
Emergence of Smooth Muscle Cell Endothelin B-Mediated Vasoconstriction in Lambs With Experimental...
10
ISSN: 1524-4539 Copyright © 2003 American Heart Association. All rights reserved. Print ISSN: 0009-7322. Online 72514 Circulation is published by the American Heart Association. 7272 Greenville Avenue, Dallas, TX DOI: 10.1161/01.CIR.0000087596.01416.2F 2003;108;1646-1654; originally published online Sep 8, 2003; Circulation Johengen and Jeffrey R. Fineman Gerrets, Karen Hendricks-Munoz, Gregory A. Ross, Janine M. Bekker, Michael J. Robert K. Fitzgerald, Olaf Reinhartz, Stefan Thelitz, Robin H. Steinhorn, Rene Stephen M. Black, Eugenia Mata-Greenwood, Robert W. Dettman, Boaz Ovadia, Blood Flow Lambs With Experimental Congenital Heart Disease and Increased Pulmonary Emergence of Smooth Muscle Cell Endothelin B–Mediated Vasoconstriction in http://circ.ahajournals.org/cgi/content/full/108/13/1646 located on the World Wide Web at: The online version of this article, along with updated information and services, is http://www.lww.com/reprints Reprints: Information about reprints can be found online at [email protected] 410-528-8550. E-mail: Fax: Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters http://circ.ahajournals.org/subscriptions/ Subscriptions: Information about subscribing to Circulation is online at by on June 11, 2007 circ.ahajournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Emergence of Smooth Muscle Cell Endothelin B-Mediated Vasoconstriction in Lambs With Experimental...

ISSN: 1524-4539 Copyright © 2003 American Heart Association. All rights reserved. Print ISSN: 0009-7322. Online
72514Circulation is published by the American Heart Association. 7272 Greenville Avenue, Dallas, TX
DOI: 10.1161/01.CIR.0000087596.01416.2F 2003;108;1646-1654; originally published online Sep 8, 2003; Circulation
Johengen and Jeffrey R. Fineman Gerrets, Karen Hendricks-Munoz, Gregory A. Ross, Janine M. Bekker, Michael J.
Robert K. Fitzgerald, Olaf Reinhartz, Stefan Thelitz, Robin H. Steinhorn, Rene Stephen M. Black, Eugenia Mata-Greenwood, Robert W. Dettman, Boaz Ovadia,
Blood FlowLambs With Experimental Congenital Heart Disease and Increased Pulmonary Emergence of Smooth Muscle Cell Endothelin B–Mediated Vasoconstriction in
http://circ.ahajournals.org/cgi/content/full/108/13/1646located on the World Wide Web at:
The online version of this article, along with updated information and services, is
http://www.lww.com/reprintsReprints: Information about reprints can be found online at
[email protected]. E-mail:
Fax:Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters
http://circ.ahajournals.org/subscriptions/Subscriptions: Information about subscribing to Circulation is online at
by on June 11, 2007 circ.ahajournals.orgDownloaded from

Emergence of Smooth Muscle Cell Endothelin B–MediatedVasoconstriction in Lambs With Experimental Congenital
Heart Disease and Increased Pulmonary Blood FlowStephen M. Black, PhD; Eugenia Mata-Greenwood, PhD; Robert W. Dettman, PhD; Boaz Ovadia, MD;
Robert K. Fitzgerald, MD; Olaf Reinhartz, MD; Stefan Thelitz, MD; Robin H. Steinhorn, MD;Rene Gerrets, PhD; Karen Hendricks-Munoz, MD; Gregory A. Ross, MD; Janine M. Bekker, BS;
Michael J. Johengen, BS; Jeffrey R. Fineman, MD
Background—Endothelin-1 (ET-1) has been implicated in the pathophysiology of pulmonary hypertension. In 1-month-old lambs with increased pulmonary blood flow, we have demonstrated early alterations in the ET-1 cascade. Theobjective of this study was to investigate the role of potential later alterations of the ET cascade in the pathophysiologyof pulmonary hypertension secondary to increased pulmonary blood flow.
Methods and Results—Eighteen fetal lambs underwent in utero placement of an aortopulmonary vascular graft (shunt) andwere studied 8 weeks after spontaneous delivery. Compared with age-matched control lambs, lung tissue ET-1 levelswere increased in shunt lambs (317.2�113.8 versus 209.8�61.8 pg/g, P�0.05). In shunt lambs (n�9), exogenous ET-1induced potent pulmonary vasoconstriction, which was blocked by the ETA receptor antagonist PD 156707 (n�3). Thispulmonary vasoconstriction was mimicked by exogenous Ala1,3,11,15 ET-1 (4 Ala ET-1), the ETB receptor agonist, andwas blocked by the ETB receptor antagonist BQ 788 (n�3). However, in control lambs (n�7), ET-1 and 4 Ala ET-1did not change pulmonary vascular tone. In contrast to 4-week-old shunt lambs, immunohistochemistry revealed theemergence of ETB receptors on smooth muscle cells in the vasculature of 8-week-old shunt lambs.
Conclusions—Over time, increased pulmonary blood flow and/or pressure results in the emergence of ETB-mediatedvasoconstriction, which coincides with the emergence of ETB receptors on smooth muscle cells. These data suggest animportant role for ETB receptors in the pathophysiology of pulmonary hypertension in this animal model of increasedpulmonary blood flow. (Circulation. 2003;108:1646-1654.)
Key Words: endothelin � pulmonary heart disease � heart defects, congenital
The development of pulmonary hypertension and its asso-ciated increased vascular reactivity is a common accom-
paniment of congenital heart disease with increased pulmo-nary blood flow.1 Recent evidence suggests that pulmonaryvascular tone and vascular smooth muscle cell proliferation isregulated by a complex interaction of vasoactive substancesthat are produced locally by the vascular endothelium, such asnitric oxide (NO) and endothelin-1 (ET-1).2,3 Endothelialinjury secondary to increased pulmonary blood flow and/orpressure disrupts these regulatory mechanisms and is apotential factor in the development of pulmonaryhypertension.4,5
ET-1 is a 21-amino-acid polypeptide produced by vascularendothelial cells that has potent vasoactive properties and ismitogenic for vascular smooth muscle cells.3,6–10 Several
studies demonstrate increased ET-1 concentrations in chil-dren with increased pulmonary blood flow and pulmonaryhypertension, suggesting a role for ET-1 in the pathophysi-ology of pulmonary hypertension.10,11 To better define therole of ET-1 in the pathogenesis of pulmonary hypertension,we established a model of pulmonary hypertension withincreased pulmonary blood flow in the lamb after in uteroplacement of an aorta-to–pulmonary vein graft. At 1 month ofage, these lambs have a pulmonary-to-systemic blood flowratio of �2:1, a mean pulmonary arterial pressure that is 50%to 75% of mean systemic arterial pressure, and pulmonaryvascular remodeling characteristic of children with pulmo-nary hypertension and increased pulmonary blood flow.12
Previously, we demonstrated that these lambs display alter-ations in the ET-1 cascade at 4 weeks of age. These include
Received June 19, 2002; de novo received November 11, 2002; revision received May 15, 2003; accepted May 19, 2003.From the Department of Pediatrics (S.M.B, E.M.-G., R.W.D., R.H.S), Northwestern University, Chicago, Ill; Departments of Pediatrics (B.O., R.K.F.,
G.A.R., J.M.B., S.T., M.J.J., J.R.F.) and Cardiothoracic Surgery (O.R.), University of California San Francisco, San Francisco, Calif; Department ofPediatrics (R.G., K.H.-M.), New York University, New York, NY; and the Cardiovascular Research Institute (J.R.F.), University of California, SanFrancisco.
Guest editor for this article was Jeffrey A. Towbin, MD, Baylor College of Medicine, Houston, Tex.Correspondence to Jeffrey R. Fineman, MD, University of California, San Francisco, 505 Parnassus Ave, Box 0106, M-680, San Francisco, CA
94143-0106.© 2003 American Heart Association, Inc.
Circulation is available at http://www.circulationaha.org DOI: 10.1161/01.CIR.0000087596.01416.2F
1646 by on June 11, 2007 circ.ahajournals.orgDownloaded from

increased plasma ET-1 levels, decreased ETB receptor proteinwith loss of ETB receptor–mediated vasodilation, and in-creased ETA receptor protein with augmentation of ETA
receptor–mediated vasoconstriction.13,14
The objective of the present study was to determinepotential later alterations of the ET-1 cascade after exposureto increased pulmonary blood flow and pressure. Therefore,we studied the hemodynamic effects of intrapulmonary in-jections of ET-1, Ala1,3,11,15 ET-1 (4 Ala ET-1) (an ETB
receptor agonist), PD 156707 (an ETA receptor antagonist),and BQ 788 (an ETB receptor antagonist) in 8-week-oldlambs with increased pulmonary blood flow and comparedthem with their effects in age-matched controls. In addition,we determined and compared lung tissue ET-1 concentrationsand prepro-ET-1, endothelin-converting enzyme-1 (ECE-1),ETA receptor, and ETB receptor protein levels by Western blotanalysis and mRNA levels by RNAse protection assays.Immunohistochemistry was also performed to localize ETA
and ETB receptors. Last, both protein determinations andlocalization studies were compared with those in 4-week-oldlambs.
MethodsSurgical Preparation and Care
EwesEighteen mixed-breed Western pregnant ewes (135 to 141 days ofgestation; term�145 days) were operated on under sterile conditionsas previously described.12–14 Through a left lateral fetal thoracotomy,an 8.0-mm Gore-tex vascular graft (�2 mm long) (W.L. Gore andAssociates) was anastomosed between the ascending aorta and mainpulmonary artery of the fetus with 7.0 proline (Ethicon Inc) with acontinuous-suture technique.
LambsEight weeks after spontaneous delivery, 35 lambs (18 shunted and 17age-matched controls) were anesthetized with ketamine hydrochlo-ride, diazepam, and fentanyl citrate, mechanically ventilated, andinstrumented to measure vascular pressures and flow as previouslydescribed12–14
Twenty-three lambs (12 shunted and 11 age-matched controls;Pozzi Livestock, Valley Ford, Calif) then underwent a hemodynamicstudy as described below. After the last protocol, the lambs werekilled by an intravenous injection of pentobarbital sodium (Eutha-nasia CII, Central City Medical), followed by bilateral thoracotomy.Twelve of the lambs (6 shunted and 6 twin controls) were killedwithout undergoing any hemodynamic study. These lungs wereremoved and prepared for tissue ET-1 levels and Western blotanalysis, RNAse protection assays, and immunohistochemistry.
All protocols and procedures were approved by the Committee onAnimal Research of the University of California, San Francisco. Allanimals were euthanized by appropriate methods as described in theNational Institutes of Health Guidelines for the Care and Use ofLaboratory Animals.
Experimental Protocols
In Vivo ET-1–Dependent ResponsesThe responses to ET-1 and 4 Ala ET-1 were determined in controllambs at rest and shunt lambs at rest with the vascular graft open.However, the responses to vasodilating agents may be dependent onthe resting tone of the vascular bed studied.15 Therefore, to ensurethat response differences were not tone dependent, the vasodilatorresponses in control lambs were also studied with increased toneduring an intravenous infusion of U46619 (a thromboxane A2
mimic), and shunted lambs were also studied after the vascular graftwas closed, when pulmonary blood flow is similar to that of controls.
Control lambs: After a 45-minute recovery period from surgery,baseline measurements of the hemodynamic variables were mea-sured. In 7 lambs, ET-1 (250 ng/kg) or 4 Ala ET-1 (1725 ng/kg) wasthen injected into the pulmonary artery in random order. Todetermine the selectivity of the agonists and the contribution of eachreceptor to basal tone, the responses to ET-1 and 4 Ala ET-1 werestudied before and during the infusion of PD 156707 (1.0 mg · kg�1
· h�1; an ETA receptor antagonist) and BQ 788 (1.0 �g · kg�1 · min�1;an ETB receptor antagonist) in 4 additional lambs. After the lambshad recovered from the last agent, an infusion of U46619 (athromboxane A2 mimic) was then begun into the inferior vena cava.After 15 minutes of steady-state pulmonary hypertension, baselinemeasurements were again obtained, ET-1 and 4 Ala ET-1 wereadministered, and the hemodynamic variables were measured asdescribed above.
Shunted lambs: After a 45-minute recovery period from surgery,baseline measurements of the hemodynamic variables were mea-sured. In 9 shunt lambs, ET-1 and 4 Ala ET-1 were then administeredas described above. In 3 additional lambs, the responses to ET-1 and4 Ala ET-1 were studied before and during the infusion of PD156707 and BQ 788 as described above. The vascular graft was thenclosed. After a 60-minute recovery, the responses to ET-1 and 4 AlaET-1 were repeated.
ET-1 DeterminationsLung tissues were homogenized in 1 mol/L acetic acid containing 10�g/mL pepstatin (Peptide International) and immediately boiled for10 minutes. The homogenates were centrifuged at 25 000g for 30minutes at 4°C, and the supernates were stored at �30°C beforebeing assayed for immunoreactive endothelin as previouslydescribed.14
Tissue PreparationThe heart and lungs were removed en bloc. The lungs were dissectedwith care to preserve the integrity of the vascular endothelium.Sections (2 to 3 g) from each lobe of the lung were removed. Thesetissues were snap-frozen in liquid N2 and stored at �70°C untilanalysis.
For RNA and protein isolation, the snap-frozen lung tissue wasprepared as previously described.16 For immunohistochemistry, thepulmonary vascular tree was rinsed with cold (4°C) PBS to removeblood and fixed by perfusion with cold (4°C) 4% paraformaldehyde.The pulmonary artery was then clamped. The airways were fixed at20 cm H2O pressure by filling the trachea with cold (4°C) 4%
General Hemodynamics
Control Lambs Shunted Lambs
Age, d 63.1�5.9 61.2�6.9
Weight, kg 22.6�4.2 18.9�4.9*
PAP, mm Hg 17.6�4.6 26.5�10.6*
SAP, mm Hg 81.1�13.8 68.3�16.2*
PAP/SAP 0.23�0.09 0.41�0.19*
Left pulmonary vascular resistance,mm Hg � mL�1 � min�1 � kg�1
0.283�0.09 0.141�0.05*
Left pulmonary blood flow,mL � kg�1 � min�1
35.4�4.8 121.7�40.0*
Heart rate, bpm 136.9�22.5 153.9�22.3
Left atrial pressure, mm Hg 7.7�3.0 10.4�4.9*
Right atrial pressure, mm Hg 7.4�2.9 5.4�2.5
Qp/Qs 1.00 3.0�0.9*
PAP indicates pulmonary arterial pressure; SAP, systemic arterial pressure.Values are mean�SD. n�15 shunt lambs and 13 age-matched controls.Values in shunted lambs were obtained before closure of the vascular graft.
*P�0.05 vs control lambs.
Black et al ETB Receptors in Pulmonary Hypertension 1647
by on June 11, 2007 circ.ahajournals.orgDownloaded from

paraformaldehyde. When the lungs were distended at this pressure,the trachea was clamped. The lungs were fixed for 24 hours at 4°Cby immersion in 4% paraformaldehyde. Representative slices fromeach lobe were removed, placed in 30% sucrose until they sank,placed in OCT, frozen on dry ice, and stored at �70°C untilsectioned. Sections (5 to 10 �m) were cut with a cryostat, transferredto aminoalkylsilane-treated slides (Superfrost Plus, Fisher Scientif-ic), and stored at �70°C.16
Western Blot AnalysisWestern blot analysis was performed as previously described.14,17
The ETA receptor antiserum was generated as previously described.14
The ETB receptor antiserum was obtained from Maine BiotechnologyServices. The prepro-ET-1 antibody was obtained from AffinityBioreagents. The specificity of the prepro-ET-1 antibody was veri-fied with a preincubation step with purified ET-1 (50 ng ET-1/15 �Lof antiserum) protein. The purified ET-1 was purchased from Sigma.ECE-1� antiserum was generated as previously described.16
Positive controls were run to demonstrate antibody specificity.The methodology and exposure times used were those that we havepreviously demonstrated to be within the linear range of theautoradiographic film and able to detect changes in lung proteinexpression.
To compare changes in ETB receptor protein levels between 4 and8 weeks, protein levels from an additional five 4-week-old controllambs and five 4-week-old shunt lambs were analyzed simulta-neously with those of 8-week-old lambs.
RNA Probe Synthesis and RNAse Protection AssayThe plasmid containing the cDNA fragment of interest was linear-ized with the appropriate restriction enzyme (Gibco-BRL). Antisense[32P]UTP radiolabeled cRNA probes (New England Nuclear) weresynthesized by in vitro transcription using either T3 or T7 RNApolymerases (Boehringer-Mannheim) in the presence of cold rCTP,rGTP, and rATP.17
RNAse protection assays were performed as previously de-scribed.17 Also included was a probe for 18S to serve as a control forthe amount of input total RNA and the recovery of protected probefragments.
ImmunohistochemistryImmunohistochemistry was performed as previously described.16
Studies were done on serial sections of control and shunt ovine lungwith our rabbit anti-ETA antibody raised as described previously.14 Inaddition, immunohistochemistry was performed with a specificrabbit antiserum raised against ETB. This antiserum was prepared byinjecting rabbits with a highly antigenic protein fragment based onrat ETB (sequence: NH2-SCLKFKANDHGYDNF-COOH). Rabbitswere bled at 6, 8, and 10 weeks, and the 8-week bleed wasimmunopurified (BioSynthesis Inc). Frozen tissue sections (7 �m)were allowed to come to room temperature. Samples were fixed for10 minutes in cold acetone, then washed 3 times with PBS. Toeliminate nonspecific binding of the primary antiserum to tissueproteins, tissue sections were incubated with 1% horse serum in PBS(blocking solution) for 1 hour. Then, tissue sections were incubatedwith anti-ETA (1:100) or anti-ETB (1:100), both in the presence ofmonoclonal smooth muscle cell–actin antibody (1:400, Sigma) bothin blocking solution at 4°C overnight. After 3 washes with PBS for5 minutes, samples were hybridized with Rhodamine Red-X goatanti-rabbit (to stain ET receptors) and Oregon Green 488 goatanti-mouse secondary antibodies (to stain smooth muscle cell–actin)(Molecular Probes) at a concentration of 1:400 in blocking solutionfor 45 minutes at room temperature. After 3 further washes withPBS, an antifading solution was added, and samples were visualizedby confocal microscopy. A minimum of 3 different sets of controland shunt lung tissue were prepared and examined.
Lung sections from four 4-week-old shunt and control lambs werealso analyzed to localize ETA and ETB receptors.
Statistical AnalysisThe mean�SD was calculated for the hemodynamic variables,systemic arterial blood gases, and pH and tissue ET-1 levels.Comparisons were made by the paired t test using the Bonferronicorrection, the unpaired t test, or ANOVA for repeated measureswith multiple-comparison testing.
Quantification of autoradiographic results was performed byscanning the bands of interest into an image editing softwareprogram (Adobe Photoshop, Adobe Systems). For RNAse protectionassays, to control for the amount of input RNA and the recovery ofprotected probe fragments, the mRNA signal of interest was normal-ized to the corresponding 18S signal for each lane. The mean�SDwas calculated for the relative RNA and protein. The unpaired t testor ANOVA was used for comparisons between 4- and 8-week-old,control and shunt lambs for repeated measures with multiple-comparison testing. A value of P�0.05 was considered statisticallysignificant.
ResultsAt 2 months of age, shunted lambs weighed significantly lessthan controls (18.9�4.9 versus 22.6�4.2 kg, P�0.05). Theratio of pulmonary to systemic blood flow (Qp/Qs) was3.0�0.9. Mean pulmonary arterial pressure was increased to41% of systemic values. This was associated with an increasein pulmonary blood flow and left atrial pressure (P�0.05).Mean systemic arterial pressure and the calculated left pul-monary vascular resistance were decreased (P�0.05) (Table).Peripheral lung tissue immunoreactive ET-1 concentrationswere greater in shunted lambs than control lambs(317.2�113.8 versus 209.8�61.8 pg/g, P�0.05).
The baseline hemodynamics and protein concentrations ofthe 4-week-old lambs used in this study were similar to thosepreviously published (data not shown).13,14 In addition, the
Figure 1. Intrapulmonary injection of both ET-1 (left) and 4 AlaET-1 (right) increase mean pulmonary arterial pressure (MPAP;top) and left pulmonary vascular resistance (LPVR; bottom) inshunted lambs with graft open but does not change these pa-rameters in control lambs at rest. Values are mean�SEM; n�9shunt and n�7 control lambs. *P�0.05 vs control lambs.
1648 Circulation September 30, 2003
by on June 11, 2007 circ.ahajournals.orgDownloaded from

baseline hemodynamics of the 4- and 8-week-old shunt lambsused in this study were similar.
Hemodynamic Study
Control LambsIn control lambs, the intrapulmonary injection of ET-1 and 4Ala ET-1 did not change mean pulmonary arterial pressure,left pulmonary blood flow, or left pulmonary vascular resis-tance (Figure 1). ET-1 increased mean systemic arterialpressure from 67.5�7.7 to 90.6�7.8 mm Hg (P�0.05).Similarly, during steady-state pulmonary hypertension in-duced by the infusion of U46619, intrapulmonary injectionsof ET-1 or 4 Ala ET-1 did not change mean pulmonaryarterial pressure or pulmonary vascular resistance (data notshown).
At rest, the infusion of PD 156707 induced a modestdecrease in mean pulmonary arterial pressure (�8.0�0.6%,P�0.05) and left pulmonary vascular resistance(�9.9�2.6%, P�0.05) (n�4). Pulmonary blood flow wasunchanged. During the infusion of PD 156707, the increase insystemic arterial pressure induced by ET-1 was blocked, andthe response to 4 Ala ET-1 was unchanged (data not shown).The infusion of BQ 788 did not change the baseline hemo-dynamic variables (n�4). In addition, during the infusion ofBQ 788, the response to ET-1 and 4 Ala ET-1 was unchanged(data not shown).
Shunted LambsIn shunted lambs with the vascular graft open, the intrapul-monary injection of ET-1 increased mean pulmonary
(25.1�8.3 to 35.2�11.8 mm Hg) and systemic (63.7�12.2 to73.3�14.2) arterial pressure and left pulmonary vascularresistance (0.13�0.04 to 0.23�0.011 mm Hg · L�1 · min�1)(P�0.05). Left pulmonary blood flow was unchanged. Sim-ilarly, the intrapulmonary injection of 4 Ala ET-1 increasedmean pulmonary (25.0�9.7 to 32.3�12.0 mm Hg) and sys-temic (66.8�12.4 to 73.6�13.1 mm Hg) arterial pressure andleft pulmonary vascular resistance (0.13�0.07 to0.22�0.16 mm Hg · L�1 · min�1) (P�0.05). Left pulmonaryblood flow decreased (132.7�41.3 to 122.1�43.7mL/min)(P�0.05).
Similarly, after shunt closure, the intrapulmonary injec-tions of ET-1 and 4 Ala ET-1 induced similar increases inmean pulmonary arterial pressure and left pulmonary vascu-lar resistance (data not shown) (P�0.05).
The percent increase in both left pulmonary vascularresistance and mean pulmonary arterial pressure induced bythe injection of ET-1 was greater in shunted lambs than incontrol lambs (P�0.05) (Figure 1). Similarly, the percentincrease in both left pulmonary vascular resistance and meanpulmonary arterial pressure induced by the injection of 4 AlaET-1 was greater in shunted lambs than in control lambs(P�0.05) (Figure 1).
The infusion of PD 156707 decreased mean pulmonaryarterial pressure (�15.4�3.7%, P�0.05) and left pulmonaryvascular resistance (�25.0�3.9%, P�0.05) (n�3). Pulmo-nary blood flow was unchanged. During the infusion of PD156707, the increase in left pulmonary vascular resistanceinduced by ET-1 was blocked (90.1�35.2% versus
Figure 2. Western blot analysis for prepro-ET-1 (left) and ECE-1� (right) in lung tissue from 8-week-old lambs. Top, Protein extracts (50�g) prepared from lung tissue from four 8-week-old lambs (2 control and 2 shunt). Also included is a positive (�ve) control consistingof a protein extract prepared from COS-7 cell transiently transfected with a mammalian expression vector containing full-length bovineECE-1� cDNA. Bottom, Densitometric values for relative prepro-ET-1 protein (left) and ECE-1� (right) (normalized to control) from 5control and 5 shunt lambs. In shunt lambs, relative prepro-ET-1 and ECE-1� protein was not significantly different from that in controllambs. Values are mean�SEM.
Black et al ETB Receptors in Pulmonary Hypertension 1649
by on June 11, 2007 circ.ahajournals.orgDownloaded from

�7.9�18.5%), but the response to 4 Ala ET-1 was un-changed. The infusion of BQ 788 did not change the baselinehemodynamic variables. However, during the infusion of BQ788, the increase in left pulmonary vascular resistance inresponse to ET-1 was partially attenuated (90.1�35.2%versus 19.2�16.5%), whereas the response to 4 Ala ET-1 wascompletely blocked (107.5�22.0% versus 1.9�5.7%) (n�3).
To determine whether the alterations in ET-1 levels andphysiological responses were associated with changes in geneexpression, we performed Western blot analysis and RNAseprotection assays. Compared with age-matched controllambs, the levels of both prepro-ET-1 and ECE-1� proteinand mRNA were unchanged (Figures 2 and 5). Protein levelsof ETA receptors were increased in shunt lambs (P�0.05),whereas the changes in ETA receptor mRNA did not reachstatistical significance (Figures 3 and 5). Protein and mRNAlevels of ETB receptors were also unchanged in shunt lambscompared with age-matched controls (Figures 3 and 5).However, compared with 4-week-old shunt lambs, proteinlevels of ETB receptor were significantly increased in 8-week-old shunt lambs (Figure 4).
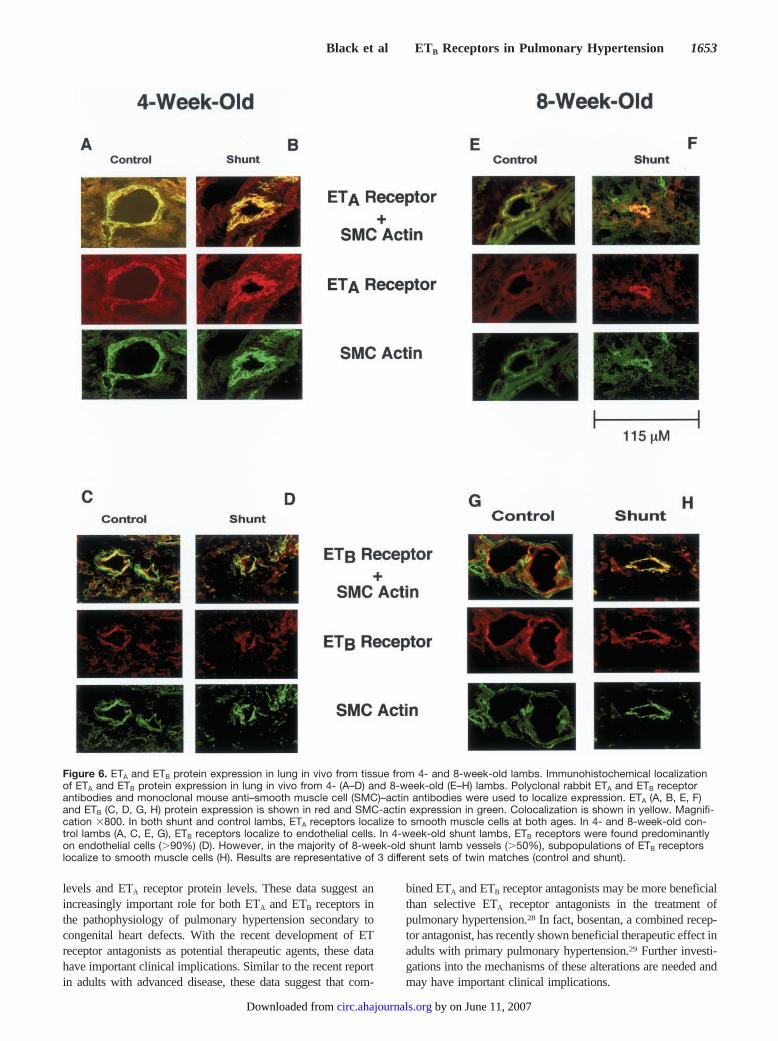
In both 4- and 8-week-old control lambs, localization byimmunohistochemistry demonstrated ETB receptors solely onvascular endothelial cells (Figure 6C and 6G) and ETA
receptors solely in the vascular smooth muscle cell layer(Figure 6A and 6E). In 4- and 8-week-old shunt lambs ETA
receptors were again found solely in the vascular smoothmuscle cell layer (Figure 6B and 6F). In 4-week-old shuntlambs, ETB receptors were found predominantly on endothe-lial cells (�90%) and occasionally on smooth muscle cells.However, in 8-week-old shunt lambs, a significant subpopu-lation of ETB receptors were localized to the vascular smoothmuscle cell layer (�50%) (Figure 6D and 6H).
DiscussionIn a follow-up investigation to our previous studies, we foundthat exposure to 8 weeks of increased pulmonary blood flowand pressure results in progressive alterations in ET-1 signal-ing. Similar to alterations displayed at 4 weeks, shuntedlambs had increased lung tissue concentrations of ET-1 andincreased protein levels of ETA receptors. However, incontrast to decreased ETB protein levels and a loss of ETB
receptor–mediated pulmonary vasodilation in 4-week-oldshunt lambs, 8-week-old shunt lambs had an upregulation ofETB receptor protein and the emergence of ETB receptor–mediated pulmonary vasoconstriction.13,14 Immunohisto-chemistry revealed that both 4- and 8-week-old control lambshad ETB receptors solely on endothelial cells, whereas8-week-old shunt lambs displayed a subpopulation of ETB
receptors present in the vascular smooth muscle cell layer.ETA receptors were found on vascular smooth muscle cells inall groups. These data suggest a potentially important role for
Figure 3. Western blot analysis for ETA receptor (left) and ETB receptor (right) in lung tissue from 8-week-old lambs. Top, Proteinextracts (50 �g) prepared from lung tissue from 8-week-old lambs. Bottom, Densitometric values for relative ETA receptor protein (left)and relative ETB receptor protein (right) (normalized to control) from 5 control and 5 shunt lambs. In shunt lambs, relative ETA receptorprotein expression was increased, and relative ETB receptor protein was not significantly different from control lambs. Values aremean�SEM. *P�0.05 vs control.
1650 Circulation September 30, 2003
by on June 11, 2007 circ.ahajournals.orgDownloaded from

smooth muscle cell ETB receptors in the pathophysiology ofpulmonary hypertension secondary to congenital heartdisease.
The hemodynamic effects of ET-1 are mediated by at least2 populations of receptors, ETA and ETB receptors. In thesystemic vasculature, the response to exogenous ET-1 isconsistently vasoconstricting, which is mediated via ETA
receptor activation.18 However, in the pulmonary vasculature,the response to exogenous ET-1 is dependent on a variety offactors, including age, species, and the underlying diseasestate.13,15,19 In the fetal and young sheep, ET-1 inducespulmonary vasodilation, which is mediated via ETB receptoractivation and the release of NO. In the adult sheep, ET-1induces pulmonary vasoconstriction, which is associated withprostaglandin release and presumed ETA receptor activa-tion.15,19 In the rabbit, rat, and human adult pulmonarycirculation, pulmonary vasoconstricting ETB receptors havebeen identified in the medial layer of pulmonary arteries.20–22
However, in response to modest doses of ET-1, ETB receptor–induced pulmonary vasoconstriction has not been reportedpreviously in the young sheep or human. With the use of 4Ala ET-1, a selective ETB receptor agonist, we demonstratesignificant pulmonary vasoconstriction in 8-week-old shuntlambs and no response in age-matched control lambs. In
addition, the infusion of BQ 788 (an ETB receptor antagonist)blocked the vasoconstriction induced by 4 Ala ET-1, whereasPD 156707 (an ETA receptor antagonist) did not, furthersuggesting that 4 Ala ET-1–induced pulmonary constrictionwas secondary to ETB receptor activation. Interestingly, theinfusion of BQ 788 did not change resting pulmonaryvascular tone in either control or shunt lambs, suggesting thatETB receptor activation participates in agonist-induced re-sponses but makes a minimal contribution to basal tone.Conversely, the increase in ETA receptor protein noted inshunt lambs was associated with an augmented pulmonaryvasodilating response to PD 156707, the ETA receptor antag-onist, suggesting that ETA receptor activation contributes tobasal tone in shunt lambs.
In association with the emergence of ETB receptor–medi-ated pulmonary vasoconstriction, there were changes inprotein levels of ETB receptors. Previously, in 4-week-oldshunt lambs, we demonstrated a decrease in ETB receptorlung protein levels compared with age-matched controllambs.14 This was associated with loss of the pulmonaryvasodilating response to the ETB receptor agonist 4 AlaET-1.13 In the present study, we demonstrate no difference inlung ETB receptor protein between 8-week-old shunt lambsand age-matched control lambs. However, compared with4-week-old shunt lambs, there was a significant increase inETB receptor protein levels. In addition, immunohistochem-istry reveals that a subset of ETB receptor expression predom-inantly in 8-week-old shunt lambs now localizes to thesmooth muscle cell layer. Taken together, these data suggestthat increased pulmonary blood flow and/or pressure causes adecrease in endothelial ETB receptors and the emergence ofsmooth muscle cell ETB receptors. Because 4 Ala ET-1–induced pulmonary vasoconstriction is now present in8-week-old shunt lambs, we speculate that the increasedprotein levels of ETB receptor at 8 weeks mediate thedemonstrated pulmonary vasoconstricting response. Al-though increasing in vitro data suggest that changes inmechanical forces alter ET-1 signaling, few data are availableon the regulation of ET receptor signaling.23,24 Therefore, thepotential mechanisms for the current receptor changes areunknown and warrant further investigation.
Changes in ETB receptors have previously been demon-strated in other animal models of pulmonary hypertension.For example, in contrast to the present study, rats exposed tomonocrotaline displayed an increase in ETB receptor–medi-ated vasodilation during the development of pulmonaryhypertension.25 In addition, in piglets exposed to hypoxia,there was loss of the normal postnatal increase in endothelialETB receptors during the first 3 days of life, and in sheep withpersistent pulmonary hypertension of the newborn, ETB receptorexpression was decreased.26,27 However, in humans with ad-vanced pulmonary vascular disease secondary to thromboem-bolic pulmonary hypertension, a recent report demonstrates aselective upregulation of ETB mRNA in the media of pulmonaryarteries.28 The present study represents the first investigation ofchanges in ET receptors during the development of pulmonaryhypertension secondary to congenital heart disease with in-creased pulmonary blood flow. Our findings are consistent withthe recent findings in adults and further suggest an important role
Figure 4. Western blot analysis for ETB receptor in lung tissue from4- and 8-week-old lambs. Top, Protein extracts (50 �g) preparedfrom lung tissue from 4 lambs (2 control and 2 shunt). Bottom,Densitometric values for relative ETB receptor protein (normalizedto 4 weeks) from five 4-week-old control and shunt lambs and five8-week-old control and shunt lambs. Four-week-old values wereassigned a value of 1. In control lambs, ETB receptor proteinexpression was decreased at 8 weeks compared with 4 weeks.However, in shunt lambs, ETB receptor protein expression wasincreased at 8 weeks compared with 4 weeks. Values aremean�SEM. *P�0.05 vs corresponding 4-week values.
Black et al ETB Receptors in Pulmonary Hypertension 1651
by on June 11, 2007 circ.ahajournals.orgDownloaded from

for both ETA and ETB receptors in the pathophysiology ofpulmonary hypertensive disorders.
In summary, in a model of increased pulmonary blood flowand pulmonary hypertension secondary to in utero aortopulmo-
nary vascular graft placement, the present study describes theemergence of smooth muscle cell, ETB receptor–mediated pul-monary vasoconstriction over the second month of postnatal life.In addition, there are persistent increases in lung tissue ET-1
Figure 5. RNAse protection assays for prepro-ET-1, ECE-1, ETA receptor, and ETB receptor. A, cRNA probe for ovine prepro-ET-1 washybridized overnight to 50 �g of total lung RNA prepared from two 8-week-old lambs (1 control and 1 shunt). Densitometric determina-tions from 5 control and 5 shunt lambs did not reveal differences (data not shown). B, cRNA probe for ovine ECE-1 was hybridizedovernight to 50 �g of total lung RNA prepared from two 8-week-old lambs (1 control and 1 shunt). Densitometric determinations from 5control and 5 shunt lambs did not reveal differences (data not shown). C, cRNA probe for ovine ETA receptor was hybridized overnightto 50 �g of total lung RNA prepared from two 8-week-old lambs (1 control and 1 shunt). Densitometric determinations from 5 controland 5 shunt lambs did not reveal differences (data not shown). D, cRNA probe for ovine ETB receptor was hybridized overnight to 50�g of total lung RNA prepared from two 8-week-old lambs (1 control and 1 shunt). Densitometric determinations from 5 control and 5shunt lambs did not reveal differences (data not shown). No protected fragments were detected in lanes in which probe was hybridizedwithout RNA (PA) or in presence of tRNA. A cRNA for ovine 18S was also hybridized to serve as a control for RNA loading.
1652 Circulation September 30, 2003
by on June 11, 2007 circ.ahajournals.orgDownloaded from
levels and ETA receptor protein levels. These data suggest anincreasingly important role for both ETA and ETB receptors inthe pathophysiology of pulmonary hypertension secondary tocongenital heart defects. With the recent development of ETreceptor antagonists as potential therapeutic agents, these datahave important clinical implications. Similar to the recent reportin adults with advanced disease, these data suggest that com-
bined ETA and ETB receptor antagonists may be more beneficialthan selective ETA receptor antagonists in the treatment ofpulmonary hypertension.28 In fact, bosentan, a combined recep-tor antagonist, has recently shown beneficial therapeutic effect inadults with primary pulmonary hypertension.29 Further investi-gations into the mechanisms of these alterations are needed andmay have important clinical implications.
Figure 6. ETA and ETB protein expression in lung in vivo from tissue from 4- and 8-week-old lambs. Immunohistochemical localizationof ETA and ETB protein expression in lung in vivo from 4- (A–D) and 8-week-old (E–H) lambs. Polyclonal rabbit ETA and ETB receptorantibodies and monoclonal mouse anti–smooth muscle cell (SMC)–actin antibodies were used to localize expression. ETA (A, B, E, F)and ETB (C, D, G, H) protein expression is shown in red and SMC-actin expression in green. Colocalization is shown in yellow. Magnifi-cation �800. In both shunt and control lambs, ETA receptors localize to smooth muscle cells at both ages. In 4- and 8-week-old con-trol lambs (A, C, E, G), ETB receptors localize to endothelial cells. In 4-week-old shunt lambs, ETB receptors were found predominantlyon endothelial cells (�90%) (D). However, in the majority of 8-week-old shunt lamb vessels (�50%), subpopulations of ETB receptorslocalize to smooth muscle cells (H). Results are representative of 3 different sets of twin matches (control and shunt).
Black et al ETB Receptors in Pulmonary Hypertension 1653
by on June 11, 2007 circ.ahajournals.orgDownloaded from

AcknowledgmentsThis research was supported by grants HL-61284 (Dr Fineman),HL-60190 (Dr Black), HD-398110 (Dr Black), and HL-67841 (DrBlack) from the National Institutes of Health and FY00-98 (DrBlack) from the March of Dimes.
References1. Hoffman JIE, Rudolph AM, Heymann MA. Pulmonary vascular disease
with congenital heart lesions: pathologic features and causes. Circulation.1981;64:873–877.
2. Furchgott RF, Vanhoutte PM. Endothelium-derived relaxing and con-tracting factors. FASEB J. 1989;3:2007–2018.
3. Yanagisawa M, Kurihara H, Kimura S, et al. A novel potent vasocon-strictor peptide produced by vascular endothelial cells. Nature. 1988;332:411–415.
4. Giaid A, Saleh D. Reduced expression of endothelial nitric oxide synthasein the lungs of patients with pulmonary hypertension. N Engl J Med.1995;333:214–221.
5. Giaid A., Yanagisawa M, Langleben D, et al. Expression of endothelin-1in the lungs of patients with pulmonary hypertension. N Engl J Med.1993;328:1732–1739.
6. Hassoun PM, Thappa V, Landman MJ, et al. Endothelin 1: mitogenicactivity on pulmonary artery smooth muscle cells and release fromhypoxic endothelial cells. Proc Exp Biol Med. 1992;199:165–170.
7. Arai H, Hori S, Aramori I, et al. Cloning and expression of a cDNAencoding an endothelin receptor. Nature. 1990;348:730–732.
8. Sakurai T, Yanagisawa M, Takuwa Y, et al. Cloning of a cDNA encodinga non-isopeptide-selective subtype of the endothelin receptor. Nature.1990;348:732–735.
9. Shetty SS, Toshikazu O, Webb RL, et al. Functionally distinct endothelinb receptors in vascular endothelium and smooth muscle. Biochem BiophysRes Commun. 1993;191:459–464.
10. Vincent JA, Ross RD, Kassab J, et al. Relation of elevated plasmaendothelin in congenital heart disease to increased pulmonary blood flow.Am J Cardiol. 1993;71:1204–1207.
11. Yoshibayashi M, Nishioka K, Nakao K, et al. Plasma endothelin concen-trations in patients with pulmonary hypertension associated with con-genital heart disease. Circulation. 1991;84:2280–2285.
12. Reddy VM, Meyrick B, Wong J, et al. In utero placement of aortopul-monary shunts: a model of postnatal pulmonary hypertension withincreased pulmonary blood flow in lambs. Circulation. 1995;92:606–613.
13. Wong J, Reddy VM, Hendricks-Munoz K, et al. Altered endothelin-1vasoactive responses in lambs with pulmonary hypertension andincreased pulmonary blood flow. Am J Physiol. 1995;269:H1965–H1972.
14. Black SM, Bekker J, Johengen M, et al. Altered regulation of the ET-1cascade in lambs with increased pulmonary blood flow and pulmonaryhypertension. Pediatr Res. 2000;47:97–106.
15. Cassin S, Kristova V, Davis T, et al. Tone-dependent responses toendothelin in isolated perfused fetal sheep pulmonary circulation in situ.J Appl Physiol. 1991;70:1228–1234.
16. Black SM, Johengen MJ, Ma ZD, et al. Ventilation and oxygenationinduce endothelial nitric oxide synthase gene expression in the lungs offetal lambs. J Clin Invest. 1997;100:1–11.
17. McMullan DM, Bekker JM, Johengen MJ, et al. Inhaled nitric oxide-induced rebound pulmonary hypertension: a role for endothelin-1. Am JPhysiol. 2001;280:H777–H785.
18. La M, Reid JJ. Endothelin-1 and the regulation of vascular tone. Clin ExpPharmacol Physiol. 1995;22:315–323.
19. Wong J, Vanderford PA, Fineman JR, et al. Developmental effects ofendothelin-1 on the pulmonary circulation in sheep. Pediatr Res. 1994;36:394–401.
20. McCulloch KM, Docherty C, MacLean MR. Endothelin receptorsmediating contraction of rat and human pulmonary resistance arteries:effect of chronic hypoxia in the rat. Br J Pharmacol. 1998;123:1621–1630.
21. McCulloch KM, Docherty CC, Morecroft I, et al. EndothelinB receptor-mediated contraction in human pulmonary resistance arteries. Br JPharmacol. 1996;119:1125–1130.
22. Docherty C, MacLean MR. Development of endothelin receptors inperinatal rabbit pulmonary resistance arteries. Br J Pharmacol. 1998;124:1165–1174.
23. Morawietz H, Talanow R, Szibor M, et al. Regulation of the endothelinsystem by shear stress in human endothelial cells. J Physiol. 2000;525:761–770.
24. Harrison VJ, Ziegler T, Bouzourene K, et al. Endothelin-1 andendothelin-converting enzyme-1 gene regulation by shear stress and flow-induced pressure. J Cardiovasc Pharmacol. 1998;31(suppl 1):S38–S41.
25. Sakai S, Miyauchi T, Hara J, et al. Hypotensive effect of endothelin-1 viaendothelin-B-receptor pathway on pulmonary circulation is enhanced inrats with pulmonary hypertension. J Cardiovasc Pharmacol. 2000;36(suppl 1):S95–S98.
26. Noguchi Y, Hislop AA, Haworth SG. Influence of hypoxia onendothelin-1 binding sites in neonatal porcine pulmonary vasculature.Am J Physiol. 1997;272:H669–H678.
27. Ivy DD, Le Cras TD, Horan MP, et al. Increased lung preproET-1 mRNAand decreased ETB-receptor gene expression in fetal pulmonary hyper-tension. Am J Physiol. 1998;274:L535–L541.
28. Bauer M, Wilkens H, Langer F, et al. Selective upregulation of endothelinB receptor gene expression in severe pulmonary hypertension. Circu-lation. 2002;105:1034–1036.
29. Rubin LJ, Badesch DB, Barst RJ, et al. Bosentan therapy for pulmonaryarterial hypertension. N Engl J Med. 2002;346:896–903.
1654 Circulation September 30, 2003
by on June 11, 2007 circ.ahajournals.orgDownloaded from




















