et al. 2005). Various factors such as angiogenin-1have been reported for their contributions to creating stabi-lised vessels (Carmeliet and Jain, 2000). Future co-culturestudies relating to the stability and perfusibility of thesevessels and their subsequent effects on bone formation needto be considered.
4.2. Quality of bone formed
4.2.1. Current co-culture systems
In current studies of co-culture systems, evidence of boneformation is often limited to early-stage mineralized andwoven bone while the study of late-stage bone is oftenneglected. The occurrence of osteogenesis in vitro is mostoften demonstrated through quantitative alkaline phospha-tase (ALP) activity measurement and gene expression ofvarious osteogenic markers such as ALP, Type 1 collagen,RunX2, osteocalcin and osteopontin. Following in vivo trans-lation, mineral deposition within these co-culture grafts isoften visually traced through von Kossa and Alizarin Redstains, with mature osteoblast morphologically identifiedthrough histology and immunochemical staining of osteogenicmarkers. Most measurements are coarsely quantitated by pixelmeasurements of the bone area,which is least suggestive of thevolume, mechanical properties and maturity of bone formed.
Of the co-culture studies for vascularised BTE, the study bySeebach et al. (2010) is one of the few that has convincinglyshown the extent of callus and bone formation in a critical sizefemoral rat defect at 8weeks. Mechanical tests performed onthe femurs transplanted with the tissue-engineered graftsshowed the highest loading in the co-cultures. However, the
lack of serial time point imaging limited the understandingof progressive vascular formation and bone healing. In an-other study, Grellier et al. (2009a, 2009b) showed intensemineralization in alginate encapsulated co-cultures at 3weekscompared to osteoprogenitor cells when imaged on a micro-CT X-ray. While conventional radiography imaging remainsthe primary approach for assessing fracture healing (Panjabiet al., 1989), it lacks resolution for visualizing and distinguish-ing fine structures during bone healing. To date, most co-cul-ture studies have been short-term investigations that haveneglected bone remodelling and quality in the long term.
4.2.2. Need for advanced imaging tools
To detect early changes during the fracture repair processsuch as callus formation and mineralization, the use of non-invasive time-scale imaging volumetric quantitative com-puted tomography and high-resolution magnetic resonanceimaging could allow better assessment of the efficacy andperformance of grafts (Kalpakcioglu et al., 2008). Of equalimportance is cross-sectional bone visualization to determinethe structural formation of the macrostructure (geometry)and microstructure (trabecular volume, trabecular spacingand connectivity) characteristics and density of bone through3D imaging and reconstructed view of the entire graft withinits orthotopic location together with the host bone. Thiscould highlight the mechanical strength of the bone formedcompared to undetected bone.
4.2.3. Use of advanced imaging tools
Random regions with emerging vessels and mineralizedtissue are oftenmeasuredwithout representative time-coursetracking of vessel morphology and bone formation in thesame location within the tissue. The use of real-time imagingcan overcome such issues and result in improved conve-nience, larger data sets and precision, as well as provide amore thorough understanding of vascular behaviour (vesselformation and regression) and vessel contributions to boneformation in both short and long terms. The same regionscan also be traced over serial time-points when a later fol-low-up is required. Furthermore, the use of real-time imagingtools can overcome current issues of fixed time-point animalharvests that lack scientific justification due to poor under-standing of time-related vascularization events. This is essen-tial when it comes to choosing suitable time-points for analy-sis, where animal numbers are often increased in each studygroup to reduce large variations between different animals;thereby contributing to increased costs, labour and unwar-ranted sacrifice of animal subjects. The use of advanced im-aging tools to track individual graft performance could beuseful for better understanding co-culture systems as we ap-ply these strategies to the clinical setting.
5. The future; conclusions
The progress of co-culture systems in traditional vascularizedtissue engineering has shown significant potential for
Y. Liu et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

improving neovascularization following the addition ofendothelial lineages, with some studies showing an accom-panying increase in bone formation. However, there remainmany challenges for tissue engineers in the mimicry of natu-ral bone tissue, and there is still much to be investigatedregarding optimal conditions for mediating effective genera-tion of large vascularized bone grafts before BTE grafts canbe successfully translated into clinical settings.
One challenge with current co-culture systems involvesthe spatial distribution of different cell types in a multicellu-lar bone tissue. In vascularized BTE, we hypothesised the for-mation of an organised vascular network with patterned cellseeding for improved cell-cell interactions and long-termfunctionality of large bone grafts. We used decellularizedvascularized bone constructs as a template subsequentlyloaded with endothelial cell monolayers through theirretained vessel networks surrounded by a customized mouldof highly osteogenic cells.
5.1. Bioprinting of 3D grafts
To accomplish this, the recent introduction of enablingtechnologies such as bioprinting of biomimetic organs offerssignificant promise as a feasible alternative strategy forsuccessful tissue engineering and regenerative medicine.Briefly, bioprinting is the printing of biological material suchas cells, growth factors and ECMs based on similar principlesas rapid prototyping. Recently, Atala et al. reported thesuccessful bioprinting of an experimental 3D kidneyprototype that offers hope to any patient on the waiting listfor kidney transplantation in Britain (BBC, 2012). For BTE,we anticipate new successful strategies for patients with largebone defects by leveraging bioprinting technology to generatea scaffoldless vascularized bone grafts to more closely mimicthe architecture and cellular distribution within naturalvascularized bone. For example, Moon et al. (2010)developed a platform that allowed printing of multilayered3D collagen-encapsulated cell patches with uniform cellseeding at regular intervals on a substrate that maintainedlong-term cell viability. More recently, Ker et al. (2011b)showed that functionalizing polystyrene fibres bioprintedwith FGF-2 and BMP-2 promoted tendon and bonedifferentiation, respectively. This method suggests the abilityof stem cell differentiation and distribution in a spatiallyoriented manner following bioprinting of growth factor-inkdroplets in an orderly arrangement on the fibres. Otherstudies have suggested the use of cell-laden self-assemblinghydrogel aggregates and tissue spheroids rather thanindividual cell printing to achieve cell density andvascularization (Mironov et al., 2011). These strategiesassisted in the geometrical control of cellular alignment anddirected them towards the lineage of interest.
Similarly, we proposed the bioprinting of patternedco-cultures and growth factors such as BMP-2 and VEGFonto substrates to promote bone and endothelial differen-tiation of multipotent MSCs. This strategy allowed the useof a single multipotent cell type that could differentiateinto multiple cellular phenotypes (Ker et al., 2011a;
Ker et al., 2011b) such as bone and endothelial cells simul-taneously, thereby generating an organised vesselnetwork within a highly osteogenic bone graft. Thismethodology also avoided the potential complexitiesinvolved in the use of co-cultures, spatial cellular distribu-tion and other issues related to choice of media.
In addition, it would be ideal to incorporate bio-patternson a mechanically stiff matrix to provide temporary strengthand support to the bone defect following implantation. In arecent study by Schuurman et al. (2011), the authorsexplored the construction of a tissue graft using alternatingdeposits of thermoplastic fibres and cell-laden hydrogelconstructs that more closely mimicked natural bone, whichis comprised of hard mineral surrounded by cells embeddedin a soft organic matrix. The use of thermoplastics alsoprovided improved mechanical properties to the hydrogelsystem and could be tailored to achieve similar strengthand architecture as native tissues.
5.2. Soft microactuators
To allow assembly and fusion of cell layers for maturation ofa thick-layered graft, the use of novel soft biodegradablemicroactuators that can provide a controlled and continuousmechanical stimulus to cells could assist in enhancement ofEC and bone differentiation towards more rapid formationof vessel networks and long-term bone remodelling. Weproposed direct adhesion of these microrobots to individualcells either by electrostatic attraction or covalent forces thatprovided direct stimuli. This approach would eliminatethe time required for graft tissue maturation in a conven-tional bioreactor, hence shortening patient waiting time forgraft implantation.
5.3. Conclusions
This novel approach of using 3D graft bioprinting in conjunc-tion with CT scans could be ideal for obtaining a customisedbiomimetic graft that fits a defect region. Advanced technol-ogies could provide great convenience to clinicians andresearchers during clinical implantation since the bone graftcan be fabricated within minutes to hours with minimalpreparation and labour. This strategy could also significantlyhelp lower patient costs and reduce waiting times for defectrepair and also provide relief and improve quality of life.The proposed strategy shows great promise towards achievingsuccessful vascularized BTE and is likely to resolve the currentchallenges and complexities with current co-culture systems,thereby closing the gap in themimicry of natural bone. Strongjustification based on its feasibility and in vivo efficacy isrequired before presenting a strong case prior to clinicaltranslation of these thick bioprinted vascularized bone grafts.
Acknowledgements
JC received salary support from the National Medical ResearchCouncil in Singapore (NMRC/CSA/012/2009).
Vascularised Bone Tissue Engineering using Coculture Systems
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

References
Aguirre A, Planell JA, Engel E. 2010; Dynamicsof bonemarrow-derived endothelial progeni-tor cell/mesenchymal stem cell interaction inco-culture and its implications in angiogene-sis. Biochem Biophys Res Commun 400(2):284–291.
Ai-Aql ZS, Alagl AS, Graves DT et al. 2008;Molecular mechanisms controlling bone for-mation during fracture healing and distrac-tion osteogenesis. J Dent Res 87(2): 107–118.
AirdWC. 2007; Phenotypic heterogeneity of theendothelium: I. Structure, function, andmechanisms. Circ Res 100(2): 158–173.
Asahara T, Murohara T, Sullivan A et al. 1997;Isolation of putative progenitor endothelialcells for angiogenesis. Science 275(5302):964–967.
Atesok K, Li R, Stewart DJ et al. 2010; Endo-thelial progenitor cells promote fracturehealing in a segmental bone defect model.J Orthop Res 28(8):1007–1014.
Bae JH, Song HR, Kim HJ et al. 2011; Discon-tinuous release of bone morphogeneticprotein-2 loaded within interconnectedpores of honeycomb-like polycaprolactonescaffold promotes bone healing in a largebone defect of rabbit ulna. Tissue Eng PartA 17(19-20): 2389–2397.
BBC. 2012; Printing a human kidney. (citation.available from http://www.bbc.com/future/story/20120621-printing-a-human-kidney.
Bensaid W, Oudina K, Viateau V et al. 2005.De novo reconstruction of functional boneby tissue engineering in the metatarsalsheep model. Tissue Eng 11(5-6): 814–824.
Bidarra SJ, Barrias CC, Barbosa MA et al. 2011.Phenotypic and proliferative modulation ofhuman mesenchymal stem cells via crosstalkwith endothelial cells. Stem Cell Res 7(3):186–197.
Biggs MJ, Richards R, Gadegaard G et al. 2009;The use of nanoscale topography to modu-late the dynamics of adhesion formation inprimary osteoblasts and ERK/MAPK signal-ling in STRO-1+ enriched skeletal stem cells.Biomaterials 30(28): 5094–5103.
Boilly B, Vercoutter-Edouart AS, HondermarckH et al. 2000; FGF signals for cell prolifer-ation and migration through differentpathways. Cytokine Growth F R 11(4):295–302.
Boraiah S, Paul O, Hawkes D et al. 2009; Com-plications of recombinant human BMP-2 fortreating complex tibial plateau fractures:a preliminary report. Clin Orthop Relat R467(12): 3257–3262.
Breitbart AS, Grande DA, Kessler R et al. 1998;Tissue engineered bone repair of calvarialdefects using cultured periosteal cells. PlastReconstr Surg 101(3): 567–574; discussion575-566.
Bruder SP, Fox BS. 1999; Tissue engineeringof bone. Cell based strategies. Clin OrthopRelat Res (367 Suppl): S68–83.
Bruder SP, Jaiswal N, Haynesworth SE. 1997;Growth kinetics, self-renewal, and the osteo-genic potential of purified human mesenchy-mal stem cells during extensive subcultiva-tion and following cryopreservation. J CellBiochem 64(2): 278–294.
Bruder SP, Kraus KH, Goldberg VM et al. 1998a;The effect of implants loaded with autolo-gous mesenchymal stem cells on the healingof canine segmental bone defects. J BoneJoint Surg Am 80(7): 985–996.
Bruder SP, Kurth AA, SheaM et al. 1998b; Boneregeneration by implantation of purified,culture-expanded humanmesenchymal stemcells. Journal of orthopaedic research: officialpublication of the Orthopaedic ResearchSociety 16(2): 155–162.
Bullens PH, Schreuder HW, de Waal MalefijtMC et al. 2010; The presence of periosteumis essential for the healing of large diaphysealsegmental bone defects reconstructed withtrabecular metal: a study in the femur ofgoats. J Biomed Mater Res B Appl Biomater92(1): 24–31.
Campagnoli C, Roberts IA, Kumar S et al. 2001;Identification ofmesenchymal stem/progeni-tor cells in human first-trimester fetalblood, liver, and bone marrow. Blood 98(8):2396–2402.
Caplan AI, Dennis JE. 2006;Mesenchymal stemcells as trophic mediators. J Cell Biochem 98:1076–1084.
Carmeliet P, Jain RK. 2000; Angiogenesis incancer and other diseases. Nature 407(6801): 249–257.
Carter DR, Beaupré GS. 2001; Skeletal functionand form: mechanobiology of skeletal develop-ment, aging, and regeneration. CambridgeUniversity Press: Cambridge; New York.
Carter DR, Beaupre GS, Giori NJ et al. 1998;Mechanobiology of skeletal regeneration.Clin Orthop Relat Res (355 Suppl):S41–55.
Colnot C. 2009; Skeletal cell fate decisionswithin periosteum and bone marrow duringbone regeneration. Journal of bone andmineral research: the official journal of theAmerican Society for Bone and MineralResearch 24(2): 274–282.
Conn PM. 2008; Sourcebook of models for bio-medical research. Humana Press: Totowa, N.J..
Connolly JG, Tiedeman R, Dehne JR. 1989;Autologous marrow injection for delayedunions of the tibia: a preliminary report.J Orthop Trauma 3: 276–282.
Cool S, Jackson R, Pincus P et al. 2002; Fibro-blast growth factor receptor 4 (FGFR4)expression in newborn murine calvaria andprimary osteoblast cultures. Int J Dev Biol46(4): 519–523.
Cool SM, Nurcombe V. 2005; The osteoblast-heparan sulfate axis: control of the bonecell lineage. Int J Biochem Cell Biol 37(9):1739–1745.
Cool SM, Nurcombe V. 2006; Heparan sul-fate regulation of progenitor cell fate.J Cell Biochem 99(4): 1040–1051.
Coulon C, Georgiadou M, Roncal C et al.2010; From vessel sprouting to normaliza-tion: role of the prolyl hydroxylase domainprotein/hypoxia-inducible factor oxygen-sensing machinery. Arterioscler ThrombVasc Biol 30(12): 2331–2336.
D’Amore PA, Thompson RW. 1987; Mechan-isms of angiogenesis. Annu Rev Physiol49:453–464.
De Bari C, Dell’Accio F, Vanlauwe J et al. 2006;Mesenchymal multipotency of adult humanperiosteal cells demonstrated by single-celllineage analysis. Arthritis Rheum 54(4):1209–1221.
Deng HW, Liu YZ. 2005; Current topics in bonebiology. World Scientific: Hackensack, NJ.
Dimmeler S, Zeiher AM. 2009; Cell therapy ofacute myocardial infarction: open questions.Cardiology 113(3): 155–160.
Dohle E, Fuchs S, Kolbe M et al. 2010; Sonichedgehog promotes angiogenesis and os-teogenesis in a co-culture system consist-ing of primary osteoblasts and outgrowthendothelial cells. Tissue Eng Part A 16(4): 1235–1237.
Drukker M, Benvenisty N. 2004; The immuno-genicity of human embryonic stem-derivedcells. Trends Biotechnol 22(3): 136–141.
Ehrlich PJ, Lanyon LE. 2002; Mechanical strainand bone cell function: a review.Osteoporosisinternational: a journal established as result ofcooperation between the European Foundationfor Osteoporosis and the National OsteoporosisFoundation of the USA 13(9): 688–700.
Eriksen EF, Axelrod D, Melsen W. 1994; Bonehistomorphometry. Raven Press: New York.
Eyre-Brook AL. 1984; The periosteum: itsfunction reassessed. Clin Orthop Relat Res(189): 300–307.
Fan W, Crawford R, Xiao Y. 2010; Enhancingin vivo vascularized bone formation by cobaltchloride-treated bone marrow stromal cellsin a tissue engineered periosteum model.Biomaterials 31(13): 3580–3589.
Fong CY, Gauthaman K, Bongso A. 2010;Teratomas frompluripotent stem cells: A clin-ical hurdle. J Cell Biochem 111(4): 769–781.
Frank O, Heim M, Jakob M et al. 2002; Real-time quantitative RT-PCR analysis of humanbonemarrow stromal cells during osteogenicdifferentiation in vitro. J Cell Biochem 85(4):737–746.
Fuchs S, Ghanaati S,OrthC et al. 2009a; Contri-bution of outgrowth endothelial cells fromhuman peripheral blood on in vivo vasculari-zation of bone tissue engineered constructsbased on starch polycaprolactone scaffolds.Biomaterials 30(4): 526–534.
Fuchs S, Hofmann A, Kirkpatrick CJ. 2007;Microvessel-like structures from outgrowthendothelial cells from human peripheralblood in 2-dimensional and 3-dimensionalco-cultures with osteoblastic lineage cells.Tissue Eng 13(10): 2577–2588.
Fuchs S, Jiang X, Schmidt H et al. 2009b;Dynamic processes involved in the pre-vascu-larization of silk fibroin constructs for boneregeneration using outgrowth endothelialcells. Biomaterials 30(7): 1329–1338.
Fujii T, UenoT, Kagawa T et al. 2006; Compari-son of bone formation ingrafted periosteumharvested from tibia and calvaria. MicroscRes Tech 69(7): 580–584.
Gawlitta D, Fledderus JO, van Rijen MH et al.2012; Hypoxia impedes vasculogenesis ofin vitro engineered bone. Tissue Eng PartA 18(1-2): 208–218.
Gerber HP, Vu TH, Ryan AM et al. 1999; VEGFcouples hypertrophic cartilage remodeling,ossification and angiogenesis during endo-chondral bone formation. Nat Med 5(6):623–628.
Germain S, Monnot C, Muller L et al. 2010;Hypoxia-driven angiogenesis: role of tipcells and extracellular matrix scaffolding.Curr Opin Hematol 17(3): 245–251.
Geuze RE, Wegman FF, Oner C et al. 2009;Influence of endothelial progenitor cellsand platelet gel on tissue-engineered boneectopically in goats. Tissue Eng Part A 15(11): 3669–3677.
Gotherstrom C, West A, Liden J et al. 2005;Difference in gene expression betweenhuman fetal liver and adult bone marrow
Y. Liu et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

mesenchymal stem cells. Haematologica90(8): 1017–1026.
Grellier M, Bareille R, Bourget C et al. 2009a;Responsiveness of human bone marrowstromal cells to shear stress. J Tissue EngRegen Med 3(4): 302–309.
Grellier M, Granja PL, Fricain JC et al. 2009b;The effect of the co-immobilization ofhuman osteoprogenitors and endothelialcells within alginate microspheres on min-eralization in a bone defect. Biomaterials30(19): 3271–3278.
Grinnemo KH, Sylven C, Hovatta O et al. 2008;Immunogenicity of human embryonic stemcells. Cell Tissue Res 331(1): 67–78.
Gruenloh W, Kambal A, Sondergaard C et al.2011; Characterization and in vivo testingof mesenchymal stem cells derived fromhuman embryonic stem cells. Tissue EngPart A 17(11-12): 1517–1525.
Grundel RE, Chapman MW, Yee T et al. 1991;Autogeneic bone marrow and porousbiphasic calcium phosphate ceramic forsegmental bone defects in the canine ulna.Clin Orthop Relat Res (266): 244–258.
Guillot PV, Gotherstrom C, Chan J et al. 2007;Human first-trimester fetalMSC express plur-ipotency markers and grow faster and havelonger telomeres than adult MSC. Stem Cells25(3): 646–654.
Guillotin B, Bareille R, Bourget C et al. 2008;Interaction between human umbilical veinendothelial cells and human osteoprogeni-tors triggers pleiotropic effect that maysupport osteoblastic function. Bone 42(6):1080–1091.
Haynesworth SE, Goshima J, Goldberg VMet al. 1992; Characterization of cells withosteogenic potential from human marrow.Bone 13(1): 81–88.
Henenfeld M. 1953; The application of Davis’Law. J Natl Assoc Chirop 43(5): 23–28.
Henrich D, Seebach C, Kaehling C et al.2009; Simultaneous cultivation of humanendothelial-like differentiated precursorcells and human marrow stromal cells onbeta-tricalcium phosphate. Tissue Eng PartC Methods 15(4): 551–560.
Hofmann A, Ritz U, Verrier S et al. 2008; Theeffect of human osteoblasts on prolifera-tion and neo-vessel formation of humanumbilical vein endothelial cells in a long-term 3D co-culture on polyurethane scaf-folds. Biomaterials 29(31): 4217–4226.
Holtorf HL, Jansen JA, Mikos AG. 2006; Modu-lation of cell differentiation in bone tissueengineering constructs cultured in a bioreac-tor. Adv Exp Med Biol 585: 225–241.
Horner EA, Kirkham J, Wood D et al. 2010;Long bone defect models for tissue engi-neering applications: criteria for choice.Tissue Eng Part B Rev 16(2): 263–271.
Hosseini A, Hogg DA. 1991a; The effects ofparalysis on skeletal development in thechick embryo. I. General effects. J Anat177: 159–168.
Hosseini A, Hogg DA. 1991b; The effects ofparalysis on skeletal development in thechick embryo. II. Effects on histogenesisof the tibia. J Anat 177: 169–178.
Hudlicka O, Brown MD, May S et al. 2006;Changes in capillary shear stress in skele-tal muscles exposed to long-term activity:role of nitric oxide. Microcirculation 13(3): 249–259.
Hutmacher DW. 2000; Scaffolds in tissue engi-neering bone and cartilage. Biomaterials 21(24): 2529–2543.
Hutmacher DW, Sittinger M. 2003; Periostealcells in bone tissue engineering. Tissue Eng9(Suppl 1): S45–64.
Illich DJ, Demir N, Stojkovic M et al. 2011;Concise review: induced pluripotent stemcells and lineage reprogramming: prospectsfor bone regeneration. Stem Cells 29(4):555–563.
Ingram DA, Mead LE, Tanaka H et al. 2004;Identification of a novel hierarchy of endo-thelial progenitor cells using human pe-ripheral and umbilical cord blood. Blood104(9): 2752–2760.
IshaugSL, CraneGM,MillerMJ et al.1997; Boneformation by three-dimensional stromal oste-oblast culture in biodegradable polymer scaf-folds. J Biomed Mater Res 36(1): 17–28.
Ito H, Koefoed M, Tiyapatanaputi P et al.2005; Remodeling of cortical bone allo-grafts mediated by adherent rAAV-RANKLand VEGF gene therapy. Nat Med 11(3):291–297.
Jackson RA, Nurcombe V, Cool SM. 2006;Coordinated fibroblast growth factor andheparan sulfate regulation of osteogenesis.Gene 379: 79–91.
Jacobsen FS. 1997; Periosteum: its relationto pediatric fractures. J Pediatr Orthop B6(2): 84–90.
Jain RK, Au P, Tam J et al. 2005; Engineeringvascularized tissue. Nat Biotechnol 23(7):821–823.
Jain RK, Munn LL, Fukumura D. 2002; Dissect-ing tumour pathophysiology using intravitalmicroscopy. Nat Rev Cancer 2(4): 266–276.
Jones EA, le Noble F, Eichmann A. 2006;What determines blood vessel structure?Genetic prespecification vs. hemodynam-ics. Physiology 21: 388–395.
Kahle M, Wiesmann HP, Berr K et al. 2010;Embryonic stem cells induce ectopic boneformation in rats. Bio-medical materialsand engineering 20(6): 371–380.
Kaigler D, Krebsbach PH, West ER et al.2005; Endothelial cell modulation of bonemarrow stromal cell osteogenic potential.The FASEB journal: official publication ofthe Federation of American Societies forExperimental Biology 19(6): 665–667.
Kaigler D, Wang Z, Horger K et al. 2006; VEGFscaffolds enhance angiogenesis and bone re-generation in irradiated osseous defects.Journal of bone and mineral research: theofficial journal of the American Society forBone and Mineral Research 21(5): 735–744.
Kaji H, Camci-Unal G, Langer R et al. 2011;Engineering systems for the generation ofpatterned co-cultures for controlling cell-cell interactions. Biochim Biophys Acta1810(3): 239–250.
Kalka C, Masuda H, Takahashi T et al. 2000;Transplantation of ex vivo expanded endothe-lial progenitor cells for therapeutic neovascu-larization. Proc Natl Acad Sci USA 97(7):3422–3427.
Kalpakcioglu BB, Morshed S, Engelke K et al.2008; Advanced imaging of bone macro-structure andmicrostructure in bone fragilityand fracture repair. J Bone Joint Surg Am 90(Suppl 1): 68–78.
Kanczler JM, Oreffo RO. 2008; Osteogenesisand angiogenesis: the potential for engi-neering bone. Eur Cell Mater 15: 100–114.
Kang KT, Allen P, Bischoff J. 2011; Bioengi-neered human vascular networks trans-planted into secondary mice reconnectwith the host vasculature and re-establishperfusion. Blood 118(25): 6718–6721.
Kempen DH, Lu L, Heijink A et al. 2009;Effect of local sequential VEGF andBMP-2 delivery on ectopic and orthoto-pic bone regeneration. Biomaterials 30(14): 2816–2825.
Ker ED, Chu B, Phillippi JA et al. 2011a; Engi-neering spatial control of multiple differenti-ation fates within a stem cell population.Biomaterials 32(13): 3413–3422.
Ker ED, Nain AS, Weiss LE et al. 2011b;Bioprinting of growth factors onto alignedsub-micron fibrous scaffolds for simulta-neous control of cell differentiationand alignment. Biomaterials 32(32):8097–8107.
Kim SS, Park MS, Cho S et al. 2010; Enhancedbone formation by marrow-derived endothe-lial and osteogenic cell transplantation. JBiomed Mater Res A 92(1): 246–253.
Kimelman-Bleich N, Seliktar D, Kallai I et al.2011; The effect of ex vivo dynamic loadingon the osteogenic differentiation of geneti-cally engineered mesenchymal stem cellmodel. J Tissue Eng Regen Med 5(5):384–393.
Kleinman HK, McGarvey ML, Liotta LA et al.1982; Isolation and characterization oftype IV procollagen, laminin, and heparansulfate proteoglycan from the EHS sar-coma. Biochemistry 21(24): 6188–6193.
Kneser U, Polykandriotis E, Ohnolz J et al.2006a; Engineering of vascularized trans-plantable bone tissues: induction of axialvascularization in an osteoconductive ma-trix using an arteriovenous loop. TissueEng 12(7): 1721–1731.
Kneser U, Schaefer D, Polykandriotis J et al.2006b; Tissue engineering of bone: thereconstructive surgeon’s point of view.J Cell Mol Med 10(1): 7–19.
Knothe Tate ML, Ritzman T, Schneider F et al.2007; Testing of a new one-stage bone-trans-port surgical procedure exploiting the perios-teum for the repair of long-bone defects.J Bone Joint Surg Am 89(2): 307–316.
Ko HC, Milthorpe BK, McFarland CD. 2007;Engineering thick tissues--the vascularisationproblem. Eur Cell Mater 14: 1–18; discussion18-19.
KolbeM, Xiang Z, Dohle E et al. 2011; Paracrineeffects influenced by cell culturemediumandconsequences on microvessel-like structuresin co-culture s of mesenchymal stem cellsand outgrowth endothelial cells. Tissue EngPart A 17(17-18): 2199–2212.
Komatsu DE, Hadjiargyrou M. 2004; Activationof the transcription factor HIF-1 and its targetgenes, VEGF, HO-1, iNOS, during fracturerepair. Bone 34(4): 680–688.
Koob S, Torio-Padron N, Stark GB et al. 2011;Bone formation and neovascularizationediated by mesenchymal stem cells andendothelial cells in critical-sized calvarialdefects. Tissue Eng Part A 17(3-4): 311–321.
Kostopoulos L, Karring T. 1995; Role of perios-teum in the formation of jaw bone. An exper-iment in the rat. J Clin Periodontol 22(3):247–254.
Kruyt MC, Dhert WJ, Yuan H et al. 2004; Bonetissue engineering in a critical size defectcompared to ectopic implantations in thegoat. J Orthop Res: 22(3): 544–551.
Kumar S, Wan C, Ramaswamy G et al. 2010;Mesenchymal stem cells expressing osteo-genic and angiogenic factors synergisticallyenhance bone formation in a mouse modelof segmental bone defect. Mol Ther 18(5):1026–1034.
Vascularised Bone Tissue Engineering using Coculture Systems
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

Kuznetsov SA, Cherman N, Robey PG. 2011; Invivo bone formation by progeny of humanembryonic stem cells. Stem Cells Dev 20(2):269–287.
Langer R, Vacanti JP. 1993. Tissue engineer-ing. Science 260(5110): 920–926.
Lanza RP, Langer RS, Vacanti J. 2007; Princi-ples of tissue engineering 3rd edn. Elsevier /Academic Press: Amsterdam; Boston.
Le Blanc K. 2003; Immunomodulatory effectsof fetal and adult mesenchymal stem cells.Cytotherapy 5(6): 485–489.
Lee DY, Cho TJ, Kim JA et al. 2008; Mobiliza-tion of endothelial progenitor cells in frac-ture healing and distraction osteogenesis.Bone 42(5): 932–941.
Lee KE, Silva A, Mooney DJ. 2011; Growthfactor delivery-based tissue engineering:general approaches and a review of recentdevelopments. J R Soc Interface 8(55):153–170.
Lelkes G. 1958; Experiments in vitro on therole of movement in the development ofjoints. J Embryol Exp Morphol 6(2):183–186.
Lemperle SM, Calhoun CJ, Curran RW et al.1998; Bony healing of large cranial andman-dibular defects protected from soft-tissueinterposition: A comparative study of sponta-neous bone regeneration, osteoconduction,and cancellous autografting in dogs. PlastReconstr Surg 101(3): 660–672.
Leondes CT. 2001; Biomechanical systems:techniques and applications. 4 vols. CRCPress: Boca Raton.
Lin K, Hsu PP, Chen BP et al. 2000; Molecularmechanism of endothelial growth arrest bylaminar shear stress. Proc Natl Acad SciUSA 97(17): 9385–9389.
Liu Y, Teoh SH, Chong M et al. 2012; Vvasculo-genic and osteogenesis-enhancing potentialof human umbilical cord blood endothelialcolony-forming cells stem cells. In press.
Lovett M, Lee K, Edwards A et al. 2009; Vascu-larization strategies for tissue engineering.Tissue Eng Part B Rev 15(3): 353–370.
Marie P, Debiais F, Cohen-Solal M et al. 2000;New factors controlling bone remodeling.Joint, bone, spine: revue du rhumatisme 67(3): 150–156.
Masuda H, Alev C, Akimaru H et al. 2011;Methodological development of a clonogenicassay to determine endothelial progenitorcell potential. Circ Res 109(1): 20–37.
Matsumoto T, Kawamoto A, Kuroda R et al.2006; Therapeutic potential of vasculogen-esis and osteogenesis promoted by peripheralblood CD34-positive cells for functional bonehealing. Am J Pathol 169(4): 1440–1457.
MatsumotoT,Mifune Y, Kawamoto A et al. 2008;Fracture inducedmobilization and incorpora-tion of bone marrow-derived endothelialprogenitor cells for bone healing. J CellPhysiol 215(1): 234–242.
McBride SH, Knothe U, Brianza S et al. 2010;Periosteal mechanics from the macro tonano scales. In: Proceedings of the UnitedStates National Applied and TheoreticalMechanics Conference, Pittsburgh.
McCloskey KE, Smith DA, Jo H et al. 2006;Embryonic stem cell-derived endothelialcells may lack complete functional matura-tion in vitro. J Vasc Res 43(5): 411–421.
McCoy RJ, O’Brien FJ. 2010; Influence of shearstress in perfusion bioreactor cultures for thedevelopment of three-dimensional bonetissue constructs: a review. Tissue Eng Part BRev 16(6): 587–601.
Meinel L, Betz O, Fajardo R et al. 2006; Silkbased biomaterials to heal critical sizedfemur defects. Bone 39(4): 922–931.
Meinel L, Fajardo R, Hofmann S et al. 2005;Silk implants for the healing of critical sizebone defects. Bone 37(5): 688–698.
Melcher AH, Accursi GE. 1971; Osteogeniccapacity of periosteal and osteoperiostealflaps elevated from the parietal bone ofthe rat. Arch Oral Biol 16(6): 573–580.
Melero-Martin JM, De Obaldia ME, Kang SYet al. 2008; Engineering robust and func-tional vascular networks in vivo with humanadult and cord blood-derived progenitorcells. Circ Res 103(2): 194–202.
Menger MD, Lehr HA. 1993; Scope andperspectives of intravital microscopy--bridgeover from in vitro to in vivo. Immunol Today14(11): 519–522.
Mifune Y, Matsumoto T, Kawamoto A et al.2008; Local delivery of granulocyte colonystimulating factor-mobilized cd34-positiveprogenitor cells using bioscaffold for modal-ity of unhealing bone fracture. Stem Cells 26(6): 1395–405.
Mironov V, Kasyanov V, Markwald RR. 2011;Organ printing: from bioprinter to organbiofabrication line. Curr Opin Biotechnol22(5): 667–673.
Moioli EK, Clark PA, Chen M et al. 2008;Synergistic actions of hematopoietic andmesenchymal stem/progenitor cells in vascu-larizing bioengineered tissues. PLoS One 3(12): e3922.
Moon S, Hasan SK, Song Y et al. 2010; Layer bylayer three-dimensional tissue epitaxy bycell-laden hydrogel droplets. Tissue Eng PartC Methods 16(1): 157–166.
Murohara T. 2010; Cord blood-derived earlyoutgrowth endothelial progenitor cells.Microvasc Res 79(3): 174–177.
Muschler GF, Nakamoto C, Griffith LG. 2004;Engineering principles of clinical cell-based tissue engineering. J Bone Joint SurgAm 86-A(7): 1541–1558.
Muschler GF, Raut VP, Patterson TE et al. 2010;The design and use of animal models fortranslational research in bone tissue engi-neering and regenerative medicine. TissueEng Part B Rev 16(1): 123–145.
Nowlan NC, Prendergast PJ, Murphy P. 2008;Identification of mechanosensitive genesduring embryonic bone formation. PLoSComput Biol 4(12): e1000250.
O’Loughlin PF, Morr S, Bogunovic L et al. 2008;Selection and development of preclinicalmodels in fracture-healing research. TheJournal of bone and joint surgery. Americanvolume 90(Suppl 1): 79–84.
Ouyang HW, CaoT, Zou XH et al. 2006; Mesen-chymal stem cell sheets revitalize nonviabledense grafts: implications for repair of large-bone and tendon defects. Transplantation 82(2): 170–174.
Ozaki A, Tsunoda M, Kinoshita S et al. 2000;Role of fracture hematoma and periosteumduring fracture healing in rats: interactionof fracture hematoma and the periosteum inthe initial step of the healing process. Journalof orthopaedic science: official journal of theJapanese Orthopaedic Association 5(1):64–70.
PanjabiMM, Lindsey RW,Walter SD et al. 1989;The clinician’s ability to evaluate the strengthof healing fractures from plain radiographs. JOrthop Trauma 3(1): 29–32.
PengH,Wright V, Usas A et al. 2002; Synergisticenhancement of bone formation and healing
by stem cell-expressed VEGF and bone mor-phogenetic protein-4. J Clin Invest 110(6):751–759.
Pittenger MF, Mackay AM, Beck S et al. 1999;Multilineage potential of adult humanmesenchymal stem cells. Science 284(5411): 143–147.
Prockop DJ. 1997; Marrow stromal cells asstem cells for nonhematopoietic tissues.Science 276(5309): 71–74.
Qi H, Aguiar DJ, Williams SM et al. 2003; Iden-tification of genes responsible for osteoblastdifferentiation from humanmesodermal pro-genitor cells. Proc Natl Acad Sci USA 100(6):3305–3310.
Quarto R, Mastrogiacomo M, Cancedda R et al.2001; Repair of large bone defects with theuse of autologous bonemarrow stromal cells.N Engl J Med 344(5): 385–386.
Reichert JC, Saifzadeh S,WullschlegerME et al.2009; The challenge of establishing preclini-cal models for segmental bone defect re-search. Biomaterials 30(12): 2149–2163.
Rho JY, Kuhn-Spearing L, Zioupos P. 1998;Mechanical properties and the hierarchicalstructure of bone. Med Eng Phys 20(2):92–102.
Robey PG. 2011; Cell sources for bone regen-eration: the good, the bad, and the ugly(but promising). Tissue Eng Part B Rev 17(6): 423–430.
Rouwkema J, de BoeJ R, Van BlitterswijkCA. 2006; Endothelial cells assemble intoa 3-dimensional prevascular network in abone tissue engineering construct. TissueEng 12(9): 2685–2693.
Rouwkema J, Rivron NC, van Blitterswijk CA.2008; Vascularization in tissue engineering.Trends Biotechnol 26(8): 434–441.
Rouwkema J, Westerweel P, de Boer E et al.2009; The use of endothelial progenitor cellsfor prevascularized bone tissue engineering.Tissue Eng Part A 15(8): 2015–2027.
Rozen N, Bick T, Bajayo A et al. 2009; Trans-planted blood-derived endothelial progenitorcells (EPC) enhance bridging of sheep tibiacritical size defects. Bone 45(5): 918–924.
Runyan CM, Jones DC, Bove KE et al. 2010;Porcine allograft mandible revitalizationusing autologous adipose-derived stemcells, bone morphogenetic protein-2, andperiosteum. Plast Reconstr Surg 125(5):1372–1382.
Salgado AJ, Coutinho OP, Reis RL. 2004;Bone tissue engineering: state of the artand future trends. Macromol Biosci 4(8):743–765.
Salter EK, Goh BC, Hung BP et al. 2011; Bonetissue engineering bioreactors - a role inthe clinic? Tissue Eng. Part B Rev 18(1):62–75.
Satoh T, Tsuchiya M, Harii K. 1983; A vascu-larised iliac musculo-periosteal free flaptransfer: a case report. Brit J Plast Surg36(1): 109–112.
Schantz JT, Woodruff MA, Lam CX et al. 2012;Differentiation potential of mesenchymal;progenitor cells following transplantationinto calvarial defects. J Mech Behav BiomedMater 11: 132–142.
Schuurman W, Khristov V, Pot M et al. 2011;Bioprinting of hybrid tissue constructswith tailorable mechanical properties.Biofabrication 3(2): 021001.
Seebach C, Henrich D, Kahling C et al. 2010;Endothelial progenitor cells and mesen-chymal stem cells seeded onto beta-TCPgranules enhance early vascularization
Y. Liu et al.
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term

and bone healing in a critical-sized bonedefect in rats. Tissue Eng Part A 16(6):1961–1970.
Shields LB, RaqueGH, Glassman SD et al. 2006;Adverse effects associated with high-doserecombinant human bone morphogeneticprotein-2 use in anterior cervical spinefusion. Spine 31(5): 542–547.
Shimizu T, Sasano Y, Nakajo S et al. 2001;Osteoblastic differentiation of periosteum-derived cells is promoted by the physicalcontact with the bone matrix in vivo. AnatRec 264(1): 72–81.
Sierra DH. 1993; Fibrin sealant adhesivesystems: a review of their chemistry,material properties and clinical applica-tions. J Biomater Appl 7: 309.
Sikavitsas VI, Bancroft GN, Holtorf HL et al.2003; Mineralized matrix deposition bymarrow stromal osteoblasts in 3D perfusionculture increases with increasing fluid shearforces. Proc Natl Acad Sci USA 100(25):14683–14688.
Simons M, Annex BH, Laham RJ et al. 2002;Pharmacological treatment of coronaryartery disease with recombinant fibroblastgrowth factor-2: double-blind, random-ized, controlled clinical trial. Circulation105(7): 788–793.
Simpson AH. 1985; The blood supply of theperiosteum. J Anat 140(Pt 4): 697–704.
Skoog, T.1965; The use of periostealflaps in therepair of clefts of the primary palate.Uppsalla, Sweden. Cleft Palate J 2: 332–339
Soltan M, Smiler D, Soltan C. 2009; Theinverted periosteal flap: a source of stemcells enhancing bone regeneration.Implant Dent 18(5): 373–379.
Song HH, Shi W, Xiang YY et al. 2005; Theloss of glypican-3 induces alterations inWnt signaling. J Biol Chem 280(3):2116–2125.
Song JW, Munn LL. 2011; Fluid forces con-trol endothelial sprouting. Proc Natl AcadSci USA 108(37): 15342–15347.
Stevenson S. 1999; Biology of bone grafts.Orthop Clin North Am 30(4): 543–552.
Stolberg S, McCloskey KE. 2009; Can shearstress direct stem cell fate? Biotechnol Prog25(1): 10–19.
Street J, Bao M, deGuzman L et al. 2002;Vascular endothelial growth factor stimu-lates bone repair by promoting angiogene-sis and bone turnover. Proc Natl Acad SciUSA 99(15): 9656–9661.
Szpalski C, Barbaro M, Sagebin F et al. 2012;Bone tissue engineering: current strategiesand techniques-part II: cell types. TissueEng Part B Rev 18(4): 258–269.
Tateishi-Yuyama E, Matsubara H, Murohara Tet al. 2002; Therapeutic angiogenesis forpatients with limb ischaemia by autologoustransplantation of bone-marrow cells: a pilotstudy and a randomised controlled trial.Lancet 360(9331): 427–435.
Tortelli F, Pujic N, Liu Y et al. 2009; Osteoblastand osteoclast differentiation in an in vitrothree-dimensional model of bone. TissueEng Part A 15(9): 2373–2383.
Tran Van PT, Vignery A, Baron R.1982; Cellularkinetics of the bone remodeling sequence inthe rat. Anat Rec 202(4): 445–451.
Trueta J, Buhr AJ. 1963; The vascular contribu-tion to osteogenesis. V. The vasculature
supplying the epiphysial cartilage in rachiticrats. J Bone Joint Surg Br 45: 572–581.
Tsigkou O, Pomerantseva I, Spencer JA et al.2010; Engineered vascularized bone grafts.Proc Natl Acad Sci USA 107(8): 3311–3316.
Tzima E, Irani-Tehrani M, Kiosses WB et al.2005; A mechanosensory complex thatmediates the endothelial cell response tofluid shear stress. Nature 437(7057):426–431.
Unger RE, Ghanaati S, Orth C et al. 2010; Therapid anastomosis between prevascularizednetworks on silk fibroin scaffolds generatedin vitro with co-culture s of human microvas-cular endothelial and osteoblast cells and thehost vasculature. Biomaterials 31(27):6959–6967.
Unger RE, Sartoris A, Peters K et al. 2007;Tissue-like self-assembly in co-cultures ofendothelial cells and osteoblasts and theformation of microcapillary-like structureson three-dimensional porous biomaterials.Biomaterials 28(27): 3965–3976.
Usami K, Mizuno H, Okada K et al. 2009;Composite implantation of mesenchymalstem cells with endothelial progenitor cellsenhances tissue-engineered bone forma-tion. J Biomed Mater Res A 90(3): 730–741.
Utvag SE, Grundnes O, Reikeraos O. 1996;Effects of periosteal stripping on healingof segmental fractures in rats. J OrthopTrauma 10(4): 279–284.
Villars F, Bordenave L, Bareille R et al. 2000;Effect of human endothelial cells onhuman bone marrow stromal cell pheno-type: role of VEGF? J Cell Biochem 79(4):672–685.
Villars F, Guillotin B, Amedee T et al. 2002;Effect of HUVEC on human osteoprogeni-tor cell differentiation needs heterotypicgap junction communication. Am J PhysiolCell Physiol 282(4): C775–785.
Wan C, Gilbert SR, Wang Y et al. 2008; Role ofhypoxia inducible factor-1 alpha pathway inbone regeneration. J Musculoskelet NeuronalInteract 8(4): 323–324.
Wan C, Shao J, Gilbert SR et al. 2010; Role ofHIF-1alpha in skeletal development. Ann NY Acad Sci 1192: 322–326.
Wang Y, Wan C, Deng L et al. 2007; Thehypoxia-inducible factor alpha pathwaycouples angiogenesis to osteogenesisduring skeletal development. J Clin Invest117(6): 1616–1626.
Warnke PH, Springer IN, Wiltfang J et al.2004; Growth and transplantation of acustom vascularised bone graft in a man.Lancet 364(9436): 766–770.
WengD, HurzelerMB,Quinones CR et al. 2000;Contribution of the periosteum to boneformation in guided bone regeneration. Astudy in monkeys. Clin Oral Implants Res 11(6): 546–554.
Werntz JR, Lane JM, Burstein AH et al. 1996;Qualitative and quantitative analysis oforthotopic bone regeneration by marrow.Journal of orthopaedic research: officialpublication of the Orthopaedic ResearchSociety 14(1): 85–93.
Wolff J.1892;Das Gezetz der Transformation derKnochen. Hirschwald: Berlin.
Wong DA, Kumar A, Jatana S et al. 2008; Neu-rologic impairment from ectopic bone in thelumbar canal: a potential complication of
off-label PLIF/TLIF use of bone morphoge-netic protein-2 (BMP-2). The spine journal:official journal of the North American SpineSociety 8(6): 1011–1018.
Wood LK, Asada R. 2011; Stochastic model-ing and identification of emergent beha-viors of an Endothelial Cell populationin angiogenic pattern formation. The In-ternational Journal of Robotics Research30(6): 659–677.
Wu DC, Boyd AS, Wood KJ. 2008; Embryonicstem cells and their differentiated derivativeshave a fragile immune privilege but stillrepresent novel targets of immune attack.Stem Cells 26(8): 1939–1950.
Xue Y, Xing Z, Hellem S et al. 2009; Endothe-lial cells influence the osteogenic potentialof bone marrow stromal cells. Biomed EngOnline 8: 34.
Yamamoto K, Sokabe T, Watabe T et al.2005; Fluid shear stress induces differ-entiation of Flk-1-positive embryonicstem cells into vascular endothelial cellsin vitro. Am J Physiol Heart Circ Physiol288(4): H1915–1924.
Yamamoto K, Takahashi T, Asahara T et al.2003; Proliferation, differentiation, and tubeformation by endothelial progenitor cells inresponse to shear stress. J Appl Physiol 95(5): 2081–2088.
Yoder MC, Mead LE, Prater D et al. 2007;Redefining endothelial progenitor cellsvia clonal analysis and hematopoieticstem/progenitor cell principals. Blood109(5): 1801–1809.
Zelzer E, McLean W, Ng Y et al. 2002; Skeletaldefects in VEGF(120/120) mice reveal multi-ple roles for VEGF in skeletogenesis. Develop-ment 129(8): 1893–1904.
Zhang ZY, Teoh SH, Chong MS et al.2010a; Neo-vascularization and boneformation mediated by fetal mesenchy-mal stem cell tissue-engineered bonegrafts in critical-size femoral defects.Biomaterials 31(4): 608–620.
Zhang ZY, Teoh SH, ChongMS et al. 2009a; Su-perior osteogenic capacity for bone tissue en-gineering of fetal compared to perinatal andadult mesenchymal stem cells. Stem Cells 27(1): 126–137.
Zhang ZY, Teoh SH, ChongWS et al. 2009b; Abiaxial rotating bioreactor for the cultureof fetal mesenchymal stem cells for bonetissue engineering. Biomaterials 30(14):2694–2704.
Zhang ZY, Teoh SH, Hui JH et al. 2012; Thepotential of human fetal mesenchymalstem cells for off-the-shelf bone tissue en-gineering application. Biomaterials 33(9):2656–2672.
Zhang ZY, Teoh SH, Teo EY et al. 2010b; Acomparison of bioreactors for culture offetal mesenchymal stem cells for bonetissue engineering. Biomaterials 31(33):8684–8695. Epub 2010 Aug 24.
ZhaoT, Zhang ZN, Rong Z et al. 2011; Immuno-genicity of induced pluripotent stem cells.Nature 474(7350): 212–215.
Zou D, Zhang Z, Ye D et al. 2011; Repairof critical-sized rat calvarial defectsusing genetically engineered bone mar-row-derived mesenchymal stem cellsoverexpressing hypoxia-inducible factor-1alpha. Stem Cells 29(9): 1380–1390.
Vascularised Bone Tissue Engineering using Coculture Systems
Copyright © 2012 John Wiley & Sons, Ltd. J Tissue Eng Regen Med (2012)DOI: 10.1002/term




















