
Response to Histamine Allows the Functional Identification of Neuronal Progenitors, Neurons,...
14
REJUVENATION RESEARCH Volume 11, Number 1, 2008 © Mary Ann Liebert, Inc. DOI: 10.1089/rej.2007.0600 Response to Histamine Allows the Functional Identification of Neuronal Progenitors, Neurons, Astrocytes, and Immature Cells in Subventricular Zone Cell Cultures Fabienne Agasse,* Liliana Bernardino,* Bruno Silva, Raquel Ferreira, Sofia Grade, and João O. Malva ABSTRACT Subventricular zone (SVZ) cell cultures contain mixed populations of immature cells, neu- rons, astrocytes, and progenitors in different stages of development. In the present work, we examined whether cell types of the SVZ could be functionally discriminated on the basis of intracellular free calcium level ([Ca 2 ] i ) variations following KCl and histamine stimulation. For this purpose, [Ca 2 ] i were measured in SVZ cell cultures from neonatal P1-3 C57Bl/6 donor mice, in single cells, after stimulation with 100 M histamine or 50 mM KCl. MAP-2-positive neurons and doublecortin-positive neuroblasts were distinguished on the basis of their se- lective ratio of response to KCl and/or histamine stimulation. Moreover, we could distinguish immature cells on the basis of the selective response to histamine via the histamine 1 recep- tor activation. Exposure of SVZ cultures to the pro-neurogenic stem cell factor (SCF) induced an increase in the number of cells responding to KCl and a decrease in the number of cells responding to histamine, consistent with neuronal differentiation. The selective response to KCl/histamine in single cell calcium imaging analysis offers a rapid and efficient way for the functional discrimination of neuronal differentiation in SVZ cell cultures, opening new per- spectives for the search of potential pro-neurogenic factors. 187 INTRODUCTION R EYNOLDS AND WEISS 1 identified a resident population of stem cells in the SVZ of the mouse brain, endowed with proliferative and self-renewal capacities and able to generate neurons and glial cells. Since then, stem cells have been identified in the SVZ of other mam- malian species, including primates. 2,3 SVZ-de- rived neuroblasts migrate along the rostral mi- gratory stream towards the olfactory bulb (OB), where they functionally differentiate into in- terneurons, involved in odor discrimination. 4–8 Characterization of SVZ neurons and neu- ronal precursors is mostly performed by im- munocytochemistry. SVZ cultures are mixed cultures of immature cells, neurons, astrocytes, oligodendrocytes, and neuronal and glial pro- Neuroprotection and Neurogenesis in Brain Repair Group, Center for Neuroscience and Cell Biology, Institute of Biochemistry, Faculty of Medicine, University of Coimbra, Coimbra, Portugal. *These authors contributed equally to this work.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Response to Histamine Allows the Functional Identification of Neuronal Progenitors, Neurons,...

REJUVENATION RESEARCHVolume 11, Number 1, 2008© Mary Ann Liebert, Inc.DOI: 10.1089/rej.2007.0600
Response to Histamine Allows the FunctionalIdentification of Neuronal Progenitors, Neurons,Astrocytes, and Immature Cells in Subventricular
Zone Cell Cultures
Fabienne Agasse,* Liliana Bernardino,* Bruno Silva, Raquel Ferreira, Sofia Grade, and João O. Malva
ABSTRACT
Subventricular zone (SVZ) cell cultures contain mixed populations of immature cells, neu-rons, astrocytes, and progenitors in different stages of development. In the present work, weexamined whether cell types of the SVZ could be functionally discriminated on the basis ofintracellular free calcium level ([Ca2�]i) variations following KCl and histamine stimulation.For this purpose, [Ca2�]i were measured in SVZ cell cultures from neonatal P1-3 C57Bl/6 donormice, in single cells, after stimulation with 100 �M histamine or 50 mM KCl. MAP-2-positiveneurons and doublecortin-positive neuroblasts were distinguished on the basis of their se-lective ratio of response to KCl and/or histamine stimulation. Moreover, we could distinguishimmature cells on the basis of the selective response to histamine via the histamine 1 recep-tor activation. Exposure of SVZ cultures to the pro-neurogenic stem cell factor (SCF) inducedan increase in the number of cells responding to KCl and a decrease in the number of cellsresponding to histamine, consistent with neuronal differentiation. The selective response toKCl/histamine in single cell calcium imaging analysis offers a rapid and efficient way for thefunctional discrimination of neuronal differentiation in SVZ cell cultures, opening new per-spectives for the search of potential pro-neurogenic factors.
187
INTRODUCTION
REYNOLDS AND WEISS1 identified a residentpopulation of stem cells in the SVZ of the
mouse brain, endowed with proliferative andself-renewal capacities and able to generateneurons and glial cells. Since then, stem cellshave been identified in the SVZ of other mam-malian species, including primates.2,3 SVZ-de-
rived neuroblasts migrate along the rostral mi-gratory stream towards the olfactory bulb (OB),where they functionally differentiate into in-terneurons, involved in odor discrimination.4–8
Characterization of SVZ neurons and neu-ronal precursors is mostly performed by im-munocytochemistry. SVZ cultures are mixedcultures of immature cells, neurons, astrocytes,oligodendrocytes, and neuronal and glial pro-
Neuroprotection and Neurogenesis in Brain Repair Group, Center for Neuroscience and Cell Biology, Institute ofBiochemistry, Faculty of Medicine, University of Coimbra, Coimbra, Portugal.
*These authors contributed equally to this work.

genitors in different stages of differentia-tion.1,9,10 Immature SVZ cells express the in-termediate filament protein nestin.9,11 Earlyneuronal commitment is detected through theexpression of the neuron-associated class IIItubulin isotype �.12 Migrating immature SVZneuroblasts express doublecortin (DCX), a microtubule-associated protein,13,14 and thepolysialylated neural cell adhesion molecule(PSA-NCAM).15,16 As maturation proceeds,SVZ-derived post-mitotic neurons expressMAP-2, neurofilament (NF), the enzyme neu-ron-specific enolase (NSE),17 and NeuN.18
However, immunocytochemistry is time con-suming, does not give information about func-tion, and is performed on fixed, or dead, cells,hampering their subsequent use for therapeu-tic purposes.
Measurement of intracellular free calciumvariations is useful to rapidly characterizefunctional cells. Indeed, membrane depolar-ization of excitable cells, such as neurons, fol-lowing exposure to high KCl concentrationsleads to the opening of voltage sensitive cal-cium channels (VSCC) and massive influx ofcalcium into the cytoplasm.19 Immature/stemcells, such as embryonic stem cells, carcinomaand astrocytoma cells express functional hista-mine receptors.20–25 Although the specific roleof histamine in neurogenesis and neuronal de-velopment is poorly understood, the specificexpression of histamine receptors may be usedas a marker for undifferentiated neural pro-genitors. Indeed, stimulation of immature/stem cells with histamine transiently increase[Ca2�]i,20,22,26 in immature precursor cells fromthe postnatal and adult SVZ.25
In the present work, we describe a rapidmethod to functionally characterize neuronaldifferentiation in SVZ cultures, based on thespecific [Ca2�]i variations according to celltype, following KCl depolarization and stimu-lation with histamine.
MATERIALS AND METHODS
All experiments were performed in accor-dance with NIH and European (86/609/EEC)guidelines for the care and use of laboratoryanimals.
SVZ cultures
SVZ cells were obtained from 1- to 3-day-oldC57Bl/6 donor mice. Brains were removed fol-lowing decapitation and placed in HBSS solu-tion (Gibco, Rockville, MD). Fragments of SVZwere dissected out of 450 �m thick coronalbrain sections, digested in 0.025% trypsin and0.265 mM EDTA (Gibco), and dissociated bygentle trituration with a P1000 pipette. The cellsuspension was diluted in serum-free culturemedium (SFM) composed of Dulbecco’s mod-ified eagle medium (D-MEM/F12 � Gluta-MAX™-I, Gibco) supplemented with 100U/mL penicillin, 100 �g/mL streptomycin(Gibco), 1% B27 (Gibco), 10 ng/mL epidermalgrowth factor (EGF; Gibco), and 10 ng/mL ba-sic fibroblast growth factor (FGF-2, Gibco). Sin-gle cells were then plated on uncoated Petridishes at a density of 3000 cells/cm2. The neu-rospheres were allowed to develop in a 95%air–5% CO2 humidified atmosphere at 37°C.
Six to eight days after plating, the neu-rospheres were collected with a Pasteur pipetteand seeded onto poly-D-lysine coated glasscover slips placed into 12-well cell cultureplates for calcium imaging experiments or 24-well cell culture plates for immunocytochem-istry, and then covered with 1 mL or 500 �L,respectively, of SFM devoid of growth factors.
For the experiments correlating single cellcalcium imaging (SCCI) with immunodetec-tion, SVZ neurospheres were seeded onto poly-D-lysine coated microgrid cover slips (Eppen-dorf CELLocate, Hamburg, Germany).
To evaluate neuronal differentiation in SVZcultures, neurospheres were allowed to de-velop over 7 days at 37°C in the absence orpresence of 10 ng/mL LIF or 20 ng/mL SCF(both from Chemicon International, Temecula,CA).
Single cell calcium imaging
To determine the differentiation pattern ofSVZ cells, we analyzed the variations of intra-cellular calcium levels following stimulationwith 50 mM KCl and 100 �M histamine (Sigma,St. Louis, MO), in accordance to the sequenceshown in Figure 1A. KCl-depolarization causesthe increase of the intracellular calcium levelsin neurons19 whereas stimulation with hista-
AGASSE ET AL.188
mine leads to the increase of intracellular cal-cium levels in stem/progenitor cells.25
SVZ cultures were loaded for 40 min at 37°Cwith 5 �M Fura-2 AM (Molecular Probes, Eu-gene, OR), 0.1% fatty acid free BSA, and 0.02%pluronic acid F-127 in Krebs (132 mM NaCl, 1mM KCl, 1 mM MgCl2, 2.5 mM CaCl2, 10 mMglucose, and 10 mM HEPES [pH 7.4]). After a10-min post-loading period at room tempera-ture the glass cover slip was mounted on RC-20 chamber in a PH3 platform (Warner Instru-ments, Hamden, CT) on the stage of an invertedfluorescence microscope Axiovert 200 (CarlZeiss, Göttingen, Germany). Cells were contin-
uously perfused with Krebs solution and stim-ulated by applying 100 �M histamine or highpotassium Krebs solution (containing 50 mMKCl, isosmotic substitution with NaCl). Solu-tions were added to the cells by a fast-pres-surized (95% air, 5% CO2 atmosphere) system(AutoMate Scientific, Berkeley, CA). The intra-cellular calcium concentration ([Ca2�]i) wasevaluated by quantifying the ratio of the fluo-rescence emitted at 510 nm following alternateexcitation (750 ms) at 340 nm and 380 nm, usinga Lambda DG4 apparatus (Sutter Instruments,Novato, CA) and a 510 nm bandpass filter (CarlZeiss)12 before fluorescence acquisition with a
FUNCTIONAL IDENTIFICATION OF NEW NEURONS 189
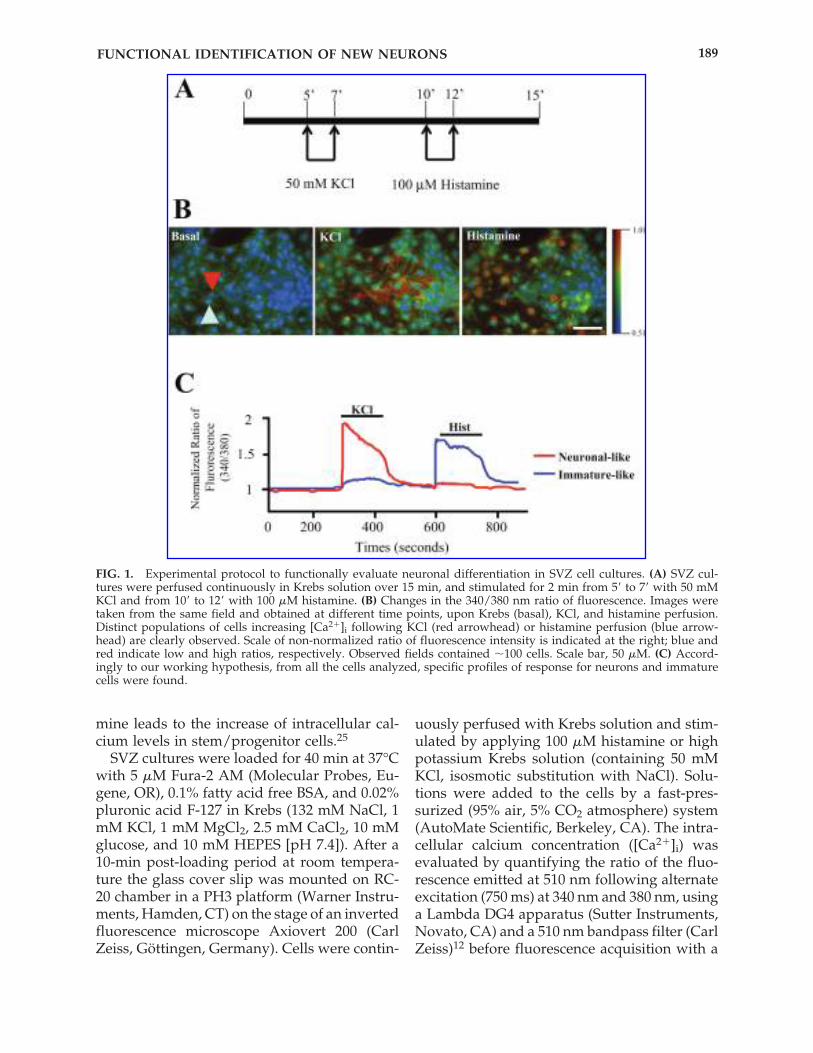
FIG. 1. Experimental protocol to functionally evaluate neuronal differentiation in SVZ cell cultures. (A) SVZ cul-tures were perfused continuously in Krebs solution over 15 min, and stimulated for 2 min from 5� to 7� with 50 mMKCl and from 10� to 12� with 100 �M histamine. (B) Changes in the 340/380 nm ratio of fluorescence. Images weretaken from the same field and obtained at different time points, upon Krebs (basal), KCl, and histamine perfusion.Distinct populations of cells increasing [Ca2�]i following KCl (red arrowhead) or histamine perfusion (blue arrow-head) are clearly observed. Scale of non-normalized ratio of fluorescence intensity is indicated at the right; blue andred indicate low and high ratios, respectively. Observed fields contained �100 cells. Scale bar, 50 �M. (C) Accord-ingly to our working hypothesis, from all the cells analyzed, specific profiles of response for neurons and immaturecells were found.

40x objective and a Coll SNAP digital camera(Roper Scientific, Tucson, AZ). Acquired val-ues were processed using the MetaFluor soft-ware (Universal Imaging, Downingtown, PA).
Histamine/KCl values for Fura-2 ratio werecalculated to determine the extent of neuronalmaturation in cultures. The results obtained inSVZ cultures were compared with those ob-tained in cortical glial cell cultures or in cul-tured hippocampal neurons.27
To determine which histamine receptor wasinvolved in the effect of histamine, we analyzedthe variations of [Ca2�]i following perfusion ofthe cells with 100 �M histamine concomitantlywith either 1 �M mepyramine or 50 �M ci-metidine (both from Tocris, Ellisville, MO),specific antagonists for the histamine receptors1 (H1R) and 2 (H2R), respectively,28 in agree-ment with the sequence shown in Figure 1B.
Immunocytochemistry
After fixation, for 1 h in 4% paraformalde-hyde, cells were permeabilized and non-specificbinding sites were blocked for 1.5 h with 0.25%Triton X-100 (Sigma) and 6% bovine serum al-bumin (BSA, Sigma) dissolved in PBS. Cellswere then subsequently incubated overnight at 4°C with the following primary antibodies:mouse monoclonal anti-nestin (1:200, Chemi-con), mouse monoclonal anti-MAP-2 antibody(1:200, Sigma), rabbit monoclonal anti-GFAPantibody (1:1000, Sigma), mouse monoclonalanti-NeuN antibody (1:100, Chemicon), rabbitpolyclonal anti-doublecortin antibody (1:200,Cell Signaling, Danvers, MA). Thereafter, thecover slips were rinsed in PBS and incubatedfor 1 h at room temperature with the secondarygoat anti-rabbit Alexa Fluor 488 antibody (1:200,Molecular Probes) or goat anti-mouse AlexaFluor 594 antibody (1:200, Molecular Probes),respectively. After rinsing with PBS, cell prepa-rations were incubated 5 min at room temper-ature with Hoechst 33342 (2 �g/mL, MolecularProbes) in PBS containing 0.25% BSA for nu-clear staining. Finally, the preparations weremounted using Dakocytomation fluorescentmedium (Dakocytomation, Carpinteria, CA).
Fluorescence images were recorded using adigital camera coupled to an Axioskop micro-scope (Carl Zeiss).
Isolation of total RNA from mouse SVZ cells
Total RNA was isolated from SVZ cells us-ing Tri Reagent (Sigma) according to the man-ufacturer’s instructions. Cells were gently ho-mogenized in guanidium thiocyanate andphenol. Chloroform was added allowing aclear isolation of RNA in the resultant aqueousphase. The RNA was precipitated with iso-propanol and the pellet washed with 75% eth-anol. The pellet was eluted in diethylpyrocar-bonate (DEPC)-treated water.
The total amount of RNA was quantified byoptical density (OD) measurements at 260 nm,and the purity was evaluated by measuring theratio of OD at 260 and 280 nm (RNA/DNA calculator GeneQuant II, Pharmacia BiotechAmersham Biosciences, Uppsala, Sweden). Ad-ditionally, RNA quality was assessed by gelelectrophoresis.
RNA extracted from splenocytes was used asa positive control for the detection of histaminereceptor 1 and 2 expression.
RT-PCR analysis
Histamine receptor 1 and 2 mRNA expres-sion was determined by reverse transcription-PCR (RT-PCR). First, cDNA was obtained fromthe transcription of 2 �g RNA using avian mi-croblastosis virus (AMV) reverse transcriptaseand Oligo-p(dT)15 primers (Roche MolecularBiochemicals, Indianapolis, IN). PCR was per-formed in a 50 �L reaction system (Roche Mo-lecular Biochemicals) containing 5 �L templatecDNA, 1 �L deoxynucleotide mix, 5 �L 10xPCR reaction buffer, 0.2 �L upstream and 0.2�L downstream primer, a variable volume ofwater, and 0.25 �LTaq DNA polymerase (35 cy-cles at 95°C for 30 s; at 56°C/58°C (histaminereceptor 1/histamine receptor 2) for 30 s andat 72°C for 30 s) (Bioron, Ludwigshafen, Ger-many).
Primers used in PCR reactions were as fol-lows. Histamine receptor 1: forward primer 5�-GGG CTC AAA GGC CAA TGA C-3� and re-verse primer 5�-TCC GCC GGC AAG TACTCA-3�. Histamine receptor 2: forward primer5�-CTG GCT GTC AGC TTG AAT CG-3� andreverse primer 5�-GCT GCC AGG GAC ACAATG A-3�. �-actin: forward primer 5�-GACTAC CTC ATG AAG ATC CT-3� and reverse
AGASSE ET AL.190

primer 5�-ATC TTG ATC ATG GTG CTG-3�(MWG-Biotech, Ebersberg, Germany).29
PCR products of each sample were sub-jected to electrophoresis in a 1.5% agarose geland stained with ethidium bromide. Pho-tographs were taken in a Versa-Doc ImagingSystem (Model 3000, Bio-Rad Laboratories,Hercules, CA).
Data analysis and statistics
In all experiments, the experimental condi-tion was run at least in triplicate. For SCCI ex-periments, the cell percentage was calculatedon the basis of one field per each coverslip con-taining about 100 cells.
Percentage of NeuN immunoreactive cellswere calculated from cell counts in five inde-pendent fields in each coverslip with a 40x ob-jective (about 200 cells per field). Because nosignificant differences were found betweencontrols, the corresponding data were pooledand expressed as mean � SEM. Statistical sig-nificance of differences was examined by one-way ANOVA followed by the Bonferroni post-test for multiple comparisons or unpaired t-test.Statistical significance level was set for (*) p val-ues � 0.05; (**) p values � 0.01; (***) p values �0.001.
RESULTS
Variations in [Ca2�]i following stimulation withKCl or histamine
We characterized the profile of [Ca2�]i vari-ations in single SVZ cells upon KCl and hista-mine stimulation. Six- to eight-day-old neu-rospheres were plated onto poly-D-lysinecoated coverslips and allowed to develop dur-ing 7 days in SFM medium devoid of growthfactors. The cells were then loaded with theFura-2 AM calcium probe, perfused continu-ously for 15 min with Krebs solution, andshortly (2 min) stimulated with 50 mM KCl orwith 100 �M histamine (Fig. 1A). Changes in340/380 nm ratios of fluorescence were moni-tored constantly during all the experiment.Figure 1B provides an example of [Ca2�]i vari-ations in SVZ cells upon KCl and histaminestimulation. Interestingly, some cells display a
rise in the [Ca2�]i following KCl stimulationbut not following histamine perfusion, consis-tent with a neuronal-like profile. Moreover, animmature-like profile, characterized by an ab-sence of response to KCl but a rise in [Ca2�]ifollowing histamine stimulation was alsofound (Fig. 1B and C).
To associate profiles of [Ca2�]i variationswith the respective cell phenotype, we moni-tored [Ca2�]i responses using SCCI and thenperformed subsequent immunocytochemicalcharacterization on SVZ cells cultivated on mi-crogrid cover slips, allowing cell localization.We show that the neuronal-like profile of[Ca2�]i variations is displayed by MAP-2 ex-pressing neurons (Fig. 2A). Another populationof cells responding to KCl, weakly howeverthan MAP-2-positive neurons, and with minorresponse to histamine, expresses the immatureneuronal marker DCX (Fig. 2B). Cells respond-ing neither to KCl nor to histamine are GFAP-positive cells (Fig. 2C).
We then calculated the ratio of responses dueto histamine or KCl exposure (Hist/KCl) in atotal of 10 MAP-2 positive neurons, 10 DCX-positive neuroblasts, and 8 GFAP expressingcells. The Hist/KCl values were significantlydifferent between groups, as shown in Figure2D, indicating that following KCl and hista-mine stimulation, SVZ-derived neurons andneuroblasts can be discriminated according tothe Hist/KCl ratio (MAP-2-positive cells,0.69 � 0.02; DCX positive cells, 0.86 � 0.02;GFAP-positive cells, 1.02 � 0.01; p � 0.001 be-tween each cell phenotype). Additionally, ra-tios displayed by both mature and immatureneurons were significantly lower than the onesfound in GFAP-positive cells, since these cellsdo not respond to stimulation with KCl, andhence display a normalized Hist/KCl ratioclose to or above one. It is possible that a sub-set of cells, mainly neuronal progenitors, stillrespond to histamine and start to respond toKCl and eventually show a net ratio of calciumtransients close to 1. However, these cells canbe distinguished from GFAP-positive cells onthe basis of their different profile of response.In the present work, two factors were indeedconsidered: the ratio of response Hist/KCl andthe profile of response. Thus, neuronal cells canbe undoubtedly discriminated from glial cells.
FUNCTIONAL IDENTIFICATION OF NEW NEURONS 191
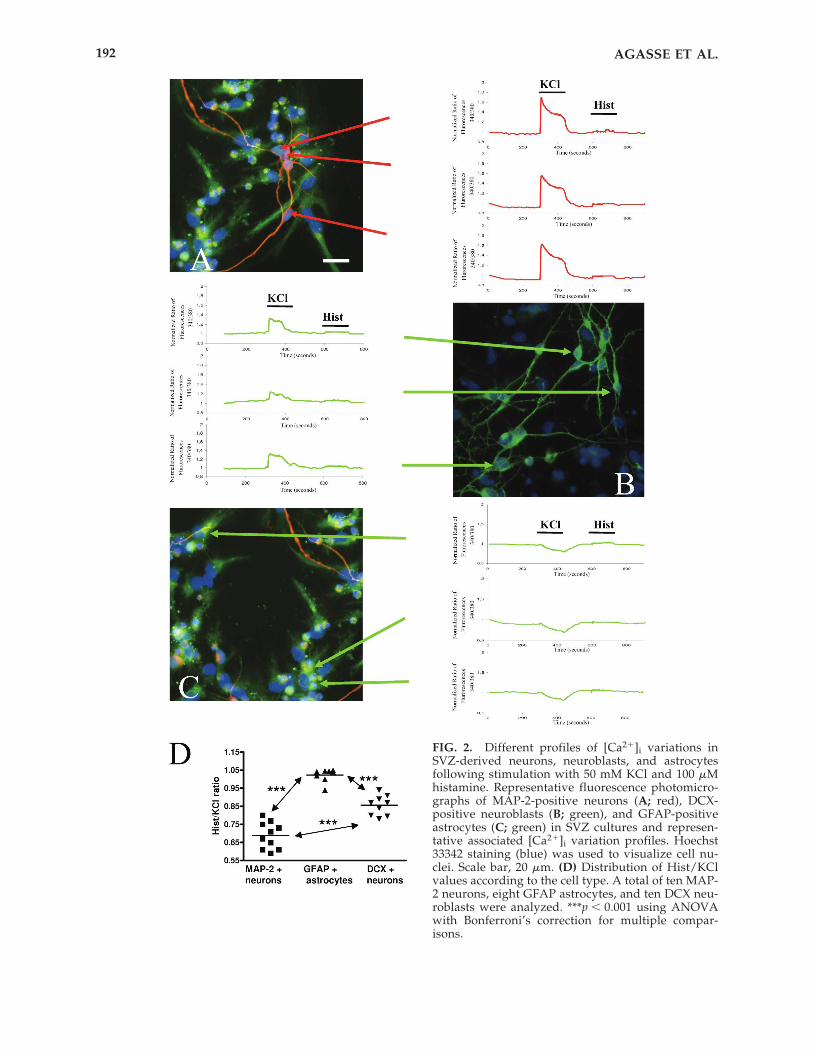
FIG. 2. Different profiles of [Ca2�]i variations inSVZ-derived neurons, neuroblasts, and astrocytesfollowing stimulation with 50 mM KCl and 100 �Mhistamine. Representative fluorescence photomicro-graphs of MAP-2-positive neurons (A; red), DCX-positive neuroblasts (B; green), and GFAP-positiveastrocytes (C; green) in SVZ cultures and represen-tative associated [Ca2�]i variation profiles. Hoechst33342 staining (blue) was used to visualize cell nu-clei. Scale bar, 20 �m. (D) Distribution of Hist/KClvalues according to the cell type. A total of ten MAP-2 neurons, eight GFAP astrocytes, and ten DCX neu-roblasts were analyzed. ***p � 0.001 using ANOVAwith Bonferroni’s correction for multiple compar-isons.
AGASSE ET AL.192

Similar SCCI experiments were performed inSVZ cultures subsequently stained for themarker of immature cells, nestin. As shown inFigure 3A, cells responding to histamine andnot to KCl express nestin, suggesting that[Ca2�]i response following histamine stimula-tion is restricted to immature/undifferentiated
cells in SVZ cultures. However, nestin is ex-pressed by a wide range of SVZ cell types.30
Accordingly, nestin-positive cells expressingGFAP, a marker for mature astrocytes, arefound in SVZ cultures (Fig. 3D, cell b), as wellas nestin�/GFAP� (Fig. 3D, cell a), andnestin�/GFAP� (Fig. 3D, cell c) cells. SCCI ex-
FUNCTIONAL IDENTIFICATION OF NEW NEURONS 193
FIG. 3. Representative fluorescence photomicrographs of nestin (A–C; red) and GFAP-positive cells (B, C; green) inSVZ cultures and the respective associated [Ca2�]i variation profiles of nestin-positive cells (A) co-expressing or not(C and B, respectively) GFAP, following stimulation with 50 mM KCl and 100 �M histamine. (D) Fluorescence con-focal photomicrograph of nestin (red) and GFAP (green) immunodetection in SVZ cell culture. Hoechst 33342 stain-ing (blue) was used to visualize cell nuclei. Scale bar, 20 �m.

periments were then conducted in SVZ cul-tures followed by double immunodetection ofnestin and GFAP markers. Immature-like pro-file of [Ca2�]i was found associated with bothnestin�/GFAP� cells as well as nestin�/GFAP� cells (Fig. 3B and C, respectively).
All the cells analyzed responding to hista-mine displayed immunoreactivity to nestin (12cells analyzed), but not all the nestin-im-munoreactive cells were found to respond tohistamine stimulation (10 cells analyzed). Thisis in accordance with the well-established factthat nestin is not a selective marker of imma-ture cells since it is also expressed by SVZ-de-rived differentiated cells.30 These results fur-ther demonstrate that increase in the bulkcytoplasmic [Ca2�]i following histamine stim-ulation is restricted to SVZ immature/undif-ferentiated cells. Furthermore, mature neuronscan be discriminated from immature cells ac-cording to their low and high Hist/KCl ratio,respectively.
Increase in [Ca2�]i in SVZ immature cells ismediated by histamine H1 receptor activation
To further investigate which histamine re-ceptor was involved in histamine-induced[Ca2�]i responses, we performed RT-PCR todetect histamine receptors H1R and H2R inSVZ cells. mRNA for both receptors werefound in SVZ cells (Fig. 4A).
To evaluate the involvement of each recep-tor in the increase of [Ca2�]i following hista-mine stimulation, we performed single cell cal-cium imaging studies in the presence of specificantagonists for H1R and H2R. Cells were con-tinuously perfused with Krebs solution for 20min and stimulated with KCl, histamine, or his-tamine plus 1 �M mepyramine, an H1R antag-onist, or with 100 �M histamine plus 50 �M cimetidine, an H2R antagonist (Fig. 4B). Rep-resentative profiles of response of immatureSVZ cells are shown in Figure 4C. Moreover,we also calculated the ratios of response fol-lowing exposure to histamine and histamine-plus antagonist. Accordingly, we evaluated thepercentage of histamine-sensitive SVZ cellsthat still responded to histamine in the pres-ence of the antagonists (ratio histamine/hista-mine�antagonist close to 1). Following stimu-lation with mepyramine, a significant decrease
of 92% in the number of immature cells re-sponding to histamine was observed, whereasstimulation with cimetidine did not block[Ca2�]i responses induced by histamine (�mepyramine, 7.67 � 1.33%; with p � 0.001 ascompared with total histamine-respondingcells (307 cells analyzed) normalized at100%; � cimetidine, 89.3 � 9.21%) (Fig. 4D).
Moreover, by performing double immuno-cytochemistry against H1R and nestin, weshowed that immature SVZ cells express thehistamine 1 receptor (Fig. 4A, right image).
Variations of [Ca2�]i in neurons, glia, andprecursor cells following stimulation with KCl orhistamine: evaluation of neuronal differentiation
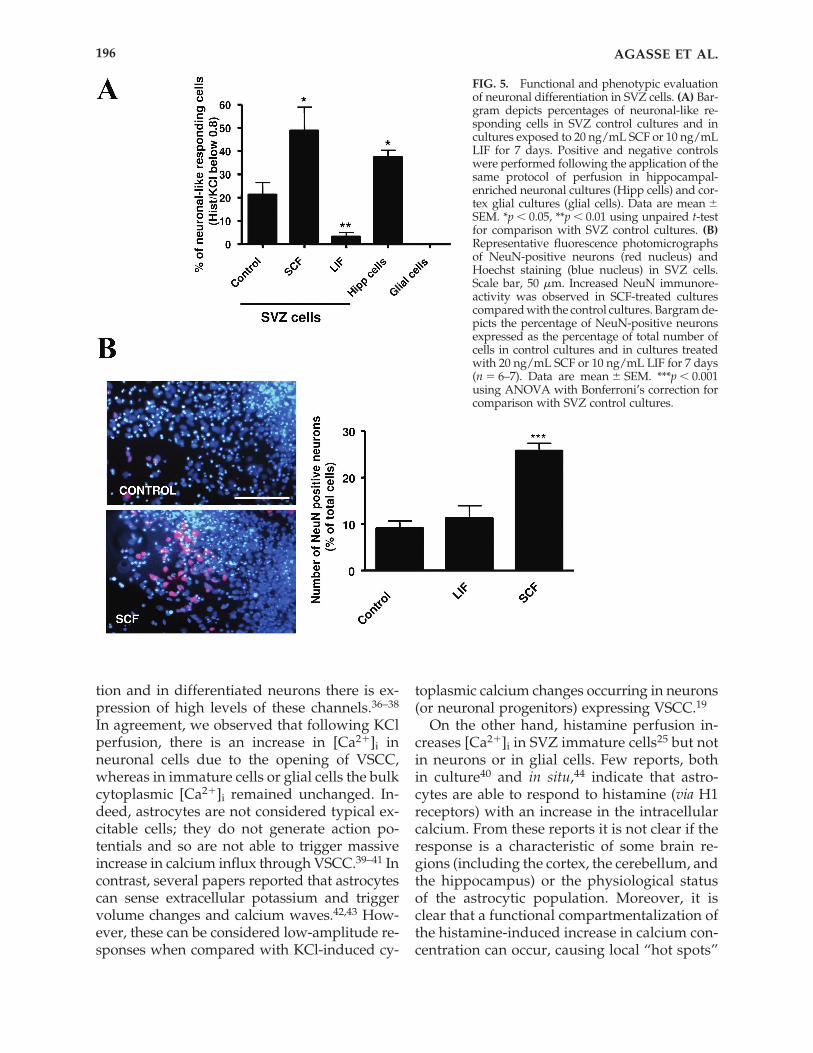
To validate the method described in thepresent work, we performed SCCI experimentson phenotypically identified cells (i.e., on neu-ronal hippocampal primary cultures and oncortical glial cell cultures). In SVZ control cul-tures, �20% of cells display a neuronal-likeprofile i.e., a Hist/KCl ratio below 0.8 (21.3 �5.3%; 974 cells analyzed) (Fig. 5A). In hip-pocampal cultures, �40% (37.4 � 3.1%; 225cells analyzed; p � 0.05 compared with controlSVZ cultures) of the cells display a lowHist/KCl ratio (below 0.8) (Fig. 5A), as ex-pected for a primary culture enriched in a neu-ronal phenotype. In cortical glial cell cultures,we did not observe any cells showing a lowHist/KCl ratio (150 cells analyzed) (Fig. 5A).
We then used this method to evaluate neu-ronal differentiation in SVZ cultures pretreatedwith the pro-neurogenic factor SCF31 or withthe gliogenic factor LIF.32–35 A 25% significantincrease in the proportion of SVZ cells present-ing a neuronal-like profile was observed in SVZcultures treated with SCF (48.8 � 10.2%; 600cells analyzed; p � 0.05 compared with controlSVZ cultures) (Fig. 5A). In contrast, followingtreatment with LIF, the relative number of neu-rons decreased (3.3 � 1.7%; 450 cells analyzed;p � 0.01 compared with control SVZ cultures)(Fig. 5A). Consistently, treatment with SCF in-creased the number of NeuN-positive neuronsto 25% (25.8 � 1.6%; in 5722 cells counted; p �0.001) as compared to 9% in the control (9.1 �1.6%; in 5140 cells counted) (Fig. 5B). Treatmentwith LIF did not modify neuronal differentia-tion (11.3 � 2.6%; in 5891 cells counted).
AGASSE ET AL.194

DISCUSSION
In the present work, we describe a rapidmethod to functionally evaluate neuronal dif-ferentiation in SVZ cell cultures. This methodis based on the profiles of [Ca2�]i variations ac-
cording to cell type, following KCl depolariza-tion and stimulation with histamine.
KCl depolarization was used as a feature ofneuronal differentiation, since VSCC are absentor expressed at very low levels in neural pre-cursors, whereas during neuronal differentia-
FUNCTIONAL IDENTIFICATION OF NEW NEURONS 195
FIG. 4. [Ca2�]i increase in immature SVZ cells following histamine stimulation is mediated by the histamine 1 re-ceptor activation. (A, left) RT-PCR detection of histamine 1 (H1R) and 2 (H2R) receptors in mouse SVZ cells (�, pos-itive controls were done in total murine spleen mRNA). (Right) Fluorescence confocal photomicrograph of nestin(red) and H1R (green) immunodetection. Hoechst 33342 staining (blue) was used to visualize cell nuclei. Scale bar,20 �m. Arrows depict double labeling. (B) SVZ cultures were perfused continuously in Krebs solution over 20 minand stimulated for 2 min from 5� to 7� with 50 mM KCl, from 10� to 12� with 100 �M histamine, and from 15� to 17�with concomitantly 100 �M histamine and either 1 �M mepyramine or 50 �M cimetidine. (C) Representative profilesof response of immature SVZ cells following perfusion according to the previously presented protocol. (D) Effect ofmepyramine and cimetidine on the percentage of immature SVZ cells responding to histamine stimulation. Data aremean � SEM. ***p � 0.001 using ANOVA with Bonferroni’s correction for comparison with the response to histaminealone.
tion and in differentiated neurons there is ex-pression of high levels of these channels.36–38
In agreement, we observed that following KClperfusion, there is an increase in [Ca2�]i in neuronal cells due to the opening of VSCC,whereas in immature cells or glial cells the bulkcytoplasmic [Ca2�]i remained unchanged. In-deed, astrocytes are not considered typical ex-citable cells; they do not generate action po-tentials and so are not able to trigger massiveincrease in calcium influx through VSCC.39–41 Incontrast, several papers reported that astrocytescan sense extracellular potassium and triggervolume changes and calcium waves.42,43 How-ever, these can be considered low-amplitude re-sponses when compared with KCl-induced cy-
toplasmic calcium changes occurring in neurons(or neuronal progenitors) expressing VSCC.19
On the other hand, histamine perfusion in-creases [Ca2�]i in SVZ immature cells25 but notin neurons or in glial cells. Few reports, bothin culture40 and in situ,44 indicate that astro-cytes are able to respond to histamine (via H1receptors) with an increase in the intracellularcalcium. From these reports it is not clear if theresponse is a characteristic of some brain re-gions (including the cortex, the cerebellum, andthe hippocampus) or the physiological statusof the astrocytic population. Moreover, it isclear that a functional compartmentalization ofthe histamine-induced increase in calcium con-centration can occur, causing local “hot spots”
AGASSE ET AL.196
FIG. 5. Functional and phenotypic evaluationof neuronal differentiation in SVZ cells. (A) Bar-gram depicts percentages of neuronal-like re-sponding cells in SVZ control cultures and incultures exposed to 20 ng/mL SCF or 10 ng/mLLIF for 7 days. Positive and negative controlswere performed following the application of thesame protocol of perfusion in hippocampal-enriched neuronal cultures (Hipp cells) and cor-tex glial cultures (glial cells). Data are mean �SEM. *p � 0.05, **p � 0.01 using unpaired t-testfor comparison with SVZ control cultures. (B)Representative fluorescence photomicrographs of NeuN-positive neurons (red nucleus) andHoechst staining (blue nucleus) in SVZ cells.Scale bar, 50 �m. Increased NeuN immunore-activity was observed in SCF-treated culturescompared with the control cultures. Bargram de-picts the percentage of NeuN-positive neuronsexpressed as the percentage of total number ofcells in control cultures and in cultures treatedwith 20 ng/mL SCF or 10 ng/mL LIF for 7 days(n � 6–7). Data are mean � SEM. ***p � 0.001using ANOVA with Bonferroni’s correction forcomparison with SVZ control cultures.

in astrocytic branches and eventual lack of bulkcalcium changes in the cytoplasm.45 In ourwork we evaluated calcium changes in themain cell bodies, excluding the astrocytic pro-cesses, and so did not detect local histamine-induced calcium rises in minor astrocyticbranches. So it is likely that in our calcium as-says the lack of detection of histamine-inducedcalcium rises in astrocyte-like cells might becaused by the fact that our evaluation of cal-cium transients is restricted to the cell bodies.
Few reports indicate that some specializedneurons can respond to histamine. However,the number of cultured sensory neurons re-sponding to histamine (100 �M) is only about15%.28 It is, therefore, likely that in non-sensoryneuronal cultures, such as in the SVZ, hista-mine receptors (in differentiated neurons) arevery minor, absent, or not functionally coupledto increase in the bulk cytoplasmic calcium.
We monitored [Ca2�]i rises using SCCI andcorrelated these data with subsequent immuno-cytochemical characterization of the same im-aged cells. [Ca2�]i increase following KCl per-fusion but not following histamine stimulationwas associated with MAP-2-positive neurons,whereas cells responding to KCl and with mi-nor response to histamine are DCX-positive im-mature neuronal precursors. Cells where [Ca2�]iwas not significantly affected by either KCl orhistamine were positive for GFAP. The analysisof the ratio of responses following histamine andKCl perfusion (Hist/KCl) supported the con-clusion that cells presenting a Hist/KCl ratiovalue below 0.80 are mature neurons, pheno-typically identifiable by MAP-2 expression.Cells responding only to histamine and thus dis-playing a high Hist/KCl ratio (higher than 1)were identified as immature nestin-positivecells. GFAP expression was found in cells re-sponding to neither KCl nor histamine, as wellas in a subpopulation of cells responding to his-tamine and co-expressing nestin, consistent withthe wide expression of nestin among cells of theSVZ, including a subpopulation of astrocytes.30
Thus, the diversity of GFAP-positive cells pres-ent in the SVZ cultures may account for the ob-served functional heterogeneity (including pro-genitors and astrocytes).
Based on the Hist/KCl ratio, we calculatedthe percentage of cells showing neuronal-like
profile in cortical glial cell cultures, neuronal-enriched hippocampal cultures, and SVZ cul-tures. No cells presenting a neuronal-like re-sponse were found in cortical glial cellcultures, whereas 40% were found in postna-tal neuronal hippocampal cultures. These datastrongly indicate that the method now de-scribed reliably allows the identification ofneurons and neuronal differentiation in SVZcultures. In non-treated SVZ (control) cul-tures, about 20% of cells show a neuronal-likeresponse. Pretreatment of SVZ cells with SCF,a trophic factor reported to stimulate neuro-genesis in an in vitro model of cerebral isch-emia, as well as in basal conditions in vivo,31
increased the percentage of cells with a neu-ronal-like response compared to control cul-tures. In contrast, pretreatment of SVZ cul-tures with LIF, a multifunctional cytokinereported to promote self-renewal of neuralstem cells or alternatively the differentiationof neural/progenitor cells into GFAP im-munoreactive cells,32–35,46 decreased the per-centage of cells with a neuronal-like responsecompared to control cultures. Differentiatedneurons expressing NeuN were quantified ineach condition and the obtained data corrob-orated the pro-neurogenic potential of SCF inpostnatal SVZ culture. Thus, the procedurethat we describe is fully adapted for thescreening of putative pro-neurogenic factors.
Using RT-PCR and histamine receptor-selec-tive inhibitors we showed that the increase ofthe [Ca2�]i in SVZ cells following stimulationwith histamine is mediated by the activation ofH1R. Indeed, it was already shown by othersthat embryonic stem and carcinoma cells ex-press functional histamine 1 receptors, sug-gesting a role for this receptor as a marker forundifferentiated neural progenitors.20,21,24,25
Our data demonstrate that the monitoringof the increase in the [Ca2�]i following KCl de-polarization and stimulation with histamine isa powerful and reliable tool to functionallyevaluate neuronal differentiation in SVZ cellcultures. SVZ culture is a mixed culture ofneurons, astrocytes, oligodendrocytes, neu-ronal and glial progenitors in different stagesof differentiation, and stem/immaturecells.1,9,10 We firstly cultivated single SVZ cellswith EGF and FGF-2 over 7 days to allow the
FUNCTIONAL IDENTIFICATION OF NEW NEURONS 197

formation of neurospheres (i.e., to increase cellproliferation). Plating onto poly-D-lysine andwithdrawal of growth factors are necessarysteps to drive the differentiation of progenitorcells into either glial or neuronal fate.47 Free-floating neurospheres adhere to the poly-D-lysine substrate, and cell differentiation oc-curs in the border of the neurospheres wheremigrating cells emerge, forming a dense car-pet of cells. All the measurements of [Ca2�]ivariations and immunocytolabeling were per-formed in these cells. In spite of the fact thatcells in the selected area might be more dif-ferentiated, it is also true that these cells arenot only phenotypically diverse (glia versusneurons) but are also different concerningtheir developmental stages. Indeed, in SVZcultures, neurons are present in different de-velopmental stages, so it is possible to findmarkers for mature neurons, such as NeuNand MAP-2, as well as markers for immaturecells, such as DCX and nestin.
Evaluation of neuronal differentiation by im-munocytochemistry involves time-consumingprocedures performed in dead cells. Single cellcalcium imaging studies provide a rapidmethod to evaluate neuronal function. Func-tionality can be analyzed by electrophysiology,looking at the ability of neurons to generatecurrents following stimulation. However,patch-clamp approaches are time consumingand allow recordings from only one cell eachtime, making impossible estimation of neu-ronal differentiation in the overall culture. Thisis in clear contrast with the possibility of imag-ing between 100 and 120 cells at the same timein 15 min using single cell calcium imaging.Moreover, this technical advance might be use-ful to perform pharmacological studies in liv-ing cells at different stages of development.
Another study describes the single isola-tion of immature cells and neuronal and glialprogenitors from the embryonic rat telen-cephalon.48 In this study, the authors used flu-orescent-activated cell sorting and the combi-nation of calcium responses to basic FGF orEFG and subsequent immunocytochemistry.However, instead of the strong response of SVZimmature cells to histamine detected using ourmethod, Barker and collaborators isolatedneural stem cells by negative flow cytometric
selection for five surface epitopes. Moreover,using calcium imaging in embryonic stem cells,the cellular response to basic FGF via FGFR1 re-ceptor activation was not specific for neuralprecursors since some neuronal and glial pro-genitors also respond with an increase of[Ca2�]i to basic FGF.48 Our method, using spe-cific increase of [Ca2�]i following histamineperfusion, proves to be faster and does not in-volved expensive FACS methods.
In conclusion, monitoring the increase of[Ca2�]i following the application of KCl or his-tamine allowed us to design a rapid method toevaluate neuronal differentiation in living SVZcultures. This method is based on the differentpatterns of increase in the [Ca2�]i, in SVZ cells,following KCl or histamine stimulation, allow-ing the characterization of an immature butfunctional population of neurons. Moreover,this method allows the rapid screening of pu-tative pro-neurogenic factors that might be rel-evant for cell-based therapy, as it allows theidentification of functional immature neurons.These cells are more plastic than fully differ-entiated neurons and so are more adequate forefficient neuronal replacement following graft-ing in the diseased brain.
ACKNOWLEDGMENTS
This work was supported by FCT Portugaland FEDER, POCI/NSE/58492/2004, PTDC/SAU-NEU/68465/2006, SFRH/BD/12731/2003,and FRH/PBD/26462/2006.
REFERENCES
1. Reynolds BA, Weiss S. Generation of neurons and as-trocytes from isolated cells of the adult mammaliancentral nervous system. Science 1992;255:1707–1710.
2. Kornack DR, Rakic P. The generation, migration, anddifferentiation of olfactory neurons in the adult pri-mate brain. Proc Natl Acad Sci USA 2001;98:4752–4757.
3. Pencea V, Bingaman KD, Freedman LJ, Luskin MB.Neurogenesis in the subventricular zone and rostralmigratory stream of the neonatal and adult primateforebrain. Exp Neurol 2001;172:1–16.
4. Lois C, Alvarez-Buylla A. Long-distance neuronal mi-gration in the adult mammalian brain. Science 1994;264:1145–1148.
AGASSE ET AL.198

5. Betarbet R, Zigova T, Bakay RA, Luskin MB.Dopaminergic and GABAergic interneurons of the ol-factory bulb are derived from the neonatal subven-tricular zone. Int J Dev Neurosci 1996;14:921–930.
6. Gheusi G, Cremer H, McLean H, Chazal G, VincentJD, Lledo PM. Importance of newly generated neu-rons in the adult olfactory bulb for odor discrimina-tion. Proc Natl Acad Sci USA 2000;97:1823–1828.
7. Baker H, Liu N, Chun HS, Saino S, Berlin R, Volpe B,Son JH. Phenotypic differentiation during migrationof dopaminergic progenitor cells to the olfactory bulb.J Neurosci 2001;21:8505–8513.
8. Shingo T, Gregg C, Enwere E, Fujikawa H, HassamR, Geary C, Cross JC, Weiss S. Pregnancy-stimulatedneurogenesis in the adult female forebrain mediatedby prolactin. Science 2003;299:117–120.
9. Levison SW, Goldman JE. Multipotential and lineagerestricted precursors coexist in the mammalian peri-natal subventricular zone. J Neurosci Res 1997;48:83–94.
10. Suslov ON, Kukekov VG, Ignatova TN, Steindler DA.Neural stem cell heterogeneity demonstrated by mo-lecular phenotyping of clonal neurospheres. Proc NatlAcad Sci USA 2002;99:14506–14511.
11. Lendahl U, Zimmerman LB, McKay RD. CNS stemcells express a new class of intermediate filament pro-tein. Cell 1990;60:585–595.
12. Lee MK, Rebhun LI, Frankfurter A. Post-translationalmodification of class III beta-tubulin. Proc Natl AcadSci USA 1990;87:7195–7199.
13. Yang HK, Sundholm-Peters NL, Goings GE, WalkerAS, Hyland K, Szele FG. Distribution of doublecortinexpressing cells near the lateral ventricles in the adultmouse brain. J Neurosci Res 2004;76:282–295.
14. Ocbina PJ, Dizon ML, Shin L, Szele FG. Doublecortinis necessary for the migration of adult subventricularzone cells from neurospheres. Mol Cell Neurosci2006;33:126–135.
15. Doetsch F, Alvarez-Buylla A. Network of tangentialpathways for neuronal migration in adult mammalianbrain. Proc Natl Acad Sci USA 1996;93:14895–14900.
16. Bonfanti L. PSA-NCAM in mammalian structuralplasticity and neurogenesis. Prog Neurobiol 2006;80:129–164.
17. Lois C, Alvarez-Buylla A. Proliferating subventricu-lar zone cells in the adult mammalian forebrain candifferentiate into neurons and glia. Proc Natl Acad SciUSA 1993;90:2074–2077.
18. Jin K, Minami M, Lan JQ, Mao XO, Batteur S, SimonRP, Greenberg DA. Neurogenesis in dentate sub-granular zone and rostral subventricular zone afterfocal cerebral ischemia in the rat. Proc Natl Acad SciUSA 2001;98:4710–4715.
19. Ambrósio AF, Silva AP, Malva JO, Mesquita JF, Car-valho AP, Carvalho CM. Role of desensitization ofAMPA receptors on the neuronal viability and on the[Ca2�]i changes in cultured rat hippocampal neurons.Eur J Neurosci 2000;2:2021–2031.
20. Bloemers SM, Leurs R, Smit MJ, Verheule S, TertoolenLG, Timmerman H, de Laat SW. Mouse P19 embry-
onal carcinoma cells express functional histamine H1-receptors. Biochem Biophys Res Commun 1993;191:118–125.
21. Bloemers SM, Verheule S, Peppelenbosch MP, SmitMJ, Tertoolen LG, de Laat S. Sensitization of the his-tamine H1 receptor by increased ligand affinity. J BiolChem 1998;273:2249–2255.
22. Panula P, Lintunen M, Karlstedt K. Histamine in braindevelopment and tumors. Semin Cancer Biol 2000;10:11–14.
23. Hernandez-Angeles A, Soria-Jasso LE, Ortega A,Arias-Montano JA. Histamine H1 receptor activationstimulates mitogenesis in human astrocytoma U373MG cells. J Neurooncol 2001;55:81–89.
24. Wiener Z, Tóth S, Gócza E, Kobolók J, Falus A. Mouseembryonic stem cells express histidine decarboxylaseand histamine H1 receptors. Inflamm Res 2003;52(Suppl 1): S53–54.
25. Tran PB, Ren D, Veldhouse TJ, Miller RJ. Chemokinereceptors are expressed widely by embryonic andadult neural progenitor cells. J Neurosci Res 2004;76:20–34.
26. Yanagida E, Shoji S, Hirayama Y, Yoshikawa F, OtsuK, Uematsu H, Hiraoka M, Furuichi T, Kawano S.Functional expression of Ca2� signaling pathways inmouse embryonic stem cells. Cell Calcium 2004;36:135–146.
27. Ahlemeyer B, Baumgart-Vogt E. Optimized protocolsfor the simultaneous preparation of primary neuronalcultures of the neocortex, hippocampus and cerebel-lum from individual newborn (P 0.5) C57Bl/6J mice.J Neurosci Methods 2005;149:110–120.
28. Nicolson TA, Bevan S, Richards CD. Characterisationof the calcium responses to histamine in capsaicin-sensitive and capsaicin-insensitive sensory neurones.Neuroscience 2002;110:329–338.
29. Terao A, Steininger TL, Morairty SR, Kilduff TS. Age-related changes in histamine receptor mRNA levelsin the mouse brain. Neurosci Lett 2004; 355:81–84.
30. Doetsch F, Garcia-Verdugo JM, Alvarez-Buylla A.Cellular composition and three-dimensional organi-zation of the subventricular germinal zone in theadult mammalian brain. J Neurosci 1997;17:5046–5061.
31. Jin K, Mao XO, Sun Y, Xie L, Greenberg DA. Stem cellfactor stimulates neurogenesis in vitro and in vivo. JClin Invest 2002;110:311–319.
32. Galli R, Pagano SF, Gritti A, Vescovi AL. Regulationof neuronal differentiation in human CNS stem cellprogeny by leukemia inhibitory factor. Dev Neurosci2000;22:86–95.
33. Viti J, Feathers A, Phillips J, Lillien L. Epidermalgrowth factor receptors control competence to inter-pret leukaemia inhibitory factor as an astrocyte in-ducer in developing cortex. J Neurosci 2003;23:3385–3393.
34. Bonaguidi MA, McGuire T, Hu M, Kan L, Samanta J,Kessler JA. LIF and BMP signaling generate separateand discrete types of GFAP-expressing cells. Devel-opment 2005;132:5503–5514.
FUNCTIONAL IDENTIFICATION OF NEW NEURONS 199

35. Nakanishi M, Niidome T, Matsuda S, Akaike A, Ki-hara T, Sugimoto H. Microglia-derived interleukin-6and leukaemia inhibitory factor promote astrocyticdifferentiation of neural stem/progenitor cells. Eur JNeurosci 2007;25:649–658.
36. Sah DW, Ray J, Gage FH. Regulation of voltage- andligand-gated currents in rat hippocampal progenitorcells in vitro. J Neurobiol 1997;32:95–110.
37. Maric D, Maric I, Barker JL. Developmental changesin cell calcium homeostasis during neurogenesis ofthe embryonic rat cerebral cortex. Cereb Cortex 2000;10:561–573.
38. Fiorio Pla A, Maric D, Brazer SC, Giacobini P, Liu X,Chang YH, Ambudkar IS, Barker JL. Canonical tran-sient receptor potential 1 plays a role in basic fibro-blast growth factor (bFGF)/FGF receptor-1-inducedCa2� entry and embryonic rat neural stem cell pro-liferation. J Neurosci 2005;25:2687–2701.
39. Parpura V, Liu F, Jeftinija KV, Haydon PG, JeftinijaSD. Neuroligand-evoked calcium-dependent releaseof excitatory amino acids from Schwann cells. J Neu-rosci 1995; 15:5831–5839.
40. Jung S, Pfeiffer F, Deitmer JW. Histamine-induced cal-cium entry in rat cerebellar astrocytes: evidence forcapacitative and non-capacitative mechanisms. JPhysiol 2000;3:549–561.
41. Hirase H, Qian L, Barthó P, Buzsáki G. Calcium dy-namics of cortical astrocytic networks in vivo. PLoSBiol 2004;2:E96.
42. O’Connor ER, Kimelberg HK. Role of calcium in as-trocyte volume regulation and in the release of ionsand amino acids. J Neurosci 1993;13:2638–2650.
43. Guerrero-Cázares H, Alatorre-Carranza Mdel P, Del-gado-Rizo V, Duenas-Jimenez JM, Mendoza-MaganaML, Morales-Villagran A, Ramirez-Herrera MA,Guerrero-Hernández A, Segovia J, Duenas-JimenezSH. Dopamine release modifies intracellular calciumlevels in tyrosine hydroxylase-transfected C6 cells.Brain Res Bull 2007;74:113–118.
44. Shelton MK, McCarthy KD. Hippocampal astrocytesexhibit Ca2�-elevating muscarinic cholinergic andhistaminergic receptors in situ. J Neurochem 2000;74:555–563.
45. Inagaki N, Fukui H, Ito S, Yamatodani A, Wada H.Single type-2 astrocytes show multiple independentsites of Ca2� signaling in response to histamine. ProcNatl Acad Sci USA 1991;88:4215–4219.
46. Bauer S, Patterson PH. Leukemia inhibitory factorpromotes neural stem cell self-renewal in the adultbrain. J Neurosci 2006;26:12089–12099.
47. Gage FH. Mammalian neural stem cells. Science2000;287:1433–1438.
48. Maric D, Maric I, Chang YH, Barker JL. Prospectivecell sorting of embryonic rat neural stem cells andneuronal and glial progenitors reveals selective ef-fects of basic fibroblast growth factor and epidermalgrowth factor on self-renewal and differentiation. JNeurosci 2003;23:240–251.
Address reprint requests to:João O. Malva
Neuroprotection and Neurogenesis in Brain Repair Group
Center for Neuroscience and Cell BiologyInstitute of Biochemistry
Faculty of MedicineUniversity of Coimbra
3004-504 CoimbraPortugal
E-mail: [email protected]
Received: July 17, 2007Accepted: September 13, 2007
AGASSE ET AL.200




















