
Reductions in serum thromboxane, prostacyclin, and leukotriene B4 levels in swine fed a fish oil...
12
EXPERIMENTAL AND MOLECULAR PATHOLOGY 55, 1-12 (1991) Reductions in Serum Thromboxane, Prostacyclin, and Leukotriene B, Levels in Swine Fed a Fish Oil Supplement to an Atherogenic Diet’ D. N. KIM,’ ‘J. SCHMEE,~ C. S. LEE,~,~ 0. SOLIS,~ J. S. ROSS,~ AND W. A. THOMAS~ ‘Department of Pathology and Laboratory Medicine, Albany Medical College, Albany, New York 12208; and ‘Statistical Unit, Union College, Schenectady, New York 12308 Received September 20, 1990, and in revised form March 22, 1991 We have reported previously that fish oil rich in omega-3 fatty acids added to a butter- cholesterol atherogenic diet for swine resulted in marked retardation of the atherosclerotic process which many regard as largely an inflammatory response to injury by excessive lipids in the intima. In this report on the same swine we present serum levels of several ei- cosanoids derived from arachidonic acid via the cyclooxygenase and lipoxygenase path- ways. The study involves six swine fed a high fat, high cholesterol diet (BT group) for 4 months, six swine fed the same diet but with 30 ml/day fish oil added (BT + FO), and five swine fed a low fat, low cholesterol mash diet (MA). The serum eicosanoids were measured by radioimmunoassay. Thromboxane Bz levels (ng/dl: means -+ SEM) were 543 2 49 for MA, 231 * 12 for BT, and 105 2 20 for BT + FO, and all differences were statistically highly significant. 6-Keto FGF,, (a relatively stable prostacyclin metabolite) levels were 249 2 31 for MA, 184 2 12 for BT, and 101 rt 10 for BT + FO, and all differences were significant. Leukotriene B, levels at 4 months were 151 5 25 for MA, 112 ‘- 11 for BT, and 84 2 11 for BT + FO. BT + FO was significantly different from both MA and BT, but BT was not significantly different from MA. Leukotriene C4 levels were not significantly different among the three groups. Of special interest was the effect of the BT diet without the FO additive in reducing several eicosanoid levels compared to MA values. The affected ei- cosanoid levels were reduced still further by the fish oil additive, indicating its ability to inhibit both the cyclooxygenase and the lipoxygenase pathways. The relation of the fish oil-induced inhibition to the observed retardation of atherogenesis is not as yet clear but there are several theoretical possibilities, including reduction in recruitment of monocytes and in proliferation of smooth muscle cells. 0 1991 Academic Press, IK. INTRODUCTION Epidemiologic studies have presented evidence suggesting that populations eat- ing large amounts of fish (and marine mammals) have death rates from athero- sclerotic complications lower than those of populations eating less fish (Bang and Dyerberg; 1985; Kromhaut et al., 1985; Editorial, 1988; Harris, 1989; Willis, 1981). Attention has been focused on the omega-3 polyunsaturated fatty acid content of the fish diet (Moncada and Vane, 1979; Moncada and Radomski, 1985). These fatty acids appear to have antithrombotic and other effects which may account for the lowered death rates from atherosclerotic heart disease (Editorial, 1988; Goodright, Jr., et al., 1982; Harris, 1989). It is also possible that they are antiatherogenic (Pomerantz and Hajjar, 1989). For practical reasons this aspect ’ Supported by NIH Grant HL404901. Also supported in part by BRSG Grant SO 7RR05394. 4 Dr. Choong Sik Lee is a Visiting Research Fellow from the Chungnam National University. School of Medicine, Taejeon. Korea. 1 0014-4800191 $3.00 Copyright 0 1991 hy Academic Press. Inc. All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Reductions in serum thromboxane, prostacyclin, and leukotriene B4 levels in swine fed a fish oil...

EXPERIMENTAL AND MOLECULAR PATHOLOGY 55, 1-12 (1991)
Reductions in Serum Thromboxane, Prostacyclin, and Leukotriene B, Levels in Swine Fed a Fish Oil Supplement to
an Atherogenic Diet’ D. N. KIM,’ ‘J. SCHMEE,~ C. S. LEE,~,~ 0. SOLIS,~ J. S. ROSS,~ AND
W. A. THOMAS~
‘Department of Pathology and Laboratory Medicine, Albany Medical College, Albany, New York 12208; and ‘Statistical Unit, Union College, Schenectady, New York 12308
Received September 20, 1990, and in revised form March 22, 1991
We have reported previously that fish oil rich in omega-3 fatty acids added to a butter- cholesterol atherogenic diet for swine resulted in marked retardation of the atherosclerotic process which many regard as largely an inflammatory response to injury by excessive lipids in the intima. In this report on the same swine we present serum levels of several ei- cosanoids derived from arachidonic acid via the cyclooxygenase and lipoxygenase path- ways. The study involves six swine fed a high fat, high cholesterol diet (BT group) for 4 months, six swine fed the same diet but with 30 ml/day fish oil added (BT + FO), and five swine fed a low fat, low cholesterol mash diet (MA). The serum eicosanoids were measured by radioimmunoassay. Thromboxane Bz levels (ng/dl: means -+ SEM) were 543 2 49 for MA, 231 * 12 for BT, and 105 2 20 for BT + FO, and all differences were statistically highly significant. 6-Keto FGF,, (a relatively stable prostacyclin metabolite) levels were 249 2 31 for MA, 184 2 12 for BT, and 101 rt 10 for BT + FO, and all differences were significant. Leukotriene B, levels at 4 months were 151 5 25 for MA, 112 ‘- 11 for BT, and 84 2 11 for BT + FO. BT + FO was significantly different from both MA and BT, but BT was not significantly different from MA. Leukotriene C4 levels were not significantly different among the three groups. Of special interest was the effect of the BT diet without the FO additive in reducing several eicosanoid levels compared to MA values. The affected ei- cosanoid levels were reduced still further by the fish oil additive, indicating its ability to inhibit both the cyclooxygenase and the lipoxygenase pathways. The relation of the fish oil-induced inhibition to the observed retardation of atherogenesis is not as yet clear but there are several theoretical possibilities, including reduction in recruitment of monocytes and in proliferation of smooth muscle cells. 0 1991 Academic Press, IK.
INTRODUCTION
Epidemiologic studies have presented evidence suggesting that populations eat- ing large amounts of fish (and marine mammals) have death rates from athero- sclerotic complications lower than those of populations eating less fish (Bang and Dyerberg; 1985; Kromhaut et al., 1985; Editorial, 1988; Harris, 1989; Willis, 1981). Attention has been focused on the omega-3 polyunsaturated fatty acid content of the fish diet (Moncada and Vane, 1979; Moncada and Radomski, 1985). These fatty acids appear to have antithrombotic and other effects which may account for the lowered death rates from atherosclerotic heart disease (Editorial, 1988; Goodright, Jr., et al., 1982; Harris, 1989). It is also possible that they are antiatherogenic (Pomerantz and Hajjar, 1989). For practical reasons this aspect
’ Supported by NIH Grant HL404901. Also supported in part by BRSG Grant SO 7RR05394. 4 Dr. Choong Sik Lee is a Visiting Research Fellow from the Chungnam National University. School
of Medicine, Taejeon. Korea.
1
0014-4800191 $3.00 Copyright 0 1991 hy Academic Press. Inc. All rights of reproduction in any form reserved.

2 KIM ET AL.
has been investigated thus far only in experimental animals and results have been mixed. Some have reported retarded atherogenesis (Davis ef al., 1987; Hollander et al., 1987; Kim et al., 1989; Weiner et al., 1986; Zhu et al., 1988; Lichtenstein and Chobanian, 1990), others no effect (Rich et al., 1989; Clubb et al., 1989), and still others enhancement (Thiery and Seidel, 1987).
Weiner et al. (1986) and ourselves (Kim et al., 1989) have reported marked retardation of atherogenesis in swine fed hyperlipidemic diets to which a fish oil (cod liver oil) supplement was added. We have suggested that the retardation observed in our study was due in part to a modest lowering of plasma cholesterol levels (circa 20%) and in part to a marked shift in lipoprotein patterns from predominantly apo B and E containing fractions (VLDL, IDL, and part of LDL) to predominantly apo B-only fractions (conventional LDL). Weiner et al. (1986) did not find terminal plasma cholesterol levels to be significantly different be- tween fish oil (FO)-treated and untreated groups. They did find serum thrombox- ane (TXB,) levels to be lowered and suggested that this might be a factor in the retardation of atherogenesis.
In our original study (Kim et al., 1989) we had not investigated levels of TXB, or any other eicosanoids. However, we still had frozen serum samples available from our experiment and with this material we have investigated levels of several eicosanoids including TXB,, 6-keto prostaglandin F,, (6-keto PGF,,), leukotriene B, (LTB,), and leukotriene C, (LTC,). The eicosanoids found in serum are in part those present in plasma and in part are generated from platelets and leukocytes during clotting. Also the antibodies available for the radioimmunoassays may have cross-reactivities that must be taken into account in making interpretations. Nevertheless, we felt that comparison of values would be useful among swine fed a low fat, low cholesterol mash diet; hyperlipidemics fed a high saturated fat, high cholesterol (BT) diet; and a treated group fed the BT diet plus a fish oil supple- ment (BT + FO), especially since we had already reported extensive quantitative atherosclerotic lesion data and plasma lipoprotein analyses on these swine.
MATERIALS AND METHODS
Details regarding most of the materials and methods on morphometry of lesions and lipoprotein analysis have been presented in two previous reports on this experiment (Kim et al., 1989, 1990). We shall only present here methods on the measurement of eicosanoid metabolites.
Experimental Design
Seventeen young male Yorkshire swine weighing 11 .O & 0.4 kg were divided into three dietary groups and fed respective diets for 4 months. Six were given a butter-cholesterol hyperlipidemic diet (BT), six were given the BT diet plus 30 ml of cod liver oil daily (BT + FO), and five were given a low fat, low cholesterol commercial mash diet (MA). Plasma cholesterol levels were monitored monthly (Leffler, 1959). Plasma lipoproteins were determined terminally and atheroscle- rotic lesions were quantified after autopsy.
Measurements of the Eicosanoid Metabolites
Fasting blood was drawn at 1 week, 1 month, and 4 months from the anterior vena cava (jugular vein) with a plastic syringe into a conical plastic tube. The blood was incubated in a water bath for 2 hr at 37°C and centrifuged for 20 min at

FISH OIL, EICOSANOIDS, AND ATHEROGENESIS 3
15OOg. Serum was divided into small aliquots and stored at - 20°C until assayed. Thromboxane B,, 6-keto PGFi,, leukotriene B,, and leukotriene C4 were mea- sured by radioimmunoassay according to the manufacturer’s (New England Nu- clear Research Products, Boston, MA) protocol using thromboxane B, 3H-RIA kit (NEK-007), 6-keto prostaglandin F,, 3H-RIA kit (NEK-008), leukotriene C, 3H- RIA kit (NEK-030), and leukotriene B, 3H-RIA kit (NEK-037). Cross-reactivity of anti-TXB, antibody with PGD, is 2.0% and it is less than 0.1% with other pros- taglandins: PGE,, PGF,,, 6-keto PGF,,, and PGA,. Cross-reactivity of anti- 6-keto PGF,, with PGE, is 2.5% and it is less than 0.3% with others: PGF,, PGE,, PGF,, PGA,, PGA,, TXB,, and 13,lCdihydro-15-keto PGF,,. Cross-reactivity of anti-LTB, with 5,12-diHETE is 3.6%, 1.3% with 20-OH-LTB,, 1.0% with 6- TransLTB,, 0.6% with 12-epi-LTB,, and less than 0.1% with many others, in- cluding LTC, and TXB,. Cross-reactivity of anti-LTC, with ll-trans-LTD, is 60.5%, with LTD, 55.3%, with LTE, 8.6%, and with all others less than 0.1%. Therefore, the result of the LTC, assay must be viewed with caution.
There exists a possibility that Al7-6-keto PGF,, (a stable metabolite of PGI,) might cross-react with an antibody for 6-keto-PGF,,. The assay of A17-6-keto PGF,, (purity 98%, Biomol, Plymouth Meeting, PA) shows cross-reactivity with the antibody for 6-keto PGF,, to be 9.5%. Similarly, assay of cross-reactivity of LTB, (purity 98%, Caymen Chemical Co., Ann Arbor, MI) with antibody for LTB, shows 6.8% cross-reactivity. TXB, might likewise cross-react with antisera of TXB,. We have not checked the cross-reactivity of TXB, yet.
Assays for TXB* and 6-keto PGF,, were carried out in duplicate first in time blocks of 1 week, 1 month, and 4 months and then assayed in samples of 1 week, 1 month, and 4 months at the same time to check if the values measured at different times were consistent. Both sets of values agreed well. LTB, and LTC, were measured at 1 week and 4 months using the same approach.
We also had available some EDTA plasma from about half of the swine for the eicosanoid part of the study. TXB, values from the plasma were much lower than those for serum and not useful in making comparisons among groups. Thus serum values were largely reflecting thromboxane formed by platelets during the clotting process. 6-Keto PGF,, values were also lower in plasma than in serum. LTB, values were generally similar in plasma and serum. We also added aspirin in a few serum samples and this markedly reduced eicosanoid values as would be ex- pected.
We decided that for our purpose of making overall comparisons among dietary groups, the serum values reflecting mainly the capacity of the platelets and leu- kocytes to produce eicosanoids were the most meaningful. They are especially relevant since the atherosclerotic lesions contain large numbers of leukocytes (monocyte/macrophages). The plasma values are not presented in this report.
Statistical Methods
Comparisons of the average levels between pairs of diet groups were based on the two-sample t test or where indicated the Mann-Whitney CJ test. For overall statements we also checked the t test results against several ANOVA post hoc analyses such as Fisher’s protected least significant difference (LSD) or Scheffe’s method (Sokal and Rohlf, 1981; Gagnon et al., 1989). The dimensionality of the data was investigated using the method of principal components (Velleman, 1989). We calculated partial association by means of multiple regression and ANCOVA.
4 KIM ET AL.
In most instances the multivariate results confirmed the univariate findings and so they are not all shown. In exploring the relationships sophisticated graphical techniques were used.
Pertinent Previous Results
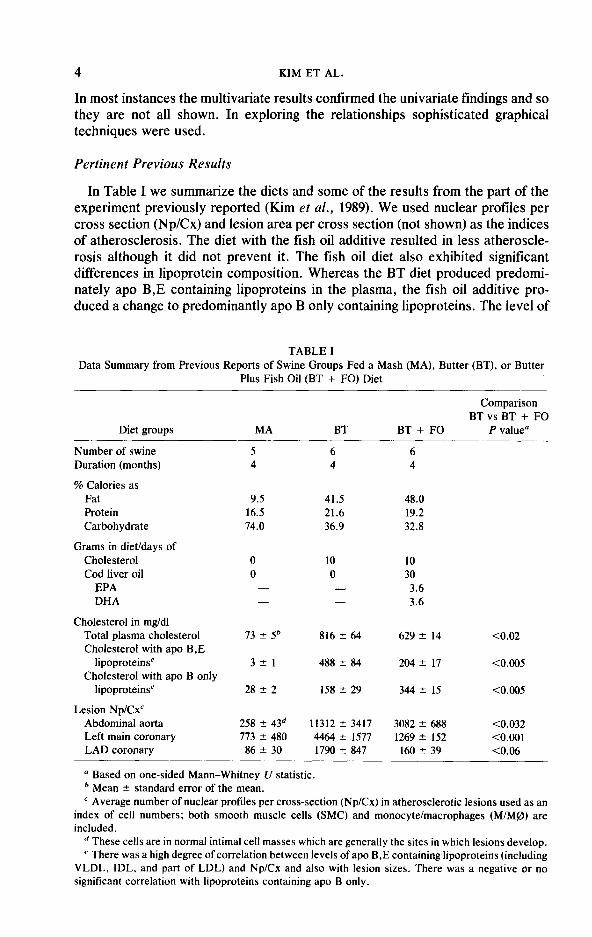
In Table I we summarize the diets and some of the results from the part of the experiment previously reported (Kim et al., 1989). We used nuclear profiles per cross section (Np/Cx) and lesion area per cross section (not shown) as the indices of atherosclerosis. The diet with the fish oil additive resulted in less atheroscle- rosis although it did not prevent it. The fish oil diet also exhibited significant differences in lipoprotein composition. Whereas the BT diet produced predomi- nately apo B,E containing lipoproteins in the plasma, the fish oil additive pro- duced a change to predominantly apo B only containing lipoproteins. The level of
TABLE I Data Summary from Previous Reports of Swine Groups Fed a Mash (MA), Butter (BT), or Butter
Plus Fish Oil (BT + FO) Diet
Diet groups
Number of swine Duration (months)
% Calories as Fat Protein Carbohydrate
Grams in diet/days of Cholesterol Cod liver oil
EPA DHA
Cholesterol in mg/dl Total plasma cholesterol Cholesterol with apo B,E
lipoproteinse Cholesterol with apo B only
lipoproteins’
Lesion Np/Cx’ Abdominal aorta Left main coronary LAD coronary
MA
5 4
BT BT + FO
Comparison BTvs BT + FO
P value”
6 6 4 4
9.5 41.5 48.0 16.5 21.6 19.2 74.0 36.9 32.8
0 0
- -
10 30 3.6 3.6
73 2 5b 816 t 64 629 k 14 co.02
321 488 2 84 204% 17 co.005
28 2 2 158 2 29 344 2 15 co.005
258 2 43d 11312 2 3417 3082 * 688 co.032 773 2 480 4464 2 1577 1269 2 152 <O.OOl
86 k 30 1790 k 847 160 2 39 co.06
a Based on one-sided Mann-Whitney U statistic. b Mean 2 standard error of the mean. ’ Average number of nuclear profiles per cross-section (Np/Cx) in atherosclerotic lesions used as an
index of cell numbers; both smooth muscle cells (SMC) and monocyte/macrophages (M/M@) are included.
d These cells are in normal intimal cell masses which are generally the sites in which lesions develop. c There was a high degree of correlation between levels of apo B,E containing lipoproteins (including
VLDL, IDL, and part of LDL) and Np/Cx and also with lesion sizes. There was a negative or no significant correlation with lipoproteins containing apo B only.

FISH OIL, EICOSANOIDS, AND ATHEROGENESIS 5
apo B,E carrying lipoproteins correlated stronger than any other terminal lipid variable with the lesion Np/Cx.
RESULTS
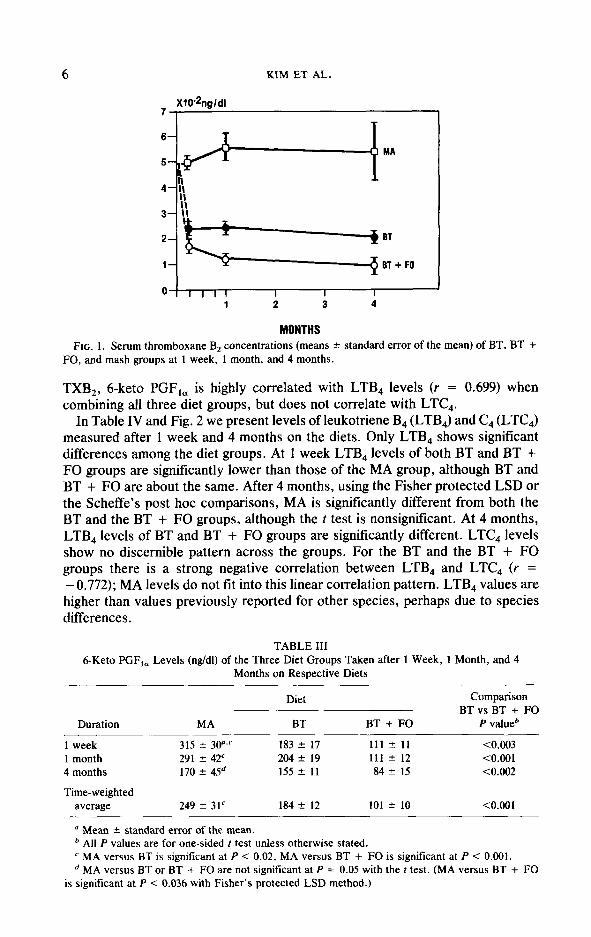
In Table II and Fig. 1 we present data on serum TXB, levels in the three dietary groups taken after 1 week, 1 month, and 4 months, as well as a time-weighted average for the entire period. The MA group consistently exhibited the highest level. The BT group showed levels significantly lower than those of the MA group, but showed levels significantly higher than those of the BT + FO group. The relationship between time-weighted averages of TXB, and total apo B,E carrying lipoprotein levels or Np/Cx (not shown in the tables) exhibited an entirely nonlinear pattern, with a negative correlation (r = 0.504 for apo B,E lipoproteins and r = 0.376 for Np/Cx) for data of all three groups and a positive correlation (r = 0.493 for apo B,E lipoproteins and r = 0.293 for Np/Cx) if data from the MA diet are excluded. The TXB, average showed a very high correlation with the 6-keto PGF,, average, whether the MA animals were included (r = 0.892) or not (r = 0.913). In the MA diet group both the TXB, and the leukotriene B, and C, levels were higher than in the hyperlipidemic diet groups. As a result the TXB, average was highly correlated with leukotriene B, levels (r = 0.760), but nonsig- nificantly so with leukotriene C, levels since MA levels were considerably less elevated compared to the other two diet groups and variables.
Table III shows data for 6-keto PGF,, (PGI, metabolite) levels. After 1 week and 1 month all three diets are significantly different in terms of 6-keto PGF,, levels. After 4 months three of the MA animals exhibited a considerable lowering of PGI, levels compared with values from earlier times, while two animals re- mained at fairly high levels. This increase in the range of the data accounts for the large standard error of the mean which produces a nonsignificant difference be- tween the MA and the BT + FO 6-keto PGF,, levels. Combining the BT and BT + FO one can observe a significant correlation (r = 0.666) between the time- weighted average of 6-keto PGF,, and the total of apo B,E lipoproteins (r = 0.476 for Np/Cx). Including the MA group produces a nonlinear relationship and the correlation is close to zero for both apo B,E lipoproteins and Np/Cx. As with
TABLE XI Serum Thromboxane B, Levels (ng/dl) of the Three Diet Groups Taken after 1 Week, 1 Month,
and 4 Months on Respective Diets
Diet Comparison BTvs BT + FO
Duration MA BT BT + FO P valueb
1 week 500 f 30”,’ 242 * 23 178 k 22 <0.037 1 month 557 k 57’ 245 + 20 124 2 27 <0.003 4 months 537 ” 126‘+ 209 2 21 94 + 34 <0.008
Time-weighted average 543 f 49’ 231 + 12 105 2 20 <O.ool
0 Mean + standard error of the mean. b All P values are for one-sided t test. ’ MA versus BT and BT + FO are significant at P < 0.001. ’ MA versus BT and BT + FO are significant at P < 0.002.
KIM ET AL.
0 ,111 I I 1
1 2 3 4
MONTHS FIG. 1. Serum thromboxane B2 concentrations (means ? standard error of the mean) of BT, BT +
FO, and mash groups at 1 week, 1 month, and 4 months.
TXB,, 6-keto PGF,, is highly correlated with LTB, levels (r = 0.699) when combining all three diet groups, but does not correlate with LTC,.
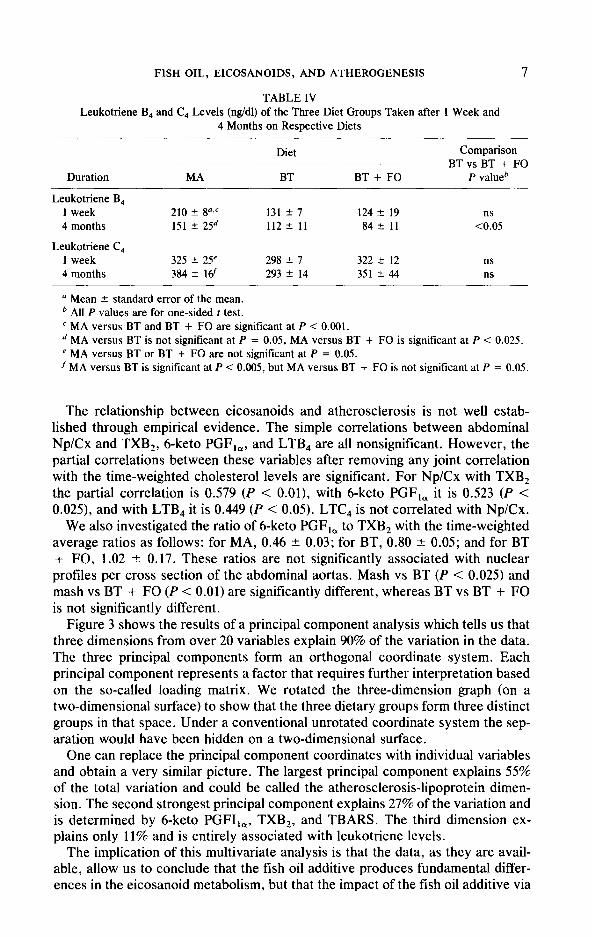
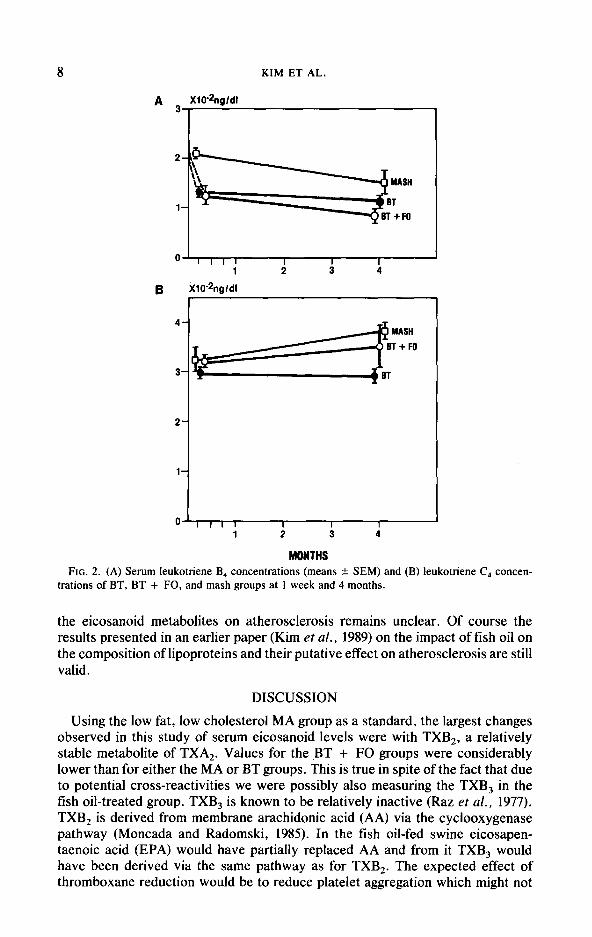
In Table IV and Fig. 2 we present levels of leukotriene B, (LTB,) and C, (LTC$ measured after 1 week and 4 months on the diets. Only LTB, shows significant differences among the diet groups. At 1 week LTB, levels of both BT and BT + FO groups are significantly lower than those of the MA group, although BT and BT + FO are about the same. After 4 months, using the Fisher protected LSD or the Scheffe’s post hoc comparisons, MA is significantly different from both the BT and the BT + FO groups, although the t test is nonsignificant. At 4 months, LTB, levels of BT and BT + FO groups are significantly different. LTC, levels show no discernible pattern across the groups. For the BT and the BT + FO groups there is a strong negative correlation between LTB, and LTC, (r = - 0.772); MA levels do not fit into this linear correlation pattern. LTB, values are higher than values previously reported for other species, perhaps due to species differences.
TABLE III 6-Keto PGF,, Levels (ng/dl) of the Three Diet Groups Taken after 1 Week, 1 Month, and 4
Months on Respective Diets
Duration MA
Diet
BT BT + FO
Comparison BTvsBT + FO
P valueb
1 week 1 month 4 months
Time-weighted average
315 !z 3cr= 183 f 17 111 2 11 <0.003 291 + 42’ 204* 19 111 * 12 <O.ool 170 + 45d 15.5 f 11 842 15 <0.002
249 k 31’ 184 2 12 101 * 10 <O.ool
L1 Mean 2 standard error of the mean. b All P values are for one-sided t test unless otherwise stated.
c MA versus BT is significant at P < 0.02, MA versus BT + FO is significant at P < 0.001. d MA versus BT or BT + FO are not significant at P = 0.05 with the t test. (MA versus BT + FO
is significant at P < 0.036 with Fisher’s protected LSD method.)
FISH OIL, EICOSANOIDS, AND ATHEROGENESIS 7
TABLE IV Leukotriene B, and C, Levels (ng/dl) of the Three Diet Groups Taken after 1 Week and
4 Months on Respective Diets
Diet Comparison BT vs BT + FO
Duration MA BT BT + FO P valueb
Leukotriene B, 1 week 210 2 8”,’ 131 t I 124 2 19 ns 4 months 151 + 25d 112 2 11 84-c 11 -co.05
Leukotriene C, 1 week 325 + 25’ 298 f 7 322 2 12 ns 4 months 384 k 16 293 f 14 351 244 ns
LI Mean 2 standard error of the mean. b All P values are for one-sided t test. ’ MA versus BT and BT + FO are significant at P < 0.001. d MA versus BT is not significant at P = 0.05, MA versus BT + FO is significant at P < 0.025. e MA versus BT or BT + FO are not significant at P = 0.05. /‘MA versus BT is significant at P < 0.005, but MA versus BT + FO is not significant at P = 0.05.
The relationship between eicosanoids and atherosclerosis is not well estab- lished through empirical evidence. The simple correlations between abdominal Np/Cx and TXB,, 6-keto PGF,,, and LTB, are all nonsignificant. However, the partial correlations between these variables after removing any joint correlation with the time-weighted cholesterol levels are significant. For Np/Cx with TXB, the partial correlation is 0.579 (P < O.Ol), with 6-keto PGF,, it is 0.523 (P < 0.025), and with LTB, it is 0.449 (P < 0.05). LTC, is not correlated with Np/Cx.
We also investigated the ratio of 6-keto PGF,, to TXB, with the time-weighted average ratios as follows: for MA, 0.46 + 0.03; for BT, 0.80 2 0.05; and for BT + FO, 1.02 * 0.17. These ratios are not significantly associated with nuclear profiles per cross section of the abdominal aortas. Mash vs BT (P < 0.025) and mash vs BT + FO (P < 0.01) are significantly different, whereas BT vs BT + FO is not significantly different.
Figure 3 shows the results of a principal component analysis which tells us that three dimensions from over 20 variables explain 90% of the variation in the data. The three principal components form an orthogonal coordinate system. Each principal component represents a factor that requires further interpretation based on the so-called loading matrix. We rotated the three-dimension graph (on a two-dimensional surface) to show that the three dietary groups form three distinct groups in that space. Under a conventional unrotated coordinate system the sep- aration would have been hidden on a two-dimensional surface.
One can replace the principal component coordinates with individual variables and obtain a very similar picture. The largest principal component explains 55% of the total variation and could be called the atherosclerosis-lipoprotein dimen- sion. The second strongest principal component explains 27% of the variation and is determined by 6-keto PGFI,,, TXB,, and TBARS. The third dimension ex- plains only 11% and is entirely associated with leukotriene levels.
The implication of this multivariate analysis is that the data, as they are avail- able, allow us to conclude that the fish oil additive produces fundamental differ- ences in the eicosanoid metabolism, but that the impact of the fish oil additive via
KIM ET AL.
00 1 2 3 4
MONTHS FIG. 2. (A) Serum Ieukotriene B, concentrations (means 5 SEM) and (B) leukotriene C4 concen-
trations of BT, BT + FO, and mash groups at 1 week and 4 months.
the eicosanoid metabolites on atherosclerosis remains unclear. Of course the results presented in an earlier paper (Kim et al., 1989) on the impact of fish oil on the composition of lipoproteins and their putative effect on atherosclerosis are still valid.
DISCUSSION
Using the low fat, low cholesterol MA group as a standard, the largest changes observed in this study of serum eicosanoid levels were with TXB,, a relatively stable metabolite of TXA,. Values for the BT + FO groups were considerably lower than for either the MA or BT groups. This is true in spite of the fact that due to potential cross-reactivities we were possibly also measuring the TXB, in the fish oil-treated group. TXB, is known to be relatively inactive (Raz et al., 1977). TXBz is derived from membrane arachidonic acid (AA) via the cyclooxygenase pathway (Moncada and Radomski, 1985). In the fish oil-fed swine eicosapen- taenoic acid (EPA) would have partially replaced AA and from it TXB, would have been derived via the same pathway as for TXB,. The expected effect of thromboxane reduction would be to reduce platelet aggregation which might not

FISH OIL, EICOSANOIDS, AND ATHEROGENESIS 9
a
FIG. 3. (A) BT, (0) BT + FO, (0) MA. This graph of the three largest principal components (PC) is rotated in such a way as to show the clear separation of the three dietary groups. The largest principal component, 1st pc, explains 55% of the total variation in the data and is primarily associated
with the atherosclerosis and lipoprotein variables. The second largest, 2nd pc, explains 27% of the total variation and is primarily associated with TBARS, 6-keto PGF,,, thromboxane B,, and LTB,. The third largest, 3rd pc, explains 11% of the total variation and is associated with LTC,. As a consequence of the high association of individual variables with the three principal components, a three-dimensional graph of average cholesterol, average PG12, and average LTC, shows a very similar pattern (not shown).
only be antithrombogenic but also antiatherogenic by reducing platelet-arterial wall interactions (Pomerantz and Hajar, 1989).
It was rather unexpected to find that the atherogenic BT diet also reduced serum TXB, levels by itself (O’Dea et al., 1988). The reasons for this are not clear, but possibly are related to the presumed increased cholesterol content of the platelet and other cell membranes. Although unlikely, the omega-3 linolenic fatty acid content of the butter may have been a contributing factor (O’Dea et al., 1988).
Human volunteers (Barradas et al., 1989) given an oleic acid supplement (20 ml of olive oil) daily for 8 weeks showed significantly reduced arachidonic acid content of platelet membrane phospholipids and inhibition of platelet aggregation. Although the authors did not measure TXB, they suggested that reduced arachi- donic acid might have resulted in a reduced substrate for TXA, synthesis. Thus various fatty acids could influence eicosanoid metabolism.
Ballou and Cheung (1985) have shown in vitro that human platelet phospholi- pase A, activity was inhibited by various unsaturated fatty acids. The significance in vivo is not known, but it is possible that the fatty acid effects on eicosanoid metabolism might not be limited to the effects on substrate concentrations.
The next largest change observed vis-a-vis the MA values was with serum 6-keto PGF,,. Values for BT + FO were lowest and those for BT were interme- diate. Cross-reactivity with PGI, metabolites was probably not a factor here since we have evidence that the cross-reactivity was less than 10%. PGI, is an anti- platelet aggregator counterbalancing TXA, (Moncada and Radomski, 1985). It also has other functions that might be antiatherogenic, including antiproliferative activity and antinecrosis features (Pomerantz and Hajjar, 1989). The most impor- tant source of PGI, is the inner portion of the arterial wall (Moncada and Radom-

10 KIM ET AL.
ski, 1985). This source is not significantly reflected in the serum since there is very little in the plasma. Most of that measured in the serum is produced by the leukocytes during the clotting process. How this capacity relates to that of the arterial wall is unknown. Lowering of PGI, production is an undesirable feature with respect to retardation of atherogenesis since it is presumed to be cytopro- tective (Moncada and Radomski, 1985). The ratio of PGI, to TXA, is presumed by some to be important but at least in the serum this was not significantly different between the BT diet and BT + FO diet. Since PGI, is as active as PGI, in contrast to TXA, and TXA,, the PGI, level must be included in the overall balance be- tween thromboxanes and prostacyclins (Dyerberg et al., 1978). At least in some situations in man PGI, + PGI, levels have been shown to be increased by fish oil over baseline PG12 levels (Fischer et al., 1986, 1987). In rats during dietary fish oil supplementation, there were increases in metabolites (Knapp and Salem, 1989). This may or may not be applicable to all other species (Morita et al., 1983).
Changes from control MA values in the serum levels of lipoxygenase products of AA were not as impressive as those with cyclooxygenase products. As with TXB, we have the problem in the fish oil-treated group with potential cross- reactivities with relatively inactive EPA derivatives of the LTB, (Terano et al., 1984; Lee et al., 1984) and LTC, series. Nevertheless serum LTB, as measured was significantly lower at 4 months in the BT + FO group than in the BT group. LTB, is a powerful chemotactic agent for monocytes and neutrophils (Soter et al., 1983; Camp et al., 1983). Neutrophils are seldom seen in atherosclerotic lesions but monocyte-derived macrophages are a prominent feature and they constituted about half of the lesion cells in these swine with smooth muscle cells (SMC) supplying most of the remainder (Kim et al., 1989). The ratio of lesion monocyte/ macrophages to SMC was not changed by the FO although total numbers were reduced considerably since the lesions were much smaller and had fewer cells. Also the number of monocytes attached to lesion endothelium was significantly reduced by the fish oil additive although numbers were still higher than in mash- fed swine (Kim et al., 1990). The latter effect has also been shown in rats (Rogers and Karnovsky, 1988). However, this is a complicated subject as there are ap- parently many chemoattractants for monocytes in lesions. Gerrity et al. (1985, 1986) have presented evidence from swine, suggesting that these include a che- motactic agent that is special for atherogenesis.
Serum levels of LTC, which is a smooth muscle relaxant did not show signif- icant differences among the three groups. As with other serum eicosanoids this does not necessarily reflect events at the arterial wall level.
We have previously shown in the same swine of this study that lesion sizes and cell numbers are highly positively correlated with mean time-weighted plasma cholesterol levels and with the apo B,E containing lipoproteins measured termi- nally. The eicosanoids TXB,, 6-keto PGF,,, and LTB, seem to explain a small portion of the atherosclerosis variable, above and beyond what can be explained by the time-weighted cholesterol levels or terminal apo B,E lipoproteins. Since these three eicosanoids are highly correlated themselves, they seem to explain a common phenomenon as indicated by the results of the principal component analysis.
In any event it seems clear that eicosanoid metabolism is inhibited by the fish oil additive as one would have expected. It is also clear that eicosanoid metabo- lism is inhibited by the high saturated fat, high cholesterol diet alone and in

FISH OIL, EICOSANOIDS, AND ATHEROGENESIS I1
directions that might not have been anticipated. If the serum eicosanoids were truly reflective of the processes in the arterial wall and if the ratio were important to lesion size, we would have expected that the difference in the ratios of TXB, to 6-keto PGF,, between the BT and the BT + FO group would have been larger. Obviously, further studies are needed to investigate PGI, and other eicosanoid levels of the arterial wall.
ACKNOWLEDGMENTS
The authors thank Mrs. Gunnel Lunden for radioimmunoassay, Mr. James Baker for lipoprotein chemistry, Mrs. Christine Sheehan and Mrs. Marty Rosner for morphometry, Ms. Betty Dougherty for typing this manuscript, and Mr. Edward Whinnery for illustrations.
REFERENCES
BALLOU, L. R., and CHEUNG, W. J. (1985). Inhibition of human platelet phospholipase A, activity by unsaturated fatty acids. Proc. Natl. Acad. Sci. USA 82, 371-375.
BANG, H. O., and DYERBERG, J. (1985). Fish oil consumption and mortality from coronary heart disease. N. Engl. J. Med. 313, 822-823.
BARRADAS, M. A., CHRISTOFIDES, J. A., JEREMY, J. Y., MIKHAILIDIS, D. P., FRY, D. E., and DAN- DONA, P. (1989). Oleic acid supplementation alters platelet function and membrane phospholipid arachidonic acid content. Biochem. Sot. Trans. 17, 505-506.
CAMP, R. D. R., COUTTS, A. A., GREAVES, M. W., KAY, A. B., and WALPORT, J. M. (1983). Re- sponse of human skin to intradermal injection of leukotrienes C,, D,, and B,. Br. J. Pharmacol. 80, 497-502.
CLUBB, F. J., SCHNITZ, J. M., BUTLER, M. M., BUJA, L. M., WILLERSON, J. T., and CAMPBELL, W. B. (1989). Effect of dietary omega-3 fatty acid on serum lipids, platelet function, and athero- sclerosis in Watanabe heritable hyperlipidemic rabbits. Arteriosclerosis 9, 529-537.
DAVIS, H. R., BRIDENSTINE, R. T., VESSELINOVITCH, D., and WISSLER, R. W. (1987). Fish oil in- hibits development of atherosclerosis in Rhesus monkeys. Arteriosclerosis 7, 441449.
DYERBERG, J., BANG, O., STOFFERSEN, E., MONCADA, S., and VANE, J. R. (1978). Eicosapentaenoic acid and prevention of thrombosis and atherosclerosis? Lancet 2, 117-I 19.
Editorial. (1988). Fish oil. Lancet 1, 1081-1083. FISCHER, S., VISCHER, A., PREAC-MURSIC, V., and WEBER, P. C. (1987). Dietary docosahexaenoic
acid is retroconverted in man to eicosapentaenoic acid, which can be quickly transformed to pros- taglandin I,. Prostaglandins 34, 367-375.
FISCHER, S., WEBER, P. C., and DYERBERG, J. (1986). The prostacyclin/thromboxane balance is favorably shifted in Greenland Eskimos. Prostaglandins 32, 235-241.
GAGNON, T., et al. (1989). “SuperAnova: Abacus Concepts.” Berkeley, CA. GERRITY, R. G. (1986). Cellular events in early atherogenesis in swine. In “Swine in Biomedical
Research” (M. E. Tumbleson, ed.), Vol. 3, pp. 1497-1509. Plenum, New York. GERRITY, R. G., Goss, J. A., and SOBY, L. (1985). Control of monocyte recruitment by chemotactic
factors(s) in lesion-prone areas of swine aorta. Arteriosclerosis 5, 5566. GOODNIGHT, S. H., JR., HARRIS, W. S., CONNOR, W. E., and ILLINGWORTH, D. R. (1982). Review:
Polyunsaturated fatty acids, hyperlipidemia, and thrombosis. Arteriosclerosis 2, 87-113. HARRIS, W. S. (1989). Fish oils and plasma lipid and lipoprotein metabolism in humans: A critical
review. J. Lipid Res. 30, 785-807. HOLLANDER, W., HONG, S., KIRKPATRICK, B. J., LEE, A., COLOMBO, M., and PRUSTY, S. (1987).
Differential effects of fish oil supplementation in atherosclerosis. Arteriosclerosis 7, 527a. KIM, D. N., Ho, H.-T., LAWRENCE, D. A., SCHMEE, J., and THOMAS, W. A. (1989). Modification of
lipoprotein patterns and retardation of atherosclerosis by a fish oil supplement to a hyperlipidemic diet for swine. Atherosclerosis 76, 35-54.
KIM, D. N., SCHMEE, J., and THOMAS, W. A. (1990). Dietary fish oil added to a hyperlipidemic diet for swine results in reduction in the excessive number of monocytes attached to arterial endothe- hum. Atherosclerosis 81, 209-216.
KNAPP, H. R., and SALEM, N., JR. (1989). Formation of PGI, in the rat during dietary fish oil supplementation. Prostaglandins 38, 509-522.
KROMHOUT, D., BOSSCHIETER, E. B., and COLJLNADER, C. L. (1985). The inverse relation between

12 KIM ET AL.
fish consumption and 20-year mortality from coronary heart disease. N. Engl. J. Med. 312, 1205- 1209.
LEE, T. H., MENCIA-HUERTA, J.-M., SHIH, C., COREY, E. J., LEWIS, R. A., and AUSTEN, K. F. (1984). Effects of exogenous arachidonic, eicosapentaenoic, and docosahexaenoic acids on the generation of S-lipoxygenase pathway products by ionophore-activated human neutrophils. J. C/in. Invest. 74, 1922-1933.
LEFFLER, H. H. (1959). Estimation of cholesterol in serum. Am. J. Clin. Puthol. 31, 310-313. LICHTENSTEIN, A. H., and CHOBANIAN, A. V. (1990). Effect offish oil on atherogenesis in Watanabe
heritable hyperlipidemic rabbit. Arteriosclerosis 10, 597-606. MONCADA, S.. and RADOMSKI, M. W. (1985). The problems and the promise of prostaglandin influ-
ences in atherogenesis. Ann. NY Acad. Sci. 454, 121-130. MONCADA, S., and VANE, J. R. (1979). Pharmacology and endogenous roles of prostaglandin and
endoperoxides, thromboxane A,, and prostacyclin. Pharmacol. Rev. 30, 293-331. MORITA, I., TAKAHASHI. R., SAITO, Y., and MUROTA, S. (1983). Effects of eicosapentaenoic acid on
archidonic acid metabolism in cultured vascular cells and platelets: Species difference. Thromb. Res. 31, 21 l-217.
O’DEA, K., STEEL. M., NAUGHTON, J., SINCLAIR, A., HOPKINS, G., ANGUS, J., HE, G.-W., NIALL, M., and MARTIN, T. J. (1988). Butter-enriched diets reduce arterial prostacyclin production in rats. Lipids 23, 234-241.
POMERANTZ, K. B., and HAJJAR, D. P. (1989). Review. Eicosanoids in regulation of arterial smooth muscle cell phenotype, proliferative capacity, and cholesterol metabolism. Arteriosclerosis 9, 413- 429.
RAZ, A., MINKES, M. S., and NEEDLEMAN, P. (1977). Endoperoxides and thromboxanes. Structural determinants for platelet aggregation and vasoconstriction. Biochim. Biophys. Acta 488, 305-3 11.
RICH, S., MILLER, J. F.. CHAROUS, S., DAVIS, H. R., SHANKS, P., GLAGOV, S., and LANDS, W. E. M. (1989). Development of atherosclerosis in genetically hyperlipidemic rabbit during chronic fish-oil ingestion. Arteriosclerosis 9, 189-194.
ROGERS, K. A., and KARNOVSKY, M. J. (1988). Dietary fish oil enhances monocyte adhesion and fatty streak formation in the hypercholesterolemic rat. Am. J. Pathol. 132, 383-388.
SOKAL, R. R., and ROHLF, F. J. (1981). “Biometry,” 2nd ed. Freeman, San Francisco. SOTER, N. A., LEWIS, R. A., COREY, E. J., and AUSTEN, K. F. (1983). Local effects of synthetic
leukotrienes (LTC,, LTD,, LTE, and LTB,) in human skin. J. Invest. Dermatol. 80, 115-l 19. TERANO, T., SALMON, J. A., and MONCADA, S. (1984). Biosynthesis and biologic activity of Ieuko-
triene B,. Prostaglandins 27, 217-232. THIERY, J., and &DEL, D. (1987). Fish oil feeding results in an enhancement of cholesterol-induced
atherosclerosis in rabbits. Atherosclerosis 63, 53-56. VELLEMAN, P. (1989). “Data Desk 3.0.” Odesta Corp., Northbrook, IL. WEINER, B. H., OCKENE, I. S., LEVINE, P H., CU~NOUD, H. F., FISHER, M., JOHNSON, B. F.,
DAOUD, A. S., JARMOLYCH, J., HOSMER, D., JOHNSON, M. H., NATALE, A., VAUDREUIL, C., and HOOGASIAN, J. J. (1986). Inhibition of atherosclerosis by cod-liver oil in a hyperlipidemic swine model. N. Engl. J. Med. 315, 841-846.
WILLIS, A. L. (1981). Nutritional and pharmacological factors in eicosanoid biology. Nutr. Rev. 39, 289-301.
ZHU, B.-Q., SMITH, D. L., SIEVERS, R. E., ISENBERG, W. M., and PARMLY, W. W. (1988). Inhibition of atherosclerosis by fish oil in cholesterol-fed rabbits. J. Am. Coil. Cardiol. 12, 1073-1078.




















