The effect on motor cortical neuronal development of focal lesions to the sub-cortical white matter...
12
Int. J. Devl Neuroscience 21 (2003) 171–182 The effect on motor cortical neuronal development of focal lesions to the sub-cortical white matter in the neonatal rat: a model for periventricular leukomalacia Claire L. Gibson 1 , Gavin J. Clowry ∗ Brain Development, Plasticity and Repair Group, School of Clinical Medical Sciences (Child Health), University of Newcastle upon Tyne, Newcastle upon Tyne NE1 4LP, UK Received 6 November 2002; received in revised form 12 March 2003; accepted 14 March 2003 Abstract Periventricular leukomalacia (PVL) is either a diffuse or cystic lesion of the periventricular white matter that leaves the overlying cortical grey matter largely intact. It is believed to result from hypoxia occurring pre- or perinatally and is a major cause of cerebral palsy. We have modelled PVL in rats comparing the effects of discrete injections of 3-nitropropionic acid (3-NP), a mitochondrial toxin, ibotenic acid (IBA), a glutamate analogue, or saline into the sub-cortical white matter on postnatal day 7 (P7). Following recovery times ranging from 3 days to 4 weeks, forebrain sections were Nissl stained or immunostained for Bax, cJun, calbindin (CB), parvalbumin (PV) or non-phosphorylated neurofilaments (NPNF). Compared to saline injections, ibotenic acid caused large lesions of both grey and white matter not characteristic of periventricular leukomalacia. 3-Nitropropionic acid injections caused small focal lesions restricted to the sub-cortical white matter. 3-Nitropropionic acid treatment initially increased expression of the apoptosis promoting proteins Bax and cJun, as well as non-phosphorylated neurofilaments in cortical layer V overlying the injection site. Non-phosphorylated neurofilament expression distal to the lesion was decreased representing a loss of cortical axons, but persisted and even increased with time within the cortex, demonstrating persistence of the parent cell bodies and local sprouting of neurites. There were significantly fewer calbindin and parvalbumin positive neurones in the motor cortex (MC) side ipsilateral to the 3-nitropropionic acid injection compared to the contralateral side. These persistent differences in expression of activity sensitive calcium binding proteins suggest alterations in local cortical circuitry without substantial loss of grey matter as is characteristic of periventricular leukomalacia. Changes in expression of Bax, cJun and non-phosphorylated neurofilaments during normal development are also described. © 2003 ISDN. Published by Elsevier Science Ltd. All rights reserved. Keywords: Periventricular leukomalacia (PVL); 3-Nitropropionic acid; Cerebral palsy; Hypoxia; Brain development 1. Introduction Periventricular leukomalacia (PVL) is characterised by a necrotic, and often cystic lesion of the periventricular white matter (Kuban, 1998) and is associated with premature ba- bies. Such a developmental lesion to the white matter results in the child displaying spastic diplegia, as seen in cerebral palsy (Miller et al., 2000) whereby spasticity is present in all four limbs but most severely in the lower limbs. The in- cidence of PVL is 0.2 per 1000 live births (Miller et al., ∗ Corresponding author. Present address: Sir James Spence Institute for Child Health, Royal Victoria Infirmary, Newcastle upon Tyne NE1 4LP, UK. Tel.: +44-191-202-3008; fax: +44-191-202-3022. E-mail address: [email protected] (G.J. Clowry). 1 Present address: Institute of Cell Signalling, Queens Medical Centre, University of Nottingham, Nottingham NG7 2UH, UK. 2000) and the risk of PVL is greatly increased with decreas- ing gestational age (Back and Volpe, 1997). The prevalence of cerebral palsy caused by PVL is increasing in the de- veloped world as a consequence of increased survival of preterm babies (Johnston, 1998) and many factors underly- ing the pathology of PVL remain unknown. The pathological features of PVL include focal periven- tricular as well as more diffuse cerebral white matter dam- age to the white matter dorsal and lateral to the lateral ventricle where the corticospinal tract is located (Paneth et al., 1990). However, areas of grey matter surrounding lesion areas are largely spared. This results in disruption of corticospinal axons, while the cortical projection neurones are left intact and subsequently make aberrant intracortical axonal projections (Marin-Padilla, 1997). The typical his- tological features of PVL are characterised by coagulation necrosis, microglial infiltration, astrocytic proliferation and 0736-5748/03/$30.00 © 2003 ISDN. Published by Elsevier Science Ltd. All rights reserved. doi:10.1016/S0736-5748(03)00041-8
Transcript of The effect on motor cortical neuronal development of focal lesions to the sub-cortical white matter...

Int. J. Devl Neuroscience 21 (2003) 171–182
The effect on motor cortical neuronal development of focal lesionsto the sub-cortical white matter in the neonatal rat:
a model for periventricular leukomalacia
Claire L. Gibson1, Gavin J. Clowry∗Brain Development, Plasticity and Repair Group, School of Clinical Medical Sciences (Child Health),
University of Newcastle upon Tyne, Newcastle upon Tyne NE1 4LP, UK
Received 6 November 2002; received in revised form 12 March 2003; accepted 14 March 2003
Abstract
Periventricular leukomalacia (PVL) is either a diffuse or cystic lesion of the periventricular white matter that leaves the overlyingcortical grey matter largely intact. It is believed to result from hypoxia occurring pre- or perinatally and is a major cause of cerebralpalsy. We have modelled PVL in rats comparing the effects of discrete injections of 3-nitropropionic acid (3-NP), a mitochondrial toxin,ibotenic acid (IBA), a glutamate analogue, or saline into the sub-cortical white matter on postnatal day 7 (P7). Following recovery timesranging from 3 days to 4 weeks, forebrain sections were Nissl stained or immunostained for Bax, cJun, calbindin (CB), parvalbumin (PV) ornon-phosphorylated neurofilaments (NPNF). Compared to saline injections, ibotenic acid caused large lesions of both grey and white matternot characteristic of periventricular leukomalacia. 3-Nitropropionic acid injections caused small focal lesions restricted to the sub-corticalwhite matter. 3-Nitropropionic acid treatment initially increased expression of the apoptosis promoting proteins Bax and cJun, as well asnon-phosphorylated neurofilaments in cortical layer V overlying the injection site. Non-phosphorylated neurofilament expression distal tothe lesion was decreased representing a loss of cortical axons, but persisted and even increased with time within the cortex, demonstratingpersistence of the parent cell bodies and local sprouting of neurites. There were significantly fewer calbindin and parvalbumin positiveneurones in the motor cortex (MC) side ipsilateral to the 3-nitropropionic acid injection compared to the contralateral side. These persistentdifferences in expression of activity sensitive calcium binding proteins suggest alterations in local cortical circuitry without substantialloss of grey matter as is characteristic of periventricular leukomalacia. Changes in expression of Bax, cJun and non-phosphorylatedneurofilaments during normal development are also described.© 2003 ISDN. Published by Elsevier Science Ltd. All rights reserved.
Keywords:Periventricular leukomalacia (PVL); 3-Nitropropionic acid; Cerebral palsy; Hypoxia; Brain development
1. Introduction
Periventricular leukomalacia (PVL) is characterised by anecrotic, and often cystic lesion of the periventricular whitematter (Kuban, 1998) and is associated with premature ba-bies. Such a developmental lesion to the white matter resultsin the child displaying spastic diplegia, as seen in cerebralpalsy (Miller et al., 2000) whereby spasticity is present inall four limbs but most severely in the lower limbs. The in-cidence of PVL is 0.2 per 1000 live births (Miller et al.,
∗ Corresponding author. Present address: Sir James Spence Institute forChild Health, Royal Victoria Infirmary, Newcastle upon Tyne NE1 4LP,UK. Tel.: +44-191-202-3008; fax:+44-191-202-3022.
E-mail address:[email protected] (G.J. Clowry).1 Present address: Institute of Cell Signalling, Queens Medical Centre,
University of Nottingham, Nottingham NG7 2UH, UK.
2000) and the risk of PVL is greatly increased with decreas-ing gestational age (Back and Volpe, 1997). The prevalenceof cerebral palsy caused by PVL is increasing in the de-veloped world as a consequence of increased survival ofpreterm babies (Johnston, 1998) and many factors underly-ing the pathology of PVL remain unknown.
The pathological features of PVL include focal periven-tricular as well as more diffuse cerebral white matter dam-age to the white matter dorsal and lateral to the lateralventricle where the corticospinal tract is located (Panethet al., 1990). However, areas of grey matter surroundinglesion areas are largely spared. This results in disruption ofcorticospinal axons, while the cortical projection neuronesare left intact and subsequently make aberrant intracorticalaxonal projections (Marin-Padilla, 1997). The typical his-tological features of PVL are characterised by coagulationnecrosis, microglial infiltration, astrocytic proliferation and
0736-5748/03/$30.00 © 2003 ISDN. Published by Elsevier Science Ltd. All rights reserved.doi:10.1016/S0736-5748(03)00041-8

172 C.L. Gibson, G.J. Clowry / Int. J. Devl Neuroscience 21 (2003) 171–182
eventual cyst formation (Perlman, 1998). Although manydifferent aetiologies may lead to PVL the common patho-logical mechanism is believed to be focal ischemic injuryto the periventricular area (Back and Volpe, 1997).
Two major factors appear to be associated with increasedrisk of white matter damage in the premature brain: vasculardevelopment and the intrinsic vulnerability of developingoligodendrocytes to hypoxia. There is an increased risk ofhypoperfusion to the white matter in the premature brain.Lesions occurring in PVL are located at the terminationof major cerebral vessels in a border zone between ante-rior and middle and posterior cerebral arteries (Takashimaand Tanaka, 1978). These termination areas or ‘watershedareas’ are located most distal from direct blood supply andare poorly vascularised (De Reuck et al., 1972). Further-more, in the final stages of foetal development the whitematter grows rapidly. This requires more energy but at thesame time distance from the blood vessels is increased.The combination of these factors explains why the whitematter is particularly vulnerable to asphyxia, hypoxia, is-chemia and trauma (Marin-Padilla, 1997). Also the tempo-ral window during which PVL occurs closes during 30–32weeks post-conceptional age, a time point coincident witha marked increase in vascular supply to the white matter(Perlman, 1998; Takashima and Tanaka, 1978).
In the white matter, the cells most at risk from hypoxia arethe oligodendrocytes which may be due, in part, to the en-hanced glucose and energy requirements for lipid synthesisof these cells (Perlman, 1998). Immature oligodendrocytesare much more vulnerable to hypoxia (Husain and Juurlink,1995), oxidative stress (Back et al., 1998; Thorburne andJuurlink, 1996), and increased glutamate levels (Follettet al., 2000) than mature oligodendrocytes. PVL occursduring a period in white matter development well before theonset of active myelination (Brody et al., 1987) and such adevelopmental loss of immature oligodendrocytes preventstheir maturation and survival. Neuropathological studiesconfirm a loss of oligodendrocytes in the area surroundingthe lateral ventricle (Iida et al., 1995) as well as evidenceof abnormal or delayed myelination (Dambska et al., 1989;Paneth et al., 1990).
In order to further investigate the underlying pathologyassociated with PVL the present study was aimed at devel-oping an animal model of PVL. We decided to compare theuse of two neurochemicals, 3-nitropropionic acid (3-NP) andibotenic acid (IBA) both of which have been shown to in-duce lesions in other models. 3-NP is a mitochondrial toxinwhich irreversibly inhibits succinate dehydrogenase (Coleset al., 1979) in the Krebs cycle and complex II of the electrontransport chain thereby impairing oxidative metabolism andthus mimicking hypoxia. 3-NP has been shown to producewhite matter lesions (McCracken et al., 2001) with the dam-age restricted to the target area when injected intracerebrally(Miller and Zaborsky, 1997). IBA is a glutamate analog thatactivates both the NMDA and metabotropic receptors butnot AMPA and kainate receptors. Under conditions where
energy metabolism is impaired, extracellular glutamate lev-els are raised. Studies have shown that intracerebral injec-tions of IBA produce large periventricular white matter cysts(Gressens et al., 1999).
Rats were injected on postnatal day 7 (P7) as this is con-sidered a stage of brain development that corresponds tohuman foetuses during the third trimester (Hagberg et al.,1997). Animals were allowed to survive for various pe-riods in order to study the development of the overlyingcortex in the presence of a lesion. Immunocytochemicalanalysis was performed using various markers looking atchanges in cell death, neuronal activity and growing ax-ons. In addition, as expression of some of these proteinsis not well characterised in normal development, observa-tions of developmental changes are also included. Some ofthe results have previously been reported in abstract form(Gibson et al., 2001).
2. Experimental procedures
2.1. Animals and surgery
Wistar rats were purchased from B&K Universal Ltd.(Hull, UK) and housed and cared for in the Newcastle Com-parative Biology Centre. The day of birth was taken as P0.In order to model PVL in rats injections of 3-NP (Sigma,n = 10) or IBA (Sigma,n = 4) were used and comparedto saline injected controls (n = 4). Surgery and anaes-thesia were performed according toClowry and Flecknell(2000). Briefly, on P7, anaesthesia was induced by inject-ing 0.3�g/kg body weight fentanyl citrate and 10�g/kg flu-anisine sub-cutaneously (Hypnorm, Janssen). The forelimbmotor cortex (MC) was exposed and under stereotaxic guid-ance a glass tip attached to a Hamilton syringe was used toinject 2�l × 1�l boluses of IBA (1�g), 3-NP (300 mM inPBS) or saline into the sub-cortical white matter (Fig. 1).The flap of bone overlying the motor cortex was replacedand the overlying scalp sutured. Anaesthesia was reversedbut analgesia maintained by an intraperitoneal injection ofnalbuthine (1�g/kg). The pups were then kept warm andreturned to their mother. All the pups survived the surgeryregardless of whether they received injections of either toxinor saline.
Animals were allowed to survive for varying times (3days, 1–4 weeks) prior to termination of the experimentsby administering a lethal overdose of sodium pentobar-bitone. They were then transcardially perfused using abrief pulse of phosphate buffered saline (PBS, pH 7.4) fol-lowed by buffered fixative (1 ml/g body weight) consistingof 4% paraformaldehyde and 0.2% picric acid in 0.1 Msodium phosphate buffer, pH 7.4. Brains were removed andpost-fixed in the same solution for 2 h before being placedinto sucrose solution (30% in PBS) overnight. In addition,a number of unoperated animals were perfused at a varietyof ages ranging from P3 up to P70.

C.L. Gibson, G.J. Clowry / Int. J. Devl Neuroscience 21 (2003) 171–182 173
Fig. 1. The location of the motor cortex (MC) containing both the pri-mary and supplementary motor areas that project to the corticospinal tract(Donoghue and Wise, 1982) is outlined in both a diagrammatic represen-tation of the forebrain and in a section immunostained for parvalbumin.The sub-cortical white matter is marked WM. The six cortical layersare labeled with Roman numerals in the somatosensory cortex and areclearly visible in this preparation. The distinction between layers is lessdiscernible in the (agranular) motor cortex and this is a characteristicfeature of this region. The site of injection in the white matter close tothe ventricle is marked with an asterisk. Scale bar represents 500�m.
2.2. Histology
Sections of forebrain (50�m) were cut on a freezingmicrotome and placed into PBS in six parallel sets. Oneset of sections were mounted immediately onto gelatincoated slides and Nissl stained. Five sets were immunos-tained free-floating for parvalbumin (PV, using monoclonalantibody obtained from Sigma, diluted 1�g/ml), calbindin(CB, monoclonal antibody from Sigma diluted 0.5�g/ml),cJun (rabbit polyclonal antibody from Calbiochem, diluted1�g/ml), Bax (monoclonal antibody from Sigma diluted2�g/ml), and non-phosphorylated neurofilaments (NPNF,monoclonal antibody SMI-32 from Sternberger Monoclon-als diluted 1�g/ml). Primary antibodies were diluted in0.1 M PBS containing 0.1% Triton X-100 (Sigma) and 3%of the appropriate blocking serum (PBST). Sections wereincubated in primary antibody solution overnight at 4◦Con a slowly rotating mixer. Sections were then washed inPBS and then incubated for 2 h in biotinylated anti-mouse(5�g/ml in PBST, Vector Labs) or biotinylated anti-rabbit(1.25�g/ml in PBST, Sigma) depending on the primaryantibody. Sections were washed and then incubated for 1 h
in streptavidin-HRP (5�g/ml in PBST, Vector Labs). Fol-lowing further washing, sections were reacted with 3�l/mlhydrogen peroxide and 50�g/ml diaminobenzidine in PBSfor 20 min and then washed in PBS. Sections were thenmounted onto gelatin-coated slides, dehydrated, cleared andcoverslipped in Entellan (Merck).
2.3. Quantitative analysis
For each animal, using the PV and CB immunostainedsections, counts were made of the number of immunoposi-tive neurones in the forelimb motor cortex overlying the in-jection site and on the contralateral side. The primary motorcortex can be distinguished from the adjacent somatosensorycortex, particularly in sections stained for PV, by its lack ofa distinct layer IV (seeFig. 1). Using a camera lucida, theprecise area in which counts were to be made was drawnout and the number of PV or CB positive neuronal profilesin this area counted. A digitising tablet was used to mea-sure this area in which the counts had been made and thenumber of PV or CB positive neurones per mm2 of forelimbmotor cortex was estimated for each animal. Counts weremade separately for the injected and control sides enablingpairedt-tests to be used to test for significant differences be-tween the ipsilateral and contralateral sides in 3-NP treatedand saline control animals.
3. Results
3.1. Normal developmental expression of Bax,cJun and NPNF
Observations of protein expression patterns were limitedto the following areas (Fig. 2G): the agranular motor cortexincluding the primary motor cortex and the supplementarymotor cortex, also the granular somatosensory cortex andcingulate cortex. At all postnatal ages studied Bax im-munoreactivity was observed predominantly in cell nuclei(Fig. 2A,B,D,E,G) although weaker Bax immunoreactivitycould also be seen in the cytoplasm. Between P3 and P7 Baximmunostaining had a patchy appearance, being generallystronger in the somatosensory cortex, where it was present inlayers III and V, present in isolated columns in the motor cor-tex, and generally absent from the cingulate cortex (Fig. 2A).Also at these younger ages, Bax immunoreactivity waspresent in the apical dendrites of pyramidal cells (Fig. 2B)however this had disappeared by P14 (Fig. 2D and E). FromP14 onwards, the strongest Bax immunoreactivity was seenin layer V (Fig. 2D and G). Bax positive cells could alsobe seen in other cortical layers, in particular the cingulatecortex (Fig. 1G) where it had been absent at younger ages.
At all postnatal ages studied, cJun immunoreactivity waspredominantly found in cell nuclei with some staining moreintensely than others. Cytoplasmic immunostaining was veryweak or not present at all. At P3-7 (Fig. 2C), cJun im-

174 C.L. Gibson, G.J. Clowry / Int. J. Devl Neuroscience 21 (2003) 171–182
Fig. 2. Normal postnatal developmental expression of Bax and cJun. Part A shows a characteristic pattern of expression in the first week postnatally.Expression is strong in layers III and V in the somatosensory cortex (SI) but patchy in the motor cortex, perhaps being confined to isolated columns ofcortical neurones (arrows). Part B shows Bax immunoreactivity at a higher magnification. It is strongest in the nucleus but also present in the cytoplasm,including the long apical dendrite of the pyramidal neurones. Expression of cJun immunoreactivity was also patchy at this stage of development (C)and not confined to the somatosensory cortex, being located mainly in the premotor and cingulate cortex () in this section. By P14, both Bax (D)and cJun (F) expression was largely confined to cortical layer V. Bax immunoreactivity had disappeared from the apical dendrite (E). Part G showsthe mature cortex annotated to show its division into both layers (I–VI) and functional regions (SI, primary somatosensory cortex; MI, primary motorcortex; MII, supplementary motor cortex; Ci, cingulate cortex; according toDonoghue and Wise (1982). G also shows mature expression pattern of Baximmunoreactivity, confined mainly to layer V with although some cells in layers II/III and VI also express the protein. Expression of cJun becomesmore widespread in maturity. It is strongly represented in layer V neurones but is present in many cells throughout the cortex (H). Scale bars represent500�m (A, C, D, F–H) 100�m (B) and 50�m (E).
munoreactivity was most strongly present in layers III andV, with some cJun immunopositive cells scattered through-out layer VI. Up until P7 (Fig. 2C), cJun immunoreactivityhad a patchy appearance with the strongest staining being inareas of the somatosensory and cingulate cortex whilst cJunimmunoreactivity in the motor cortex was relatively weak.By P14, cJun immunoreactivity is largely confined to layerV (Fig. 2F). However, by P28 many cJun positive cells werefound throughout all cortical layers and many cJun positivecells persisted into adulthood.
The use of the monoclonal antibody SMI-32 revealedNPNF immunoreactivity present in cell body cytoplasm,primary dendrites and axons at various postnatal ages(Fig. 3A–E). At all postnatal ages studied, NPNF im-munoreactivity was found predominantly in layer III andV pyramidal neurons and also in some large multipolar
nerve cells. At P3, NPNF immunoreactivity was presentthroughout the somatosensory and motor cortex. It wasfound both in cell bodies and apical dendrites of pyramidalneurones but not in their axons at this age (Fig. 3A and B).By P5, a similar pattern of NPNF immunoreactivity couldbe seen although except that staining was now detectablein cortical axons projecting to the corpus callosum and inbundles through the striatum (Fig. 3C). By P14, NPNF im-munoreactivity was confined to layers III and V throughoutsomatosensory and motor areas (Fig. 3D) although therewas a marked absence of NPNF immunoreactivity fromthe cingulate cortex. By P70, there was lack of axonalstaining in all cortical areas and NPNF immunoreactiv-ity was confined to the cell body and primary dendritesof large neurones, mainly in layer V but also in layer III(Fig. 3E).

C.L. Gibson, G.J. Clowry / Int. J. Devl Neuroscience 21 (2003) 171–182 175
Fig. 3. Normal developmental expression of non-phosphorylated neurofilaments (NPNF). Parts A–C illustrate expression in the first week postnatally,which can be irregular, and generally stronger in the somatosensory cortex, although it is clearly expressed also in the motor areas. The higher powermicrograph (B) demonstrates that it is localised to dendrites as well as cell bodies. It was only observed in cortical axons from P5 onwards (C). By P14(D), NPNF immunoreactivity is strongly and consistently expressed by large neurones throughout the somatosensory and motor cortex, predominantlyinlayers III and V. Axonal immunostaining in the sub-cortical white matter is also prominent. However, expression in the cingulate cortex is still weak. Inthe mature animal (E) expression is present throughout layers III and V of the cortex in the cell bodies and primary dendrites of large neurones but hasdisappeared from their axons in the sub-cortical white matter. Scale Bars represents 500�m (A, C–E) and 100�m. (B).
3.2. A comparison of the effects of IBA versus 3-NPinjection
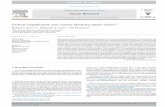
Neither toxin had an effect on the development of eithergross motor behaviour or use of the contralateral forelimb.Within 3 days following injections of either IBA (Fig. 4A) or3-NP (Fig. 4D) there was histological evidence of destruc-tion of the periventricular white matter. However, injectionsof IBA also resulted in large lesions and necrosis of theoverlying grey matter (Fig. 4B). In the area of cortex overly-ing the IBA injection sites, there was evidence of a reduceddensity of neurones and those that were present showed ev-idence of dying including darkly stained nuclei and vacuo-lated cytoplasm (Fig. 4C). These features are not generallycharacteristic of PVL whereas 3-NP injections producedmuch smaller and focal lesions restricted to the periventric-ular white matter and did not result in the death of overlyinggrey matter (Fig. 4D), which is a more characteristic model.
Due to this, the rest of the study was continued using 3-NPinjected animals and these were compared to saline controls.
3.3. Short-term effects of 3-NP injections
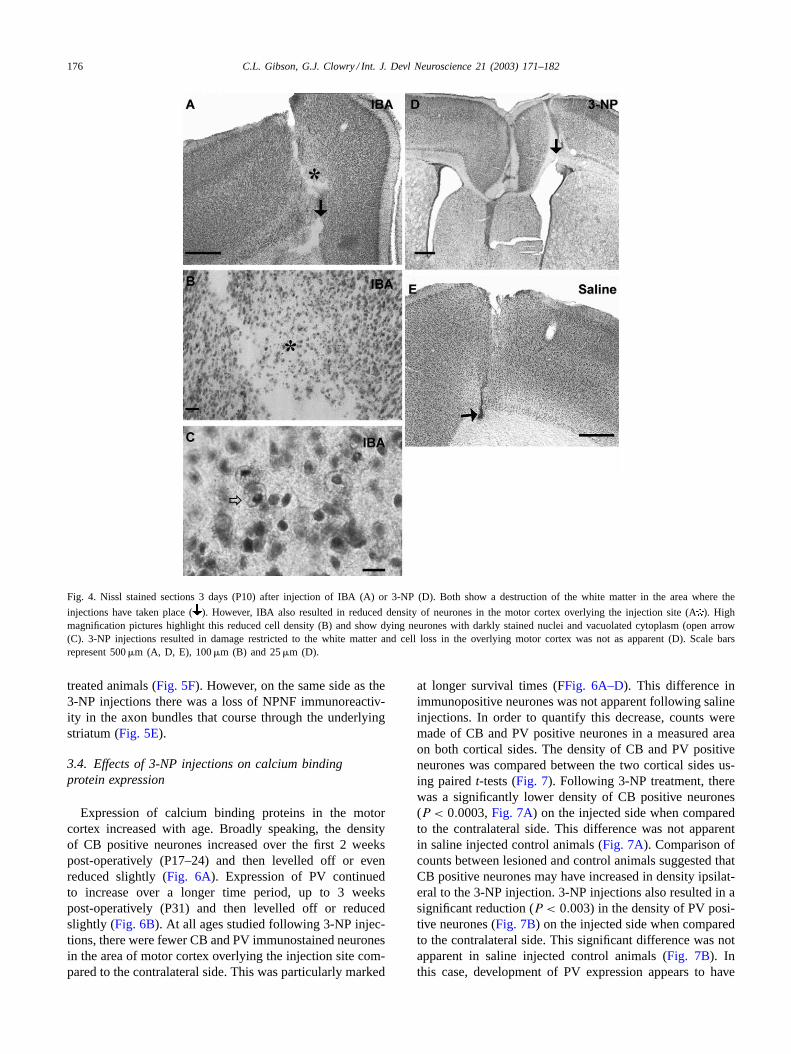
Short-term survival (1 and 2 weeks) of animals follow-ing 3-NP injections resulted in increased expression of Bax(Fig. 5A) and cJun (Fig. 5C) in layer V of the motor cortexoverlying the injection sites, compared to the contralateralside, nearby regions of the ipsilateral cortex, and normal an-imals that did not receive any interventions. Furthermore,these differences in immunostaining were not apparent innormal animals or following injection of saline (Fig. 5B andD). In addition, there was increased expression of Bax andcJun immunoreactivity in the cingulate gyrus on the sidecontralateral to the injection site. 3-NP injections also causedincreased expression of NPNF in the motor cortex overlyingthe injection site (Fig. 5E) which was not apparent in saline
176 C.L. Gibson, G.J. Clowry / Int. J. Devl Neuroscience 21 (2003) 171–182
Fig. 4. Nissl stained sections 3 days (P10) after injection of IBA (A) or 3-NP (D). Both show a destruction of the white matter in the area where the
injections have taken place (). However, IBA also resulted in reduced density of neurones in the motor cortex overlying the injection site (A). Highmagnification pictures highlight this reduced cell density (B) and show dying neurones with darkly stained nuclei and vacuolated cytoplasm (open arrow(C). 3-NP injections resulted in damage restricted to the white matter and cell loss in the overlying motor cortex was not as apparent (D). Scale barsrepresent 500�m (A, D, E), 100�m (B) and 25�m (D).
treated animals (Fig. 5F). However, on the same side as the3-NP injections there was a loss of NPNF immunoreactiv-ity in the axon bundles that course through the underlyingstriatum (Fig. 5E).
3.4. Effects of 3-NP injections on calcium bindingprotein expression
Expression of calcium binding proteins in the motorcortex increased with age. Broadly speaking, the densityof CB positive neurones increased over the first 2 weekspost-operatively (P17–24) and then levelled off or evenreduced slightly (Fig. 6A). Expression of PV continuedto increase over a longer time period, up to 3 weekspost-operatively (P31) and then levelled off or reducedslightly (Fig. 6B). At all ages studied following 3-NP injec-tions, there were fewer CB and PV immunostained neuronesin the area of motor cortex overlying the injection site com-pared to the contralateral side. This was particularly marked
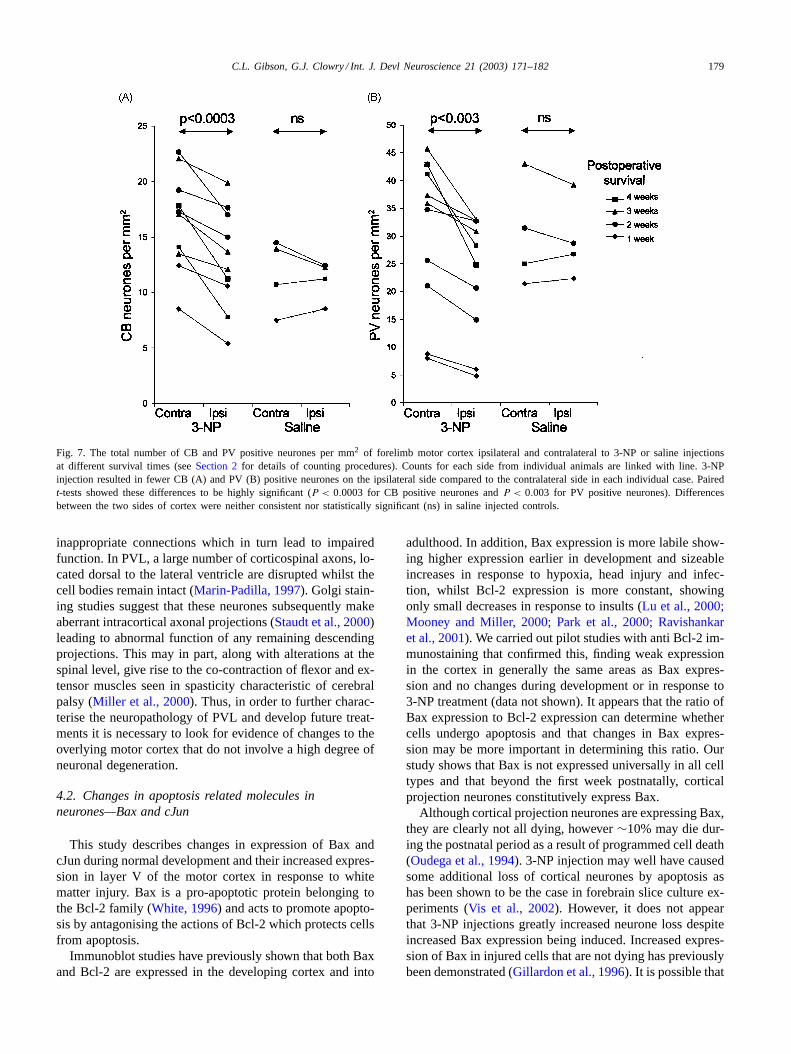
at longer survival times (FFig. 6A–D). This difference inimmunopositive neurones was not apparent following salineinjections. In order to quantify this decrease, counts weremade of CB and PV positive neurones in a measured areaon both cortical sides. The density of CB and PV positiveneurones was compared between the two cortical sides us-ing pairedt-tests (Fig. 7). Following 3-NP treatment, therewas a significantly lower density of CB positive neurones(P < 0.0003,Fig. 7A) on the injected side when comparedto the contralateral side. This difference was not apparentin saline injected control animals (Fig. 7A). Comparison ofcounts between lesioned and control animals suggested thatCB positive neurones may have increased in density ipsilat-eral to the 3-NP injection. 3-NP injections also resulted in asignificant reduction (P < 0.003) in the density of PV posi-tive neurones (Fig. 7B) on the injected side when comparedto the contralateral side. This significant difference was notapparent in saline injected control animals (Fig. 7B). Inthis case, development of PV expression appears to have
C.L. Gibson, G.J. Clowry / Int. J. Devl Neuroscience 21 (2003) 171–182 177
Fig. 5. The short-term effects, 7 days after treatment (P14) following damage (d) to the periventricular white matter as a result of 3-NP injections.Expression of Bax (A, ) and cJun (C, ) was greatly increased in layer V of the overlying motor cortex in comparison to normal animals that had notreceived any injections (B, D). However, 3-NP injection also resulted in an increased contralateral expression of Bax (A) and cJun (C), particularlyinthe cingulate gyrus (arrow) and also in a narrow band between the primary and supplementary motor cortex. NPNF expression was also increased inthe motor cortex following 3-NP injection (E,) in comparison to the normal expression (F). There was also a loss of NPNF immunoreactivity in axonbundles the underlying striatum (E, open arrow). Scale bars represent 500�m.
been retarded on both sides of the cortex initially following3-NP injection, but more so ipsilateral to the lesion. Threeto four weeks post-operatively, the difference between thetwo sides becomes more accentuated and involves both areduction in PV ipsilateral to the injection and an increaseon the contralateral side.
3.5. Other long-term changes following 3-NPinjections
With long-term survival (3 and 4 weeks), there was a per-sistence of increased NPNF expression in the motor cortexoverlying the injection site following 3-NP (Fig. 6F).
Bax and cJun expression appeared relatively normal inthe longer term.
4. Discussion
The present study has described and compared the use oftwo neurotoxic agents in order to develop a realistic animalmodel of PVL. Following both IBA and 3-NP injections,there were obvious signs of destruction of the periventricu-lar white matter as seen in the human condition PVL. How-ever, IBA injections also resulted in the large-scale death ofneurones by necrosis in the overlying grey matter which isless characteristic of PVL. Following injection of 3-NP at

178 C.L. Gibson, G.J. Clowry / Int. J. Devl Neuroscience 21 (2003) 171–182
Fig. 6. Four weeks after 3-NP injections various long-term changes in expression of different immunocytochemical markers were evident. This figure iscomposed from adjacent sections from the same animal. There were fewer PV (B,) and CB (D, ) positive neurones in the motor cortex ipsilateralto the injection, compared to the contralateral side. The decreased expression of PV is apparent throughout all layers of the cortex whereas loss of CBexpression seems confined to cells in more superficial layers. However, there was an increased expression of NPNF in the motor cortex overlying theinjection site (F) compared with the contralateral cortex (E). Scale bars represent 500�m (A, B) and 250�m (C, D).
all survival times studied, white matter lesions were presentwithout any signs of large-scale destruction of the overlyinggrey matter. No obvious signs of motor impairment wereseen, even after 4 weeks, however we have previously foundthat extensive removal of the forelimb motor cortex at P7in the rat fails to produce easily detectable effects upon de-velopment of motor control in the rat (Gibson et al., 2000).Nevertheless, it is unlikely that such lesions to the whitematter did not impact upon the grey matter and in order toinvestigate this, we investigated changes in expression ofproteins using immunocytochemistry[DoH1][DoH2][CG3].These included the pro-apoptotic proteins Bax and cJun,NPNF, a marker for large cortical projection neurones andtheir axons, and calcium binding proteins, the expressionof which is activity dependent. Reports of studies of Bax,cJun, and NPNF immunocytochemistry during developmentare limited therefore we also studied the localisation of ex-pression of immunoreactivity of these proteins postnatally.
4.1. Choice of model
The majority of the present study was carried out us-ing 3-NP injection as a model for PVL because the greymatter was relatively spared. Other groups using IBA havereported the presence of large periventricular white mattercysts however some (Gressens et al., 1999) have also de-scribed neuronal loss affecting all cortical layers. This isperhaps not surprising. Although immature oligodendro-cytes express glutamate receptors, these are predominantlyof the AMPA sensitive sub-type (Gallo et al., 1994; Follettet al., 2000). IBA is primarily an agonist of NMDA andmetabotropic glutamate receptors, which are expressed byneurones, along with AMPA receptors.
Although a realistic model of PVL would not involvelarge-scale destruction of the grey matter its compositionshould be altered due to the deprivation of afferent inputs andnormal efferent output. As a result, neurons may form new,
C.L. Gibson, G.J. Clowry / Int. J. Devl Neuroscience 21 (2003) 171–182 179
Fig. 7. The total number of CB and PV positive neurones per mm2 of forelimb motor cortex ipsilateral and contralateral to 3-NP or saline injectionsat different survival times (seeSection 2for details of counting procedures). Counts for each side from individual animals are linked with line. 3-NPinjection resulted in fewer CB (A) and PV (B) positive neurones on the ipsilateral side compared to the contralateral side in each individual case. Pairedt-tests showed these differences to be highly significant (P < 0.0003 for CB positive neurones andP < 0.003 for PV positive neurones). Differencesbetween the two sides of cortex were neither consistent nor statistically significant (ns) in saline injected controls.
inappropriate connections which in turn lead to impairedfunction. In PVL, a large number of corticospinal axons, lo-cated dorsal to the lateral ventricle are disrupted whilst thecell bodies remain intact (Marin-Padilla, 1997). Golgi stain-ing studies suggest that these neurones subsequently makeaberrant intracortical axonal projections (Staudt et al., 2000)leading to abnormal function of any remaining descendingprojections. This may in part, along with alterations at thespinal level, give rise to the co-contraction of flexor and ex-tensor muscles seen in spasticity characteristic of cerebralpalsy (Miller et al., 2000). Thus, in order to further charac-terise the neuropathology of PVL and develop future treat-ments it is necessary to look for evidence of changes to theoverlying motor cortex that do not involve a high degree ofneuronal degeneration.
4.2. Changes in apoptosis related molecules inneurones—Bax and cJun
This study describes changes in expression of Bax andcJun during normal development and their increased expres-sion in layer V of the motor cortex in response to whitematter injury. Bax is a pro-apoptotic protein belonging tothe Bcl-2 family (White, 1996) and acts to promote apopto-sis by antagonising the actions of Bcl-2 which protects cellsfrom apoptosis.
Immunoblot studies have previously shown that both Baxand Bcl-2 are expressed in the developing cortex and into
adulthood. In addition, Bax expression is more labile show-ing higher expression earlier in development and sizeableincreases in response to hypoxia, head injury and infec-tion, whilst Bcl-2 expression is more constant, showingonly small decreases in response to insults (Lu et al., 2000;Mooney and Miller, 2000; Park et al., 2000; Ravishankaret al., 2001). We carried out pilot studies with anti Bcl-2 im-munostaining that confirmed this, finding weak expressionin the cortex in generally the same areas as Bax expres-sion and no changes during development or in response to3-NP treatment (data not shown). It appears that the ratio ofBax expression to Bcl-2 expression can determine whethercells undergo apoptosis and that changes in Bax expres-sion may be more important in determining this ratio. Ourstudy shows that Bax is not expressed universally in all celltypes and that beyond the first week postnatally, corticalprojection neurones constitutively express Bax.
Although cortical projection neurones are expressing Bax,they are clearly not all dying, however∼10% may die dur-ing the postnatal period as a result of programmed cell death(Oudega et al., 1994). 3-NP injection may well have causedsome additional loss of cortical neurones by apoptosis ashas been shown to be the case in forebrain slice culture ex-periments (Vis et al., 2002). However, it does not appearthat 3-NP injections greatly increased neurone loss despiteincreased Bax expression being induced. Increased expres-sion of Bax in injured cells that are not dying has previouslybeen demonstrated (Gillardon et al., 1996). It is possible that

180 C.L. Gibson, G.J. Clowry / Int. J. Devl Neuroscience 21 (2003) 171–182
cells have an increased susceptibility to apoptosis followinginjury and this is reflected by increases in Bax expression.Although such proteins were originally identified as mark-ers of cell death their expression is not always a precursor toapoptosis and has been linked to regeneration. Also, it hasbeen shown that Bcl-2 is inactivated upon phosphorylation(Haldar et al., 1995). Due to the close homology betweenBcl-2 and Bax, it remains possible that if Bax is similarlyregulated in this way. We conclude that the expression ofBax alone is not sufficient to induce cell death.
Immediate early gene (IEG) products such as cJun aretranscription factors that affect the expression of other genesover longer periods of time. Although induction of cJunexpression can be a precursor to apoptosis it has also beenlinked to regeneration (Herdegen et al., 1997). Following anerve transection, neurones either die or regenerate depend-ing on the proximity of the transection to the cell soma.Proximal transections of the optic nerve lead to vigorousregeneration and a marked increase in cJun expression(Herdegen et al., 1997), suggesting an association betweenincreased levels of cJun expression and increased metabolicactivity necessary to promote regeneration. Expression lev-els of immediate early genes such as cJun can also be anindicator of any alterations in the levels of neuronal activitypresent as their expression has been shown to increase duringconditions of increased synaptic activity such as long-termpotentiation (Hughes et al., 1999). In addition, conditioningischemia selectively induces cJun in hippocampal CA1 neu-rons and many of the cells positive for cJun not only survivethe initial ischemia, but become protected from cell deathfollowing a subsequent ischemic insult (Sommer et al.,1995). Our developmental studies have shown that, as withBax, cortical projection neurones express cJun during devel-opment and continue to do so in maturity. Therefore, in thepresent study, the increased expression of Bax and cJun mayindicate that injured neurones are primed to make a choicebetween either cell death or an attempt at regeneration.
4.3. Changes in neurofilament expression
Neurofilaments are strands of protein belonging to one ofthe three major cytoskeletal elements found in neurons andare believed to serve an important structural role in neurons.Neurofilaments sustain the axonal and dendritic branchingpatterns and promote axonal growth or thickening (Leterrieret al., 1996). They also participate in the dynamic propertiesof the axonal cytoskeleton during neuronal differentiation,axon outgrowth and guidance (Nixon and Shea, 1992). In themature neocortex SMI-32, antibodies label a sub-populationof pyramidal cells that contain the non-phosphorylated neu-rofilament triplet protein (Hiscock and Mackenzie, 1998).
The monoclonal antibody SMI-32 recognises an epitopepresent in neurofilaments in the non-phosphorylated stateand visualises neuronal cell bodies, dendrites and somethick axons (Sternberger and Sternberger, 1983). Early onin development neurofilaments are non-phosphorylated and
function to maintain cytoskeletal plasticity for axonal out-growth and neuronal migration (Grant and Pant, 2000). Thisexplains why in the present study shows large amounts ofNPNF expression earlier on in development when comparedto adulthood. The present study also reported a loss of NPNFpositive axons in the striatum underlying the injection sitethat probably reflects a loss of cortical axons in this area as aresult of lesioning. Neurofilaments are assembled in the cellbody of neurones, transported down the length of the axonand degraded in the nerve terminal. Axotomy will in turnlead to accumulation of neurofilament in cell bodies andproximal stump. This was seen in the short-term followinglesioning in the present study. In the longer term, high NPNFexpression in the cortex persisted.King et al. (2001)haveprovided evidence that neurofilaments play a role in axonalsprouting following brain injury. Therefore, persistence ofNPNF expression observed in our study may represent theintracortical sprouting of axons, as has been reported inhuman PVL (Marin-Padilla, 1997, seeSection 1). Patholog-ical changes in NPNF expression including accumulationof protein in the cell body have also been seen in multiplesclerosis as a result axonal disruption (Trapp et al., 1998).
4.4. Levels of neuronal activity—changes in calciumbinding proteins
CB and PV are calcium binding proteins expressed mainlybe inhibitory interneurones (Celio, 1990). In the cerebralcortex, PV is found in the majority of fast spiking interneu-rones with over half of these cells also expressing calbindin(Cauli et al., 1997). Thus, the expression of such calciumbinding proteins is normally associated with highly activeneurones (Kawaguchi et al., 1987) and the changed expres-sion observed in the present study is taken to reflect differ-ences in activity levels in those neurones. In addition, theexpression of calcium binding proteins has been shown to bedependent upon the development of normal afferent input inthe rat barrel cortex. Thus, there is reduced expression of PVand CB by cortical interneurones if inputs from the thalamusare disrupted during development (Alcantara et al., 1996).Similarly, neurones in the spinal cord fail to express PV inresponse to decreased corticospinal input during develop-ment (Gibson et al., 2000). Expression of PV by the humanthalamocortical system is dependent upon the establishmentof functional circuitry and there is evidence of alterations inexpression of PV immunoreactivity by cortical neurones inresponse to white matter lesions arising from PVL (Iai andTakashima, 1999). In the present study, there was a persis-tence of differential CB and PV expression between the twosides of the cortex following unilateral 3-NP injection. Thisprobably reflects an activity dependent change of phenotypein those cells rather than any loss of cells. An interestingobservation from this study is that increases in expressionon the contralateral side contributed at least as much as re-ductions ipsilateral to the injection to the overall imbalancein expression between the two sides. This may reflect two

C.L. Gibson, G.J. Clowry / Int. J. Devl Neuroscience 21 (2003) 171–182 181
different phenomena. Firstly, it illustrates that the two cor-tices are functionally linked such that alterations to one sidehave knock-on effects on the other side. Secondly, disrup-tion of one corticospinal pathway during development maylead to a retention and upregulation of the ipsilateral path-way from the contralateral cortex in both rat (Aisaka et al.,1999; Reinoso and Castro, 1989) and human (Eyre et al.,2001). This might result in increased activity and expressionof activity dependent proteins in the contralateral motorcortex. However, in man, there is no evidence as yet thatupregulation of the ipsilateral pathway following a unilat-eral perinatal lesion correlates with a decreased expressionof hemiplegia in the affected individual (J. Eyre, personalcommunication).
4.5. Long-term aims of developing a lesion model
This study has employed 3-NP in order to produce a con-sistent and reproducible lesion of the periventricular whitematter during a time period of direct relevance to PVL. Sucha model was characterised by destruction of the white matterwithout any significant loss of overlying grey matter. How-ever, as seen in PVL, the grey matter was not left unaffectedand we have reported various cellular changes in responseto the lesion.
One of the main characteristics of PVL is the damagecaused by the death of oligodendrocytes. However, theseglial cells and the myelin they produce, along with laterformed glial scars provide an inhibitory environment inwhich to promote regeneration. A more detailed studyof the glial response to 3-NP injections in this model, incomparison to the response of glial cells in the human con-dition is required. However, one approach to treating thesymptoms of PVL, i.e. spasticity, would be promoting earlyregeneration of corticospinal projections and their appropri-ate innervation of spinal cord areas. The corticospinal tractretains a high degree of plasticity throughout human devel-opment (Eyre et al., 2000). Therefore, early interventionrepair strategies that could persuade disrupted corticospinalaxons to re-grow through the cystic areas of PVL, coupledwith the neutralisation of the inhibitory properties of myelin(Raineteau and Scwab, 2001) may allow corticospinal ax-ons find their appropriate targets (Terashima, 1995). Thedevelopment of an appropriate model increases the possibil-ity of revealing the conditions necessary to promote axonalregeneration and restore appropriate function.
Acknowledgements
Mr. Gordon Arnott provided skilled technical supportthroughout this study. We are grateful to the staff of theNewcastle Comparative Biology Centre for their care of theexperimental animals, and to Dr. Bert Joosten for helpfuladvice and suggestions. This work was supported by theNewcastle University Hospitals Special Trustees.
References
Aisaka, A., Aimi, Y., Yasuhara, O., Tooyama, I., Kimura, H., Shimada,M., 1999. Two modes of corticospinal reinnervation occur close tospinal targets following unilateral lesion to the motor cortex in neonatalhamsters. Neuroscience 90, 53–67.
Alcantara, S., Soriano, E., Ferrer, I., 1996. Thalamic and basal forebrainafferents modulate the development of parvalbumin and calbindin D28kimmunoreactivity in the barrel cortex of the rat. Eur. J. Neurosci. 8,1522–1534.
Back, S.A., Gan, X., Li, Y., Rosenberg, P.A., Volpe, J.J., 1998. Maturationdependent vulnerability of oligodendrocytes to oxidative stress-induceddeath caused by glutathione depletion. J. Neurosci. 18, 6241–6253.
Back, S.A., Volpe, J.J., 1997. Cellular and molecular pathogenesis ofperiventricular white matter. Ment. Retard. Dev. Disabil. Res. Rev. 3,96–107.
Brody, B.A., Kinney, H.C., Kloman, A.S., 1987. Sequence of centralnervous system myelination in human infancy. I. An autopsy study ofmyelination. J. Neuropathol. Exp. Neurol. 46, 283–301.
Cauli, B., Audinat, E., Lamobolez, B., Angulo, M.C., Ropert, N., Tsuzuki,K., Hestrin, S., Rossier, J., 1997. Molecular and physiological diversityof cortical non-pyramidal cells. J. Neurosci. 17, 3894–3906.
Celio, M.R., 1990. Calbindin D28K and parvalbumin in the rat nervoussystem. Neuroscience 35, 375–475.
Clowry, G.J., Flecknell, P.A., 2000. The successful use offentanyl/fluanisone (‘Hypnorm’) as an anaesthetic for intracranialsurgery in neonatal rats. Lab. Anim. 34, 260–264.
Coles, C.J., Edmondson, D.E., Singer, T.P., 1979. Inactivation of succinatedehydrogenase by 3-nitropropionate. J. Biol. Chem. 254, 5161–5167.
Dambska, M., Laure-Kamionowska, M., Schmidt-Sidor, B., 1989. Earlyand late pathological changes in perinatal white matter damage. J.Child Neurol. 4, 29–38.
De Reuck, J., Chatta, A.S., Richardson, E.P., 1972. Pathogenesis andevolution of periventricular leukomalacia in infancy. Arch. Neurol. 27,229–236.
Donoghue, J.P., Wise, S.P., 1982. The motor cortex of the rat. J.Comp. Neurol. 212, 76–88.
Eyre, J.A., Miller, S., Clowry, G.J., Conway, E.A., Watts, C., 2000.Functional corticospinal projections are established prenatally in thehuman foetus permitting involvement in the development of spinalmotor centres. Brain 123, 51–64.
Eyre, J.A., Taylor, J.P., Villagra, F., Smith, M.R., Miller, S., 2001. Evidenceof activity dependent withdrawal of corticospinal projections duringhuman development. Neurology 57, 1543–1554.
Follett, P.L., Rosenberg, P.A., Volpe, J.J., Jensen, F.E., 2000. NBQXattenuated excitotoxic injury in developing white matter. J. Neurosci.20, 9235–9241.
Gallo, V., Patneau, D.K., Mayer, M.L., Vaccarino, F.M., 1994. Excitatoryamino acid receptors in glial progenitor cells: molecular and functionalproperties. Glia 11, 94–101.
Gibson, C.L., Arnott, G.A., Clowry, G.J., 2000. Plasticity in the rat spinalcord seen in response to lesions to the motor cortex during developmentbut not to lesions in maturity. Exp. Neurol. 166, 422–434.
Gibson, C.L., Arnott, G.A., Clowry, G.J., 2001. Towards developing a ratmodel of periventricular leucomalacia. Br. Neurosci. Assoc. Abstr. 16,43.
Gillardon, F., Lenz, C., Waschke, K.F., Krajewski, S., Reed, J.C.,Zimmermann, M., Kuschinsky, W., 1996. Altered expression of Bcl-2,Bcl-X, Bax and c-Fos colocalizes with DNA fragmentation andischemic cell damage following middle cerebral artery occlusion inrats. Mol. Brain Res. 40, 254–260.
Grant, P., Pant, H.C., 2000. Neurofilament protein synthesis andphosphorylation. J. Neurocytol. 29, 843–872.
Gressens, P., Besse, L., Robberecht, P., Gozes, I., Fridkin, M., Evrard,P., 1999. Neuroprotection of the developing brain by systemicadministration of vasoactive intestinal peptide derivatives. J. Pharmacol.Exp. Ther. 288, 1207–1213.

182 C.L. Gibson, G.J. Clowry / Int. J. Devl Neuroscience 21 (2003) 171–182
Hagberg, H., Bona, E., Gilland, E., Puka-Sundvall, M., 1997.Hypoxia-ischaemia model in the 7-day-old rat: possibilities andshortcomings. Acta Paediatr. Suppl. 422, 85–88.
Haldar, S., Jena, N., Croce, C.M., 1995. Inactivation of Bcl-2 byphosphorylation. Proc. Natl. Acad. Sci. U.S.A. 92, 4507–4511.
Herdegen, T., Skene, P., Bahr, M., 1997. The c-Jun transcription factor—bipotential mediator of neuronal death, survival and regeneration.Trends Neurosci. 20, 227–231.
Hiscock, J.J., Mackenzie, L., 1998. Laminar distribution of Fos/calcium-binding protein and Fos/neurofilament protein-labeled neurons inrat motor and sensory cortex after picrotoxin-induced seizures.Exp. Neurol. 149, 373–383.
Hughes, P.E., Alexi, T., Walton, M., Williams, C.E., Dragunow, M., Clark,R.G., Gluckman, P.D., 1999. Activity and injury dependent expressionof inducible transcription factors, growth factors and apoptosis relatedgenes within the central nervous system. Prog. Neurobiol. 57, 421–450.
Husain, J., Juurlink, B.H., 1995. Oligodendroglial precursor cellsusceptibility to hypoxia is related to poor ability to cope with reactiveoxygen species. Brain Res. 698, 86–94.
Iai, M., Takashima, S., 1999. Thalamocortical development ofparvalbumin neurons in normal and periventricular leukomalacia brains.Neuropediatrics 30, 14–18.
Iida, K., Takashima, S., Ueda, K., 1995. Immunohistochemical studyof myelination and oligodendrocytes in infants with periventricularleukomalacia. Pediatr. Neurol. 13, 296–304.
Johnston, M.V., 1998. Selective vulnerability in the neonatal brain. Ann.Neurol. 44, 155–156.
Kawaguchi, Y., Katsamuru, H., Kosaka, T., Heizmann, C.W., 1987. Fastspiking cells in the rat hippocampus (CA1 region) contain the calciumbinding protein parvalbumin. Brain Res. 416, 369–374.
King, C.E., Canty, A.J., Vickers, J.C., 2001. Alterations in neurofilamentsassociated with reactive brain changes and axonal sprouting followingacute physical injury to the rat neocortex. Neuropathol. Appl.Neurobiol. 27, 115–126.
Kuban, K.C.K., 1998. White-matter disease of prematurity, periventricularleukomalacia, and ischemic lesions. Dev. Med. Child Neurol. 40, 571–573.
Leterrier, J.F., Hartwig, J., Kas, J., Vegnrs, R., Janmey, P., 1996.Mechanical effects of neurofilament crossbridges: modulation byphosphorylation, lipids and interactions with F-actin. J. Biol. Chem.271, 15687–15694.
Lu, J., Moochhala, S., Kaur, C., Ling, E.A., 2000. Changes inapoptosis-related protein (p53, Bax, Bcl-2 and Fos) expression withDNA fragmentation in the central nervous system of rats after closedhead injury. Neurosci. Lett. 290, 89–92.
Marin-Padilla, M., 1997. Developmental neuropathology and impact ofperinatal brain damage. II. White matter lesions of the neocortex. J.Neuropathol. Exp. Neurol. 56, 219–235.
McCracken, E., Dewar, D., Hunter, A.J., 2001. White matterdamage following systemic injection of the mitochondrial inhibitor3-nitropropionic acid in rat. Brain Res. 892, 329–335.
Miller, P.J., Zaborsky, L., 1997. 3-Nitropropionic acid neurotoxicity:visualization by silver staining and implications for use as an animalmodel of Huntington’s disease. Exp. Neurol. 146, 212–229.
Miller, S.P., Shevell, M.I., Patenaude, Y., O’Gorman, A.M., 2000.Neuromotor spectrum of periventricular leukomalacia in children bornat term. Pediatr. Neurol. 23, 155–159.
Mooney, S.M., Miller, M.W., 2000. Expression of bcl-2, bax and caspase-3in the brain of the developing rat. Dev. Brain Res. 123, 103–117.
Nixon, R.A., Shea, T.B., 1992. Dynamics of neuronal intermediatefilaments: a developmental perspective. Cell. Motil. Cytoskeleton 22,81–91.
Oudega, M., Varon, S., Haag, T., 1994. Distribution of corticospinal motorneurons in the postnatal rat: quantitative evidence for massive collateralelimination and modest cell death. J. Comp. Neurol. 347, 115–126.
Paneth, N., Rudeli, R., Monte, W., Rodriguez, E., 1990. White matternecrosis in the very low birth weight infants: neuropathologic andultrasonographic findings in infants surviving 6 days or longer. J.Pediatr. 116, 975–984.
Park, S.K., Choi, S.I., Jin, J.K., Choi, E.K., Kim, J.I., Carp, R.I., Kim,Y.S., 2000. Differential expression of Bax and BCl-2 in the brains ofhamsters infected with 263K scrapie agent. NeuroReport 11, 1677–1682.
Perlman, J.M., 1998. White matter injury in the preterm infant: animportant determination of abnormal developmental outcome. EarlyHum. Dev. 53, 99–120.
Raineteau, O., Scwab, M.E., 2001. Plasticity of motor systems afterincomplete spinal cord injury. Nat. Rev. Neurosci. 2, 263–273.
Ravishankar,S., Ashraf, Q.M., Fritz, K., Mishra, O.P., Delivoria-Papadopoulos, M.,2001. Expression of Bax and Bcl-2 proteins during hypoxia in cerebralcortical neuronal nuclei of newborn piglets: effects of administrationof magnesium sulfate. Brain Res. 901, 23–29.
Reinoso, B.S., Castro, A.J., 1989. A study of corticospinal remodellingusing retrograde fluorescent tracers in rats. Exp. Brain Res. 74, 387–394.
Sommer, C., Gass, P., Kiessling, M., 1995. Selective cJun expression inCA1 neurons of the gerbil hippocampus during and after acquisitionof an ischemia-tolerant state. Brain Pathol. 5, 135–144.
Staudt, M., Niemann, G., Grodd, W., Krageloh-Mann, I., 2000. Thepyramidal tract in congenital hemiparesis: relationship betweenmorphology and function in periventricular lesions. Neuropediatrics31, 257–264.
Sternberger, L.A., Sternberger, N.H., 1983. Monoclonal antibodiesdistinguish phosphorylated and non-phosphorylated forms ofneurofilaments in situ. Proc. Natl. Acad. Sci. U.S.A. 80, 6126–6130.
Takashima, S., Tanaka, K., 1978. Development of cerebrovasculararchitecture and its relationship to periventricular leukomalacia. Arch.Neurol. 35, 11–16.
Terashima, T., 1995. Anatomy, development and lesion-induced plasticityof rodent corticospinal tract. Neurosci. Res. 22, 139–161.
Thorburne, S.K., Juurlink, B.H., 1996. Low glutathione and high irongovern the susceptibility of oligodendroglial precursors to oxidativestress. J. Neurochem. 67, 1014–1022.
Trapp, B.D., Peterson, J., Ransohoff, R.M., Rudick, R., Mork, S., Bo, L.,1998. Axonal transection in the lesions of multiple sclerosis. N. Engl.J. Med. 338, 278–285.
Vis, J.C., de Boer-van Huizen, R.T., Verbeek, M.M., de Waal, R.M.W.,ten Donkelaar, H.J., Kremer, B., 2002. 3-Nitropropionic acid inducescell death and mitochondrial dysfunction in rat corticostriatal slices.Neurosci. Lett. 329, 86–90.
White, E., 1996. Life, death and the pursuit of apoptosis. Genes Dev. 10,1–15.