
Renal lymphatic ultrastructure and translymphatic transport
14
MICROVASCULAR RESEARCH 19, 338-351 (1980) Renal Lymphatic Ultrastructure and Translymphatic Transport1 KURT H. ALBERTINE AND CHARLES C. C. O’MORCHOE Department of Anatomy, Loyola University Stritch School of Medicine, Maywood, Z//inois 60153 Received May 30, 1979 Information on the mechanisms of transport across renal lymphatic endothelium in the dog was derived by qualitative and quantitative ultrastructural analyses. Interlobular and intralobular renal lymphatics were studied in dogs with ureteric obstruction and lymphatic ligation, in dogs with lymphatic ligation only, and in dogs with unimpeded lymph and urine flow. Minor differences were detected in the ultrastructural features of lymphatic en- dothelium among the three groups of animals and between the two types of lymphatics studied within each group. Approximately 5% of the endothelial intracytoplasmic volume was occupied by vesicles, the majority of which were of the small uncoated variety. On an average 65% of these vesicles lay within the cytoplasm, and of the remainder, most were associated with the luminal border. Interendothelial contacts were of three varieties--end- to-end, overlapping, and complexly interdigitating. One-third of these lacked any specialized junctional complexes, whereas about 60% of the identifiable complexes revealed a fascia occludens. The remainder (approximately 10%) possessed fascia adhaerentes. Only 2 exam- ples, out of 240 contacts studied, could be described as open junctions (greater than 50 nm). The conclusions drawn from this study were that (1) interlobular and intralobular renal lymphatics subserve similar functions in lymph formation, (2) at least in the absence of local irritation or trauma, open junctions do not play a significant role in translymphatic transport of fluid and protein, and (3) for poorly diffusible substances the primary pathways into the lymphatic lumen are intravesicular transport and the normal intercellular channels between contiguous cells. INTRODUCTION The mechanisms by which transport from blood to lymph occur have been subject to structural and functional investigations. Structural studies have focused on the identification of morphologically visible transport pathways (Casley-Smith, 1964, 1965, 1969; Casley-Smith et al., 1975; Casley-Smith and Sims, 1976; Dob- bins and Rollins, 1970; Leak and Burke, 1966, 1968). Other studies have attempted to characterize the process from a functional standpoint (Renkin, 1964; Garlick and Renkin, 1970; Renkin et al., 1977a, b; Lassen et al., 1974; Taylor et al., 1977; Brace et al., 1977). Although both types of study seek to elucidate the relative importance of the intracytoplasmic, the vesicular and the intercellular routes, there is as yet little agreement on the issue. Morphological studies are hampered by the problem of extrapolation from static observations to dynamic changes. Interpretation of physiological experiments, on the other hand, is hindered by the 1 This work was supported by Grant AM 18473from the National Institutes of Health. 338 CKI26-2862/80/03033&14$.00/0 Copyright @ 1980 by Academic Press, Inc. All tights of reproduction in any form reserved. kinted in U.S.A.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Renal lymphatic ultrastructure and translymphatic transport

MICROVASCULAR RESEARCH 19, 338-351 (1980)
Renal Lymphatic Ultrastructure and Translymphatic Transport1
KURT H. ALBERTINE AND CHARLES C. C. O’MORCHOE
Department of Anatomy, Loyola University Stritch School of Medicine, Maywood, Z//inois 60153
Received May 30, 1979
Information on the mechanisms of transport across renal lymphatic endothelium in the dog was derived by qualitative and quantitative ultrastructural analyses. Interlobular and intralobular renal lymphatics were studied in dogs with ureteric obstruction and lymphatic ligation, in dogs with lymphatic ligation only, and in dogs with unimpeded lymph and urine flow. Minor differences were detected in the ultrastructural features of lymphatic en- dothelium among the three groups of animals and between the two types of lymphatics studied within each group. Approximately 5% of the endothelial intracytoplasmic volume was occupied by vesicles, the majority of which were of the small uncoated variety. On an average 65% of these vesicles lay within the cytoplasm, and of the remainder, most were associated with the luminal border. Interendothelial contacts were of three varieties--end- to-end, overlapping, and complexly interdigitating. One-third of these lacked any specialized junctional complexes, whereas about 60% of the identifiable complexes revealed a fascia occludens. The remainder (approximately 10%) possessed fascia adhaerentes. Only 2 exam- ples, out of 240 contacts studied, could be described as open junctions (greater than 50 nm). The conclusions drawn from this study were that (1) interlobular and intralobular renal lymphatics subserve similar functions in lymph formation, (2) at least in the absence of local irritation or trauma, open junctions do not play a significant role in translymphatic transport of fluid and protein, and (3) for poorly diffusible substances the primary pathways into the lymphatic lumen are intravesicular transport and the normal intercellular channels between contiguous cells.
INTRODUCTION
The mechanisms by which transport from blood to lymph occur have been subject to structural and functional investigations. Structural studies have focused on the identification of morphologically visible transport pathways (Casley-Smith, 1964, 1965, 1969; Casley-Smith et al., 1975; Casley-Smith and Sims, 1976; Dob- bins and Rollins, 1970; Leak and Burke, 1966, 1968). Other studies have attempted to characterize the process from a functional standpoint (Renkin, 1964; Garlick and Renkin, 1970; Renkin et al., 1977a, b; Lassen et al., 1974; Taylor et al., 1977; Brace et al., 1977). Although both types of study seek to elucidate the relative importance of the intracytoplasmic, the vesicular and the intercellular routes, there is as yet little agreement on the issue. Morphological studies are hampered by the problem of extrapolation from static observations to dynamic changes. Interpretation of physiological experiments, on the other hand, is hindered by the
1 This work was supported by Grant AM 18473 from the National Institutes of Health.
338
CKI26-2862/80/03033&14$.00/0 Copyright @ 1980 by Academic Press, Inc. All tights of reproduction in any form reserved. kinted in U.S.A.

LYMPHATIC ULTRASTRUCTURE AND TRANSPORT 339
interplay of dynamic factors as they occur in the blood capillary wall, the in- terstitium, and the lymphatic endothelium. While most attention has been paid to the blood capillary, two principal theories on translymphatic transport have emerged. One is that protein and fluid movement occurs primarily by convective forces (Casley-Smith, 1976, 1977; Lassen et al., 1974). The second theory is that the bulk of lymph formation occurs by dissipative forces-either diffusion or vesicular (Dobbins and Rollins, 1970). Functional experiments by Renkin et al. (1977a, b), Garlick and Renkin (1970), and Per1 (1975) on the dog’s paw have tended to support this theory. For example, Renkin et al. (1977b) provided evidence that only 30% of the total flux of albumin from plasma into lymph occurred by convection (solvent drag), the remainder being accounted for by dissipative forces.
The present study was concerned with ultrastructural evidence of transport across the wall of renal lymphatics in the dog. Renal lymphatics were selected because they are accessible to in vivo fixation with comparatively little direct trauma to the capillary bed, and because renal lymph continues to flow in the anesthetized animal. With this preparation the morphologically visible routes of translymphatic transport were analyzed indirectly by a qualitative and quantita- tive approach.
METHODS AND MATERIALS
The study was performed on the left kidneys of nine mongrel dogs of either sex subdivided into three groups. One group of three dogs was subjected to left ureteric obstruction 3 days prior to removal of the kidney, and to efferent renal lymphatic ligation 4 to 6 hr before renal fixation. The second group was subjected to the renal lymphatic ligation procedure only. A third group of three dogs was not subjected to experimental maneuver so that both urine and lymph flowed freely until the time of fixation. Sodium pentobarbital was used during all surgical procedures.
All nine kidneys were fixed by the in vivo drip technique using 5% acrolein (Kodak or MCB) in Millonig’s phosphate buffer (Albertine and O’Morchoe, 1979). By this technique 250 ml of fixative was dripped onto the ventral surface of the kidneys of anesthetized animals and simultaneously 100 ml of fixative was perfused retrogradely through a double cannula in the ureter. The second limb of the cannula served as an outlet for the fixative, thereby preventing a pressure buildup in the renal pelvis. At the end of this step, 30 tissue strips (1 x 1 x 4 mm) were cut from the cortex of each kidney. Ten of these strips were taken from each pole and 10 from the central region. The tissue strips were further fixed for 1 hr at 4” in 2% glutaraldehyde in Millonig’s phosphate buffer and postfixed in 1% osmium tetroxide at 4” for 1 hr. They were subsequently flat-embedded in Epon 812 and sectioned parallel to the surface, starting at the capsule. Sections 1 pm thick were stained with 1% toluidine blue and examined for potential lymphatics. Tissue containing lymphatics was trimmed and sectioned with an LKB utrami- crotome. These thin sections were stained with uranyl acetate and Reynold’s lead citrate and examined with an Hitachi HU- 1 lB-2 electron microscope.
Quantitative data were obtained from two types of cortical lymphatics,-those

340 ALBERTINE AND O’MORCHOE
that were associated with interlobular blood vessels (interlobular lymphatics) and those deemed to be in the substance of the renal lobules (intralobular lymphatics). The volume density (V,) of intraendothelial cytoplasmic vesicles was obtained by the point-counting method of Weibel(1973). By this method a transparent grid was superimposed over electron micrographs of standard magnification and the vol- ume density derived from the formula V, = Pi/P,, where Pi is the number of points which overlay vesicles and PT equals the total number of points over cyto- plasm. In addition, the size of small uncoated cytoplasmic vesicles was estimated by measuring the external and internal diameter in two planes perpendicular to one another. The position of the vesicles in the cell was also recorded according to whether they lay within the cytoplasm or were in contact with or opened onto the luminal and abluminal surface of the cells. The numerical density (NV) of intracy- toplasmic vesicles was obtained by the formula N, = nJA (D + T) (Elias and Hennig, 1967), where nA is the number of vesicles counted in an area (A) of 2 pm2 of section, b is the mean vesicular diameter (84 nm), and T is the section thickness (60 nm).
Lymphatic endothelial cell thickness was measured several times along the length of the cells to obtain the best estimate of the maximal and minimal non- nuclear endothelial cell thicknesses as well as the perinuclear cell thickness. The number of endothelial cells seen in typical cross sections of lymphatics was also recorded. Lymphatic endothelial cell contacts between contiguous cells were enumerated and classified according to the specialized intercellular junctions present.
During the collection of all the above data care was taken to ensure that values were obtained from both polar and central regions of the renal cortex. Statistical analysis was performed where appropriate using Student’s t test.
RESULTS
Lymphatics were found in interlobular and intralobular locations as previously described (Albertine and O”Morchoe, 1979). Both types of vessels revealed an ultrastructure which is now well recognized as being characteristic of lymphatics generally (Casley-Smith and Florey, 1961; Fraley and Weiss, 1961; Rhodin, 1%5; Leak and Burke, 1966, 1968). Most important for their identification was the lack of fenestrations and the absence of a basal lamina (Fig. l), features that were in marked contrast to those of peritubular blood capillaries. The typical appearance of the cytoplasm, including organelles, of the lymphatic endothelial cells is shown in Fig. 2. No obvious lymphatic ultrastructural differences were found among the different regions of the kidney nor among the different kidney groups.
Quantitative data on the number and size of cells forming lymphatic endothelial walls also failed to reveal a difference among the three groups’of animals (Tables 1 and 2). Table 1 shows that interlobular lymphatics contained parts of approxi- mately eight endothelial cells in any one cross section, in contrast to intralobular lymphatics which showed only four or five. This finding conformed with an earlier observation that cross sections of interlobular lymphatics have about twice the surface area of intralobular vessels (Albertine and O’Morchoe, 1979). Table 2 provides evidence on endothelial cell thickness which in nonnuclear areas varied

LYMPHATIC ULTRASTRUCTURE AND TRANSPORT 341
FIG. 1. Electron micrograph of a renal cortical lymphatic capillary (L) from a kidney with unim- peded lymph flow. The lymphatic has a non-fenestrated endothelial wall that lacks a subjacent basal lamina. x 5730.
-, . A.“.., d junctional complex is present a. VLlr and not at the other (arrows). x 43,630.

342 ALBERTINE AND O’MORCHOE
TABLE 1 MEAN NUMBER OF CELLS IN A TYPICAL CROSS SECTION
Group Region No. LY” Number
Ureter-obstructed (k SE)
Lymphatic ligated (k SE)
Free lymph flow (rt SE)
a Lymphatic. * p < 0.025. c p < 0.025.
Interlobular 12 8 (1) Intralobular 5 5 (1)
Interlobular 12 8 (I)* Intralobular 5 4 (I)*
Interlobular 12 8 (1Y Intralobular 5 4 (1Y
from about 0.058 to 1.0 pm. Thickness varied from one lymphatic to another, as well as in different parts of the same vessel.
Two subpopulations of vesicles, differentiated by size, were found. The larger heterogeneous population (200-400 nm in diameter) were infrequently encoun- tered. On occasion, especially in ureter-obstructed kidneys, flocculent material which was similar to that seen in the interstitium was found in these vesicles (Figs. 3 and 4). The smaller, more homogeneous, type of vesicles constituted a much more prominent feature of lymphatic cytoplasm. Most of the vesicles had a comparatively simple type of membranous wall (uncoated). However, occasional vesicles of both sizes appeared to have a thicker coat with a more fuzzy lining (coated vesicles). Quantitative data on the size, position, number, and volume density of the smaller uncoated vesicles is given in Tables 3,4, and 5. These show that the average vesicle was about 84 nm in external diameter, with no significant difference among the various groups (Table 3), and that about three quarters of all the vesicles were within the cytoplasm (Table 4). Of the remaining vesicles, there were more associated with the luminal border than with the abluminal border of the cells. Volume density measurements (Table 5) showed that between 3 and 5% of the endothelial cytoplasm was occupied by the small vesicles. Table 5 also shows that between 6 and 11 vesicles occupied each cubic micrometer of nonnuclear cytoplasm.
The regions where adjacent endothelial cells came into contact belonged to
TABLE 2 MEAN LYMPHATIC END~THELIAL CELL THICKNESS (pm)
Group Region Maximum nonnuclear
Minimum nonnuclear
At nucleus
Ureter-obstructed (2 SE)
Lymphatic ligated (k SE)
Free lymph flow (-c SE)
Interlobular (N= 16) Intralobular (N=4)
Interlobular (N= 16) Intralobular (N=4)
Interlobular (N= 16) Intmlobular (N=4)
0.91 (0.12) 0.10 (0.013) 3.2 (0.22) 0.66 (0.35) 0.058 (0.023) 2.7 (0.21)
0.85 (0.082) 0.10 (0.0894) 3.1 (0.16) 0.85 (0.17) 0.088 (0.017) 2.8 (0.17)
0.82 (0.076) 0.085 (0.0075) 3.2 (0.20) 0.81 (0.15) 0.069 (0.016) 2.8 (0.23)

LYMPHATIC ULTRASTRUCTURE AND TRANSPORT 343
FIG. 3. Electron micrograph of a portion of a lymphatic (L) from a kidney with ureteric obstruc- tion and renal lymphatic ligation. Interstitial flocculent material has filled an abluminal cytoplasmic invagination (CI) and membrane-bound vesicles (large arrows). The small arrows indicate small coated vesicles. x 22,230.
FIG. 4. Electron micrograph of apart of a lymphatic capillary (L) from a kidney with unimpeded lymph flow. Flocculent material (arrow) is located at the luminal opening of two coalesced vesicles. x
64,620.

344 ALBERTINE AND O’MORCHOE
TABLE 3 MEAN DIAMETER OF VESICLES (pm)
Group Region
Free” Attachedb
ED ID ED ID
Ureter-obstructed Interlobular 0.085 0.074 0.084 0.070 Intralobular 0.083 0.072 0.082 0.073
Lymphatic ligated Interlobular 0.082 0.071 0.084 0.073 Intral6bular 0.083 0.073 0.085 0.074
Free lymph flow Interlobular 0.089 0.077 0.085 0.074 Intralobular 0.085 0.073 0.085 0.073
a “Free” vesicles were in the interior of the cytoplasm. Ten vesicles measured for each mean determination. ED, external diameter; ID, internal diameter.
b “Attached” vesicles were in contact with the endothelial cell membrane.
TABLE 4 FREQUENCY OF VESICLE POSITION (%)
Group Region No. % % % % % V” L-T L-O Freeb Ab-T Ab-0
Ureter-obstructed Interlobulti 329 9 7 77 3 1 Intralobulald 29 7 3 83 3 3
Lymphatic ligated Interlobular 208 7 17 63 6 7 Intralobular 59 10 8 68 7 7
Free lymph flow Interlobular 116 11 19 55 6 9 Intralobular 29 10 14 62 7 7
B V, Vesicle; L, luminal; Ab, abluminal; T, touch; 0, open. b “Free” vesicles were in the interior of the cytoplasm. c 40 pm of cell length examined per region. d 8 pm of cell length examined per region.
TABLE 5 MEAN V, AND N, FOR VESICLES”
Group
Ureter-obstructed (k SE)
Lymphatic ligated (2 SE)
Free lymph flow (2 SE)
Region
Interlobular Intralobular
Interlobular Intralobular
Interlobular Intralobular
No. V” NV LY” bmSIpm3) (No./Fm3)
28 0.049 (0.0067) 11 (2) 5 0.030 (0.0045) 6 (1)
29 0.044 (0.0064) 8 (1) 7 0.038 (0.0070) 9 (3)
28 0.040 (0.0033) 8 (1) 5 0.035 (0.0054) 6 (2)
a These values are for nonnuclear cytoplasm. b Lymphatic.

LYMPHATIC ULTRASTRUCTURE AND TRANSPORT 345
three morphological patterns. The simplest of these was where an end-to-end type of contact was established (Fig. 5). A second format was where contiguous ceils overlapped one another for varying distances (Fig. 6). The third pattern was more complex in that the adjacent cells interdigitated with one another (Fig. 7). Specialized junctions (Fig. 8) were often found in association with all three types of contact, and these are quantified in Tables 6 and 7. Although the junctions could be classified according to the standard nomenclature, the term fascia seemed more appropriate than zonula (Bloom and Fawcett, 1975) since in most instances these junctional complexes did not appear to encircle the cells but rather to exist along a segment of their borders (Fig. 2). At a number of cell contacts it was not possible to identify or classify a junctional complex because of the plane of section in relation to that of the membranes. Table 7 shows that of the identifiable lymphatic endothelial cell contacts, between 21 and 38% lacked any form of specialized junctional complex. The remainder of the contacts were seen to have fascia occludentes and/or fascia adhaerentes.
DISCUSSION
Although the three groups of animals used in the present study provided examples of lymphatics under different physiological conditions, the primary purpose of using these groups was to assist in the identification of intrarenal lymphatics. Both ureteric obstruction and lymphatic ligation were employed to dilate the lymphatic vessels and thus simplify their recognition in tissue sections.
FIG. 5. Electron micrograph of lymphatic (L) endothelial cell contacts from a kidney with unim- peded lymph flow. An end-to-end abutment of adjacent cellular processes (arrows) is illustrated. Small uncoated vesicles (V) are located within the cytoplasm. M, mitochondria. x 57,590.

346 ALBERTINE AND O’MORCHOE
FIG. 6. Electron micrograph of an overlapping (arrows) type of cell contact from a kidney with unimpeded lymph flow. Note the absence of a specialized junctional complex. x 47,120.
FIG. 7. Electron micrograph of a complex type of lymphatic (L) interendothelial cell contact (large arrows) from,a kidney that had undergone renal lymphatic ligatiod’alone. Note the prominent continu- ous basal lamina (small arrow))of the neighboring blood capillary (BC) whereas a discontinuous segment of a basal lamina is evident subjacent to thd lytiphatic endothelium (small arrow). M, mitochondria. x 47,642.
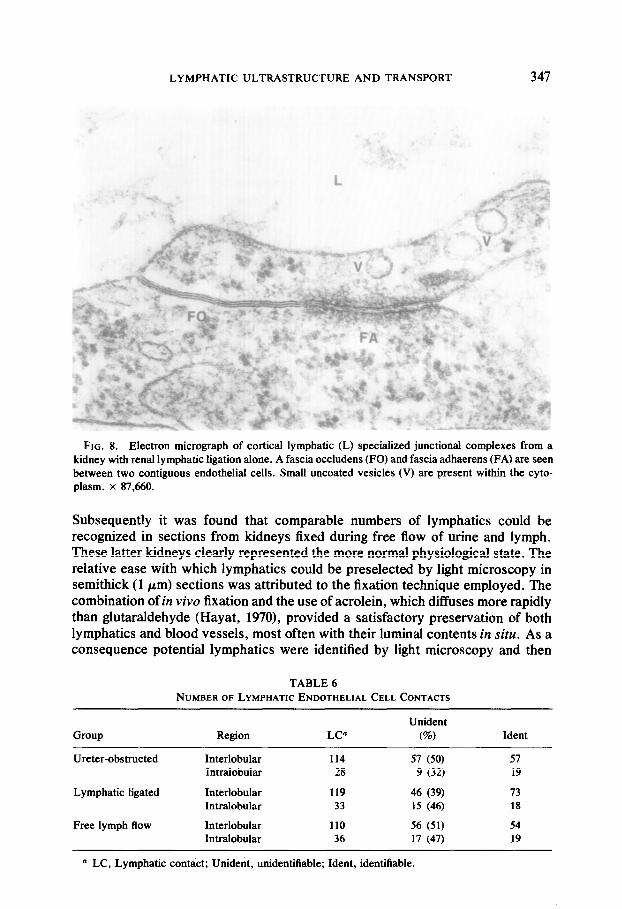
LYMPHATIC ULTRASTRUCTURE AND TRANSPORT 347
FIG. 8. Electron micrograph of cortical lymphatic (L) specialized junctional complexes from a kidney with renal lymphatic ligation alone. A fascia occludens (FO) and fascia adhaerens (FA) are seen between two contiguous endothelial cells. Small uncoated vesicles (V) are present within the cyto- plasm. x 87,660.
Subsequently it was found that comparable numbers of lymphatics could be recognized in sections from kidneys fixed during free flow of urine and lymph. These latter kidneys clearly represented the more normal physiological state. The relative ease with which lymphatics could be preselected by light microscopy in semithick (1 pm) sections was attributed to the fixation technique employed. The combination of in vivo fixation and the use of acrolein, which diffuses more rapidly than glutaraldehyde (Hayat, 1970), provided a satisfactory preservation of both lymphatics and blood vessels, most often with their luminal contents in situ. As a consequence potential lymphatics were identified by light microscopy and then
TABLE 6 NUMBEROF LYMPHATIC ENDOTHELIAL CELL CONTACTS
Group Region LC” Unident
m Ident
Ureter-obstructed Interlobular 114 57 (50) 57 Intralobular 28 9 (32) 19
Lymphatic ligated Interlobular 119 46 (39) 73 Intralobular 33 15 (46) 18
Free lymph flow Interlobular 110 56 (51) 54 Intralobular 36 17 (47) 19
D LC, Lymphatic contact; Unident, unidentifiable; Ident, identifiable.

348 ALBERTINE AND O’MORCHOE
TABLE I TYPEOF LYMPHATICENDOTHELIAL CELL CONTACTS
Group Region Ident FO” FA No JC Open
Ureter-obstructed Interlobular 57 41 (72)” 4 (7) 12 (21) 0 Intralobular 19 14 (74) 0 5 (26) 0
Lymphatic ligated Interlobular 73 35 (48) 8 (11) 28 (38) 2 (3) Intralobular 18 11 (62) 1 (5) 6 (33) 0
Free lymph flow Interlobular 54 32 (59) 6 (11) 16 (30) 0 Intralobular 19 11 (58) 2 (10) 6 (32) 0
n FO, Fascia occludens; FA, fascia adhaerens; JC, junctional complex. * The percentage is in parentheses.
confirmed in thin sections by their ultrastructural characteristics (Casley-Smith and Florey, 1961; Fraley and Weiss, 1961; Rhodin, 1965; Rojo-Ortega& al., 1973; Frank et al., 1977).
Comparatively little difference was detected in the ultrastructural features of transendothelial lymphatic transport among the three groups of animals. One striking difference, however, was in the numerical density of uncoated small vesicles in the ureter-obstructed group of kidneys. This may be explained in part by the relative dilatation of the intralobular lymphatics which followed occlusion of the ureter-a maneuver that brings about edema of the kidney. Edema in turn induces lymphatic dilatation through the action of anchoring filaments (Leak, 1976). As lymphatic endothelium becomes thin and attenuated it would appear that the vesicular component of the cytoplasm decreases. Why this occurred in the intralobular rather than the interlobular lymphatics is not evident.
Of special interest in this study is the otherwise close similarity in structure between interlobular and intralobular lymphatics. It was anticipated that the two might differ functionally, and therefore morphologically, on the supposition that the initial lymphatics lying within the lobule would reflect the transport process, whereas the interlobular vessels might portray more the characteristics of collect- ing vessels. The fact that no morphological difference was apparent between the two groups of lymphatics leads us to conclude that both types of lymphatic vessels may be engaged in lymph formation.
The question on the mechanism(s) of lymph formation remains. Casley-Smith et al. (1975), Casley-Smith and Sims (1976), and Casley-Smith (1977) have main- tained that gaps between contiguous endothelial cells (in excess of 0.1 pm) provide sufficient pathways to account for total lymph formation, at least in certain regions of the body. Morphological evidence for the existence of such pathways has been provided by Casley-Smith (1%4,1965), Leak and Burke (1966, 1%8), and others. However, care in interpretation of this evidence must be exercised since trauma of any sort, including depilation of the skin overlying subcutaneous sites of study or injections of tracer particles, may stimulate his- tamine release with subsequent edema. It is known that the incidence of intercellu- lar gaps increases as a consequence of trauma and this is true of the kidney (Ohkuma, 1973; Nordquist et al., 1973) as elsewhere. The kidney, however, is amenable to in vivo drip fixation with little or no direct trauma to the capillary bed.

LYMPHATIC ULTRASTRUCTURE AND TRANSPORT 349
Under such circumstances the incidence of intercellular gaps was found to be so low in the present study (Table 7) that it is difflcult to attribute much importance to them. It can be argued that the lymphostasis induced in two of the groups of dogs could have served to minimize the presence of gaps (open junctions) because lymph formation may have been reduced. Alternatively the edema associated with ureteric and lymphatic occlusion could bring about the formation of such gaps. In any event the lack of open junctions or gaps in free flow kidneys argues against their importance in normal lymph formation-that is, in the absence of edema or increased lymph flow.
The typical intercellular pathways, approximately 15 to 20 nm wide, provide a viable transport route. As demonstrated in Figs. 6,7, and 8 they vary in complex- ity, and presumably the end-to-end type of contact provides a simpler mode of entry than do the overlapping or complexly interdigitating types. Certainly, not all of these various regions of proximity reveal the specialized junctional complexes which might impede the transport of fluid (Table 7).
Dissipative forces may be reflected in diffusion or vesicular transport across endothelial cells. Diffusion does not readily lend itself to morphological study. Vesicular transport has been the subject of numerous morphological (Dobbins and Rollins, 1970; Casley-Smith, 1969; Bruns and Palade, 1968) and functional studies. In the present study the volume density of small uncoated vesicles, which formed the major vesicular component of lymphatic endothelial cytoplasm, amounted to about 5%. This contrasts with a figure of 15% obtained by Dobbins and Rollins (1970) in mouse lacteals and 36% by Casley-Smith (1969) in murine diaphragmatic lymphatics. It is not at the moment clear whether these disparities are attributable to regional, methodological, or to species differences. The disposition of the vesicles seen in the present study (Table 4) suggests that they are involved in the transport process. More definitive studies on this topic using tracers are in prog- ress. Examples of vesicles forming chains which might provide a continuous channel across the lymphatic wall were not seen in the present study. Thus it was concluded that the principal morphologically visible routes of transport across renal lymphatic endothelium are provided by intracytoplasmic vesicles and by the typical narrow intercellular spaces between contiguous endothelial cells. No in- formation was obtained on the relative importance of these two routes in the formation of lymph. However, it was concluded that open intercellular gaps do not play an important role under conditions of relatively normal lymph formation.
ACKNOWLEDGMENTS
Thanks are expressed to Ms. H. M. Jarosz and Mrs. L. M. Fox for their technical assistance.
REFERENCES
ALBERTINE, K. H., AND O’MORCHOE, C. C. C. (1979). The distribution and density of the canine renal cortical lymphatic system. Kidney Int. 16,470-480.
BLOOM, W., AND FAWCETT, D. W. (eds.) (1975). Blood and lymph vascular systems. In “A Textbook of Histology,” p. 391. Saunders, Philadelphia.
BRACE, R. A., GRANGER, D. N., AND TAYLOR, A. E. (1977). Analysis of lymphatic protein flux data. II Effect of capillary heteroporosity on estimates of reflection coefficients and ps products. Microvasc. Res. 14, 215-226.

350 ALBERTINE AND O’MORCHOE
BRUNS, R. R., AND PALADE, G. E. (1968). Studies on blood capillaries. I. General organization of blood capillaries in muscle. J. Cell Biol. 37, 244-276.
CASLEY-SMITH, J. R. (1976). Calculations relating to the passage of fluid and protein out of arterial- limb fenestrae, through basement membranes and connective tissue channels, and into venous-limb fenestrae and lymphatics. Microvasc. Res. 12, 13-34.
CASLEY-SMITH, J. R. (1964). Endothelial permeability-the passage of particles into and out of diaphragmatic lymphatics. Quart. J. Exp. Physiol. 49, 365-383.
CASLEY-SMITH, J. R. (1%5). Endothelial permeability. II. The passage of particles through the lymphatic endothelium of normal and injured ears. Brit. J. Exp. Pathol. 46, 35-49.
CASLEY-SMITH, J. R. (1977). Lymph and lymphatics. In “Microcirculation” (G. Kaley and B. M. Altura, eds.), Vol. I, pp. 423-502. University Park Press, Baltimore.
CASLEY-SMITH, J. R. (1%9). The dimensions and numbers of small vesicles in cells, endothelial and mesothelial and the significance of these for endothelial transport. J. Microsc. 90, 251-269.
CASLEY-SMITH, J. R., AND FLOREY, H. W. (l%l). The structure of normal small lymphatics. Quart. J. Exp. Physiol. 46, 101-106.
CASLEY-SMITH, J. R., AND SIMS, M. A. (1976). Protein concentrations in regions with fenestrated and continuous blood capillaries and in initial collecting lymphatics. Microvasc. Res. 12, 245-257.
CASLEY~MITH, J. R., GREEN, H. S., HARRIS, J. L., AND WADEY, P. J. (1975). The quantitative morphology of skeletal muscle capillaries in relation to permeability. Microvnsc. Res. 10, 43-64.
DOBBINS, W. O., AND ROLLINS, E. L. (1970). Intestinal mucosal lymphatic permeability: An electron microscopic study of endothelial vesicles and cell junctions. J. Ultrastruct. Res. 33, 29-59.
ELHAY, S., AND CASLEY-SMITH, J. R. (1976). Mathematical model of the initial lymphatics. Mic- rovasc. Res. 12, 121-140.
ELIAS, H., AND HENNIG, A. (1967). Stereology of the human renal glomerulus. In “Quantitative Methods in Morphology” (E. R. Weibel and H. Elias, eds.), pp. 129-166. Springer-Verlag, Berlin.
FRALEY, E. E., AND WEISS, L. (l%l). An electron microscopic study of the lymphatic vessels in the penile skin of the rat. Amer. J. Anat. 109, 85-101.
FRANK, R. M., WIEDEMANN, P., AND FELLINGER, E. (1977). Ultrastructure of lymphatic capillaries in the human dental pulp. Cell Tissue Res. 178, 229-238.
GARLICK, D. G., AND RENKIN, E. M. (1970). Transport of large molecules from plasma to interstitial fluid and lymph in dogs. Amer. J. Physiol. 219, 1595-1605.
HAYAT, M. A. (1970). In “Principles and Techniques of Electron Microscopy, Biological Applica- tions” (M. A. Hayat, ed.), Vol. 1, pp. 81-84. Van Nostrand Reinhold, New York.
LASSEN, N. A., PARVING, H. H., AND ROSSING, N. (1974). Filtration as the main mechanism of overall transcapillary protein escape from the plasma. Microvasc. Res. 7, i-iv.
LEAK, L. V. (1970). Electron microscopic observations on lymphatic capillaries and the structural components of the connective tissue-lymph interface. Microvasc. Res. 2, 361-394.
LEAK, L. V. (1976). The structure of lymphatic capillaries in lymph formation. Fed. Proc. 35, 1863-1871.
LEAK, L. V., AND BURKE, J. F. (1%8). Electron microscopic study of lymphatic capillaries in the removal of connective tissue fluids and particulate substances. Lymphology 1, 39-52.
LEAK, L. V., AND BURKE, J. F. (1966). Fine structure of the lymphatic capillary and the adjoining connective tissue area. Amer. J. Anat. 118, 785-810.
NORDQUIST, R. E., BELL, R. D., SINCLAIR, R. J., AND KEYL, M. J. (1973). The distribution and ultrastructural morphology of lymphatic vessels in the canine renal cortex. Lymphology 6, 13-19.
OHKUMA, M. (1973). Electron microscopic observation of the renal lymphatic capillary after injection of ink solution. Lymphology 6, 175-181.
PERL, W. (1975). Convection and permeation of albumin between plasma and inter&mm. Microvasc. Res. 10, 83-94.
RENKIN, E. M. (1!364). Transport of large molecules across capillary walls. Physiologist 7, 13-28. RENKIN, E. M., WATSON, P. D., SLOOP, C. H., JOYNER, W. M., AND CURRY, F. E. (1977a). Transport
pathways for fluid and large molecules in microvascular endothelium of the dog’s paw. Microvasc. Res. 14, 205-214.
RENKIN, E. M., JOYNER, W. M., SLOOP, C. H., AND WATSON, P. D. (1977b). Influence of venous pressure on plasma-lymph transport in the dog’s paw: Convective and dissipative mechanisms. Microvasc. Res. 14, 191-204.

LYMPHATIC ULTRASTRUCTURE AND TRANSPORT 351
RHODIN, J. A. G. (1965). Fine structure of the peritubular capillaries of the human kidney. In “Progress in Pyelonephritis” (E. H. Kass, ed.) pp. 391-458. Davis, Philadelphia.
ROJO-ORTEGA, J. M., YEGHIAYAN, E., AND GENEST, J. (1973). Lymphatic capillaries in the renal cortex of the rat. Lab. Invest. 29, 336-341.
TAYLOR, A. E., GRANGER, D. N., AND BRACE, R. A. (1977). Analysis of lymphatic flux data. I. Estimation of the reflection coefficient and permeability surface area product for total protein. Microvasc. Res. 13, 297-313.
WEIBEL, E. R. (1973). Stereological techniques for electron microscopy. In “Principles and Tech- niques of Electron Microscopy, Biological Applications” (M. A. Hayat, ed.), Vol. 3, pp. 237-296. Van Nostrand Reinhold, New York.




















