
Formation of F-ring Isoprostane-like Compounds (F3-Isoprostanes) in Vivo from Eicosapentaenoic Acid
8
Formation of F-ring Isoprostane-like Compounds (F 3 -Isoprostanes) in Vivo from Eicosapentaenoic Acid * Received for publication, February 2, 2006, and in revised form, March 16, 2006 Published, JBC Papers in Press, March 28, 2006, DOI 10.1074/jbc.M601035200 Ling Gao ‡ , Huiyong Yin ‡ , Ginger L. Milne ‡ , Ned A. Porter § , and Jason D. Morrow ‡1 From the ‡ Division of Clinical Pharmacology, Departments of Medicine and Pharmacology and the § Department of Chemistry, Vanderbilt University, Nashville, Tennessee 37232 Eicosapentaenoic acid (EPA, C20:5, -3) is the most abundant polyunsaturated fatty acid (PUFA) in fish oil. Recent studies suggest that the beneficial effects of fish oil are due, in part, to the genera- tion of various free radical-generated non-enzymatic bioactive oxi- dation products from -3 PUFAs, although the specific molecular species responsible for these effects have not been identified. Our research group has previously reported that pro-inflammatory prostaglandin F 2 -like compounds, termed F 2 -isoprostanes (IsoPs), are produced in vivo by the free radical-catalyzed peroxidation of arachidonic acid and represent one of the major products resulting from the oxidation of this PUFA. Based on these observations, we questioned whether F 2 -IsoP-like compounds (F 3 -IsoPs) are formed from the oxidation of EPA in vivo. Oxidation of EPA in vitro yielded a series of compounds that were structurally estab- lished to be F 3 -IsoPs using a number of chemical and mass spectro- metric approaches. The amounts formed were extremely large (up to 8.7 1.0 g/mg EPA) and greater than levels of F 2 -IsoPs gener- ated from arachidonic acid. We then examined the formation of F 3 -IsoPs in vivo in mice. Levels of F 3 -IsoPs in tissues such as heart are virtually undetectable at baseline, but supplementation of ani- mals with EPA markedly increases quantities up to 27.4 5.6 ng/g of heart. Interestingly, EPA supplementation also markedly re- duced levels of pro-inflammatory arachidonate-derived F 2 -IsoPs by up to 64% ( p < 0.05). Our studies provide the first evidence that identify F 3 -IsoPs as novel oxidation products of EPA that are gen- erated in vivo. Further understanding of the biological conse- quences of F 3 -IsoP formation may provide valuable insights into the cardioprotective mechanism of EPA. Isoprostanes (IsoPs) 2 are prostaglandin (PG)-like compounds that are formed non-enzymatically from the free radical-induced oxidation of arachidonic acid. Formation of these compounds proceeds through the generation of bicyclic endoperoxide PGH 2 -like intermediates that are reduced to PGF 2 -like compounds termed F 2 -IsoPs (1) or undergo rearrangement to PGD 2 and PGE 2 -like compounds (D 2 /E 2 -IsoPs) (2) and thromboxane-like compounds (isothromboxanes) (3). Unlike cyclooxygenase-derived PGs, IsoPs are formed in situ esterified to phos- pholipids and are subsequently released, presumably by the action of a phospholipase A 2 (4). Quantification of F 2 -IsoPs has been shown to be one of the most reliable markers to assess oxidant stress status and lipid peroxidation in vivo (5–7). In addition, F 2 -IsoPs have been shown to exert potent vasoconstrictive activities in the vasculature (8, 9), suggest- ing a pathogenic role for these compounds in diseases such as athero- sclerosis (10, 11). Eicosapentaenoic acid (C20:5 -3; EPA) is among the more abundant omega-3 polyunsaturated fatty acids (PUFA) present in fish oils. The other important PUFA in fish oil is docosahexaenoic acid (C22:6 -3; DHA). Studies in animals as well as human epidemiological studies, and more recently, clinical intervention trials, suggest that fish consumption or dietary fish oil supplementation reduces the incidence of important diseases including atherosclerosis and sudden death, neurodegenera- tion, and various inflammatory disorders (12, 13). The mechanism(s) by which these beneficial effects occurs is unknown, but it has been hypothesized that various enzymatically and non-enzymatically gener- ated peroxidation products of EPA possess anti-inflammatory activities (14 –19). Previously, A ¨ nggård and colleagues (20) provided limited evidence that F-ring IsoP-like compounds (F 3 -IsoPs) can be formed from the oxidation of EPA in vitro. These compounds are termed F 3 -IsoPs because they are structurally analogous to F 2 -IsoPs yet contain three double bonds. We therefore considered the possibility that IsoP-like compounds could be formed by the free radical-induced peroxidation of EPA in vivo. The rationale for undertaking these studies, in part, was based on the hypothesis that F 3 -IsoPs contribute to the beneficial bio- logical effects of EPA and fish oil supplementation in that they may exert biological activities that are anti-inflammatory as compared with F 2 -IsoPs. Indeed, one report states that the EPA-derived IsoP, 15-F 3t - IsoP, possesses activity that is different from 15-F 2t -IsoP in that it does not affect human platelet shape change or aggregation (21). 15-F 2t -IsoP is a ligand for the thromboxane receptor and induces platelet shape change and also causes vasoconstriction (1, 22). The lack of activity of 15-F 3t -IsoP is consistent with observations regarding EPA-derived PGs in that these latter compounds exert either weaker agonist effects or no effects in comparison with arachidonate-derived PGs (12, 23, 24). Based on studies of arachidonic acid oxidation, the mechanism by which F 3 -IsoPs could be formed is outlined in Fig. 1. There are four bis-allylic positions in EPA at carbons 7, 10, 13, and 16 as opposed to three bis-allylic positions in arachidonate where hydrogen abstraction can occur. Depending on the position of hydrogen abstraction and oxy- gen insertion, eight hydroperoxides are formed. These eight hydroper- oxides subsequently generate six F 3 -IsoP regioisomers. Each regioiso- mer is theoretically comprised of eight racemic diastereomers for a total of 96 compounds. A nomenclature system for the IsoPs has been estab- lished and approved by the Eicosanoid Nomenclature Committee in which the different regioisomer classes are designated by the carbon number on which the side chain hydroxyl is located with the carboxyl carbon designated as C-1 (25). Thus, in accordance with this nomen- * This work was supported by National Institutes of Health Grants GM15431, ES31125, RR00096, DK48831, and CA77839. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 To whom correspondence should be addressed. Tel.: 615-343-1124; Fax: 615-322-3669; E-mail: [email protected]. 2 The abbreviations used are: IsoP, isoprostane; PG, prostaglandin; EPA, eicosapentae- noic acid; PUFA, polyunsaturated fatty acid; AAPH, 2,2-azobis(2-amidinopropane) hydrochloride; GC, gas chromatography; NICI, negative ion chemical ionization; MS, mass spectrometry; LC, liquid chromatography; ESI, electrospray ionization; CID, col- lision-induced dissociation; SRM, selective reaction monitoring; RT, retention time. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 281, NO. 20, pp. 14092–14099, May 19, 2006 © 2006 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. 14092 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 20 • MAY 19, 2006
-
Upload
vanderbilt -
Category
Documents
-
view
5 -
download
0
Transcript of Formation of F-ring Isoprostane-like Compounds (F3-Isoprostanes) in Vivo from Eicosapentaenoic Acid

Formation of F-ring Isoprostane-like Compounds(F3-Isoprostanes) in Vivo from Eicosapentaenoic Acid*
Received for publication, February 2, 2006, and in revised form, March 16, 2006 Published, JBC Papers in Press, March 28, 2006, DOI 10.1074/jbc.M601035200
Ling Gao‡, Huiyong Yin‡, Ginger L. Milne‡, Ned A. Porter§, and Jason D. Morrow‡1
From the ‡Division of Clinical Pharmacology, Departments of Medicine and Pharmacology and the §Department of Chemistry,Vanderbilt University, Nashville, Tennessee 37232
Eicosapentaenoic acid (EPA, C20:5, �-3) is the most abundantpolyunsaturated fatty acid (PUFA) in fish oil. Recent studies suggestthat the beneficial effects of fish oil are due, in part, to the genera-tion of various free radical-generated non-enzymatic bioactive oxi-dation products from �-3 PUFAs, although the specific molecularspecies responsible for these effects have not been identified. Ourresearch group has previously reported that pro-inflammatoryprostaglandin F2-like compounds, termed F2-isoprostanes (IsoPs),are produced in vivo by the free radical-catalyzed peroxidation ofarachidonic acid and represent one of the major products resultingfrom the oxidation of this PUFA. Based on these observations, wequestioned whether F2-IsoP-like compounds (F3-IsoPs) areformed from the oxidation of EPA in vivo. Oxidation of EPAin vitro yielded a series of compounds that were structurally estab-lished to be F3-IsoPs using a number of chemical andmass spectro-metric approaches. The amounts formed were extremely large (upto 8.7 � 1.0 �g/mg EPA) and greater than levels of F2-IsoPs gener-ated from arachidonic acid. We then examined the formation ofF3-IsoPs in vivo in mice. Levels of F3-IsoPs in tissues such as heartare virtually undetectable at baseline, but supplementation of ani-mals with EPA markedly increases quantities up to 27.4 � 5.6 ng/gof heart. Interestingly, EPA supplementation also markedly re-duced levels of pro-inflammatory arachidonate-derived F2-IsoPs byup to 64% (p < 0.05). Our studies provide the first evidence thatidentify F3-IsoPs as novel oxidation products of EPA that are gen-erated in vivo. Further understanding of the biological conse-quences of F3-IsoP formationmay provide valuable insights into thecardioprotective mechanism of EPA.
Isoprostanes (IsoPs)2 are prostaglandin (PG)-like compounds thatare formed non-enzymatically from the free radical-induced oxidationof arachidonic acid. Formation of these compounds proceeds throughthe generation of bicyclic endoperoxide PGH2-like intermediates thatare reduced to PGF2-like compounds termed F2-IsoPs (1) or undergorearrangement to PGD2 and PGE2-like compounds (D2/E2-IsoPs) (2)and thromboxane-like compounds (isothromboxanes) (3). Unlikecyclooxygenase-derived PGs, IsoPs are formed in situ esterified to phos-pholipids and are subsequently released, presumably by the action of a
phospholipase A2 (4). Quantification of F2-IsoPs has been shown to beone of the most reliable markers to assess oxidant stress status and lipidperoxidation in vivo (5–7). In addition, F2-IsoPs have been shown toexert potent vasoconstrictive activities in the vasculature (8, 9), suggest-ing a pathogenic role for these compounds in diseases such as athero-sclerosis (10, 11).Eicosapentaenoic acid (C20:5�-3; EPA) is among themore abundant
omega-3 polyunsaturated fatty acids (PUFA) present in fish oils. Theother important PUFA in fish oil is docosahexaenoic acid (C22:6 �-3;DHA). Studies in animals as well as human epidemiological studies, andmore recently, clinical intervention trials, suggest that fish consumptionor dietary fish oil supplementation reduces the incidence of importantdiseases including atherosclerosis and sudden death, neurodegenera-tion, and various inflammatory disorders (12, 13). Themechanism(s) bywhich these beneficial effects occurs is unknown, but it has beenhypothesized that various enzymatically and non-enzymatically gener-ated peroxidation products of EPA possess anti-inflammatory activities(14–19).Previously, Anggård and colleagues (20) provided limited evidence
that F-ring IsoP-like compounds (F3-IsoPs) can be formed from theoxidation of EPA in vitro. These compounds are termed F3-IsoPsbecause they are structurally analogous to F2-IsoPs yet contain threedouble bonds. We therefore considered the possibility that IsoP-likecompounds could be formed by the free radical-induced peroxidation ofEPA in vivo. The rationale for undertaking these studies, in part, wasbased on the hypothesis that F3-IsoPs contribute to the beneficial bio-logical effects of EPA and fish oil supplementation in that theymay exertbiological activities that are anti-inflammatory as compared withF2-IsoPs. Indeed, one report states that the EPA-derived IsoP, 15-F3t-IsoP, possesses activity that is different from 15-F2t-IsoP in that it doesnot affect human platelet shape change or aggregation (21). 15-F2t-IsoPis a ligand for the thromboxane receptor and induces platelet shapechange and also causes vasoconstriction (1, 22). The lack of activity of15-F3t-IsoP is consistent with observations regarding EPA-derived PGsin that these latter compounds exert either weaker agonist effects or noeffects in comparison with arachidonate-derived PGs (12, 23, 24).Based on studies of arachidonic acid oxidation, the mechanism by
which F3-IsoPs could be formed is outlined in Fig. 1. There are fourbis-allylic positions in EPA at carbons 7, 10, 13, and 16 as opposed tothree bis-allylic positions in arachidonate where hydrogen abstractioncan occur. Depending on the position of hydrogen abstraction and oxy-gen insertion, eight hydroperoxides are formed. These eight hydroper-oxides subsequently generate six F3-IsoP regioisomers. Each regioiso-mer is theoretically comprised of eight racemic diastereomers for a totalof 96 compounds. A nomenclature system for the IsoPs has been estab-lished and approved by the Eicosanoid Nomenclature Committee inwhich the different regioisomer classes are designated by the carbonnumber on which the side chain hydroxyl is located with the carboxylcarbon designated as C-1 (25). Thus, in accordance with this nomen-
* This work was supported by National Institutes of Health Grants GM15431, ES31125,RR00096, DK48831, and CA77839. The costs of publication of this article weredefrayed in part by the payment of page charges. This article must therefore behereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
1 To whom correspondence should be addressed. Tel.: 615-343-1124; Fax: 615-322-3669;E-mail: [email protected].
2 The abbreviations used are: IsoP, isoprostane; PG, prostaglandin; EPA, eicosapentae-noic acid; PUFA, polyunsaturated fatty acid; AAPH, 2,2�-azobis(2-amidinopropane)hydrochloride; GC, gas chromatography; NICI, negative ion chemical ionization; MS,mass spectrometry; LC, liquid chromatography; ESI, electrospray ionization; CID, col-lision-induced dissociation; SRM, selective reaction monitoring; RT, retention time.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 281, NO. 20, pp. 14092–14099, May 19, 2006© 2006 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
14092 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 20 • MAY 19, 2006

clature system, the F3-IsoP regioisomers are designated as 5-, 8-, 11-,12-, 15-, and 18-series F3-IsoPs. Herein, we present evidence thatF3-IsoPs are, in fact, formed in significant amounts in vitro and in vivofrom the free radical-catalyzed peroxidation of EPA.
EXPERIMENTAL PROCEDURES
Materials—Eicosapentaenoic acid and d4-15-F2t-IsoP (8-iso-PGF2�)were purchased fromCayman Chemical Co. (Ann Arbor, MI). Dimeth-ylformamide and undecane were obtained from Aldrich. Pentafluoro-benzyl bromide and diisopropylethylamine were from Sigma. 2,2�-Azo-bis(2-amidinopropane) hydrochloride (AAPH) was from EastmanKodakCo.N,O-bis (trimethylsilyl) trifluoroacetamidewas fromSupelco(Bellefonte, PA). [2H9]N,O-bis (trimethylsilyl) acetamide was fromCDN Isotopes (Pointe-Claire, PQ). C-18 and silica Sep-Pak cartridgeswere from Waters Associates (Milford, MA). 60ALK6D TLC plateswere fromWhatman (Maidstone, UK).
Oxidation of EPA—Five milligram of fresh EPA was dissolved in 100�l of ethanol and added immediately to 4.9 ml of phosphate-buffered
saline solution (pH 7.4) containing 10 mM AAPH. The EPA oxidationreaction mixture was incubated in a shaking water bath at 37 °C forvarying amounts of time, after which it was placed immediately at�80 °C until further processing.
Purification and Analysis of F3-IsoPs—Free and esterified F3-IsoPswere extracted using C-18 and Silica Sep-Pak cartridges, converted to apentafluorobenzyl ester, purified by TLC, converted to a trimethylsilylether derivative, and quantified by stable isotope dilution gas chroma-tography (GC)/negative ion chemical ionization (NICI)mass spectrom-etry (MS) with d4-15-F2t-IsoP as an internal standard using a modifica-tion of the method described for the quantification of F2-IsoPs (26).Instead of scraping 1 cm below to 1 cm above where PGF2� methyl estermigrates on TLC for analysis of F2-IsoPs, the scraped area was extendedto 3.5 cm above where the PGF2� methyl ester migrates. This extendedarea of the TLC plate was determined to contain F3-IsoPs by analyzingsmall 5-mm cuts using approaches for their identification describedbelow. The M–�CH2C6F5 ions were monitored for quantification (m/z567 for F3-IsoPs andm/z 573 for d4-15-F2t-IsoP). Quantification of the
FIGURE 1. Pathway for the formation of F3-IsoPsby non-enzymatic peroxidation of EPA (A and B).
Characterization of Eicosapentaenoic Acid Eicosanoids
MAY 19, 2006 • VOLUME 281 • NUMBER 20 JOURNAL OF BIOLOGICAL CHEMISTRY 14093

total amount of F3-IsoPs was determined by integrating the peak area ofmaterial in them/z 567 channel in comparison with them/z 573 chan-nel. GC/NICI/MSwas carried out using anAgilent Technologies 6890NNetwork GC/MS system. Sample work using this method does not leadto the generation of F3-IsoPs during purification and derivatization. Insome studies, F2-IsoPs were quantified as described previously (5).
Analysis of F3-IsoPs by Liquid Chromatography (LC)/ElectrosprayIonization (ESI)/MS/MS—F3-IsoPs generated by the in vitro and in vivooxidation of EPAwere extracted by C-18 and Silica Sep-Pak cartridge asdescribed above. The samples were then analyzed by negative ionESI-MS coupled with reverse-phase high pressure liquid chromatogra-phy separation using a SupelcoDiscovery C18 column (15 cm� 2.1mm,5 �m) at a flow rate of 0.2 ml/min with a linear gradient starting with80% solvent A (2 mM ammonium acetate) to 65% in 20 min. Mobilephase B consisted of acetonitrile/methanol (95:5). The ESI source wasfitted with a deactivated fused silica capillary (100 �m inner diameter).Nitrogenwas used as both the sheath gas and the auxiliary gas, at 45 and17 p.s.i., respectively. The capillary temperature was 280 °C. The sprayvoltage was 4.3 kV, and the tube lens voltage was 80 V. Collision-in-duced dissociation (CID) of the molecular ion of putative F3-IsoPs wasperformed from 20 to 30 eV under 1.5 millitorrs of argon. Spectra thatare shown were obtained at 25 eV. Spectra were displayed by averagingscans across chromatographic peaks. Selective reaction monitoring(SRM) was performed according to characteristic fragmentation pat-terns of F2-IsoPs (27). The collision energy for SRM was 30 eV. LC/MS
was carried out using a ThermoFinnigan TSQ Quantum 1.0 SR 1 massspectrometer. Data acquisition and analysis were performed usingXcaliber software, version 1.3.
Preparation of F3-IsoPs from Rodent Tissue—Mice (strain C57BL/6J(B6)) were fed with a rodent AIN-93 diet supplemented with 0, 0.28,0.56, or 0.84% EPA (by weight). In some studies, animals were adminis-tered CCl4 (1 ml/kg) intraperitoneally to induce an oxidant stress. After8 weeks of feeding, the mice were sacrificed. Heart and other tissueswere removed and immediately flash-frozen in liquid nitrogen andstored in�80 °C. Analyzed tissue samples were homogenized in 5ml ofice-cold chloroform:methanol (2:1, v/v) containing butylated hydroxy-toluene (0.005%) to prevent ex vivo auto-oxidation. Esterified F3-IsoPsin phospholipids were hydrolyzed to liberate free F3-IsoPs. As notedpreviously (28), the addition of various PUFAs including arachidonicacid, EPA, or DHA to tissues during workup does not increase the levelsof isoprostane-like compounds in tissue extracts. F3-IsoPs in sampleswere then purified and analyzed as described above. F2-IsoPs werequantified as described previously (5). Fatty acid content in tissues wasdetermined by GC (29). For some studies, male Sprague-Dawley ratswere fed a rodent AIN-93 diet supplemented with 0.56% EPA. After 8weeks of feeding, the rats were sacrificed. F3-IsoPs in liver tissues wereanalyzed by LC/ESI/MS/MS as described above.
RESULTS
Formation of F3-IsoPs in Vitro—A representative selective ion moni-toring chromatogram obtained from the AAPH-induced oxidation ofEPA in vitro is shown in Fig. 2. The chromatographic peak in the lowerm/z 573 ion current chromatogram represents the internal standardd4-15-F2t-IsoP. In the upper m/z 567 ion current chromatogram are aseries of chromatographic peaks eluting over approximately a 1-mininterval. These compounds possess a molecular mass predicted forF3-IsoPs. In addition, these compounds elute at later retention timesthan the deuterated 15-F2t-IsoP internal standard; it would be predictedthat the retention time of F3-IsoPs on GC should be longer than that ofF2-IsoPs because F3-IsoPs contain onemore double bond. Furthermore,it should be noted that the retention times over which the F3-IsoPs elutemay differ somewhat in the different figures because these analyseswereperformed on different days using different columns that vary some-what in length.Additional experimental approaches were then performed to provide
further evidence that the compounds represented by the chromato-graphic peaks in the m/z 567 ion current chromatogram are F3-IsoPs.
FIGURE 2. Selected ion current chromatograms obtained from the GC/MS analysisof F3-IsoPs generated during AAPH-induced oxidation of EPA in vitro. The series ofpeaks in the m/z 567 ion current chromatogram represent putative F3-IsoPs, and the singlepeak in the m/z 573 ion current chromatogram represents the d4-15-F2t-IsoP internal stand-ard. Compounds are analyzed as pentafluorobenzyl ester, trimethylsilyl ether derivatives.
FIGURE3.AnalysisofputativeF3-IsoPsasa[2H9]-trimethylsilyl ether derivative. Before [2H9]trime-thylsilyl ether derivatization, intense peaks are pres-ent in the m/z 567 ion current chromatogram (A)representing F3-IsoPs. There are no peaks present 27atomic mass units higher at m/z 594. Following[2H9]trimethylsilyl ether derivatization, intensepeaks appear in the m/z 594 ion current chromato-gram (B), indicating that compounds represented bythe peaks in the m/z 567 ion current chromatogramhave three hydroxyl groups. There are no peakspresent at m/z 567.
Characterization of Eicosapentaenoic Acid Eicosanoids
14094 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 20 • MAY 19, 2006
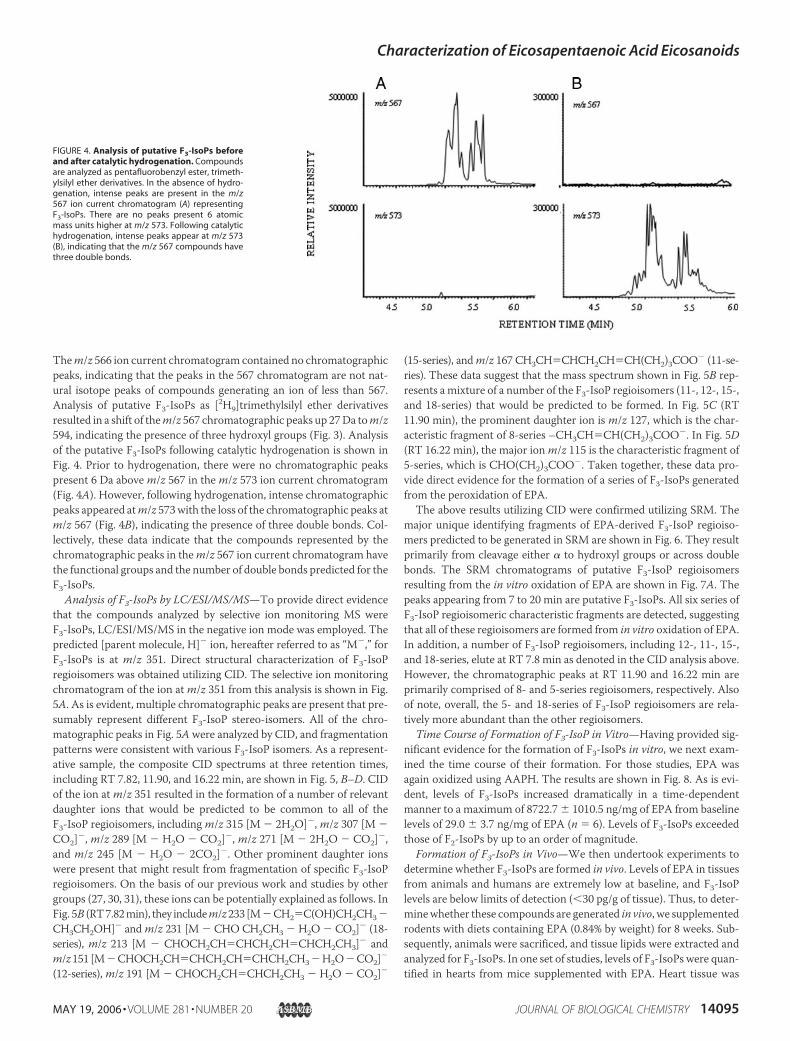
Them/z 566 ion current chromatogram contained no chromatographicpeaks, indicating that the peaks in the 567 chromatogram are not nat-ural isotope peaks of compounds generating an ion of less than 567.Analysis of putative F3-IsoPs as [2H9]trimethylsilyl ether derivativesresulted in a shift of them/z 567 chromatographic peaks up 27Da tom/z594, indicating the presence of three hydroxyl groups (Fig. 3). Analysisof the putative F3-IsoPs following catalytic hydrogenation is shown inFig. 4. Prior to hydrogenation, there were no chromatographic peakspresent 6 Da abovem/z 567 in them/z 573 ion current chromatogram(Fig. 4A). However, following hydrogenation, intense chromatographicpeaks appeared atm/z 573with the loss of the chromatographic peaks atm/z 567 (Fig. 4B), indicating the presence of three double bonds. Col-lectively, these data indicate that the compounds represented by thechromatographic peaks in them/z 567 ion current chromatogram havethe functional groups and the number of double bonds predicted for theF3-IsoPs.
Analysis of F3-IsoPs by LC/ESI/MS/MS—To provide direct evidencethat the compounds analyzed by selective ion monitoring MS wereF3-IsoPs, LC/ESI/MS/MS in the negative ion mode was employed. Thepredicted [parent molecule, H]� ion, hereafter referred to as “M�,” forF3-IsoPs is at m/z 351. Direct structural characterization of F3-IsoPregioisomers was obtained utilizing CID. The selective ion monitoringchromatogram of the ion atm/z 351 from this analysis is shown in Fig.5A. As is evident, multiple chromatographic peaks are present that pre-sumably represent different F3-IsoP stereo-isomers. All of the chro-matographic peaks in Fig. 5Awere analyzed by CID, and fragmentationpatterns were consistent with various F3-IsoP isomers. As a represent-ative sample, the composite CID spectrums at three retention times,including RT 7.82, 11.90, and 16.22 min, are shown in Fig. 5, B–D. CIDof the ion atm/z 351 resulted in the formation of a number of relevantdaughter ions that would be predicted to be common to all of theF3-IsoP regioisomers, includingm/z 315 [M � 2H2O]�,m/z 307 [M �CO2]�, m/z 289 [M � H2O � CO2]�, m/z 271 [M � 2H2O � CO2]�,and m/z 245 [M � H2O � 2CO2]�. Other prominent daughter ionswere present that might result from fragmentation of specific F3-IsoPregioisomers. On the basis of our previous work and studies by othergroups (27, 30, 31), these ions can be potentially explained as follows. InFig. 5B (RT7.82min), they includem/z233 [M�CH2�C(OH)CH2CH3�CH3CH2OH]� andm/z 231 [M � CHO CH2CH3 � H2O � CO2]� (18-series), m/z 213 [M � CHOCH2CH�CHCH2CH�CHCH2CH3]� andm/z151 [M�CHOCH2CH�CHCH2CH�CHCH2CH3�H2O�CO2]�
(12-series),m/z 191 [M � CHOCH2CH�CHCH2CH3 � H2O � CO2]�
(15-series), andm/z 167 CH3CH�CHCH2CH�CH(CH2)3COO� (11-se-ries). These data suggest that the mass spectrum shown in Fig. 5B rep-resents a mixture of a number of the F3-IsoP regioisomers (11-, 12-, 15-,and 18-series) that would be predicted to be formed. In Fig. 5C (RT11.90 min), the prominent daughter ion is m/z 127, which is the char-acteristic fragment of 8-series –CH3CH�CH(CH2)3COO�. In Fig. 5D(RT 16.22 min), the major ionm/z 115 is the characteristic fragment of5-series, which is CHO(CH2)3COO�. Taken together, these data pro-vide direct evidence for the formation of a series of F3-IsoPs generatedfrom the peroxidation of EPA.The above results utilizing CID were confirmed utilizing SRM. The
major unique identifying fragments of EPA-derived F3-IsoP regioiso-mers predicted to be generated in SRM are shown in Fig. 6. They resultprimarily from cleavage either � to hydroxyl groups or across doublebonds. The SRM chromatograms of putative F3-IsoP regioisomersresulting from the in vitro oxidation of EPA are shown in Fig. 7A. Thepeaks appearing from 7 to 20 min are putative F3-IsoPs. All six series ofF3-IsoP regioisomeric characteristic fragments are detected, suggestingthat all of these regioisomers are formed from in vitro oxidation of EPA.In addition, a number of F3-IsoP regioisomers, including 12-, 11-, 15-,and 18-series, elute at RT 7.8 min as denoted in the CID analysis above.However, the chromatographic peaks at RT 11.90 and 16.22 min areprimarily comprised of 8- and 5-series regioisomers, respectively. Alsoof note, overall, the 5- and 18-series of F3-IsoP regioisomers are rela-tively more abundant than the other regioisomers.
Time Course of Formation of F3-IsoP in Vitro—Having provided sig-nificant evidence for the formation of F3-IsoPs in vitro, we next exam-ined the time course of their formation. For those studies, EPA wasagain oxidized using AAPH. The results are shown in Fig. 8. As is evi-dent, levels of F3-IsoPs increased dramatically in a time-dependentmanner to a maximum of 8722.7 � 1010.5 ng/mg of EPA from baselinelevels of 29.0 � 3.7 ng/mg of EPA (n � 6). Levels of F3-IsoPs exceededthose of F2-IsoPs by up to an order of magnitude.
Formation of F3-IsoPs in Vivo—We then undertook experiments todetermine whether F3-IsoPs are formed in vivo. Levels of EPA in tissuesfrom animals and humans are extremely low at baseline, and F3-IsoPlevels are below limits of detection (�30 pg/g of tissue). Thus, to deter-minewhether these compounds are generated in vivo, we supplementedrodents with diets containing EPA (0.84% by weight) for 8 weeks. Sub-sequently, animals were sacrificed, and tissue lipids were extracted andanalyzed for F3-IsoPs. In one set of studies, levels of F3-IsoPs were quan-tified in hearts from mice supplemented with EPA. Heart tissue was
FIGURE 4. Analysis of putative F3-IsoPs beforeand after catalytic hydrogenation. Compoundsare analyzed as pentafluorobenzyl ester, trimeth-ylsilyl ether derivatives. In the absence of hydro-genation, intense peaks are present in the m/z567 ion current chromatogram (A) representingF3-IsoPs. There are no peaks present 6 atomicmass units higher at m/z 573. Following catalytichydrogenation, intense peaks appear at m/z 573(B), indicating that the m/z 567 compounds havethree double bonds.
Characterization of Eicosapentaenoic Acid Eicosanoids
MAY 19, 2006 • VOLUME 281 • NUMBER 20 JOURNAL OF BIOLOGICAL CHEMISTRY 14095

examined because supplementation of animals and humanswith fish oilmarkedly increases levels of EPA in this organ. In addition, fish oil sup-plementation is associated with a marked decrease in cardiovasculardisease. A representative GC/MS ion current chromatogram obtainedfromone of these analyses is shown in Fig. 9. The chromatographic peakin the lowerm/z 573 ion current chromatogram represents the internalstandard d4-15-F2t-IsoP. In the upper m/z 567 ion current chromato-gram are a series of chromatographic peaks that have a molecular massand retention times expected for the F3-IsoPs. The pattern of peaksrepresenting F3-IsoPs is very similar to that obtained from the oxidationof EPA in vitro. In addition, as predicted, these compounds were foundto contain three hydroxyl groups and three double bonds.Analogous to LC/ESI/MS/MS studies performed in vitro, experi-
ments were then carried out to obtain further evidence that the chro-matographic peaks in the m/z 567 ion current chromatogram of Fig. 9represent F3-IsoPs formed in vivo. For these studies, liver tissue fromrats fed a diet supplemented with 0.56% EPA was utilized to obtainadequate amounts of F3-IsoPs for analysis by LC/MS. In CID experi-ments, all of daughter ions that would be predicted to be common to all
of the F3-IsoP regioisomers, including m/z 333 [M � H2O]�, m/z 307[M � CO2]�, m/z 289 [M � H2O � CO2]�, m/z 271 [M � 2H2O �CO2]�, andm/z 245 [M � H2O � 2CO2]�, were detected. In addition,all 6-series regioisomeric characteristic fragments were formed in bothCID (data not shown) and SRM experiments (Fig. 7B). As is evident inFig. 7B, the relative abundance of different regioisomers in vivo is similarto in vitro studies. Taken together, these experiments provide evidencethat F3-IsoPs are formed in abundance in vivo and that the regioiso-meric distribution is similar to that observed in vitro.We performed studies to examine the effect of different concen-
trations of EPA on the generation of F3-IsoPs in vivo. Mice wereadministered 0, 0.28, 0.56, or 0.84% EPA (by weight) in their diets for8 weeks, and levels of EPA and F3-IsoPs were measured in hearttissue. As shown in Fig. 10, A and B, as the EPA concentration inheart tissues increases, levels of F3-IsoPs increase in a concentration-dependent manner. In addition, we also quantified the effect of EPAsupplementation on arachidonate content and F2-IsoP levels in thesame heart tissue. Interestingly, supplementation with EPA decreasedarachidonate content (Fig. 10C). Importantly, in addition, levels of
FIGURE 5. Analysis of putative F3-IsoPs as a free acid by LC/ESI/MS/MS. A, selected ion monitoring chromatogram of the [parent molecule, H]� (M�) ion at m/z 351 from LC/ESI/MSanalysis of putative F3-IsoPs obtained from AAPH-induced oxidation of EPA in vitro. Peaks that were subjected to CID are denoted by the arrows. The m/z 351 ion was subjected to CIDat 25eV, and daughter ions were scanned from 50 to 400. B, CID spectrum obtained by summing scans over the chromatographic peak at retention time 7.82 min. C, CID spectrumobtained by summing scans over the chromatographic peak at retention time 11.90 min. D, CID spectrum obtained by summing scans over the chromatographic peak at retentiontime 16.22 min.
Characterization of Eicosapentaenoic Acid Eicosanoids
14096 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 20 • MAY 19, 2006

F2-IsoPs decreased dramatically by up to 64% (p � 0.05) (Fig. 10D),suggesting that EPA effectively decreases levels of proinflammatoryF2-IsoPs formed from arachidonate.Finally, we also determined the effect of enhanced oxidant stress on
endogenous F3-IsoPs in EPA-supplemented (0.56%) mice. Levels ofthese compounds increased from 86.27 � 11.01 ng/g of tissue to
311.5 � 60.79 ng/g of tissue in livers after administration of CCl4 toinduce an oxidant stress.
DISCUSSION
Our studies have elucidated a novel class of F2-IsoP-like compounds,F3-IsoPs, formed in vivo from the free radical-induced peroxidation ofEPA. Previous studies by Anggård and colleagues (20) have describedthe formation of F3-IsoPs in vitro, although little structural informationwas provided, and it was not determined whether these compoundscould be generated in vivo (5). Levels of compounds generated signifi-cantly exceed those of F2-IsoPs derived from arachidonate, perhapsbecause EPA contains more double bonds and is more easily oxidized.The present studies describe the formation of F-type prostane ring-containing molecules. Free radical-induced peroxidation of arachido-nate results not only in the formation of F-ring IsoPs but also E/D-ringIsoPs, A/J-ring IsoPs, and thromboxane-like molecules (isothrombox-anes) (3, 32). Thus, although it remains the subject of future studies, it islikely that analogous compounds, in addition to F3-IsoPs, are alsoformed of products of the non-enzymatic peroxidation of EPA.A major impetus for undertaking the present studies regarding EPA
oxidation has been the fact that supplementation of humanswith fish oilcontaining large amounts of EPA andDHAhas been shownbeneficial inthe prevention of important human diseases such as atherosclerosis andsudden death, neurodegeneration, and various inflammatory disorders(12, 13). Although the mechanism(s) by which these beneficial effectsoccurs is unknown, a potentially important anti-atherogenic and anti-inflammatory mechanism of �-3 PUFAs is their interference with thearachidonic acid cascade that generates pro-inflammatory eicosanoids(12, 33). EPA not only can replace arachidonic acid in phospholipidbilayers but is also a competitive inhibitor of cyclooxygenase, reducingthe production of 2-series PGs and thromboxane, in addition to the4-series leukotrienes. As noted, the 3- and 5-series eicosanoids (respec-
FIGURE 6. Scheme for SRM fragmentation of different F3-IsoP regioisomers. Themajor route of fragmentation involves cleavage � to hydroxyl groups or across doublebonds.
FIGURE 7. SRM analysis of putative F3-IsoPs obtained from AAPH-induced oxidation of EPA. A, in vitro; B, from rat livers after 8 weeks of EPA supplementation (0.56%). Please referto Fig. 6 for specific ions being monitored for different F3-IsoP regioisomers.
Characterization of Eicosapentaenoic Acid Eicosanoids
MAY 19, 2006 • VOLUME 281 • NUMBER 20 JOURNAL OF BIOLOGICAL CHEMISTRY 14097

tively) derived from EPA are either less biologically active or inactive ascompared with the former products and are thus considered to exerteffects that are less inflammatory (34, 35). Further, Serhan and col-
leagues (14–17) have described a group of polyoxygenated DHA andEPA derivatives termed resolvins that are produced in various tissues.These compounds inhibit cytokine expression and other inflammatoryresponses in microglia, skin cells, and other cell types.Recently, there has been significant interest in the biological activities
of non-enzymatic free radical-initiated peroxidation products of �-3PUFAs. Sethi et al. (18, 19) reported that EPAoxidized in the presence ofCu2�, but not native EPA, significantly inhibits human neutrophil andmonocyte adhesion to endothelial cells, a process linked to the develop-ment of atherosclerosis and other inflammatory disorders. This effectwas induced via inhibition of endothelial adhesion receptor expressionand was modulated by the activation of the peroxisome proliferator-activated receptor-� by EPA oxidation products. In addition, oxidizedEPAmarkedly reduced leukocyte rolling and adhesion to venular endo-thelium of lipopolysaccharide-treated mice in vivo, and the effect wasnot observed in peroxisome proliferator-activated receptor-�-deficientmice. These studies suggest that the beneficial effects of �-3 fatty acidsmay be mediated, in part, by the anti-inflammatory effects of oxidizedEPA. Similarly, Vallve et al. (36) have shown that various non-enzymat-ically generated aldehyde oxidation products of EPA andDHAdecreasethe expression of the CD36 receptor in human macrophages. Up-regu-lation of this receptor has been linked to atherosclerosis. Additionalrecent reports have suggested that other related biological effects ofEPA and DHA, such as modulation of endothelial inflammatory mole-cules, are related to their peroxidation products (18, 37). Arita et al. (38)have also shown that non-enzymatically oxidized EPA enhances apo-ptosis in HL-60 leukemia cells, supporting the contention that oxidized�-3 PUFAs are both anti-proliferative and anti-inflammatory. Similarfindings have been reported in HepG2 (human hepatoma) cells andAH109A (rat liver cancer) cells (39, 40). In virtually none of thesereports, however, have the specific peroxidation products responsiblefor these effects been structurally identified. Further studies to charac-terize these molecules are needed. Thus, a rationale for undertakingthese studies was based on the hypothesis that F3-IsoPs are formed fromthe peroxidation of EPA and contribute to the beneficial biologicaleffects of EPA and fish oil supplementation in that they exert biological
FIGURE 8. Time course of formation of F3-IsoPs during oxidation of EPA in vitro byAAPH. Data are expressed as means � S.D. (n � 6).
FIGURE 9. Selected ion current chromatogram obtained from the GC/MS analysis forF3-IsoPs esterified in mouse hearts. The series of peaks in the m/z 567 ion currentchromatogram represent putative F3-IsoPs, and the single peak in the m/z 573 ion cur-rent chromatogram represents the d4-15-F2t-IsoP internal standard.
FIGURE 10. Levels of fatty acids and isopros-tanes from hearts of mice after 8 weeks of EPAsupplementation (0, 0.28, 0.56, 0.84% byweight). A, EPA; B, F3-IsoPs; C, arachidonate; D,F2-IsoPs. Data are expressed as means � S.D. (n �5). *, p � 0.05 as compared with control.
Characterization of Eicosapentaenoic Acid Eicosanoids
14098 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 20 • MAY 19, 2006

activities that are anti-inflammatory. Studies assessing the biologicalactivity of 15-F3t-IsoP could test this important hypothesis but will haveto await the chemical synthesis of the molecule.The studies reported herein have begun to systematically define the
oxidation of EPA in vivo and in vitro for the first time. F3-IsoPs wereidentified using a variety of complementary chemical and MS ap-proaches, including LC/ESI/MS/MS. As expected, 6-series of F3-IsoPregioisomers were identified from both in vitro and in vivo sources. Thefragmentation patterns of these regioisomers are similar to F2-IsoPregioisomers, and indeed, information that we have previously acquiredwith F2-IsoPs was extremely useful in the present studies (27). Of note,the relative abundance of 5- and 18-series F3-IsoPs predominates overthe other series. Such regioisomeric predominance has also beenreported for F2-IsoP regioisomers in which 5- and 15-series compoundsare formed in greater abundance than 8- and 12-series molecules (27).At least part of the reason for this is likely due to the fact that precursorsof 8- and 12-series F2-IsoPs can undergo further oxidation and cycliza-tion to yield a novel class of compounds termed dioxolane endoperox-ides (41). Although undetermined at present, it is likely that a similarmechanism may account for the predominance of 5- and 18-seriesF3-IsoPs.
Another interesting finding from the above studies relates to theobservation that EPA reduces the formation of F2-IsoPs in vivo. Asmentioned, F2-IsoPs are generally considered to be pro-inflammatorymolecules and have been implicated in the pathophysiological conse-quences of oxidative stress. It is thus intriguing to propose that part ofthe mechanism by which EPA prevents certain diseases associated withincreased inflammation relates to its ability to decrease F2-IsoPgeneration.In summary, we report the discovery that F-ring IsoP-like com-
pounds, termed F3-IsoPs, are formed in vivo in large amounts as prod-ucts of the non-enzymatic free radical-catalyzed peroxidation of EPA, amajor long chain polyunsaturated fatty acid in fish oil. Further under-standing of the biological consequences of the formation of these novelcompounds and factors influencing their formation and metabolismmay provide valuable insights into the role of EPA in human physiologyand pathophysiology.
REFERENCES1. Morrow, J. D., Hill, K. E., Burk, R. F., Nammour, T. M., Badr, K. F., and Roberts, L. J.,
II (1990) Proc. Natl. Acad. Sci. U. S. A. 87, 9383–93872. Morrow, J. D., Minton, T. A., Mukundan, C. R., Campbell, M. D., Zackert, W. E.,
Daniel, V. C., Badr, K. F., Blair, I. A., and Roberts, L. J. (1994) J. Biol. Chem. 269,4317–4326
3. Morrow, J. D., Awad, J. A., Wu, A., Zackert, W. E., Daniel, V. C., and Roberts, L. J., II(1996) J. Biol. Chem. 271, 23185–23190
4. Morrow, J. D., Awad, J. A., Boss, H. J., Blair, I. A., and Roberts, L. J., II (1992) Proc. Natl.Acad. Sci. U. S. A. 89, 10721–10725
5. Morrow, J. D., and Roberts, L. J., II (2002)Methods Mol. Biol. 186, 57–666. Morrow, J. D. (2000) Drug Metab. Rev. 32, 377–3857. Roberts, L. J., II, and Morrow, J. D. (2000) Free Radic. Biol. Med. 28, 505–5138. Kromer, B. M., and Tippins, J. R. (1996) Br. J. Pharmacol. 119, 1276–12809. Cracowski, J. L., Stanke-Labesque, F., Devillier, P., Chavanon, O., Hunt, M., Souvig-
net, C., and Bessard, G. (2000) Eur. J. Pharmacol. 397, 161–16810. Cracowski, J. L., Devillier, P., Durand, T., Stanke-Labesque, F., and Bessard, G. (2001)
J. Vasc. Res. 38, 93–10311. Habib, A., and Badr, K. F. (2004) Chem. Phys. Lipids 128, 69–7312. Kris-Etherton, P. M., Harris, W. S., and Appel, L. J. (2002) Circulation 106,
2747–275713. Ruxton, C. (2004) Nurs. Stand. 18, 38–4214. Hong, S., Gronert, K., Devchand, P. R., Moussignac, R. L., and Serhan, C. N. (2003)
J. Biol. Chem. 278, 14677–1468715. Serhan, C. N., and Levy, B. (2003) Chem. Immunol. Allergy 83, 115–14516. Serhan, C. N., Clish, C. B., Brannon, J., Colgan, S. P., Chiang, N., and Gronert, K.
(2000) J. Exp. Med. 192, 1197–120417. Serhan, C. N., Hong, S., Gronert, K., Colgan, S. P., Devchand, P. R., Mirick, G., and
Moussignac, R. L. (2002) J. Exp. Med. 196, 1025–103718. Sethi, S., Eastman, A. Y., and Eaton, J. W. (1996) J. Lab. Clin. Med. 128, 27–3819. Sethi, S. (2002) Redox. Rep. 7, 369–37820. Nourooz-Zadeh, J., Halliwell, B., and Anggård, E. (1997) Biochem. Biophys. Res. Com-
mun. 236, 467–47221. Pratico, D., Smyth, E. M., Violi, F., and FitzGerald, G. A. (1996) J. Biol. Chem. 271,
14916–1492422. Morrow, J. D., Minton, T. A., and Roberts, I. I. (1992) Prostaglandins 44, 155–16323. Balapure, A. K., Rexroad, J., Kawada, K., Watt, D. S., and Fitz, T. A. (1989) Biochem.
Pharmacol. 38, 2375–238124. Kulkarni, P. S., and Srinivasan, B. D. (1985) Investig. Ophthalmol. Vis. Sci. 26,
1178–118225. Taber, D. F., Morrow, J. D., and Roberts, L. J., II (1997) Prostaglandins 53, 63–6726. Morrow, J. D., and Roberts, L. J., II (1999)Methods Enzymol. 300, 3–1227. Waugh, R. J.,Morrow, J. D., Roberts, L. J., II, andMurphy, R. C. (1997) Free Radic. Biol.
Med. 23, 943–95428. Roberts, L. J., II,Montine, T. J.,Markesbery,W. R., Tapper, A. R., Hardy, P., Chemtob,
S., Dettbarn, W. D., and Morrow, J. D. (1998) J. Biol. Chem. 273, 13605–1361229. Morrison, W. R., and Smith, L. M. (1964) J. Lipid Res. 53, 600–60830. Kerwin, J. L., and Torvik, J. J. (1996) Anal. Biochem. 237, 56–6431. Kerwin, J. L., Wiens, A. M., and Ericsson, L. H. (1996) J. Mass Spectrom. 31, 184–19232. Morrow, J. D., and Roberts, L. J. (1997) Prog. Lipid Res. 36, 1–2133. Uauy, R., Mena, P., and Valenzuela, A. (1999) Eur. J. Clin. Nutr. 53, Suppl. 1, S66–S7734. Calder, P. C. (2001) Lipids 36, 1007–102435. Yang, P., Chan, D., Felix, E., Cartwright, C., Menter, D. G., Madden, T., Klein, R. D.,
Fischer, S. M., and Newman, R. A. (2004) J. Lipid Res. 45, 1030–103936. Vallve, J. C., Uliaque, K., Girona, J., Cabre, A., Ribalta, J., Heras, M., and Masana, L.
(2002) Atherosclerosis 164, 45–5637. Das, U. N. (1999) Prostaglandins Leukotrienes Essent. Fatty Acids 61, 157–16338. Arita, K., Yamamoto, Y., Takehara, Y., Utsumi, T., Kanno, T., Miyaguchi, C.,
Akiyama, J., Yoshioka, T., and Utsumi, K. (2003) Free Radic. Biol. Med. 35, 189–19939. Kiserud, C. E., Kierulf, P., and Hostmark, A. T. (1995) Thromb. Res. 80, 75–8340. Kokura, S., Nakagawa, S., Hara, T., Boku, Y., Naito, Y., Yoshida, N., and Yoshikawa, T.
(2002) Cancer Lett. 185, 139–14441. Yin, H., Morrow, J. D., and Porter, N. A. (2004) J. Biol. Chem. 279, 3766–3776
Characterization of Eicosapentaenoic Acid Eicosanoids
MAY 19, 2006 • VOLUME 281 • NUMBER 20 JOURNAL OF BIOLOGICAL CHEMISTRY 14099




















