
Augmented Vasoconstriction and Thromboxane Formation by 15-F2t-Isoprostane (8-Iso-Prostaglandin F2a)...
10
Augmented Vasoconstriction and Thromboxane Formation by 15-F 2t -Isoprostane (8-Iso-Prostaglandin F 2a ) in Immature Pig Periventricular Brain Microvessels Xin Hou, MD, PhD; Fernand Gobeil, Jr, PhD; Krishna Peri, PhD; Giovanna Speranza, BSc; Anne Marilise Marrache, BSc; Pierre Lachapelle, PhD; Jackson Roberts II, MD; Daya R. Varma, MD, PhD; Sylvain Chemtob, MD, PhD Background and Purpose—Oxidant stress, especially in the premature, plays a major role in the pathogenesis of hypoxic-ischemic encephalopathies mostly manifested in the periventricular region. We studied the vasomotor mode of actions of the peroxidation product 15-F 2t -isoprostane (15-F 2t -IsoP) (8-iso-prostaglandin F 2a ) on periventricular region during development. Methods—Effects of 15-F 2t -IsoP on periventricular microvessels of fetal, newborn, and juvenile pigs were studied by video imaging and digital analysis techniques. Thromboxane formation and intracellular Ca 21 were measured by radioimmu- noassay and by using the fluorescent indicator fura 2-AM. Results—15-F 2t -IsoP–mediated constriction of periventricular microvessels decreased as a function of age such that in the fetus it was ’2.5-fold greater than in juvenile pigs. 15-F 2t -IsoP evoked more thromboxane formation in the fetus than in the newborn, which was greater than that in the juvenile periventricular region; this was associated with immunoreactive thromboxane A 2 (TXA 2 ) synthase expression in the fetus that was greater than that in newborn pigs, which was greater than that in juvenile pigs. 15-F 2t -IsoP–induced vasoconstriction was markedly inhibited by TXA 2 synthase and receptor blockers (CGS12970 and L670596). Vasoconstrictor effects of the TXA 2 mimetic U46619 on fetal, neonatal, and juvenile periventricular microvessels did not differ. 15-F 2t -IsoP increased TXA 2 synthesis by activating Ca 21 influx through non–voltage-gated channels in endothelial cells (SK&F96365 sensitive) and N-type voltage-gated channels (v-conotoxin sensitive) in astrocytes; smooth muscle cells were not responsive to 15-F 2t -IsoP but generated Ca 21 transients to U46619 via L-type voltage-sensitive channels. Conclusions—15-F 2t -IsoP causes periventricular brain region vasoconstriction in the fetus that is greater than that in the newborn, which in turn is greater than that in the juvenile due to greater TXA 2 formation generated through distinct stimulatory pathways, including from endothelial and astroglial cells. The resulting hemodynamic compromise may contribute to the increased vulnerability of the periventricular brain areas to oxidant stress–induced injury in immature subjects. (Stroke. 2000;31:516-525.) Key Words: fetus n ischemia n newborn n peroxidation n prostaglandins O xidant stress plays a major role in the pathogenesis of various disorders such as hypoxic-ischemic encephalop- athies, 1–3 including periventricular leukomalacia in premature subjects. 2,4–6 Free radicals can alter brain hemodynamics by causing vasoconstriction 7,8 and increase thromboxane A 2 (TXA 2 ). 9–13 Although TXA 2 has been implicated in peroxidation-induced vasoconstriction, 9 –12 the mechanisms of TXA 2 production by brain vasculature during oxidant stresses remain unknown. A series of prostaglandin F 2 –like compounds produced nonenzymatically by free radical–induced peroxidation of See Editorial Comment, page 525 arachidonic acid independent of cyclooxygenase have been shown to be produced in vivo and their formation to increase markedly during oxidant injury. 13–19 15-F 2t -Isoprostane (15- F 2t -IsoP) (8-iso-prostaglandin F 2a ) 20 is one of the abundantly generated isoprostanes in vivo; it is a potent renal, coronary, pulmonary, retinal, and cerebral vasoconstrictor. 7,18,21–24 Al- though the vasomotor effects of 15-F 2t -IsoP are largely inhibited by TXA 2 receptor blockers, 7,22–24 15-F 2t -IsoP does not seem to bind directly with the TXA 2 receptor. 25–27 We Received August 27, 1999; final revision received October 28, 1999; accepted November 1, 1999. From the Centre de Recherche de l’Ho ˆpital Sainte-Justine, Department of Pediatrics and Pharmacology, Universite ´ de Montre ´al, Montre ´al, Que ´bec, Canada (X.H., F.G., K.P., G.S., A.M.M., S.C.); Departments of Pharmacology and Therapeutics (G.S., A.M.M., D.R.V., S.C.) and Ophthalmology (P.L.), McGill University, Montre ´al, Que ´bec, Canada; and Departments of Pharmacology and Medicine, Vanderbilt University, Nashville, Tenn (J.R.). Correspondence to Sylvain Chemtob, MD, PhD, Research Center, Ho ˆpital Sainte-Justine, Department of Pediatrics and Pharmacology, 3175 Co ˆte Sainte-Catherine, Montre ´al, Que ´bec, Canada, H3T 1C5. E-mail [email protected] © 2000 American Heart Association, Inc. Stroke is available at http://www.strokeaha.org 516 by guest on May 25, 2015 http://stroke.ahajournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Augmented Vasoconstriction and Thromboxane Formation by 15-F2t-Isoprostane (8-Iso-Prostaglandin F2a)...

Augmented Vasoconstriction and Thromboxane Formationby 15-F2t-Isoprostane (8-Iso-Prostaglandin F2a) in Immature
Pig Periventricular Brain MicrovesselsXin Hou, MD, PhD; Fernand Gobeil, Jr, PhD; Krishna Peri, PhD; Giovanna Speranza, BSc;
Anne Marilise Marrache, BSc; Pierre Lachapelle, PhD; Jackson Roberts II, MD;Daya R. Varma, MD, PhD; Sylvain Chemtob, MD, PhD
Background and Purpose—Oxidant stress, especially in the premature, plays a major role in the pathogenesis ofhypoxic-ischemic encephalopathies mostly manifested in the periventricular region. We studied the vasomotor mode ofactions of the peroxidation product 15-F2t-isoprostane (15-F2t-IsoP) (8-iso-prostaglandin F2a) on periventricular regionduring development.
Methods—Effects of 15-F2t-IsoP on periventricular microvessels of fetal, newborn, and juvenile pigs were studied by videoimaging and digital analysis techniques. Thromboxane formation and intracellular Ca21 were measured by radioimmu-noassay and by using the fluorescent indicator fura 2-AM.
Results—15-F2t-IsoP–mediated constriction of periventricular microvessels decreased as a function of age such that in thefetus it was'2.5-fold greater than in juvenile pigs. 15-F2t-IsoP evoked more thromboxane formation in the fetus thanin the newborn, which was greater than that in the juvenile periventricular region; this was associated withimmunoreactive thromboxane A2 (TXA2) synthase expression in the fetus that was greater than that in newborn pigs,which was greater than that in juvenile pigs. 15-F2t-IsoP–induced vasoconstriction was markedly inhibited by TXA2
synthase and receptor blockers (CGS12970 and L670596). Vasoconstrictor effects of the TXA2 mimetic U46619 onfetal, neonatal, and juvenile periventricular microvessels did not differ. 15-F2t-IsoP increased TXA2 synthesis byactivating Ca21 influx through non–voltage-gated channels in endothelial cells (SK&F96365 sensitive) and N-typevoltage-gated channels (v-conotoxin sensitive) in astrocytes; smooth muscle cells were not responsive to 15-F2t-IsoP butgenerated Ca21 transients to U46619 via L-type voltage-sensitive channels.
Conclusions—15-F2t-IsoP causes periventricular brain region vasoconstriction in the fetus that is greater than that in thenewborn, which in turn is greater than that in the juvenile due to greater TXA2 formation generated through distinctstimulatory pathways, including from endothelial and astroglial cells. The resulting hemodynamic compromise maycontribute to the increased vulnerability of the periventricular brain areas to oxidant stress–induced injury in immaturesubjects.(Stroke. 2000;31:516-525.)
Key Words: fetusn ischemian newbornn peroxidationn prostaglandins
Oxidant stress plays a major role in the pathogenesis ofvarious disorders such as hypoxic-ischemic encephalop-
athies,1–3 including periventricular leukomalacia in prematuresubjects.2,4–6 Free radicals can alter brain hemodynamics bycausing vasoconstriction7,8 and increase thromboxane A2
(TXA 2).9 –13 Although TXA2 has been implicated inperoxidation-induced vasoconstriction,9–12 the mechanismsof TXA2 production by brain vasculature during oxidantstresses remain unknown.
A series of prostaglandin F2–like compounds producednonenzymatically by free radical–induced peroxidation of
See Editorial Comment, page 525
arachidonic acid independent of cyclooxygenase have beenshown to be produced in vivo and their formation to increasemarkedly during oxidant injury.13–19 15-F2t-Isoprostane (15-F2t-IsoP) (8-iso-prostaglandin F2a)20 is one of the abundantlygenerated isoprostanes in vivo; it is a potent renal, coronary,pulmonary, retinal, and cerebral vasoconstrictor.7,18,21–24Al-though the vasomotor effects of 15-F2t-IsoP are largelyinhibited by TXA2 receptor blockers,7,22–24 15-F2t-IsoP doesnot seem to bind directly with the TXA2 receptor.25–27 We
Received August 27, 1999; final revision received October 28, 1999; accepted November 1, 1999.From the Centre de Recherche de l’Hopital Sainte-Justine, Department of Pediatrics and Pharmacology, Universite de Montreal, Montreal, Quebec,
Canada (X.H., F.G., K.P., G.S., A.M.M., S.C.); Departments of Pharmacology and Therapeutics (G.S., A.M.M., D.R.V., S.C.) and Ophthalmology (P.L.),McGill University, Montreal, Quebec, Canada; and Departments of Pharmacology and Medicine, Vanderbilt University, Nashville, Tenn (J.R.).
Correspondence to Sylvain Chemtob, MD, PhD, Research Center, Hopital Sainte-Justine, Department of Pediatrics and Pharmacology, 3175 CoˆteSainte-Catherine, Montreal, Quebec, Canada, H3T 1C5. E-mail [email protected]
© 2000 American Heart Association, Inc.
Stroke is available at http://www.strokeaha.org
516 by guest on May 25, 2015http://stroke.ahajournals.org/Downloaded from

recently reported that 15-F2t-IsoP causes marked constrictionof retinal vessels of piglets by stimulating TXA2 synthesis,21
but such cyclooxygenase-dependent action of this isoprostanehas not been uniformly observed in other vascular beds.28
Moreover, the direct effects of 15-F2t-IsoP on brain intrapa-renchymal vasculature, which is intimately involved in thegenesis of encephalopathies, are not known.
Because oxidant stress–induced encephalopathies aremostly localized in the periventricular region in immaturecompared with older subjects,2 we postulated that the con-strictor effects of the product of peroxidation 15-F2t-IsoP onmicrovessels of the periventricular brain region are morepronounced in immature than in older subjects. In this processwe evaluated the effects of 15-F2t-IsoP on these microvesselsas well as the role and cellular sources of TXA2 in thesevascular responses. It was found that 15-F2t-IsoP causedperiventricular microvessel constriction in fetal more than innewborn more than in juvenile animals by stimulating in-creased formation of TXA2 through distinct pathways inendothelial and astroglial cells from brains of youngersubjects.
Materials and MethodsTissue PreparationAnimals were used according to a protocol of the Animal CareCommittee of the Hopital Sainte-Justine in accordance with theprinciples of the Guide for the Care and Use of ExperimentalAnimals of the Canadian Council on Animal Care. Fetal pigs (78 to90 days’ gestation [term, 114 days]) were obtained from an abattoir(St-Helene, Quebec, Canada) immediately after exsanguination ofsows. Newborn (1 to 3 days old) and juvenile pigs (4 to 5 weeks old)were obtained from Fermes Menard Inc (L’Ange-Gardien, Quebec,Canada). Animals were anesthetized with halothane ('2.5% to 5%),and india ink (1.5 mL/kg) was injected intracardially to facilitatevisualization of the cerebral microvessels; animals were killed withpentobarbital (120 mg/kg), and brains were removed and placedimmediately in ice-cold Krebs’ buffer (pH 7.4) of the followingcomposition (mmol/L): NaCl 120, KCl 4.5, CaCl2 2.5, MgSO4 1.0,NaHCO3 27, KH2PO4 1.0, and glucose 10; 1.5 U/mL heparin wasadded to the buffer. For biochemical measurements, tissues werefrozen in liquid N2 and stored at280°C.
Vasomotor Response of BrainPeriventricular MicrovesselsSlices of brain (1 mm thick) exposing the periventricular brain regionwere prepared as previously described29,30 to study relatively undis-turbed penetrating microvessels (30 to 50mm), reported to contributesignificantly in the control of cerebral vascular resistance.31 Thebrain slices were pinned securely to a wax base of a 20-mL bathcontaining Krebs’ buffer (pH 7.4) equilibrated with 21% O2, 5%CO2, and 74% N2 and maintained at 37°C. The preparations werewashed 2 to 3 times with fresh buffer and allowed to equilibrate for45 minutes before the start of the experiment.
Cerebral microvessels were visualized and recorded with a videocamera (model CCD72, MTI) mounted on a dissecting microscope(model M-400, Nikon) as previously reported.29,30Vascular diameterwas measured with a digital image analyzer (Sigma Scan software,Jandel Scientific) and repeated 3 times with a variability of,1%;pilot experiments indicated that the inert india ink did not modifyvascular responses to constrictors (eg, U46619 and phenylephrine)and relaxants (eg, carbaprostacyclin and sodium nitroprusside).Vascular diameter was recorded before and after topical applicationof increasing concentration of test agents (15-F2t-IsoP, thromboxanemimetic U46619, and prostaglandin F2a [PGF2a]) in the presence andabsence of 30 minutes’ pretreatment with the following agents at
known effective concentrations9,21,32: nonselective cyclooxygenaseinhibitor ibuprofen (10mmol/L); nonselective phospholipase A2
blocker oleoyloxyethyl phosphocholine33 (OPPC) (50 mmol/L);TXA2 synthase inhibitor CGS12970 (1mmol/L); TXA2 receptorantagonist L670596 (0.1mmol/L); endothelin ETA receptor blockerBQ-123 (1 mmol/L); non–voltage-dependent Ca21 entry and
Figure 1. Concentration-vasoconstrictor response to 15-F2t-IsoP, PGF2a, and U46619 on brain periventricular microvesselsof fetal, neonatal, and juvenile pigs. Constriction is the percentreduction in vascular diameter from basal values, which were36.563.2, 34.562.6, and 35.162.8 mm, respectively, for fetus,newborn, and juvenile, as previously reported.27,28 Effects ofagents were studied in situ on brain slices, as described inMethods. Data are mean6SEM of 5 to 6 separate experiments.*P,0.05 compared with fetus and newborn; †P,0.05 comparedwith fetus and juvenile (2-way ANOVA and comparison amongmeans test).
Hou et al 15-F2t-Isoprostane Effects on Brain Microvessels 517
by guest on May 25, 2015http://stroke.ahajournals.org/Downloaded from
receptor-mediated Ca21 channel blocker SK&F9636534 (20mmol/L);L-type voltage-gated Ca21 channel blocker nifedipine (5mmol/L);and N-type voltage-gated Ca21 channel blockerv-conotoxin35
(10 mmol/L). Focus was placed on receptor-operated and N- as wellas L-type voltage-gated Ca21 channels since endothelial cells are notexcitable and are mostly devoid of voltage-gated Ca21 channels,36
whereas astrocytes contain voltage-gated Ca21 channels primarily ofthe N- and L-types.37,38
Microvascular Endothelial and Smooth MuscleCell CultureMicrovessels from fetal brain were prepared as previously de-scribed.29,30,39Microvessels were suspended in selective endothelialor smooth muscle growth media (Clonetics). Confluent individualendothelial and smooth muscle cells were trypsinized, centrifuged,reseeded in culture flasks, and subcultured; cell viability was verifiedby trypan blue exclusion and was.90%. Endothelial cells wereidentified by their cobblestone morphology at confluence, positivereactivity to factor VIII antibody, and negative reactivity to smoothmuscle–specific actin and glial fibrillary acidic protein (GFAP)antibodies (Dako). Smooth muscle cells were recognized by theirspindle-shaped appearance, positive reactivity to smooth muscle–specific actin antibody, and negative reactivity to factor VIII andGFAP antibodies. Confluent cultures of endothelial and smoothmuscle cells of passages 5 to 15 were used for experiments.
Astroglial Cell CultureAstrocytes were cultured from fetal brain.40 Brains were collected inHam’s F-12 medium containing penicillin (50 U/mL) and strepto-mycin (50 mg/mL). Brain homogenate was sequentially filteredthrough 230- and 150-mm nylon mesh, and the filtrate was centri-fuged at 1000gfor 7 minutes and resuspended in DMEM with 10%fetal calf serum and incubated in air and 5% CO2 at 37°C. Mixedglial cultures were grown for 9 to 11 days, and loosely attachedmacrophages were removed with a rotary shaker at 225 rpm for 3hours. Media were changed, and culture was equilibrated for 6 hoursand shaken thereafter for 18 hours to remove oligodendrocyteprogenitors. Cultures were trypsinized and reseeded. Purity ofastrocytes was assessed by immunoreactivity to GFAP, which was.95%.
Thromboxane AssayThe effects of 15-F2t-IsoP on thromboxane formation were studied infetus, newborn, and juvenile pig brain slices stimulated (15 minutes)with 15-F2t-IsoP (1mmol/L); the reaction was stopped with liquid N2.Thromboxane B2 (stable TXA2 metabolite) was determined on thehomogenized tissue by radioimmunoassay, as previously de-scribed.9,21 TXB2 concentration was also measured in the culturemedia of astroglial, endothelial, and smooth muscle cells stimulatedfor 15 minutes with 15-F2t-IsoP (1 mmol/L) in the absence orpresence of ibuprofen (10mmol/L), CGS12970 (1mmol/L),
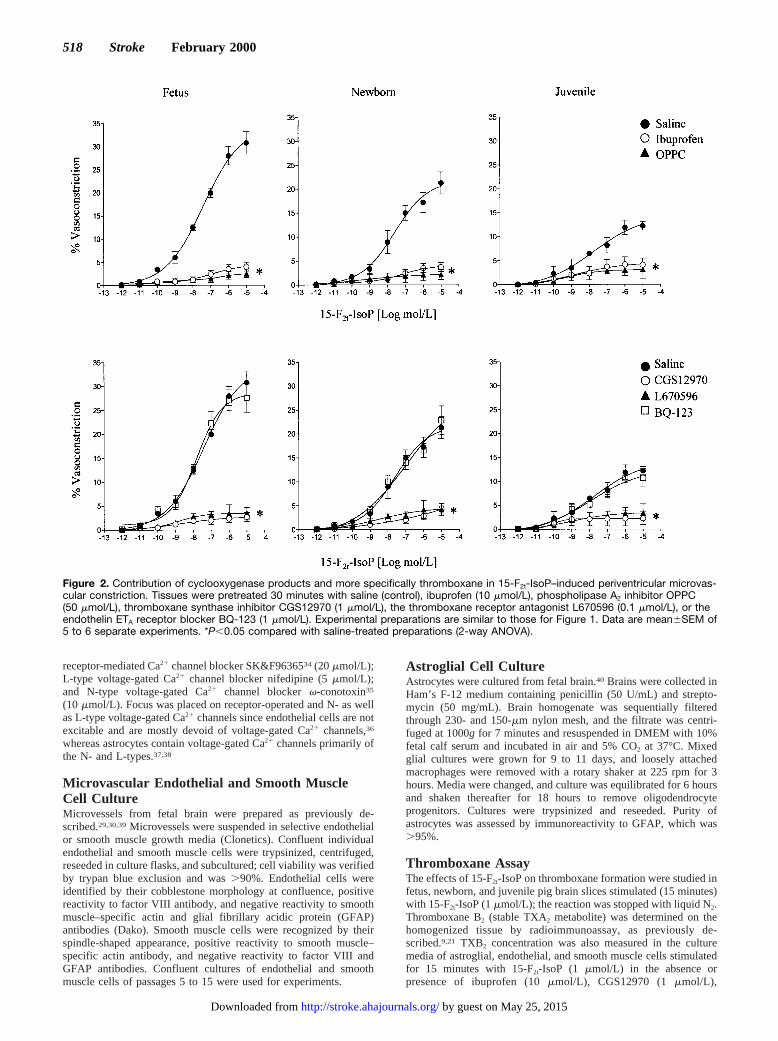
Figure 2. Contribution of cyclooxygenase products and more specifically thromboxane in 15-F2t-IsoP–induced periventricular microvas-cular constriction. Tissues were pretreated 30 minutes with saline (control), ibuprofen (10 mmol/L), phospholipase A2 inhibitor OPPC(50 mmol/L), thromboxane synthase inhibitor CGS12970 (1 mmol/L), the thromboxane receptor antagonist L670596 (0.1 mmol/L), or theendothelin ETA receptor blocker BQ-123 (1 mmol/L). Experimental preparations are similar to those for Figure 1. Data are mean6SEM of5 to 6 separate experiments. *P,0.05 compared with saline-treated preparations (2-way ANOVA).
518 Stroke February 2000
by guest on May 25, 2015http://stroke.ahajournals.org/Downloaded from
SK&F96365 (20 mmol/L), nifedipine (5 mmol/L), v-conotoxin(10 mmol/L), or EGTA (100mmol/L).
Ca21 SignalsIntracellular Ca21 ([Ca21]i) signals were measured with the fluores-cent indicator fura 2-AM, as we reported.21 For this purpose,confluent endothelial, smooth muscle, and astroglial cells of fetalpigs were trypsinized in a solution containing 0.05% trypsin and0.02% EDTA for 2 minutes, then 5 mL of HBSS was added. Cellswere centrifuged at 250gfor 10 minutes and resuspended in a buffercontaining (in mmol/L) HEPES 20,D-glucose 10, KCl 4.6, NaCl118, and CaCl2 0.5, as well as 1% fetal bovine serum. Cell viabilitywas determined by trypan blue exclusion and was.90%. Fura2-AM (2 mmol/L) and 0.2% Pluronic F-127 were added to cellsuspensions, which were incubated at 37°C for 30 minutes. Theloaded cells were then washed twice and resuspended in HBSS withCa21 (2.5 mmol/L) and 1% fetal bovine serum with or withoutpretreatment for 15 minutes with SK&F96365 (20mmol/L), nifedi-pine (5mmol/L), v-conotoxin (10mmol/L), or EGTA (100mmol/L)followed by stimulation with 15-F2t-IsoP (1 mmol/L) or U46619(1 mmol/L). The [Ca21]i was determined in 2 mL of fura 2–loadedcell suspension ('23106 cells per milliliter) continuously stirred andmeasured with a spectrofluorometer (model LS 50, Perkin-Elmer) byusing excitation wavelengths of 340 and 380 nm and emission at 510nm. Calibration of the fluorescent signal was determined on 2 mL of
cell suspension by sequential addition of 10 mmol/L ionomycin toobtain the maximal fluorescence ratio (Rmax) and to 5 mmol/L EGTAplus 0.2% Triton X-100 to obtain the minimal fluorescence ratio(Rmin). Autofluorescence was determined by measuring fluorescencefrom nonloaded cells and subtracting it from the fluorescenceproduced by fura 2-AM–loaded cells to calculate the fluorescenceratio R corresponding to the values produced at 340 and 380 nm. The[Ca21]i was calculated from the equation of Grynkiewicz et al41:[Ca21]i5Kd[(R2Rmin)/(Rmax2R)](Sf2/Sb2), whereKd (224 nmol/L) isthe effective dissociation constant of fura 2-AM/Ca21 complex andSf2/Sb2 is the ratio of fluorescence intensity at 380-nm wavelength inthe presence of EGTA to that in the presence of Triton X-100.
Immunoblotting of Thromboxane SynthaseTXA2 synthase immunoreactivity on brain was determined as wepreviously described for other membrane-bound enzymes.42 Briefly,homogenized tissues of periventricular regions of all age groupsstudied were preabsorbed with 50 mL of immunoprecipitin for 30minutes and then centrifuged at 12 000gfor 10 minutes to removeimmunoprecipitin. The supernatant was incubated with porcineTXA2 synthase–specific polyclonal antibodies (Cayman Chemicals)for 1.5 hours, and immune complexes were collected by incubationwith 50 mL immunoprecipitin for 30 minutes followed by centrifu-gation. Immune precipitates were denatured in SDS buffer andcentrifuged at 12 000gfor 15 minutes to remove the immunopreci-
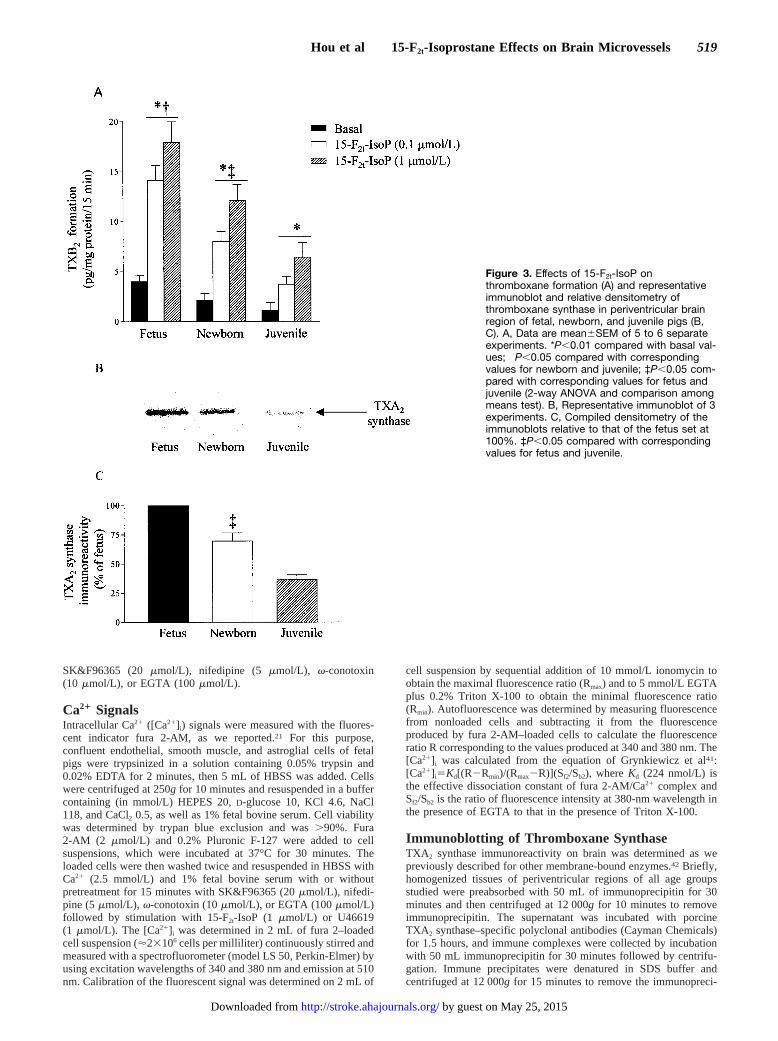
Figure 3. Effects of 15-F2t-IsoP onthromboxane formation (A) and representativeimmunoblot and relative densitometry ofthromboxane synthase in periventricular brainregion of fetal, newborn, and juvenile pigs (B,C). A, Data are mean6SEM of 5 to 6 separateexperiments. *P,0.01 compared with basal val-ues; †P,0.05 compared with correspondingvalues for newborn and juvenile; ‡P,0.05 com-pared with corresponding values for fetus andjuvenile (2-way ANOVA and comparison amongmeans test). B, Representative immunoblot of 3experiments. C, Compiled densitometry of theimmunoblots relative to that of the fetus set at100%. ‡P,0.05 compared with correspondingvalues for fetus and juvenile.
Hou et al 15-F2t-Isoprostane Effects on Brain Microvessels 519
by guest on May 25, 2015http://stroke.ahajournals.org/Downloaded from

pitin; samples were loaded on SDS-polyacrylamide gels. Proteinswere electrophoretically transferred to nitrocellulose membranes andincubated with TXA2 synthase–specific antibodies. After they werewashed, membranes were incubated with horseradish peroxidase–conjugated anti-rabbit IgG antibody followed by several washes.Immunoreactive bands were visualized by enhanced chemilumines-cence (Amersham Canada), as recommended by the supplier, andanalyzed by densitometry.
ChemicalsL670596 and CGS12970 were generous gifts from Merck-Frosst(Pointe-Claire, Quebec, Canada) and Ciba-Geigy (Summit, NJ),respectively. The following products were purchased: 15-F2t-IsoP(.99% pure), U46619, and TXA2 synthase polyclonal antibodies(Cayman Chemicals); ATP, EDTA, EGTA, OPPC, ibuprofen, iono-mycin, nifedipine, Triton X-100,v-conotoxin, and Tris-HCl (SigmaChemical); SK&F96365 (BioMol); fura 2-AM (Calbiochem); TXB2
radioimmunoassay kits (Amersham); endothelial cell, smooth mus-cle cell, and astrocyte growth medium (Clonetics); factor VIIIantibody, smooth muscle–specific actin antibody, and GFAP anti-body (Dako); and all other chemicals (Fisher Scientific).
Statistical AnalysisAll results are expressed as mean6SEM. Results were analyzed withStudent’st test and 2-way ANOVA, factoring for concentrations andage or treatments. Post-ANOVA comparisons among means wereperformed with the Turkey-Kramer method. Statistical significancewas set atP#0.05.
ResultsEffects of 15-F2t-IsoP on BrainPeriventricular Microvessels15-F2t-IsoP caused concentration-dependent constriction ofperiventricular microvessels from all animals (Figure 1). TheEC50 values of 15-F2t-IsoP on fetal, newborn, and juvenile pigmicrovessels were comparable: 38.764.8, 24.863.6, and17.663.1 nmol/L, respectively. On the other hand, maximalconstriction to 15-F2t-IsoP in fetus (30.861.7%) was greater
than in newborn (21.362.3%), which was greater than injuvenile pig microvessels (12.262.1%). In contrast, PGF2a
was more effective on juvenile than on fetal and neonatal pigmicrovessels (P,0.05).
Relationship Between Vasoconstrictor Effects of15-F2t-IsoP and TXA2
The vasoconstrictor effects of 15-F2t-IsoP on microvesselsfrom all 3 groups of animals were almost fully inhibited byibuprofen, phospholipase A2 inhibitor OPPC, TXA2 synthaseinhibitor CGS12970, and TXA2 receptor antagonist L670596(Figure 2); the endothelin ETA receptor blocker BQ-123 wasineffective. PGF2a-induced constriction was unaffected by allthe above inhibitors. Correspondingly, 15-F2t-IsoP increasedTXB2 generation in periventricular tissue of fetal, newborn,and juvenile pigs; basal and 15-F2t-IsoP–induced TXA2 pro-duction exhibited an age-dependent profile that was greater infetal than in newborn than in juvenile microvessels (Figure3A). A similar developmental pattern of expression of immu-noreactive TXA2 synthase was observed (Figure 3B and 3C).On the other hand, vasoconstriction to the TXA2 mimeticU46619 did not differ between the 3 age groups (Figure 1C).Hence, developmental changes in 15-F2t-IsoP–induced con-striction seem largely dependent on ontogenic differences inTXA 2 formation, which is highest in the fetus.
Effects of 15-F2t-IsoP on Thromboxane Formationby Cultured Cerebral Endothelial, Smooth Muscle,and Astroglial CellsTo determine the potential source of increased thromboxanein response to 15-F2t-IsoP in immature animals (a principalinterest of this study), thromboxane formation was measuredon primary cultures of fetal cerebrovascular cells, namely,endothelial and smooth muscle cells, as well as astroglial
Figure 4. Effects of 15-F2t-IsoP on TXB2 production by cultured endothelial (A) and astroglial cells (A) from fetal pig brain. Cells werepretreated 20 minutes with saline or one of the following: ibuprofen (10 mmol/L), CGS12970 (1 mmol/L), nifedipine (5 mmol/L),SK&F96365 (20 mmol/L), v-conotoxin (10 mmol/L), and EGTA (100 mmol/L). Data are mean6SEM of 5 to 6 separate experiments.*P,0.01 compared with all other values without asterisk.
520 Stroke February 2000
by guest on May 25, 2015http://stroke.ahajournals.org/Downloaded from

cells, which are not only perivascular but also are the mostabundant cell type in brain; experiments were conducted onlyon cells from fetal animals since fetus exhibited responses to15-F2t-IsoP and immunoreactive expression of TXA2 synthasethat were similar to those of newborn (Figures 1 to 3).15-F2t-IsoP stimulated formation of TXB2 by endothelial andastroglial cells (Figure 4); effects of 15-F2t-IsoP on TXA2
generation by smooth muscle cells were negligible (,2pg/106 cells per 15 minutes). The stimulatory effects of15-F2t-IsoP on TXB2 formation in endothelial and astroglialcells were diminished by inhibitors of TXA2 synthesis (ibu-profen and CGS12970).
Because prostanoid formation is Ca21 dependent throughphospholipase A2 requirements, we attempted to identify thetype of channel involved in 15-F2t-IsoP–induced TXA2 genera-tion. We focused on receptor-operated and N- as well as L-typevoltage-gated Ca21 channels since endothelial cells are notexcitable and are mostly devoid of voltage-gated Ca21 chan-
nels,36 whereas astrocytes contain voltage-gated channels, pri-marily of the N- and L-types.37,38,43 The putative receptor-operated Ca21 channel blocker SK&F9636534 selectivelyreduced TXB2 formation in endothelial cells, and the selectiveN-voltage–gated Ca21 channel blockerv-conotoxin35 caused asimilar effect only in astrocytes (Figure 4); Ca21 chelator EGTAwas effective in both cells. Therefore, TXB2 formation inducedby 15-F2t-IsoP is dependent on extracellular Ca21, which seemsto influx through activation of distinct Ca21 channels in endo-thelial and astroglial cells.
Effects of 15-F2t-IsoP on Ca21 TransientsThe effects of 15-F2t-IsoP on Ca21 transients (using fura2-AM) corroborated data on TXB2 formation. 15-F2t-IsoPinduced an increase in Ca21 signals in endothelial cells, whichwas significantly reduced by SK&F96365 and EGTA but notby nifedipine orv-conotoxin (Figure 5A and 5B). This effectof 15-F2t-IsoP on astrocytes was not significantly affected by
Figure 5. Intracellular calcium transients[Ca21]i in fetal pig brain endothelial (A, B)and astroglial cells (C, D) in response to15-F2t-IsoP, and in smooth muscle cells(E, F) in response to U46619. Cells werepretreated 20 minutes with saline (con-trol), nifedipine (5 mmol/L), SK&F96365(20 mmol/L), v-conotoxin (10 mmol/L),and EGTA (100 mmol/L). Intracellular cal-cium transients were measured with fura2-AM, as described in Methods. Arrowsin B, D, F point to moment of addition of15-F2t-IsoP (B, D) and U46619 (F). Valuesare mean6SEM of 4 to 5 separateexperiments. *P,0.05 compared with allother values without asterisk.
Hou et al 15-F2t-Isoprostane Effects on Brain Microvessels 521
by guest on May 25, 2015http://stroke.ahajournals.org/Downloaded from
nifedipine and SK&F96365 but was markedly inhibited byv-conotoxin and EGTA (Figure 5C and 5D). In contrast,15-F2t-IsoP did not affect Ca21 transients in smooth musclecells (Figure 5E and 5F). On the other hand, TXA2 mimeticU46619 induced Ca21 transients in smooth muscle cells,which was inhibited by nifedipine but not by SK&F96365(Figure 5E and 5F).
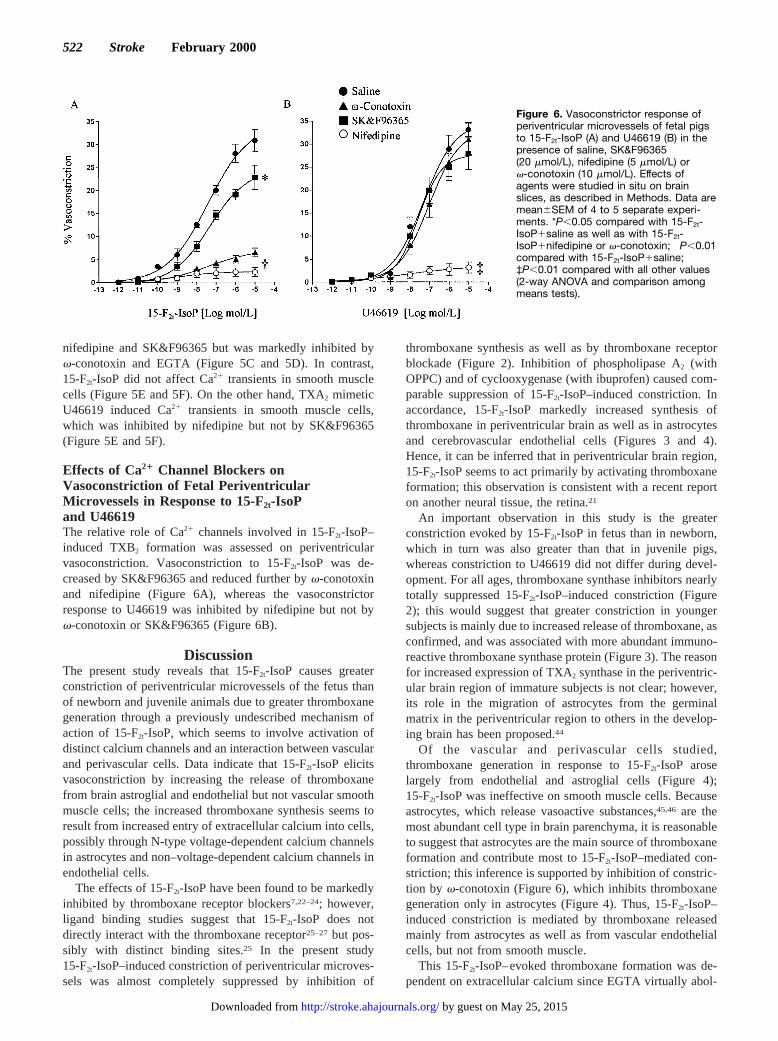
Effects of Ca21 Channel Blockers onVasoconstriction of Fetal PeriventricularMicrovessels in Response to 15-F2t-IsoPand U46619The relative role of Ca21 channels involved in 15-F2t-IsoP–induced TXB2 formation was assessed on periventricularvasoconstriction. Vasoconstriction to 15-F2t-IsoP was de-creased by SK&F96365 and reduced further byv-conotoxinand nifedipine (Figure 6A), whereas the vasoconstrictorresponse to U46619 was inhibited by nifedipine but not byv-conotoxin or SK&F96365 (Figure 6B).
DiscussionThe present study reveals that 15-F2t-IsoP causes greaterconstriction of periventricular microvessels of the fetus thanof newborn and juvenile animals due to greater thromboxanegeneration through a previously undescribed mechanism ofaction of 15-F2t-IsoP, which seems to involve activation ofdistinct calcium channels and an interaction between vascularand perivascular cells. Data indicate that 15-F2t-IsoP elicitsvasoconstriction by increasing the release of thromboxanefrom brain astroglial and endothelial but not vascular smoothmuscle cells; the increased thromboxane synthesis seems toresult from increased entry of extracellular calcium into cells,possibly through N-type voltage-dependent calcium channelsin astrocytes and non–voltage-dependent calcium channels inendothelial cells.
The effects of 15-F2t-IsoP have been found to be markedlyinhibited by thromboxane receptor blockers7,22–24; however,ligand binding studies suggest that 15-F2t-IsoP does notdirectly interact with the thromboxane receptor25–27 but pos-sibly with distinct binding sites.25 In the present study15-F2t-IsoP–induced constriction of periventricular microves-sels was almost completely suppressed by inhibition of
thromboxane synthesis as well as by thromboxane receptorblockade (Figure 2). Inhibition of phospholipase A2 (withOPPC) and of cyclooxygenase (with ibuprofen) caused com-parable suppression of 15-F2t-IsoP–induced constriction. Inaccordance, 15-F2t-IsoP markedly increased synthesis ofthromboxane in periventricular brain as well as in astrocytesand cerebrovascular endothelial cells (Figures 3 and 4).Hence, it can be inferred that in periventricular brain region,15-F2t-IsoP seems to act primarily by activating thromboxaneformation; this observation is consistent with a recent reporton another neural tissue, the retina.21
An important observation in this study is the greaterconstriction evoked by 15-F2t-IsoP in fetus than in newborn,which in turn was also greater than that in juvenile pigs,whereas constriction to U46619 did not differ during devel-opment. For all ages, thromboxane synthase inhibitors nearlytotally suppressed 15-F2t-IsoP–induced constriction (Figure2); this would suggest that greater constriction in youngersubjects is mainly due to increased release of thromboxane, asconfirmed, and was associated with more abundant immuno-reactive thromboxane synthase protein (Figure 3). The reasonfor increased expression of TXA2 synthase in the periventric-ular brain region of immature subjects is not clear; however,its role in the migration of astrocytes from the germinalmatrix in the periventricular region to others in the develop-ing brain has been proposed.44
Of the vascular and perivascular cells studied,thromboxane generation in response to 15-F2t-IsoP aroselargely from endothelial and astroglial cells (Figure 4);15-F2t-IsoP was ineffective on smooth muscle cells. Becauseastrocytes, which release vasoactive substances,45,46 are themost abundant cell type in brain parenchyma, it is reasonableto suggest that astrocytes are the main source of thromboxaneformation and contribute most to 15-F2t-IsoP–mediated con-striction; this inference is supported by inhibition of constric-tion by v-conotoxin (Figure 6), which inhibits thromboxanegeneration only in astrocytes (Figure 4). Thus, 15-F2t-IsoP–induced constriction is mediated by thromboxane releasedmainly from astrocytes as well as from vascular endothelialcells, but not from smooth muscle.
This 15-F2t-IsoP–evoked thromboxane formation was de-pendent on extracellular calcium since EGTA virtually abol-
Figure 6. Vasoconstrictor response ofperiventricular microvessels of fetal pigsto 15-F2t-IsoP (A) and U46619 (B) in thepresence of saline, SK&F96365(20 mmol/L), nifedipine (5 mmol/L) orv-conotoxin (10 mmol/L). Effects ofagents were studied in situ on brainslices, as described in Methods. Data aremean6SEM of 4 to 5 separate experi-ments. *P,0.05 compared with 15-F2t-IsoP1saline as well as with 15-F2t-IsoP1nifedipine or v-conotoxin; †P,0.01compared with 15-F2t-IsoP1saline;‡P,0.01 compared with all other values(2-way ANOVA and comparison amongmeans tests).
522 Stroke February 2000
by guest on May 25, 2015http://stroke.ahajournals.org/Downloaded from

ished it (Figure 4). Astrocytes contain various calciumchannels.37,38,43The increase in thromboxane formation andcalcium signals in astrocytes was inhibited by the selectiveN-type voltage-gated calcium channel blockerv-conotoxin35
but not by L-type voltage-gated channel blocker nifedipineand putative inhibitor of non–voltage-gated calcium channelsSK&F96365.34 This finding would suggest that in astrocytes,15-F2t-IsoP stimulates thromboxane formation by enhancingentry of calcium mainly via N-type voltage-gated calciumchannels. In contrast in endothelial cells, SK&F96365 wasfound to inhibit 15-F2t-IsoP–induced thromboxane formationand the increase in intracellular calcium (Figures 4 and 5).SK&F96365 has been reported to inhibit receptor-mediatedcalcium entry at#30 mmol/L concentration, whereas atconcentrations.100 mmol/L, SK&F-96365 also blocksvoltage-gated calcium channels.34 In this study SK&F96365was used at 20mmol/L. Hence, influx of calcium in cerebro-vascular endothelial cells in response to 15-F2t-IsoP is possi-bly via receptor-operated channels; similar observations havebeen made in retinal endothelial cells.21 Collectively, datasuggest that 15-F2t-IsoP increases influx of calcium throughdistinct channels in astrocytes and endothelial cells, and thisin turn leads to activation of phospholipase A2 and metabo-lism of arachidonic acid into thromboxane. The involvementof separate pathways (channel activation) resulting inthromboxane formation would be consistent with existence ofdistinct receptor sites for 15-F2t-IsoP, which remain to becharacterized.
Taken together, these data suggest that 15-F2t-IsoP exertslittle direct effects on cerebrovascular smooth muscle; rather,its vasoconstrictor effects are mediated indirectly by releaseof thromboxane from other vascular or perivascular cells,including astroglial and endothelial cells. This inference isbased on a number of observations: (1) 15-F2t-IsoP stimulatedthromboxane production and calcium signals from astroglialand endothelial cells but was ineffective on smooth muscle(Figures 4 and 5); (2) periventricular vasoconstriction to15-F2t-IsoP is thromboxane dependent (Figure 2);
(3) thromboxane mimetic U46619 evoked a nifedipine-sensitive calcium transient in smooth muscle and a vasocon-striction (Figures 5E, 5F, and 6); and (4) 15-F2t-IsoP–inducedvasoconstriction was nearly totally abolished by nifedipineand partly by v-conotoxin and SK&F96365 (Figure 6),according to the relative abundance of cells containingcorresponding channels (astroglial and endothelial cells).
In conclusion, the present study shows that 15-F2t-IsoPcauses more pronounced vasoconstriction in the periventric-ular brain region of the fetus than in that of older subjectsbecause of greater thromboxane formation, dependent on anewly described complex mechanism involving interactionbetween astrocytes and endothelial and smooth muscle cells;a model depicting this interrelationship is shown in Figure 7.We speculate that 15-F2t-IsoP may be a contributory factor inthe hemodynamic compromise and periventricular brain in-jury in the premature neonate during oxidant stress; cycloox-ygenase inhibitors, thromboxane synthase, and/or receptorblockers may attenuate the deleterious effects of oxidantstress.47,48
AcknowledgmentsThis work was supported by grants from the Medical ResearchCouncil of Canada, the Hospital for Sick Children Foundation, theMarch of Dimes Birth Defects Foundation, the Heart and StrokeFoundation of Quebec, and the Fonds de la Recherche en Sante´ duQuebec. Drs Gobeil and Chemtob are recipients of fellowship andscientist awards, respectively, from the Medical Research Council ofCanada. We wish to thank Hensy Fernandez for her technicalassistance.
References1. Southorn PA, Powis G. Free radicals in medicine, I: chemical nature and
biological reaction; II: involvement in human disease.Mayo ClinProc. 1988;63:381–408.
2. Volpe JJ. Brain injury in the premature infant: overview of clinicalaspects, neuropathology, and pathogenesis.Semin Pediatr Neurol. 1998;5:135–151.
3. Sullivan JL. Iron, plasma antioxidants, and the oxygen radical disease ofprematurity.Am J Dis Child. 1988;142:1341–1344.
Figure 7. Model based on data presented,depicting mechanism of action of 15-F2t-IsoP on periventricular brain microvesselsof immature subjects involving activationof specific channels in endothelial andastroglial cells resulting in thromboxaneformation and their interaction with smoothmuscle cells to elicit 15-F2t-IsoP–inducedvasoconstriction, found to be greater infetal and newborn than juvenile animals(Figure 1). F indicates fetus; NB, newborn;A, adult; R, receptive site for 15-F2t-IsoP(unknown); ROC, receptor-operated Ca21
channel; PLA2, phospholipase A2; COX,cyclooxygenase; AA, arachidonic acid;N-VOC, N-type voltage-gated Ca21 chan-nels; TP, thromboxane receptor; andL-VOC, L-type voltage-gated Ca21
channels.
Hou et al 15-F2t-Isoprostane Effects on Brain Microvessels 523
by guest on May 25, 2015http://stroke.ahajournals.org/Downloaded from

4. Taylor DL, Edwards AD, Mehmet H. Oxidative metabolism, apoptosisand perinatal brain injury.Brain Pathol. 1999;9:93–117.
5. Noseworthy MD, Bray TM. Effect of oxidative stress on brain damagedetected by MRI and in vivo 31P-NMR.Free Radic Biol Med. 1998;24:942–951.
6. Vannucci RC, Perlman JM. Interventions for perinatal hypoxic-ischemicencephalopathy.Pediatrics. 1997;100:1004–1014.
7. Hoffman SW, Moore S, Ellis EF. Isoprostanes: free radical-generatedprostaglandins with constrictor effects on cerebral arterioles.Stroke.1997;28:844–849.
8. Sano K, Asano T, Tanishima T, Sasaki T. Lipid peroxidation as a causeof cerebral vasospasm.Neurol Res. 1980;2:253–272.
9. Abran D, Varma DR, Chemtob S. Increased thromboxane-mediated con-tractions of retinal vessels of newborn pigs to peroxides.Am J Physiol.1995;268(Heart Circ Physiol. 37):H628–H632.
10. Chemtob S, Hardy P, Abran D, Li DY, Peri K, Cuzzani O, Varma DR.Peroxide cyclooxygenase interactions in postasphyxial changes in retinaland choroidal hemodynamics.J Appl Physiol. 1995;78:2039–2046.
11. Leffler CW, Busija DW, Armstead WM, Mirro R. H2O2 effects oncerebral prostanoids and pial arteriolar diameter in piglets.Am J Physiol.1990;258(Heart Circ Physiol. 27):H1382–HI387.
12. Tate RM, Morris HG, Schroeder WR, Repine JE. Oxygen metabolitesstimulate thromboxane production and vasoconstriction in isolated saline-perfused rabbit lungs.J Clin Invest. 1984;74:608–613.
13. Morrow JD, Roberts LJ II. The isoprostanes: current knowledge anddirections for future research.Biochem Pharmacol. 1996;51:1–9.
14. Roberts LJ II, Morrow JD. Generation and actions of isoprostanes.Biochim Biophys Acta. 1997;1345:121–135.
15. Morrow JD, Roberts LJ II. The isoprostanes: unique bioactive products oflipid peroxidation.Prog Lipid Res. 1997;36:1–36.
16. Morrow JD, Awad JA, Boss HJ, Blair IA, Roberts LJ II. Non-cycloox-ygenase-derived prostanoids (F2-isoprostanes) are formed in situ on phos-pholipids.Proc Natl Acad Sci U S A. 1992;89:10721–10725.
17. Longmire AW, Swift LL, Roberts LJ II, Awad JA, Burk RF, Morrow JD.Effect of oxygen tension on the generation of F2-isoprostanes and mal-ondialdehyde in peroxidizing rat liver microsomes.Biochem Pharmacol.1994;47:1173–1177.
18. Morrow JD, Hill KE, Burk RF, Nammour TM, Badr KF, Roberts LJ II.A series of prostaglandin F2-like compounds are produced in vivo inhumans by a non-cyclooxygenase, free radical-catalyzed mechanism.Proc Natl Acad Sci U S A. 1990;87:9383–9387.
19. Salabudeen A, Badr KF, Morrow JD, Roberts LJ II. Hydrogen peroxideinduces 21-aminosteroid-inhibitable F2-isoprostane production andcytolysis in renal tubular epithelial cells.J Am Soc Nephrol. 1995;6:1300–1303.
20. Taber DF, Morrow JD, Roberts LJ II. A nomenclature system for theisoprostanes.Prostaglandins. 1997;53:63–67.
21. Lahaie I, Hardy P, Hou X, Hassessian H, Asselin P, Lachapelle P,Almazan G, Varma DR, Morrow JD, Roberts LJ II, Chemtob S. A novelmechanism for vasoconstrictor action of 8-isoprostaglandin F2 alpha onretinal vessels.Am J Physiol. 1998;274(pt 2):R1406–R1416.
22. Banerjee M, Kang KH, Morrow JD, Roberts LJ, Newman JH. Effects ofa novel prostaglandin, 8-epi-PGF2a, in rabbit lung in situ.Am J Physiol.1992;263(Heart Circ Physiol. 32):H660–H663.
23. Kang KH, Morrow JD, Roberts II LJ, Newman JH, Banerjee M. Airwayand vascular effects of 8-epi-prostaglandin F2a in isolated perfused ratlung. J Appl Physiol. 1993;74:460–465.
24. Kromer BM, Tippins JR. Coronary artery constriction by the isoprostane8-epi prostaglandin F2a. Br J Phamacol. 1996;119:1276–1280.
25. Fukunaga M, Makita N, Roberts LJ II, Morrow JD, Takahashi K, BadrKF. Evidence for the existence of F2-isoprostane receptors on rat vascularsmooth muscle cells.Am J Physiol. 1993;264(Cell Physiol. 33):C1619–C1624.
26. Pratico D, Smyth EM, Violi F, Fitzgerald A. Local amplification ofplatelet function by 8-epi prostaglandin F2a is not mediated bythromboxane receptor isoforms.J Biol Chem. 1996;271:14916–14924.
27. Yura T, Fukunaga M, Grygorczyk R, Makita N, Takahashi K, Badr KF.Molecular and functional evidence for the distinct nature ofF2-isoprostane receptors from those of thromboxane A2. Adv Prostaglan-din Thromboxane Leukot Res. 1995;23:237–239.
28. Wagner RS, Weare C, Jin N, Mohler ER, Rhoades RA. Characterizationof signal transduction events stimulated by 8-epi-prostaglandin (PG)F2
alpha in rat aortic rings.Prostaglandins. 1997;54:581–599.29. Li DY, Hardy P, Abran D, Martinez-Bermudez AK, Guerguerian AM,
Bhattacharya M, Almazan G, Menezes R, Peri KG, Varma DR, ChemtobS. Key role for cyclooxygenase-2 in PGE2 and PGF2 alpha receptorregulation and cerebral blood flow of the newborn.Am J Physiol. 1997;273(pt 2):R1283–R1290.
30. Li DY, Abran D, Peri KG, Varma DR, Chemtob S. Inhibition of prosta-glandin synthesis in newborn pigs increases cerebral microvessel prosta-glandin F2 alpha and prostaglandin E2 receptors, their second messengersand vasoconstrictor response to adult levels.J Pharmacol Exp Ther.1996;278:370–377.
31. Harper SL, Bohlen HG, Rubin MJ. Arterial and microvascular contri-butions to cerebral cortical autoregulation in rats.Am J Physiol. 1984;246:H17–H24.
32. Fox JA. Novel omega-conopeptides reduced field potential amplitudes inthe rat hippocampal slice.Neurosci Lett. 1994;165:157–160.
33. Magolda RL, Ripka WC, Galbraith W. Novel synthesis of potent site-specific phospholipase A2 inhibitors. In: Bailey JM, ed.Prostaglandins,Leukotrienes and Lipoxins. New York, NY: Plenum Press; 1985:669–676.
34. Merritt JE, Armstrong WP, Benham CD, Hallam TJ, Jacob R, Jaxa-Chamiec A, Leigh BK, McCarthy SA, Moores KE, Rink TJ. SK&F96365, a novel inhibitor of receptor-mediated calcium entry.Biochem J.1990;271:515–522.
35. Rivier J, Galyean R, Gray WR, Azimi-Zonooz A, McIntosh JM, Cruz LJ,Olivera BM. Neuronal calcium channel inhibitors: synthesis of omega-conotoxin GVIA and effects on 45Ca uptake by synaptosomes.J BiolChem. 1987;262:1194–1198.
36. Himmel HM, Whorton AR, Strauss HC. Intracellular calcium, currents,and stimulus-response coupling in endothelial cells.Hypertension. 1993;21:112–127.
37. Fern R. Intracellular calcium and cell death during ischemia in neonatalrat white matter astrocytes in situ.J Neurosci. 1998;18:7232–7243.
38. Porter JT, McCarthy KD. Hippocampal astrocytes in situ respond toglutamate released from synaptic terminals.J Neurosci. 1996;16:5073–5081.
39. Li DY, Varma DR, Chemtob S. Ontogenic increase in PGE2 and PGF2alpha receptor density in brain microvessels of pigs.Br J Pharmacol.1994;112:59–64.
40. Almazan G, Afar DE, Bell JC. Phosphorylation and disruption of inter-mediate filament proteins in oligodendrocyte precursor cultures treatedwith calyculin A. J Neurosci Res. 1993;36:163–172.
41. Grynkiewicz G, Poenie M, Tsien RY. A new generation of Ca21 indi-cators with greatly improved fluorescence properties.J Biol Chem. 1985;260:3440–3450.
42. Peri K, Hardy P, Li DY, Varma DR, Chemtob S. Prostaglandin G/Hsynthase-2 is a major contributor of brain prostaglandins in the newborn.J Biol Chem. 1995;270:24615–24620.
43. Westenbroek RE, Bausch SB, Lin RC, Franck JE, Noebels JL, CatterallWA. Upregulation of L-type Ca21 channels in reactive astrocytes afterbrain injury, hypomyelination, and ischemia.J Neurosci. 1998;18:2321–2334.
44. McDonough W, Tran N, Giese A, Norman SA, Berens ME. Altered geneexpression in human astrocytoma cells selected for migration, I:thromboxane synthase.J Neuropathol Exp Neurol. 1998;57:449–455.
45. Murphy S, Pearce B, Jeremy J, Dandona P. Astrocytes as eicosanoid-producing cells.Glia. 1988;1:241–245.
46. Harder DR, Alkayed NJ, Lange AR, Gebremedhin D, Roman RJ. Func-tional hyperemia in the brain: hypothesis for astrocyte-derived vasodilatormetabolites.Stroke. 1998;29:229–234.
47. Ment LR, Stewart WB, Petroff OA, Duncan CC. Thromboxane synthesisinhibitor in a beagle pup model of perinatal asphyxia.Stroke. 1989;20:809–814.
48. Iijima T, Sawa H, Shiokawa Y, Saito I, Ishii H, Nakamura Z, Sankawa H.Thromboxane synthetase inhibitor ameliorates delayed neuronal death inthe CA1 subfield of the hippocampus after transient global ischemia ingerbils.J Neurosurg Anesthesiol. 1996;8:237–242.
524 Stroke February 2000
by guest on May 25, 2015http://stroke.ahajournals.org/Downloaded from

Lachapelle, Jackson Roberts II, Daya R. Varma and Sylvain ChemtobXin Hou, Fernand Gobeil, Jr, Krishna Peri, Giovanna Speranza, Anne Marilise Marrache, Pierre
) in Immature Pig Periventricular Brain Microvesselsα2(8-Iso-Prostaglandin F-Isoprostane2tAugmented Vasoconstriction and Thromboxane Formation by 15-F
Print ISSN: 0039-2499. Online ISSN: 1524-4628 Copyright © 2000 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Stroke doi: 10.1161/01.STR.31.2.516
2000;31:516-524Stroke.
http://stroke.ahajournals.org/content/31/2/516World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://stroke.ahajournals.org//subscriptions/
is online at: Stroke Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer process is available in the
Request Permissions in the middle column of the Web page under Services. Further information about thisOnce the online version of the published article for which permission is being requested is located, click
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office.Strokein Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on May 25, 2015http://stroke.ahajournals.org/Downloaded from




















