
Responses to flooding intensity in Leontodon taraxacoides
10
Printed from the CJO service for personal use only by... New Phytol. (1999), 141, 119–128 Responses to flooding intensity in Leontodon taraxacoides A. A. GRIMOLDI*, P. INSAUSTI, G. G. ROITMAN A. SORIANO Departamento de Ecologı U a, Facultad de Agronomı U a, Universidad de Buenos Aires, Avenida San Martı U n 4453, 1417 Buenos Aires, Argentina Received 19 May 1998 ; accepted 11 September 1998 Natural flooding is one of the major factors affecting vegetation dynamics in many regions of the world. The Flooding Pampa Grasslands (Argentina) are frequently exposed to flooding events of diverse intensity and duration, some of which Leontodon taraxacoides, an exotic dicot. frequent in these grasslands, seems to survive. Its responses to four different water depths (0, 1, 7 and 13 cm) were studied. The results indicate that plants in conditions of total submergence (depth of 13 cm) did not survive. In less severe flood conditions, increases in the leaf insertion angle resulted in the maintenance of a large proportion of the total leaf area above the water. Differences in leaf length and a decrease in the width and the proportion of lobes per leaf were also found under partial submergence conditions (depth of 7 cm). Root and leaf aerenchyma, present in unflooded plants, showed a significant increase in flood conditions. In spite of the anatomical and morphological responses, total biomass and leaf area were severely affected by water depth. Control plants allocated more biomass to reproductive organs, while partly submerged plants allocated more to leaves and less to reproductive organs. Mature L. taraxacoides plants presented a wide range of plastic adjustment as a survival strategy in soil anaerobiosis, and appear to be able to survive short spring floods in a vegetative state ; in contrast, they might not tolerate total submergence conditions imposed by more intense and long-lasting floods. Key words : flood, water depth, aerenchyma, leaf morphology, allocation, Leontodon taraxacoides, Flooding Pampa Grasslands. In certain types of grassland, natural flooding is a major factor in the disturbance regime. In these systems, plants have to cope with unpredictable periods of flooding at different times of the year. The interaction between plant responses and the charac- teristics of the flooding event, such as duration, seasonal timing, water depth and frequency, influences floristic composition and plant community dynamics (Menges & Waller, 1983 ; Voesenek et al., 1992). Responses to increasing flooding intensity vary widely among plant species and are generally associated with anatomical and morphological changes (Jackson & Drew, 1984 ; Blom & Voesenek, 1996). For example, it has been found that in the genus Rumex, aerenchyma formation and leaf elongation are important for the recovery of contact with the aerial environment and allow oxygen transport to the submerged tissues (Laan et al., *Author for correspondence (e-mail grimoldi!ifeva.edu.ar). 1990b; Van der Sman et al., 1991). In conditions of high flooding, when the water surface is not reached by plants, other responses such as underwater photosynthesis (Laan & Blom, 1990a), and metabolic adaptations or dormancy are critical for plant survival (Armstrong et al., 1994). In some cases, plant responses to flooding involve changes in biomass allocation patterns, enabling tolerant plants to remain vegetative during flooding periods (Van der Sman, et al. 1993; Blom & Voesenek, 1996). The Flooding Pampa Grassland (Argentina) is currently exposed to cattle grazing and periodic flooding events of diverse intensity and duration (Chaneton et al., 1988 ; Soriano, 1991). Natural floods cause most dicots in the lowland communities to decrease their biomass formation or to die, while native monocots generally increase their cover and biomass (Chaneton et al., 1988 ; Insausti, 1996). Recent studies in this grassland found that native grasses present plastic responses to flooding, such as aerenchyma tissue formation and increase in plant height (Rubio et al., 1995 ; Loreti & Oesterheld,
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Responses to flooding intensity in Leontodon taraxacoides

Printed from the CJO service for personal use only by...
New Phytol. (1999), 141, 119–128
Responses to flooding intensity in
Leontodon taraxacoides
A. A. GRIMOLDI*, P. INSAUSTI, G. G. ROITMAN A. SORIANO
Departamento de EcologıUa, Facultad de AgronomıUa, Universidad de Buenos Aires,
Avenida San MartıUn 4453, 1417 Buenos Aires, Argentina
Received 19 May 1998; accepted 11 September 1998
Natural flooding is one of the major factors affecting vegetation dynamics in many regions of the world. The
Flooding Pampa Grasslands (Argentina) are frequently exposed to flooding events of diverse intensity and
duration, some of which Leontodon taraxacoides, an exotic dicot. frequent in these grasslands, seems to
survive. Its responses to four different water depths (0, 1, 7 and 13 cm) were studied. The results indicate that
plants in conditions of total submergence (depth of 13 cm) did not survive. In less severe flood conditions,
increases in the leaf insertion angle resulted in the maintenance of a large proportion of the total leaf area above
the water. Differences in leaf length and a decrease in the width and the proportion of lobes per leaf were also found
under partial submergence conditions (depth of 7 cm). Root and leaf aerenchyma, present in unflooded
plants, showed a significant increase in flood conditions. In spite of the anatomical and morphological
responses, total biomass and leaf area were severely affected by water depth. Control plants allocated more biomass
to reproductive organs, while partly submerged plants allocated more to leaves and less to reproductive organs.
Mature L. taraxacoides plants presented a wide range of plastic adjustment as a survival strategy in soil
anaerobiosis, and appear to be able to survive short spring floods in a vegetative state; in contrast, they might not
tolerate total submergence conditions imposed by more intense and long-lasting floods.
Key words: flood, water depth, aerenchyma, leaf morphology, allocation, Leontodon taraxacoides, Flooding
Pampa Grasslands.
In certain types of grassland, natural flooding is a
major factor in the disturbance regime. In these
systems, plants have to cope with unpredictable
periods of flooding at different times of the year. The
interaction between plant responses and the charac-
teristics of the flooding event, such as duration,
seasonal timing, water depth and frequency,
influences floristic composition and plant community
dynamics (Menges & Waller, 1983; Voesenek et al.,
1992). Responses to increasing flooding intensity
vary widely among plant species and are generally
associated with anatomical and morphological
changes (Jackson & Drew, 1984; Blom & Voesenek,
1996). For example, it has been found that in the
genus Rumex, aerenchyma formation and leaf
elongation are important for the recovery of contact
with the aerial environment and allow oxygen
transport to the submerged tissues (Laan et al.,
*Author for correspondence (e-mail grimoldi!ifeva.edu.ar).
1990b; Van der Sman et al., 1991). In conditions of
high flooding, when the water surface is not reached
by plants, other responses such as underwater
photosynthesis (Laan & Blom, 1990a), and metabolic
adaptations or dormancy are critical for plant
survival (Armstrong et al., 1994). In some cases,
plant responses to flooding involve changes in
biomass allocation patterns, enabling tolerant plants
to remain vegetative during flooding periods (Van
der Sman, et al. 1993; Blom & Voesenek, 1996).
The Flooding Pampa Grassland (Argentina) is
currently exposed to cattle grazing and periodic
flooding events of diverse intensity and duration
(Chaneton et al., 1988; Soriano, 1991). Natural
floods cause most dicots in the lowland communities
to decrease their biomass formation or to die, while
native monocots generally increase their cover and
biomass (Chaneton et al., 1988; Insausti, 1996).
Recent studies in this grassland found that native
grasses present plastic responses to flooding, such as
aerenchyma tissue formation and increase in plant
height (Rubio et al., 1995; Loreti & Oesterheld,

Printed from the CJO service for personal use only by...
120 A. A. Grimoldi et al.
1996). We have little information regarding the
effects of flooding on exotic dicot. species naturalized
in the region, but it is known that after floods some
exotic dicots such as Leontodon taraxacoides persist,
possibly representing 10% of the total biomass in
lowland communities (Insausti, 1996).
Plastic responses that could minimize environ-
mental constraints to ensure survival, as a solution to
temporally heterogeneous environments, have been
discussed previously (Bradshaw, 1965; Via et al.,
1995). These responses could be important for the
survival of such exotic dicots as L. taraxacoides in the
Flooding Pampa Grasslands. Our objective was to
identify the anatomical, morphological and allo-
metric responses of L. taraxacoides measured in
different depths of water, and to analyse the effects of
flooding intensity on the survival of the plant,
particularly relevant when one considers the natural
flooding of grasslands which are invaded by this
species.
Species characteristics and study site
Leontodon taraxacoides (Vill) Merat is a short-lived
European perennial herb from the Asteraceae family
(tribe: Cichorieae). In Europe it is a common plant in
poor and overgrazed grasslands (Rivas & Rivas,
1963) found in dry open sites and sand dunes as well
as in pond borders (Pignatti, 1982). In the Flooding
Pampa Grassland, it is present only in plant
communities in humid lowland (Burkart, et al.
1990); it is characterized by self-compatibility,
bractless scapes with solitary heads, lobate leaves
disposed in a rosette, and a short erect truncate
stock. The reproductive phase runs from the end of
spring until the beginning of autumn.
Plants used in this study were taken from a field
under continuous grazing in Pila, in the Province of
Buenos Aires, Argentina (36° 30« S, 58° 30« W).
Plants were collected from a stand of Piptochaetium
montevidense, Ambrosia tenuifolia, Eclipta bellidioides,
Mentha pulegium and Briza subaristata, one of the
most widespread communities of the Flooding
Pampa (Burkart et al., 1990); found in flat areas, it is
associated with typical Natraquoll soils, with fine
texture that causes imperfect drainage. Short-lived
(1 month) floods of 7 cm or less occur at the
beginning of almost every spring. In contrast, large
floods are unusual and cover grasslands with water
10–30 cm deep for 3–5 months (Chaneton et al., 1988;
Soriano, 1991). At the same site, field experiments
showed soil redox potential (Eh() (A1 horizon) values
of 223 mV in unflooded conditions and ®153 mV
in flooded (Taboada & Lavado, 1986). The reductive
status of the soil during flooding corresponds with
conditions that reduce growth of most flood-
intolerant species (Ponnamperuma, 1984).
Experimental design
By the end of winter, similarly sized individuals of L.
taraxacoides were extracted in rectangular soil blocks
0±08 m#¬0±2 m, in which the selected plant was
central. In the field, depth of the root system does
not exceed 0±2 m (data not shown). Blocks were
transported to the experimental garden and were put
in groups of three, separated by a plastic membrane,
in a plastic container 0±24 m#¬0±4 m, each of which
represented an experimental unit ; there were five
replications. By this method the difficulty of simu-
lating floods in the field or maintaining experimental
controls during a natural flood were avoided.
After 2 months of acclimatization, treatments
consisting of different flood levels were randomly
assigned and applied for 49 d: (1) control ; (2)
waterlogging; (3) partial submergence; and (4) total
submergence. Controls were well drained and were
watered daily up to field capacity. Waterlogging
consisted of a constant water depth above soil level of
1 cm. At the beginning of the experiment, the partial
submergence and total submergence treatments were
flooded to a depth of 7 cm, the plants remaining
totally submerged. Afterwards the water level was
maintained constant in the partial submergence
treatment and was gradually increased in the total
submergence treatment, according to the height of
the plants, up to 13–14 cm above soil level at the end
of the experiment. The containers were kept in the
open air and were rotated every 7 d. In the flooded
treatments, water was gradually replaced.
Anatomical analysis
Root and leaf porosity were determined using the
pycnometer method (Sojka, 1988), based on the
weight increase which occurs when root or leaf air
volume is replaced by water after maceration. This
analysis was performed using young leaves and
lateral roots. The lateral roots used were produced
from the tap root and were 8–10 cm long, extracted
from a soil depth of 0–5 cm. In addition, some roots
and leaves were fixed and preserved in formalin-
acetic-alcohol. Segments of lateral roots from at least
2 cm from the tip, and leaves from 1 cm from the
plant stock were dehydrated in a series of increasing
ethanol concentrations and embedded in paraffin
wax. Sections 15–20 µm thick were cut, stained with
safranin and fast green and mounted in Canada
balsam. The percentage of root aerenchyma per total
cross-sectional area was calculated with a Minimop
Digitizer Prov. attached to an ocular microscope
(Sojka, 1988). These variables were measured in five
randomly selected plants at the beginning of the
experiment and 15 d after the first measurement, and
at the end of the study in one plant of each
replication. Root aerenchyma and root and leaf
porosity were also measured in five plants grown for
18 months in drained conditions.

Printed from the CJO service for personal use only by...
Flooding intensity and Leontodon taraxacoides 121
L
Lb
La
Lc
1 cm
7 cm
HAa
α
Figure 1. Description of morphological analysis. Leaf
lengths (L) present in three layers were calculated for all
treatments: (La) 0–1 cm, (Lb) 1–7 cm and (Lc) above 7 cm
depth. Leaf height (H), leaf insertion angle in relation to
soil surface (£), maximal (A) and minimal (a) leaf width at
the mid-point of the leaf.
Morphological analysis
Leaf length and insertion angle in relation to the soil
surface were periodically measured. Leaf height was
calculated by: H ¯ sin (£)¬L (Fig. 1). Analyses
were performed by calculating the averages of all
adult leaves and the three maximum values of each
plant. Since results were the same for both analyses,
only the results from adult leaves are presented.
After the plant harvest, length (L), area, mid-rib
diameter, maximal (A) and minimal (a) width at the
mid-point of the leaf were measured in all adult leaves
(Fig. 1). The outlines of 10 randomly selected leaves
were copied onto white paper, the extremes of each
lobe were joined with straight lines and the leaf
dissection index was calculated as the quotient of leaf
area and the area of the empty space surface between
the lobes. All areas were measured with a Li-3000
leaf area meter (Li-Cor, Inc. Lincoln, NE, USA).
The vertical arrangement of leaf area was quantified
by an arbitrary division into three height layers, in
accordance with the imposed experimental water
depths. For all treatments the leaf length per layer
was determined by: La (0–1 cm) ¯ 1 cm}sin (£),
Lb (1–7 cm) ¯ (7 cm}sin £)®La and Lc ("7 cm)
¯ L®Lb®La (Fig. 1). For each individual, the
relationship between leaf length and leaf area was
estimated by linear regression (P!0±0001; r#"0±8)
and the leaf area present in each layer was calculated.
The number of leaves extending beyond each layer
was also calculated at the last measurement.
Biomass and survival
At the end of the experiment, two plants of each
replication were harvested and separated into three
different parts: heads and scapes; leaves; and stock
and roots. After maintaining soil blocks in 0±4%
sodium pyrophosphate solution for 48 h, they were
washed carefully and stock-roots material separated
as completely as possible. The material was weighed
after oven-drying for 72 h at 70°C. The biomass
allocation was analysed as the ratio of each part to the
total plant biomass. The total leaf length:leaf
biomass ratio was also calculated.
Statistical analysis
Morphological variables were analysed by repeated
measures ANOVA (n ¯ 5), with water level as the
between-subject main effect and time as the within-
subject factor. Greenhouse-Geyser adjustment was
used when mild violations of the hypothesis of
sphericity of the covariance matrices occurred. The
analysis was performed in two different ways: all
treatments within four measurement days (5, 10, 19,
25) and excluding the submerged treatment; within
six measurement days (5, 10, 19, 25, 32, 46).
Individual contrasts were made by the Bonferroni
test at a probability level of 0±05 (Von Ende, 1993).
Variables measured once were analysed using one-
way (n ¯ 5) and a posteriori means-
comparison were performed with Tukey tests at a
probability level of 0±05. Leaf area, biomass, leaf
number and scape number data were transformed
(log x) to comply with the assumptions of the
ANOVA. Porosity (%), biomass allocation (%), leaf
area (%) and leaf number (%) per layer data were
(arcsin ox)-transformed. All results are presented as
mean³SE.
Anatomical analysis
Roots and leaves of plants growing for 18 month
under conditions of free drainage showed porosity
values, determined by pycnometry, of 10±5³2±1%
and 11±7³1±8% respectively. The porosity of the
plants growing under field conditions was 20±93³1±7% in roots and 15±87³1±5% in leaves. After 15 d
of flooding, no differences were found in leaf
porosity among treatments (P ¯ 0±2954), but
root porosity decreased in conditions of total sub-
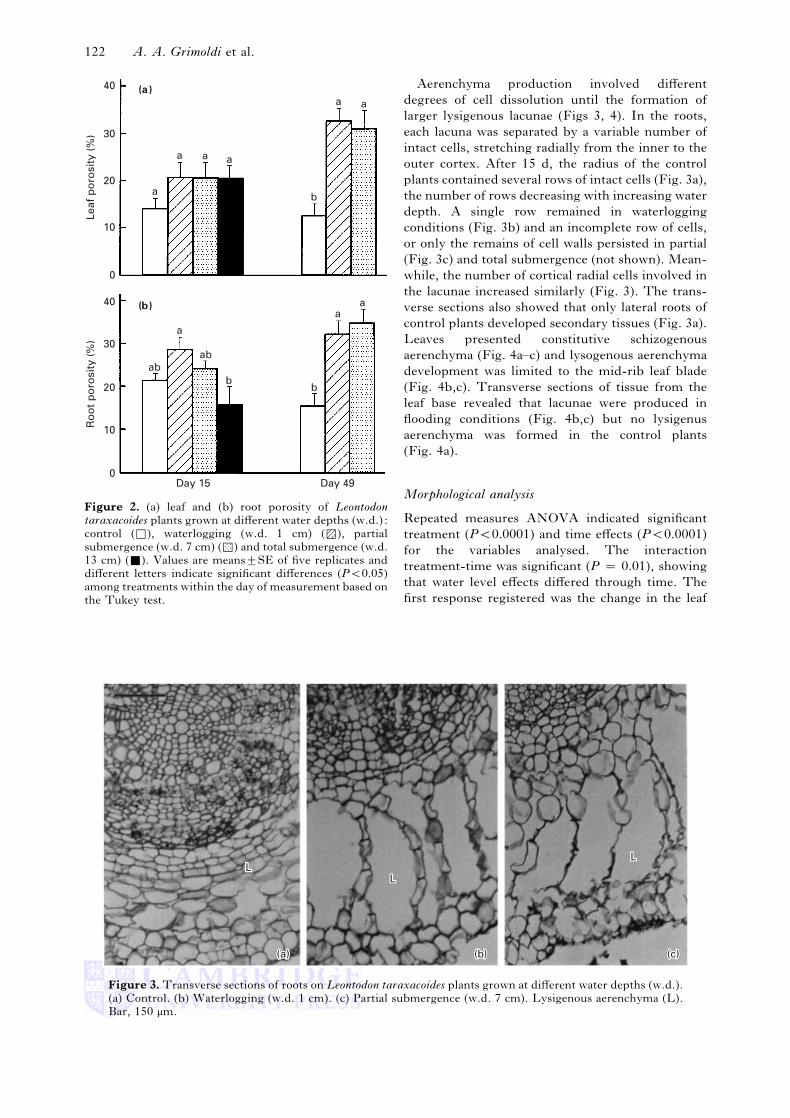
mergence (P ¯ 0±0342) (Fig. 2). After 49 d,
considerable increases in leaf (P ¯ 0±0008) and root
porosity (P ¯ 0±0009) were recorded in plants in the
waterlogging and partial submergence treatments
compared with the controls (Fig. 2). In drained
conditions, porosity values were lower than those
measured at the beginning of the experiment (Fig.
2). Root porosity values, based on pycnometer
measurements, did not differ from those found by
visual cross-sectional area analysis. However, on day
15, the root aerenchyma of totally submerged plants,
calculated by visual analysis, was greater
(31±94³1±58%) than the value obtained by the
pycnometer method (Fig. 2).
Printed from the CJO service for personal use only by...
122 A. A. Grimoldi et al.
(a)40
30
20
10
0
a
a a a
Leaf
po
rosi
ty (
%)
b
a a
(b)40
30
20
10
0
a
ab
b
Ro
ot
po
rosi
ty (
%)
b
aa
Day 15 Day 49
ab
Figure 2. (a) leaf and (b) root porosity of Leontodontaraxacoides plants grown at different water depths (w.d.) :
control (*), waterlogging (w.d. 1 cm) (8), partial
submergence (w.d. 7 cm) (7) and total submergence (w.d.
13 cm) (+). Values are means³SE of five replicates and
different letters indicate significant differences (P!0±05)
among treatments within the day of measurement based on
the Tukey test.
(a)(a)(a) (b)(b)(b) (c)(c)(c)
LLLLLL
LLL
Figure 3. Transverse sections of roots on Leontodon taraxacoides plants grown at different water depths (w.d.).
(a) Control. (b) Waterlogging (w.d. 1 cm). (c) Partial submergence (w.d. 7 cm). Lysigenous aerenchyma (L).
Bar, 150 µm.
Aerenchyma production involved different
degrees of cell dissolution until the formation of
larger lysigenous lacunae (Figs 3, 4). In the roots,
each lacuna was separated by a variable number of
intact cells, stretching radially from the inner to the
outer cortex. After 15 d, the radius of the control
plants contained several rows of intact cells (Fig. 3a),
the number of rows decreasing with increasing water
depth. A single row remained in waterlogging
conditions (Fig. 3b) and an incomplete row of cells,
or only the remains of cell walls persisted in partial
(Fig. 3c) and total submergence (not shown). Mean-
while, the number of cortical radial cells involved in
the lacunae increased similarly (Fig. 3). The trans-
verse sections also showed that only lateral roots of
control plants developed secondary tissues (Fig. 3a).
Leaves presented constitutive schizogenous
aerenchyma (Fig. 4a–c) and lysogenous aerenchyma
development was limited to the mid-rib leaf blade
(Fig. 4b,c). Transverse sections of tissue from the
leaf base revealed that lacunae were produced in
flooding conditions (Fig. 4b,c) but no lysigenus
aerenchyma was formed in the control plants
(Fig. 4a).
Morphological analysis
Repeated measures ANOVA indicated significant
treatment (P!0±0001) and time effects (P!0±0001)
for the variables analysed. The interaction
treatment-time was significant (P ¯ 0±01), showing
that water level effects differed through time. The
first response registered was the change in the leaf

Printed from the CJO service for personal use only by...
Flooding intensity and Leontodon taraxacoides 123
(a)(a)(a) (b)(b)(b) (c)(c)(c)
LLL
LLL
Figure 4. Transverse sections of leaves of Leontodon taraxacoides plants grown at different water depths (w.d.).
(a) Control. (b) Waterlogging (w.d. 1 cm). (c) Partial submergence (w.d. 7 cm). Arrows point to schizogenous
aerenchyma; lysigenous aerenchyma (L). Bar, 150 µm.
angle and height from a prostrate to a more erect
position in flood conditions. The differences were
observed within 24 h of the beginning of the flood
treatments and corresponded with the water height
imposed in each one of them (Fig. 5). Leaf angle and
height in partially and totally submerged conditions
showed a marked initial change, but little subsequent
change (Fig. 5a,c). The morphology of plants under
total submergence changed steadily until death (Fig.
5). The angle measurements differed significantly
after 10 d (Fig. 5a), as did the height measurements
after 30 d (Fig. 5c), among all treatments. After 10 d,
length of mature leaves were significantly shorter in
plants in waterlogged conditions than in those in the
rest of the treatments (Fig. 5b). After 19 d the leaves
of the controls were longer than those under partial
and total submergence (Fig. 5b).
At the end of the experiment, a great proportion of
the leaves were above the height of 7 cm under
partial submergence conditions (P!0±0001); under
waterlogged conditions, most leaves were positioned
within the 1–7-cm layer (P!0±0001), and in control
conditions the proportion present in the lower layer
was significantly greater than in the rest of the
treatments (P ¯ 0±0094) (Table 1). At harvest, plants
under partial submergence had a graminoid-like
morphology (Fig. 6). Narrowness of blades and
absence of lobes were indicated by the differences
between the maximal (P!0±0001) and minimal (P ¯0±0073) width and in the leaf dissection index
(P!0±0001) (Table 1). Leaves of the waterlogged
plants were narrower but had the same lobe
proportion as the control plants (Fig. 6; Table 1).
Under partial submergence, the diameter of the leaf
mid-rib was larger than in the rest of the treatments
(P ¯ 0±0052), and the area of the mature leaf was
greater in the control plants than in the rest of the
treatments (P ¯ 0±0004) (Table 1).
Biomass and survival
At the beginning of the experiment, the total number
of leaves per plant was 14±2³0±4. Plants continued
producing leaves, except under total submergence,
which caused the death of 50% of individuals after
25 d and of 100% after 40 d of treatment. At the final
measurement the control plants had more leaves and
flower scapes than the plants in the rest of the
treatments. The numbers of leaves and scapes were
greater in the waterlogged plants than in those under
partial submergence (P!0±0001) (Table 2).
The total leaf area per plant was higher in control
plants (P!0±0001), and differences were not found
between the two flood treatments (Table 2). At the
same time, leaf area of the top layer ("7 cm) was
greater under partial submergence than in water-
logged conditions (P ¯ 0±0002). Shallow water
depth corresponded to higher values of plant biomass
(P!0±0001) and higher allocation to reproductive
organs (P ¯ 0±0002) (Table 2). Accordingly, partly
submerged plants allocated more biomass to leaves

Printed from the CJO service for personal use only by...
124 A. A. Grimoldi et al.
(a)90
80
70
60
0
(b)14
12
10
8
12
Mea
n le
af h
eig
ht
(cm
)
Days
50
40
30
20
10
6
9
6
3
00 10 20 4030 50
Mea
n le
af le
ng
th (
cm)
Mea
n le
af a
ng
le (
°)
(c)
Figure 5. Adult leaves of Leontodon taraxacoides plants grown under different water depths (w.d.) : control (+),
waterlogging (w.d. 1 cm) (E), partial submergence (w.d. 7 cm) (*), total submergence (w.d. 13 cm) (D).
(a) Mean leaf insertion angle (°) in relation to soil surface (b) Length (cm). (c) height (cm). Values are
means³SE of five replications.
(P ¯ 0±0297) and stock-root organs (P ¯ 0±0103)
than plants in the rest of the treatments. The total
leaf length:leaves biomass ratio was smaller in the
control plants than in those in conditions of partial
submergence (P ¯ 0±0094) (Table 2).
The proportion of leaf area and biomass positioned
in higher layers increased with higher water levels.
The total leaf area above 7 cm was 2±7³1% in
control, 1±8³0±5% in waterlogged, and, significantly
different from these, 19±0³3% in partly submerged
plants. Waterlogged and partly submerged plants
had greater total leaf area in the 1–7 cm layer
(66³3%) than the controls (47±6³3±9%, P!0±0001).
All treatments differed significantly in the pro-
portion of the leaf area in the 0–1 cm layer
(P!0±0001), control plants had the highest,
49±7³4±5%, and partially submerged plants the
lowest, 14±1³0±7%, (P!0±0001). Under partial
submergence, leaf area in the upper layer
corresponded to 11% of the aboveground biomass,
whereas in the rest of the treatments the proportion
represented 1% (P!0±0001).
The results show a strong negative relationship
between water depth and growth and survival of L.
taraxacoides. In waterlogged and conditions of
partial submergence, plant growth and biomass were
severely affected and, under totally submerged

Printed from the CJO service for personal use only by...
Flooding intensity and Leontodon taraxacoides 125
Table 1. Leaf morphological traits of Leontodon taraxacoides plants
grown at different water depths (w.d.): control, waterlogging (w.d. 1 cm)
and partial submergence (w.d. 7 cm)
Variable Control Waterlogging
Partial
submergence
Maximal leaf width (cm) 1±67³0±10 a 0±91³0±03 b 0±64³0±09 c
Minimal leaf width (cm) 0±54³0±04 a 0±35³0±01 b 0±41³0±05 ab
Leaf dissection index 1±41³0±16 b 1±95³0±26 b 6±49³0±53 a
Leaf area (cm#) 7±90³0±94 a 3±60³0±13 b 3±90³0±65 b
Mid-rib diameter (cm) 0±13³0±01 b 0±12³0±01 b 0±15³0±01 a
Leaves per layer (%)
0–1 cm 32±8³4±6 a 13±8³2±9 b 14±9³4±5 b
1–7 cm 54±8³2±4 b 73±4³4±2 a 24±8³5±5 c
"7 cm 12±4³3±5 b 12±8³4±8 b 60±2³3±3 a
Values are means³SE of five plants harvested at the end of the experiment.
Different letters indicate significant differences (P!0±05) between treatments
based on Tukey test.
(a) (b)
(c)
Figure 6. Shape of leaves of Leontodon taraxacoidesplants grown for 49 d at different water depths (w.d.).
(a) Control. (b) waterlogging (w.d. 1 cm). (c) partial
submergence (w.d. 7 cm). Bar, 1 cm.
conditions, plants could not survive. The anatomical,
morphological and allometric traits of plants growing
in flooded conditions show a wide range of plastic
adjustment, associated with survival under anaerobic
conditions (Blom & Voesenek, 1996).
Anatomical and morphological plastic responses
to flooding in herbaceous plants have been widely
studied (Jackson & Drew, 1984; Armstrong et al.,
1994). Hormones play a central role in the expression
of these traits (Voesenek et al., 1992). Some studies
have found that, in conditions of flooding, the
internal ethylene concentration influences lysigenous
aerenchyma tissue production (Drew et al., 1979),
epinastic curvature of the leaves (Jackson & Drew,
1984) and shoot elongation (Van der Sman et al.,
1991). Accordingly, the vital contribution of
increased leaf height to better aerated and
illuminated zones above or close to the water surface,
and the presence of aerenchyma tissue, which
reduces the resistance to oxygen transport and the
oxygen demand of the plant tissues, has been
demonstrated by various authors (Armstrong, 1979;
Laan & Blom 1990a; Laan et al., 1990b).
In L. taraxacoides, increase of root porosity in
conditions of flooding and root aerenchyma type are
similar to those found in wetland species (Smirnoff
& Crawford, 1983; Justin & Armstrong, 1987). In
some cases, shoot aerenchyma increased significantly
and was apparently accompanied by an increase in
the diameter of the organs involved, such as the
increment of the mid-rib diameter in L. taraxacoides
in conditions of partial submergence (Moon et al.,
1993; Coops et al., 1996). The contrast between the
low root-porosity values found by the pycnometer
method under submergence conditions with those
found by the visual cross-sectional technique sug-
gests a possible collapse of the root radius cells.
Some of the lacunae might be filled with water, which
would affect the pycnometer analysis and root
survival (Sojka, 1988). L. taraxacoides has con-
stitutive schizogenous leaf aerenchyma and develops
lysogenous leaf aerenchyma in response to flooding.
The results showed that lateral roots present
lysogenous aerenchyma that showed a significant
increase in flood conditions; and that root porosity
might gradually decrease in drained-soil conditions.
In this case, the presence of secondary tissues
and less porous organs would probably confer more
resistance to soil mechanical constraints (Engelaar
& Blom, 1995), especially in the dry summer season
when trampling by cattle produces greater increases
in soil bulk density (Taboada & Lavado, 1993). This
fluctuation pattern contrasts with the data available
for Paspalum dilatatum (Rubio et al., 1995; Loreti
& Oesterheld, 1996), a native grass with high flood
tolerance that contains large proportions of aeren-
chyma throughout the year. In this species, the
presence of a sclerenchymatous sheath in the outer
layers of the cortex (Rubio et al., 1995) might provide

Printed from the CJO service for personal use only by...
126 A. A. Grimoldi et al.
Table 2. Leaf area and biomass traits of Leontodon taraxacoides plants
grown at different water depths (w.d.): control, waterlogging (w.d. 1 cm)
and partial submergence (w.d. 7 cm)
Variable Control Waterlogging
Partial
submergence
Scape number 77³5 a 7³1 b 3³1 c
Leaf number 137³12 a 24³2 b 17³2 c
Plant leaf area (cm#)
0–1 cm 479³62 a 23³4 b 7³1 c
1–7 cm 485³94 a 47³4 b 35³2 b
"7 cm 26³11 a 2³1 b 11³2 a
Total 990³150 a 72³7 b 53³3 b
Plant biomass (g)
Heads and scapes 5±57³0±89 a 0±44³0±09 b 0±08³0±04 c
Leaves 4±89³0±75 a 0±51³0±07 b 0±29³0±04 c
Stock and roots 1±95³0±18 a 0±34³0±05 b 0±19³0±07 c
Total 12±41³1±44 a 1±29³0±16 b 0±56³0±09 c
Leaf length}leaf biomass
(cm}g)
246±1³21±2 b 321±4³37±2 ab 474±7³95±0 a
Values are means³SE of five plants harvested at the end of the experiment.
Different letters indicate significant differences (P!0±05) between treatments
based on Tukey test.
a stronger root structure which protects the lacunae
from collapse, which occurs in many flood-tolerant
monocotyledons (Smirnoff & Crawford, 1983).
Several traits of leaf morphology responded to
water depth and, as a result, a high proportion of the
leaf area was positioned relative to the water height
in each treatment. The short reaction time indicates
that plastic changes in epinasty and leaf length
occurred in response to flooding (Van der Sman et
al., 1991). The leaf insertion angle appears to be the
principal factor enabling leaves to emerge above the
water surface. In addition, between flooded treat-
ments leaf elongation was promoted only in those
plants in conditions of partial or total submergence.
Like those for Rumex maritimus, our experimental
data showed complete survival only of the plants
whose leaf tips emerged above the water surface,
showing the vital importance of recovering air–shoot
contact to flood-survival ability (Laan & Blom,
1990a; Van der Sman et al., 1991). Furthermore, in
L. taraxacoides, these responses were followed by a
complete change of leaf shape, involving leaf width
reduction and total absence of lobes under conditions
of partial submergence. Leaf shape responses have
also been documented, especially in aquatic species
in relation to water depth (Bostrack & Millington,
1962; Wooten, 1986). These results show that the
intensity of the stimulus determines the degree and
the number of the morphological responses to
flooding, increasing with environmental stress.
In spite of the reported mechanisms of flood
resistance, reduction in leaf growth rate and plant
biomass indicates a stress condition in flooding
treatments (Chapin, 1980). In L. taraxacoides,
anaerobic and reductive soil conditions seem to
produce metabolic constraints that severely affect
root and shoot growth (Jackson & Drew, 1984). Leaf
growth rate is extremely sensitive to a variety of
stresses (Chapin, 1980), including flooding (Jackson
& Drew, 1984). Leaves of waterlogged plants were
shorter, but changes in leaf angle were sufficient to
raise them above the water level. By contrast, partly
and totally submerged plants elongated their leaves
in a magnitude similar to controls but with less
biomass, as demonstrated by higher leaf length:leaf
biomass ratio. This response indicated that leaf
elongation is the priority for recovery of contact with
the air and survival in conditions of partial sub-
mergence.
Biomass allocation was markedly affected by the
different experimental treatments. Control plants
allocated a great proportion of biomass to repro-
ductive structures; in contrast, flooded plants
increased leaf or stock-root biomass. The greater leaf
biomass proportion in conditions of partial sub-
mergence could be part of a homeostatic strategy to
maximize the acquisition of oxygen, the most
limiting resource under these conditions (Bloom
et al., 1985). It is known that perennial plants com-
monly respond to stress by reducing allocation
to reproductive structures, and do the reverse in
conditions of high resource availability (Chapin,
1980; Bloom et al., 1985). In the Netherlands River
Forelands, it has been found that some perennial
species delay or postpone flowering to the next year
in flooding conditions (Van der Sman et al., 1993).
This strategy was described as stress-tolerant-
ruderal (S-R) (sensu Grime, 1979; Van der Sman
et al., 1993) and corresponds with the established
secondary strategy proposed for L. taraxacoides in

Printed from the CJO service for personal use only by...
Flooding intensity and Leontodon taraxacoides 127
the functional approach to common British species
(Grime et al., 1988). At a greater scale of analysis, it
was proposed that stress-tolerant species have
morphologically fixed traits under stress conditions
(Grime, 1979), but in the case of L. taraxacoides,
phenotypic plasticity might play a central role in
reducing adverse environmental effects (Bradshaw,
1965; Via et al., 1995), especially in conditions of
partial flooding (Laan & Blom, 1990a; Laan et al.,
1990b).
It has been shown that plastic responses could
influence the process of species invasion (Meerts,
1992; Williams et al. 1995). L. taraxacoides occurs in
dry and humid open sites of European grasslands
(Rivas & Rivas, 1963; Pignatti, 1982, Grime et al.,
1988), but in the Flooding Pampa Grassland it is
present only in humid-lowland plant communities
(Burkart et al., 1990). The constitutive and plastic
features related to flood-survival could indicate an
adjustment of this species to the characteristics of
those invaded communities (Noble, 1989). Grazing
facilitates invasion and subsequent dominance of
alien dicots, such as L. taraxacoides in the Flooding
Pampa Grasslands (Oesterheld & Sala, 1990).
However, after flooding, native monocots generally
increase their presence and exotic dicots suffer
significant decreases in biomass and abundance
(Chaneton et al., 1988; Insausti, 1996). Adult
L. taraxacoides plants, nevertheless, can survive
slight spring floods, through a certain ability to
survive under root anaerobiosis. In contrast, L.
taraxacoides does not endure intense and long-lasting
floods when totally submerged. The flooding stress
effects that were detected in this study corroborate
the results found in experiments at the community
scale (Chaneton et al., 1988; Insausti, 1996) and
show that flooding intensity strongly influences the
demography of this species.
We especially thank Dorita Grimoldi for her technical
assistance in all phases of this work. We thank Professor
Mo! nica Tourn, who helped with the anatomical analysis,
and P. Quinos, J. Loreti, M. Omacini, M. Nogue! s Loza
and two anonymous reviewers, who helped to improve the
manuscript. We also thank the Bordeau family, owners of
Estancia Las Chilcas, who facilitated our work on their
land. Agustı!n Grimoldi was supported by a fellowship
from CONICET.
Armstrong W. 1979. Aeration in higher plants. Advances in
Botanical Research 7 : 225–332.
Armstrong W, Bra$ ndle R, Jackson MB. 1994. Mechanisms of
flood tolerance in plants. Acta Botanica Neerlandica 43 :
307–358.
Blom CWPM, Voesenek LACJ. 1996. Flooding: the survival
strategies of plants. Trends in Ecology and Evolution 11 :
290–295.
Bloom AJ, Chapin III FS, Mooney HA. 1985. Resource
limitation in plants – an economic analogy. Annual Review of
Ecology and Systematics 16 : 363–392.
Bostrack JM, Millington WF. 1962. On the determination of
leaf form in an aquatic heterophyllous species of Ranunculus.
Bulletin of the Torrey Botanical Club 89 : 1–20.
Bradshaw AD. 1965. Evolution significance of phenotypic
plasticity in plants. Advances in Genetics 13 : 115–155.
Burkart SE, Leo! n RJC, Movia CP. 1990. Inventario
fitosociolo! gico del pastizal de la Depresio! n del Salado (Prov. de
Bs. As.) en un a! rea representativa de sus principales ambientes.
Darwiniana 30 : 27–69.
Chaneton EJ, Facelli JM, Leo! n RJC. 1988. Floristic changes
induced by flooding on grazed and ungrazed lowland grasslands
in Argentina. Journal of Range Management 41 : 497–501.
Chapin III FS. 1980. The mineral nutrition of wild plant. Annual
Review of Ecology and Systematics 11 : 233–260.
Coops H, Van der Brink FWB, Van der Velde G. 1996. Growth
and morphological responses of four helophyte species in an
experimental water-depth gradient. Aquatic Botany 54 : 11–24.
Drew MC, Jackson MB, Giffard S. 1979. Ethylene-promoted
adventitious rooting and development of cortical air spaces
(aerenchyma) in roots may be adaptive responses to flooding in
Zea mays L. Planta 147 : 83–88.
Engelaar WMHG, Blom CWPM. 1995. Effects of flooding and
trampling on the performance of river foreland species of
Rumex and Plantago. Acta Botanica Neerlandica 44 : 225–245.
Grime JP. 1979. Plant strategies and vegetation processes. New
York, USA: J Wiley and Sons.
Grime JP, Hodgson JG, Hunt R. 1988. Comparative plant
ecology. A functional approach to common British species.
London, UK: Unwin Hyman.
Insausti P. 1996. Respuestas estructurales y funcionales a las
inundaciones de un pastizal de la DepresioU n del Salado (Prov. de
Buenos Aires). M.Sc. thesis, Universidad de Buenos Aires,
Argentina.
Jackson MB, Drew MC. 1984. Effects of flooding on growth and
metabolism of herbaceous plants. In: Kozlowski TT, ed.
Flooding and plant growth. London, UK: Academic Press,
49–128.
Justin HFW, Armstrong W. 1987. The anatomical charac-
teristics of roots and plant response to soil flooding. New
Phytologist 106 : 465–495.
Laan P, Blom CWPM. 1990a. Growth and survival responses of
Rumex species to flooded and submerged conditions: the
importance of shoot elongation, underwater photosynthesis and
reserves carbohydrates. Journal of Experimental Botany 41 :
775–783.
Laan P, Tosserams M, Blom CWPM, Veen BW. 1990b.
Internal oxygen transport in Rumex species and its significance
for respiration under hypoxic conditions. Plant and Soil 122 :
39–46.
Loreti J, Oesterheld M. 1996. Intraspecific variation in the
resistance to flooding and drought in populations of Paspalum
dilatatum from different topographic positions. Oecologia 108 :
279–84.
Meerts P. 1992. An experimental investigation of life history and
plasticity in two cytotypes of Polygonum aviculare L. subsp.
avidulare that coexist in an abandoned arable field. Oecologia
92 : 442–449.
Menges ES, Waller DM. 1983. Plant strategies in relation to
elevation and light in floodplain herbs. The American Naturalist
122 : 454–473.
Moon M, Rattray MR, Putz FE, Bowes G. 1993.
Acclimatization to flooding of the herbaceous vine, Mikania
scandens. Functional Ecology 7 : 610–615.
Noble IR. 1989. Attributes of invaders and the invading process:
terrestrial and vascular plants. In: Drake JA, ed. Biological
invasions: a global perspective. Chichester, UK: John Wiley &
Sons, 301–313.
Oesterheld M, Sala OE. 1990. Effects of grazing on seedling
establishment: the role of seed and safe-site availability. Journal
of Vegetation Science 1 : 353–358.
Pignatti S. 1982. Flora D’Italia. Bologna, Italy: Edagricole.
Ponnamperuma FN. 1984. Effects on flooding on soils. In:

Printed from the CJO service for personal use only by...
128 A. A. Grimoldi et al.
Kozlowski TT, ed. Flooding and plant growth. London, UK:
Academic Press Inc., 9–45.
Rivas S, Rivas S. 1963. Estudio y clasificacioU n de los pastizales
espanh oles. Madrid, Spain: MAPA.
Rubio G, Casasola G, Lavado RS. 1995. Adaptations and
biomass production of two grasses in response to waterlogging
and soil nutrient enrichment. Oecologia 102 : 102–105.
Smirnoff N, Crawford RMM. 1983. Variation in the structure
and response to flooding of root aerenchyma in some wetland
plants. Annals of Botany 51 : 237–249.
Sojka RE. 1988. Measurement of root porosity (volume of root air
space). Environmental and Experimental Botany 28 : 275–280.
Soriano A. 1991. Rio de La Plata grasslands. In: Coupland RT,
ed. Temperate subhumid grasslands. Ecosystems of the world.
Natural Grasslands, vol 8A. Amsterdam, The Netherlands:
Elsevier, 379–382.
Taboada ME, Lavado RS. 1986. Caracterı!sticas del re!gimen
acuico de un Natracuol de la Pampa Deprimida. Ciencia del
Suelo 4 : 66–71.
Taboada ME, Lavado RS. 1993. Influence of cattle trampling on
soil porosity under alternate dry and ponded conditions. Soil
Use and Management 9 : 139–143.
Van der Sman AJM, Joosten NN, Blom CWPM. 1993.
Flooding regimes and life-history characteristics of short-lived
species in river forelands. Journal of Ecology 81 : 121–103.
Van der Sman AJM, Voesenek LACJ, Blom CWPM, HarrenFJM, Reuss J. 1991. The role of ethylene in shoot elongation
with respect to survival and seed output of flooded Rumex
maritimus L. plants. Functional Ecology 5 : 304–313.
Via S, Gomulkiewicz R, De Jong G, Sheiner SM, SchlichtingCD, Van Tienderen PH. 1995. Adaptive phenotypic plasticity:
consensus and controversy. Trends in Ecology and Evolution 10 :
212–217.
Voesenek LACJ, Van der Sman AJM, Harren FJM and BlomCWPM. 1992. An amalgamation between hormone physiology
and plant ecology: a review on flooding resistance and ethylene.
Journal of Plant Growth Regulation 11 : 171–188.
Von Ende CN. 1993. Repeated-measures analysis : growth and
other time-dependent measures. In: Scheiner SM, Gurevitch J,
eds. Design and analysis of ecological plants. New York, USA:
Chapman & Hall, 113–137.
Williams DG, Mack RN, Black RA. 1995. Ecophysiology of
introduced Pennisetum setaceum on Hawaii : the role of pheno-
typic plasticity. Ecology 76 : 1569–1580.
Wooten JW. 1986. Variations in leaf characteristics of six species
of Sagitaria (Alismataceae) caused by various water levels.
Aquatic Botany 23 : 321–327.




















