
Rajeshkumar Final Thesis.pdf - Bharathidasan University ...
247
ISOLATION, IDENTIFICATION AND OPTIMIZATION OF PHOSPHATE SOLUBILIZING MICROORGANISMS BY PHYSICAL AND CHEMICAL MUTAGENS, CARBON AND NITROGEN SOURCES ON THE PRODUCTION OF PHOSPHATASE AND LIPASE THESIS SUBMITTED TO BHARATHIDASAN UNIVERSITY FOR THE AWARD OF THE DEGREE OF DOCTOR OF PHILOSOPHY IN BOTANY By J. RAJESH KUMAR, M.Sc., M.Phil., Research Supervisor Dr. M.H. Muhammad Ilyas M.Sc., Ph.D., Associate Professor of Botany P.G. AND RESEARCH DEPARTMENT OF BOTANY JAMAL MOHAMED COLLEGE (AUTONOMOUS) (ACCREDITED WITH ‘A’ GRADE BY NAAC- CGPA 3.6 OUT OF 4.0) TIRUCHIRAPPALLI- 620 020 TAMIL NADU, INDIA July 2011
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Rajeshkumar Final Thesis.pdf - Bharathidasan University ...

ISOLATION, IDENTIFICATION AND OPTIMIZATION OF PHOSPHATE SOLUBILIZING MICROORGANISMS BY
PHYSICAL AND CHEMICAL MUTAGENS, CARBON AND NITROGEN SOURCES ON THE PRODUCTION OF
PHOSPHATASE AND LIPASE
THESIS SUBMITTED TO
BHARATHIDASAN UNIVERSITY
FOR THE AWARD OF THE DEGREE OF
DOCTOR OF PHILOSOPHY
IN
BOTANY
By
J. RAJESH KUMAR, M.Sc., M.Phil.,
Research Supervisor Dr. M.H. Muhammad Ilyas M.Sc., Ph.D.,
Associate Professor of Botany
P.G. AND RESEARCH DEPARTMENT OF BOTANY JAMAL MOHAMED COLLEGE (AUTONOMOUS)
(ACCREDITED WITH ‘A’ GRADE BY NAAC- CGPA 3.6 OUT OF 4.0)
TIRUCHIRAPPALLI- 620 020 TAMIL NADU, INDIA
July 2011

Date:
CERTIFICATE
This is to certify that the thesis entitled “Isolation, identification
and optimization of phosphate solubilizing microorganisms by
physical and chemical mutagens, carbon and nitrogen sources
on the production of phosphatase and lipase”, submitted to
Bharathidasan University, Tiruchirappalli - 620 024, for the award of the degree
of Doctor of Philosophy in Botany, is an authentic record of original work
carried out by Mr. J. RAJESHKUMAR under my guidance and supervision
during the study period at the PG and Research Department of Botany, Jamal
Mohamed College, Tiruchirappalli- 620 020, Tamil Nadu, India.
I further certify that no part of this thesis has been submitted any where
else for the award of any degree, diploma, associateship, fellowship or other
similar titles to any candidate.
(M.H. MUHAMMAD ILYAS)
Dr. M.H. MUHAMMAD ILYAS Associate Professor
Jamal Mohamed College (Autonomous)
(Nationally reaccredited with ‘A’ grade by NAAC in 2009)
PG & Research Department of Botany

Date:
DECLARATION
I do hereby declare that this work entitled “Isolation, identification and
optimization of phosphate solubilizing microorganisms by
physical and chemical mutagens, carbon and nitrogen sources
on the production of phosphatase and lipase”, has been originally
carried out by me under the guidance of Dr. M.H. Muhammad Ilyas, Associate
Professor, PG and Research Department of Botany, Jamal Mohamed College,
Tiruchirappalli- 620 020, and this work has not been submitted elsewhere for
any other degree.
(J. RAJESHKUMAR)

ACKNOWLEDGEMENTS
I have pleasure in writing this page in my dissertation to express my happiness
which I feel to acknowledge all those who helped me to complete this work
successfully.
It gives me great pleasure expressing my deep sense of indebtedness and
gratitude to my research guide Dr. M.H. Muhammad Ilyas, M.Sc., Ph.D., Associate
Professor, PG and Research Department of Botany, Jamal Mohamed College
(Autonomous), Tiruchirapalli – 620 020 for all the encouragement and valuable
guidance received by me at every stage of the work. The useful discussion I had with
him, both inside and outside the college, were always a source of inspiration to me.
I sincerely thank Hajee Dr. M. Sheik Mohamed, M.Com., M.Phil., Ph.D,
PGDCA, FICWA, FMSPI, PGDFM., Dip. In M.A. former Principal, Jamal
Mohamed College, Tiruchirapalli – 620 020 for having given me this wonderful
opportunity to do my Ph.D work at Jamal Mohamed College , Tiruchirapalli – 620
020.
My earnest thanks and deep of gratitude to Hajee Dr. R. Kadher
Mohiadeen, M.Com., M.Phil., Ph.D. Principal, Jamal Mohamed College,
Tiruchirapalli – 620 020 for his encouragement.
I am very grateful to Dr.S.Ahemed John, M.Sc., M.Phil., Ph.D., Associate
Professor and Head, PG and Research Department of Botany, Jamal Mohamed
College (Autonomous), Tiruchirapalli– 620 020 for their constant encouragement and
support.
I sincerely thank both Teaching and Non teaching staff members of PG and
Research Department of Botany, Jamal Mohamed College (Autonomous),
Tiruchirapalli – 620 020 for their generous help and Co-operation.

My special heartfelt thanks are due to Dr. N.Sengottain, M.Sc., M.Phil.,
Ph.D. Associate Professer and Head, Department of Botany and Microbiology,
Urumu Dhanalakhsmi college, Tiruchirapalli for having provided me with all the
facilities and encouragement throughout the course of this work.
I extent my sincere gratitude to Mr. P. Malaiyarasa Pandian, M.Sc.,
M.B.A., M.Phil. Lecturer, Department of Microbiology, Hindusthan College of Arts
and Science, Chennai, and Sangeetha Menan for his keen interest and valuable
suggestions in the progress of my work.
I extend my thanks to Mr. A. Shanmugasundaram, Lecturer, Department of
Microbiology, Urumu Dhanalakshmi College, for their appreciable help during the
course my research.
I express my thanks to Dr. Subramaniyam V. Mantha, Ph.D. R&D Director,
Green Signal BioPharma Private Ltd., Chennai for his encouragement and valuable
suggestions. I express my profound sense of gratitude to Dr. Srinivasan, Ph.D for his
valuable enthusiastic support and encouragement.
I also thank Mr.Shafi for his computer typing and designing.
Finally, no words of gratitude will be sufficient to pay sincere thanks to my
Mother whose inspiration and dedication helped me in finishing the project work
J. RAJESHKUMAR

CONTENTS
Page No.
1. INTRODUCTION 1
2. REVIEW OF LITERATURE 11
3. MATERIALS AND METHODS 66
4. RESULTS 82
5. DISCUSSION 138
6. SUMMARY AND CONCLUSION 159
7. REFERENCES

LIST OF TABLES
Table No.
Title
1 Rhizosphere Microbial Density.
2 Ratio between Rhizosphere microbes and PSB
3 Cultural, Morphological and biochemical characteristics of isolated bacteria
4 Efficacy of phosphate solubilization of UV treated Aspergillus niger
5 Efficacy of phosphate solubilization of UV treated Aspergillus fumigatus
6 Efficacy of phosphate solubilization of UV treated Penicillium sp
7 Efficacy of phosphate solubilization of UV treated Pseudomonas strains
8 Efficacy of phosphate solubilization of Sodium azide treated Aspergillus niger
9 Efficacy of phosphate solubilization of Sodium azide treated Aspergillus fumigatus
10 Efficacy of phosphate solubilization of Sodium azide treated Penicillium sp
11 Efficacy of phosphate solubilization of Sodium azide treated Pseudomonas strains
12 Efficacy of phosphate solubilization of Ethyl Methane Sulphonate (EMS) Aspergillus niger
13 Efficacy of phosphate solubilization of Ethyl Methane Sulphonate (EMS) Aspergillus fumigatus
14 Efficacy of phosphate solubilization of Ethyl Methane Sulphonate (EMS) treated Penicillium sp
15 Efficacy of phosphate solubilization of Ethyl Methane Sulphonate (EMS) treated Pseudomonas strains
16 Efficacy of phosphate solubilization of Aspergillus niger (ANems120) grown on different Carbon sources
17 Efficacy of phosphate solubilization of Aspergillus niger (ANems120) grown on different Nitrogen sources

Table No.
Title
18 Efficacy of phosphate solubilization of Aspergillus niger (ANsa120) grown on different Carbon sources
19 Efficacy of phosphate solubilization of Aspergillus niger (ANsa120) grown on different Nitrogen sources
20 Efficacy of phosphate solubilization of Penicillium sp.(PEsa150) grown on different Carbon sources
21 Efficacy of phosphate solubilization of Penicillium sp.(PEsa150) grown on different Nitrogen sources
22 Efficacy of phosphatase activity of UV treated Aspergillus niger
23 Efficacy of phosphatase activity of fungal strains UV treated Aspergillus fumigatus
24 Efficacy of phosphatase activity of Penicillium sp.
25 Efficacy of phosphatase activity of UV treated Pseudomonas sp.
26 Efficacy of phosphatase activity of Sodium azide treated Aspergillus niger strains
27 Efficacy of phosphatase activity of Sodium azide treated Aspergillus fumigatus strains
28 Efficacy of phosphatase activity of Sodium azide treated Penicillium sp.
29 Efficacy of phosphatase activity of Sodium azide treated Pseudomonas sp.
30 Efficacy of phosphatase activity of Ethyl Methane Sulphonate (EMS) Aspergillus niger
31 Efficacy of phosphatase activity of Ethyl Methane Sulphonate (EMS) treated Aspergillus fumigatus
32 Efficacy of phosphatase activity of Ethyl Methane Sulphonate (EMS) treated Penicillium sp.
33 Efficacy of phosphatase activity Ethyl Methane Sulphonate (EMS) treated Pseudomonas sp.
34 Efficacy of phosphatase activity of Aspergillus niger (ANems120) grown on different Carbon sources
35 Efficacy of phosphatase activity of Aspergillus niger (ANems120) grown on different Nitrogen sources
36 Efficacy of phosphatase activity of Aspergillus niger (ANsa120)grown on different Carbon sources

Table No.
Title
37 Efficacy of phosphatase activity of Aspergillus niger (ANsa120) grown on different Nitrogen sources
38 Efficacy of phosphatase activity of Penicillium sp.(PEsa150) grown on different Carbon sources
39 Efficacy of phosphatase activity of Penicillium sp.(PEsa150) grown on different Nitrogen sources
40 Efficacy of Lipase activity of UV treated Aspergillus niger
41 Efficacy of Lipase activity of UV treated Aspergillus fumigatus
42 Efficacy of Lipase activity of UV treated Penicillium sp
43 Efficacy of lipase activity of UV treated Pseudomonas sp.
44 Efficacy of lipase activity of Sodium azide treated Aspergillus niger
45 Efficacy of lipase activity of Sodium azide treated Aspergillus fumigatus
46 Efficacy of lipase activity of Sodium azide treated Penicillium sp.
47 Efficacy of lipase activity of Sodium azide treated Pseudomonas sp.
48 Efficacy of lipase activity of Ethyl Methane Sulphonate (EMS) treated Aspergillus niger
49 Efficacy of lipase activity of Ethyl Methane Sulphonate (EMS) treated Aspergillus fumigatus
50 Efficacy of lipase activity of Ethyl Methane Sulphonate (EMS) treated Penicillium sp
51 Efficacy of lipase activity of Ethyl Methane Sulphonate (EMS) treated Pseudomonas sp.
52 Efficacy of lipase activity of fungal strains grown on different Carbon sources
53 Efficacy of lipase activity of Bacterial strains grown on different Carbon sources
54 Efficacy of lipase activity of fungal strains grown on different Nitrogen sources
55 Efficacy of lipase activity of Bacterial strains grown on different Nitrogen sources
56 Screening for IAA by Wild and Mutated Bacterial and Fungal strains
57 Quantification study of IAA by Wild and Mutated Bacterial and Fungal strains

LIST OF FIGURES
Figure No
Title
1 Phosphate solubilization efficacy by wild strains
2 Phosphate solubilization efficacy by Fungal strains treated with chemical mutagen
3 Efficacy of phosphatase activity by wild strains
4 Efficacy of phosphatase activity by Fungal strains treated with chemical mutagen
5 Efficacy of Lipase activity by wild strains
6 Efficacy of Lipase activity by Chemical treated Fungal Strains
LIST OF PLATES
Plate No
Title
1 Biochemical Characterization of Bacterial culture
2 Growth of Fungal colonies on Sabouraud agar plates
3 Phosphate solubilization of Bacterial and Fungal cultures in Pikovskaya agar
4 Phosphate solubilization and phosphatase production by fungal cultures in Pikovskaya broth
5 Indole Acetic Acid production by bacterial and fungal cultures in Czepekdox broth and thin layer chromatogram for IAA

1
Introduction
Microorganisms play a very important role in almost every sector. One
can find applications (uses) of microorganisms in agriculture, forestry, food,
industry, medicine, and environment. The scope and significance of
microbiology has enlarged manifold, particularly when importance of
environment was realized globally and the word environment was used in a
much wider sense in terms of totality to include almost everything, every bit of
nature.
Besides being important in biogeochemical cycling of nutrients, microbes
play vital role in maintenance of soil fertility and in crop protection. Microbes
are being exploited in two important ways as biofertilizers, and creating new
nitrogen-fixing organisms. Most of the chemical reactions that take place in the
soil, leading to increased availability of several major and micronutrients often
have active contribution of microbes. The nitrogen-fixing bacteria, blue green
algae, and phosphate solubilizing bacteria are well known to enhance availability
of major nutritional elements like nitrogen and phosphorus to plants whereas the
decomposer bacteria are instrumental for recycling, and thereby increasing, the
availability of carbon and several micronutrients from plant residues to soil.
Phosphate Solubilization
Phosphorus has been called “the key of life” because it is directly
involved in most of the life processes. Next to nitrogen it is invariably classified
as one of the macronutrients and it is a key element in frequency of use as
fertilizer. It serves as a primary energy source for microbial oxidation. It is a

2
constituent substance in life processes. Soil cannot give high yields if it is
deficient in phosphate.
Soil contains both organic and inorganic forms of phosphorus. The
organic forms of the element in soil are compounds of phytins, phospholipids
and nucleic acids that come mainly by the way of decaying vegetation.
Plants take phosphate in the form of soluble orthophosphate ions but due
to the presence of Ca, Mg, K, Na, Al and Fe ions in soil, the soluble
orthophosphate is converted to insoluble form. Because of this process plants
utilize very little amount of phosphate, even though phosphorus containing
fertilizers are added (Vivekkumar et al., 1999).
This unmanaged excess of phosphate application is known to cause
environmental, economic problems and leads to pollution due to soil erosion and
runoff water containing large amounts of soluble phosphate (Brady, 1990). The
runoff from phosphate loaded soil is accepted as the main factor in
eutrophication of natural water reservoirs (Del Campillo et al., 1999).
Traditional phosphate fertilizer production is based on chemical processing of
insoluble mineral phosphate high-grade ore, which includes an energy intensive
treatment with sulfuric acid at high temperature. This process is environmentally
undesirable, not least because of the release of contaminants into the main
product, gas streams and byproducts. These contaminants must be dealt with as
potential air and water environmental pollutants (Vassilev and Vassileva, 2003).
Furthermore, use of phosphate fertilizers has become a costly affair and there is
a need for alternative sources.

3
Some microorganisms are known to be involved in the solubilization of
insoluble phosphates (Alexander, 1977). These phosphate-solubilizing
microorganisms render insoluble phosphate into soluble form through the
process of acidification, chelation and exchange reactions. This process not only
compensates for higher cost of manufacturing fertilizers in industry but also
mobilizes the fertilizers added to soil (Rodriguez and Reynaldo, 1999).
Therefore, many researchers have tried to increase the plant-available phosphate
fraction by means of Phosphate solubilizing microorganisms (PSMs) such as
Achromobacter sp, Agrobacterium sp, Alcaligens sp, Bacillus cereus,
B. polymyxa, B. megaterium, B. subtilis, Pseudomonas striata and Xanthomonas
sp and Fungi like Aspergillus niger, A. flavus, A. fumigatus, Penicillium sp and
Rhizopus sp.
Soil microorganisms are not only associated in the cycling of phosphorus
but also participate in solubilization of inorganic phosphorus and in the
mineralization of organic phosphorus (Agnihothori, 1970; Ostwal and Bhide,
1972).
Mechanism of Phosphate Solubilization
Different mechanisms have been suggested for the solubilization of
inorganic phosphorus by phosphate solubilizers. It is generally accepted that the
major mechanism of mineral phosphate solubilization is the action of organic
acids synthesized by soil microorganisms (Halder et al., 1990). Production of
organic acids results in acidification of the microbial cell and its surroundings.
Consequently, inorganic phosphate may be released from a mineral phosphate
by proton substitution for Calcium ion (Goldstein, 1994). The production of
organic acids by phosphate solubilizing bacteria has been well documented.
Among them, gluconic acid seems to be the most frequent agent of mineral

4
phosphate solubilization. It is reported as the principal organic acid produced by
phosphate solubilizing bacteria such as Pseudomonas sp., Erwinia herbicola,
Pseudomonas cepacia and Burkholderia cepacia. Another organic acid
identified in strains with phosphate-solubilizing ability is 2-ketogluconic acid,
which is present in Rhizobium leguminosarum, Rhizobium meliloti, Bacillus
firmus, Strains of Bacillus liqueniformis and Bacillus amyloliquefaciens were
found to produce mixtures of lactic, isovaleric, isobutyric, and acetic acids.
Other organic acids, such as glycolic, oxalic, malonic, and succinic acid, have
also been identified among phosphate solubilizers. (Rodriguez and Fraga,
1999).
There is also experimental evidence that supports the role of organic acids
in mineral phosphate solubilization. Halder et al. (1990) showed that the organic
acids isolated from a culture of Rhizobium leguminosarum solubilized an amount
of P nearly equivalent to the amount that was solubilized by the whole culture.
Besides this, treatment of the culture filtrates from several Rhizobium strains
with pepsin or removal of proteins by acetone precipitation did not affect
phosphate release capacity, showing that this was not an enzymatic process.
However, neutralization with NaOH destroyed the solubilization activity.
Alternative possibilities, other than organic acids for mineral phosphate
solubilization have been proposed based on the lack of a linear correlation
between pH and the amount of solubilized phosphate. In addition, no significant
amounts of organic acid production could be detected from a phosphate
solubilizing fungus, Penicillium sp. Studies have shown that the release of
hydrogen ions to the outer surface in exchange for cation uptake or with the help
of hydrogen ion translocation ATPase could constitute alternative ways for
solubilization of mineral phosphates.

5
Other mechanisms have been considered, such as the production of
chelating substances by microorganisms as well as the production of inorganic
acids, such as sulfuric, nitric, and carbonic acid. However, the effectiveness of
these processes has been questioned and their contribution to phosphate release
in soil appears to be negligible. The phosphatase enzyme plays a key role in
transforming organic forms of Phosphate in to plant available inorganic form.
They provide to be active in all three components of the rhizosphere soil,
microbes and plant root. There are two types of phosphatase, according to their
pH optima.
Both acid and alkaline phosphatase have been found as external and
internal enzyme in algae and bacteria. Alkaline phosphatase are metalo proteins.
The organic function of soil phosphorus has received relatively little attention
because of its complex nature, although this fraction may account for 15-20% of
phosphate on soil. In order to become available these phosphate compounds
must be hydrolyzed by phosphatase. This enzyme may therefore be very
important in phosphorus nutrition of plants (Tarafdar et al., 1992). Chelating
substances and inorganic acids such as sulfuric, nitric, and carbonic acid are
considered as other mechanisms for phosphate solubilization. However the
effectiveness and their contribution to P release in soils seems to be less than
organic acid production.
Lipase
Lipases are the enzymes that catalyze the hydrolysis of triglycerides to
glycerol and fatty acids. Microbial lipases are relatively stable and are capable of
catalyzing a variety of reactions. They are potential important for diverse
industrial applications.

6
Degreasing is an essential step in the production of glove and clothing
leather. Degreasing helps to obtain soft and pliable leather for garment
manufacture. Degreasing is carried out by emulsification with detergents or by
solvent extraction. It is well known that organic solvents like kerosene, petrol,
perchloroethylene and trichloroethylene are highly unsafe, hazardous to the
workers and heavily pollute the environment. Lipase which are projected as
alternatives for solvents and detergents, catalyse the breakdown of fats and can
be obtained from animal, microbial and plant sources. (Kamini et al., 1999).
Lipase enzymes play a vital role in many fields such as food, dairy
Leather, paper, pharmaceutical, detergent, textile and cosmetic industries. Lipase
are secreted by microorganisms like bacteria, yeasts, molds and a few protozoa.
The production of lipase by microorganisms depends largely on the species,
strains and culture conditions. Microbial lipases are diverse in their enzymatic
properties, substrate specificity and are usually more thermo stable than animal
or plant lipases (Vanitha, 2002).
Microbial lipases represent the major commercial source of this enzyme.
In recent years, research on lipases, mainly of microbial origin, has increased
because of their great commercial potential and also due to the potential of this
enzyme to both hydrolyze fats and synthesize specific esters with desirable
flavor properties for industrial application. Lipases are of considerable
commercial and industrial potential. Furthermore there is an increasing interest
in the development of new application for these enzymes in products and
processes (Paul and Carles, 1992).

7
Indole Acetic Acid
Plant growth promoting rhizobacteria are considered to promote plant
growth directly or indirectly. They can exhibit a variety of characteristics
responsible for influencing plant growth. Plant hormones and other regulatory
chemicals are now used in a variety of applications for commercial reasons to
control some aspects of plant development. Auxins were the first plant hormones
to be discovered. The principle auxin in plants is indole-3-acetic acid (Yurekli et
al., 2003).
Diverse soil microorganisms including bacteria, fungi and algae are
capable of producing physiologically active quantities of auxins, which may
exert pronounced effects on plant growth and establishment. Many of them can
produce auxins in axenic cultures. Most of the species use tryptophan to produce
indole-3-acetic acid (IAA), mainly through the indole-3-pyruvic acid and
tryptamine pathways (Tudzynski and Sharon. 2002).
For many years it was assumed that tryptophan (Trp) was the only
precursor of IAA. However, work with tryptophan-auxotrophic mutants and
isotope labeling has established that IAA biosynthesis can occur via a
tryptophan- independent route also (Normanly, 1997; Venis and Napier, 1991),
although in the presence of Trp, microbes release greater quantities of IAA and
related compounds. There is firm evidence that indole-3-acetic acid (IAA)
(Arshad and Frankenberger, 1991; Sarwar and Frankenberger, 1994; Barea and
Brown, 1974; Brown, 1972; Brown and Burlingham, 1968; Lee et al., 1970;
Scott, 1972), gibbe-rellins and cytokinins (Sarwar and Frankenberger, 1994;
Barea and Brown, 1974), all produced by plants and essential to their growth and
development, are produced also by various bacteria which live in association
with plants. There is also evidence that the growth hormones produced by the

8
bacteria can in some instances increase growth rates and improve yields of the
host plants (Arshad and Frankenberger, 1991; Sarwar and Frankenberger, 1994;
Barea and Brown, 1974). It is possible that bacteria capable of phosphate
solubilization may improve plant productivity both by hormonal stimulation and
by supplying phosphate.
It is generally agreed that indole-3-acetic acid (IAA) is the major and
most abundant auxin in plants and plays a key role in the regulation of plant
growth and development (Moore, 1989, Luthen et al., 1999 and Davies, 1995). It
is presumed that plant growth promoting rhizobacteria producing plant growth
regulators play a critical role in plant growth promotion. To assess this
hypothesis, local isolates of phosphate solubilizing microbes were screened for
their intrinsic ability to produce IAA in the presence and absence of L-
tryptophan (Shahab et al., 2009).
Mutation
UV irradiation and Ethyl Methane Sulphonate (EMS) have long been
used as a model DNA damaging agent to study not only UV specific repair
processes but also general repair and DNA damage tolerance pathways. Several
UV sensitive mutants have been found to be defective in both nucleotide
excision repair and in DNA damage tolerance pathways. The mutants can
provide valuable insights into DNA repair process and mechanism of
mutagenesis (Srinorakutara et al., 2008).
Mutation alters the genotype of microorganisms, when it expresses that
leads to alter the character or death of microorganisms. The Ultra Violet
radiation forms thymine dimer in gene sequence. But the photolyase enzymes
present in living system break the thymine dimer and correct it. The increasing

9
exposure time to UV radiation may form the thymine dimer in gene sequence
that code photolyase enzyme. In this situation the thymine dimer can not break
by the enzyme of living system. The chemical agents such as Sodium Azide,
Ethyl Methane Sulphonate may alter the gene sequence by altering the base
pairs. The azide ion alters the structure of cytosine such that it forms hydrogen
bonds with adenine, rather than guanine. This produces a C -> T transition. Ethyl
Methane Sulphonate is a strong mutagenic agent. It alkylates N7 of Guanine and
severely alters the base pairing.
In the recent past, a variety of studies are conducted to improve the
productivity of enzymes and microbial conversion process using random
mutagenesis, typically which involve production of microbial enzymes, plant
hormones and phosphate solubilization.
The potential mechanism for phosphate solubilization might be
acidification either by proton extrusion associated with ammonium assimilation
(De Freitas et al., 1997 and Reyes et al., 1999) or by organic acid production
(Cunningham and Kuiack, 1992). Acid phosphatases and phytases secreted by
these microorganisms also have an important role in phosphate solubilization
(Richardson et al., 2000). Reyes et al. (1999) isolated the mutants of Penicillium
rugulosum which showed high phosphate solubilizing activity compared to wild
type strain. A significant increase in soluble phosphate level was observed in
case of UV induced mutants of A. tubingensis compared with the wild type.
There might be a possibility of alteration at genetic level in case of mutants
(Achal et al., 2007).
In the present research work, the bacterial and fungal strains were tried to
improve for their phosphate solubilizing capacity by physical and chemical

10
treatments. Their ability in the production of phosphatase, lipase and Indole
Acetic Acid were analysed. The effect of different carbon and nitrogen sources
on the production of phosphatase, lipase and on phosphate solubilization were
studied.

11
Review of Literature
In recent years, attention has been paid to the microbial products, which
have significant agricultural value. Microorganisms produce secondary
metabolites and some of which have phosphate solubilizing capabilities. Some
of the microorganisms are isolated from the soil and their phosphate
solubilization capabilities were improved by using physical and chemical
mutagenesis.
Phosphate solubilization
Soil generally contains adequate amount of organic and inorganic
phosphorus. But most of these remain unavailable to plants. The efficiency of
water soluble phosphorus is usually low and it’s recovery does not exceed 20%
(Darmwal et al., 1989). But by the help of phosphate solubilizers, phosphate
availability in soil and the utilization of phosphate by plants are increased.
(Gaur and Sacher, 1980). Microbial solubilization of insoluble phosphate,
especially low grade and it’s use in agriculture is receiving greater attention.
This process not only compensates for higher cost of manufacturing fertilizers in
industry but also mobilizes the fertilizers added to the soil.
Isolation and identification
Johnston (1954) isolated fungal cultures from rhizosphere soil and
identified as Aspergillus niger, A. terreus and Penicillum digitatum based on
their spore structure and colony morphology. A number of reports are available
related to phosphate solubilization in different soils. Sen and Paul (1957)
reported that the solubilization of calcium phosphate and iron phosphate in
liquid media by four Bacillus sp, which were isolated from the glands of Cassia
occidentalis. Casida (1959) isolated phosphate solubilizing fungi Aspergillus

12
niger, A. flavus, A. nidulans, A. terreus and Penicillium liliacinum from
rhizosphere soil. Chhonokar and Subba Rao (1967) isolated fungal cultures from
legume root nodules and identified as Aspergillus flavus, A. niger and A.
nidulans based on their spore structure and colony morphology.
Ahamed and Jha (1968) isolated Phosphate Solubilizing Bacteria, fungi
and actinomyces from soil samples of Bihar on carrot extract agar. From their
studies, it is identified that bacteria were more in number than other types of
microbes whereas, the fungi performed better solubilization than other types of
microbes. Bhurat and Sen (1968) examined the leaf surface of wheat, pea, barley
and gram for the presence of Phosphate Solubilizing Bacteria and isolated nearly
15 isolates of various species.
Sethi and Subba Rao (1968) screened 48 isolates of fungi from Delhi and
Ludhiana soils for their potentialities to solubilize tri calcium phosphate and
calcium phosphate in culture media. The results indicated that about seventeen
species belonging to genera like Aspergillus, Penicillum, Cladosporium,
Fusarium, and Pacecilomyces were found to be significantly effective. Metha
and Bhide (1970) selected 42 isolates with solubilization capacity of Tri Calcium
Phosphate in culture out of 149 soil fungal cultures. Most efficient solubilizers
were of Aspergillus sp., Penicillum sp., Pythium sp., Curvularia sp.,
Chaetomium sp. and Humicola sp.
Agnihothori (1970) evaluated phosphate solubilizing potential of several
fungi isolated from forest tree seedbeds. Aspergillus niger, A. flavus, Fusarium
oxysporum, Sclerotium roltsii, Clindracladium sp. and Penicillium sp. were
significant to solubilize phosphate. Gaur et al. (1973) isolated Aspergillus
carbonum, Aspergillus flavus, Aspergillus fumigatous, and Aspergillus wentii

13
from Mussourie, Jhamarkotra and Maton Rock Phosphates and studied the
solubilization capacity. Rao et al. (1982) isolated phosphate dissolving
actinomycetes from soil and this organism was identified as Streptomyces
species from its slide culture.
Venkateswaralu et al. (1984) isolated phosphate-solubilizing
actinomycetes from orchid soils using modified Pikovskaya medium. Singh et
al. (1984) identified phosphate solubilizing fungal cultures viz., Aspergillus
awamori and A. niger based on their colony color, colony morphology and
spore structure. Darmwal et al. (1989) isolated phosphate solubilizing fungal
cultures from different samples of rhizosphere of wheat, gram, garden soils and
composed materials. Thakker et al. (1993) isolated phosphate solubilizing gram
positive and gram negative bacteria from different crops and compost samples
and identified the pure culture on the basis of cell morphology, staining reaction,
biochemical and physiological characteristics as outlined in 8th edition of
Bergey’s Manual of Determinative and Systemic Bacteriology.
Phosphate solubilization assay
Arora and Gaur (1979) inoculated the bacterial and fungal culture into
Pikovskaya broth and after incubation the water-soluble phosphate in the
supernatant was estimated by the King’s B method improved by Sherman with
Klett-Sommerson colorimeter using red filter. Thakker et al. (1993) inoculated
the broth culture containing phosphate-solubilizing bacteria, adjusted with
optical density 1.0 into pikovskaya broth. The phosphate solubilization was
assayed interval of 3 days by chlorostannous reduced molybdophosphoric acid
method.

14
Varshanarishan et al. (1995) cut 80mm mycelial disc (=75 x 106 spores
per ml) from four days old culture, grown on Czapek’s dox agar by sterilized
cork borer under aseptic condition into 50ml sterile pikovskaya broth. After
incubation, the water soluble phosphorus in the culture filtrate was estimated
every 24 hours up to 7days by chlorostannous reduced molybdophosphoric acid
method. Seshadri et al. (2000) inoculated a single colony of Azospirillum
halopraeferans from nutrient agar to sterile broth containing water insoluble
phosphorus. After incubation, the cultures were harvested on alternate days,
centrifuged at 10, 000 rpm for 15 minutes and then subjected for phosphate
estimation by paramolybdate blue method.
Metabolic mechanism of Phosphate solubilization
The potential mechanism for phosphate solubilization might be
acidification either by proton extrusion associated with ammonium assimilation
(De Freitas et al., 1997; Reyes et al., 1999) or by organic acid production
(Cunningham and Kuiack, 1992). Acid phosphatase and phytases secreted by
microorganisms also have an important role in phosphate solubilization
(Richardson et al., 2000).
Katznelson and Bose (1959) isolated rhizosphere bacteria from wheat,
that oxidized Glucose and alanine and they observed over one third of cultures
tested were capable of dissolving insoluble phosphate in the form of calcium
phosphate. Goswami and Sen (1962) isolated three bacterial strains and
examined for their Tri Calcium Phosphate solubilizing activity in Pikovskaya
broth.
Kucey (1984) Phosphate-solubilizing, total bacterial and fungal
populations were determined by serial dilution and plate counting. He reported

15
that the fungi were superior than bacteria in solubilizing both freshly precipitated
calcium phosphate and Idaho Rock Phosphate. Fungi also retained this ability
over many sub culturing transfers. A high percentage of the bacterial isolates lost
their solubilizing ability when sub cultured. A significant correlation was found
between an organism's ability to solubilize freshly precipitated calcium
phosphate in agar plates and Idaho Rock Phosphate in solution culture.
Cerezine et al. (1988) grew the fungus Aspergillus niger in a stationary
culture containing modified citrate medium supplemented with fluoroapatite.
Solubilization of insoluble phosphate was increased with fungi growth, reaching
maximum after 11 days of the inoculation. Soluble phosphate levels were
correlated with pH of the culture medium but not with titrable acidity values,
probably due to metabolic activity of fungus resulting from consumption of
sugar in the culture medium, They also studied the effect of carbon and nitrogen
source on phosphate solubilization.
Halder et al. (1991) also studied the phosphate solubilization efficacy of
Bradirhizobium strains by using different amount of phosphates of both
hydroxyapitite and tri-calcium phosphate in liquid cultures. Phosphate
solubilization was related to pH decrease caused by the strains. Nitrogen in the
form of ammonium effected high solubilization of phosphate in the medium.
However yeast Extract of nitrate was also remarkably effective as a nitrogen
source to support PO4 solubilization, Glucose was best carbon source for the
process. Two fungal isolates, Penicillium bilaji and Penicillium fuscum were
found to solubilize different amounts of Rock Phosphate in liquid culture and the
relationship between inorganic P solubilization and showed pH drop by each
isolate. The also reported the role of Nitrogen in P solubilization metabolism of
P. bilaji. (Asea et al., 1988).

16
Cunningham and Kuiack (1992) isolated Penicillium bilaii to study the
solubilization of mineral phosphates and enhance plant uptake of phosphate.
Using agar media with calcium phosphate and the pH indicator alizarin red S,
the influence of the medium composition on phosphate solubility and medium
acidification was recorded. Three gram positive and four gram negative
Phosphate Solubilizing Bacteria were isolated from rhizosphere of different
crops and compost samples. Among the bacteria, Enterobacter aerogens, was
found to be most effective tri Calcium Phosphate (TCP) & Rock Phosphate
solubilizer. In presence of tri Calcium Phosphate and Rock Phosphate, the effect
of carbon, nitrogen sources and pH of the medium on phosphate solubilization
were also studied. (Thakkar et al. 1993).
Singal et al. (1994) Aspergillus japonicus and A. fetidus were found to
solubilize five types of Indian Rock Phosphates at pH 8 and 9. Solubilization
was higher in the presence of pyrite than in controls lacking either pyrite or
fungal inoculum. Both the Aspergillus were found to be good pyrite solubilizers
and could grow over a wide pH range. Solubilization of Rock Phosphates was
the result of organic acid release and pyrite oxidation. Aspergillus aculeatus
isolated from rhizosphere of gram, Solubilized Phosphate from TCP in
Pikovskaya Medium. The phosphate solubilizing activity was highest after 48h
of fungal growth in the presence of Glucose and ammonium sulphate as the best
carbon and nitrogen sources. They also reported the growth of the
microorganisms and the pH of the growth medium showed no correlation with
phosphate solubilizing activity (Varsanarsian et al.1995).
Two species like Penicillium aurantiogriseum and Pseudomonas sp.
having high abilities in solubilizing inorganic phosphates (hydroxylapatite and

17
brushite) were used to examine solubilization mechanisms. No direct contact
between microorganisms and Calcium phosphates (Ca-Ps) were necessary for
effective solubilization (Illmer and Schinner, 1995).
Vassilev et al. (1996) successfully cultivated phosphate solubilizing
Aspergillus niger on sugar beet-waste material supplemented with Rock
Phosphate. Nine Streptosporangium isolates of Eudilius eugeniae (earthworm)
casts were found to be acid tolerant, Rock Phosphate solubilizers which could
grow on synthetic Glucose or carboxyl methyl-cellulose nitrogen free or
ammonium chloride enriched media, as the sole carbon source. These isolates
could be exploited in the industrial production of microbial phosphate fertilizers
which would enhance organic residues and plant nutrients recycling in acid soils
of Nigeria. (Mba, 1996).
Two soil types, loamy sand and sandy soils, were treated with atrazine,
pyrethrin and a mixture of metobromuron and metolachor for eight weeks in the
laboratory to determine the effect of the chemicals on soil microbial populations
and their mineralization activities (Taiwo and Oso, 1997).
Aspergillus niger, citric acid producing strain grown on olive cake-
based medium, was able to solubilize Rock Phosphate. Solubilization of
insoluble phosphate increased during the solid-state fermentation process.
Various combinations of olive cake and Rock Phosphate, previously treated or
untreated by the fungus, were introduced into a calcareous, phosphorus
deficient soil to improve the growth of Trifolium repens in a greenhouse
experiment. Greater growth and phosphate uptake of Mycorrhizal and Non-
Mycorrhizal plants were achieved when microbe treated olive cake and Rock

18
Phosphate were applied to soil compared with all other treatments. (Vassileva
et al., 1997).
The interactive effect of phosphate-solubilizing bacteria and Arbuscular
Mycorrhizal fungi on plant use of low bioavailable soil phosphate sources was
evaluated using soil microorganisms which integrated isotopic dilution
techniques. The microbial inoculum consisted of the Arbuscular Mycorrhizal
fungus Glomus intraradices and two phosphate-solubilizing rhizobacterial
isolates Enterobacter sp. and Bacillus subtilis. The Mycorrhizosphere
interactions between bacterial and fungal plant associates was identified and
it’s contribution to the biogeochemical phosphorus cycling was also
investigated. (Toro et al., 1997).
The effect of immobilization technique on phosphate-solubilization by
Enterobacter sp., was reported. Various amounts of the immobilized
bioparticles were applied in a repeated batch fermentation process in order to
solubilize Venezuelan Rock Phosphate. Phosphate solubilization was
significantly higher compared to a free-cell single-batch control. Whole and
destroyed immobilized bioparticles were introduced into a soil enriched with
Rock Phosphate to improve the growth of onion plants. (Vassileve et al 1998).
Spores of Aspergillus niger were encapsulated in Agar, Calcium Alginate
and Potassium Carrageenan and the encapsulated-fungus were tested for it’s
phosphate solubilizing capability using the culture medium supplemented with
different concentrations of Rock Phosphate. The highest average soluble
phosphate concentration was obtained with agar-cell beads as compared with
other encapsulated systems (Vassileve et al. 1998).

19
Kim et al. (1998) examined the phosphate solubilization capability of
Enterobacter agglomerans by organic energy source in unsterilized soil. After
the enrichment of organic energy source, they examined the phosphatase activity
and available phosphate concentration. Pany et al. (1998) collected surface soil
samples, treated with different phosphate sources and analyzed for microbial
population with respect to total bacteria and Phosphate Solubilizing Bacteria.
Anusuya and Jayarajan (1998) identified Trichoderma viride fungal culture
isolated from rhizosphere soil and compare the phosphate solubilizing capacity
of the fungus against Bacillus sp.
Pseudomonas cepacia is known as a Rock Phosphate solubilizer in
bioreactors and in soils. BarYosef et al. (1999) determine the production rates of
gluconic acid and 2-ketogluconic acid by the bacteria in the presence of clay
minerals which prevail in soils, and the resulting rate and extent of
orthophosphate release into the suspension solutions. An isolate of A. niger was
found to be very effective in solubilizing Rock Phosphate. The influence of
nutritional requirements such as carbon sources, nitrogen sources and different
concentrations of KH2PO4 on Rock Phosphate solubilization was extensively
studied. Moreover, the effect of heavy metals, such as Mn2+, Co2+, Zn2+, Cu2+ as
metal chloride and Al3+ as aluminium sulphate, on the solubilization rate of Rock
Phosphate was illustrated. A biotic effect of organic acids such as citric and
oxalic acids, as major metabolites of A. niger, on the solubilization of Rock
Phosphate, was studied. The obtained results revealed that there is a relationship
between the rate of solubilization of Rock Phosphate and the extracellular
exudates including organic acids, alkaline and acid phosphatase.
Penicillium radicum, a phosphate-solubilizing fungus isolated from the
rhizosphere of wheat roots and its ability to solubilize inorganic phosphate was

20
studied in vitro. The fungus was grown in liquid medium cultures containing
either ammonium or nitrate as the sole source of nitrogen. The titratable acidity,
pH and concentrations of organic acids and soluble phosphate were determined
periodically during incubation. Phosphate solubilization was generally higher
when ammonium rather than nitrate was the sole source of nitrogen. Soluble
phosphate concentrations in the culture medium were directly proportional to the
titrable acidity and organic acid concentration and inversely related to pH.
(Whitelaw et al., 1999).
Sudhansupal (1999) isolated 23 Phosphate Solubilizing Bacterial cultures.
Of the 23 isolates the acid tolerant strain was identified as Bacillus sp on the
basis of morphological and biochemical tests as described in Bacteriological
manual. Artidave and Patel (1999) isolated Phosphate Solubilizing Bacteria
from soil. They identified the bacteria up to genus level by positive test of the
organism for pigment production on King’s B medium and by cultural,
morphological and biochemical characteristics. Based on the above results and
oxidase positive reaction in King’s B medium, they identified the isolated
culture was Pseudomonades.
Phosphate-solubilizing bacteria and solubilization of Mussoorie
phosphate rock were examined in simulated fish ponds enriched with compost
and exogenous introduction of phosphate-solubilizing bacteria. They found that
the combined effects of the Phosphate-solubilizing bacterial population of both
exogenous and compost origin with short generation time. The relationship
between the phosphorus level of water and Phosphate-solubilizing bacteria
population was expressed in exponential equations (Sahu and Jana, 2000).
Vazquez et al. (2000) isolated thirteen Phosphate Solubilizing Bacterial strains
from rhizosphere of mangroves using culture media containing tri basic calcium

21
phosphate and they analyzed the bacterial culture for the production of organic
acids.
Shekhar et al. (2000) isolated Phosphate Solubilizing Bacterial strains
from the rhizosphere of chickpea and alkaline soils. These bacterial strains were
screened for the phosphate solubilizing ability in the presence of 10 percentage
salt, pH 12 and temperature 45oC. Four strains of Phosphate Solubilizing
Bacteria were isolated from the rhizosphere of chickpea and alkaline soils. All
four strains demonstrated diverse levels of phosphate solubilization activity
under in vitro conditions in the presence of various carbon and nitrogen sources.
Acid production may have contributed to phosphate solubilization, but was not
the only reason for phosphate release into the medium. The strains showed
varied levels of phosphate solubilization when the effects of different sources of
nitrogen were examined during growth (Nautiyal et al., 2000).
Fenice et al. (2000) studied the relationship between gluconic acid
production and phosphate solubilization by the encapsulated fugal strain
Penicillium variabile. They also reported that the degree of phosphate
solubilization was not influenced by decreasing the Glucose concentration in the
cultivation medium.
Vassileve et al. (2001) examined phosphate solubilization and organic
acid production by using immobilized microbial cells. The immobilized cells
showed higher degree of phosphate solubilization and organic acid production
than free cells. Kang et al. (2002) screened for phosphate solubilizing fungal
strains in the field soil at Taegu, South Korea. Such strains were identified as
Fomitopsis sp. Phosphate solubilizing ability of Fomitopsis sp. was studied on
four different insoluble phosphates, viz. Tri Calcium Phosphate, Rock

22
Phosphate, Aluminium phosphate and Hydroxyapatite. Tri Calcium Phosphate
was found to be solubilized maximally, while Hydroxyapatite could not be
solubilized by the isolated fungal strain. Further, the effect of salinity under in
vitro conditions on the solubilization activity of Rock Phosphate was also
observed.
Three isolates of Aspergillus tubingensis and two isolates of Aspergillus
niger isolated from rhizospheric soils were tested on solubilization of different
Rock Phosphates. All the isolates of Aspergillus were capable of solubilizing all
the natural Rock Phosphates. A. tubingensis showed maximum percent of
solubilization in all the Rock Phosphates tested in this study when compared to
other isolates. (Reddy et al. 2002). Pseudomonas corrugata, a soil isolate,
initially obtained from a temperate location in Sikkim, was examined for its Tri
Calcium Phosphate solubilizing ability along a wide temperature range, from
psychrophillic to mesophillic. The study has implications for developing carrier-
based microbial inoculants for improved growth of plants in the mountains
(Pandey et al. 2002).
A bacterial strain isolated from rhizospheric soil of grasses growing
spontaneously in Spanish soil, actively solubilized phosphates in vitro when
calcium phosphate was used as a phosphorus source. This strain was Gram-
negative, strictly aerobic, rod-shaped and motile. The strain produced catalase,
but not oxidase. Cellulose, Casein, Starch, Gelatin, Aesculin and Urea were not
hydrolyzed. Growth was observed with many carbohydrates as the carbon
source. (Peix et al. 2003).
The Mineral Phosphate Solubilization (MPS) was studied in ten
Aspergillus niger strains. MPS activity was measured in solid (Pikovskaya's

23
medium) as well as liquid media using different phosphate sources, carbon
sources and nitrogen sources All the strains showed a zone of clearance of
Tricalcium Phosphate in Pikovskaya's medium in plates and solubilized
Dicalcium and Tricalcium Phosphates in broth efficiently. Among the carbon
sources Aspergillus niger preferred mannitol for higher phosphate solubilization.
Nitrogen in the form of nitrate was very effective in solubilizing inorganic
phosphates. Xylose and urea were the poorest sources of carbon and nitrogen for
all the strains of Aspergillus. Phosphate release was associated with reduction in
pH (Seshadri et al., 2004).
A phosphate-solubilizing microorganism isolated from rhizospheric soil
and temporarily identified as Burkholderia glathei produced gluconate and
acetate using Glucose as a carbon source and its metabolic activity caused the
pH of the liquid medium to decrease as low as 4.4. Whole-cell fatty acids
methyl ester profile and 16S rDNA sequence analysis were employed to isolate
and identify the bacterial groups that actively solubilized phosphates in vitro
from rhizosphere soil of various crops of Korea. Out of several hundred colonies
that grew on Pikovskaya’s medium 13 best isolates were selected based on the
solubilization of insoluble phosphates in liquid culture and further characterized
and identified (Chung et al. 2005).
The fungal strains were isolated from agriculture soil, having potential to
solubilize insoluble inorganic phosphates were characterized. Two fungal
isolates were tested for their Tricalcium Phosphate solubilization efficiency in
both solid and liquid medium. Isolates were identified as Aspergillus sp. and
Penicillium sp. Phosphate solubilization activity in liquid broth culture in
presence of various carbon and nitrogen sources, relation between pH and
phosphate solubilization were recorded (Pradhan and Sukla, 2005).

24
The phosphate-solubilizing bacterium was isolated from soil in Pingdong
and Taiwan and identified as B. cepacia based on its 16S ribosomal DNA
(rDNA) sequence. The supernatant was filtered through a 0.22 µm Millipore
filter and assessed for bacterial population, pH, organic acids, and phosphate
solubilizing capability (Lin et al., 2006). The phosphate solubilizing fungi were
isolated to assess the solubilization ability of phosphate fractions in various soil.
The highest numbers of isolates with high solubilization capacity were detected
in pasture soil, followed by tropical rain forest and forest patch soils. Pasture soil
presented both the largest contents of insoluble phosphates and the largest
number of fungal isolates with phosphate-solubilizing ability. The range and size
of phosphate fractions influenced the number of fungi and their ability to
solubilize hardly soluble phosphates (Barroso et al., 2006).
The possible action of Phosphate Solubilizing Bacteria on the
leguminous–rhizobia symbiosis was studied in the region where the available
phosphorus distribution is not uniform. Sinorhizobium meliloti,
Bradyrhizobium japonicum and two phosphorus-solubilizing strains of
Pseudomonas putida were used for growth promotion treatments. (Rosas et al.,
2006). Fankem et al. (2006) collected seven rhizosphere soil samples from oil
palm tree of Cameroon. The collected soil samples were air dried, crushed to
pass through 2mm sieves and the Phosphate Solubilizing Bacteria were isolated
by using the soil samples. For the estimation of phosphate solubilization Reyas
basal medium having Calcium, Aluminum and Iron phosphates was used. At the
end of incubation time, it appeared that, phosphate solubilization resulted from a
combined effect of pH decrease of the media and organic acids production.
Furthermore, each of the tested isolates was able to produce at least one of the
most important organic acids such as Citrate, Malate and Tartrate. Among the

25
ten isolates tested, three were identified as Pseudomonas fluorescens and would
be considered as potential biofertilizers.
To develop environment friendly biofertilizer, solubilizing insoluble
phosphates, salt- and pH-tolerant, insoluble inorganic phosphate solubilizing
bacterium was isolated from soybean rhizosphere. On the basis of its
physiological and biochemical characteristics, this bacterium was identified as
Pantoea agglomerans. Glucose medium consisting of ammonium nitrate was the
optimal and best medium for phosphate solubilization (Son et al., 2006). Two
phosphate and potassium solubilizing strains were isolated from the soil of
Tianmu Mountain, Zhejiang Province (China) and they were phenotypically and
phylogenetically characterized (Hu et al., 2006).
The isolates from different composts such as farm waste compost, rice
straw compost, Gliricidia vermicompost, and macrofauna, showed Rock
Phosphate solubilization in buffered medium in plate culture. When tested in
Rock Phosphate broth medium, all the isolates, Enterobacter cloacae, Serratia
marcescens, Serratia sp, Pseudomonas sp., and Pseudomonas sp. showed
gluconic acid production and solubilized Rock Phosphate. In the presence of
different carbon sources, cellulose degrading and Phosphate solubilizing strains
showed a drop in pH and solubilized Rock Phosphate. Significantly, these
bacteria isolated from composts and macrofauna solubilized Rock Phosphate in
the presence of various pure carbon substrates and crop residues and their
importance in soil/rhizosphere conditions are discussed. (Hameeda et al., 2006).
A field experiment has been conducted with four systemic herbicides at
their recommended field rates to investigate their effects on growth and
activities of aerobic non-symbiotic N2-fixing bacteria and Phosphate
Solubilizing Microorganisms in relation to availability of nitrogen and

26
phosphorus in the rhizosphere soils as well as yield of the rice crop. Application
of herbicides, in general, highly stimulated the population and activities of the
target microorganisms, which resulted in a greater amount of atmospheric
nitrogen fixation and phosphate solubilization in the rhizosphere soils of the test
crop. (Das and Debnath, 2006).
Chen et al. (2006) isolated, screened and characterized 36 strains of
Phosphate Solubilizing Bacteria (PSB) from Central Taiwan. Mineral
Phosphate Solubilizing (MPS) activities of all isolates were tested on tri
calcium phosphate medium by analyzing the soluble phosphate content after
72 h of incubation at 30 °C. Biosolubilization of Rock Phosphate using a
Penicillium sp., an Aspergillus sp., Pleurotus ostreatus, Bradyrhizobium
elkanii and their fungal–rhizobial biofilms was investigated. The study
identified an effective method of fungal–rhizobial biofilm mediated
solubilization of Rock Phophate. (Jayasinghearachchi and Seneviratne, 2006).
The phosphate solubilizing fungi Aspergillius spp., Penicillium spp. and
Fusarium spp. and bacteria B.subtilis, and B.megatherium were collected from
saline affected area of Amravati district. The strains were used to reduce the
salinity of soil by utilizing their organic acid metabolic activity. (Rajankar et
al., 2007). Vassilev et al. (2007) utilized dry olive waste, as a substrate for
phytase production and Rock Phosphate solubilization by Citric Acid
producing Aspergillus niger. Corn steep liquor, Yeast Extract and Ammonium
Nitrate were used as nitrogen source for phosphate solubilization. Both
enzyme production and phosphate solubilization depended on water medium
content, type of nitrogen source, inoculum size and the presence and initial
concentration of Phosphate in the medium.

27
Perez et al. (2007) conducted a survey of Phosphate-solubilizing bacteria
naturally colonizing a limonitic crust in the south-east region of Venezuela. A
total of 130 heterotrophic bacterial isolates showing different degrees of mineral
Tri Calcium Phosphate (Ca3 (PO4)2) solubilizing activities were isolated and
their phosphate solubilizing efficacy were detected with different phosphate
source. The 10 best Ca3 (PO4)2 solubilizers were characterized by partial
sequencing analysis of their respective 16S rRNA genes.
Eupenicillium parvum, a phosphate-solubilizing microorganism was
isolated from the tea rhizosphere. The fungus developed a phosphate
solubilization zone on modified Pikovskaya agar, supplemented with tri calcium
phosphate. Quantitative estimation of phosphate solubilization in Pikovskaya
broth showed high solubilization of Tri Calcium Phosphate and Aluminium
phosphate. The fungus also solubilized north Carolina Rock Phosphate and
Mussoorie Rock Phosphate, and exhibited high levels of tolerance against
desiccation, acidity, salinity, Aluminium, and Iron. Solubilization of inorganic
phosphates by the fungus was also observed under high stress levels of
Aluminium, Iron, and desiccation, though the significant decline in phosphate
solubilization was marked in the presence of Aluminium than Iron (Vyas et al.
2007).
Strain Paecilomyces marquandii was isolated from soil deficient in
phosphate on Pikovskaya’s medium. To study the effect of different carbon
sources on phosphate solubilization, Pikovskaya medium was modified where tri
calcium phosphate was replaced with other phosphate source and Glucose was
replaced with 9 individual carbon compounds viz. Fructose, Galactose, Glycerol,
Lactose, Maltose, Mannose, Sorbitol, Starch and Sucrose. The nitrogen sources
were evaluated similarly by replacing Ammonium Sulphate with six different
nitrogen sources viz. Ammonium Chloride, Asparagine, Calcium Nitrate,

28
Potassium Nitrate, Sodium Nitrate and Urea and their effect on phosphate
solubilization were reported. (Ahuja et al. 2007).
Narula et al. (2007) isolated Phosphate Solubilizing Bacteria from the
rhizosphere soil of eternal rays and reference strains of A. chroococcum,
Pantoea, Pseudomonas were compared for phosphate solublizing ability over a
period of 18 days. Without much effect on pH, cumulative effect of these entire
factors might have had an indirect effect on phosphate solubilization.
Shahab and Ahmed (2008) studied that the phosphate solubilization
efficiency of ten soil bacteria for various parameters like carbon sources such as
Glucose, Fructose, Sucrose and Lactose, variable concentration of Sodium
Chloride and Glucose. Glucose was the most favorable carbon source for
solubilization while lactose is the least favorable carbon source. Acinetobacter
lwoffi, Pseudomonas aeruginosa and Bacillus thuringiensis were found to be the
most promising isolates.
Kang et al. (2008) stated that Aspergillus sp, a soil isolate had excellent
potential to solubilize Rock Phosphate in vitro. The process was influenced by
the presence of various concentrations of local loess (red soil). The simultaneous
occurrence, in our experiment, of high levels of solubilized phosphate and
synthesized citric acid, together with the lowest reached pH values, confirmed
the role of citric acid in the phosphate solubilization mechanism. When the soil
was present, phosphate release was better correlated than citrate synthesis with
Hydrogen ion concentration. Changes in soluble phosphate concentration did not
follow a sigmoid pattern. The ability of organism to release phosphatase was
also studied.

29
The mechanisms involved in the weathering processes of insoluble Rock
Phosphate of Moroccan phosphate mine by phosphate solublizing strains
isolated from Moroccan phosphate mine indicated that the isolates produce
siderophore but not organic acids. (Hamdali et al. 2008). Thermo-tolerant
phosphate-solubilizing microbes including bacteria, actinomycetes, and fungi
were isolated from different compost plants and biofertilizers to prepare the
multi-functional biofertilizer. Most of the isolates possessed amylase, Carboxy
Methyl Cellulase, Chitinase, Pectinase, Protease, Lipase, and Nitrogenase
activities. All isolates could solubilize calcium phosphate and Israel Rock
Phosphate. Adding these microbes can shorten the period of maturity, improve
the quality, increase the soluble phosphorus content, and enhance the
populations of phosphate-solubilizing and proteolytic microbes in biofertilizers
(Chang and Yang. 2009).
Park et al. (2009) investigated the ability of Pseudomonas fluorescens to
solubilize insoluble phosphate via two possible mechanisms: proton excretion by
ammonium assimilation and organic acid production. There were no clear
differences in pH and phosphate solubilization between Glucose-ammonium and
Glucose-nitrate media. phosphate solubilization was significantly promoted with
Glucose compared to Fructose. Regardless of nitrogen sources used,
Pseudomonas fluorescens solubilized little insoluble phosphate with Fructose.
High performance liquid chromatography analysis showed that Pseudomonas
fluorescens produced mainly gluconic and tartaric acids with small amounts of
2-ketogluconic, formic and acetic acids. During the culture, the pH was reduced
with increase in gluconic acid concentration and was inversely correlated with
soluble phosphate concentration.
Khan et al. (2009) described the occurrence, mechanisms and role of
Phosphorus solubilizing bacteria in crop production. Plants acquire phosphorus

30
from soil solution as phosphate anion. It is the least mobile element in plant and
soil contrary to other macronutrients. It precipitates in soil as orthophosphate or
is absorbed by Ferric and Aluminum oxides through legend exchange.
Phosphorus Solubilizing Bacteria play role in phosphorus nutrition by enhancing
its availability to plants through release from inorganic and organic soil
phosphate pools by solubilization and mineralization. Principal mechanism in
soil for mineral phosphate solubilization is lowering of soil pH by microbial
production of organic acids and mineralization of organic Phosphates by acid
phosphatase. Use of phosphorus solubilizing bacteria as inoculants increases
phosphate uptake. These bacteria also increase prospects of using phosphatic
rocks in crop production. Greater efficiency of Phosphate Solubilizing Bacteria
has been shown through co-inoculation with other beneficial bacteria and
mycorrhiza.
Srividya et al. (2009) isolated and characterized the fungal strains from
agriculture soil, having potential to solubilize insoluble inorganic phosphates on
Pikovskya’s medium with Tricalcium Phosphate. Aspergillus niger and
Penicillium sp. showed high phosphate solubilisation efficiency on Pikovskya’s
medium with Tricalcium Phosphate in liquid broth in 5 days of growth. A. niger,
showed maximum phosphate solubilization efficiency on Pikovskya’s agar solid
and liquid medium in 5 days of growth. Aspergillus sp showed diverse levels of
phosphate solubilization activity in both solid and liquid broth culture in
presence of various carbon and nitrogen sources and different media. Phosphate
Solubilizing Microorganisms convert insoluble phosphates into soluble forms
generally through the process of acidification, chelation and exchange reactions.
Joseph and Jisha (2009) isolated Phosphate Solubilizing Bacteria
possessing the ability to solubilize insoluble inorganic phosphates from
rhizosphere soil. Eighty-one potential PSBs thus obtained were quantitatively

31
screened for phosphate solubilization. Of these, four bacteria such as
Acetobacter liquefaciens, Acetobacter sp., Pseudomonas gladioli and one
unidentified strain found to be efficient phosphate solubilizers. They were
selected for further evaluation, and found that they solubilize Tricalcium
Phosphate in buffered as well as non-buffered media. The efficiency of
phosphate solubilization was decreased in buffered media compared to non-
buffered media. The buffering capacity of the medium reduced the effectiveness
of Phosphate Solubilizing Bacterias in releasing Phosphate from Tricalcium
Phosphates.
Bacterial solubilization of insoluble inorganic phosphate has been studied
as a means of providing available phosphorus for crop production. Bacterial
abilities to solubilize Calcium Phosphate and Rock Phosphate have been
identified to be related with their abilities to produce Gluconic acid and
Ketogluconic acid. However, there is no information regarding the relationship
between bacterial ability to solubilize Aluminum phosphate and their ability to
produce organic acids. Bacterial ability to solubilize Calcium and Aluminum
phosphates were determined as the concentration of soluble phosphate in the
filtrate of bacterial cultivation media, while bacterial ability to produce organic
acids were assessed from the accumulated organic acids in its. Organic acids
related with the solubilization of calcium phosphate differ from the ones related
with the solubilization of Aluminum Phosphate. Moreover, there is similarity in
the production of organic acids related to the solubilization of Aluminum
Phosphates and Iron phosphate (Prijambada et al., 2009).
Saha and Biswas (2009) isolated, purified and characterized Phosphate
Solubilizing Bacteria from different fertility gradient with regards to N, P and K
status of soil through their insoluble mineral phosphate-source utilization
patterns. Four insoluble phosphate sources; purulia Rock Phosphate, Mussourie

32
Rock Phosphate, crystalline iron and Aluminum Phosphate were charged in
basic Pikovskaia solid medium. Growth pattern of the isolates on those
phosphate sources was recorded. Different communities utilized different
Phosphate sources in different magnitudes.
Kumar et al. (2010) isolated six Phosphate Solubilizing Bacteria from
paddy fields of Eastern Uttar Pradesh, India and identified as members of
Enterobacter and Exiguobacterium genera. Of the six isolates, Enterobacter sp.
exhibited high level of phosphate solubilization in liquid medium.
Exiguobacterium sp. showed increased phosphate solubilization efficiency under
alkaline pH.
Ekin. (2010) evaluated the effect of application of Phosphate Solubilizing
Bacteria, Bacillus Sp, with and without varying amounts of phosphorus fertilizer
on growth and yield of sunflower under field conditions. The application
Phosphate Solubilizing Bacteria was able to mobilize phosphate efficiently in
the sunflower and improved seed quality and oil yield. It also enhanced the head
diameter, seed weight, kernel ratio and oil content and increases in oil yield.
However, when phosphate solubilizing bacteria was used in conjunction with
phosphatic fertilizers, a much greater effect was observed.
Kapri and Tewari. (2010) isolated fourteen strains of Trichoderma sp
from the forest tree rhizospheres of Pinus, Deodar, Bamboo, Guava and Oak on
Trichoderma selective medium. The isolates were tested for their in-vitro P-
solubilizing potential using National Botanical Research Institute Phosphate
(NBRIP) broth containing Tricalcium Phosphate as the sole phosphate source,
and compared with a standard culture of T. harzianum. All the cultures were
found to solubilize Tricalcium Phosphate but with varying potential. Extra-
cellular acid and alkaline phosphatases of the fungus were induced only in the

33
presence of insoluble phosphorus source. The study explores high Phosphate
solubilizing potential of Trichoderma sp., which can be exploited for the
solubilization of fixed phosphates present in the soil, thereby enhancing soil
fertility and plant growth.
Chakraborty et al. (2010) collected four hundred isolates from soil
samples from forest, river basin, agricultural fields and rhizosphere of plantation
crops of North Bengal. The isolates were screened for phosphate solubilizing
activity on Pikovskaya’s agar medium. Among the screened isolates, ninety
showed phosphate solubilizing activity. Out of these, ten isolates belonging to
Aspergillus niger, A. melleus and A. clavatus were selected for further in vitro
evaluation of phosphate solubilization using Tricalcium Phosphate and Rock
Phosphate. The study revealed that the isolates could solubilize Tricalcium
Phosphate better than Rock Phosphate. Selected isolates were mass multiplied
using farm-yard manure and were tested in vivo for their growth promoting
activity in soybean. While all the isolates promoted growth, A. niger was found
to be most effective.
The importance of rhizospheric microbial phosphate solubilization has
now been well documented. However, the performance of these microbes is
greatly affected by various environmental stresses such as salt stress, pH stress,
temperature stress etc. In this study, two stress tolerant phosphate solubilizing
rhizobacteria Arthrobacter sp. and Bacillus sp. have been isolated from tomato
rhizosphere and characterized with various morphological and biochemical tests.
Phosphate Solubilizing Bacteria were screened on the basis of their phosphate
solubilization and strains with high phosphate solubilizing ability were then
tested against wide range of temperature, pH, and salt stresses. Their ability to
solubilize other insoluble phosphates, such as ferric phosphate and aluminum
phosphate was also studied. In addition to phosphate solubilizing ability these

34
strains also demonstrated various plant growth promoting and biocontrol
activities including Indole Acetic Acid production. These two strains have the
potential to be used as plant growth promoting rhizobacteria (Samiran et al.,
2010).
The ability to solubilize insoluble inorganic phosphate compounds by
Gluconacetobacter diazotrophicus was studied using different culture
approaches. Qualitative plate assays using Tricalcium Phosphate as the sole
phosphate source showed that G. diazotrophicus produced solubilization only
when aldoses were used as the carbon source. In batch cultures with
hydroxyapatite as the phosphate source and Glucose as the carbon source, more
than 98% of insoluble phosphate was solubilized. Continuous cultures of G.
diazotrophicus showed significant activities under carbon or phosphate
limitation. It was suggest that G. diazotrophicus is an excellent candidate to be
used as biofertilizer because in addition to the already described plant growth-
promoting abilities of this organism, it shows a significant mineral phosphate
solubilization capacity (Crespo et al., 2011).
Mutation
Mutations are heritable changes in genetic material. The mutation can
cause Macroalterations or Microalterations. Sometimes it causes a silent
mutation that does not result in a changed amino acid sequence, i.e., the new
codon just happens to code for the same amino acid. Macroalterations are large
changes, such as duplications, deletions, inversions or rearrangements of a large
number of bases. Microalterations involve single base pairs. Transitions are
Purine changes to an alternate Purine, Pyrimidine changes to an alternate
Pyrimidine. Transversions is a position with a Pyrimidine changes to have a
Purine; or, Purine to Pyrimidine.

35
Morphological Mutants have altered shape. Lethal Mutants die as a result
of having the mutation. Conditional Mutants are normal under one condition
(permissive), but abnormal under another (restrictive). Biochemical Mutants
cause defects in biochemical pathways for a substance, which is then deficient.
Mutation mechanism
A mutagen or mutagenic agent is a substance that increases the mutation
rate more than the naturally occurring rate. These are substances, conditions and
forms of energy that significantly increase the frequency of mutations. Examples
of forms of energy are Ultraviolet light, X-rays, Cosmic energy, Gamma
radiation, Alpha particles, Beta particles and Neutrons. Examples of substances
that are mutagens are Nitrous acid, Hydroxylamine, Ethyl Ethane Sulfonate, 5-
bromouracil, Celery, Benzo (a) Pyrene, Acridine dyes.
Ultraviolet Radiation causes Pyrimidine dimers. Gamma and X-Rays:
These can act directly on DNA. Alkylating agents are chemicals that donate
alkly groups to other molecules. Ethyl Methane Sulfonate (EMS) is an example.
Base analogs are similar to the actual correct base and so get incorporated into
the DNA as would its natural counterpart. The problem is that, if they are more
prone to tautomeric shifts than the natural base, the frequency for mutation goes
up, substantially. The compound 5-bromouracil is an example of an analog to
thymine. It undergoes a tautomeric shift to base pair with guanine instead of
adenine, causing a transition. Deaminating agents cause the loss of the amino
group. Deaminating agents would increase the frequency of Cytosine
deamination, greatly. Nitrous acid is a deaminating agent. Intercalation agents
are compounds that can slide between the nitrogenous bases in a DNA molecule.
This tends to cause a greater likelihood for slippage during replication, resulting

36
in an increase in frameshift mutations. Hydroxylating agents add an OH group to
a position on the DNA base cytosine, causing a G:C to A:T transition.

37
The mechanisms of action of mineral phosphate solubilization, the effect
of carbon, nitrogen and phosphate sources on phosphate solubilization (MPS)
were studied in the wild-type Mps+ Penicillium rugulosum strain and in
negative (Mps−) and superpositive (Mps++) mutants derived from it. Mps+
phenotype was strongly associated with the production of gluconic or citric
acids. (Reyes et al. 1999)
Ray et al. (1999) isolated Corynebacterium species from spoiled coconut,
was found capable of producing extracellular Lipase. The Lipase activity of the
strain was tried to improve by mutagenic technique using a chemical mutagen N-

38
methyl, N-nitro, N-nitroso guanidine. They also investigated the superior carbon
and nitrogen sources for Lipase production by the mutant strain.
A pot experiment was conducted in the green house to investigate the
establishment of phosphate solubilizing strains of Azotobacter chroococcum,
including soil isolates and their N-methyl-Nnitro-N-nitrosoguanidine mutants,
in the rhizosphere. The solubilization capacity and IAA production ability of
mutants were also studied (Kumar et al., 2001).
The high mineral phosphate solublizing wild strain Penicillium
rugulosum and two of its mutants were used to investigate phosphate
solubilization in liquid cultures contains two Venezuelan phosphate rocks and
Utah variscite. Solubilization of the various P sources by the wild-type and the
first mutant resulted mostly from the action of organic acids. The organic acids
are likely involved in the solubilization of the other phosphate rock (Reyes et al.,
2001).
The enhancement of Lipase production from Aspergillus niger was
attempted by ultraviolet (UV) and nitrous acid mutagenesis, and the mutants
were selected on media containing bile salts. Nitrous acid mutants exhibited
increased efficiency for Lipase production when compared with UV mutants in
submerged fermentation. The hyper producing UV and nitrous acid mutants
were further subjected to a second step of mutagenesis to devise an economical
and ecofriendly technique for Lipase production by the effective use of
hydrocarbons (Mala et al., 2001).
An indigenous strain of Aspergillus niger isolated from an oil mill waste
was used to analyze Lipase activity. The mutant strains were isolated by

39
exposing the parent using ultra-violet and N-methyl, N-nitro, N-nitroso
guanidine. The effect of different carbon and nitrogen sources were also studied
on the mutant which produce high Lipase activity (Ellaiah et al. 2002).
Pseudomonas fluorescens strains and their cold tolerant mutants were
examined for their Tri Calcium Phosphate solubilizing activity in broth media at
10°Cand 25°C. Invariably, all the cold tolerant mutants were found more
efficient than their respective wild type counterparts for phosphate solubilization
activity at 10°C as compared to 25°C (Das et al. 2003).
Top of Form
Bottom of Form
A study for screening and selection of cold tolerant mutants of Pseudomonas
fluorescens strains based on phosphate solubilization ability and subsequent
effect on plant growth promotion under in vitro and in situ conditions was
conducted. Of all the mutants tested, two were selected, as there was a 21-fold
increase in one mutant and a 10-fold decrease in another mutant over their
respective wild types. (Katiyar and Goel. 2003).
Chakraborty et al. (2003) made a attempt to isolate hyper motile mutant
of Bradyrhizobium spp. through UV mutagenisis. A 30W germicidal lamp
exiting most light rays at 256 nm was used as UV source for the purpose of
inducing mutations. A 10 ml suspension of washed cells from an exponential
phase in phosphate buffer was taken in a sterilized petri dish and exposed to
UV rays in a dark from a distance of 20 cm with continuous shaking. Samples
of 0.1 ml were withdrawn from the plate at an interval of 10 sec and inoculated
into Yeast Extract Mannitol broth to examine the growth of suffering cells.
Bapiraju et al. (2004) investigated the Lipase activity by Rhizopus species
isolated from coconut oil mill waste. They improved the Lipase activity of the

40
strain by natural selection and random mutagenesis by UV and N-Methyl, N-
Nitro, N-Nitroso Guanidine.
Mutants of Aspergillus niger NCIM 1207, isolated by subjecting conidia
to UV-irradiation, were tested for the production of Lipase. Mutants UV-10 and
ANCR-1 showed seven fold and five fold enhanced productivity of enzyme,
respectively, over the wild strain in shake flask culture when grown in SOB
medium containing 1% olive oil. Maximum Lipase activity was obtained in the
culture broth when UV-10 was grown in medium supplemented with 0.5%
Triton X-100. A higher concentration of oil in the medium did not help Lipase
production in the case of mutant UV-10. Similarly no increase in enzyme levels
was observed when mutant UV-10 was grown in medium supplemented with
Glucose. However, the addition of Glucose in the medium resulted in increased
levels of Lipase production by wild strain, Aspergillus niger NCIM 1207
(Mahadik et al., 2004).
Azospirillum sp was isolated from soil sample paddy field at
Thirumangalam, Madurai. The isolated Azospirillum sp. was treated with UV
rays Acridine Orange and Ethidium Bromide. The mutants were tested for the
production of auxin under trp (-) & trp (+) condition compared with wild strains.
The mutants produced more auxin than the wild type. The results revealed that
the Auxin production by mutant was independent of Tryptophan (Kulanthaivel et
al., 2006).
Twenty-three bacterial isolates were screened for their mineral
phosphate–solubilizing ability on Pikovskaya and National Botanical Research
Institute’s Phosphate (NBRIP) agar. The majority of the isolates exhibited a
strong ability to solubilize Hydroxyapatite in both solid and liquid media. The

41
solubilization in liquid medium corresponded with a decrease in the pH of the
medium. Serratia marcescens GPS-5, known for its biocontrol of late leaf spot
in groundnut, emerged as the best solubilizer. S. marcescens GPS-5 was
subjected to Ethyl Methane Sulfonate mutagenesis, and a total of 1700 mutants,
resulting after 45 minutes of exposure, were screened on buffered NBRIP
medium for alterations in mineral phosphate solubilizing ability compared with
that of the wild type (Tripura et al., 2007).
Phenotypic mutants of Aspergillus tubingensis were obtained by UV
irradiation and phosphate solubilization ability of these mutants were studied and
compared with the wild type strain. These mutants also showed maximum Acid
Phosphatase and Phytase activity. These results suggest that phosphate
solubilization by these isolates is due to lowering of pH of the culture filtrate and
also the activity of Acid Phosphatase and Phytase (Achal et al. 2007).
The ability of several Pseudomonas spp. to solubilize Ca3 (PO4)2 was
compared. While all Pseudomonas sp were found to facilitate a decrease in pH
and solubilize inorganic phosphate by the production of extracellular organic
acids, strains varied by producing either gluconic or 2-ketogluconic acid. It was
further investigated that the genetic mechanisms involved in inorganic phosphate
solubilization by Pseudomonas spp., a transposon mutant library of P.
fluorescens was screened for mutants with reduced Ca3 (PO4)2 solubilization
ability. Mutations in the gcd and pqqE genes greatly reduced the solubilization
ability, whereas mutations in the pqqB gene only moderately reduced this ability.
The combination of biochemical analysis and genomic comparisons revealed
that alterations in the pqq biosynthetic genes, and the presence/absence of the
gluconate dehydrogenase (gad) gene, fundamentally affect phosphate
solublization in strains of P. fluorescens (Miller et al., 2010).

42

43
Phosphatase
The effect of phosphate on the production of phosphatases by Aspergillus
awamori var. kawachii was studied. The concentration of phosphatase,
produced, was distinguished between high and low phosphate medium. The
phosphatase fractions produced in the low phosphate medium were different
from those of the high phosphate medium (Ohta et al., 1968).
A simple method of assaying soil phosphatase activity was described. It
involves colorimetric estimation of the p-nitrophenol released by phosphatase
activity when soil is incubated with buffered (pH 6·5) Sodium p-nitrophenyl
phosphate solution and toluene at 37° C for 1 hr. The method is rapid and
precise, and it has significant advantages over methods previously proposed for
assay of soil phosphatase activity (Tabatabai and Bremner. 1969).
The occurrence and distribution of a special group of bacteria, capable
of dissolving insoluble phosphates, were studied in marine environments,
especially in sediments. The phosphatase activity was also investigated. There
was a positive correlation between the total phosphate content and the
phosphatase activity. The phosphatase activity was recorded in all samples,
irrespective of salinity variations (Ayyakkannu and Chandramohan, 1971).
A group of yeasts isolated from burned children and 14 reference strains
were tested for phosphatase activity by using phenolphthalein phosphate
substrates. Phosphatase activity was widely distributed among various species
and strains representing seven genera. The greater enzyme activity of the yeast
cell was not related to more rapid or greater cell growth or decrease in the pH of
culture media. Extracellular constitutive heat-labile acid phosphatase was found
in broth filtrates of some strains (Smith et al., 1973).

44
The bacteria Lysobacter enzymogenes was studied for it’s extracellular
phosphatase during the stationary phase of growth. The cells also produced a
cell-associated alkaline phosphatase. This enzyme is found in the particulate
fraction of cell extracts and may be membrane bound. The production of both
phosphatases, especially the extracellular enzyme, is reduced by inorganic
phosphate (Richard and Tigerstrom, 1984).
The thermostability properties of three Acid Phosphatases viz, Aspergillus
fumigatus Phytase, Aspergillus niger Phytase, and A. niger Acid Phosphatase
(pH 2.5), were investigated by measuring enzymatic activity. The Phytases of
A. fumigatus and A. niger were both denatured at temperatures between 50 and
70°C. A. niger pH 2.5 acid phosphatase displayed considerably higher
thermostability, denaturation, conformational changes, and irreversible
inactivation were observed only at temperatures of 80°C (Wyss et al., 1998).
A Phosphatase overproducing Citrobacter sp. was grown in an air-lift
reactor in steady state continuous culture under limitation of Carbon, Phosphorus
or Nitrogen. Substantial biofilm formation and the highest Phosphatase activity,
were observed under Lactose limitation (Allan et al., 2002). The fungus
Rhizopus delemar produced extracellular and cellular acid phosphatase during
the growth in starch-supplemented medium in the presence or absence of copper
ions. The levels of both acid phosphatase activities were observed. The pH
optimum of the enzyme was determined to be in the range of 3.5–4.5 using p-
nitrophenyl phosphate as a substrate (Tsekova and Galabova. 2003).
The effect of several carbon sources on the production of Alkaline
Phosphatase by the thermotolerant Aspergillus caespitosus was analysed. The

45
pH of the medium likely affects the process of enzyme secretion according to the
carbon source used. (Guimaraes et al., 2003). Sarapatka et al. (2004) studied
Acid Phosphatase activity on the root systems of cereal varieties and also in
nutrient medium on which crops were planted under conditions of changing pH
and phosphorus supply. After 10 days of cultivation the plant roots were
harvested, homogenized and the acid phosphatase activity was measured. The
correlation of increasing pH and available phosphorus level in the nutrient
medium with acid phosphatase activity in the root system of various species and
cereal cultivars was recorded. The Phosphatase Activity of Ectomycorrhizal
roots and nutrient status of Pine seedlings inoculated with Mycorrhizal fungi
were studied under different physical factors, namely relative humidity, light
intensity and concentration of acid phosphatase activity (Jha et al. 2006).
Padmapriya and Anand (2010) studied nine filamentous cyanobacterial
isolates for the screening for amylase, protease, beta lactamae and phosphatase
enzymes. They stated that the extracellular activity of acid phosphatase enzyme
in the non-heterocystous forms, Phormidium minnesotensis and P. boryanum,
showed a decrease as the growth increased, reaching negligible levels on the 20th
day. In the heterocystous forms, both the acid phosphatase extracellular and
intracellular activities were absent up to a period of 8 days. In the case of
alkaline phosphatase intracellular activities, a gradual decrease in the enzyme
content from the first interval is observed in almost all the forms.
Phosphatase and Phosphate Solubilization
Bacterial strains were obtained from a soil sample and culture collection
center and they were used for enzyme production in a low-concentration
phosphate medium. Various concentrations of Sodium Phosphate or Calcium
were added in experiments to study the regulation of phosphatase synthesis.

46
Alkaline Phosphatase activity was measured spectrophotometrically by
monitoring the release of para nitro phenol from para nitro phenyl phosphate
(PNPP) at 400 nm (Prada et al. 1996).
The use of Phosphate Solubilizing Bacteria as inoculants simultaneously
increases phosphate uptake by the plant and crop yield. Strains from the genera
Pseudomonas, Bacillus and Rhizobium are among the most powerful phosphate
solubilizers. The principal mechanism for mineral phosphate solubilization is the
production of organic acids, and Acid Phosphatases play a major role in the
mineralization of organic phosphorous in soil. Several Phosphatase encoding
genes have been cloned and characterized and a few genes involved in Mineral
Phosphate Solubilization have been isolated. Therefore, genetic manipulation of
Phosphate Solubilizing Bacteria to improve their ability to improve plant growth
may include cloning genes involved in both mineral and organic phosphate
solubilization, followed by their expression in selected rhizobacterial strains
(Rodriguez and Fraga.1999).
Seawater and sediment samples were collected and were screened for
phosphate solubilization by streaking them on to Hydroxyapatite medium11. The
phosphate solubilization was expressed as positive and negative depending on
the halo formation. The cultures which showed halo formation around their
colonies were considered to be the phosphate solubilizers. These solubilizers of
inorganic phosphate were further screened for Phosphatase production (De
Souza et al., 2000).
The strains, viz., phosphobacteria, yeast and mildew were compared for
their Phosphorus Dissolving Mechanism and phosphorus solubilizing abilities.
Acids, Acid Phosphatase, and Alkaline Phosphatase which have synergic effects

47
on phosphate dissolution were studied. The variation in activities of Acid
Phosphatase and Alkaline Phosphatase of Phosphate Solubilizing Microbes were
also examined (ChuanQing and WeiYi.2005).
Ponmurugan and Gopi (2006) conducted an experiment to enumerate the
population density of phosphobacteria in the rhizosphere soils of various crops
using Ketznelson and Bose medium following dilution plate technique. Efforts
have been made to isolate phosphobacteria from these soils and isolated strains
were inoculated in specific media containing specific substrates to produce
growth regulating substances such as Indole Acetic Acid and Gibberellic Acid
and Phosphatase enzyme.
Aspergillus niger was studied for its Acid Phosphatase and Invertase
activity in defined media supplemented with various sucrose concentrations.
Ontogenic changes in extracellular, cytoplasmic, and wall-bound enzyme
activities of A. niger were studied. Growth in terms of fresh weight showed
inverse correlation with pH. At higher pH values, both enzyme activities were
higher in the medium supplemented with low sucrose concentration. It was
observed that the more the fresh weight of fungi decreased, the greater was the
enzyme activity observed (Pawar and Thaker, 2009).
Lipase
Lipases are produced by a number of microorganisms, including bacteria,
fungi, and yeasts. In recent years, research on microbial Lipases has increased
because of their practical applications in industry, as in the hydrolysis of fats,
production of fatty acids and food additives, synthesis of esters and peptides,
resolution of racemic mixtures, or additives in detergents.

48
Constein and Thomas (1902) reported that the difference in the sources of
Lipase used by different workers, the diversity of the data report on this subject
seems to partly due to the fact that the many efforts thus far made to purify the
enzyme have not met with definite success.
Juichiro et al. (1963) reported that the Lipase is one of the enzymes
whose chemical and enzymological properties have not yet been fully elucidated
despite the vast literature accumulated in the past. Conflicting reports exists
concerning its requirement for co-factors, specific to substrates, mechanism of
reactions and other properties started their experiment with an extensive
screening test of moulds which led to selection of several strains of Rhizopus and
Aspergillus niger as most potent Lipase formers. Successful isolation of the
enzyme could be made from the bran-koji culture of Aspergillus niger, and the
enzyme was finally obtained in a crystalline form.
Wen Hsiung et al. (1972) isolated a thermophilic fungus which can
produce a thermostable Lipase (glycerol ester hydrolyses) from compost soil and
identified this as Humicola lanuginose. The culture conditions and some
properties of crude Lipase preparation were described in the previous report. In
an attempt to obtain more comprehensive information on the thermostable
Lipase, a lightly purified enzyme preparation is an essential pre-requisite of the
purified Lipase of Humicola lanuginose.
Wen Hsiung et al. (1973) reported on substrate specificity and mode of
action of the Lipase of thermophilic fungus Humicola lanuginose s-38. It was
made clear that mode of action of Humicola Lipase on triolein and on
phosphatidyls ethanolamine was identical. The Humicola Lipase had no activity
of lipoprotein Lipase. Determination of the lipolytic activity after a certain time

49
of incubation, the maximal lipolytic activity and a time integrated lipolytic
activity are compared as estimations for potential hydrolytic capacity of
microorganisms. (Jonsson and Snygg. 1974).
Sugiura et al. (1975) reported that long chain fatty acids generally had an
effect on Lipase production by Candida paralipolytica in both the cells and the
broth. A study on the influence of olive oil in Lipase production by Rhizopus
species, Penicillium roquefortii and Mucor species showed that the cellular
activity increased in some cases and decreased in others. Nobuhiro and Omar
(1977) isolated many microorganisms which are known to produce different
types of Lipases. Almost all microbial Lipases which have been reported can be
regarded as acid Lipases or neutral Lipases when they are classified by their
optimum pH values.
Watanabe et al. (1977) identified the alkaline Lipase producing
microorganisms, their culturing conditions and some properties of the crude
enzymes obtained. Oso (1978) carried out a study on the ability of Tararomyces
emersionstolk to produce extracellular Lipase in stationary liquid medium under
various conditions. The best temperatures for Lipase synthesis and activity were
40-45°C, and Lipase production was tested at all the temperature (37-55°).
Akhtar and Nadeem (1983) proposed the effect of triglycerides on the
growth of Mucor hiemalis and the production of Lipase and mycelial lipids on
addition of 1% triglycerides to the fermentation medium. The added
triglycerides seemed to be utilized through the formation of free fatty acids. The
lipids produced per gram mycelia were high initially (260mg/g dry weight at 48
hrs), and showed reduced production later. With increase in growth the

50
maximum mycelia lipids per 100ml of culture medium was obtained from
176mg/100ml.
Recent investigations have shown that enzymes have potential in large
scale processing of lipid material, particularly in the areas of fat splitting,
synthesis reversal of hydrolysis and interiesterification. (Posorske. 1984).
Warner et al. (1984) set out to determine the kinetics of lipolysis of olive oil,
coconut oil and tallow and the effects of temperature concentration of Lipase and
pH on commercially available Lipase from Candida rugosa. The effects of
calcium and sodium ions in olive oil lipolysis at room temperature were
determined in the present study.
Dow and Fhee (1986) developed simple and rapid calorimetric method to
determine the Lipase activity for fat splitting. A further simplification of the
method of Lowry and finally for determination of free fatty acids by eliminating
the solvent evaporation and centrifugation steps for Lipase assay. Macrae and
Hammond (1985) reported that Lipase produced by thermophilic fungi have
drawn great deal of attention lately due to their possible application in the food,
pharmaceutical and detergent industries. With newer applications in light, more
active Lipases are being sought from microorganisms.
Nahas (1988) initiated to investigate the effect of different growth media
and conditions on Lipase production by Rhizopus oligosporus, after studying the
factors influencing the growth and production of extracellular Lipase by
Rhizopus oligosporus. The yields of Lipase was maximum at 25°C and pH 6.5
after 3 days shaking culture which enhanced growth but decreased Lipase
production.

51
Johri et al. (1990) studied extracellular Lipase of Sporotrichum
thermophile. It was found that 15%-20% of the enzyme remained cell bound. It
was therefore of interest to examine Lipase secretion by protoplasts derived from
the mycelium. Since immobilization of protoplasts can improve stability and
reusability, entrapment of protoplasts of Sporotrichum thermophile in alginate
was attempted.
Martin (1990) presented his study on the Lipase catalyzed alcoholysis
sunflower oil in order to optimize conditions for the synthesis. Gerald and
Tsuneo (1991) presented that it is possible to obtain nearly theoretical yields of
monoglycerides (approximately 90% wt) using Pseudomonas sp. Lipase and a
yield of approximately 80% wt using Mucor miehei Lipase under appropriate
conditions.
Valerie et al. (1992) isolated strains of Penicillium cyclopium,
Aspergillius niger Aspergillis flavus and Rhizopus arrhizus secrete extracellular
enzymes exhibited good synthetic activities in organic solvent. A loop fixed-bed
reactor was designed for the continuous synthesis of esters.
Lipase biosynthesis occurred in medium without lipids, but for improved
production an inducer was needed. The source and concentration of an inducer
had no significant effect. Starch as an additional carbon source stimulated Lipase
biosynthesis when used in small amounts. Addition of NH4NO3 as a nitrogen
source, KH2PO4 as a phosphate source as well as Mg ions to the medium with
initial pH 5.0 gave the best yield (Pokorny et al., 1994).
There are other reports on the application of microbial Lipase to the
hydrolysis of racemic esters transesterification and to racemization in situ to

52
yield optically pure enantiomers for the manufacture of chiral symptoms. In
addition to racemization in situ Lipases are also capable of catalysing synthetic
reactions, which lead to the production of life-saving drugs (Lee, 1995).
Parmar et al. (1996) reported that because of their excellent capability for
specific region selective reaction in a variety of organic solvents with broad
substrate recognition, Lipases have emerged as an important biocatalyst in
biomedical application. Hilda and Gobinith (1997) reported that oil pollution is a
serious obstacle to photosynthesis, a fundamental life process in plant kingdom,
which affects the food chain and productivity of the sea. Oil is also absorbed by
fish eventually to reach and endanger human life. The lipolytic activity of
physiologically diverse microorganism can be used to degrade oil spills in
marine environment. The fungus Acremonium alternate exhibited varied
efficiency in hydrolyzing the various oils. It degraded olive and ground nut oils
to a maximum efficiency.
Gombert et al. (1999) studied growth and enzyme production by a
Brazilian strain of Penicillium restrictum. Solid waste from the babassu oil
industry was used as the basic nutrient source and was supplemented with
peptone, olive oil or starch at different C:N ratios. Lipase activity was very
sensitive to the kind and the level of supplementation, and decreased as protease
level and pH in the media increased.
Different carbon sources affecting growth and Lipase production in
Candida rugosa were studied by using batch cultures on defined medium.
Carbohydrates and acids non-related to fats and lipids or fatty acids were used
as a carbon sources to induce Lipase production. The highest yields of enzyme
were obtained with lipids or fatty acids as carbon sources (Dalmau et al., 2000).

53
A response surface approach has been used to study the production of
extracellular Lipase by Candida cylindracea. Medium constituents were
optimized for the production of Lipase by central composite design. A 25 design
matrix was used in the experiments with only the carbon source being varied.
The Glucose and olive oil were used as the carbon source All the other medium
constituents namely yeast extract, malt extract, peptone and tween 80 were
simultaneously optimized (Muralidhar et al., 2001).
Coca et al. (2001) used strains from bacteria, fungi and yeast for a
quantitative screening of lipase-production using 2% of olive oil as carbon
source. The production and kinetic characteristics of extracellular lipases
expressed by the best producers such as Aspergillus niger and A. fumigatus were
studied. The more ideal carbon source for lipase synthesis was selected. The
optimum pH and temperature for extract enzymatic activities were also
investigated.
The Lipase production abilty of Rhizopus arrhizus NCIM 877, 878, 879
and Aspergillus niger NCIM 1207 were studied in synthetic oil based medium
under submerged conditions. Rhizopus strains showed major intracellular
activity while A. niger NCIM 1207 produced mainly extracellular activity. The
pH and temperature optima for Lipase production and activity and the efficacy
of Lipase production by submerged and solid state fermentation in A. niger
NCIM 1207 were also studied (Mahadik et al., 2002).
The n-alkane degrading Pseudomonas pseudomallei was investigated for
its extracellular Lipase activity. The stability of the immobilized Lipase in

54
organic solvents, retention of activity, pH and thermo stability of Lipase
produced were studied (Kanwar and Goswami, 2002).
Arya et al. (2003) stated that commercial porcine pancreas Lipase was
immobilized covalently onto alkyl amine glass-beads affixed on the inner wall of
a plastic beaker by an adhesive. The immobilized enzyme retained 10.8% of the
initial activity of free enzyme with a conjugation yield of 52-mg/g supports. The
optimum pH and incubation temperature were decreased, while time for linearity
and λm for triolein of enzyme were increased after immobilization.
Achamma et al. (2003) isolated a bacterium, producing extracellular
Lipases was isolated from coconut oil cake and identified as Bacillus mycoides
by morphological and biochemical characteristics. The incubation period for
maximum growth and Lipase production, optimum pH and temperature for the
production of Lipase were optimized. Shake condition induced more Lipase
production. They also studied the best carbon, nitrogen and mineral source for
Lipase production.
The effect of different lipid sources added to basal medium was verified
to improve Lipase production. Maximum growth and Lipase activity was studied
in the presence of different surfactants. The best temperature for Lipase
production was also recorded (Silva et al., 2005).
A thermophilic isolate Bacillus coagulans produced an extracellular
alkaline Lipase, the production of which was substantially enhanced when the
type of carbon source, nitrogen source and the initial pH of culture medium were
consecutively optimized (Satyendrakumar et al., 2005).

55
The non-conventional yeast Yarrowia lipolytica produces an extracellular
Lipase encoded by the LIP2 gene. Mutant strains with enhanced productivity
were previously obtained either by chemical mutagenesis or genetic engineering.
One of these mutants, named LgX64.81 to select new overproducing strains was
used for amplification of the LIP2 gene. A process for Lipase production in
bioreactors and compared Lipase production levels in batch and fed-batch
cultures were also developed (Fickers et al., 2005).
Rodriques et al. (2006) reported that Rhizopus homothalicus was
cultivated for Lipase production in Solid State Fermentation (SSF) using
sugarcane bagasse as a support and impregnated with a liquid medium. The
production of an extra cellular Lipase from Rhizopus homothallicus was
improved by modification of media nutrients. These results are promising
because these strain producing high Lipase concentrations in an in expensive and
simple medium, which facilitates its recovery and purification. Lipase extraction
from the fermented solid was also studied. Efficient recovery of the enzyme was
used.
The microbial production of extra-cellular Lipase on olive-mill
wastewater by Geotrichum candidum, Rhizopus arrhizus, Rhizopus oryzae,
Aspergillus oryzae, Aspergillus niger, Candida cylindracea and Penicillium
citrinum were investigated. All strains were able to grow on the undiluted olive-
mill wastewater, producing extra-cellular Lipase activity. The efficacy of Lipase
production by the best strain Candida cylindracea on different and nitrogen and
oil source were recorded (Annibale et al., 2006).
Two Lipase-producing yeast strains named Saccharomyces cerevisiae and
Williopsis californica were isolated from Olive oil. The ratio of the aqueous to

56
the organic phase which influence the maximum enzyme activity was
determined. The microbiological analysis carried out on commercial extra virgin
olive oil, produced in four different geographic areas, demonstrated that the
presence of Lipase-producing yeast varied from zero to 56% of the total yeasts
detected, according to the source of oil samples (Ciafardini et al., 2006).
Lipase production in Aspergillus niger was tested by Falony et al. (2006)
using both submerged fermentation and solid-state fermentation on a mineral
culture medium and wheat bran, respectively. The optimization of the culture
medium was carried out for both fermentation. The maximum lipase activity was
obtained during the submerged fermentation in a medium containing Glucose
and olive oil. The lipase activity was reached maximum in solid-state
fermentation process with a medium containing ammonium sulphate and urea.
The optimum pH and temperature for enzymatic activity were also investigated.
The bacteria that could grow on media containing olive mill waste water
(OMW) were isolated and their Lipase production capacities were investigated.
The strain possessing high Lipase activity among 17 strains grown on tributyrin
agar medium was identified as Bacillus sp. The effect of initial pH on the Lipase
activity was investigated in tributyrin medium and the liquid medium
composition was improved by replacing tributyrin, with different carbon sources
(Sevagi et al., 2007).
The strain for Lipase production was Penicillium expansum, isolated from
the waste of a rap oil manufactory in China. The strain was used to produce
Lipase by submerged fermentation. The enzyme so produced was purified from
the fermentation broth by DEAE-Sepharose chromatography (Lianghua et al.,
2007).

57
An extracellular Lipases producing thermophilic Bacillus sp. was
isolated. The strain was used to produce Lipase by submerged fermentation. The
Lipases so produced were purified and characterized by novel techniques.
(Nawani and Kaur, 2007).
Damaso et al. (2008) aimed to produce lipases by solid-state fermentation
using, as substrate, agroindustrial residue supplemented with by-products from
corn oil refining process or olive oil. For a group of ten fungi strains selected in
the first steps, the lipase activity obtained by solid-state fermentation varied from
dry substrate. Among the evaluated strains, the Aspergillus niger mutant was
selected by presenting the best enzymatic production. For the fermentation tests,
two substrates were also investigated. Wheat bran and corn cob, both
supplemented with olive oil. Additionally, three industrial by-products from
corn oil refining (soapstock, stearin and fatty acids) were evaluated as substitutes
to the olive oil in the function of lipases production inducer.
Amara and Salem (2009) studied that two strains of Pseudomonas
aeruginosa for lipase production from Castor oil aiming to control the different
castor oil wastes and technical lipase production. Both strains are able to grow
on media consists of Castor oil and Yeast extract at 37°C. The lipase enzyme
activity at 37, 60 and 75°C in presence of p-nitrophenyl palmitate were
investigated. Lipase produced from the two strains revealed mesophilic and
thermophilic activities. They recommended for using P. aeruginosa lipases for
castor oil waste control and for the production of mesophilic and thermophilic
lipases under mesophilic conditions.
Rani and Panneerselvam (2009) selected and tested certain organisms
for lipase activity. Among the organism tested, Aspergillus fumigatus, A. terreus,

58
Penicillium chrysogenum, P. funiculosum and Fusarium moniliforme were
reported as a high lipase producer. Various environmental parameters like pH,
temperature and nutritional parameters such as carbon and nitrogen sources were
also investigated to optimize the production of lipase.
Pogaku et al. (2010) isolated a bacterial strain from an oil contaminated
soil and was identified as Staphylococcus sp. It was screened for lipase activity
on tributyrin agar and spirit blue agar medium. Maximum lipase production was
observed at 48 h of growth. Peptone was found to be as an ideal nitrogen source
for lipase production. Addition of any nitrogen source other than peptone to the
medium resulted in a significant reduction of enzyme production. Lower lipase
production was noted when an inorganic nitrogen source was used as the sole
nitrogen source. Starch was used as a major carbon source for optimum
production of lipase. Of the natural oils, olive oil was able to induce more lipase
rather than the oils like groundnut, coconut, castor oils. The pH and temperature
effect on lipase production were also investigated.
Out of 64 fungal isolates obtained, 43 exhibited lipase activity. It was
found that maximum lipase activity was obtained in liquid medium by Fusarium
oxysporum. Various physicochemical parameters such as pH, temperature,
incubation period, volume ratio, agitation and effect of different carbon and
nitrogen sources were studied in order to determine the optimum conditions for
lipase production. The lipase present in the broth was partially purified with
ammonium sulphate, ethanol and acetone (Rifaat et al., 2010).
Chouhan and Dawande (2010) studied that the production of extracellular
lipase from P. aeruginosa on Rhodamine B agar plates containing olive oil as
ingredient, analysed spectrophotometrically using olive oil as well as Tween-20

59
as substrate, purified and precipitated by 30% saturated ammonium sulphate,
showed the maximum lipase activity as compared to the crude enzyme.
Salihu et al. (2011) screened ten microorganisms for their potential to
produce lipase in palm oil mill effluent based medium. Among the 10 organisms,
the most promising strain was Candida cylindracea which showed appreciable
activity both on agar plates and liquid cultures. Medium supplementation by
NH4Cl and olive oil led to high enzyme activity. However, supplementation
with organic nitrogen sources resulted in better enzyme activity. Addition of
malt extract, peptone and olive oil into the medium greatly influenced the lipase
production. Among the oils that were tested, olive oil was found to be the best
for the expression of extracellular lipase activity in an optimized palm oil mill
effluent supplemented medium.
The culture and desired growth conditions for the production of
extracellular lipase from Aspergillus niger was extensively investigated by
Hosseinpour et al. (2011). Enzyme production was carried out in a submerged
culture using major nutrients from soya flour as main constituent of the media.
The optimum weight percentage of soya flour, Glucose and olive oil
concentrations on lipase production were defined. Combination of nitrogen
sources such as yeast extract and peptone were found suitable nitrogen sources.
Effect of pH, Temperature and Calcium ions on lipase activity were also
investigated.
Kakde and Chavan (2011) screened the lipolytic activity abnormal
safflower seeds. Dominant fungi were isolated from abnormal oilseeds on Potato
Dextrose Agar. Total twenty fungi were isolated. Out of that lipase enzyme
activity of ten dominant fungi other than Aspergillus sp. was studied by using

60
different nutritional sources like carbon, nitrogen, phosphorus, sulphur,
antibiotic and vitamin sources. It was found that carbon sources like Fructose
and sucrose induces lipase activity while starch, lactose and carboxyl methyl
cellulose inhibits lipase activity. Nitrogen sources like casein and peptone which
are organic forms stimulated maximum lipase enzyme production of storage
fungi. Sulphur sources like calcium sulphate and ferrus sulphate reduced the
lipase enzyme production by storage fungi while, phosphorus source like
disodium hydrogen ortho-phosphate, ammonium phosphate and potassium di-
hydrogen ortho-phosphate stimulated lipase enzyme production. Antibiotic like
ampicillin, norfloxacin and tetracycline reduced the lipase production of storage
fungi. Lipase activity of storage fungi was reduced in presence of vitamin source
like riboflavin while, folic acid and vitamin C stimulated the lipase enzyme
production.
Indole Acetic Acid
Pathogenic strains of Xanthomonas campestris pv. glycines, and
nonpathogenic strains were compared for their ability to produce indole
compounds, including the plant hormone Indole Acetic Acid (IAA) in liquid
media with or without supplementation with L-Tryptophan. Several additional
strains of plant-pathogenic Xanthomonads and Pseudomonads were also tested
for Indole Acetic Acid production to determine whether in vitro production of
Indoles present in culture filtrates were identified by thin-layer chromatography,
high-performance liquid chromatography, UV spectroscopy, mass spectroscopy,
and gas chromatography-mass spectrometry and were quantitated by high-
performance liquid chromatography. All strains examined, produced Indole
Acetic Acid when liquid media were supplemented with L-Tryptophan (Fett et
al. 1987).

61
Bric et al. (1991) developed a new assay that differentiates between
Indole Acetic Acid (IAA) producing and nonproducing bacteria on a colony
plate lift. Medium supplemented with 5 mM L-Tryptophan is inoculated with
isolates of interest, overlaid with a nitrocellulose membrane, and then incubated
until bacterial colonies reach 1 to 2 mm in diameter. The membrane is removed
to a filter paper saturated with Salkowski reagent and incubated until distinct red
haloes form around the colonies. The colorimetric reaction to Indole Acetic Acid
is limited to a region immediately surrounding each colony, is specific to isolates
producing Indole Acetic Acid, occurs within 1 h after the membrane is placed in
the reagent, and is sensitive to as little as 50 p mol of Indole Acetic Acid in a 2-
mm spot. This assay, used for quantifying epiphytic and endophytic populations
of Indole Acetic Acid producing isolates of Pseudomonas syringae subsp.
savastanoi and for detecting Indole Acetic Acid producing colonies of other
Pseudomonads and Erwinia herbicola. The assay provides a rapid and
convenient method to screen large numbers of bacteria.
Auxin production by 131 strains of Pseudomonas syringae subsp.
savastanoi was investigated with the aim of looking for correlations among this
characteristic and the origin of the strains, the types of symptoms, and the host
plant. Most of the P. syringae subsp. savastanoi strains, except those isolated
from ash, produced auxin and harbored Indole Acetic Acid genes. Among ash
strains, which were pathogenic only on ash were found to produce auxin and to
harbor Indole Acetic Acid genes (Garden et al., 1992).
Pseudomonas sp., a potent phosphorus solublizer, also produced
significant levels of siderophore and Indole Acetic Acid. Siderotyping indicated
it was P. aeruginosa siderovar 1. Cadmium, Nickel, and Chromium resistant
mutants were developed and characterized for their PGPR properties. Mutants

62
were stable under non-selective pressure. However, they were able to promote
root and shoot elongation in soybeans at a significant level in the presence of
metals unfamiliar to the wild type (Gupta et al., 2002).
Patten and Glick. (2002) isolated the growth-promoting bacterium
Pseudomonas putida and an Indole Acetic Acid deficient mutant constructed by
insertional mutagenesis. The canola seedling primary roots from seeds treated
with wild-type P. putida were on average 35 to 50% longer than the roots from
seeds treated with the Indole Acetic Acid deficient mutant and the roots from
uninoculated seeds. In addition, exposing mung bean cuttings to the wild-type
strain stimulated the formation of many, very small, adventitious roots.
Formation of fewer roots was stimulated by treatment with the Indole Acetic
Acid deficient mutant. These results suggest that bacterial Indole Acetic Acid
plays a major role in the development of the host plant root system.
Yurekli et al. (2003) determined the physical and chemical conditions
necessary for optimal biosynthesis of Indole Acetic Acid by Lentinus sajor-caju.
Glucose was determined to be superior to sucrose as carbon and energy source.
The synthesis of Indole Acetic Acid in a nitrogen-free medium or in a medium
with low external phosphate was substantially reduced. Light exposed and non-
agitated cultures grown in dark had also reduced levels of Indole Acetic Acid
compared to agitated cultures grown in dark. The highest Indole Acetic Acid
level was determined in cultures grown in Glucose containing medium (pH 7.5)
on a rotary shaker (150 rpm) at 30 °C in dark. The biological activity of Indole
Acetic Acid obtained from the extra-cellular culture of Lentinus sajor-caju was
determined using oat coleptile growth test.

63
Maor et al. (2004) studied the effects of tryptophan, Indole-3-acetic Acid,
and Indole-3-acetamide on Indole-3-acetic Acid biosynthesis in fungal axenic
cultures and on in planta indole-3-acetic acid production by the fungus. Indole-3-
acetic Acid biosynthesis was strictly dependent on external tryptophan and was
enhanced by tryptophan and indole-3-acetamide.
The bacteria Sphingomonas sp, Microbacterium sp, Mycobacterium sp,
Bacillus sp, Rhizobium sp. Rhodococcus sp, Cellulomonas sp, Pseudomonas sp,
and Micrococcus luteus isolated from the roots of greenhouse tropical orchids
were shown to produce Indole-3-acetic Acid (IAA) and to excrete it into the
culture liquid. The presence and activity of Indole-3-acetic Acid were
demonstrated colorimetrically, by thin-layer chromatography, and by biotests.
The associated bacteria varied in their ability to excrete indole compounds (1–
28μg/ml nutrient broth). Addition of tryptophan to the growth medium enhanced
the production of phytohormone (Tsavkelova et al., 2005).
Bhuvaneswari et al. (2006) extracted Indole-3-acetic acid (IAA) and
estimated from three species of Camarosporium and Colleatotrichum falcatum.
Auxin generally occurs bound with an amino acid or sugar. Over six different
precursor molecules for auxins have been reported. Auxin production was
tryptophan dependant. No IAA or any other indoles were detected in the
absence of tryptophan, while in the presence of tryptophan IAA production was
high. IAA production was more in culture filtrate when compared to the
mycelium of the fungi.
Rhizobium isolates from root region of Sesbania procumbens and stem
nodules of S. rostrata and S. procumbens were shown to produce indole-3-acetic
acid (IAA) in culture supplemented with L-Tryptophan. Production of IAA was

64
maximal after 72 h of incubation when the bacteria reached stationary phase of
growth. The cultural requirements were optimized for maximum IAA
production. The effect of carbon and nitrogen sources revealed that Glucose and
potassium nitrate were best promoters for IAA production over controls. The
IAA from this isolate was extracted, purified and identified by thin layer
chromatography. (Sridevi and Mallaiah. 2007).
The microbial community Streptomyces, Bacillus, Erwinia and
Pseudomonas genera were isolated from Paphiopedilum appletonianum and
Pseudomonas, Bacillus and Flavobacterium genera were isolated from
Pholidota articulate, produced indole-3-acetic acid (IAA). Variations in its
biosynthesis among the strains of the same genus were also observed. The
highest auxin level was detected during the stationary growth phase (Tsavkelova
et al., 2007).
The bacterial strains including Mesorhizobium, Azotobacter, and
phosphate-solubilizing bacteria were isolated and tested for siderophore,
ammonia, Indole Acetic Acid production and phosphate solubilization in vitro.
Among the isolates, M. ciceri, A. chrococcum and Bacillus were positive for
IAA, The diameter of the P solubilization zone, phosphate released in liquid
medium and concomitant drop in pH were also recorded (Wani et al., 2007).
Phytohormone-like acting compounds have been suggested to be involved
in the phytostimulatory action exerted by the plant-beneficial rhizobacterium
Bacillus amyloliquefaciens. Analyses with culture filtrates of Bacillus
amyloliquefaciens demonstrated the presence of indole-3-acetic acid (IAA),
corroborating it as one of the pivotal plant-growth-promoting substances

65
produced by this bacterium. In the presence of 5 mM tryptophan, a fivefold
increase in IAA secretion was registered. (Idris et al., 2007).
Forty three strains were isolated from knots induced by Pseudomonas
savastanoi in different olive cultivars. All the selected bacteria were shown to
produce variable amounts of the plant growth hormone indole-3-acetic acid
(IAA). Their gene sequences were analysed by 16s rDNA method (Ouzari et al.,
2008).
The effect of phosphate-solubilizing fungi such as strains of Aspergillus
awamori, and Penicillium citrinum isolated from rhizosphere of various crops,
was observed on the growth and seed production of chickpea plants in pot
experiments. All phosphate-solubilizing fungi isolates were biocompatible and
produced growth-promoting hormone, Indole Acetic Acid (IAA), varying in
concentration (Mittal et al., 2008).
About on hundred rhizobial bacterial strains were isolated, identified and
tested for Indole-3-acetic Acid in Luria Broth medium supplemented with
tryptophan. They used modified nitrocellulose membrane method as a qualitative
test for the ability of IAA production by the isolates (Etesami et al., 2009).
Indigenous soil samples were tested for Phosphate solubilization.
Efficient phosphate solubilizngbacteria were isolated. Effect of four different
media on phosphate solubilization were determined. Auxin production by these
bacteria were determined via bioassay and high performance liquid
chromatography by the bacteria in liquid culture. Indole Acetic Acid and indole
butyric acid were produced by these bacteria in varying concentration with and
without the addition of tryptophan. These bacteria showed stimulatory effects on

66
the growth of root and shoot elongation of Mung beans (Vigna radiata). Three
promising isolates were investigated to establish the effect on plant growth
(Shahab et al., 2009).
The filamentous cyanobacterium Arthrospira platensis strain was isolated
from a rice field. The ability of this strain to synthesize the bioactive compound
Indole Acetic Acid was demonstrated. IAA was extracted from the culture of
Arthrospira platensis strain and its identity was confirmed by thin-layer
chromatography as well as by high-performance liquid chromatography. The
IAA precursor L-tryptophan was required for IAA biosynthesis (Mehboob et al.,
2010).
A total of 216 bacterial strains were isolated from rice rhizospheric soils
in Northern Thailand. The bacterial strains were initially tested for solubilization
of inorganic phosphate, Indole Acetic Acid production, selected strains were
then tested for optimized conditions for Indole Acetic Acid production. It was
found that all strains had solubilized inorganic phosphate. The best Indole Acetic
Acid producer was identified by biochemical testing and DNA sequence
analysis as Klebsiella. In addition to being the best Indole Acetic Acid
producer, this strain was a high phosphate solubilizer. The production of Indole
Acetic Acid was further confirmed by extraction of crude Indole Acetic Acid
from this isolate and subsequent Thin Layer Chromatography analysis
(Chaiharn and Lumyong, 2011).

67
Materials and Methods
Glass wares cleaning
All the glass wares were soaked in Potassium dichromate solution (100 g
of Potassium dichromate in 1 liter of distilled water followed by slow addition of
500 mL concentrated Sulfuric acid) for about 12 hrs and rinsed in tap water.
Finally, they were washed in distilled water, dried and used.
Sterilization
The glass wares were sterilized at 180ºC for one hour in hot air oven. All
the media were autoclaved at 15 lbs pressure for 15 min.
Chemicals
All the chemicals and media used in this work were purchased from Hi-
media, Ranbaxy and Sigma limited and distilled water was used throughout the
study.
pH adjustment
The pH of the medium was adjusted with 0.1N NaOH or 0.1N HCl.
Chemicals used
1. Sodium Azide (Sigma) Chemical Mutagen.
2. Ethyl Methane Sulphonate (Sigma) Chemical Mutagen.
3. Ammonium Molybdate (Ranbaxy) for Chloromolybdic acid reagent
preparation.
4. Stannous Chloride (Ranbaxy) for Chlorostannous acid reagent
preparation.
5. Hydrochloric acid (Ranbaxy) Solution preparation and pH adjustment.

68
6. Phosphate Buffer (pH 7.0)
Dissolve 1.20g of sodium dihydrogen phosphate and 0.885g of disidium
hydrogen phosphate in 1 liter volume distilled water.
7. Chloromolybdic acid reagent for Phosphate Estimation
Ammonium molybdate (15g) was dissolved in 400ml of warm distilled
water. The solution was cooled and 300ml of 10.0N Hydrochloric acid
was added slowly. This was diluted to 1000ml after cooling.
8. Chlorostannous acid reagent for Phosphate Estimation
Stannous chloride (25g) was dissolved in 50ml of concentrated
Hydrochloric acid and was diluted to 500ml using recently boiled
distilled water. This solution was diluted with 1.2N HCl to about one
liter.
9. Sulphuric acid (Ranbaxy) (7N) for Phosphate estimation
196 ml concentrated sulphuric acid in 1000 ml distilled water.
10. Staining solutions Kit (Himedia) contains solutions of Gram Crytal Violet, Gram Iodine andGram Safranin for Gram reaction of bacteria.
11. Kovac reagent (Himedia) for Biochemical identification of bacteria.
12. Barrit’s reagent (Himedia) for Biochemical identification of bacteria.
13. Lacto Phenol Cotton Blue (Himedia) Spore staining of fungal cultures.
14. Potassium di-hydrogen orthophosphate (Ranbaxy)for Standard graph
preparation for Phosphate estimation.
15. Paranitro phenyl phosphate (PNP-P) (Sigma) (1%) : Substrate for Acid
Phosphatase assay.
Dissolve 1 g of paranitro phenyl phosphate in 100 ml of distilled water.
16. Para nitro phenol (PNP) (sigma): Preparation of standard graph for Acid
Phosphatase assay
Dissolve 1.5 g of paranitro phenol in 100 ml of distilled water.
17. Magnesium chloride (Ranbaxy) (1mM) for Acid Phosphatase assay
Dissolve 20.331 mg of Magnesium Chloride in 100 ml of distilled water.

69
18. Sodium Hydroxide (Ranbaxy) (0.2M) for Acid Phosphatase assay
Dissolve 8 g of pellets Sodium hydroxide in 1000 ml of distilled water.
19. Sodium Acetate buffer (Ranbaxy) (0.1 M, pH 5.8) for Acid Phosphatase
assay
a) Dissolve 0.6g of Sodium acetate in 100 ml of Distilled water.
b) Dissolve 0.6 ml of acetic acid in 100 ml of distilled water.
From ‘a’ take 90 ml and from ‘b’ take 10 ml make the final volume of
100 ml.
20. Gum Arabic (Ranboxy) for Lipase Assay
3g Gum Arabic in 100 ml distilled water.
21. Olive Oil (Himedia) for Lipase Assay.
22. O.1 M Phosphate buffer (pH 7.0) for Lipase Assay.
23. Alcoholic Phenolphthalein indicator (Himedia) for Lipase Assay.
24. Sodium Hydroxide (Ranbaxy) (0.02N)
Diluted from 1.0N Sodium hydroxide stock solution. (20ml stock made
up to 1000ml with distilled water.
25. Sodium Hydroxide (Ranbaxy) (1.0N)
40 g Sodium Hydroxide pellets dissolved in distilled water and made up
to 1000ml with distilled water.
26. Silica gel G (Ranbaxy) for Thin Layer Chromatograpy.
27. Salkowski reagent for Indole Acetic Acid estimation
2.03 g Ferric chloride dissolved in 500 ml water and 300 ml concentrated
sulphuric acid.
28. Ethyl acetate, Chloroform and Formic acid (Ranbaxy) solvent system for
Indole acetic Acid identification by Thinlayer Chromatograpy.
29. Glucose, Sucrose, Lactose and Mannitol (Ranboxy) act as carbon sources.
30. Ammonium sulphate, Potassium Nitrate, Sodium Nitrate and Urea
(Ranboxy) act as nitrogen sources.

70
Media used
Nutrient agar medium (Hi-media) for isolation and culture maintenance.
Sabouraud agar medium (Hi-media) for isolation and culture
maintenance.
Starch agar medium (Hi-media) for biochemical identification of bacteria.
Casein agar medium (Hi-media) for biochemical identification of
bacteria.
Certimide agar (Hi-media) for biochemical identification of bacteria.
MR-VP broth (Hi-media) for biochemical identification of bacteria.
Christensen citrate agar medium (Hi-media) for biochemical
identification of bacteria.
TSI agar medium (Hi-media) for biochemical identification of bacteria.
SIM agar medium (Hi-media) for biochemical identification of bacteria.
Urea agar base (Hi-media) for biochemical identification of bacteria.
Gelatin agar (Hi-media) for biochemical identification of bacteria.
Trypticase Nitrate broth (Hi-media) for biochemical identification of
bacteria.
Asparagine- Proline broth (Hi-media) for biochemical identification of
bacteria.
Pikovskaya medium (Hi-media) for isolation of Phosphate solubilizing
bacteria.
Pikovskaya broth (Hi-media) for Phosphate estimation and Acid
Phosphatase assay.
Czapek’s dox broth (Hi-media) Lipase and Indole Acetic Acid assay.
Carbohydrate fermentation medium contains Glucose or Sucrose or
Lactose or mannitol as a carbon source for biochemical identification of
bacteria.

71
Soil sampling
Different types of rhizosphere soil were collected from the rice fields in
and around Mannachanalur area of Trichy ditrict, Tamilnadu for the isolation of
bacterial and fungal cultures.
Isolation of phosphate solubilizing Microorganism
(Manjugupta and Bhriguvansji, 1997).
These soils were mixed and subjected to serial dilution technique using
the stock soil suspension which contains 10 g of soil sample in 100 ml of sterile
distilled water. From the serial diluted suspension, 1 mL of suspension from
10-3 to 10-6 dilution was taken for pour plate technique in Nutrient agar,
Sabouraud agar and Pikovskaya agar medium. The Nutrient agar plates were
observed after 2 days of incubation for enumeration of bacterial colonies, the
Sabouraud agar plates were observed after 3days of incubation for enumeration
of fungal colonies. After 2-3 days of incubation, the plates were observed for
phosphate solubilizing bacterial and fungal cultures from Pikovskaya agar
medium. The bacterial cultures were then transferred to nutrient agar slants and
fungal cultures to the Sabouraud agar slants for further studies.
Identification of Bacteria (Cowan, 1974) and Fungi (Josephgilman, 2001)
The bacterial culture was identified up to genus level by cultural,
morphological and biochemical characteristics. From the 24 hrs broth culture of
the isolated bacteria, 1ml of the culture was inoculated into Kings B agar,
Cetrimide agar, Asparagine –Proline broth, MR-VP broth and carbohydrate
fermentation tubes. One loop ful of culture was inoculated into Christensan
citrate agar, Urea agar, Gelatin agar, Starch agar, SIM medium (Stab) along with
Oxidase and Catalase test.

72
After 24 – 48 hrs incubation, the results were observed in the Kings B
agar and Cetrimide agar for colony morphology and pigmentation of the
bacteria. Asparagine –Proline broth was observed for its fluorescents activity in
UV transilluminator and other tests for the biochemical characterization purpose.
The bacterial culture was identified based on the observed results using
bacteriological manual.
The fungal cultures were identified based on the colony morphology and
spore structure. The spore structure was identified microscopically by Lacto
phenol cotton blue method.
The above isolated bacterial and fungal cultures were tested for its
Phosphate solubilization property and Phosphatase, Lipase and Indole Acetic
Acid producton cabablity in liquid broth.
Phosphate solubilization in liquid broth
Inoculation (Sudhansupal, 1999; Varshanarishan et al., 1995)
The pure bacterial culture from nutrient agar slant was inoculated into
nutrient broth. After 18 hrs of incubation, 0.5 mL of nutrient broth culture (1
O.D) was inoculated into 100 mL Pikovskaya broth containing 1g of Tricalcium
Phosphate. The flask was incubated at 37 ºC. The Pure fungal cultures from
Sabouraud agar slants were transferred to Sabouraud agar plates. After four
days of incubation at 27 ºC, two 8 mm disc were cut from the plates by sterile
borer from all three fungal plates and were inoculated into 100 ml Pikovskaya
broth separately. The flasks were incubated at 27 ºC. Uninoculated Pikovskaya
broth served as control in each case. Each experiment was done in triplicate set.

73
Phosphate estimation (Artidave and Patel, 1999; Seshadri, 1995)
Growth medium was withdrawn aseptically at three days interval from
each flask and centrifuged at 10, 000 rpm for 20 min. The supernatant was
analyzed for phosphate content by Chlorostannous Reduced Molybdophosphoric
Blue Color Method. The clear 10 mL of supernatant was collected in a 50 mL
volumetric flask and 10 mL of chloromolybdic acid reagent was added along the
sides to the flask. The content was diluted to 40 mL and 5 drops of
chlorostannous acid reagent was added. After thorough mixing the volume was
made up to 50 mL quickly. The optical density was measured at 600 nm and the
value was obtained with the help of the standard curve. Pigments produced by
some of the cultures interfere with color development which was overcome by
the addition of 1g of activated charcoal to each flask. The solution was filtered
and the filtrate was taken for further analysis.
Preparation of standard graph
Potassium di-hydrogen orthophosphate (0.2195g) was dissolved in
400 mL distilled water and 25 mL of 7N H2SO4 was added. This solution was
made up to 1000 mL with distilled water, which served as stock phosphate
solution. From that 20 mL solution was taken and diluted to 500 mL, which was
secondary phosphate solution. From the secondary solution 0.5 mL, 1 mL, 2.5
ml, 5 mL, 7.5ml, 10.0ml, 12.5ml, 15.0ml, 20.0ml and 25ml solutions was taken
and Phosphate estimation was carried out and Optical Density (O.D) value was
obtained as described above. The O.D values (X-axis) and phosphate in ppm
(Y-axis) were plotted on a graph.
Random UV mutational studies for Bacteria (Kumar and Dhruv, 1990)
The 24 hrs old parent culture was used for the preparation of bacterial
suspension, which was diluted with phosphate buffer (pH 7.0) contains 108

74
cells/ml. The above culture was inoculated over the surface of nutrient agar
plates. Then the plates were exposed to UV light (2600 Ao) at a distance of 15cm
in different time intervals and one plate was kept as growth control without
exposing to UV light for control. After the above process the plates were
covered with black paper to avoid light induced repair mechanism and were
incubated at 37ºC for 2 days.
Then the cultures were tested for phosphate solubilization as described
above.
Random UV mutational studies for Fungi (Ellaiah et al., 2002)
The growth of 72 hrs old fungal cultures were scrapped off in 5ml sterile
distilled water and diluted with 45 ml of sterile distilled water containing Tween
80 (1:4000). Sterile glass beads were added and were shaked on a rotary shaker
for 30 min to break the hyphal mycelium. The suspension was filtered by using
sterile cotton cloth to remove the mycelium. The spore suspension was prepared
in phosphate buffer (pH 7.0) containing 106 spores per ml. Five ml quantities of
the spore suspension was pipette aseptically into sterile petri dishes of 90 mm
diameter having a flat bottom. Then the plates were exposed to UV light (2600
Ao) at a distance of 15cm from the center of the Germicidal lamp (UV light
source) in different time intervals.
The exposure times were 10, 20, 30, 40, 50, 60, 70 and 80 min. Each UV
exposed spore suspension was stored in dark overnight to avoid photo
reactivation, then was serially diluted in phosphate buffer and plated on
Sabouraud agar medium. The plates were incubated for 5 days at 27°C and the
number of colonies in each plate were counted. Each colony was assumed to be
formed from a single spore.

75
Then the cultures were tested for phosphate solubilization as described
earlier.
The unexposed fungal suspension was inoculated into the Sabouraud
agar medium that served as a growth (positive) control.
Random Chemical mutational study (Bapiraju et al., 2004)
Sodium Azide and Ethyl Methane Sulphonate (EMS) were the chemical
mutagens used in the present study for the strain improvement of phosphate
solubilizers. The spore suspension of fungal cultures was prepared by using
phosphate buffer pH 7.0 as described earlier. To 9 ml of spore suspension, 1 ml
of sterile solution of Sodium Azide (250 μg ml-1 in phosphate buffer) and Ethyl
Methane Sulphonate (EMS) (150 μg ml-1 in phosphate buffer) was added. The
24 hrs old bacterial culture was used for the preparation of bacterial suspension,
which was diluted with phosphate buffer (pH 7.0) contains 108 cells/ml. To a 9
ml of bacterial suspension, 1 ml of sterile solution of Sodium Azide (250 μg ml-1
in phosphate buffer) and Ethyl Methane Sulphonate (EMS) (150 μg ml-1 in
phosphate buffer) was added. The above culture was inoculated and Samples
were withdrawn from the reaction mixture at an interval of 30, 60, 90, 120 and
150 min. and centrifuged for 10 min. at 5000 rpm. The cells were washed three
times with sterile distilled water and again re-suspended in 10 ml sterile buffer.
The samples were serially diluted in the same buffer and plated over Nutrient
agar and Sabouraud agar medium for bacteria and fungi respectively.
Then the cultures were tested for phosphate solubilization as described
earlier.

76
The unexposed fungal suspension was inoculated into the Sabouraud
agar medium that served as a growth (positive) control.
Effect of different carbon sources
To study the effect of different carbon sources on phosphate
solubilization, the carbon source of Pikovskaya broth i. e. Glucose was replaced
by Lactose, Sucrose and Mannitol. The mutated cultures which were superior in
Phosphate solubilization were inoculated, incubated and Phosphate
solubilization assay was carried out as described earlier.
Effect of Different Nitrogen Sources
To study the effect of different nitrogen sources on phosphate
solubilization, the nitrogen source of Pikovskaya broth i.e. Ammonium Sulphate
was replaced by Potassium Nitrate, Sodium Nitrate and Urea. The mutated
cultures which were superior in Phosphate solubilization were inoculated,
incubated and Phosphate solubilization assay was carried out as described
earlier.
Acid phosphatase Production
Inoculation (Sudhansupal, 1999; Varshanarishan et al., 1995)
The pure bacterial culture from nutrient agar slant was inoculated on
nutrient broth. After 18 hours incubation, 0.5 ml of nutrient broth culture (1
O.D) was inoculated in 100 ml Pikovskaya broth containing 1g of Tricalcium
phosphate. The flask was incubated at 37oC. The pure fungal cultures from
Sabouraud agar slants were transferred to Sabouraud agar plates. After four
days incubation at 27oC, two 8 mm disc were cut from the plates of all three
fungal plates and were inoculated into 100 ml Pikovskaya broth. The flasks

77
were incubated at 27oC. Uninoculated Pikovskaya broth served as control in
each case. Each experiment was done in triplicate set.
Acid phosphatase assay (Eileen ingham et al., 1979)
The enzyme Acid phosphatase was assayed using para- nitrophenyl
phosphate (PNP-P) as substrate. The reaction mixture contained 2.5 ml (0.1M)
sodium acetate buffer (pH 5.8), I ml (1mM) Magnisium chloride, 0.5 ml 1%
PNP-P and 0.5ml of a suitable dilution of enzyme preparation. One ml of the
reaction mixture was transferred to 2ml of 0.2M Sodium Hydroxide before and
after 15 min incubation at 370C to stop the reaction. The Sodium Hydroxide
solution added before incubation act as a control sample for each analysis. The
amount of Para- nitro phenol (PNP) liberated was measured by recording the
absorbance at 420 nm using an appropriate calibration curve. Activity is
expressed as μmol PNP liberated min-1 .The blank was run in a similar manner
using distilled water.
Preparation of Standard curve
Dissolve 1.0 g of Para- nitro phenol in water, and dilute the solution to
1000 ml. This is standard Para nitro phenol solution. Store the solution in a
refrigerator. To prepare the standard graph, dilute 1 ml of the standard Para-
nitro phenol solution to 100 ml in a volumetric flask and mix the solution
thoroughly. Then pipette 0, 1, 2, 3, 4 and 5-ml aliquots (equaling to 0, 10, 20, 30,
40 and 50 μg of Para- nitro phenol) of this diluted standard solution into 5 ml
volumetric flask and made up to 5 ml by addition of water, and proceed as
described above. The O.D value (Y axis) and corresponding Para nitro phenol
concentration (X axis) were plotted in a graph.

78
Random mutational studies
Physical and chemical mutations were carried out as described earlier.
The bacterial and fungal cultures, obtained after Physical and chemical mutation
were tested for Acid Phosphatase as described earlier.
Effect of Different Carbon Sources
To study the effect of different carbon sources on Phosphate
solubilization, the carbon source of Pikovskaya broth i. e. Glucose was replaced
by Latose, Sucrose and Mannitol. Then the mutated cultures which were
superior in Acid Phosphatase were inoculated, incubated and Acid Phosphatase
assay was carried out as described earlier.
Effect of Different Nitrogen Sources
To study the effect of different nitrogen sources on Phosphate
solubilization, the nitrogen source of Pikovskaya broth i.e. Ammonium Sulphate
was replaced by Potassium Nitrate, Sodium Nitrate and Urea. The mutated
cultures which were superior in Acid Phosphatase actvty in Pikovskaya broth
were inoculated, incubated and Acid Phosphatase assay was carried out as
described earlier.
Lipase Production
Inoculation (Sudhansupal, 1999; Varshanarishan et al., 1995)
The pure bacterial culture from nutrient agar slant was inoculated on
nutrient broth. After 18 hours incubation, 0.5 ml of nutrient broth culture (1
O.D) was inoculated in 100 ml Czapek’s dox broth containing 1% olive oil as
inductor and also 10% olive oil in 100ml Czapek’s dox broth instead of Sucrose.
The pure fungal culture from Sabouraud agar slant were transferred to
Sabouraud agar plates. After four days incubation at 27°C, two 8mm disc were
cut from all three fungal plates and inoculated into 100ml Czapek’s dox broth

79
containing 1% olive oil as inductor and also 10% olive oil in 100ml Czapek’s
dox broth instead of Sucrose. The flasks were incubated at 27° C. Uninoculated
Czapek’s dox broth served as a control in each case. Each experiment was done
in triplicate set.
Lipase assay (Ray et al., 1999)
Growth medium was withdrawn aseptically at three days interval from
each flask and centrifuged at 3000rpm for 15 minutes and the supernatant was
collected and used for lipase activity.
The enzyme solution prepared by mixing the supernatant solution with
1ml of olive oil and 3% gum Arabic in 250ml conical flask and the contents
were emulsified using magnetic stirrer for 5 minutes at top speed.
After emulsification, 10ml of enzyme solution was taken in a flask
followed by 20ml of double distilled water and 5ml of 0.1M phosphate buffer
(pH 7). This mixture was kept on rotary shaker for 30 minutes at 120rpm.
Simultaneously the same protocol was followed except for the enzyme solution
and this was treated as a control. After 30 minutes a drop of 1% alcoholic
phenolphthalein solution was added and titrated against 0.02 N Sodium
Hydroxide (NaOH) till the appearance of pale pink colour.
Lipase activity
One unit of lipase activity was defined as the amount of enzyme required
to release one µ mol of free fatty acid in one min under standard assay
conditions.
Volume of Normality of NaOH consumed × NaOH Lipase activity = ––––––––––––––––––––––––––––––––––––––––––– × 1000 Time of incubation × Volume of enzyme solution expressed as Unit/ Substrate

80
Random mutational studies
Physical and chemical mutation was carried out as described earlier. The
bacterial and fungal cultures, obtained after Physical and chemical mutation
were tested for Lipase as described above.
Effect of different Carbon Sources
To study the effect of different carbon sources on lipase production, the
carbon source of Czapek‘s dox broth i.e. Sucrose was replaced by Lactose,
Glucose and Mannitol. The mutated cultures which were superior in Lipase
activity were inoculated, incubated and Lipase assay was carried out as
described earlier. When Sucrose, Lactose, Glucose and Mannitol as a carbon
source in Czapek’s dox broth, 1% olive oil was used as a inducer.
Effect of different Nitrogen Sources
To study the effect of different nitrogen sources on lipase production, the
Nitrogen source of Czapek’s dox broth i.e. Sodium Nitrate was replaced by
Potassium Nitrate, Ammonium Sulphate and Urea. The mutated cultures which
were superior in Lipase activity were inoculated, incubated and Lipase assay was
carried out as described earlier. When Sodium Nitrate Potassium Nitrate,
Ammonium Sulphate and Urea as a nitrogen source in Czapek’s dox broth, 1%
olive oil was used as a inducer.
Indole Acetic Acid (IAA)
Indole Acetic Acid Production (Maria Guineth et al., 2000)
The isolated bacterial and fungal cultures were subjected to Indole Acetic
Acid production by inoculating the cultures in Czapek’s dox broth.

81
The pure bacterial culture from nutrient agar slants was inoculated on
nutrient broth. After 18 hrs incubation, 0.5 mL of nutrient broth culture (1 O.D)
was inoculated in 100 mL Czapek’s dox broth and incubated at 280C for 3 days.
The pure fungal cultures from sabouraud agar slants were transferred to
Sabouraud agar plates. After 5 days incubation at 270C two 8mm discs were cut
from the fungal plates & were inoculated separately into 100ml Czapek’s dox
broth. The flask were incubated at 300C for 4 days. (Mittal et al., 2008)
Screening of Indole Acetic Acid producer (Maria Guineth et al., 2000)
Culture broth was withdrawn aseptically after three days incubation for
bacteria and five days incubation for fungi from each flask and centrifuged at
3000 rpm for 15 minutes and the supernatant was collected and used for IAA
assay by Thin layer chromatography. The centrifuged sample of each broth were
brought to pH 3.0 and extracted three times with ethyl acetate. The organic
phase was concentrated to dryness and the diluted with 0.5 ml methanol.
Application of this solution on silica gel G plate (20cm X 5cm) and
chromatogram was developed with Chloroform –ethyl acetate-formic acid
(5:3:2) solvent system and then developed with Salkowiski reagent.
Quantitative estimation of Indole Acetic Acid (Tsavkelova et al., 2007)
The quantitative estimation of Indole Acetic Acid in broth cultures were
determined by a colorimetric assay technique using Salkwoski reagent. The two
part of the supernatant, one part of the Salkowski’s reagent was added. The O.D
was determined at 530nm in a spectrophotometer. A standard curve was
prepared from serial dilutions of IAA stock solution.

82
Preparation of standard graph for Indole Acetic Acid
1. 100mg of Indole Acetic Acid was weighed and dissolved in 100ml of
distilled water by slight heating. From this stock solution, Indole Acetic
Acid standard solutions were as prepared in different concentrations.
2. 8ml of the Indole Acetic Acid standard solution was taken with 2ml
Salkowski reagent. It was incubated at room temperature in dark for 30
minutes. Blank was prepared by taking 8ml of distilled water with 2ml of
Salkowski reagent.
3. Optical density was read at 530nm and plotted a graph for the
concentration of Indole Acetic Acid against optical density.
Estimation of Indole Acetic Acid
1. 8ml of culture filtrate was taken and 2ml of Salkowski reagent was added
to it. It was incubated at room temperature in dark for 30 minutes for the
development of pink colour.
2. The optical density was read at 530nm.
3. The results were read from standard graph and expressed µg/L
Random mutational studies
Physical and chemical mutation was carried out as described earlier. The
bacterial and fungal cultures, obtained after Physical and chemical mutation
were tested for IAA qualitatively and quantitatively as described above.

83
Results
ISOLATION AND IDENTIFICATION
The Nutrient agar plates were observed after 2days incubation for
bacterial enumeration and the Sabouraud agar plates were observed after 3days
incubation for fungal enumeration. It was found that more number of bacterial
and fungal colonies in 103 and 104 dilution, 255 colony forming units of bacteria
and 25 colony forming units of fungi in 105 dilution, 29 colony forming units of
bacteria and 3 colony forming units of fungi in 105 dilution and in 107 dilution, it
was 3 colony forming units of bacteria and less than 1 colony forming units
fungi after incubation. Finally, it was calculated the initial mother suspension
contains 3 x 107 colony forming units of bacteria per mL and 3 x 106 colony
forming units of fungi per mL. The results were presented in Table 1.
The same serial diluted suspension (103 to 107 dilutions) were inoculated
into Pikovskaya agar the plates were observed after 3 days of incubation for
phosphate solubilizing microorganisms (PSB). The plates were inoculated with
the suspensions such as 103 and 104 dilution, showed more colonies and were too
numerous to count for both bacteria and fungi. In 105 dilution, 16 bacterial and 7
fungal colonies and in 106 dilution, 2 bacterial and 1fungal colonies were found
to be phosphate solubilizers. Based on the results, the between rhizosphere
microbes and phosphate solubilizers were calculated which was 15:1 for bacteria
and 3.5:1 for fungi. The results were presented in Table 2.
The Phosphate solubilizing bacterial cultures were found to be single type
based on colony morphology in Kings B agar and cetrimide agar and gram
reaction by gram staining. Pale white, translucent colonies Circular, convex, and
smooth colonies and Green mucoid colonies were observed in Kings B agar and

84
Cetrimide agar respectively. The gram staining showed gram negative reaction.
Further, it was observed as Indole, Urease, Methyl Red, Vogus Proskauer and
Hydrogen sulphide production were negative and Citrate, Gelatin hydrolysis,
Nitrate reduction, Starch hydrolysis, Catalase and Oxisdase were positive. The
isolated bacteria was motile, produced green color pigment in Cetrimide agar
and Asparagine –proline broth which fluorescents in UV Trans illuminator.
Carbohydrate fermentation tubes showed no gas production and acid production
found on top of the tube.
The isolated bacterial cultures were identified as Pseudomonas sp. by
cultural, morphological and biochemical characteristics. The results were
presented in Table 3.
Plate-1 showes the colonies of Pseudomonas sp. on Nutrient and
Cetrimide agar Asparagine –Proline broth tube in UV Trans illuminator and
Oxidase test.
Among the fungal cultures isolated, three fungal colonies were shown
clear zone around them in Pikovskaya agar medium were selected for further
studies. The fungal cultures were identified as Aspergillus niger, Aspergillus
fumigatus and Penicillium sp. based on the below observations in macroscopic
and microscopic methods.
Aspergillus niger showed black surface pigmentation with a dense felt of
conidiophores. Conidiophores are long, smooth walled and have round shaped
terminal vesicle which support a biseriate of phialides on the vesicle. Over the
phialides are the round conidia forming radial chains.

85
Aspergillus fumigatus showed green surface pigmentation with a dense
felt of conidiophores. Conidiophores are colorless, rough walled and have round
shaped terminal vesicle which support a biseriate of phialides on the vesicle.
Over the phialides are the round conidia forming radial chains.
Penicillium sp. showed green color colonies consisting of a dense felt of
conidiophores. Microscopically, conodiophores show branching, and phialides
produced in groups from branched metulae, giving brush-like appearance.
Conidia are globuse, greenish and smooth.
Plate-2 shows the growth of fungal colonies on Sabouraud agar plates
and Plate-3 shows the Phosphate solubilization of fungal colonies in Pikovskaya
agar.
Random UV Mutagenesis
The bacterial colonies gradually decreased with increasing exposure
time. There was no growth on plates which were exposed to UV light more than
8 minutes. Four bacterial strains resistant for 7 min UV exposure were selected,
designated as PSUV 1, PSUV 2, PSUV 3 and PSUV 4 and transferred to nutrient
broth and agar slants for further study.
The fungal cultures were selected from the plates exposed to 30, 40, 50
and 60 min UV exposure time and they were given the code numbers as
ANuv30, ANuv40, ANuv50 and ANuv60 for Aspergillus niger, Aspergillus
fumigatus mutants were AFuv30, AFuv40, AFuv50 and AFuv60 whereas the
Penicillium species mutants were given the code numbers PEuv30, PEuv40,
PEuv50 and PEuv60. There was no growth observed the plates inoculated with
fungal cultures at 70 and 80 miniutes UV exposure time.

86
Random Chemical Mutagenesis
The Aspergillus niger, Aspergillus fumigatus mutants cultures were
selected from the plates exposed to 30, 60, 90 and 120 min exposure time where
as Penicillium species mutants cultures from 60, 90, 120 and 150 min exposure
time of Sodium azide.
The Sodium Azide treated Aspergillus niger mutants were given the code
numbers ANsa30, ANsa60, ANsa90 and ANsa120, Aspergillus fumigatus
mutants were AFsa30, AFsa60, AFsa90 and AFsa120 whereas the Penicillium
species mutants were given the code numbers PEsa60, PEsa90, PEsa120 and
PEsa150.
The Aspergillus niger, Aspergillus fumigatus mutants cultures were
selected from the plates exposed to 30, 60, 90 and 120 min exposure time where
as Penicillium species mutants cultures from 60, 90, 120 and 150 min exposure
time of Ethyl Methane Sulphonate.
The Ethyl Methane Sulphonate treated Aspergillus niger mutants were
given the code numbers ANems30, ANems60, ANems90 and ANems120,
Aspergillus fumigatus mutants were AFems30, AFems60, AFems90 and
AFems120 whereas the Penicillium species mutants were given the code
numbers PEems60, PEems90, PEems120 and PEems150.
Then the cultures were tested for Phosphate estimation, Acid
Phosphatase, Lipase and Indole Acetic acid production.

87
Phosphate Solubilization
Phosphate Solubilization Capacity of UV Treated Aspergillus niger
The results of Phosphate Solubilization capacity of UV treated
Aspergillus niger were presented in Table 4.
The phosphate solubilization capacity of UV treated Aspergillus niger
(ANuv60) on day three recorded maximum efficacy in solubilization (3.05ppm
of Phosphate) followed by ANuv50 strain (2.85 ppm of Phosphate), ANuv30
(2.65 ppm of Phosphate) and ANuv40 (2.50 ppm of Phosphate). The wild
Aspergillus niger which was used as a standard control solubilized 2.20ppm of
Phosphate in the third day of incubation.
The phosphate solubilization capacity of UV treated Aspergillus niger
(ANuv60) on day six recorded maximum efficacy in solubilization (4.65ppm of
Phosphate) followed by ANuv50 strain (4.35 ppm of Phosphate), ANuv30 (4.05
ppm of Phosphate) and ANuv40 (3.85 ppm of Phosphate). The wild Aspergillus
niger which was used as a standard control solubilized 3.65 ppm of Phosphate in
the sixth day of incubation.
The phosphate solubilization capacity of UV treated Aspergillus niger
(ANuv60) on day nine recorded maximum efficacy in solubilization (5.85ppm of
Phosphate) followed by ANuv50 strain (5.50 ppm of Phosphate), ANuv30 (5.00
ppm of Phosphate) and ANuv40 (4.75 ppm of Phosphate). The wild Aspergillus
niger which was used as a standard control solubilized 4.60 ppm of Phosphate in
the ninth day of incubation.
The phosphate solubilization efficacy of ANuv60 strain increased 38.64%
on third day, 27.40% phosphate solubilization on sixth day and 27.17%

88
phosphate solubilization on ninth day of incubation using wild type as a standard
(100%).
Phosphate Solubilization Capacity of UV Treated Aspergillus fumigatus
The results of Phosphate Solubilization capacity of UV treated
Aspergillus fumigatus were presented in Table 5.
The phosphate solubilization capacity of UV treated Aspergillus
fumigatus (AFuv50) on day three recorded maximum efficacy in solubilization
(3.55ppm of Phosphate) followed by AFuv60 strain (3.35 ppm of Phosphate),
AFuv30 (3.25 ppm of Phosphate) and AFsa40 (3.15 ppm of Phosphate). The
wild Aspergillus fumigatus which was used as a standard control solubilized 2.95
ppm of Phosphate in the third day of incubation.
The phosphate solubilization capacity of UV treated Aspergillus
fumigatus (AFuv50) on day six recorded maximum efficacy in solubilization
(3.95ppm of Phosphate) followed by AFuv60 strain (3.85 ppm of Phosphate),
AFuv30 (3.65 ppm of Phosphate) and AFsa40 (3.65 ppm of Phosphate). The
wild Aspergillus fumigatus which was used as a standard control solubilized 3.25
ppm of Phosphate in the sixth day of incubation.
The phosphate solubilization capacity of UV treated Aspergillus
fumigatus (AFuv50) on day nine recorded maximum efficacy in solubilization
(4.55ppm of Phosphate) followed by AFuv60 strain (4.45 ppm of Phosphate),
AFuv40 (4.15 ppm of Phosphate) and AFsa30 (3.85 ppm of Phosphate). The
wild Aspergillus fumigatus which was used as a standard control solubilized 3.65
ppm of Phosphate in the ninth day of incubation.

89
The phosphate solubilization efficacy of AFuv50 strain increased 20.34%
on third day, 21.54% phosphate solubilization on sixth day and 24.66%
phosphate solubilization on ninth day of incubation using wild type as a standard
(100%).
Phosphate Solubilization Capacity of UV Treated Penicillium sp
The results of Phosphate Solubilization capacity of UV treated
Penicillium sp were presented in Table 6.
The phosphate solubilization capacity of UV treated Penicillium sp
(PEuv50) on day three recorded maximum efficacy in solubilization (3.75 ppm
of Phosphate) followed by PEuv40 strain (3.65 ppm of Phosphate), PEuv30
(3.45 ppm of Phosphate) and PEuv60 (3.40 ppm of Phosphate). The wild
Penicillium sp which was used as a standard control solubilized 3.30 ppm of
Phosphate in the third day of incubation.
The phosphate solubilization capacity of UV treated Penicillium sp
(PEuv50) on day six recorded maximum efficacy in solubilization (4.40 ppm of
Phosphate) followed by PEuv40 strain (4.25 ppm of Phosphate), PEuv60 (4.20
ppm of Phosphate) and PEuv30 (4.10 ppm of Phosphate). The wild Penicillium
sp which was used as a standard control solubilized 3.85 ppm of Phosphate in
the sixth day of incubation.
The phosphate solubilization capacity of UV treated Penicillium sp
(PEuv50) on day nine recorded maximum efficacy in solubilization (4.75 ppm of
Phosphate) followed by PEuv40 strain (4.45 ppm of Phosphate), PEuv60 (4.35
ppm of Phosphate) and PEuv30 (4.25 ppm of Phosphate). The wild Penicillium

90
sp which was used as a standard control solubilized 4.00 ppm of Phosphate in
the sixth day of incubation.
The phosphate solubilization efficacy of PEuv50 strain increased 13.64%
on third day, 14.29% phosphate solubilization on sixth day and 18.75%
phosphate solubilization on ninth day of incubation using wild type as a standard
(100%).
Phosphate Solubilization Capacity of UV Treated Pseudomonas sp.
The results of Phosphate Solubilization capacity of UV treated
Pseudomonas sp were presented in Table 7.
The phosphate solubilization capacity of UV treated Pseudomonas sp
(PSuv3) on day three recorded maximum efficacy in solubilization (4.15 ppm of
Phosphate) followed by PSuv4 strain (4.05 ppm of Phosphate), PSuv2 (3.95 ppm
of Phosphate) and PSuv1 (3.85 ppm of Phosphate). The wild Pseudomonas sp
which was used as a standard control solubilized 3.80 ppm of Phosphate in the
third day of incubation.
The phosphate solubilization capacity of UV treated Pseudomonas sp
(PSuv4) on day six recorded maximum efficacy in solubilization (4.35 ppm of
Phosphate) followed by PSuv3 strain (4.25 ppm of Phosphate), PSuv2 (4.10 ppm
of Phosphate) and PSuv1 (3.95 ppm of Phosphate). The wild Pseudomonas sp
which was used as a standard control solubilized 4.15 ppm of Phosphate in the
sixth day of incubation.
The phosphate solubilization capacity of UV treated Pseudomonas sp
(PSuv4) on day nine recorded maximum efficacy in solubilization (4.65 ppm of

91
Phosphate) followed by PSuv3 strain (4.45 ppm of Phosphate), PSuv2 (4.30 ppm
of Phosphate) and PSuv1 (4.15 ppm of Phosphate). The wild Pseudomonas sp
which was used as a standard control solubilized 4.75 ppm of Phosphate in the
ninth day of incubation.
All mutants of Pseudomonas strains treated with UV on day nine showed
decreased phosphate solubilization activity when compared to wild type.
Phosphate Solubilization Capacity of Sodium Azide Treated Aspergillus
niger
The results of Phosphate Solubilization capacity of sodium azide treated
Aspergillus niger were presented in Table 8.
The phosphate solubilization capacity of sodium azide treated Aspergillus
niger (ANsa120) on day three recorded maximum efficacy in solubilization
(3.45ppm of Phosphate) followed by ANsa90 strain (2.95 ppm of Phosphate),
ANsa60 (2.50 ppm of Phosphate) and ANsa30 (2.40 ppm of Phosphate). The
wild Aspergillus niger which was used as a standard control solubilized 2.20ppm
of Phosphate in the third day of incubation.
The phosphate solubilization of sodium azide treated Aspergillus niger
(ANsa120) on day six recorded maximum efficacy in solubilization (4.95 ppm of
Phosphate) followed by ANsa90 strain (4.65 ppm of Phosphate) ANsa60 (4.30
ppm of Phosphate) and ANsa30 (4.10 ppm of Phosphate). The wild Aspergillus
niger which was used as a standard control solubilized 3.65 ppm of Phosphate in
the sixth day of incubation.

92
The phosphate solubilization of sodium azide treated Aspergillus niger
(ANsa120) on day nine recorded maximum efficacy in solubilization (6.15 ppm
of Phosphate) followed by ANsa90 strain (5.80 ppm of Phosphate) ANsa60
(5.55 ppm of Phosphate) and ANsa30 (5.05 ppm of Phosphate). The wild
Aspergillus niger which was used as a standard control solubilized 4.60ppm of
Phosphate in the ninth day of incubation.
The phosphate solubilization efficacy of ANsa120 strain increased 56.8%
on third day, 35.6% phosphate solubilization on sixth day and 33.6% phosphate
solubilization on ninth day of incubation using wild type as a standard (100%).
Here the percentage of phosphate solubilization is decreased with the increasing
the day of incubation.
Phosphate Solubilization Capacity of Sodium Azide Treated Aspergillus
fumigatus
The results of Phosphate Solubilization capacity of sodium azide treated
Aspergillus fumigatus were presented in Table 9.
The phosphate solubilization capacity of sodium azide treated Aspergillus
fumigatus (AFsa120) on day three recorded maximum efficacy in solubilization
(4.05ppm of Phosphate) followed by AFsa90 strain (3.45 ppm of Phosphate),
AFsa60 (3.25 ppm of Phosphate) and AFsa30 (3.10 ppm of Phosphate). The
wild Aspergillus fumigatus which was used as a standard control solubilized 2.95
ppm of Phosphate in the third day of incubation.
The phosphate solubilization capacity of sodium azide treated Aspergillus
fumigatus (AFsa120) on day six recorded maximum efficacy in solubilization
(4.65ppm of Phosphate) followed by AFsa90 strain (4.00 ppm of Phosphate),

93
AFsa60 (3.75 ppm of Phosphate) and AFsa30 (3.25 ppm of Phosphate). The
wild Aspergillus fumigatus which was as a standard control solubilized 3.45 ppm
of Phosphate in the sixth day of incubation.
The phosphate solubilization capacity of sodium azide treated Aspergillus
fumigatus (AFsa120) on day nine recorded maximum efficacy in solubilization
(5.50ppm of Phosphate) followed by AFsa90 strain (5.30 ppm of Phosphate),
AFsa60 (4.25 ppm of Phosphate) and AFsa30 (3.95 ppm of Phosphate). The
wild Aspergillus fumigatus which was used as a standard control solubilized 3.65
ppm of Phosphate in the ninth day of incubation.
The phosphate solubilization efficacy of AFsa120 strain increased 37.0%
on third day, 43.0% phosphate solubilization on sixth day and 50.6% phosphate
solubilization on ninth day of incubation using wild type as a standard (100%).
Here the percentage of phosphate solubilization is increased with the increasing
the day of incubation.
Phosphate Solubilization Capacity of Sodium Azide Treated Penicillium sp
The results of Phosphate Solubilization capacity of sodium azide treated
Penicillium sp were presented in Table 10.
The phosphate solubilization capacity of sodium azide treated Penicillium
sp (PEsa150) on day three recorded maximum efficacy in solubilization (3.80
ppm of Phosphate) followed by PEsa120 strain (3.70 ppm of Phosphate), PEsa90
(3.60 ppm of Phosphate) and PEsa60 (3.50 ppm of Phosphate). The wild
Penicillium sp which was used as a standard control solubilized 3.30 ppm of
Phosphate in the third day of incubation.

94
The phosphate solubilization capacity of sodium azide treated Penicillium
sp (PEsa150) on day six recorded maximum efficacy in solubilization (4.85 ppm
of Phosphate) followed by PEsa120 strain (4.65 ppm of Phosphate), PEsa90
(4.50 ppm of Phosphate) and PEsa60 (4.15 ppm of Phosphate). The wild
Penicillium sp which was used as a standard control solubilized 3.85 ppm of
Phosphate in the sixth day of incubation.
The phosphate solubilization capacity of sodium azide treated Penicillium
sp (PEsa150) on day nine recorded maximum efficacy in solubilization (5.95
ppm of Phosphate) followed by PEsa120 strain (5.65 ppm of Phosphate), PEsa90
(5.05 ppm of Phosphate) and PEsa60 (4.35 ppm of Phosphate). The wild
Penicillium sp which was used as a standard control solubilized 4.00 ppm of
Phosphate in the ninth day of incubation.
The phosphate solubilization efficacy of PEsa150 strain increased 15.1%
on third day, 25.9% phosphate solubilization on sixth day and 48.7% phosphate
solubilization on ninth day of incubation using wild type as a standard (100%).
Phosphate Solubilization Capacity of Sodium Azide Treated Pseudomonas sp
The results of Phosphate Solubilization capacity of sodium azide treated
Pseudomonas sp were presented in Table 11.
The phosphate solubilization of sodium azide treated Pseudomonas sp.
(PSsa120) recorded maximum efficacy in solubilization (3.45 ppm of Phosphate)
followed by PSsa90 strain (3.35 ppm of Phosphate), PSsa60 strain (3.15 ppm of
Phosphate) and PSsa30 strain (2.85 ppm of Phosphate). The wild Pseudomonas
sp which was used as a standard control solubilized 3.80 ppm of Phosphate in
the third day of incubation.

95
The phosphate solubilization of sodium azide treated Pseudomonas sp.
(PSsa120) recorded maximum efficacy in solubilization (4.25 ppm of Phosphate)
followed by PSsa90 strain (4.05 ppm of Phosphate), PSsa60 strain (3.95 ppm of
Phosphate) and PSsa30 strain (3.65 ppm of Phosphate). The wild Pseudomonas
sp. which was used as a standard control solubilized 4.15 ppm of Phosphate in
the sixth day of incubation.
The phosphate solubilization of sodium azide treated Pseudomonas sp
(PSsa120) recorded maximum efficacy in solubilization (4.65 ppm of Phosphate)
followed by PSsa3 strain (4.50 ppm of Phosphate), PSsa60 strain (4.45 ppm of
Phosphate) and PSsa30 strain (4.15 ppm of Phosphate).. The wild Pseudomonas
sp which was used as a standard control solubilized 4.75 ppm of Phosphate in
the ninth day of incubation.
All mutants of Pseudomonas strains treated with sodium azide on day
three, six and nine showed decreased phosphate solubilization activity when
compared to wild type.
Phosphate Solubilization Capacity of Ethyl Methane Sulphonate Treated
Aspergillus niger
The results of Phosphate Solubilization capacity of Ethyl Methane
Sulphonate treated Aspergillus niger were presented in Table 12.
The phosphate solubilization capacity of Ethyl Methane Sulphonate
treated Aspergillus niger (ANems120) on day three recorded maximum efficacy
in solubilization (2.85ppm of Phosphate) followed by ANems90 strain (2.65
ppm of Phosphate), ANems60 (2.50 ppm of Phosphate) and ANems30 (2.30

96
ppm of Phosphate). The wild Aspergillus niger which was used as a standard
control solubilized 2.20ppm of Phosphate in the third day of incubation.
The phosphate solubilization capacity of Ethyl Methane Sulphonate
treated Aspergillus niger (ANems120) on day sixth recorded maximum efficacy
in solubilization (4.75ppm of Phosphate) followed by ANems90 strain (4.45
ppm of Phosphate), ANems60 (4.25 ppm of Phosphate) and ANems30 (3.90
ppm of Phosphate). The wild Aspergillus niger which was used as a standard
control solubilized 3.65ppm of Phosphate in the sixth day of incubation.
The phosphate solubilization capacity of Ethyl Methane Sulphonate
treated Aspergillus niger (ANems120) on day ninth recorded maximum efficacy
in solubilization (6.25ppm of Phosphate) followed by ANems90 strain (5.65
ppm of Phosphate), ANems60 (5.40 ppm of Phosphate) and ANems30 (4.75
ppm of Phosphate). The wild Aspergillus niger which was used as a standard
control solubilized 4.60 ppm of Phosphate in the ninth day of incubation.
The phosphate solubilization efficacy of ANems120 strain increased
29.5% on third day, 30.1% phosphate solubilization on sixth day and 35.8%
phosphate solubilization on ninth day of incubation using wild type as a standard
(100%).
Phosphate Solubilization Capacity of Ethyl Methane Sulphonate Treated
Aspergillus fumigatus
The results of Phosphate Solubilization capacity of Ethyl Methane
Sulphonate treated Aspergillus fumigatus were presented in Table 13.

97
The phosphate solubilization capacity of Ethyl Methane Sulphonate
treated Aspergillus fumigatus (AFems120) on day three recorded maximum
efficacy in solubilization (3.95ppm of Phosphate) followed by AFems90 strain
(3.75 ppm of Phosphate), AFems60 (3.55 ppm of Phosphate) and AFems30
(3.25 ppm of Phosphate). The wild Aspergillus fumigatus which was used as a
standard control solubilized 2.95 ppm of Phosphate in the third day of
incubation.
The phosphate solubilization capacity of Ethyl Methane Sulphonate
treated Aspergillus fumigatus (AFems120) on day six recorded maximum
efficacy in solubilization (4.45ppm of Phosphate) followed by AFems90 strain
(4.15 ppm of Phosphate), AFems60 (3.80 ppm of Phosphate) and AFems30
(3.45 ppm of Phosphate). The wild Aspergillus fumigatus which was used as a
standard control solubilized 3.25 ppm of Phosphate in the sixth day of
incubation.
The phosphate solubilization capacity of Ethyl Methane Sulphonate
treated Aspergillus fumigatus (AFems120) on day nine recorded maximum
efficacy in solubilization (5.15ppm of Phosphate) followed by AFems90 strain
(4.65 ppm of Phosphate), AFems60 (4.15 ppm of Phosphate) and AFems30
(3.85 ppm of Phosphate). The wild Aspergillus fumigatus which was used as a
standard control solubilized 3.65 ppm of Phosphate in the ninth day of
incubation.
The phosphate solubilization efficacy of AFems120 strain increased
33.8% on third day, 36.9% phosphate solubilization on sixth day and 41.0%
phosphate solubilization on ninth day of incubation using wild type as a standard
(100%).

98
Phosphate Solubilization Capacity of Ethyl Methane Sulphonate Treated
Penicillium sp
The results of Phosphate Solubilization capacity of Ethyl Methane
Sulphonate treated Penicillium sp were presented in Table 14.
The phosphate solubilization capacity of Ethyl Methane Sulphonate
treated Penicillium sp (PEems150) on day three recorded maximum efficacy in
solubilization (4.80 ppm of Phosphate) followed by PEems120 strain (4.55 ppm
of Phosphate), PEems90 (4.05 ppm of Phosphate) and PEems60 (3.65 ppm of
Phosphate). The wild Penicillium sp which was used as a standard control
solubilized 3.30 ppm of Phosphate in the third day of incubation.
The phosphate solubilization capacity of Ethyl Methane Sulphonate
treated Penicillium sp (PEems150) on day six recorded maximum efficacy in
solubilization (5.70 ppm of Phosphate) followed by PEems120 strain (5.10 ppm
of Phosphate), PEsa90 (4.90 ppm of Phosphate) and PEems60 (4.70 ppm of
Phosphate). The wild Penicillium sp which was used as a standard control
solubilized 3.85 ppm of Phosphate in the sixth day of incubation.
The phosphate solubilization capacity of Ethyl Methane Sulphonate
treated Penicillium sp (PEems150) on day six recorded maximum efficacy in
solubilization (5.95 ppm of Phosphate) followed by PEems120 strain (5.45 ppm
of Phosphate), PEems90 (5.30 ppm of Phosphate) and PEems60 (5.15 ppm of
Phosphate). The wild Penicillium sp which was used as a standard control
solubilized 4.00 ppm of Phosphate in the ninth day of incubation.
The phosphate solubilization efficacy of PEems150 strain increased
45.4% on third day, 48.0% phosphate solubilization on sixth day and 48.7%

99
phosphate solubilization on ninth day of incubation using wild type as a standard
(100%).
Phosphate Solubilization Capacity of Ethyl Methane Sulphonate Treated
Pseudomonas sp
The results of Phosphate Solubilization capacity of Ethyl Methane
Sulphonate treated Pseudomonas sp were presented in Table 15.
The phosphate solubilization of Ethyl Methane Sulphonate treated
Pseudomonas sp (PSems150) on day three recorded maximum efficacy in
solubilization (2.45 ppm of Phosphate) followed by PSems120 strain (2.40 ppm
of Phosphate), PSems90 (2.35 ppm of Phosphate) and PSems60 (2.30 ppm of
Phosphate). The wild Pseudomonas sp which was used as a standard control
solubilized 3.80 ppm of Phosphate in the third day of incubation.
The phosphate solubilization of Ethyl Methane Sulphonate treated
Pseudomonas sp (PSems150) on day six recorded maximum efficacy in
solubilization (3.25ppm of Phosphate) followed by PSems120 strain (3.05 ppm
of Phosphate) PSems90 (2.90 ppm of Phosphate) and PSems60 (2.80 ppm of
Phosphate). The wild Pseudomonas sp which was used as a standard control
solubilized 4.15 ppm of Phosphate in the sixth day of incubation.
The phosphate solubilization of Ethyl Methane Sulphonate treated
Pseudomonas sp (PSems150) on day nine recorded maximum efficacy in
solubilization (4.25ppm of Phosphate) followed by PSems120 strain (4.15 ppm
of Phosphate) PSems90 (3.55 ppm of Phosphate) and PSems60 (3.30 ppm of
Phosphate). The wild Pseudomonas sp which was used as a standard control
solubilized 4.75 ppm of Phosphate in the ninth day of incubation.

100
All mutants of Pseudomonas strains treated with Ethyl Methane
Sulphonate on day three, six and nine showed decreased phosphate
solubilization activity when compared to wild type.
Effect of Different Carbon and Nitrogen Sources on the Efficacy of
Phosphate Solubilization
The superior mutated cultures of Phosphate solubilization in Pikovskaya
broth were identified as Ethyl Methane Sulphonate treated Aspergillus niger
(ANems120), Sodium Azide Treated Aspergillus niger (ANsa120) and Sodium
Azide Treated Penicillium sp (PEsa150) and their efficacy of Phosphate
solubilization on different Carbon and Nitrogen Sources were studied.
Effect of Different Carbon Sources on the Efficacy of Phosphate
Solubilization of Ethyl Methane Sulphonate Treated Aspergillus niger
(ANems120)
The results of different carbon sources on the efficacy of phosphate
solubilization of Ethyl Methane Sulphonate treated Aspergillus niger
(ANems120) were presented in Table 16
The efficacy of phosphate solubilization of ANems120 was maximum in
the medium containing Glucose (2.85 ppm of Phosphate) followed by Sucrose
(2.40 ppm of Phosphate), Lactose (2.20 ppm of Phosphate) and Mannitol (2.05
ppm of Phosphate) in the third day of incubation.
The efficacy of phosphate solubilization of ANems120 was maximum in
the medium containing Glucose (4.75 ppm of Phosphate) followed by Sucrose

101
(3.75 ppm of Phosphate), Lactose (3.60 ppm of Phosphate) and Mannitol (3.05
ppm of Phosphate) in the sixth day of incubation.
The efficacy of phosphate solubilization of ANems120 was maximum in
the medium containing Glucose (6.25 ppm of Phosphate) followed by Sucrose
(4.85 ppm of Phosphate), Lactose (4.60 ppm of Phosphate) and Mannitol (4.35
ppm of Phosphate) in the ninth day of incubation.
Effect of Different Nitrogen Sources on the Efficacy of Phosphate
Solubilization of Ethyl Methane Sulphonate Treated Aspergillus niger
(ANems120)
The results of different nitrogen sources on the efficacy of phosphate
solubilization of Ethyl Methane Sulphonate treated Aspergillus niger
(ANems120) were presented in Table 17.
The efficacy of phosphate solubilization of ANems120 was maximum in
the medium containing Ammonium sulphate (2.85 ppm of Phosphate) followed
by Urea (2.40 ppm of Phosphate), Potassium nitrate (2.35 ppm of Phosphate)
and Sodium nitrate (1.90 ppm of Phosphate) in the third day of incubation.
The efficacy of phosphate solubilization of ANems120 was maximum in
the medium containing Ammonium sulphate (4.75 ppm of Phosphate) followed
by Urea (3.70 ppm of Phosphate), Potassium nitrate (3.70 ppm of Phosphate)
and Sodium nitrate (2.65 ppm of Phosphate) in the sixth day of incubation.
The efficacy of phosphate solubilization of ANems120 was maximum in
the medium containing Ammonium sulphate (6.25 ppm of Phosphate) followed
by Urea (4.75 ppm of Phosphate), Potassium nitrate (4.70 ppm of

102
Phosphate)and Sodium nitrate (3.20 ppm of Phosphate) in the ninth day of
incubation.
Effect of Different Carbon Sources on the Efficacy of Phosphate
Solubilization of Sodium Azide Treated Aspergillus niger (ANsa120).
The results of different carbon sources on the efficacy of phosphate
solubilization of sodium azide treated Aspergillus niger (ANsa120) were
presented in Table 18
The efficacy of phosphate solubilization of ANsa120 was maximum in
the medium containing Sucrose (3.55 ppm of Phosphate) followed by Glucose
(3.45 ppm of Phosphate), Lactose (3.05 ppm of Phosphate) and Mannitol (1.85
ppm of Phosphate) in the third day of incubation.
The efficacy of phosphate solubilization of ANsa120 was maximum in
the medium containing Glucose (4.95 ppm of Phosphate) followed by Sucrose
(4.53 ppm of Phosphate), Lactose (3.55 ppm of Phosphate) and Mannitol (2.15
ppm of Phosphate) in the sixth day of incubation.
The efficacy of phosphate solubilization of ANsa120 was maximum in
the medium containing Glucose (6.15 ppm of Phosphate) followed by Sucrose
(4.95 ppm of Phosphate), Lactose (3.95 ppm of Phosphate) and Mannitol (2.90
ppm of Phosphate) in the ninth day of incubation.

103
Effect of Different Nitrogen Sources on the Efficacy of Phosphate
Solubilization of Sodium Azide Treated Aspergillus niger (ANsa120).
The results of different nitrogen sources on the efficacy of phosphate
solubilization of sodium azide treated Aspergillus niger (ANsa120) were
presented in Table 19.
The efficacy of phosphate solubilization of ANsa120 was maximum in
the medium containing Ammonium sulphate (3.45 ppm of Phosphate) followed
by Urea (2.45 ppm of Phosphate), Potassium nitrate (2.40 ppm of Phosphate)
and Sodium nitrate (1.75 ppm of Phosphate) in the third day of incubation.
The efficacy of phosphate solubilization of ANsa120 was maximum in
the medium containing Ammonium sulphate (4.95 ppm of Phosphate) followed
by Urea (3.85 ppm of Phosphate), Potassium nitrate (3.70 ppm of Phosphate)
and Sodium nitrate (2.30 ppm of Phosphate) in the sixth day of incubation.
The efficacy of phosphate solubilization of ANsa120 was maximum in
the medium containing Ammonium sulphate (6.15 ppm of Phosphate) followed
by Urea (4.70 ppm of Phosphate), Potassium nitrate (4.65 ppm of
Phosphate)and Sodium nitrate (2.75 ppm of Phosphate) in the ninth day of
incubation.
Effect of Different Carbon Sources on the Efficacy of Phosphate
Solubilization of Sodium Azide Treated Penicillium sp (PEsa150).
The results of different carbon sources on the efficacy of phosphate
solubilization of sodium azide treated Penicillium sp (PEsa150) were presented
in Table 20

104
The efficacy of phosphate solubilization of PEsa150 was maximum in the
medium containing Glucose (3.80 ppm of Phosphate) followed by Sucrose (3.35
ppm of Phosphate), Lactose (3.15 ppm of Phosphate) and Mannitol (3.05 ppm of
Phosphate) in the third day of incubation.
The efficacy of phosphate solubilization of PEsa150 was maximum in the
medium containing Glucose (4.85 ppm of Phosphate) followed by Sucrose (3.90
ppm of Phosphate), Lactose (3.80 ppm of Phosphate) and Mannitol (3.75 ppm of
Phosphate) in the sixth day of incubation.
The efficacy of phosphate solubilization of PEsa150 was maximum in the
medium containing Glucose (5.95 ppm of Phosphate) followed by Sucrose (4.60
ppm of Phosphate), Lactose (4.35 ppm of Phosphate) and Mannitol (4.25 ppm of
Phosphate) in the ninth day of incubation.
Effect of Different Nitrogen Sources on the Efficacy of Phosphate
Solubilization of Sodium Azide Treated Penicillium sp (PEsa150)
The results of different nitrogen sources on the efficacy of phosphate
solubilization of sodium azide treated Penicillium sp (PEsa150) were presented
in Table 21
The efficacy of phosphate solubilization of PEsa150 was maximum in the
medium containing Ammonium sulphate (3.80 ppm of Phosphate) followed by
Urea (3.70 ppm of Phosphate), Potassium nitrate (3.35 ppm of Phosphate) and
Sodium nitrate (3.30 ppm of Phosphate) in the third day of incubation.
The efficacy of phosphate solubilization of PEsa150 was maximum in the
medium containing Ammonium sulphate (4.85 ppm of Phosphate) followed by

105
Urea (3.95 ppm of Phosphate), Potassium nitrate (3.90 ppm of Phosphate) and
Sodium nitrate (3.75 ppm of Phosphate) in the sixth day of incubation.
The efficacy of phosphate solubilization of PEsa150 was maximum in the
medium containing Ammonium sulphate (5.95 ppm of Phosphate) followed by
Urea (4.70 ppm of Phosphate), Potassium nitrate (4.60 ppm of Phosphate)and
Sodium nitrate (4.05 ppm of Phosphate) in the ninth day of incubation.
Figure- 1 showes the comparsion of efficacy of Phosphatate solublization
by wild strains and Figure-2 showes the comparsion of efficacy of Phosphatate
solublization by chemical treated fungal strains.
Phosphatase Activity
Efficacy of UV Treated Aspergillus niger on Phosphatase Activity
The results of Phosphatase activity of UV treated Aspergillus niger were
presented in Table 22.
The maximum efficacy of Phosphatase activity of UV treated Aspergillus
niger (ANuv60) on day three recorded (0.165 µmol min-1) followed by ANuv50
strain (0.134 µmol min-1), ANuv30 (0.121 µmol min-1) and ANuv40 (0.103
µmol min-1). The Phosphatase activity of Wild Aspergillus niger which was used
as a standard control produced 0.097µmol min-1 in the third day of incubation.
The maximum efficacy of Phosphatase activity of UV treated Aspergillus
niger (ANuv60) on day six recorded (0.275µmol min-1) followed by ANuv50
strain (0.260 µmol min-1), ANuv30 (0.223 µmol min-1) and ANuv40 (0.218
µmol min-1). The Phosphatase activity of Wild Aspergillus niger which was used
as a standard control produced 0.192 µmol min-1 in the sixth day of incubation.

106
The maximum efficacy of Phosphatase activity of UV treated Aspergillus
niger (ANuv60) on day nine recorded (0.424µmol min-1) followed by ANuv50
strain (0.399 µmol min-1), ANuv30 (0.345 µmol min-1) and ANuv40 (0.341
µmol min-1). The Phosphatase activity of Wild Aspergillus niger which was used
as a standard control produced 0.303 µmol min-1 in the ninth day of incubation.
The Phosphatase activity of ANuv60 strain increased 70.10% on third
day, 43.23% Phosphatase activity on sixth day and 39.93% Phosphatase
activity on ninth day of incubation using wild type as a standard (100%).
Efficacy of UV Treated Aspergillus fumigatus on Phosphatase Activity
The results of Phosphatase activity of UV treated Aspergillus fumigatus
were presented in Table 23.
The maximum efficacy of Phosphatase activity of UV treated Aspergillus
fumigatus (AFuv50) on day three recorded (0.195 µmol min-1) followed by
ANuv60 strain (0.173 µmol min-1), ANuv30 (0.171 µmol min-1) and ANuv40
(0.160 µmol min-1). The Phosphatase activity of Wild Aspergillus fumigatus
which was used as a standard control produced 0.156µmol min-1 in the third
day of incubation.
The maximum efficacy of Phosphatase activity of UV treated Aspergillus
fumigatus (AFuv50) on day six recorded (0.227 µmol min-1) followed by
AFuv60 strain (0.224 µmol min-1), AFuv40 (0.205 µmol min-1) and AFuv30
(0.203 µmol min-1). The Phosphatase activity of Wild Aspergillus fumigatus
which was used as a standard control produced 0.178µmol min-1 in the sixth
day of incubation.

107
The maximum efficacy of Phosphatase activity of UV treated Aspergillus
fumigatus (AFuv50) on day nine recorded (0.269 µmol min-1) followed by
ANuv60 strain (0.265 µmol min-1), AFuv40 (0.232 µmol min-1) and AFuv30
(0.221 µmol min-1). The Phosphatase activity of Wild Aspergillus fumigatus
which was used as a standard control produced 0.197µmol min-1 in the ninth
day of incubation.
The Phosphatase activity of AFuv50 strain increased 25.00% on third
day, 27.53% Phosphatase activity on sixth day and 36.55% Phosphatase activity
on ninth day of incubation using wild type as a standard (100%).
Efficacy of UV Treated Penicillium sp on Phosphatase Activity
The results of Phosphatase activity of UV treated Penicillium sp were
presented in Table 24.
The maximum efficacy of Phosphatase activity of UV treated Penicillium
sp (PEuv50) on day three recorded (0.209 µmol min-1) followed by PEuv40
strain (0.195 µmol min-1), PEuv30 (0.180 µmol min-1) and PEuv60 (0.178 µmol
min-1). The Phosphatase activity of Wild Penicillium sp which was used as a
standard control produced 0.169µmol min-1 in the third day of incubation.
The maximum efficacy of Phosphatase activity of UV treated Penicillium
sp (PEuv40) on day six recorded (0.245 µmol min-1) followed by PEuv50 strain
(0.237 µmol min-1), PEuv60 (0.234 µmol min-1) and PEuv30 (0.228 µmol min-
1). The Phosphatase activity of Wild Penicillium sp which was used as a
standard control produced 0.213 µmol min-1 in the sixth day of incubation.

108
The maximum efficacy of Phosphatase activity of UV treated Penicillium
sp (PEuv50) on day nine recorded (0.267 µmol min-1) followed by PEuv40
strain (0.264 µmol min-1), PEuv60 (0.257 µmol min-1) and PEuv30 (0.239 µmol
min-1). The Phosphatase activity of Wild Penicillium sp which was used as a
standard control produced 0.236 µmol min-1 in the ninth day of incubation.
The Phosphatase activity of PEuv50 strain increased 23.67% on third day,
11.27% Phosphatase activity on sixth day and 13.14% Phosphatase activity on
ninth day of incubation using wild type as a standard (100%).
Efficacy of UV Treated Pseudomonas sp on Phosphatase Activity
The results of Phosphatase activity of UV treated Pseudomonas sp were
presented in Table 25.
The maximum efficacy of Phosphatase activity of UV treated
Pseudomonas sp (PSuv3) on day three recorded (0.227 µmol min-1) followed by
PSuv2 strain (0.215 µmol min-1), PSuv4 (0.210 µmol min-1) and PSuv1 (0.205
µmol min-1). The Phosphatase activity of Wild Pseudomonas sp which was
used as a standard control produced 0.200µmol min-1 in the third day of
incubation.
The maximum efficacy of Phosphatase activity of UV treated
Pseudomonas sp (PSuv3) on day six recorded (0.233 µmol min-1) followed by
PSuv2 strain (0.227 µmol min-1), PSuv4 (0.226 µmol min-1) and PSuv1 (0.225
µmol min-1). The Phosphatase activity of Wild Pseudomonas sp which was
used as a standard control produced 0.234µmol min-1 in the six day of
incubation.

109
The maximum efficacy of Phosphatase activity of UV treated
Pseudomonas sp (PSuv4) on day nine recorded (0.286 µmol min-1) followed by
PSuv3 strain (0.275 µmol min-1), PSuv2 (0.236 µmol min-1) and PSuv1 (0.231
µmol min-1). The Phosphatase activity of Wild Pseudomonas sp which was
used as a standard control produced 0.287µmol min-1 in the ninth day of
incubation.
All mutants of Pseudomonas strains treated with UV on day six and nine
showed decreased Phosphatase activity activity when compared to wild type.
Efficacy of Sodium Azide Treated Aspergillus niger on Phosphatase Activity
The results of Phosphatase activity of sodium azide treated Aspergillus
niger were presented in Table 26.
The maximum efficacy of Phosphatase activity of sodium azide treated
Aspergillus niger (ANsa120) on day three recorded (0.184 µmol min-1) followed
by ANsa90 strain (0.152 µmol min-1), ANsa60 (0.110 µmol min-1) and ANsa30
(0.107 µmol min-1). The Phosphatase activity of Wild Aspergillus niger which
was used as a standard control produced 0.097µmol min-1 in the third day of
incubation.
The maximum efficacy of Phosphatase activity of sodium azide treated
Aspergillus niger (ANsa120) on day six recorded (0.340 µmol min-1) followed
by ANsa90 strain (0.268 µmol min-1), ANsa60 (0.234 µmol min-1) and ANsa30
(0.218 µmol min-1). The Phosphatase activity of Wild Aspergillus niger which
was used as a standard control produced 0.192µmol min-1 in the sixth day of
incubation.

110
The maximum efficacy of Phosphatase activity of sodium azide treated
Aspergillus niger (ANsa120) on day nine recorded (0.474 µmol min-1) followed
by ANsa90 strain (0.421 µmol min-1), ANsa60 (0.401 µmol min-1) and ANsa30
(0.335 µmol min-1). The Phosphatase activity of Wild Aspergillus niger which
was used as a standard control produced 0.303µmol min-1 in the ninth day of
incubation.
The Phosphatase activity of ANsa120 strain increased 89.69% on third
day, 77.08% Phosphatase activity on sixth day and 56.43% Phosphatase activity
on ninth day of incubation using wild type as a standard (100%).
Efficacy of Sodium Azide Treated Aspergillus fumigatus on Phosphatase
Activity
The results of Phosphatase activity of sodium azide treated Aspergillus
niger were presented in Table 27.
The maximum efficacy of Phosphatase activity of sodium azide treated
Aspergillus fumigatus (AFsa120) on day three recorded (0.232 µmol min-1)
followed by AFsa90 strain (0.185 µmol min-1), AFsa60 (0.174 µmol min-1) and
AFsa30 (0.165 µmol min-1). The Phosphatase activity of Wild Aspergillus
fumigatus which was used as a standard control produced 0.156µmol min-1 in
the third day of incubation.
The maximum efficacy of Phosphatase activity of sodium azide treated
Aspergillus fumigatus (AFsa120) on day six recorded (0.274 µmol min-1)
followed by AFsa90 strain (0.229 µmol min-1), AFsa60 (0.203 µmol min-1) and
AFsa30 (0.189 µmol min-1). The Phosphatase activity of Wild Aspergillus

111
fumigatus which was used as a standard control produced 0.178µmol min-1 in
the sixth day of incubation.
The maximum efficacy of Phosphatase activity of sodium azide treated
Aspergillus fumigatus (AFsa120) on day nine recorded (0.365 µmol min-1)
followed by AFsa90 strain (0.351 µmol min-1), AFsa60 (0.241 µmol min-1) and
AFsa30 (0.213 µmol min-1). The Phosphatase activity of Wild Aspergillus
fumigatus which was used as a standard control produced 0.197µmol min-1 in
the ninth day of incubation.
The Phosphatase activity of AFsa120 strain increased 48.71% on third
day, 53.93% Phosphatase activity on sixth day and 85.27% Phosphatase activity
on ninth day of incubation using wild type as a standard (100%).
Efficacy of Sodium Azide Treated Penicillium sp on Phosphatase Activity
The results of Phosphatase activity of sodium azide treated Penicillium sp
were presented in Table 28.
The maximum efficacy of Phosphatase activity of sodium azide treated
Penicillium sp (PEsa150) on day three recorded (0.210 µmol min-1) followed by
PEsa120 strain (0.200 µmol min-1), PEsa90 (0.191 µmol min-1) and PEsa60
(0.181 µmol min-1). The Phosphatase activity of Wild Penicillium sp which was
used as a standard control produced 0.169 µmol min-1 in the third day of
incubation.
The maximum efficacy of Phosphatase activity of sodium azide treated
Penicillium sp (PEsa150) on day six recorded (0.276 µmol min-1) followed by
PEsa120 strain (0.261 µmol min-1), PEsa90 (0.249 µmol min-1) and PEsa60

112
(0.233 µmol min-1). The Phosphatase activity of Wild Penicillium sp used as a
standard control produced 0.213 µmol min-1 in the sixth day of incubation.
The maximum efficacy of Phosphatase activity of sodium azide treated
Penicillium sp (PEsa150) on day nine recorded (0.420 µmol min-1) followed by
PEsa120 strain (0.400 µmol min-1), PEsa90 (0.349 µmol min-1) and PEsa60
(0.261 µmol min-1). The Phosphatase activity of Wild Penicillium sp which was
used as a standard control produced 0.236 µmol min-1 in the ninth day of
incubation.
The Phosphatase activity of PEsa150 strain increased 24.26% on third
day, 29.57% Phosphatase activity on sixth day and 77.96% Phosphatase activity
on ninth day of incubation using wild type as a standard (100%).
Efficacy of Sodium Azide Treated Pseudomonas sp on Phosphatase Activity
The results of Phosphatase activity of sodium azide treated Pseudomonas
sp were presented in Table 29.
The maximum efficacy of Phosphatase activity of sodium azide treated
Pseudomonas sp (PSsa120) on day three recorded (0.185 µmol min-1) followed
by PSsa90 strain (0.180 µmol min-1), PSsa60 (0.171 µmol min-1) and PSsa30
(0.169 µmol min-1). The Phosphatase activity of Wild Pseudomonas sp which
was used as a standard control produced 0.200µmol min-1 in the third day of
incubation.
The maximum efficacy of Phosphatase activity of sodium azide treated
Pseudomonas sp (PSsa120) on day six recorded (0.241 µmol min-1) followed by
PSsa90 strain (0.232 µmol min-1), PSsa60 (0.220 µmol min-1) and PSsa30

113
(0.199 µmol min-1). The Phosphatase activity of Wild Pseudomonas sp which
was used as a standard control produced 0.234µmol min-1 in the six day of
incubation.
The maximum efficacy of Phosphatase activity of sodium azide treated
Pseudomonas sp (PSsa120) on day nine recorded (0.275 µmol min-1) followed
by PSsa90 strain (0.260 µmol min-1), PSsa60 (0.256 µmol min-1) and PSsa30
(0.224 µmol min-1). The Phosphatase activity of Wild Pseudomonas sp which
was used as a standard control produced 0.287µmol min-1 in the ninth day of
incubation.
All mutants of Pseudomonas strains treated with sodium azide on day
three, six and nine showed decreased Phosphatase activity activity when
compared to wild type.
Efficacy of Ethyl Methane Sulphonate Treated Aspergillus niger on
Phosphatase Activity
The results of Phosphatase activity of Ethyl Methane Sulphonate treated
Aspergillus niger were presented in Table 30.
The maximum efficacy of Phosphatase activity of Ethyl Methane
Sulphonate treated Aspergillus niger (ANems120) on day three recorded (0.131
µmol min-1) followed by ANems90 strain (0.125 µmol min-1), ANems60 (0.104
µmol min-1) and ANems30 (0.099 µmol min-1). The Phosphatase activity of
Wild Aspergillus niger which was used as a standard control produced
0.097µmol min-1 in the third day of incubation.
The maximum efficacy of Phosphatase activity of Ethyl Methane
Sulphonate treated Aspergillus niger (ANems120) on day six recorded (0.273

114
µmol min-1) followed by ANems90 strain (0.253 µmol min-1), ANems60 (0.222
µmol min-1) and ANems30 (0.210 µmol min-1). The Phosphatase activity of
Wild Aspergillus niger which was used as a standard control produced
0.192µmol min-1 in the sixth day of incubation.
The maximum efficacy of Phosphatase activity of Ethyl Methane
Sulphonate treated Aspergillus niger (ANems120) on day nine recorded (0.452
µmol min-1) followed by ANems90 strain (0.412 µmol min-1), ANems60 (0.365
µmol min-1) and ANems30 (0.316 µmol min-1). The Phosphatase activity of
Wild Aspergillus niger which was used as a standard control produced 0.303
µmol min-1 in the ninth day of incubation.
The Phosphatase activity of ANems120 strain increased 35.05% on third
day, 42.18% Phosphatase activity on sixth day and 49.17% Phosphatase activity
on ninth day of incubation using wild type as a standard (100%).
Efficacy of Ethyl Methane Sulphonate Treated Aspergillus fumigatus on
Phosphatase Activity
The results of Phosphatase activity of Ethyl Methane Sulphonate treated
Aspergillus fumigatus were presented in Table 31.
The maximum efficacy of Phosphatase activity of Ethyl Methane
Sulphonate treated Aspergillus fumigatus (AFems120) on day three recorded
(0.219 µmol min-1) followed by AFems90 strain (0.206 µmol min-1), AFems60
(0.189 µmol min-1) and AFems30 (0.175 µmol min-1). The Phosphatase activity
of Wild Aspergillus fumigatus which was used as a standard control produced
0.156 µmol min-1 in the third day of incubation.

115
The maximum efficacy of Phosphatase activity of Ethyl Methane
Sulphonate treated Aspergillus fumigatus (AFems120) on day six recorded
(0.258 µmol min-1) followed by AFems90 strain (0.236 µmol min-1), AFems60
(0.212 µmol min-1) and AFems30 (0.197 µmol min-1). The Phosphatase activity
of Wild Aspergillus fumigatus which was used as a standard control produced
0.178 µmol min-1 in the sixth day of incubation.
The maximum efficacy of Phosphatase activity of Ethyl Methane
Sulphonate treated Aspergillus fumigatus (AFems120) on day nine recorded
(0.318 µmol min-1) followed by AFems90 strain (0.261 µmol min-1), AFems60
(0.231 µmol min-1) and AFems30 (0.214 µmol min-1). The Phosphatase activity
of Wild Aspergillus fumigatus which was used as a standard control produced
0.197 µmol min-1 in the ninth day of incubation.
The Phosphatase activity of AFems120 strain increased 40.38% on third
day, 44.94% Phosphatase activity on sixth day and 61.42% Phosphatase activity
on ninth day of incubation using wild type as a standard (100%).
Efficacy of Ethyl Methane Sulphonate Treated Penicillium sp on
Phosphatase Activity
The results of Phosphatase activity of Ethyl Methane Sulphonate treated
Penicillium sp were presented in Table 32.
The maximum efficacy of Phosphatase activity of Ethyl Methane
Sulphonate treated Penicillium sp (PEems150) on day three recorded (0.278
µmol min-1) followed by PEems120 strain (0.245 µmol min-1), PEems90 (0.227
µmol min-1) and PEems60 (0.197 µmol min-1). The Phosphatase activity of Wild

116
Penicillium sp which was used as a standard control produced 0.169 µmol min-1
in the third day of incubation.
The maximum efficacy of Phosphatase activity of Ethyl Methane
Sulphonate treated Penicillium sp (PEems150) on day six recorded (0.381 µmol
min-1) followed by PEems120 strain (0.324 µmol min-1), PEems90 (0.325 µmol
min-1) and PEems60 (0.275 µmol min-1). The Phosphatase activity of Wild
Penicillium sp which was used as a standard control produced 0.213 µmol min-1
in the sixth day of incubation.
The maximum efficacy of Phosphatase activity of Ethyl Methane
Sulphonate treated Penicillium sp (PEems150) on day nine recorded (0.425
µmol min-1) followed by PEems120 strain (0.371 µmol min-1), PEems60 (0.370
µmol min-1) and PEems90 (0.369 µmol min-1). The Phosphatase activity of Wild
Penicillium sp which was used as a standard control produced 0.236 µmol min-1
in the ninth day of incubation.
The Phosphatase activity of PEems150 strain increased 64.49% on third
day, 78.87% Phosphatase activity on sixth day and 80.00% Phosphatase activity
on ninth day of incubation using wild type as a standard (100%).
Efficacy of Ethyl Methane Sulphonate Treated Pseudomonas sp on
Phosphatase Activity
The results of Phosphatase activity of Ethyl Methane Sulphonate treated
Pseudomonas sp were presented in Table 33.
The maximum efficacy of Phosphatase activity of Ethyl Methane
Sulphonate treated Pseudomonas sp (PSsa150) on day three recorded (0.113
µmol min-1) followed by PSsa120 strain (0.100 µmol min-1), PSsa90 (0.095

117
µmol min-1) and PSsa60 (0.091 µmol min-1). The Phosphatase activity of Wild
Pseudomonas sp which was used as a standard control produced 0.200µmol
min-1 in the third day of incubation.
The maximum efficacy of Phosphatase activity of Ethyl Methane
Sulphonate treated Pseudomonas sp (PSsa150) on day six recorded (0.175 µmol
min-1) followed by PSsa120 strain (0.167 µmol min-1), PSsa90 (0.161 µmol
min-1) and PSsa60 (0.150 µmol min-1). The Phosphatase activity of Wild
Pseudomonas sp which was used as a standard control produced 0.234µmol
min-1 in the six day of incubation.
The maximum efficacy of Phosphatase activity of Ethyl Methane
Sulphonate treated Pseudomonas sp (PSsa150) on day nine recorded (0.229
µmol min-1) followed by PSsa120 strain (0.220 µmol min-1), PSsa90 (0.191
µmol min-1) and PSsa60 (0.180 µmol min-1). The Phosphatase activity of Wild
Pseudomonas sp which was used as a standard control produced 0.287µmol
min-1 in the nine day of incubation.
All mutants of Pseudomonas strains treated with Ethyl Methane
Sulphonate on day three, six and nine showed decreased Phosphatase activity
activity when compared to wild type.
Effect of Different Carbon and Nitrogen Sources on the Efficacy of Acid
Phosphatase Production.
The superior mutated cultures of Phosphatase in Pikovskaya broth were
identified as Ethyl Methane Sulphonate treated Aspergillus niger (ANems120),
Sodium Azide Treated Aspergillus niger (ANsa120) and Sodium Azide Treated

118
Penicillium sp (PEsa150) and their efficacy of Phosphatase on different Carbon
and Nitrogen Sources were studied.
Effect of Different Carbon Sources on the Efficacy of Phosphatase Activity
of Ethyl Methane Sulphonate Treated (ANems120).
The results of different carbon sources on the efficacy of Phosphatase
activity of Ethyl Methane Sulphonate treated Aspergillus niger (ANems120)
were presented in Table 34.
The efficacy of Phosphatase activity of ANems120 was maximum in the
medium containing Glucose (0.131 µmol min-1) followed by Sucrose (0.107
µmol min-1), Lactose (0.094 µmol min-1) and Mannitol (0.083 µmol min-1) in the
third day of incubation.
The efficacy of Phosphatase activity of ANems120 was maximum in the
medium containing Glucose (0.273 µmol min-1) followed by Sucrose (0.206
µmol min-1), Lactose (0.182 µmol min-1) and Mannitol (0.167 µmol min-1) in the
sixth day of incubation.
The efficacy of Phosphatase activity of ANems120 was maximum in the
medium containing Glucose (0.452 µmol min-1) followed by Sucrose (0.279
µmol min-1), Lactose (0.257 µmol min-1) and Mannitol (0.229 µmol min-1) in the
ninth day of incubation.

119
Effect of Different Nitrogen Sources on the Efficacy of Phosphatase Activity
of Ethyl Methane Sulphonate Treated Aspergillus niger (ANems120).
The results of different nitrogen sources on the efficacy of Phosphatase
activity of Ethyl Methane Sulphonate treated Aspergillus niger (ANems120)
were presented in Table 35.
The efficacy of Phosphatase activity of ANems120 was maximum in the
medium containing Ammonium sulphate (0.131 µmol min-1) followed by Urea
(0.109 µmol min-1), Potassium nitrate (0.102 µmol min-1) and Sodium nitrate
(0.074 µmol min-1) in the third day of incubation.
The efficacy of Phosphatase activity of ANems120 was maximum in the
medium containing Ammonium sulphate (0.273 µmol min-1) followed by Urea
(0.203 µmol min-1), Potassium nitrate (0.200 µmol min-1) and Sodium nitrate
(0.122 µmol min-1) in the sixth day of incubation.
The efficacy of Phosphatase activity of ANems120 was maximum in the
medium containing Ammonium sulphate (0.452 µmol min-1) followed by Urea
(0.275 µmol min-1), Potassium nitrate (0.271 µmol min-1) and Sodium nitrate
(0.173 µmol min-1) in the ninth day of incubation.
Effect of Different Carbon Sources on the Efficacy of Phosphatase Activity
of Sodium Azide Treated Aspergillus niger (ANsa120).
The results of different carbon sources on the efficacy of Phosphatase
activity of sodium azide treated Aspergillus niger (ANsa120) were presented in
Table 36.

120
The efficacy of Phosphatase activity of ANsa120 was maximum in the
medium containing Sucrose (0.186 µmol min-1) followed by Glucose (0.184
µmol min-1), Lactose (0.165 µmol min-1) and Mannitol (0.076 µmol min-1) in the
third day of incubation.
The efficacy of Phosphatase activity of ANsa120 was maximum in the
medium containing Glucose (0.340 µmol min-1) followed by Sucrose (0.248
µmol min-1), Lactose (0.187 µmol min-1) and Mannitol (0.094 µmol min-1) in the
sixth day of incubation.
The efficacy of Phosphatase activity of ANsa120 was maximum in the
medium containing Glucose (0.474 µmol min-1) followed by Sucrose (0.330
µmol min-1), Lactose (0.213 µmol min-1) and Mannitol (0.134 µmol min-1) in the
ninth day of incubation.
Effect of Different Nitrogen Sources on the Efficacy of Phosphatase Activity
of Sodium Azide Treated Aspergillus niger (ANsa120).
The results of different nitrogen sources on the efficacy of Phosphatase
activity of sodium azide treated Aspergillus niger (ANsa120) were presented in
Table 37.
The efficacy of Phosphatase activity of ANsa120 was maximum in the
medium containing Ammonium sulphate (0.184 µmol min-1) followed by Urea
(0.112 µmol min-1), Potassium nitrate (0.108 µmol min-1) and Sodium nitrate
(0.008 µmol min-1) in the third day of incubation.
The efficacy of Phosphatase activity of ANsa120 was maximum in the
medium containing Ammonium sulphate (0.340 µmol min-1) followed by Urea

121
(0.216 µmol min-1), Potassium nitrate (0.203 µmol min-1) and Sodium nitrate
(0.103 µmol min-1) in the sixth day of incubation.
The efficacy of Phosphatase activity of ANsa120 was maximum in the
medium containing Ammonium sulphate (0.474 µmol min-1) followed by Urea
(0.273 µmol min-1), Potassium nitrate (0.254 µmol min-1) and Sodium nitrate
(0.125 µmol min-1) in the ninth day of incubation.
Effect of Different Carbon Sources on the Efficacy of Phosphatase Activity
of Sodium Azide Treated Penicillium sp (PEsa150).
The results of different carbon sources on the efficacy of Phosphatase
activity of sodium azide treated Penicillium sp (PEsa150) were presented in
Table 38.
The efficacy of Phosphatase activity of PEsa150 was maximum in the
medium containing Glucose (0.210 µmol min-1) followed by Sucrose (0.171
µmol min-1), Lactose (0.168 µmol min-1) and Mannitol (0.162 µmol min-1) in the
third day of incubation.
The efficacy of Phosphatase activity of PEsa150 was maximum in the
medium containing Sucrose (0.288 µmol min-1) followed by Glucose (0.276
µmol min-1), Lactose (0.214 µmol min-1) and Mannitol (0.204 µmol min-1) in the
sixth day of incubation.
The efficacy of Phosphatase activity of PEsa150 was maximum in the
medium containing Glucose (0.420 µmol min-1) followed by Sucrose (0.342
µmol min-1), Mannitol (0.261 µmol min-1) and Lactose (0.253 µmol min-1) in the
ninth day of incubation.

122
Effect of Different Nitrogen Sources on the Efficacy of Phosphatase Activity
of Sodium Azide Treated Penicillium sp (PEsa150).
The results of different nitrogen sources on the efficacy of Phosphatase
activity of sodium azide treated Penicillium sp (PEsa150) were presented in
Table 39.
The efficacy of Phosphatase activity of PEsa150 was maximum in the
medium containing Ammonium sulphate (0.210 µmol min-1) followed by Urea
(0.205 µmol min-1), Potassium nitrate (0.165 µmol min-1) and Sodium nitrate
(0.163 µmol min-1) in the third day of incubation.
The efficacy of Phosphatase activity of PEsa150 was maximum in the
medium containing Ammonium sulphate (0.276 µmol min-1) followed by Urea
(0.223 µmol min-1), Potassium nitrate (0.218 µmol min-1) and Sodium nitrate
(0.209 µmol min-1) in the sixth day of incubation.
The efficacy of Phosphatase activity of PEsa150 was maximum in the
medium containing Ammonium sulphate (0.420 µmol min-1) followed by Urea
(0.270 µmol min-1), Potassium nitrate (0.261 µmol min-1) and Sodium nitrate
(0.229 µmol min-1) in the ninth day of incubation.
Plate-4 shows the Phosphate solubilization and Phosphatase production
by fungal cultures in Pikovskaya broth.
Figure- 3 showes the comparsion of efficacy of Phosphatase activity by
wild strains and Figure-4 showes the comparsion of efficacy of Phosphatase
activity by chemical treated fungal strains.

123
Lipase Activity
Efficacy of UV Treated Aspergillus niger on Lipase Activity
The results of Lipase activity of UV treated Aspergillus niger were
presented in Table 40.
The maximum efficacy of Lipase activity of UV treated Aspergillus niger
(ANuv50) grown in Sucrose medium recorded (2.15 unit g-1 of substrate)
followed by ANuv40 strain (1.95 unit g-1 of substrate), ANuv60 (1.89 unit g-1 of
substrate) and ANuv30 (1.86 unit g-1 of substrate). The Lipase activity of wild
Aspergillus niger which was used as a standard control produced 1.62 unit g-1 of
substrate after 96 hrs incubation.
The maximum efficacy of Lipase activity of UV treated Aspergillus niger
(ANuv50) grown in olive oil medium recorded (0.90 unit g-1 of substrate)
followed by ANuv40 strain (0.62 unit g-1 of substrate), ANuv60 (0.60 unit g-1 of
substrate) and ANuv30 (0.49 unit g-1 of substrate). The Lipase activity of wild
Aspergillus niger which was used as a standard control produced 0.47 unit g-1 of
substrate after 96 hrs incubation.
The Lipase activity of ANuv50 strain increased 32.72% followed by
ANuv40 (20.37%), ANuv60 (16.66%) and ANuv30 (14.81%) after 96 hrs
incubation in Sucrose medium using wild type as a standard (100%). The
Lipase activity of ANuv50 strain increased 91.49% followed by ANuv40
(31.91%), ANuv60 (27.66%) and ANuv30 (4.26%) after 96 hrs incubation in
olive medium using wild type as a standard (100%).

124
Efficacy of UV Treated Aspergillus fumigatus on Lipase Activity
The results of Lipase activity of UV treated Aspergillus fumigatus were
presented in Table 41.
The maximum efficacy of Lipase activity of UV treated Aspergillus
fumigatus (AFuv50) grown in Sucrose medium recorded (2.09 unit g-1 of
substrate) followed by AFuv60 strain (1.75 unit g-1 of substrate), AFuv40 (1.39
unit g-1 of substrate) and AFuv30 (1.22 unit g-1 of substrate). The Lipase activity
of wild Aspergillus fumigatus which was used as a standard control produced
1.01 unit g-1 of substrate after 96 hrs incubation.
The maximum efficacy of Lipase activity of UV treated Aspergillus
fumigatus (AFuv50) grown in olive oil medium recorded (0.60 unit g-1 of
substrate) followed by AFuv60 strain (0.56 unit g-1 of substrate), AFuv40 (0.38
unit g-1 of substrate) and AFuv30 (0.33 unit g-1 of substrate). The Lipase activity
of wild Aspergillus fumigatus which was used as a standard control produced
0.27 unit g-1 of substrate after 96 hrs incubation.
The Lipase activity of AFuv50 strain increased 106.93% followed by
AFuv60 (73.26%), AFuv40 (37.62%) and AFuv30 (20.79%) after 96 hrs
incubation in Sucrose medium using wild type as a standard (100%). The
Lipase activity of AFuv50 strain increased 122.22% followed by AFuv60
(107.40%), AFuv40 (40.74%) and AFuv30 (22.22%) after 96 hrs incubation in
olive medium using wild type as a standard (100%).
Efficacy of UV Treated Penicillium sp on Lipase Activity
The results of Lipase activity of UV treated Penicillium sp were presented
in Table 42.

125
The maximum efficacy of Lipase activity of UV treated Penicillium sp
(PEuv60) grown in Sucrose medium recorded (1.69 unit g-1 of substrate)
followed by PEuv50 strain (1.62 unit g-1 of substrate), PEuv40 (1.43 unit g-1 of
substrate) and PEuv30 (1.28 unit g-1 of substrate). The Lipase activity of wild
Penicillium sp which was used as a standard control produced 1.20 unit g-1 of
substrate after 96 hrs incubation.
The maximum efficacy of Lipase activity of UV treated Penicillium sp
(PEuv60) grown in olive oil medium recorded (0.85 unit g-1 of substrate)
followed by PEuv40 strain (0.73 unit g-1 of substrate), PEuv50 (0.69 unit g-1 of
substrate) and PEuv30 (0.48 unit g-1 of substrate). The Lipase activity of wild
Penicillium sp which was used as a standard control produced 0.46 unit g-1 of
substrate after 96 hrs incubation.
The Lipase activity of PEuv60 strain increased 40.83% followed by
PEuv50 (35.00%), PEuv40 (19.17%) and PEuv30 (6.67%) after 96 hrs
incubation in Sucrose medium using wild type as a standard (100%). The
Lipase activity of PEuv60 strain increased 84.78% followed by PEuv40
(50.00%), PEuv50 (58.7%) and PEuv30 (4.35%) after 96 hrs incubation in olive
medium using wild type as a standard (100%).
Efficacy of UV Treated Pseudomonas sp on Lipase Activity
The results of Lipase activity of UV treated Pseudomonas sp were
presented in Table 43.
The maximum efficacy of Lipase activity of UV treated Pseudomonas sp
(PSuv3) grown in Sucrose medium recorded (2.49 unit g-1 of substrate) followed

126
by PSuv4 strain (2.29 unit g-1 of substrate), PSuv1 (2.22 unit g-1 of substrate)
and PSuv2 (2.19 unit g-1 of substrate). The Lipase activity of wild Pseudomonas
sp which was used as a standard control produced 2.10 unit g-1 of substrate after
96 hrs incubation.
The maximum efficacy of Lipase activity of UV treated Pseudomonas sp
(PSuv3) grown in olive oil medium recorded (0.38 unit g-1 of substrate) followed
by PSuv4 strain (0.34 unit g-1 of substrate), PSuv2 (0.33 unit g-1 of substrate)
and PSuv1 (0.23 unit g-1 of substrate). The Lipase activity of wild Pseudomonas
sp which was used as a standard control produced 0.17 unit g-1 of substrate after
96 hrs incubation.
The Lipase activity of PSuv3 strain increased 18.57% followed by
PSuv4 (9.04%), PSuv1 (5.71%) and PSuv2 (4.28%) after 96 hrs incubation in
Sucrose medium using wild type as a standard (100%). The Lipase activity of
PSuv3 strain increased 123.53% followed by PSuv4 (100.00%), PSuv2 (94.12%)
and PSuv1 (35.29%) after 96 hrs incubation in olive medium using wild type as
a standard (100%).
Efficacy of Sodium Azide Treated Aspergillus niger on Lipase Activity
The results of Lipase activity of sodium azide treated Aspergillus niger
were presented in Table 44.
The maximum efficacy of Lipase activity of sodium azide treated
Aspergillus niger (ANsa90) grown in Sucrose medium recorded (2.61 unit g-1 of
substrate) followed by ANsa60 strain (2.09 unit g-1 of substrate), ANsa30 (1.90
unit g-1 of substrate) and ANsa120 (1.85 unit g-1 of substrate). The Lipase activity

127
of wild Aspergillus niger which was used as a standard control produced 1.62
unit g-1 of substrate after 96 hrs incubation.
The maximum efficacy of Lipase activity of sodium azide treated
Aspergillus niger (ANsa90) grown in olive oil medium recorded (0.98 unit g-1 of
substrate) followed by ANsa60 strain (0.68 unit g-1 of substrate), ANsa120 (0.63
unit g-1 of substrate) and ANsa30 (0.51 unit g-1 of substrate). The Lipase activity
of wild Aspergillus niger which was used as a standard control produced 0.47
unit g-1 of substrate after 96 hrs incubation.
The Lipase activity of ANsa90 strain increased 61.11% followed by
ANsa60 (29.01%), ANsa30 (17.28%) and ANsa120 (14.19%) after 96 hrs
incubation in Sucrose medium using wild type as a standard (100%). The
Lipase activity of ANsa90 strain increased 108.61% followed by ANsa60
(44.68%), ANsa120 (34.04%) and ANsa30 (8.51%) after 96 hrs incubation in
olive medium using wild type as a standard (100%).
Efficacy of Sodium Azide Treated Aspergillus fumigatus on Lipase Activity
The results of Lipase activity of sodium azide treated Aspergillus
fumigatus were presented in Table 45.
The maximum efficacy of Lipase activity of sodium azide treated
Aspergillus fumigatus (AFsa120) grown in Sucrose medium recorded (1.60 unit
g-1 of substrate) followed by AFsa30 strain (1.45 unit g-1 of substrate), AFsa60
(1.25 unit g-1 of substrate) and AFsa90 (1.22 unit g-1 of substrate). The Lipase
activity of wild Aspergillus fumigatus which was used as a standard control
produced 1.01 unit g-1 of substrate after 96 hrs incubation.

128
The maximum efficacy of Lipase activity of sodium azide treated
Aspergillus fumigatus (AFsa120) grown in olive oil medium recorded (0.44 unit
g-1 of substrate) followed by AFsa30 strain (0.41 unit g-1 of substrate), AFsa60
(0.38 unit g-1 of substrate) and AFsa90 (0.30 unit g-1 of substrate). The Lipase
activity of wild Aspergillus fumigatus which was used as a standard control
produced 0.27 unit g-1 of substrate after 96 hrs incubation.
The Lipase activity of AFsa120 strain increased 58.42% followed by
AFsa30 (43.56%), AFsa60 (23.76%) and AFsa90 (20.79%) after 96 hrs
incubation in Sucrose medium using wild type as a standard (100%). The
Lipase activity of AFsa120 strain increased 62.96% followed by AFsa30
(51.85%), AFsa60 (40.74%) and AFsa90 (11.11%) after 96 hrs incubation in
olive medium using wild type as a standard (100%).
Efficacy of Sodium Azide Treated Penicillium sp on Lipase Activity
The results of Lipase activity of sodium azide treated Penicillium sp were
presented in Table 46.
The maximum efficacy of Lipase activity of sodium azide treated
Penicillium sp (PEsa120) grown in Sucrose medium recorded (2.01 unit g-1 of
substrate) followed by PEsa90 strain (1.76 unit g-1 of substrate), PEsa30 (1.56
unit g-1 of substrate) and PEsa60 (1.43 unit g-1 of substrate). The Lipase activity
of wild Penicillium sp which was used as a standard control produced 1.20 unit
g-1 of substrate after 96 hrs incubation.
The maximum efficacy of Lipase activity of sodium azide treated
Penicillium sp (PEsa120) grown in olive oil medium recorded (0.99 unit g-1 of
substrate) followed by PEsa90 strain (0.73 unit g-1 of substrate), PEsa60 (0.69

129
unit g-1 of substrate) and PEsa30 (0.48 unit g-1 of substrate). The Lipase activity
of wild Penicillium sp which was used as a standard control produced 0.46 unit
g-1 of substrate after 96 hrs incubation.
The Lipase activity of PEsa120 strain increased 67.50% followed by
PEsa90 (46.67%), PEsa30 (30.00%) and PEsa60 (19.17%) after 96 hrs
incubation in Sucrose medium using wild type as a standard (100%). The
Lipase activity of PEsa120 strain increased 115.22% followed by PEsa90
(58.70%), PEsa60 (50.00%) and PEsa30 (4.35%) after 96 hrs incubation in olive
medium using wild type as a standard (100%).
Efficacy of Sodium Azide Treated Pseudomonas sp on Lipase Activity
The results of Lipase activity of sodium azide treated Pseudomonas sp
were presented in Table 47.
The maximum efficacy of Lipase activity of sodium azide treated
Pseudomonas sp (PSsa90) grown in Sucrose medium recorded (3.23 unit g-1 of
substrate) followed by PSsa120 strain (3.04 unit g-1 of substrate), PSsa60 (2.93
unit g-1 of substrate) and PSsa30 (2.28 unit g-1 of substrate). The Lipase activity
of wild Pseudomonas sp which was used as a standard control produced 2.10
unit g-1 of substrate after 96 hrs incubation.
The maximum efficacy of Lipase activity of sodium azide treated
Pseudomonas sp (PSsa90) grown in olive oil medium recorded (0.39 unit g-1 of
substrate) followed by PSsa120 strain (0.39 unit g-1 of substrate), PSsa60 (0.35
unit g-1 of substrate) and PSsa30 (0.28 unit g-1 of substrate). The Lipase activity
of wild Pseudomonas sp which was used as a standard control produced 0.17
unit g-1 of substrate after 96 hrs incubation.

130
The Lipase activity of PSsa90 strain increased 53.80% followed by
PSsa120 (44.76%), PSsa60 (39.52%) and PSsa30 (8.57%) after 96 hrs
incubation in Sucrose medium using wild type as a standard (100%). The
Lipase activity of PSsa90 strain increased 129.41% followed by PSsa120
(129.41%), PSsa60 (105.88%) and PSsa30 (64.71%) after 96 hrs incubation in
olive medium using wild type as a standard (100%).
Efficacy of Ethyl Methane Sulphonate Treated Aspergillus niger on Lipase
Activity
The results of Lipase activity of Ethyl Methane Sulphonate treated
Aspergillus niger were presented in Table 48.
The maximum efficacy of Lipase activity of Ethyl Methane Sulphonate
treated Aspergillus niger (ANems150) grown in Sucrose medium recorded (3.85
unit g-1 of substrate) followed by ANems120 strain (3.10 unit g-1 of substrate),
ANems90 (2.09 unit g-1 of substrate) and ANems60 (1.90 unit g-1 of substrate).
The Lipase activity of wild Aspergillus niger which was used as a standard
control produced 1.62 unit g-1 of substrate after 96 hrs incubation.
The maximum efficacy of Lipase activity of Ethyl Methane Sulphonate
treated Aspergillus niger (ANems150) grown in olive oil medium recorded (0.94
unit g-1 of substrate) followed by ANems120 strain (0.87 unit g-1 of substrate),
ANems90 (0.77 unit g-1 of substrate) and ANems60 (0.62 unit g-1 of substrate).
The Lipase activity of wild Aspergillus niger which was used as a standard
control produced 0.47 unit g-1 of substrate after 96 hrs incubation.
The Lipase activity of ANems150 strain increased 137.65% followed by
ANems120 (91.35%), ANems90 (29.01%) and ANems60 (17.28%) after 96 hrs

131
incubation in Sucrose medium using wild type as a standard (100%). The
Lipase activity of ANems150 strain increased (100.00%) followed by
ANems120 (85.11%), ANems90 (63.83%) and ANems60 (31.91%) after 96 hrs
incubation in olive medium using wild type as a standard (100%).
Efficacy of Ethyl Methane Sulphonate Treated Aspergillus fumigatus on
Lipase Activity
The results of Lipase activity of Ethyl Methane Sulphonate treated
Aspergillus fumigatus were presented in Table 49.
The maximum efficacy of Lipase activity of Ethyl Methane Sulphonate
treated Aspergillus fumigatus (AFems120) grown in Sucrose medium recorded
(2.76 unit g-1 of substrate) followed by AFems150 strain (2.60 unit g-1 of
substrate), AFems90 (2.26 unit g-1 of substrate) and AFems60 (1.85 unit g-1 of
substrate). The Lipase activity of wild Aspergillus fumigatus which was used as
a standard control produced 1.01 unit g-1 of substrate after 96 hrs incubation.
The maximum efficacy of Lipase activity of Ethyl Methane Sulphonate
treated Aspergillus fumigatus (AFems120) grown in olive oil medium recorded
(0.45 unit g-1 of substrate) followed by AFems150 strain (0.41 unit g-1 of
substrate), AFems90 (0.40 unit g-1 of substrate) and AFems60 (0.33 unit g-1 of
substrate). The Lipase activity of wild Aspergillus fumigatus which was used as
a standard control produced 0.27 unit g-1 of substrate after 96 hrs incubation.
The Lipase activity of AFems120 strain increased 173.26% followed by
AFems150 (157.43%), AFems90 (123.76%) and AFems60 (83.17%) after 96 hrs
incubation in Sucrose medium using wild type as a standard (100%). The
Lipase activity of AFems120 strain increased 66.67% followed by AFems150

132
(51.85%), AFems90 (48.15%) and AFems60 (22.22%) after 96 hrs incubation in
olive medium using wild type as a standard (100%).
Efficacy of Ethyl Methane Sulphonate Treated Penicillium sp on Lipase
Activity
The results of Lipase activity of Ethyl Methane Sulphonate treated
Penicillium sp were presented in Table 50.
The maximum efficacy of Lipase activity of Ethyl Methane Sulphonate
treated Penicillium sp (PEems150) grown in Sucrose medium recorded (1.97
unit g-1 of substrate) followed by PEems120 strain (1.89 unit g-1 of substrate),
PEems90 (1.79 unit g-1 of substrate) and PEems60 (1.65 unit g-1 of substrate).
The Lipase activity of wild Penicillium sp which was used as a standard control
produced 1.20 unit g-1 of substrate after 96 hrs incubation.
The maximum efficacy of Lipase activity of Ethyl Methane Sulphonate
treated Penicillium sp (PEems150) grown in olive oil medium recorded (0.77
unit g-1 of substrate) followed by PEems120 strain (0.63 unit g-1 of substrate),
PEems90 (0.58 unit g-1 of substrate) and PEems60 (0.50 unit g-1 of substrate).
The Lipase activity of wild Penicillium sp used as a standard control produced
0.46 unit g-1 of substrate after 96 hrs incubation.
The Lipase activity of PEems150 strain increased 64.17% followed by
PEems120 (57.50%), PEems90 (49.17%) and PEems60 (36.67%) after 96 hrs
incubation in Sucrose medium using wild type as a standard (100%). The
Lipase activity of PEems150 strain increased 67.39% followed by PEems120
(36.96%), PEems90 (26.09%) and PEems60 (8.70%) after 96 hrs incubation in
olive medium using wild type as a standard (100%).

133
Efficacy of Ethyl Methane Sulphonate Treated Pseudomonas sp on Lipase
Activity
The results of Lipase activity of Ethyl Methane Sulphonate treated
Pseudomonas sp were presented in Table 51.
The maximum efficacy of Lipase activity of Ethyl Methane Sulphonate
treated Pseudomonas sp (PSems120) grown in Sucrose medium recorded (4.45
unit g-1 of substrate) followed by PSems150 strain (4.12 unit g-1 of substrate),
PSems90 (3.53 unit g-1 of substrate) and PSems60 (2.42 unit g-1 of substrate).
The Lipase activity of wild Pseudomonas sp which was used as a standard
control produced 2.21 unit g-1 of substrate after 96 hrs incubation.
The maximum efficacy of Lipase activity of Ethyl Methane Sulphonate
treated Pseudomonas sp (PSems120) grown in olive oil medium recorded (0.61
unit g-1 of substrate) followed by PSems150 strain (0.52 unit g-1 of substrate),
PSems90 (0.47 unit g-1 of substrate) and PSems60 (0.34 unit g-1 of substrate).
The Lipase activity of wild Pseudomonas sp which was used as a standard
control produced 0.22 unit g-1 of substrate after 96 hrs incubation.
The Lipase activity of PSems120 strain increased 101.36% followed by
PSems150 (86.43%), PSems90 (59.72%) and PSems60 (9.50%) after 96 hrs
incubation in Sucrose medium using wild type as a standard (100%). The
Lipase activity of PSems120 strain increased 177.27% followed by PSems150
(136.36%), PSems90 (113.64%) and PSems60 (54.55%) after 96 hrs incubation
in olive medium using wild type as a standard (100%).

134
Effect of Different Carbon and Nitrogen Sources on the Efficacy of Lipase
Activity.
The superior mutated cultures of Lipase in Czapek’s dox broth were
identified as Ethyl Methane Sulphonate treated Aspergillus niger (ANems120)
and Aspergillus fumigatus (AFems120), Aspergillus niger (ANems150),
Pseudomonas sp. (PSems150) and (PSems120) and their efficacy of lipase
activity were studied by using different Carbon and Nitrogen sources.
Effect of Different Carbon Sources on the Efficacy of Lipase Activity.
The results of different carbon sources on the efficacy of lipase activity of
Ethyl Methane Sulphonate treated Aspergillus niger (ANems120) and
Aspergillus fumigatus (AFems120), Aspergillus niger (ANems150),
Pseudomonas sp. (PSems150) and (PSems120) were presented in Tables 52 &
53.
The efficacy of lipase activity of Aspergillus niger (ANems120) was
maximum in the medium containing Sucrose (3.10 Unit/g of substrate) followed
by Glucose (2.94 Unit/g of substrate), Mannitol (1.85 Unit/g of substrate) and
Lactose (1.65 Unit/g of substrate) after 96 hrs incubation.
The efficacy of lipase activity of Aspergillus fumigatus (AFems120) was
maximum in the medium containing Sucrose (2.76 Unit/g of substrate) followed
by Glucose (2.54 Unit/g of substrate), Mannitol (1.69 Unit/g of substrate) and
Lactose (1.25 Unit/g of substrate) after 96 hrs incubation.
The efficacy of lipase activity of Aspergillus niger (ANems150) was
maximum in the medium containing Sucrose (3.85 Unit/g of substrate) followed

135
by Glucose (2.44 Unit/g of substrate), Mannitol (1.98 Unit/g of substrate) and
Lactose (1.34 Unit/g of substrate) after 96 hrs incubation.
The efficacy of lipase activity of Pseudomonas sp. (PSems120) was
maximum in the medium containing Sucrose (4.45 Unit/g of substrate) followed
by Glucose (2.95 Unit/g of substrate), Mannitol (2.74 Unit/g of substrate) and
Lactose (1.80 Unit/g of substrate) after 96 hrs incubation.
The efficacy of lipase activity of Pseudomonas sp. (PSems150) was
maximum in the medium containing Sucrose (4.12 Unit/g of substrate) followed
by Glucose (2.45 Unit/g of substrate), Mannitol (2.05 Unit/g of substrate) and
Lactose (1.65 Unit/g of substrate) after 96 hrs incubation.
Effect of Different Nitrogen Sources on the Efficacy of Lipase Activity
The results of different nitrogen sources on the efficacy of lipase activity
of Ethyl Methane Sulphonate treated Aspergillus niger (ANems120) and
Aspergillus fumigatus (AFems120) Aspergillus niger (ANems150) Pseudomonas
sp. (PSems150) and (PSems120)were presented in Tables 54 & 55.
The efficacy of lipase activity of Aspergillus niger (ANems120) was
maximum in the medium containing Sodium nitrate (3.10 Unit/g of substrate)
followed by Potassium nitrate (2.85 Unit/g of substrate), Ammonium sulphate
(2.01 Unit/g of substrate) and urea (1.55 Unit/g of substrate) after 96 hrs
incubation.
The efficacy of lipase activity of Aspergillus fumigatus (AFems120) was
maximum in the medium containing Sodium nitrate (2.76 Unit/g of substrate)
followed by Potassium nitrate (2.67 Unit/g of substrate), Ammonium sulphate

136
(2.13 Unit/g of substrate) and urea (1.54 Unit/g of substrate) after 96 hrs
incubation.
The efficacy of lipase activity of Aspergillus niger (ANems150) was
maximum in the medium containing Sodium nitrate (3.85 Unit/g of substrate)
followed by Potassium nitrate (2.24 Unit/g of substrate), Ammonium sulphate
(1.98 Unit/g of substrate) and urea (1.27 Unit/g of substrate) after 96 hrs
incubation.
The efficacy of lipase activity of Pseudomonas sp. (PSems120) was
maximum in the medium containing Sodium nitrate (4.45 Unit/g of substrate)
followed by Potassium nitrate (2.94 Unit/g of substrate), Ammonium sulphate
(2.71 Unit/g of substrate) and urea (1.45 Unit/g of substrate) after 96 hrs
incubation.
The efficacy of lipase activity of Pseudomonas sp. (PSems150) was
maximum in the medium containing Sodium nitrate (4.12 Unit/g of substrate)
followed by Potassium nitrate (2.99 Unit/g of substrate), Ammonium sulphate
(2.64 Unit/g of substrate) and urea (1.40 Unit/g of substrate) after 96 hrs
incubation.
Figure- 5 showes the comparsion of efficacy of lipase activity by wild
strains and Figure-6 showes the comparsion of efficacy of lipase activity by
chemical treated fungal strains.

137
Indole Acetic Acid (IAA)
Screening Of IAA Production by Wild And Mutated Strains of Aspergillus
niger, Aspergillus fumigatus and Penicillium sp.
The results of IAA activity in wild and mutated strains of Aspergillus
niger, Aspergillus fumigatus and Penicillium sp were presented in Table 56 and
Plate-5 shows the IAA production by bacterial and fungal cultures in Czapek’s
dox broth and Thinlayer chromatogram for IAA.
The wild and mutated cultures were analysed for IAA production
qualitatively by Thin layer chromatography method. All the wild strains except
Aspergillus fumigatus were positive for IAA production. Among the mutated
strains EMS treated Aspergillus niger and UV, sodium azide and Ethyl
Methane Sulphonate treated Aspergillus fumigatus were negative for IAA
production where as rest of all mutated strains were positive for IAA production.
Quantitative Estimation of IAA Production by Wild And Mutated Strains
of Aspergillus niger, Penicillium sp. and Pseudomonas sp.
The wild and mutated cultures were analysed for IAA production
quantitatively by Spectrophotometric method. The results were presented in
Table 57.
The efficacy of Indole Acetic Acid production of Aspergillus niger was
maximum in ANsa120 (17.53µg/L) followed by ANsa90 (16.73µg/L), ANsa60
(15.65µg/L), ANuv30 (13.01µg/L), ANsa30 (12.75µg/L), ANuv40 (12.47µg/L)
and ANuv60 (11.85µg/L).
The wild Aspergillus niger produced 12.33 µg/L of Indole Acetic Acid.

138
The efficacy of Indole Acetic Acid production of Pencillium sp. was
maximum in PEsa120 (10.64µg/L) followed by PEsa90 (9.75µg/L), PEsa60
(8.88µg/L), PEsa150 (8.83µg/L), PEuv30 (8.67µg/L), PEems60 (8.35µg/L),
PEuv40 (7.55µg/L), PEems90 (7.45µg/L), PEuv50 (7.07µg/L), PEems120
(6.97µg/L), PEuv60 (6.83µg/L) and PEems150 (6.35µg/L).
The wild Pencillium sp. produced 7.68 µg/L of Indole Acetic Acid.
The efficacy of Indole Acetic Acid production of Pseudomonas sp. was
maximum in PSuv3 (8.60µg/L) followed by PSems150 (8.45µg/L), PSems120
(8.35µg/L), PSsa60 (8.23µg/L), PSsa30 (8.08µg/L), PSuv4 (8.08µg/L), PSsa90
(8.00µg/L), PSems90 (8.00µg/L), PSuv2 (7.55µg/L), PSems60 (7.07µg/L),
PSsa120 (6.75µg/L) and PEems150 (6.35µg/L).
The wild Pseudomonas sp. produced 7.50 µg/L of Indole Acetic Acid.

139
Discussion
The Phosphate Solubilizing Microorganisms were isolated from rice
rhizosphere soil of Mannachanalur area, Trichy district, Tamilnadu. by using
Pikovskaya agar. Zone formation around the colony in Pikovskaya agar is the
basic criteria for the selection of Phosphate Solubilizing Microorganisms. The
earlier of reports were available for screening of Phosphate Solubilizing
Microorganisms, using Pikovskaya agar. (Hardy et al., 1998; Puente et al., 2004)
showed that the bacteria isolated from the rhizoplane of desert plants, was very
effective in dissolving Calcium phosphates on solid medium.
Phosphate Solubilization
In the present study, the phosphate solubilization efficacy of isolated
fungal strains was higher when compared to the bacterial strains in general.
These results were in coincidence with the earlier findings of Arora and Gaur
(1979);Kucey (1984). Among the fungal strains isolated in the present study, the
Aspergillus niger was found to be predominant phosphate solubilizer followed
by Penicillium sp., Pseudomonas sp. and Aspergillus fumigatus. This may be
due to the production of more organic acid in culture medium by the Phosphate
Solubilizing Microorganism. The more organic acid produced in the culture
medium decrease the pH which is the key indication for more phosphate
solubilization. The Ammonium Sulphate is a nitrogen source of Pikavskaya’s
medium, the acid production in response to the assimilation of cations such as
ammonium is well known fungal phenomenon. The uptake of ammonium by
fungi in a liquid medium commonly leads to a rapid drop in pH of the medium.
Whitelaw et al. (1999) reported that the soluble phosphate concentrations in the
culture medium was directly proportional to organic acid concentration and
inversely related to pH. The main mechanism for phosphate solubilisation by

140
Penicillium radicum was acid production leading to a decrease in pH. Halder et
al. (1991) reported the phosphate solubilization was related to pH decrease
caused by the Bradyrhizobium strains. Cerezine et al. (1988) reported that the
soluble phosphate levels were correlated with pH of the culture medium,
probably due to metabolic activity of Aspergillus niger resulting from
consumption of sugar in the culture medium. Pradhan and Sukla. (2005)
described that the Aspergillus sp. solubilized higher phosphate comparing
Penicillium sp from Tricalcium Phosphate. Drop in pH during growth was more
prominent in absence of Tricalcium Phosphate in the liquid medium. This
indicates that absence of soluble phosphate in media induces the acid production.
Illmer et al. (1995) stated that the production of detectable amount of
organic acids by A. niger is an important factor for phosphate solubilization
mechanism. Nevertheless, organic acids alone are involved, although they are
less effective compared to biotic leaching. Narsian and Patel (2000) explained
that the acidic pH was accompanied with phosphate solubilization Aspergillus
aculeatus in all the experiments and concluded no specific correlation could be
established between maximum phosphate solubilization, growth or pH. Seshadri
et al. (2004) also stated that the release of phosphate by Aspergillus niger in the
liquid culture was associated with reduction in pH of culture medium. Barroso et
al. (2006); Achal et al. (2007) studied the relationship between phosphate
solubilization, extensive growth, acid production, and decrease in pH. They
concluded the phosphate solubilization was related to acid production, pH drop
and Aspergillus thubingensis growth in the culture medium.
Sperber (1957), Das (1963), Sethi and Subba Rao (1968) and Ahmed and
Jha (1968) were reported there is no significant relationship could be established
between the quantity of phosphate solubilized and drop in pH. The results of pH

141
drop also support the data of phosphate solubilization by Pseudomonas, Bacillus
and Rhizobium, which is associated with acidification of the culture medium, but
the extent of phosphate solubilization and pH drop are not proportionally co-
related (Rodriguez and Fraga, 1999).
Bolan et al. (1994), Sagoe et al. (1997), Alam et al. (2002), and Kumari
et al. (2008) have reported that Malic, Tartaric, Oxalic, and Citric acids have
high capability to release soluble phosphate from insoluble Tricalcium
Phosphate as well as Rock Phosphate. Kang et al. (2008) suggested that the
solubilizing ability is not related to organic acid produced but to the nature of
organic acid produced by Aspergillus sp. Kapri and Tewari (2010) highlighted
the comparative phosphate solubilizing potential of Trichoderma sp. isolated
from Pinus, Deoder, Bamboo and Oak rhizospheres. It was found that all the
isolates were capable of differentially utilizing Tricalcium Phosphate from liquid
medium. This was indicated by the soluble phosphate concentrations and
increase in acidity of the growth medium.
In the present study, the phosphate solubilization efficacy of the wild
strains was observed in the descending order of Aspergillus niger >Penicillium
sp. > Aspergillus fumigatus. Among the UV irradiated fungal strains, ANuv60
was found as the predominant phosphate solubilizing strain followed by
ANuv50, PEuv60, ANuv30, ANuv40, PEuv50 and AFuv50. In chemical
mutagenesis of phosphate solubilizing strains, ANems120 showed the highest
phosphate solubilization activity followed by ANsa120, PEsa150, PEems150,
ANsa90, ANems90, PEsa120, ANsa60 and AFsa120. Achal et al. (2007)
reported a significant increase in soluble phosphate level was observed in case of
UV induced mutants of Aspergillus strains compared with wild strains. The
present study also revealed that UV induced mutant enhanced the phosphate

142
solubilization compared with wild strains. There might be a possibility of
alteration at genetic levels in case of mutants. Tripura et al. (2007) reported that
the Ethyl Methane Sulphonate treated microbial strain Serratia marcescens have
increased phosphate solubilization efficiency compared to the wild strains.
Reyes et al. (2001) studied that phosphate solubilization of two UV induced
Penicillium rugulosam along with wild strain. Among the studied strains, MP++
strain solubilized high amount of phosphate and they stated that the
solubilization depends on the type of phosphate source.
In the present study, UV induced Penicillium sp. showed less
solubilization when compared to UV induced Aspergilus niger and the same
showed high solubilization when compared to wild Penicillium sp. This may be
due to more production of organic acid, Acid Phosphatase and Phytase enzyme
by mutant strains, which was supported by Achal et al. (2007). The mechanisms
of action of mineral phosphate solubilization (MPS) were studied in the wild-
type Mps+ Penicillium rugulosum strain and in negative (Mps−) and
superpositive (Mps++) mutants derived from it. MPS activities were measured in
liquid media using Sucrose as carbon source and nitrite, ammonium plus nitrate,
ammonium and arginine as nitrogen sources. Ammonium significantly decreased
phosphate solubilization, and this activity was higher in the Mps++ mutant than
in the wild-type depending on the N sources used. The Mps+ phenotype was
strongly associated with the production of gluconic or citric acids. The results
also suggest for the MPS− mutant the involvement of the H+ pump mechanism in
the solubilization of small amounts of phosphates. (Reyes et al. 1999). Vassileve
et al. (1998) reported the encapsulated spores of Aspergillus niger solubilized
Rock Phosphate in the culture medium due to the production of organic acids.

143
In the present study, Pseudomonas sp. was found to be the lowest
phosphate solubilizer, when compared to the fungal strains. The UV and
chemical mutated Pseudomonas strains were showed decreased phosphate
solubilizing capacity when compared to wild strains. These results were in
acceptable with the findings of Miller et al. (2010), who stated that mutants with
showed calcium phosphate solubilization ability. Mutations in the gcd and pqqE
genes greatly reduced the solubilization ability, whereas mutations in the pqqB
gene only moderately reduced this ability. The combination of biochemical
analysis and genomic comparisons revealed that alterations in the pqq
biosynthetic genes, and the presence/absence of the gluconate dehydrogenase
(gad) gene, fundamentally affect phosphate solublization in strains of
P. fluorescens. Goldstein and Liu (1987) have shown that mineral phosphate
solubilizing activity by Erwinia herbicola is genetically coded in a gene cluster
on the plasmids of microorganisms possessing this activity. It was also reported
that gene expression and mineral phosphate solubilization of bacteria is affected
by the presence of soluble phosphate due to feed back regulation. Kumar et al.
(2001) suggested that it is possible to select physiologically efficient strains of
A. chroococcum through mutagenesis starting from soil isolates and that
microbial inoculants can be used as an economic input to increase crop
productivity with lower fertilizer levels.
The present study indicates that the solubilization efficacy of the fungal
strain Aspergillus niger treated with Sodium Azide (ANsa120) and Penicillium
sp. treated with Ethyl Methane Sulphonate (PEems150) were about 1.5-fold
higher when compared to wild strain. The chemical mutagenic agents such as
Sodium Azide, Ethyl Methane Sulphonate may alter the gene sequence by
altering the base pairs. The azide ion alters the structure of cytosine such that it
forms hydrogen bonds with adenine, rather than guanine. This produces a

144
cytosine to thymine transition. Ethyl Methane Sulphonate is a strong mutagenic
agent. It alkylates N7 of Guanine and severely alters the base pairing.
The phosphate solubilization efficacy of mutant fungal strains
(ANems120, ANsa120 and PEsa150) were demonstrated in the presence of
various carbon and nitrogen sources. The effect of different carbon sources on
phosphate solubilization efficiency of fungal strains was found to be in the
ascending order Mannitol < Lactose < Sucrose < Glucose. This is in
disagreement with the findings of Seshadri et al. (2004) who reported that the
Mannitol was the best carbon source utilized by Aspergillus niger preferred for
higher phosphate solubilization. The present study revealed that Mannitol was
found to be the poorest carbon source and the Urea as one of the predominant
nitrogen sources for phosphate solubilization by Aspergillus niger. This in
confirmation with the findings of Seshadri et al. (2004) who reported the
nitrogen in the form nitrate was very effective and Urea was the poorest source
of nitrogen by Aspergillus niger. This may due to the genetic alteration in the
fungus due to chemical mutation. Reyes et al. (1999) studied that the Sucrose
appeared to be the best carbon source for phosphate solubilization by UV
induced Penicillium rugulosum. Nautiyal et al. (2000) reported that the Glucose
and Lactose were the best carbon source and Sucrose, Sorbitol were identified as
poor carbon source for phosphate solubilization, ammonium and nitrate source
to be equally effective for phosphate solubilization. This is in contrary with the
earlier report of Halder et al (1991) who stated that Glucose decreased the
medium pH most and caused highest solubilization of phosphate followed by
arabinose, sodium acetate, sodium gluconate and sodium succinate. Mohamed et
al (1985); Abd-Alla (1994) reported that ammonium salts induced phosphate
solubilization whereas the nitrate salts repress the phosphate solubilization
activity.

145
Similarly, in the present study, the addition of various nitrogen sources in
the medium to determine their effect on phosphate solubilization efficiency of
the inoculated strains (ANems120, ANsa120 and PEsa150) gave the following
results in the ascending order Sodium Nitrate < Potassium Nitrate < Urea <
Ammonium Sulphate. Whitelaw et al. (1999) reported that the Penicillium strain
solubilized the insoluble phosphate in the culture medium containing ammonium
or nitrate as sole source of nitrogen. Vassileve et al. (2007) compared the
phosphate solubilization efficacy of Aspergillus niger by using corn steep liquor,
Ammonium Sulphate and yeast extract as nitrogen source. The phosphate
solubilization activity of Xanthomonas campestris was measured in both the
wild type and mutant strains using various carbon and nitrogen sources. Glucose
was found to be the best in both (wild 39.9%; mutant 67.1%) strains followed by
Sucrose (46.8%) in the mutant and molasses (36.0%) in the wild type.
Ammonium Sulphate was the best nitrogen source for both the strains, followed
by and Urea. (Sharan et al., 2008). Son et al. (2006) investigated the effect of
carbon sources on the insoluble phosphate solubilization Pseudomonas species.
It solubilized insoluble phosphate with all carbon sources except Adonitol and
Lactose. They also studied all nitrogen sources except Sodium Nitrite, increased
the level of soluble phosphate production. Among the carbon and nitrogen
sources used, Glucose and Ammonium Nitrate was the best carbon and nitrogen
source respectively for the insoluble phosphate solubilization by Pseudomonas
species. In all cases insoluble phosphate solubilization was accompanied by a
distinct pH decrease.
Cerezine et al. (1988) reported the soluble phosphate levels were
correlated with pH of the culture medium probably due to metabolic activity of
fungus resulting from consumption of sugar in the Aspergillus niger culture
medium. Fructose, Glucose, xylose and Sucrose were the carbohydrates that

146
favored ‘P’ solubilization when compared to Galactose and Maltose. Among the
nitrogen sources tested ammonium salts favored the production of larger
amounts of soluble phosphate than peptone, Urea and Nitrate. The results were
positively correlated with the present study. Phosphate solubilization was
decreased with decreasing Glucose concentration; Solubilization was increased
when the nitrogen source was Nitrate by the soil fungus Aspergillus niger
(Hardy et al. 1998). The present study revealed that the carbon source Glucose
and nitrogen source Ammonium Sulphate were the best for phosphate
solubilization, which confirm results of earlier reports. Goenadi et al. (2000)
have reported that the accumulation of more insoluble phosphate using
Aspergillus niger and Glucose as the carbon source. Illmer and Schinner (1992)
have shown that the accumulation of high insoluble phosphate using Glucose
utilizing Penicillium sp. and Pseudomonas sp. respectively. According to
Srividya et al. (2009) the Aspergillus niger utilized the nitrogen salts having
either ammonium group or nitrate group or both as nitrogen source. Ammonium
Sulphate was found to be best in reducing the medium pH and simultaneous
solubilization of phosphate, out of the entire nitrogen sources used and observed
that Aspergillus niger was able to utilize Ammonium Sulphate most efficiently
to decrease the pH of the medium for phosphate solubilization. Fasim et al.
(2002) have reported that the bacterial isolates from the air environments of
tannery solubilize the Zinc Phosphate only in presence of Glucose. Disimmine,
(1998) reported solubilization of Zinc Phosphate by Pseudomonas fluorescens
only in presence of Glucose and slight solubilization in presence of mannose
while no solubilization was detected in presence of Gluconate, Galactose,
Glycerol, Sorbitol and Fructose.
The present study reported that the poorest carbon source and nitrogen
source were Mannitol and Sodium Nitrate respectively for phosphate

147
solubilization. The results of present study were negatively correlated with
Barroso et al. (2006) who stated that solubilization of CaHPO4 was enhanced by
Aspergillus niger when the carbon source was Mannitol, Maltose, Galactose and
Glucose (in that order) and this was positively correlated with the present study
when evaluating with nitrogen sources. The solubilization of CaHPO4 or AlPO4
decreased in the following order: Glycine > NH4Cl > NaNO3 and NH4NO3 >
Urea > (NH4)2SO4, respectively. Ammonical nitrogen (NH4+-N) sources were
the most effective in the production of acids and in lowering of the pH.
Previous reports on phosphorus solubilizing microorganisms like
Ectomycorrhizal fungi by Lapeyrie et al., (1991) who attributed the differences
in phosphate solubilization (when ammonium and nitrate were used) to the use
of different mechanisms for the generation of acidity in the culture. Production
of organic acids is an important mechanism, but is not the only possibility for
phosphate solubilization. The other mechanism involved in solubilization of
insoluble phosphates is extrusion of H+ ions during ammonium ions assimilation
by Parks et al. (1990) for Penicillium like fungus, Illmer and Schinner (1995) for
Pseudomonas sp. and Penicillium aurantiogriseum Illmer et al. (1995) for
Aspergillus niger, Penicillium simplicissimum, Pseudomonas sp. and Penicillium
aurantiogriseum. In the present study, the phosphate solubilization using
ammonium salts as the nitrogen source was substantially higher than with other
nitrogen sources. Thus, the results suggested the second mechanism which
acidification of the medium by proton extrusion during ammonium ions was
involved in phosphate solubilization by these isolates.
Phosphatase
The Phosphatase production capacity of isolated bacterial and fungal
cultures were analysed by inoculating the bacterial and fungal isolates into

148
pikovskaya broth. In the present study, para nitro phenyl phosphate was used as
a substrate for Acid Phosphatase assay. The earlier reports were available for the
utilization of para nitro phenyl phosphate as a substrate for Acid Phosphatase
assay (Prada et al. 1996;Puruchothaman, 1994).
In the present study, the Phosphatase production efficacy of the wild
strains were observed in the descending order of Aspergillus niger>
Pseudomonas sp >Penicillium sp. > Aspergillus fumigatus. The UV irradiated
and chemical treated bacterial strains in the present study showed that decrease
in Phosphatase production, when compared to wild strains. Among the UV
irradiated fungal strains, ANuv60 was found the best Phosphatase producing
strain followed by ANuv50, ANuv30, ANuv40, AFuv50, PEuv50, AFuv60,
PEuv40, PEuv60, PEuv30, AFuv40 and AFuv30.
Among the chemical treated fungal strains, ANsa120 was the
predominant fungal strain followed by ANems120, PEems150, ANsa90,
PEsa150, ANems90, PEsa120, AFsa120, PEems120, PEems60, PEems90,
ANems60, AFsa120, AFsa90, PEsa90, ANsa30, AFems120, ANems30,
AFems90, PEsa60, AFsa60, AFems60, AFems30 and AFsa30. The result of the
present study was in agreement with Relwani et al. (2008) who reported that the
enzyme activities such as Acid Phosphatase and Phytase was increased
significantly in mutants of A. tubingensis when compared to the wild type. Achal
et al. (2007) reported that Acid Phosphatase production by UV induced mutants
of Aspergillus tubingensis, might be the reason for highest phosphate
solubilization by mutant strains. The production of the Acid Phosphatase was
favoured by low pH values.

149
Among all physical and chemical treated fungal strains, the best
Phosphatase producing strains such as ANsa120, ANems120 and PEems150
were demonstrated in the presence of various carbon and nitrogen sources in the
present study. The effect of different carbon sources on Phosphatase production
efficacy of fungal strains were found to be in the ascending order of Mannitol<
Lactose < Sucrose < Glucose. Andrea et al. (1990) reported that Pseudomonas
aeruginosa and Rhizobium meliloti, utilized several choline derivatives as the
sole carbon and nitrogen source and increased the production of Acid
Phosphatase activity. Spicers and McGill (1979) demonstrated an increase in
Phosphatase activity during incubation of soil amended with Glucose and
Ammonium Sulphate.
The effect of different nitrogen sources on Phosphatase production
efficacy of fungal strains were found to be in the ascending order of Sodium
Nitrate < Potassium Nitrate < Urea < Ammonium Sulphate in the present study.
The results were in agreement with Semenova et al. (1986) who stated that the
synthesis of the secreted enzyme depended on the sources of carbon and nitrogen
nutrition. The enzyme yield was highest in a medium with Sucrose as a carbon
source and Ammonium salt as a nitrogen source. The secretion of Acid
Phosphatase is stimulated by an increase in the sugar content and a deficiency of
the nitrogen source in the medium.
Semenova et al. (1986) studied that Acid Phosphatase production from
Saccharomyces cerevisiae in presence of phosphate concentration of 10 mM
which showed repression of the Phosphatase production. Sudha and
Purushothaman (2000) reported that Pseudomonas strain isolated from the
freshwater pond system showed repression. Casida (1959) isolated several
species of Aspergillus sp. capable of producing Acid Phosphatase active against

150
organic phosphate in soils. Smith et al. (1973) reported the addition of gradient
amount of Potassium phosphate to Phosphatase test broth was the least
inhibitory to Phosphatase activity by Candida tropicalis. Sarapatka et al. (2004)
stated that the increasing amount of available phosphate increasing the
Phosphatase activity in cereal roots. Pedregosa et al. (1991) reported that
Aspergillus strains produce non repressible, repressible and phosphate inducible
Phosphotases. It is accepted that culture condition may cause fluctuations and
multiplicity of fungal Phosphatases. Ponmurugan and Gopi (2006) stated that
there was a positive correlation between phosphate solubilizing capacity and
Phosphatase enzyme activity. These earlier reports were in agreement with
present results which showed that the increase in inorganic phosphate
concentration due to phosphate solubilization were not repressed the
Phosphatase production by all strains used in this study. But disagreement with
the earlier reports of Semenova et al. (1986) who studied that Acid Phosphatase
production from Saccharomyces cerevisiae in presence of phosphate
concentration of 10 mM which showed repression of the Phosphatase
production. Ohta et al. (1968) stated that the production of Phosphatase by the
black koji fungus Aspergillus awamori is a repressible enzyme.
The earlier reports explained that there are two systems of regulation in
Phosphatase induction in microorganisms. This may be the reason for repressible
and non repressible nature of Phosphatases by inorganic phosphate. Singleton
and Sainsberry (1988) explained that about the phosphate controlled genes or
pho genes, which is the gene cluster, consist of Pho A encodes Phosphatase, Pho
S encodes inorganic phosphate binding site, PhoE encodes porin and Pho B
encodes the regulator gene as like the operon model. Wagner et al. (1995)
reported that Synechoccus strain system of phosphate irrepressible Acid
Phosphatase in addition to the normal repressible system.

151
Lipase
The Lipase production capacity of isolated bacterial and fungal cultures
were analysed by inoculating the isolates in Czapek’s dox broth enriched with
1% Olive oil as a inducer and the same were inoculated in Czapek’s dox broth
replaced carbon source as 10% Olive oil for Lipase production. The number of
reports were available for the addition of inducer to induce the production of
Lipases. Salihu et al. (2011) utilized Olive oil and Palm oil as an inducer. Olive
oil by Rifaat et al. (2010); Hosseinpour et al. (2011); Cocunut oil by Rani and
Panneerselvam (2009) and different lipid sources namely Olive oil, Coconut oil,
Groundnut oil, Triacetin, Tributyrin and Surfactants namely Tween 20, Tween
40, Triton –X was used for acceleration of Lipase by Pogaku et al. (2010). It was
suggested that the Lipase formed might be derived from carbohydrates in the
medium with small quantities of lipid material as inducer (Minoda and Ota,
1980; Iwai et al., 1973).
In the present study, the Lipase activity of isolated bacterial strains were
higher compared to the fungal strains in the present study. Rani and
Panneerselvam (2009) showed that Aspergillus fumigatus, A. terreus,
Penicillium chrysogenum, P. funiculosum and Fusarium moniliforme were
selected as the highest Lipase producers. In the present study, the Lipase
production efficacy of the wild strains was observed in the descending order of
Pseudomonas sp.>Aspergillus niger >Penicillium sp. > Aspergillus fumigatus.
Juichiro et al. (1963) reported that several strains of Rhizopus and Asphergillus
niger as most potent Lipase formers. Successful isolation of the enzyme could be
made from the bran-koji culture of Aspergillus niger. Ogundero (1981) stated
that A. fumigatus and A. nidulans were able to degrade vegetable oils and
triglycerides.

152
Among the UV irradiated bacterial strains in the present study, PSuv3
was found to be the predominant Lipase producing strain grown in both Sucrose
and 10% Olive oil containing medium followed by PSuv4, PSuv2 and PSuv1.
Among the UV irradiated fungal strains, ANuv50 was found the best Lipase
producing strain grown in both Sucrose containing medium followed by
AFuv50, ANuv40, ANuv60, ANuv30, AFuv60, PEuv60, PEuv50, PEuv40,
AFuv40, PEuv30 and AFuv30.
Lawrence et al. (1967) studied the Lipase activity was considerably
increased by nutritional and physical conditions from Pseudomonad. Suzuki et
al. (1988) used the Olive oil as a carbon source for microbial growth and Lipase
production by Pseudomonas sp. Ray et al. (1999) reported the Lipase activity of
the isolated bacterial strain, Corynebacterium sp. was increased 2.3 fold by
mutagenic technique. This was agreed with our present study, since all mutant
strains tested were found to produce more Lipase activity, when compared to
wild strains. The present study also indicated, the strain Pseudomonas sp.
(PSems120) treated with Ethyl Methane Sulphonate, produced 2-fold higher
Lipase activity when compared to wild strain. PSems120 which was the
predominant Lipase producing strain, among all the strains used in the present
study and the bacterial strains PSems150, PSems90, PSsa90 and PSsa120 were
followed the strain PSems120. ANems150 was the best strain for Lipase
production among the chemical mutated fungal strains followed by ANems120,
AFems120, Ansa90, AFems150, AFems90, ANsa60, PEsa120 and PEems150
grown in Sucrose containing medium. Ellaiah et al. (2002) isolated a fungal
strain, which produced Lipase constitutively was used to produce mutants using
physical and chemical agents and the mutant strain with Lipase productivity of
2-fold higher was obtained. Bapiraju et al. (2004) reported that the the UV and
chemical treated fungal strains of Rhizopus sp. showed 133% to 232% higher

153
than the wild strains. Caob and Zhanga (2000) reported that the Lipase
production of UV and chemical treated Pseuomonas mutant 3.25 fold higher
than the wild strain.
The present study stated that the mutant strain PSems120, ANsa90,
PSsa90, AFuv50 and PEsa120 showed 2 fold and PSems150, PEuv40, PEsa90,
AFsa120 and PSsa90 showed 1.5 fold enhanced productivity of enzyme, over
the wild strain. This was in agreement with findings of Mahadik et al. (2004),
who reported that Mutant strains of Aspergillus niger showed seven to five fold
enhanced productivity of Lipase, over the wild strain. Mala et al. (2001) isolated
UV and nitrous acid derived mutants of A. niger selected on media containing
bile salts. Nitrous acid mutants exhibited increased efficiency of Lipase
production compared with wild strain. Mutation alters the genotype of
microorganisms, when it expresses that leads to alter the character or death of
microorganisms. The ultra-violet radiation forms thymine dimer in gene
sequence. But the photolyase enzymes present in living system break the
thymine dimer and correct it. The increasing exposure time to UV radiation may
forms the thymine dimer in gene sequence that code photolyase enzyme. In this
situation the thymine dimer can not break by the enzyme of living system
(Freifelder, 1990; Radman, 1999). The chemical agents such as Sodium Azide,
Ethyl Methane Sulphonate may alter the gene sequence by altering the base
pairs. The azide ion alters the structure of cytosine such that it forms hydrogen
bonds with adenine, rather than guanine. This produces a cytosine to thymine
transition. Ethyl Methane Sulphonate is a strong mutagenic agent. It alkylates
N7 of Guanine and severely alters the base pairing.
In the present study, all the bacterial strains were showed less Lipase
activity, when compared to fungal strains grown in 10% Olive oil containing

154
medium and moreover all the strains, grown in 10% Olive oil containing
medium were showed poorest Lipase activity. This may due to the inoculated
strains failed to utilize more 10% Olive oil as a carbon source for their growth
and subsequently reduction in Lipase activity. This was in agreement with
Mahadik et al. (2004) who stated, higher concentration of oil in the medium did
not help Lipase production in the case of mutant. On the other hand, Sucrose,
Lactose, Mannitol and Glucose may support the growth initially and the inducer
1% Olive oil induced the inoculated strains for more Lipase production in latter
stages. Many researchers have reported the positive effect of sugars on Lipase
production by Aspergillus niger (Pal et al., 1978), by Rhizopus chinensis
(Nakashima et al., 1988) and by Rhizopus oryzae (Salleh et al., 1993). On the
contrary of present study, Muralidhar et al. (2001) stated that their experimental
results indicate that Olive oil is a better carbon source for Lipase production by
Candida cylindraces compared to Glucose. This is supported by Dalmau et al.,
2000; Gordillo et al., 1998 for Candida rugosa. Optimization of the quantity of
Olive oil in the fermentation medium along with the other nutrients resulting in
the rise in enzyme production also contradicts the fact that large quantities of
Olive oil decrease the Lipase activity by for Candida rugosa (Gordillo et al.,
1995).
Nahas (1988) reported that Carbohydrates were good sources of carbon
for growth but low Lipase production was obtained by Rhizopus oligoporus.
Lipase production was strongly repressed by higher concentration of Glucose. In
present study revealed that fungal culture utilizing Olive oil as a carbon source
exhibited lowest Lipase producing activity. This result was not in agreement
with those of Paul and Carles (1992). Falony et al. (2006) reported that the A.
niger strain showed more Lipase activity among the test strains used. The
production of Lipase was more significant in culture medium added with lipids

155
as the carbon source than in the culture medium without lipids. Previous works
on the physiology of Lipase production showed that the mechanisms regulating
biosynthesis vary widely in different microorganisms. Results obtained with
Calvatia (Christakopoulos et al., 1992), Rhizopus (Salleh et al., 1993),
Aspergillus (Pokorny et al., 1994), and Rhodotorula (Papaparaskevas et al.,
1992), showed that Lipase production seems to be constitutive and independent
of the addition of lipid substrates to the culture medium, although their presence
enhanced the level of Lipase activity produced. On the other hand, (Shimada et
al., 1992; Rapp, 1995), lipid substrates are necessary for Lipase production by
Geotrichium candidum and Fusarium oxysporum respectively and also,
carbohydrates can act as repressors of its biosynthesis.
In the present study, Lipase production efficacy of mutant fungal strains
(ANems120, AFems120, ANems150, PSems120 and PSems150) were
demonstrated in the presence of various carbon and nitrogen sources. The effect
of different carbon sources on Lipase production efficiency of bacterial and
fungal strains was found to be in the ascending order Mannitol < Lactose <
Glucose < Sucrose. In present study Aspergillus niger, and Penicillium sp.,
utilized Sucrose as a carbon source exhibited highest Lipase producing activities,
these results are in agreement with those of Susumu and Yashio (1975). Elliah
et al. (2002) reported that the Aspergillus niger utilized dextrose as carbon
source for highest Lipase activity. The Lipase production was decreased in the
order of Sucrose < Lactose < Mannitol < Dextrose by the mutant strain of
Aspergillus niger. These results were in contrary to the present study results.
Petrovic et al. (1990) reported maximum Lipase production when
Glucose and peptone were incorporated in the production medium using
Penicillium roquefortii. In the present study, Glucose favoured the production

156
of Lipase. This is in contrary with Mahadik et al. (2004) who reported that there
was no increase in enzyme levels was observed when mutant UV-10 was grown
in medium supplemented with Glucose. However, the addition of Glucose in the
medium resulted in increased levels of Lipase production by wild strain,
Aspergillus niger. The maximum Lipase activity was found with the
combination of Sucrose with Olive oil inducer and Glucose with Olive oil
inducer in the present study. But the repressive effect of Glucose with fatty oil is
reported by Dalmau et al. (2000) for Candida rugosa, Nahas (1988) for
Rhizopus oligosporus, Baillargeon et al. (1989) for Geotrichium candidum, and
Rapp (1995) for Fusarium oxysporum. but differ from those obtained for C.
rugosa by Chang et al. (1994). Cordova (1998) reported that the carbon sources
such as Glucose, Fructose, Glycerol, Xylose, Sucrose and Lactose are probably
utilized before Lipase production, there were little or no differences with these
substrates as carbon source.
Damaso et al. (2008) reported that the Aspergillus niger was found to be
the best Lipase producer. Soap stock was the best substrate and inducer compare
to Olive oil and they also explained the repressive effect Olive oil on Lipase
production. Coca et al. (2001) stated that the Aspergillus niger and Aspergillus
fumigatus were the best Lipase producer in the medium containing Olive oil as a
carbon source. This is in contrary with our present results. The present study
revealed that Aspergillus fumigatus utilizing Mannitol and Lactose as a carbon
sources and exhibiting lowest Lipase producing activity. In case of Sucrose and
Glucose exhibited highest Lipase producing activity.
Kakde and Chavan. (2011) found that carbon sources like Fructose and
Sucrose induced Lipase activity while Lactose, Starch and Carboxyl Methyl
Cellulose inhibited Lipase activity by Penicillium chrysogenum. Nitrogen

157
sources like Nitrate, Nitrite, Amide, Ammonium, and Protein showed its effect
on Lipase enzyme of fungi. Casein and Peptone which are organic forms
stimulated maximum Lipase enzyme production of storage fungi. Both the
species of Penicillium viz. Penicillium notatum and Penicillium chrysogenum
showed maximum extracellular Lipase activity in presence of Casein and
Peptone. Urea which is an amide form and Ammonium Phosphate which is an
ammonium form hampered the extracellular Lipase enzyme production of fungi.
These reports were found to be similar to our present study results. Chavan and
Kakde, (2009) studied that the Lipase enzyme activity of storage fungi under the
influence of carbon and nitrogen sources. They found that carbon sources as like
Fructose and Sucrose induces Lipase activity while Starch, Lactose and
Carboxyl Methyl Cellulose inhibit Lipase activity. Nitrogen sources as like
Nitrate, Nitrite, Amide, Ammonium, and Protein affect in different ways on
Lipase enzyme of fungi. The results of the present study revealed that the
Lactose and Urea was a poor carbon and nitrogen sources respectively in the
tested bacterial strains like PSems120 and PSems150 and fungal strains like
ANems120, AFems120 and ANems150.
The effect of different nitrogen sources on Lipase production efficiency
of bacterial and fungal strains was found to be in the ascending order Urea
<Ammonium Sulphate < Potassium Nitrate < Sodium nitrite in the present
study. This was in accordance with Lima et al. (2003), who stated that the
Lipase activity by Penicillium sp. was higher when the medium containing only
the inorganic nitrogen sources and in the presence of Olive oil, Ammonium
Sulphate produced less mycelial growth subsequently less Lipase activity. But
the Ammonium Sulphate and Potassium Nitrate combination produced higher
Lipase activity in short fermentation time. Other workers (Freire et al., 1997;
Pereira-Meirelles et al., 1997;Samad et al., 1990) employed Candida lipolytica,

158
Penicillium restrictum and Rhizopus sp. reported maximum Lipase production,
when organic nitrogen was used as a nitrogen source. Waller and Comeau
(1990) reported that the incorporation of corn steep liquor into the production
medium gave good Lipase activity by Candida sp. Rodriguez et al. (2006)
reported that Ammonium Sulphate, Ammonium Nitrate and Sodium Nitrate
except Urea reduced the production of Lipase activity compared to the initial
medium containing yeast extract. With Urea, the activity was around six times
higher. A similar observation has been reported using Penicillium sp. (Gombert
et al., 1999). On the other hand, Lima et al. (2003) found that Lipase production
in Penicillium sp. was stimulated using Ammonium Sulphate.
Indole Acetic Acid
The phosphate solubilizing wild and mutant strains of bacteria and fungi
were analysed for their Indole Acetic Acid production efficacy qualitatively by
Thin Layer Chromatography. Phosphate Solubilizing Bacteria are capable of
producing physiologically active Auxins that may.have pronounced effects on
plant growth. (Ponmurugan and Gopi. 2006). Shahab et al. (2009) reported that
the phosphate solubilizing microbes from Mung beans excreted the
phytohormones like Auxin and ethylene.
The results of present study revealed that the wild strains of Pseudomonas
sp., Aspergillus niger and Penicillium sp. have the ability to produce the plant
hormone Indole Acetic Acid except Aspergillus fumigatus. Among the mutated
bacterial and fungal strains, all the strains were able to produce Indole Acetic
Acid except Ethyl Methane Sulphonate treated Aspergillus niger and all the
strains of Aspergillus fumigatus. Since the wild strains failed to produce Indole
Acetic Acid, the mutant Aspergillus fumigatus might not produced the Indole
Acetic Acid.

159
Kulanthaivel et al. (2006) reported that Azosprillum sp. were treated with
UV, Acridine orange and Ethidium bromide. The production of Indole Acetic
Acid by most of mutant strains were higher when compared to wild strains. This
is in agreement with the present study results, which stated most of the mutant
produce more amount of Indole Acetic Acid when compared to wild strains.
Garcia et al. (1980) stated that the mutant Azosprillum brasilense excreated
Indole Acetic Acid more when compare to wild strains. Normanly et al. (1983)
confirmed that the orangepericap mutant Arabidopsis thaliana, accumulate
Auxin greater than wild type.
Tien et al. (1979) reported that Azospirillum and phosphobacteria isolated
from the soil of pearl millet produced Indole Acetic Acid, gibberellic acid and
cytokinin like substances. (Brown, 1972), and some of them are capable of
dissolving phosphate (Barea et al., 1976). Production of Indole Acetic Acid
varies greatly among different species and is also influenced by culture
conditions, growth stage and availability of substrate (s) (Brown, 1972; Vijila,
2000).

160
Summary and Conclusion
The Phosphate solubilizing microorganisms were isolated from rice
rhizosphere soil of Mannachanalur area, Trichy district, Tamilnadu. by
using Pikovskaya agar.
The efficacy of Phosphate solubilization assay and Phosphatase activity
assay of isolated bacterial and fungal strains were analysed by
spectrophotometrically using Pikovskaya broth.
The efficacy of Lipase activity by the isolated bacterial and fungal strains
were analysed spectrophotometrically using Czapadox broth.
Screening of Indole Acetic Acid production of isolated bacterial and
fungal strains were analysed by Thin Layer Chromatography and
spectrophotometric estimation using Czapadox broth.
Random mutational studies such as UV and chemical mutation of isolated
bacterial and fungal strains to improve their phosphate solubilization,
Phosphatase activity, Lipase activity and Indole Acetic Acid production
capability were carried out.
The results of present study revealed that the phosphate solubilization
efficacy of isolated fungal strains was higher when compared to the
bacterial strains.
The phosphate solubilization efficacy of the wild strains was observed in
the descending order of Aspergillus niger >Penicillium sp. > Aspergillus
fumigatus.
Among the UV irradiated fungal strains, ANuv60 was found as the
predominant phosphate solubilizing strain.
ANems120 showed the highest phosphate solubilization activity among
the chemical mutated fungal strains.

161
The solubilization efficacy of the fungal strain Aspergillus niger treated
with Sodium Azide (ANsa120) and Penicillium sp. treated with Ethyl
Methane Sulphonate (PEems150) were about 1.5-fold higher when
compared to wild strain.
The effect of different carbon sources on phosphate solubilization
efficiency of fungal strains was found to be in the ascending order
Mannitol < Lactose < Sucrose < Glucose.
The effect of different nitrogen sources on phosphate solubilization
efficiency of fungal strains was found to be in the ascending order
Sodium Nitrate < Potassium Nitrate < Urea < Ammonium Sulphate.
Lipase activity of isolated bacterial strain Pseudomonas sp. higher when
compared to the isolated fungal strains.
The Lipase production efficacy of the wild microbial strains were
observed in the descending order of Pseudomonas sp.>Aspergillus niger
>Penicillium sp. > Aspergillus fumigatus.
Among the UV irradiated bacterial strains in the present study, PSuv3
was found to be the predominant Lipase producing strain grown in both
Sucrose and 10%Olive oil containing medium.
The Pseudomonas sp. (PSems120) was the predominant Lipase producing
strain, among all the strains, which produced 2-fold higher Lipase
activity when compared to wild strain.
The effect of different carbon sources on Lipase production efficiency of
bacterial and fungal strains was found to be in the ascending order of
Mannitol < Lactose < Glucose < Sucrose.
The effect of different nitrogen sources on Lipase production efficiency
of bacterial and fungal strains was found to be in the ascending order of
Urea <Ammonium Sulphate < Potassium Nitrate < Sodium nitrite.

162
The Phosphatase production efficacy of the wild strains were observed in
the descending order of Aspergillus niger.> Pseudomonas sp
>Penicillium sp. > Aspergillus fumigatus.
Among the UV irradiated fungal strains, ANuv60 was found to be the
best Phosphatase producing strain.
Among the chemical treated fungal strains, ANsa120 was the
predominant fungal strain for Phosphatase activity.
The effect of different carbon sources on Phosphatase production efficacy
of fungal strains were found to be in the ascending order of Mannitol<
Lactose < Sucrose < Glucose.
The effect of different nitrogen sources on Phosphatase production
efficacy of fungal strains were found to be in the ascending order of
Sodium Nitrate < Potassium Nitrate < Urea < Ammonium Sulphate.
The wild and mutated strains of Pseudomonas sp., Aspergillus niger and
Penicillium sp. have the ability to produce the plant hormone Indole
Acetic Acid except Aspergillus fumigatus.
It can be concluded from the findings of present study that the wild and
mutant bacterial and fungal strains like Pseudomonas sp., Aspergillus niger,
Penicillium sp. and Aspergillus fumigatus can be exploited biotechnologically
for the production of Phosphatase and Lipase.
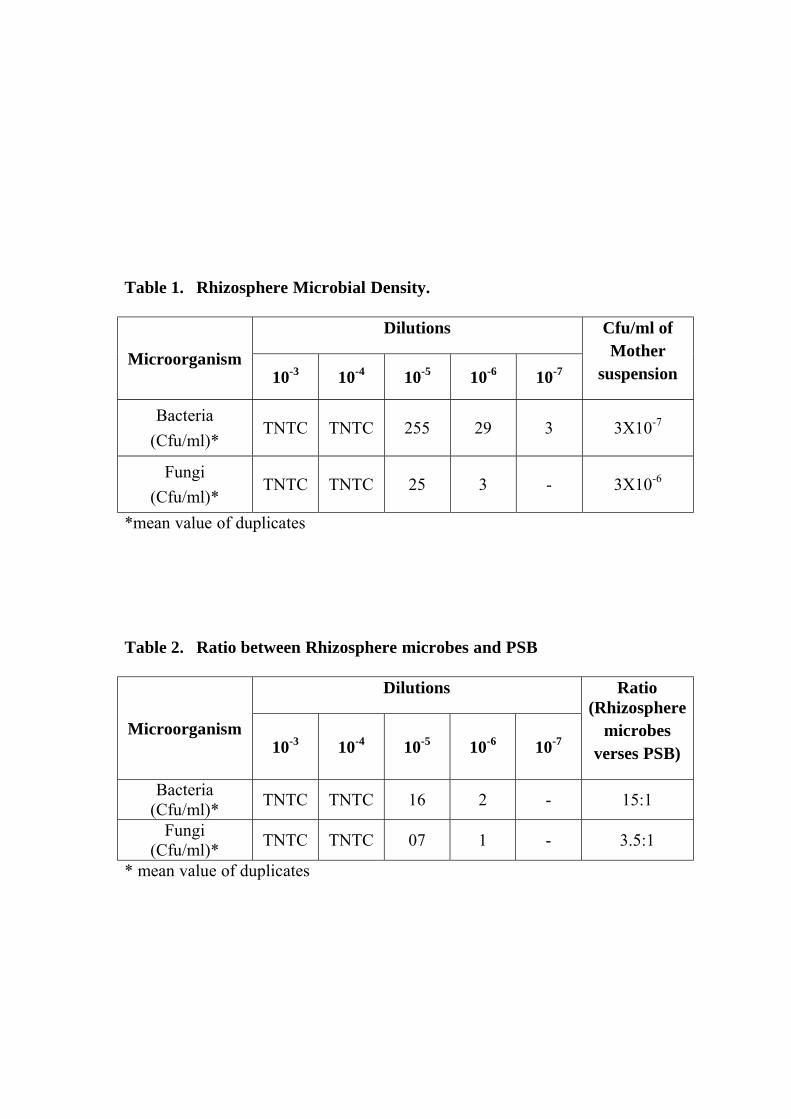
Table 1. Rhizosphere Microbial Density.
Microorganism
Dilutions Cfu/ml of Mother
suspension 10-3 10-4 10-5 10-6 10-7
Bacteria
(Cfu/ml)* TNTC TNTC 255 29 3 3X10-7
Fungi
(Cfu/ml)* TNTC TNTC 25 3 - 3X10-6
*mean value of duplicates
Table 2. Ratio between Rhizosphere microbes and PSB
Microorganism
Dilutions Ratio (Rhizosphere
microbes verses PSB) 10-3 10-4 10-5 10-6 10-7
Bacteria (Cfu/ml)*
TNTC TNTC 16 2 - 15:1
Fungi (Cfu/ml)*
TNTC TNTC 07 1 - 3.5:1
* mean value of duplicates
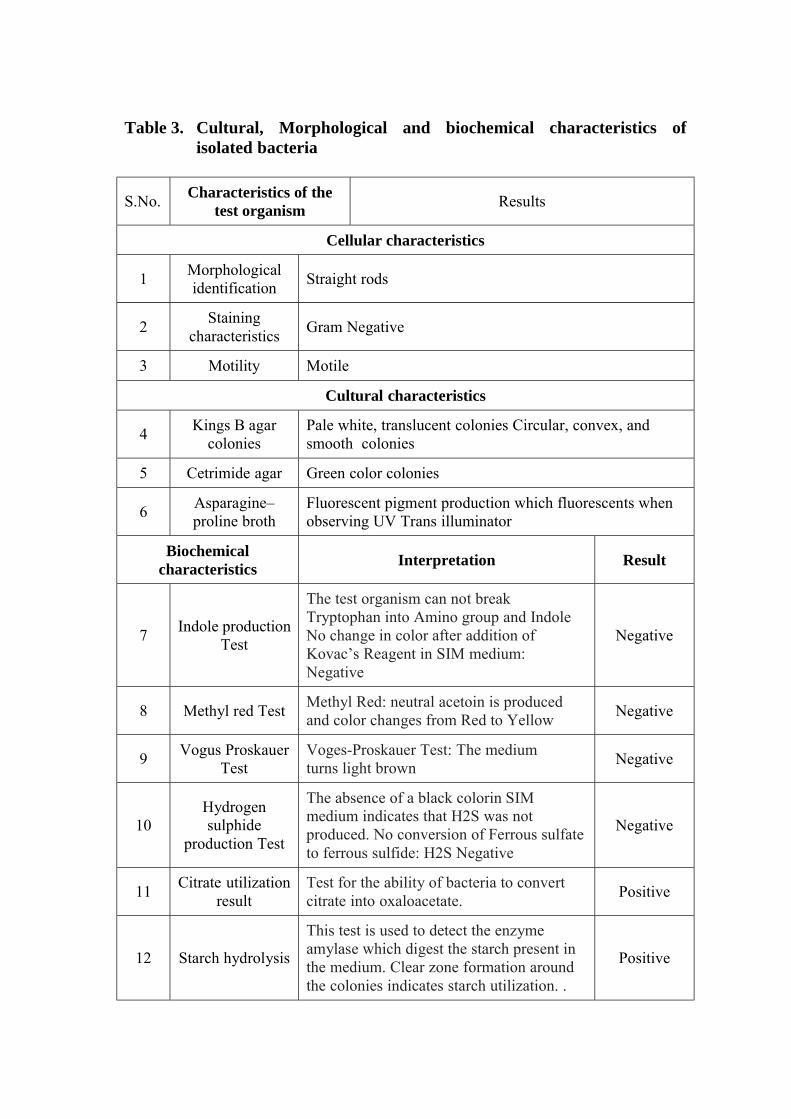
Table 3. Cultural, Morphological and biochemical characteristics of isolated bacteria
S.No. Characteristics of the
test organism Results
Cellular characteristics
1 Morphological identification
Straight rods
2 Staining
characteristics Gram Negative
3 Motility Motile
Cultural characteristics
4 Kings B agar
colonies Pale white, translucent colonies Circular, convex, and smooth colonies
5 Cetrimide agar Green color colonies
6 Asparagine–proline broth
Fluorescent pigment production which fluorescents when observing UV Trans illuminator
Biochemical characteristics
Interpretation Result
7 Indole production
Test
The test organism can not break Tryptophan into Amino group and Indole No change in color after addition of Kovac’s Reagent in SIM medium: Negative
Negative
8 Methyl red Test Methyl Red: neutral acetoin is produced and color changes from Red to Yellow
Negative
9 Vogus Proskauer
Test Voges-Proskauer Test: The medium turns light brown
Negative
10 Hydrogen sulphide
production Test
The absence of a black colorin SIM medium indicates that H2S was not produced. No conversion of Ferrous sulfate to ferrous sulfide: H2S Negative
Negative
11 Citrate utilization
result Test for the ability of bacteria to convert citrate into oxaloacetate.
Positive
12 Starch hydrolysis
This test is used to detect the enzyme amylase which digest the starch present in the medium. Clear zone formation around the colonies indicates starch utilization. .
Positive

Biochemical characteristics
Interpretation Result
13 Gelatin
liquefaction
This test is used to detect the enzyme Gelatinase which digest the Gelatin present in the medium. . Medium liquefied
Positive
14 Urease
production
This test is used to detect the enzyme Urease, which breaks down urea into ammonia. Lack of color change represent negative
Negative
15 Catalase Test
This test is used to detect the enzyme catalase. Liberation of bubbles (O2) immediately after the addition of hydrogen peroxide solution
Positive
16 Oxidase test
This test is based on detecting the production of the enzyme cytochrome oxidase. Appearance of Blue colur after immediate addition of culture in oxidase disc
Positive
17 Nitrate reduction
Test
Add one ml of sulfanilic acid to each tube, then add one ml of dimethyl 1-naphthylamine solution Development of a red color indicates nitrate has been reduced
Positive
18
Acid and gas production in carbohydrate fermentation
Medium
Test organism showed Respiration of sugars - Acid seen in upper part of the tube
showed no gas
production and acid
production found on top of the tube

Table 4. Efficacy of phosphate solubilization of UV treated Aspergillus niger
UV irradiated Aspergillus
niger strains
*‘P’ ppm of filtrate (Day) Increased % of phosphate
Solubilization
(Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
ANuv30 2.65±0.07 4.05±0.09 5.00±0.05 20.45 10.96 8.70
ANuv40 2.50±0.09 3.85±0.11 4.75±0.07 13.64 5.48 3.26
ANuv50 2.85±0.05 4.35±0.06 5.50±0.04 29.55 19.18 19.57
ANuv60 3.05±0.06 4.65±0.07 5.85±0.09 38.64 27.40 27.17
Wild type 2.20±0.05 3.65±0.1 4.60±0.04
*Mean value ± Standard Deviation of triplicates Table 5. Efficacy of phosphate solubilization of UV treated Aspergillus
fumigatus
UV irradiated Aspergillus fumigatus
strains
*‘P’ ppm of filtrate (Day) Increased % of phosphate
solubilization (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
AFuv30 3.25±0.07 3.65±0.06 3.85±0.05 10.17 12.31 5.48
AFuv40 3.15±0.12 3.65±0.05 4.15±0.07 6.78 12.31 13.70
AFuv50 3.55±0.09 3.95±0.08 4.55±0.08 20.34 21.54 24.66
AFuv60 3.35±0.05 3.85±0.08 4.45±0.06 13.56 18.46 21.92
Wild type 2.95±0.04 3.25±0.06 3.65±0.08
*Mean value ± Standard Deviation of triplicates

Table 6. Efficacy of phosphate solubilization of UV treated Penicillium Strains
UV irradiated Penicillium
Strains
*‘P’ ppm of filtrate (Day) Increased % of phosphate
solubilization (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
PEuv30 3.45±0.08 4.10±0.05 4.25±0.06 4.55 6.49 6.25
PEuv40 3.65±0.07 4.25±0.07 4.45±0.08 10.61 10.39 11.25
PEuv50 3.75±0.06 4.40±0.08 4.75±0.05 13.64 14.29 18.75
PEuv60 3.40±0.08 4.20±0.1 4.35±0.11 3.03 9.09 8.75
Wild type 3.30±0.05 3.85±0.06 4.00±0.09
*Mean value ± Standard Deviation of triplicates Table 7. Efficacy of phosphate solubilization of UV treated Pseudomonas
strains
UV treated Pseudomonas
strains
*‘P’ ppm of filtrate (Day) Increased % of
phosphate solubilization (Day)
3rd 6th 9th 3rd
(%) 6th (%) 9th (%)
PSuv1 3.85±0.09 3.95±0.07 4.15±0.08 1.32 -4.8 -12.6
PSuv2 3.95±0.08 4.10±0.04 4.30±0.03 3.95 -1.2 -9.4
PSuv3 4.15±0.07 4.25±0.06 4.45±0.05 9.21 2.41 -6.3
PSuv4 4.05±0.09 4.35±0.09 4.65±0.04 6.58 4.82 -2.1
Wild type 3.80±0.06 4.15±0.07 4.75±0.06
*Mean value ± Standard Deviation of triplicates

Table 8. Efficacy of phosphate solubilization of Sodium azide treated Aspergillus niger
Sodium
azide treated
Aspergillus niger
strains
*‘P’ ppm of filtrate (Day) Increased % of phosphate
solubilization (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
ANsa30 2.40±0.06 4.10±0.05 5.05±0.09 9.0 12.3 9.7
ANsa60 2.50±0.05 4.30±0.09 5.55±0.08 13.6 17.8 20.6
ANsa90 2.95±0.09 4.65±0.1 5.80±0.08 34.0 27.4 26.0
ANsa120 3.45±0.06 4.95±0.11 6.15±0.05 56.8 35.6 33.6
Wild type 2.20±0.05 3.65±0.1 4.60±0.04
*Mean value ± Standard Deviation of triplicates Table 9. Efficacy of phosphate solubilization of Sodium azide treated
Aspergillus fumigatus
Sodium azide
treated Aspergillus fumigatus
strains
*‘P’ ppm of filtrate (Day) Increased % of phosphate
solubilization (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
AFsa30 3.10±0.05 3.45±0.06 3.95±0.1 5.0 6.1 8.2
AFsa60 3.25±0.07 3.75±0.06 4.25±0.9 10.1 15.3 16.4
AFsa90 3.45±0.04 4.00±0.04 5.30±0.12 16.9 23.0 45.0
AFsa120 4.05±0.06 4.65±0.09 5.50±0.05 37.0 43.0 50.6
Wild type 2.95±0.04 3.25±0.06 3.65±0.08
*Mean value ± Standard Deviation of triplicates

Table 10. Efficacy of phosphate solubilization of Sodium azide treated Penicillium strains
Sodium
azide treated
Penicillium strains
*‘P’ ppm of filtrate (Day) Increased % of phosphate
solubilization (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
PEsa60 3.50±0.06 4.15±0.05 4.35±0.07 6.0 7.7 8.7
PEsa90 3.60±0.03 4.50±0.05 5.05±0.04 9.0 16.8 26.2
PEsa120 3.70±0.08 4.65±0.08 5.65±0.09 12.1 20.7 41.2
PEsa150 3.80±0.09 4.85±0.04 5.95±0.06 15.1 25.9 48.7
Wild type 3.30±0.05 3.85±0.06 4.00±0.09
*Mean value ± Standard Deviation of triplicates Table 11. Efficacy of phosphate solubilization of Sodium azide treated
Pseudomonas strains
Sodium azide treated Pseudomonas
strains
*‘P’ ppm of filtrate (Day) Increased % of
phosphate solubilization (Day)
3rd 6th 9th 3rd
(%) 6th (%) 9th (%)
PSsa30 2.85±0.05 3.65±0.1 4.15±0.07 25.00 -12.0 -12.6
PSsa60 3.15±0.07 3.95±0.08 4.45±0.09 -17.1 -4.8 -6.3
PSsa90 3.35±0.05 4.05±0.05 4.50±0.04 -11.8 -2.4 -5.2
PSsa120 3.45±0.08 4.25±0.08 4.65±0.06 -9.2 2.41 -2.1
Wild type 3.80±0.06 4.15±0.07 4.75±0.06
*Mean value ± Standard Deviation of triplicates

Table 12. Efficacy of phosphate solubilization of Ethyl Methane Sulphonate (EMS) Aspergillus niger
EMS
treated Aspergillus
niger strains
*‘P’ ppm of filtrate (Day) Increased % of phosphate
solubilization (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
ANems30 2.30±0.07 3.90±0.06 4.75±0.06 4.5 6.8 3.2
ANems60 2.50±0.05 4.25±0.05 5.40±0.04 13.6 16.4 17.3
ANems90 2.65±0.09 4.45±0.07 5.65±0.06 20.4 21.9 22.8
ANems120 2.85±0.09 4.75±0.07 6.25±0.08 29.5 30.1 35.8
Wild type 2.20±0.05 3.65±0.1 4.60±0.04
*Mean value ± Standard Deviation of triplicates
Table 13. Efficacy of phosphate solubilization of Ethyl Methane Sulphonate (EMS) Aspergillus fumigatus
EMS
treated Aspergillus fumigatus
strains
*‘P’ ppm of filtrate (Day) Increased % of phosphate
solubilization (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
AFems30 3.25±0.03 3.45±0.09 3.85±0.05 10.1 6.1 5.4
AFems60 3.55±0.07 3.80±0.06 4.15±0.06 20.3 16.9 13.6
AFems90 3.75±0.05 4.15±0.06 4.65±0.09 27.1 27.6 27.3
AFems120 3.95±0.08 4.45±0.07 5.15±0.08 33.8 36.9 41.0
Wild type 2.95±0.04 3.25±0.06 3.65±0.08
*Mean value ± Standard Deviation of triplicates

Table 14. Efficacy of phosphate solubilization of Ethyl Methane Sulphonate (EMS) treated Penicillium strains
EMS treated
Penicillium strains
*‘P’ ppm of filtrate (Day) Increased % of phosphate
solubilization (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
PEems60 3.65±0.11 4.70±0.07 5.15±0.06 10.6 22.0 28.7
PEems90 4.05±0.07 4.90±0.1 5.30±0.08 22.7 27.2 32.5
PEems120 4.55±0.09 5.10±0.1 5.45±0.05 37.8 32.4 36.2
PEems150 4.80±0.06 5.70±0.09 5.95±0.04 45.4 48.0 48.7
Wild type 3.30±0.05 3.85±0.06 4.00±0.09
*Mean value ± Standard Deviation of triplicates
Table 15. Efficacy of phosphate solubilization of Ethyl Methane Sulphonate (EMS) treated Pseudomonas strains
EMS treated Pseudomonas
Strains
*‘P’ ppm of filtrate (Day) Increased % of
phosphate solubilization (Day)
3rd 6th 9th 3rd
(%) 6th (%) 9th (%)
PSems60 2.30±0.08 2.80±0.04 3.30±0.07 -39.4 -32.5 -30.5
PSems90 2.35±0.06 2.90±0.05 3.55±0.09 -38.1 -30.0 -25.2
PSems120 2.40±0.08 3.05±0.05 4.15±0.06 -36.8 -26.5 -12.6
PSems150 2.45±0.09 3.25±0.08 4.25±0.05 -35.5 -21.6 -10.5
Wild type 3.80±0.06 4.15±0.07 4.75±0.06
*Mean value ± Standard Deviation of triplicates

Table 16. Efficacy of phosphate solubilization of Aspergillus niger (ANems120) grown on different Carbon sources
Carbon sources vs ANems120 Strains
*‘P’ ppm of filtrate (Day)
3rd 6th 9th
Glucose 2.85±0.09 4.75±0.07 6.25±0.08
Sucrose 2.40±0.05 3.75±0.08 4.85±0.06
Mannitol 2.05±0.04 3.05±0.06 4.35±0.09
Lactose 2.20±0.08 3.60±0.06 4.60±0.06
*Mean value ± Standard Deviation of triplicates; Table 17. Efficacy of phosphate solubilization of Aspergillus niger
(ANems120) grown on different Nitrogen sources
Nitrogen sources vs ANems120 strains
*‘P’ ppm of filtrate (Day)
3rd 6th 9th
Ammonium sulphate 2.85±0.09 4.75±0.07 6.25±0.08
Sodium nitrate 1.90±0.05 2.65±0.09 3.20±0.05
Potassium nitrate 2.35±0.07 3.70±0.09 4.70±0.08
Urea 2.40±0.04 3.70±0.04 4.75±0.05
*Mean value ± Standard Deviation of triplicates; Table 18. Efficacy of phosphate solubilization of Aspergillus niger
(ANsa120) grown on different Carbon sources
Carbon sources vs ANsa120 Strains
*‘P’ ppm of filtrate (Day)
3rd 6th 9th
Glucose 3.45±0.06 4.95±0.11 6.15±0.05
Sucrose 3.55±0.09 4.55±0.04 4.95±0.06
Mannitol 1.85±0.1 2.15±0.08 2.90±0.07
Lactose 3.05±0.06 3.55±0.07 3.95±0.09
*Mean value ± Standard Deviation of triplicates;

Table 19. Efficacy of phosphate solubilization of Aspergillus niger (ANsa120) grown on different Nitrogen sources
Nitrogen sources vs AFsa120 Strains
*‘P’ ppm of filtrate (Day)
3rd 6th 9th
Ammonium sulphate 3.45±0.06 4.95±0.11 6.15±0.05
Sodium nitrate 1.75±0.09 2.30±0.1 2.75±0.05
Potassium nitrate 2.40±0.05 3.70±0.06 4.65±0.07
Urea 2.45±0.06 3.85±0.08 4.70±0.05
*Mean value ± Standard Deviation of triplicates; Table 20. Efficacy of phosphate solubilization of Penicillium sp.(PEsa150)
grown on different Carbon sources
Carbon sources PEsa150 *‘P’ ppm of filtrate (Day)
3rd 6th 9th
Glucose 3.80±0.09 4.85±0.04 5.95±0.06
Sucrose 3..35±0.1 3.90±0.06 4.60±0.05
Mannitol 3.05±0.11 3.75±0.05 4.25±0.04
Lactose 3.15±0.06 3.80±0.07 4.35±0.07
*Mean value ± Standard Deviation of triplicates;
Table 21. Efficacy of phosphate solubilization of Penicillium sp.(PEsa150) grown on different Nitrogen sources
Nitrogen sources PEsa150 *‘P’ ppm of filtrate (Day)
3rd 6th 9th
Ammonium sulphate 3.80±0.09 4.85±0.04 5.95±0.06
Sodium nitrate 3.30±0.07 3.75±0.05 4.05±0.06
Potassium nitrate 3.35±0.04 3.90±0.07 4.60±0.05
Urea 3.70±0.06 3.95±0.04 4.70±0.08
*Mean value ± Standard Deviation of triplicates;

Table 22. Efficacy of phosphatase activity of UV treated Aspergillus niger
UV irradiated Aspergillus
niger strains
*Phosphatase activity µmol min-1 (Day)
Increased % of phosphatase activity (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
ANuv30 0.121±0.04 0.223±0.05 0.345±0.04 24.74 16.15 13.86
ANuv40 0.103±0.02 0.218±0.04 0.341±0.03 6.19 13.54 12.54
ANuv50 0.134±0.03 0.260±0.05 0.399±0.05 38.14 35.42 31.68
ANuv60 0.165±0.02 0.275±0.05 0.424±0.06 70.10 43.23 39.93
Wild type 0.097±0.05 0.192±0.03 0.303±0.04
*Mean value ± Standard Deviation of triplicates Table 23. Efficacy of phosphatase activity of fungal strains UV treated
Aspergillus fumigatus
UV irradiated Aspergillus fumigatus
strains
*Phosphatase activity µmol min-1 (Day)
Increased % of phosphatase activity (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
AFuv30 0.171±0.05 0.203±0.06 0.221±0.06 9.62 14.04 13.86
AFuv40 0.160±0.05 0.205±0.05 0.232±0.08 2.56 15.17 12.54
AFuv50 0.195±0.04 0.227±0.07 0.269±0.04 25.00 27.53 31.68
AFuv60 0.173±0.05 0.224±0.04 0.265±0.03 10.90 25.84 39.93
Wild type 0.156±0.04 0.178±0.04 0.197±0.06
*Mean value ± Standard Deviation of triplicates

Table 24. Efficacy of phosphatase activity of Penicillium strains
UV irradiated Penicillium
strains
*Phosphatase activity µmol min-1 (Day)
Increased % of phosphatase activity (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
PEuv30 0.180±0.04 0.228±0.06 0.239±0.06 6.51 7.04 1.27
PEuv40 0.195±0.05 0.245±0.08 0.264±0.04 15.38 15.02 11.86
PEuv50 0.209±0.05 0.237±0.07 0.267±0.06 23.67 11.27 13.14
PEuv60 0.178±0.05 0.234±0.04 0.257±0.05 5.33 9.86 8.90
Wild type 0.169±0.04 0.213±0.06 0.236±0.06
*Mean value ± Standard Deviation of triplicates Table 25. Efficacy of phosphatase activity of UV treated Pseudomonas sp.
UV treated Pseudomonas
strains
*Phosphatase activity µmol min-1 (Day)
Increased % of phosphatase activity (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
PSuv1 0.205±0.04 0.225±0.06 0.231±0.05 2.5 -3.85 -19.51
PSuv2 0.215±0.04 0.227±0.07 0.236±0.05 7.5 -2.95 -17.77
PSuv3 0.227±0.07 0.233±0.04 0.275±0.06 13.5 -0.43 -4.18
PSuv4 0.210±0.03 0.226±0.06 0.286±0.05 5.0 -3.42 -0.35
Wild type 0.200±0.05 0.234±0.05 0.287±0.07
*Mean value ± Standard Deviation of triplicates
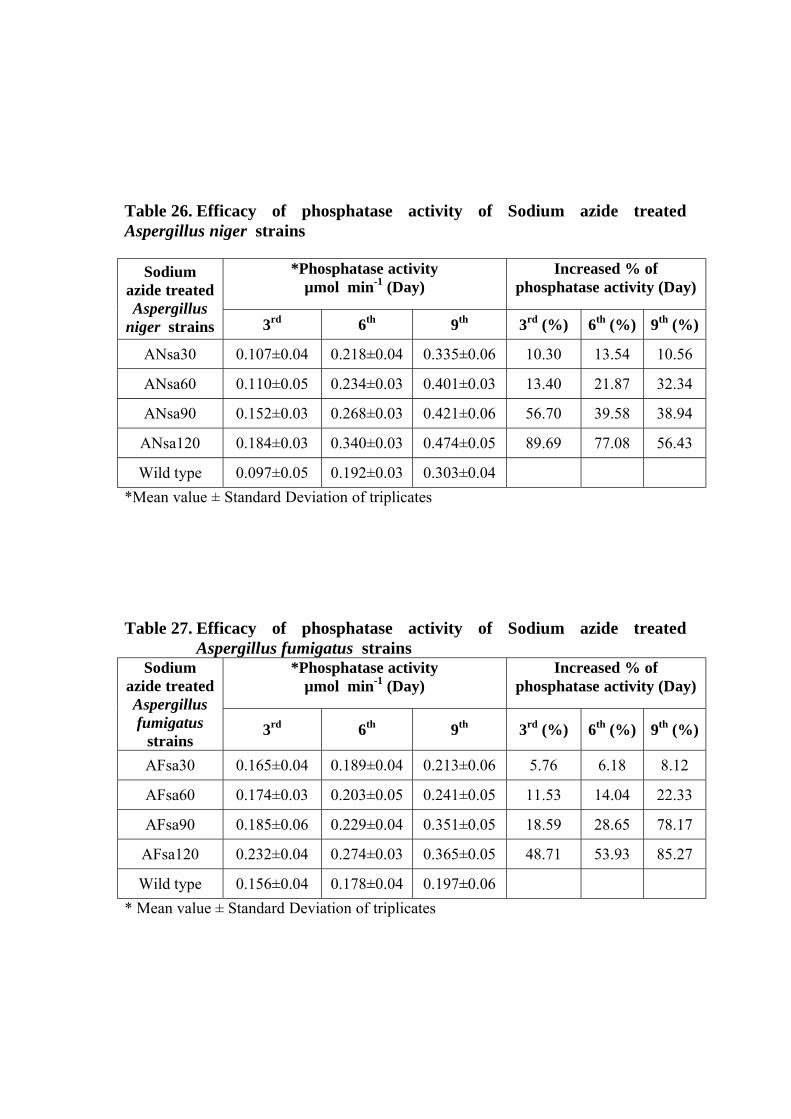
Table 26. Efficacy of phosphatase activity of Sodium azide treated Aspergillus niger strains
Sodium azide treated Aspergillus
niger strains
*Phosphatase activity µmol min-1 (Day)
Increased % of phosphatase activity (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
ANsa30 0.107±0.04 0.218±0.04 0.335±0.06 10.30 13.54 10.56
ANsa60 0.110±0.05 0.234±0.03 0.401±0.03 13.40 21.87 32.34
ANsa90 0.152±0.03 0.268±0.03 0.421±0.06 56.70 39.58 38.94
ANsa120 0.184±0.03 0.340±0.03 0.474±0.05 89.69 77.08 56.43
Wild type 0.097±0.05 0.192±0.03 0.303±0.04
*Mean value ± Standard Deviation of triplicates Table 27. Efficacy of phosphatase activity of Sodium azide treated
Aspergillus fumigatus strains Sodium
azide treated Aspergillus fumigatus
strains
*Phosphatase activity µmol min-1 (Day)
Increased % of phosphatase activity (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
AFsa30 0.165±0.04 0.189±0.04 0.213±0.06 5.76 6.18 8.12
AFsa60 0.174±0.03 0.203±0.05 0.241±0.05 11.53 14.04 22.33
AFsa90 0.185±0.06 0.229±0.04 0.351±0.05 18.59 28.65 78.17
AFsa120 0.232±0.04 0.274±0.03 0.365±0.05 48.71 53.93 85.27
Wild type 0.156±0.04 0.178±0.04 0.197±0.06
* Mean value ± Standard Deviation of triplicates

Table 28. Efficacy of phosphatase activity of Sodium azide treated Penicillium strains
Sodium azide treated Penicillium
strains
*Phosphatase activity µmol min-1 (Day)
Increased % of phosphatase activity (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
PEsa60 0.181±0.06 0.233±0.05 0.261±0.04 7.10 9.39 10.59
PEsa90 0.191±0.04 0.249±0.04 0.349±0.04 13.01 16.90 47.88
PEsa120 0.200±0.05 0.261±0.05 0.400±0.06 18.34 22.53 69.49
PEsa150 0.210±0.04 0.276±0.06 0.420±0.08 24.26 29.57 77.96
Wild type 0.169±0.04 0.213±0.06 0.236±0.06
*Mean value ± Standard Deviation of triplicates Table 29. Efficacy of phosphatase activity of Sodium azide treated
Pseudomonas strains
Sodium azide treated Pseudomonas
strains
*Phosphatase activity µmol min-1 (Day)
Increased % of phosphatase activity (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
PSsa30 0.169±0.06 0.199±0.07 0.224±0.03 -15.5 9.39 10.59
PSsa60 0.171±0.04 0.220±0.06 0.256±0.06 -14.5 16.90 47.88
PSsa90 0.180±0.05 0.232±0.07 0.260±0.04 -10.0 22.53 69.49
PSsa120 0.185±0.06 0.241±0.03 0.275±0.04 -7.5 29.57 77.96
Wild type 0.200±0.05 0.234±0.05 0.287±0.07
*Mean value ± Standard Deviation of triplicates

Table 30. Efficacy of phosphatase activity of Ethyl Methane Sulphonate (EMS) Aspergillus niger
EMS treated Aspergillus
niger strains
*Phosphatase activity µmol min-1 (Day)
Increased % of phosphatase activity (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
ANems30 0.099± 0.210± 0.316± 2.06 9.37 4.29
ANems60 0.104±0.03 0.222±0.05 0.365±0.04 7.21 15.62 20.46
ANems90 0.125±0.04 0.253±0.06 0.412±0.03 28.86 31.77 35.97
ANems120 0.131±0.05 0.273±0.05 0.452±0.07 35.05 42.18 49.17
Wild type 0.097±0.05 0.192±0.03 0.303±0.04
*Mean value ± Standard Deviation of triplicates
Table 31. Efficacy of phosphatase activity of Ethyl Methane Sulphonate (EMS) treated Aspergillus fumigatus
EMS treated Aspergillus
niger strains
*Phosphatase activity µmol min-1 (Day)
Increased % of phosphatase activity (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
AFems30 0.175±0.04 0.197±0.06 0.214±0.05 12.17 10.67 8.62
AFems60 0.189±0.05 0.212±0.04 0.231±0.07 21.15 19.10 17.25
AFems90 0.206±0.05 0.236±0.07 0.261±0.05 32.05 32.58 32.48
AFems120 0.219±0.05 0.258±0.04 0.318±0.04 40.38 44.94 61.42
Wild type 0.156±0.04 0.178±0.04 0.197±0.06
*Mean value ± Standard Deviation of triplicates

Table 32. Efficacy of phosphatase activity of Ethyl Methane Sulphonate (EMS) treated Penicillium strains
EMS treated Penicillium
strains
*Phosphatase activity µmol min-1 (Day)
Increased % of phosphatase activity (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
PEems60 0.197±0.05 0.275±0.05 0.370±0.05 16.56 29.10 56.78
PEems90 0.227±0.05 0.325±0.05 0.369±0.05 34.32 52.58 56.35
PEems120 0.245±0.06 0.324±0.03 0.371±0.05 44.97 52.11 57.20
PEems150 0.278±0.04 0.381±0.07 0.425±0.07 64.49 78.87 80.08
Wild type 0.169±0.04 0.213±0.06 0.236±0.06
*Mean value ± Standard Deviation of triplicates
Table 33. Efficacy of phosphatase activity Ethyl Methane Sulphonate (EMS)
treated Pseudomonas strains
EMS treated Pseudomonas
strains
*Phosphatase activity µmol min-1 (Day)
Increased % of phosphatase activity (Day)
3rd 6th 9th 3rd (%) 6th (%) 9th (%)
PSems60 0.091±0.05 0.150±0.07 0.180±0.06 -54.5 29.10 56.78
PSems90 0.095±0.05 0.161±0.05 0.191±0.05 -52.5 52.58 56.35
PSems120 0.100±0.03 0.167±0.04 0.220±0.05 -50.0 52.11 57.20
PSems150 0.113±0.04 0.175±0.08 0.229±0.06 -43.5 78.87 80.08
Wild type 0.200±0.05 0.234±0.05 0.287±0.07
*Mean value ± Standard Deviation of triplicates
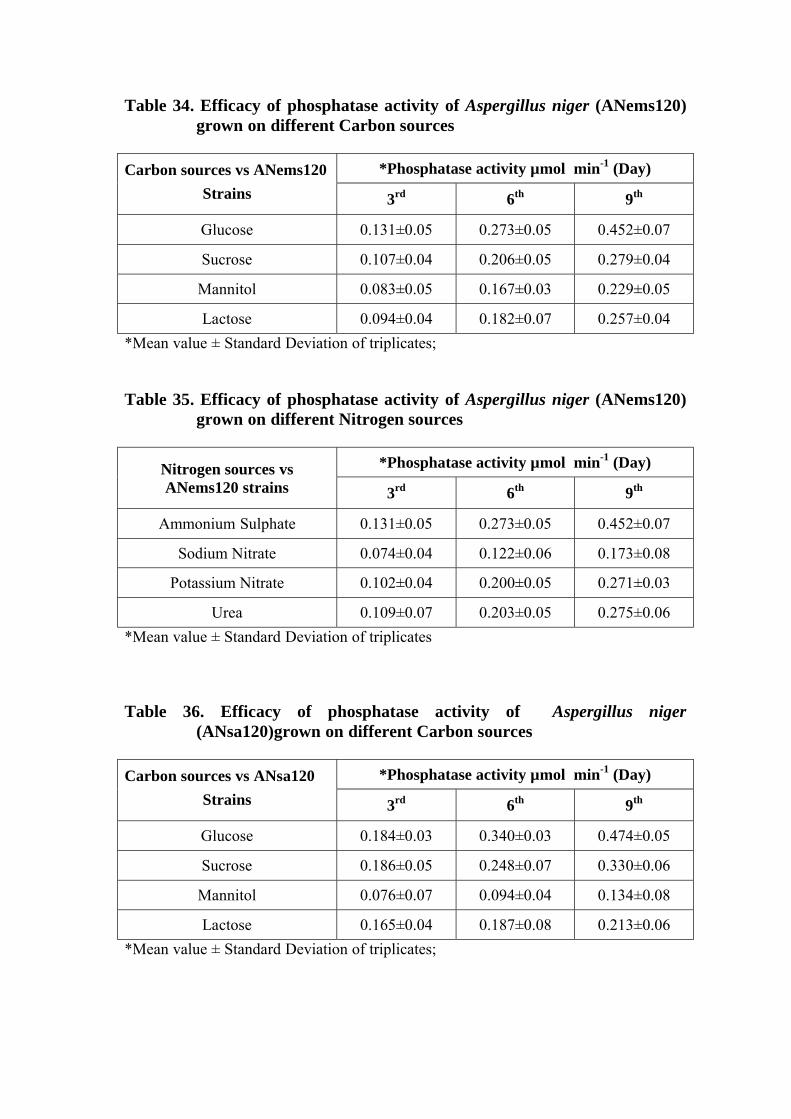
Table 34. Efficacy of phosphatase activity of Aspergillus niger (ANems120) grown on different Carbon sources
Carbon sources vs ANems120 Strains
*Phosphatase activity µmol min-1 (Day)
3rd 6th 9th
Glucose 0.131±0.05 0.273±0.05 0.452±0.07
Sucrose 0.107±0.04 0.206±0.05 0.279±0.04
Mannitol 0.083±0.05 0.167±0.03 0.229±0.05
Lactose 0.094±0.04 0.182±0.07 0.257±0.04
*Mean value ± Standard Deviation of triplicates; Table 35. Efficacy of phosphatase activity of Aspergillus niger (ANems120)
grown on different Nitrogen sources
Nitrogen sources vs ANems120 strains
*Phosphatase activity µmol min-1 (Day)
3rd 6th 9th
Ammonium Sulphate 0.131±0.05 0.273±0.05 0.452±0.07
Sodium Nitrate 0.074±0.04 0.122±0.06 0.173±0.08
Potassium Nitrate 0.102±0.04 0.200±0.05 0.271±0.03
Urea 0.109±0.07 0.203±0.05 0.275±0.06
*Mean value ± Standard Deviation of triplicates Table 36. Efficacy of phosphatase activity of Aspergillus niger
(ANsa120)grown on different Carbon sources
Carbon sources vs ANsa120 Strains
*Phosphatase activity µmol min-1 (Day)
3rd 6th 9th
Glucose 0.184±0.03 0.340±0.03 0.474±0.05
Sucrose 0.186±0.05 0.248±0.07 0.330±0.06
Mannitol 0.076±0.07 0.094±0.04 0.134±0.08
Lactose 0.165±0.04 0.187±0.08 0.213±0.06
*Mean value ± Standard Deviation of triplicates;

Table 37. Efficacy of phosphatase activity of Aspergillus niger (ANsa120) grown on different Nitrogen sources
Nitrogen sources vs ANsa120 Strains
*Phosphatase activity µmol min-1 (Day)
3rd 6th 9th
Ammonium Sulphate 0.184±0.03 0.340±0.03 0.474±0.05
Sodium Nitrate 0.08±0.02 0.103±0.06 0.125±0.07
Potassium Nitrate 0.108±0.08 0.203±0.08 0.254±0.04
Urea 0.112±0.07 0.216±0.07 0.273±0.06
*Mean value ± Standard Deviation of triplicates
Table 38. Efficacy of phosphatase activity of Penicillium sp.(PEsa150)
grown on different Carbon sources
Carbon sources PEsa150 *Phosphatase activity µmol min-1 (Day)
3rd 6th 9th
Glucose 0.210±0.04 0.276±0.06 0.420±0.08
Sucrose 0.171±0.08 0.288±0.06 0.342±0.03
Mannitol 0.162±0.04 0.204±0.04 0.261±0.04
Lactose 0.168±0.05 0.214±0.06 0.253±0.04
*Mean value ± Standard Deviation of triplicates Table 39. Efficacy of phosphatase activity of Penicillium sp.(PEsa150)
grown on different Nitrogen sources
Nitrogen sources PEsa150 *Phosphatase activity µmol min-1 (Day)
3rd 6th 9th
Ammonium Sulphate 0.210±0.04 0.276±0.06 0.420±0.08
Sodium Nitrate 0.163±0.05 0.209±0.03 0.229±0.05
Potassium Nitrate 0.165±0.04 0.218±0.04 0.261±0.05
Urea 0.205±0.05 0.223±0.07 0.270±0.03
*Mean value ± Standard Deviation of triplicates

Table 40. Efficacy of Lipase activity of UV treated Aspergillus niger
UV treated Aspergillus niger strains
Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of Lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
ANuv30 1.86±0.08 0.49±0.04 14.81 4.26
ANuv40 1.95±0.06 0.62±0.08 20.37 31.91
ANuv50 2.15±0.05 0.90±0.06 32.72 91.49
ANuv60 1.89±0.05 0.60±0.04 16.66 27.66
Wild type 1.62±0.07 0.47±0.04
*Mean value ± Standard Deviation of triplicates
Table 41. Efficacy of Lipase activity of UV treated Aspergillus fumigatus
UV treated Aspergillus fumigatus strains
Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of Lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
AFuv30 1.22±0.09 0.33±0.04 20.79 22.22
AFuv40 1.39±0.08 0.38±0.04 37.62 40.74
AFuv50 2.09±0.09 0.60±0.04 106.93 122.22
AFuv60 1.75±0.06 0.56±0.07 73.26 107.40
Wild type 1.01±0.07 0.27±0.04
*Mean value ± Standard Deviation of triplicates

Table 42. Efficacy of Lipase activity of UV treated Penicillium strains
UV treated Pencillum strains
Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of Lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
PEuv30 1.28±0.06 0.48±0.05 6.67 4.35
PEuv40 1.43±0.06 0.73±0.04 19.17 58.70
PEuv50 1.62±0.08 0.69±0.06 35.00 50.00
PEuv60 1.69±0.08 0.85±0.04 40.83 84.78
Wild type 1.20±0.10 0.46±0.04
*Mean value ± Standard Deviation of triplicates
Table 43. Efficacy of lipase activity of UV treated Pseudomonas strains
UV treated Pseudomonas strains
Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of Lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
PSuv1 2.22±0.01 0.23±0.02 5.71 8.70
PSuv2 2.19±0.09 0.33±0.08 4.28 26.09
PSuv3 2.49±0.07 0.38±0.03 18.57 36.96
PSuv4 2.29±0.02 0.34±0.05 9.04 67.39
Wild type 2.10±0.08 0.17±0.06
*Mean value ± Standard Deviation of triplicates

Table 44. Efficacy of lipase activity of Sodium azide treated Aspergillus niger
Sodium azide treated Aspergillus niger
strains
Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of Lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
ANsa30 1.90±0.08 0.51±0.07 17.28 8.51
ANsa60 2.09±0.05 0.68±0.05 29.01 44.68
ANsa90 2.61±0.07 0.98±0.04 61.11 108.61
ANsa120 1.85±0.06 0.63±0.03 14.19 34.04
Wild type 1.62±0.07 0.47±0.04
*Mean value ± Standard Deviation of triplicates±
Table 45. Efficacy of lipase activity of Sodium azide treated Aspergillus
fumigatus
Sodium azide treated Aspergillus fumigatus
strains
Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of Lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
AFsa30 1.45±0.05 0.41±0.04 43.56 51.85
AFsa60 1.25±0.05 0.38±0.06 23.76 40.74
AFsa90 1.22±0.07 0.30±0.03 20.79 11.11
AFsa120 1.60±0.07 0.44±0.07 58.42 62.96
Wild type 1.01±0.07 0.27±0.04
*Mean value ± Standard Deviation of triplicates±

Table 46. Efficacy of lipase activity of Sodium azide treated Penicillium strains
Sodium azide treated Pencillilum strains
Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of Lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
PEsa30 1.56±0.06 0.48±0.04 30.00 4.35
PEsa60 1.43±0.04 0.69±0.06 19.17 50.00
PEsa90 1.76±0.08 0.73±0.04 46.67 58.70
PEsa120 2.01±0.07 0.99±0.03 67.50 115.22
Wild type 1.20±0.10 0.46±0.04
*Mean value ± Standard Deviation of triplicates
Table 47. Efficacy of lipase activity of Sodium azide treated Pseudomonas
strains
Sodium azide treated Pencillilum strains
Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of Lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
PSsa30 2.28±0.04 0.28±0.11 8.57 64.71
PSsa60 2.93±0.03 0.35±0.02 39.52 105.88
PSsa90 3.23±0.05 0.39±0.08 53.80 129.41
PSsa120 3.04±0.09 0.39±0.04 44.76 129.41
Wild type 2.10±0.06 0.17±0.06
*Mean value ± Standard Deviation of triplicates

Table 48. Efficacy of lipase activity of Ethyl Methane Sulphonate (EMS) treated Aspergillus niger
EMS treated Aspergillus niger
strains
Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of Lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
ANems60 1.90±0.09 0.62±0.06 17.28 31.91
ANems90 2.09±0.10 0.77±0.07 29.01 63.83
ANems120 3.10±0.08 0.87±0.05 91.35 85.11
ANems150 3.85±0.07 0.94±0.03 137.65 100.00
Wild type 1.62±0.07 0.47±0.04
*Mean value ± Standard Deviation of triplicates
Table 49. Efficacy of lipase activity of Ethyl Methane Sulphonate (EMS) treated Aspergillus fumigatus
EMS treated Aspergillus fumigatus
strains
Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of Lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
AFems60 1.85±0.05 0.33±0.05 83.17 22.22
AFems90 2.26±0.09 0.40±0.04 123.76 48.15
AFems120 2.76±0.08 0.45±0.06 173.26 66.17
AFems150 2.60±0.09 0.41±0.03 157.43 51.85
Wild type 1.01±0.07 0.27±0.04
*Mean value ± Standard Deviation of triplicates

Table 50. Efficacy of lipase activity of Ethyl Methane Sulphonate (EMS) treated Penicillium strains
EMS treated Penicillium strains
Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of Lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
PEems60 1.64±0.06 0.50±0.03 36.67 8.70
PEems90 1.79±0.05 0.58±0.03 49.17 26.09
PEems120 1.89±0.04 0.63±0.04 57.50 36.96
PEems150 1.97±0.08 0.77±0.06 64.17 67.39
Wild type 1.20±0.10 0.46±0.04
*Mean value ± Standard Deviation of triplicates
Table 51. Efficacy of lipase activity of Ethyl Methane Sulphonate (EMS) treated Pseudomonas strains
EMS treated Penicillium strains
Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of Lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
PSems60 2.42±0.10 0.34±0.06 9.50 54.55
PSems90 3.53±0.05 0.47±0.09 59.72 113.64
PSems120 4.45±0.07 0.61±0.05 101.36 177.27
PSems150 4.12±0.04 0.52±0.05 86.43 136.36
Wild type 2.21±0.09 0.22±0.12
*Mean value ± Standard Deviation of triplicates

Table 52. Efficacy of lipase activity of fungal strains grown on different Carbon sources
Carbon sources vs ANems120
Strain
Lipase activity
after 96 hrs (Unit/g of
substrate)*
Carbon sources vs AFems120
Strain
Lipase activity
after 96 hrs (Unit/g of
substrate)*
Carbon sources vs ANems150
Strain
Lipase activity
after 96 hrs (Unit/g of
substrate)*
Glucose 2.94±0.06 Glucose 2.54±0.05 Glucose 2.44±0.03
Sucrose 3.10±0.08 Sucrose 2.76±0.08 Sucrose 2.61±0.07
Mannitol 1.85±0.07 Mannitol 1.69±0.07 Mannitol 1.98±0.07
Lactose 1.65±0.09 Lactose 1.25±0.07 Lactose 1.34±0.09
*Mean value ± Standard Deviation of triplicates Table 53. Efficacy of lipase activity of Bacterial strains grown on different
Carbon sources
Carbon sources vs PSems120 Strain
Lipase activity after 96 hrs
(Unit/g of substrate)*
Carbon sources vs PSems150 Strain
Lipase activity after 96 hrs
(Unit/g of substrate)*
Glucose 2.95±0.07 Glucose 2.45±0.08
Sucrose 4.12±0.04 Sucrose 4.12±0.04
Mannitol 2.74±0.06 Mannitol 2.05±0.04
Lactose 1.80±0.04 Lactose 1.65±0..05
*Mean value ± Standard Deviation of triplicates

Table 54. Efficacy of lipase activity of fungal strains grown on different Nitrogen sources
Nitrogen sources vs ANems120
Strain
Lipase activity
after 96 hrs
(Unit/g of substrate)*
Nitrogen sources vs AFems120
Strain
Lipase activity
after 96 hrs
(Unit/g of substrate)*
Nitrogen sources vs ANems150
Strain
Lipase activity
after 96 hrs
(Unit/g of substrate)*
Sodium nitrate
3.10±0.08 Sodium nitrate
2.76±0.08 Sodium nitrate
2.61±0.07
Potassium nitrate
2.85±0.05 Potassium nitrate
2.67±0.09 Potassium nitrate
2.24±0.06
Ammonium sulphate
2.01±0.05 Ammonium sulphate
2.13±0.10 Ammonium sulphate
1.98±0.08
Urea 1.55±0.07 Urea 1.54±0.06 Urea 1.27±0.08
*Mean value ± Standard Deviation of triplicates Table 55. Efficacy of lipase activity of Bacterial strains grown on different
Nitrogen sources
Carbon sources vs PSems120 Strain
Lipase activity after 96 hrs
(Unit/g of substrate)*
Carbon sources vs PSems150 Strain
Lipase activity after 96 hrs
(Unit/g of substrate)*
Sodium nitrate 4.12±0.04 Sodium nitrate 4.12±0.04
Potassium nitrate 2.94±0.07 Potassium nitrate 2.99±0.05
Ammonium sulphate 2.71±0.09 Ammonium sulphate 2.13±0.06
Urea 1.45±0.06 Urea 1.54±0.04
*Mean value ± Standard Deviation of triplicates

Table 56. Screening for IAA by Wild and Mutated Bacterial and Fungal strains
Wild and Mutated strains of
Aspergillus niger
IAA activity
Wild and Mutated strains of
Aspergillus fumigatus
IAA activity
Wild and Mutated strains of
Penicillium sp
IAA activity
Wild and Mutated strains of
Pseudomonas sp
IAA activity
Wild type + Wild type - Wild type + Wild type +
ANuv30 + AFuv30 - PEuv30 + PSuv1 +
ANuv40 + AFuv40 - PEuv40 + PSuv2 +
ANuv50 + AFuv50 - PEuv50 + PSuv3 +
ANuv60 + AFuv60 - PEuv60 + PSuv4 +
ANsa30 + AFsa30 - PEsa60 + PSsa30 +
ANsa60 + AFsa60 - PEsa90 + PSsa60 +
ANsa90 + AFsa90 - PEsa120 + PSsa90 +
ANsa120 + AFsa120 - PEsa150 + PSsa120 +
ANems30 - AFems30 - PEems60 + PSems60 +
ANems60 - AFems60 - PEems90 + PSems90 +
ANems90 - AFems90 - PEems120 + PSems120 +
ANems120 - AFems120 - PEems150 + PSems150 +
Key: + activity; - No activity;

Table 57. Quantification study of IAA by Wild and Mutated Bacterial and Fungal strains
Wild and Mutated strains of
Aspergillus niger
*IAA activity (µg/L)
Wild and Mutated strains of
Penicillium sp
*IAA activity (µg/L)
Wild and Mutated strains of
Pseudomonas sp
*IAA activity (µg/L)
Wild type 12.33±0.12 Wild type 7.68±0.14 Wild type 7.50±0.17
ANuv30 13.01±0.15 PEuv30 8.67±0.17 PSuv1 6.50±0.18
ANuv40 12.47±0.15 PEuv40 7.55±0.18 PSuv2 7.55±0.17
ANuv50 12.33±0.17 PEuv50 7.07±0.11 PSuv3 8.60±0.18
ANuv60 11.85±0.13 PEuv60 6.83±0.17 PSuv4 8.08±0.15
ANsa30 12.75±0.10 PEsa60 8.88±0.15 PSsa30 8.08±0.19
ANsa60 15.65±0.19 PEsa90 9.75±0.19 PSsa60 8.23±0.15
ANsa90 16.73±0.13 PEsa120 10.64±0.12 PSsa90 8.00±0.17
ANsa120 17.53±0.15 PEsa150 8.83±0.10 PSsa120 6.75±0.11
PEems60 8.35±0.11 PSems60 7.07±0.18
PEems90 7.45±0.09 PSems90 7.09±0.17
PEems120 6.97±0.18 PSems120 8.35±0.19
PEems150 6.35±0.14 PSems150 8.45±0.14
*Mean value ± Standard Deviation of triplicates

Plate 1
Biochemical Characterization of Bacterial culture
A. Growth on Cetrimide agar B. Growth on Nutrient agar
C. Asparagine -Proline broth culture tube showed fluorescence in UV-trans illuminator (Magnified image –Right)
D. Oxidase test ( Positive – Left Disc and Control – Right disc)

Plate 2 Growth of Fungal colonies on Sabouraud agar plates
Aspergillus niger
Penicillium sp.
Aspergillus fumigatus

Plate 3 Phosphate solubilization of Bacterial and Fungal cultures in Pikovskaya agar
A. Control
B. Phosphate solubilization by Penicillium sp.
C. Phosphate solubilization by Aspergillus niger
D. Phosphate solubilization by
Aspergillus fumigatus
E. Phosphate solubilization by Pseudomonas sp.

Plate 4 Phosphate solubilization and phosphatase production by fungal cultures in
Pikovskaya broth
Phosphate solubilization by Penicillium sp.
A. Wild strain B. PEems150
Phosphate solubilization by Aspergillus niger
C. Wild strain D. ANsa120
Phosphate solubilization by
Aspergillus fumigatus E. AFsa120 F. Wild strain

Plate 5
Indole Acetic Acid production by bacterial and fungal cultures in Czepekdox broth and thin layer chromatogram for IAA
IAA production by Wild Bacterial and Fungal cultures.
A. Aspergillus niger, B. Pseudomonas sp., C. Penicillium sp., D. Aspergillus fumigatus, E. Control
Thin layer chromatogram for IAA
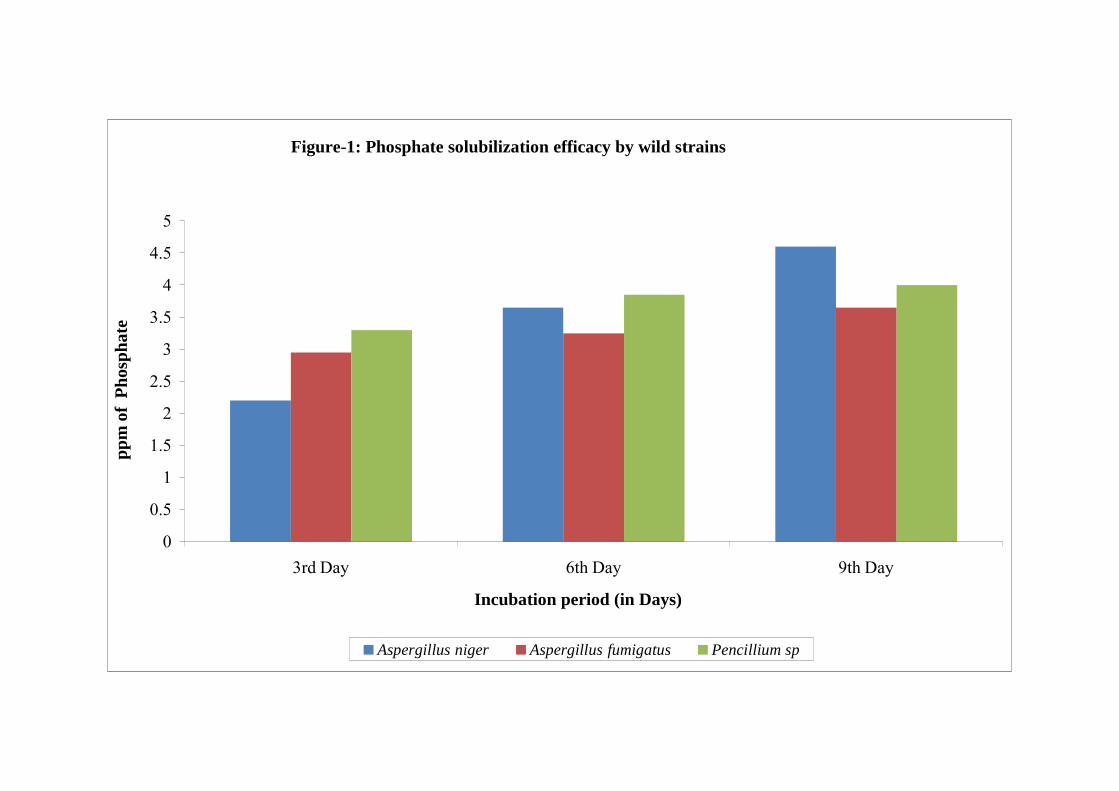
Figure-1: Phosphate solubilization efficacy by wild strains
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
3rd Day 6th Day 9th Day
Incubation period (in Days)
pp
m o
f P
hos
ph
ate
Aspergillus niger Aspergillus fumigatus Pencillium sp
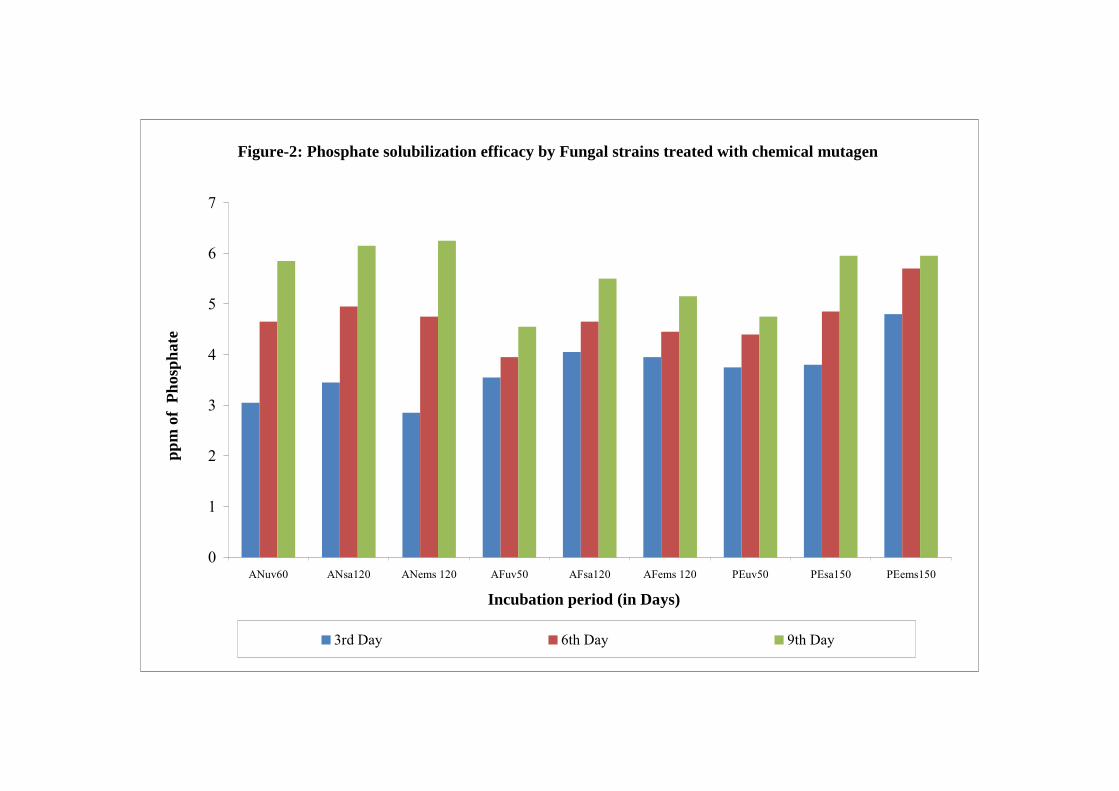
Figure-2: Phosphate solubilization efficacy by Fungal strains treated with chemical mutagen
0
1
2
3
4
5
6
7
ANuv60 ANsa120 ANems 120 AFuv50 AFsa120 AFems 120 PEuv50 PEsa150 PEems150
Incubation period (in Days)
ppm
of
Ph
osph
ate
3rd Day 6th Day 9th Day

Figure-3: Efficacy of phosphatase activityby wild strains
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
3rd Day 6th Day 9th Day
Incubation period (in Days)
µm
ol m
in-1
Aspergillus niger Aspergillus fumigatus Pencillium sp

Figure -4: Efficacy of phosphatase activityby Fungal strains treated with chemical mutagen
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
0.5
ANuv60 ANsa120 ANems120 AFuv50 AFsa120 AFems PEuv50 PEsa150 PEems150
Incubation period (in Days)
µm
ol m
in-1
3rd Day 6th Day 9th Day

Figure-5: Efficacy of Lipase activity by wild strains
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
Sucrose medium Olive oil medium
un
it g
-1 o
f su
bst
rate
Aspergillus niger Aspergillus fumigatus Pencillium sp

Figure-6: Efficacy of Lipase activity by Chemical treated Fungal Strains
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
ANuv50 ANsa90 ANems150 AFuv50 AFsa120 AFems120 PEuv60 PEsa120 PEems150
un
it g
-1of
su
bst
rate
Sucrose medium Olive oil medium

i
References
Abd-Alla, M.H., 1994. Phosphatases and the utilization of organic P by Rhizobium
leguminosarum biovar viceae. Biology and Fertility of Soils, 18: 294-296.
Achal, V., Savant V.V., Reddy, M.S., 2007. Phosphate solubilization by a wild type
strain and UV-induced mutants of Aspergillus tubingensis. Soil Biology &
Biochemistry, 39, 695–699.
Achamma, T., Manoj Mathews, A. K.Valsa, S. Mohan and R.Manjula.2003.
Optimizatiopn of growth conditions for the production of extracelluar Lipase
by Bacillus mycoides. Indian Journal of Microbiology, 43 : 67-69
Agnihothori, V.P. 1970. Solubilizaion of insoluble phosphate by some soil fungi
isolated from nursery seed beds. Canadian Journal of Microbiology, 16: 877-
880.
Ahamed, N. and K.K. Jha. 1968. Solubilization of rock phosphate by microorganisms
isolated from Bihar soils. Journal of Canadian Applied Microbiology, 14:89-95
Ahuja, A., Ghosh, S.B., DeSouza, S.F., 2007. Isolation of a starch utilizing, phosphate
solubilizing fungus on buffered medium and its characterization. Bioresource
Technology, 98 : 3408–3411
Akhtar, W. and M.N. Nawazish, M. 1983. Effect of triglycerides on the production Of
lipids lipase by Mucor hiemails. Division of Biochemistry, University of
Punjab, 329 : 664-669.
Alam, S., Khalil, S., Ayub, N. and Rashid. M., 2002. In vitro solubilization of inorganic
phosphate by phosphate solubilizing microorganisms (PSM) from maize
rhizosphere. Int. J. Agri. Biol., 4: 454-458.
Alexander, M., 1977. Introduction to Soil Microbiology. John Wiley and Sons, New
York.
Allan, V.J.M., Callow, M.E., Macaskie, L.E. and Beedle, M.P. 2002. Effect of nutrient
limitation on biofilm formation and phosphatase activity of a Citrobacter sp.
Microbiology, 148: 277–288
Amara, A.A and Salem, S.R. 2009. Degradation of Castor Oil and Lipase Production
by Pseudomonas aeruginosa. American-Eurasian Journal of Agric. & Environ.
Sci., 5(4): 556-563.

ii
Andrea E. Lucchini, Teresita A. Lisa and Carlos E. Domenech 1990. Choline
derivatives increase two different acid phosphatases in Rhizobium meliloti and
Pseudomonas aeruginosa. Archives of Microbiology, 153(6): 596-599.
Annibale, A., Sermanni, F.F. and Petruccioli, T., 2006. Olive-mill wastewaters: a
promising substrate for microbial lipase production. Bioresource Technology,
97: 1828-1833.
Arora, A. and Gaur, A.C., 1979. Microbial solubilization of different inorganic
phosphates. Indian Journal of Experimental Biology, 17: 1258 – 1260.
Arshad, M. and Frankenberger, W.T., 1991. Microbial production of plant hormones.
Plant Soil, 133: 1-8.
Artidaue, and Patel, H.H., 1999. Inorganic phosphate solubilizing soil Pseudomonas.
Indian Journal of Microbiology, 39: 161 – 164.
Arya, S., Kalia, V., Chandran, P. and Pundir, C.S., 2003. Lipase immobilized on walls
of plastic beaker: kinetic properties and application in washing of oil stained
cloth. Indian Journal of Chemical Technology, 10, 598-602
Asea, P.E.A., Kucey, R.M.N. and Stewart, J.W.B. 1988. Inorganic phosphate solubilization by two Penicillium species in culture solution and soil. Soil
Biochemistry, 20: 459-464.
Aunsuya, D. and Jayarajan, R. 1998. Solubilization of phosphorus by Trichoderma
viride. Current Science, 74(5): 464-465.
Ayyakkannu, K. and Chandramohan, D., 1971. Occurrence and distribution of phosphate solubilizing and phosphatase in marine sediments of Porto-Novo.
Mar. Biol., 11 : 201-205.
Baillargeon M.W., Bistline, R.G. and Sonnet, P.E., 1989. Evaluation of strains of
Geotrichum candidum for lipase production and fatty acid specificity. Appl
Microbiol Biotechnol., 30: 92– 96.
Bapiraju, K.V.V.S.N., Sujatha, P. and Ramana, T., 2004. Mutation induced enhanced
biosynthesis of lipase. African Journal of Biotechnology, 3: 618-621.
Barea, J.M. and Brown, M.E., 1974. Effects on plant growth by Azotobacter paspali related to synthesis of plant growth regulating substances. J. Appl. Bacteriol., 37: 583-593.
Barea, J.M., Navaro, E., Montoya, E., 1976: Production of plant growth regulators by
rhizosphere phosphate-solubilizing bacteria. J. Appl. Bacteriol., 40: 129–134.

iii
Barea, J.M., Ocampo, J.A., Azcon, R., Olivares, J. and Montoya, E., 1978. Effect of
ecological factors on the establishment of Azotobacter in the rhizosphere.
Ecological Bulletin (Stockholm), 26: 325-330.
Barrosol, C.B., Pereira, G.T., and Nahas, E., 2006. Solubilization of CaHPO4 and
AlPO4 by Aspergillus niger in culture media with different carbon and nitrogen
sources. Brazilian Journal of Microbiology, 37:434-438.
Bar-Yosef, B., Rogers, R.D., Wolfram, J.H. and Richman, E. 1999. Pseudomonas
cepacia–Mediated Rock Phosphate Solubilization in Kaolinite and
Montmorillonite Suspensions. Soil Science Society of American Journal, 63:
1703-1708
Bhurat, M.C. and Sen, A. 1968. Phosphate solubilizing bacteria in phyllosphere of
winer crops. Indian Journal of Microbiology, 8: 255-256.
Bhuvaneswari,V., Kathiravan, G., Gangadevi, V. and Muthumary, J. 2006. Extraction
and estimation of Indo, E-3-acetic acid (IAA) from some Endophytic and
pathogenic Coelomycetes. Asian Journal of Microbial. Biotech. Env. Sci., 8(2):
243-248.
Bolan, N.S., Naidu, R., Mahimairaja, S. and Baskaran, S., 1994. Influence of low-molecular-weight organic acids on the solubilization of phosphates. Biol. Fertil.
Soils, 18: 311-319.
Brady, N.C., 1990. The Nature and Properties of Soils. Macmillan, New York.
Bric, J.M., Bostock, R.M. and Silverstone, S.E., 1991. Rapid in situ assay for
indoleacetic acid production by bacteria immobilized on a nitrocellulose
membrane. Applied and Environmental Microbiology, 57: 535-538
Brown, M.E. 1972. Plant growth substances produced by microorganism of soil and
rhizosphere. J. Appl. Bacteriol., 35 : 443.
Brown, M.E. and Burlingham, S.W. 1968. production of plant growth substances by
Azotobacter chroococcum. J. Gen. Microbiol., 53: 135-144.
Brown, M.E., 1972. Plant growth substances produced by microorganisms of soil and
rhizosphere. J. Appl. Bacteriol. 43: 443-451.
Caob, S.G. and Zhanga, K.C., 2000. Production properties and application to non aqueous enzymatic catalysis of lipase from newly isolated Pseudomonas strain.
Enzyme Microbial Technology, 27: 74-82.

iv
Casida, L.E. 1959. Phosphate activity of common soil fungi. Soil Science, 87: 305-
310.
Cerezine, P.C., Nahas, E. and Banzattro, D.A., 1988. Soluble phosphate accumulation by Aspergillus niger from fluorapatite. Applied Microbiology and
Biotechnology, 29: 501-501.
Chaiharn, M. and Lumyong, S. 2011. Screening and optimization of indole-3-acetic
acid production and phosphate solubilization from rhizobacteria aimed at
improving plant growth. Current Microbiology, 62(1): 173-81.
Chakraborty, B.N., Chakraborty, U., Saha, A., Sunar, K. and Dey, P.L., 2010.
Evaluation of phosphate solubilizers from soils of North Bengal and their
diversity analysis. World J. Agric. Science, 6(2): 195-200.
Chakraborty, M., Khan, H. and Ramakrishna, K., 2003. Symbiotic properties of
hypermotile mutants of Bradyrhizobium sp. (Vigna aconitifolia). Indian Journal
of Microbiology, 43(2): 101-103.
Chang, C.H. and Yang, S.S., 2009. Thermolerent phosphate solubilizing microbes for
multifunctional biofertilizer preparation. Bioresources Technology, 100: 1648-
1658.
Chang, R.C., Chou, S.J. and Shaw, J.F., 1994. Multiple forms and functions of Candida
rugosa lipase. Biotechnol. Appl. Biochem., 19: 93–97.
Chavan A.M. and Kakde, R.B., 2009. Impact of carbon and nitrogen sources on lipase
enzyme activity of fungi isolated from abnormal oilseeds. Ind. J. Appl. and Pure
Bio., 25: 31-34.
Chelius, M.K. and Wodzinski, R.J., 1994. Strain Improvement of Aspergillus niger for
phytase production. Applied Microbiology and Biotechnology, 41: 79-83.
Chen, Y.P., Rekha, P.D., Arun, A.B., Shen, F.T., Lai, W.A. and Young, C.C., 2006.
Phosphate solubilizing bacteria from subtropical soil and their tricalcium
phosphate solubilizing abilities. Appl. Soil Ecol., 34: 33-41.
Chhonokar, P.K. and Subba Rao, N.S., 1967. Phosphate solubilization of fungi
associated with legume root nodules. Canadian Journal of Microbiology, 13:
743-753.
Chouhan, M. and Dawande, A.Y., 2010. Partial purification, characterization of lipase produced from Pseudomonas aeruginosa. Asiatic Journal of Biotechnology
Resources, 01: 29-34.

v
Christakopoulos, P., Tzia, C., Kekos, D. and Macris, B.J., 1992. Production and
characterization of extracellular lipase from Calvatia gigantea. Appl. Microbiol.
Biotechnol., 38:194 –97.
ChuanQing, Z., WeiYi, H., 2005. Comparison in P-solubilizing effects between
different P-solubilizing microbes and variation of activities of their
phosphatases. Acta Pedologica Sinica, 42(2): 286-294
Chung, H., Park, M., Madhaiya, M., Seshadri, S., Son, J., Cho, H. and Sa, T., 2005.
Isolation and characterization of phosphate solubilizing bacteria from the
rhizosphere of crop plants of Korea. Soil Biology and Biochemistry, 37: 1970-
1974.
Ciafardini, G., Zullo, B.A. and Iride, A., 2006. Lipase production by yeasts from extra
virgin olive oil. Food Microbiology, 23: 60–67
Coca, J., Hernandez, O., Berrio, R., Martinez, S., Diaz, E. and Dustet, J.C., 2001.
Production and characterization of lipases from Aspergillus niger and A.
fumigatus. Biotecnologia Applicada., 18: 216-220.
Constein, W. and Thomas. A., 1902. Purification and crystallization of lipase.
Agricultural Biology and Chemistry, 15 : 202-207.
Cordova J., 1998. Isolement identification et physiologie des champignons
thermophiles en vue de la production de lipases par fermentation en milieu
solide Montpellier, France: Universite´ de Montpellier II.
Cowan, S.T., and Steel, K.J, 1970. Manual for the identification of medical bacteria.
Cambridge University Press, London.
Crespo,J.M., Boiardi, J.L. and Luna, M.F., 2011. Mineral phosphate solubilization
activity of gluconacetobacter diazotrophicus under P-limitation and plant root
environment. Agricultural Sciences, 2(1): 16-22.
Cunningham, J.E. and Kuiack, C., 1992. Production of citric and oxalic acids and
solubilization of calcium phosphate by Penicillium billai. Applied and
Environmental Microbiology, 52: 1451–1458.
Dalmau, E., Montesinos, J.L., Lotti, M. and Casas, C., 2000. Effect of different carbon
sources on lipase production by Candida rugosa. Enzyme Microb. Technol., 26
(9/10): 657–663.
Damaso, M.C.T., Passianoto, M.A., Freitas, S.,C.D., Freire, D.M.G., Lago, R.C.A.,
and Couri, S., 2008. Utilization of agro industrial residues for lipase production
by solid-state fermentation. Brazilian Journal of Microbiology, 39: 676-681.

vi
Darmwal, N.S., Singh, R.S. and Raj, R., 1989. Isolation of phosphate solubilizers
from different sources. Current Science, 58(10): 570-571.
Das, A.C. and Debnath, A., 2006. Effect of systemic herbicides on N2-fixing and phosphate solubilizing microorganisms in relation to availability of nitrogen and
phosphorus in paddy soils of West Bengal. Chemosphere, 65: 1082–1086.
Das, A.C., 1963. Utilization of insoluble phosphate by soil fungi. J. Indian Soc. Soil
Sci., 11: 195–207.
Das, K., Katiyar, V. and Goel, R., 2003. ‘P’ solubilization potential of plant growth
Promoting Pseudomonas mutants at low temperature. Microbiol. Res., 158: 359
– 362.
Davies, P.J., 1995. Plant hormones physiology, biochemistry and molecular biology,
2nd edn. Dordrecht: Kluwer Academic Publishers, 112.
De Freitas, J.R., Banerjee, M.R. and Germida, J.J., 1997. Phosphatesolubilizing
rhizobacteria enhance the growth and yield but not phosphorus uptake of canola
(Brassica napus L.). Biology and Fertility of Soils, 24: 358–364.
De Souza, M.J.B.D., Nair, S. and Chandramohan, D., 2000. Phosphate solubilizing
bacteria around Indian peninsula. Indian Journal of Marine Sciences, 29, 48-51.
Del Campillo, S.E., Van der Zee, S., Torrent, J., 1999. Modelling longterm phosphorus
leaching and changes in phosphorus fertility in excessively fertilized acid sandy
soils. Eur. J. Soil Sci., 50: 391–399.
Disimine, C., Sayer, J.A., Gadd, G.M., 1998. solubilization of zinc phosphate by strain
of Pseudomonas fluorescens isolated from forest soil. Biol. Fertil. Soils, 28: 87-
94.
Dow, Y.K and Fhee, J.S., 1986. A simple and rapid colorimetric method for
determination of free fatty acids for lipase assay. Journal American oil
Chemist’s Society, 63 : 89-92.
Eileen Ingham., K.T. Holland., G. Gowland and W.J. Cunliffe. 1979. Purification and
partial characterization of an acid phosphatase produced by Propionibacterium
acnes. Journal of General Microbiology, 118: 59 – 65.
Ekin, Z. 2010. Performance of phosphate solubilizing bacteria for improving growth
and yield of sunflower (Helianthusannuus L.) in the presence of phosphorus
fertilizer. African Journal of Biotechnology, 9(25): 3794-3800.

vii
Ellaiah, P., Prabhakar, T., Ramakrishna, B., Thaer Taleb, A. and Adinarayana, K.,
2002. Strain improvement of Aspergillus niger for the production of lipase.
Indian Journal of Microbiology, 42: 151-153.
Etesami, H., Alikhani, H.A. and Akbari, A.A., 2009. Evaluation of plant growth
hormones production (IAA) ability by Iranian soils rhizobial strains and effects
of superior strains application on wheat growth indexes. World Applied
Sciences Journal, 6 (11): 1576-1584
Falony, G., Armas, J.C., Mendoza, J.C.D. and Hernandez, J.L.M., 2006. Production of
extracellular lipase from Aspergillus niger by Solid-state fermentation. Food
Technol. Biotechnol., 44 (2). 235–240.
Fankem, H., Nwaga, D., Deubel, A., Dieng, L., Merbach, W. and Etoa, F.X. 2006.
Occurrence and functioning of phosphate solubilizing microorganisms from oil
palm tree (Elaeis guineensis) rhizosphere in Cameroon. African Journal of
Biotechnology, 5 (24): 2450-2460.
Fasim, F., Ahmed, N., Parsons, R. and Gadd, G.M., 2002. Solubilization of zinc salts
by bacterium isolated by the air environment of tannery. FEMS Microb. Lett.,
213: 1-6.
Fenice, M., Selbman, L., Federici, F. and Vassilev, N., 2000. Application of
encapsulated Penicillium variable P16 in solubilization of rock phosphate.
Bioresource Technology, 73(2): 157-162.
Fett, W., Osman, S. and Dunn, M., 1987. Auxin production by plant pathogenic
Pseudomonas and Xanthomonas. Applied Environmental Microbiology,
53:1839-1845.
Fickers, P., Fudalej, F., Nicaud, J.M., Destain, J. and Thonart, P. 2005. Selection of
new over-producing derivatives for the improvement of extracellular lipase
production by the non-conventional yeast Yarrowia lipolytica. Journal of
Biotechnology, 115:379–386.
Freifelder, D., 1990. Microbial Genetics. Marosa Publishing House, New Delhi, India,
pp. 191 – 210.
Freire, D.M.G., Teles, E.M.F., Bon, E.P.S. and Sant’Anna, G.L. 1997. Lipase
production by Penicillium restrictum in a bench-scale fermenter. effect of carbon and nitrogen nutrition agitation and aeration. Appl. Biochem.
Biotechnol., 63/64: 409–21.
Garcia, M., Hubbell, D.H., Gaskinsn, M.H. and Dazzo, F.B., 1980. Association of
Azospirillum with grass roots. Appl. Environ. Microbiol., 37: 1016-1024.

viii
Gardan, L., David, C., Morel, M., Glickmann, E., Ghorrah, M.A., Petit, A. and
Dessaux, Y., 1992. Evidence for a correlation between auxin production and
host plant species among strains of Pseudomonas syringae subsp. savastanoi.
Applied and Environmental Microbiology, 50(8): 1780-1783
Gaur, A.C. and Sachar, S., 1980. Effect of rock Phosphate and Glucose concentration
of phosphate solubilization by Aspergillus awamori, Current Science, 49: 553 –
554.
Gaur, A.C., Madhan, M. and Ostwal, K.P., 1973. Solubilization of phosphoric
compounds by native microflora of rock phosphates. Indian Journal of
Experimental Biology, 10: 393-394.
Gerald, P.M., and Tsuneo, Y., 1991. Further improvements in the yield of
monoglycerides during enzymatic glycerolysis of fats and oils. Journal
American Oil Chemist’s Society, 68 : 6-10.
Goenadi, D.H., Siswanto and Sugiarto, Y., 2000. Bioactivation of poorly soluble
phosphate rocks with a phosphorus solubilizing fungus. Soil Sci. Soc. Am. J.,
64: 927–932.
Goldstein A.H., 1994. Involvement of the quinoprotein glucose dehydrogenase in the solubilization of exogenous phosphates by gram-negative bacteria. In: Torriani-
Gorini A, Yagil E, Silver, S, editors. Phosphate in Microorganisms:Cellular and
Molecular Biology. Washington, DC: ASM Press, pp. 197–203.
Goldstein, A.H. and Liu, S.T., 1987. Molecular cloning and regulation of a mineral
phosphate solubilizing gene from Erwinia herbicola. Biotechnol., 5: 72-74.
Gombert, A.K., Pinto, A.L., Castilho, L.R. and Freire, D.M.G., 1999. Lipase
production by Penicillium restrictum in solid-state fermentation using babassu
oil cake as substrate. Process Biochemistry, 35: 85–90.
Gordillo, M.A., Obradors, N., Montesinos, J.L., Valero, F., Lafuente, J. and Sola, C.,
1995. Stability studies and effect of the initial oleic acid concentration on lipase
production by Candida rugosa. Appl. Microbiol. Biotechnol., 43(1): 38–41.
Gordillo, M.A., Sanz, A., Sanchez, A., Valero, F., Montesinos, J.L., Lafuente, J., and
Sola, C., 1998. Enhancement of Candida rugosa lipase production by using
different control fed-batch operational strategies. Biotechnol. Bioeng., 60(2):
156–168.
Goswami, K.P. and Sen, A., 1962. Solubilization of calcium phosphate by three
strains of phosphobacterin. Indian Journal of Agricultural Science, 32: 9-10.

ix
Guimaraes, L.H., Jorge, J.A., Terenzi, H.F., Jamur, M.C., Oliver, C., De Lourdes
Teixeira, De Moraes Polizeli, M., 2003. Effect of carbon source on alkaline
phosphatase production and excretion in Aspergillus caespitosus. Journal of
Basic Microbiology, 43(3):210-217.
Gupta, A., Meyer, J.M. and Goel, R., 2002. Development of heavy metal-resistant mutants of phosphate solubilizing Pseudomonas sp. NBRI 4014 and their
characterization. Current Microbiology, 45(5) : 323 - 327.
Halder, A.K., Mishra, A.K. and Chakarbarthy, P.K., 1991. Solubilization of inorganic
phosphate by Bradyrhizobium. Indian J. Exp. Biol., 29: 28-31.
Halder, A.K., Mishra, A.K., Bhattacharyya, P. and Chakrabartty, P.K., 1990.
Solubilization of rock phosphate by Rhizobium and Bradyrhizobium. J. Gen.
Appl. Microbiol., 36: 81–92.
Hamdali, H., Hafidi, M., Virolle, M.J. and Ouhdouch, Y., 2008. Rock
phosphatesolubilizing Actinomycetes: screening for plant growth-promoting
activities. World J. Microbiol. Biotechnol., 24: 2565-2575.
Hameeda, B., Harini, G., Rupela, O. P., Wani, S. P. and Reddy, G. 2006. Growth
promotion of maize by phosphate solubilizing bacteria isolated from composts and macrofauna. Microbiol. Res., 163(2): 234-242
Hardy, D.J.E., Karamushka, V.I., Gruzina, Nikovska, G.N., Sayer, J.A. and Gadd,
G.M., 1998. Influence of carbon and nitrogen and phosphorus source on the
solubilization of insoluble metal compounds by Aspergillus niger. Mycol. Res.,
102: 1050-1054.
Hilda, K. A and Gobinith, S., 1997. Hydrolysis of oil and marine environment ethics.
Center for Advanced Studies in Botany, 15 : 21-28.
Hosseinpour, M.N., Najafpour, G.D., Younesi, H. and Khorrami, M., 2011. Submerged
culture studies for lipase production by Aspergillus niger NCIM 584 on soya
flour. Middle East Journal of Scientific Research, 7(3). 362-366.
Hu, X., Chen, J. and Guo, J., 2006. Two phosphate- and potassium-solubilizing bacteria
isolated from Tianmu Mountain, Zhejiang, China. World Journal of
Microbiology & Biotechnology, 22:983–990
Idris, E.E., Iglesias, D.J., Talon, M. and Borriss, R., 2007. Tryptophan-Dependent
Production of Indole-3-Acetic Acid (IAA) Affects Level of Plant Growth
Promotion by Bacillus amyloliquefaciens FZB42. The American
Phytopathological Society, 20 (6): 619–626.

x
Illmer, P. and Schinner, F., 1995. Solubilization of inorganic calcium phosphates
solubilization mechanisms. Soil Biol. Biochem., 27: 257–263.
Illmer, P., Barbato, A., Schinner, F., 1995. Solubilization of hardlysoluble AlPO4, with
P-solubilizing microorganisms. Soil Biol. Biochem., 27: 265–270.
Illmer, P., Schinner, F., 1992. Solubilization of inorganic phosphate by microorganisms
isolated from forest soils. Soil Biol. Biochem., 24: 389–395.
Iwai M., Tsujisaka Y., Okamoto, Y. and Fukumoto, J., 1973. Lipid requirement for the
lipase production by Geotrichurn candidum Link. Agric. Biol. Chem., 37: 929.
Jayasinghearachchi, H.S. and Seneviratne, G., 2006. Fungal solubilization of rock
phosphate is enhanced by forming fungal–rhizobial biofilms. Soil Biology &
Biochemistry, 38: 405–408.
Jha, B.N., Sharma, G.D. and Shukla, A.K., 2006. Phosphatase activity of
ectomycorrhizal root and nutrient status of pine seedlings under different
physical factors. Asian Journal of Microbial Biotechnology and Environmental
Science, 8(3): 561-566.
Johnston, H.W., 1954. The solubilization of insoluble phosphate - A quantitative and
comparative study of the action of selected aliphatic acids on TCP. Journal of
Science and Technology, 36: 49-65.
Johri, S.N., Alurralde, J.D. and Klein. J., 1990. Lipase production by free and
Immobilized protoplast of Sporotriclum (chrysoporium) Thermophile Apinis.
Applied Microbiology and Biotechnology, 33: 367-371.
Jonsson, U and Snygg, B.G., 1974. Lipase production and activity as a function of time
pH and temperature of four lipolytic microorganism. Journal of Applied
Bacteriology, 37 : 571-581.
Joseph, S. and Jisha, M.S., 2009. Buffering reduces phosphate solubilizing ability of
selected strains of bacteria. World Journal of Agricultural Sciences, 5 (1). 135-
137.
Josephgilman, 2001. A Manual of Soil Fungi, 2nd Edition, Biotechnology Books,
Delhi.
Juichiro, F., Mikeo, I. and Yoshio, T., 1963. Purification and Crystallization of a lipase
secreated by Aspergillus niger. Journal of Applied Microbiology, 9 : 353-361.

xi
Kakde, R.B. and Chavan, A.M., 2011. Effect of carbon, nitrogen, sulphur, phosphorus,
antibiotic and vitamin sources on hydrolytic enzyme production by storage
fungi. Recent Research in Science and Technology, 3(5): 20-28.
Kamini, N.R., Hemachander, C. and Mala, J.G.S., 1999. Microbial enzyme technology
as an alternative to conventional chemicals in leather industry. Current Science,
77: 80-86.
Kang, S.C., Ha, C.G., Lee, T.G. and Maheshwari, D.K., 2002. Solubilization of
insoluble inorganic phosphates by a soil-inhabiting fungus Fomitopsis sp.
PS102. Current Science, 82: 439–442.
Kang, S.C., Pandey, P., Khillon, R. and Maheshwari, D.K., 2008. Process of rock
phosphate solubilization by Aspergillus sp. PS 104 in soil amended medium.
J. Environ. Biol., 29(5):743-746.
Kanwar, L. and Goswami, P., 2002. Isolation of Pseudomonas lipase produced in pure
hydrocarbon substrate and its application in the synthesis of isoamyl acetate
using membrane immobilized lipase. Enzyme and Microbial Technology, 31:
727-735.
Kapri, A. and Tewari, L., 2010. Phosphate solubilization potential and phosphatase activity of rhizospheric Trichoderma sp. Brazilian Journal of Microbiology.
(Online Journal).
Katiyar, V. and Goel, R., 2003. Solubilization of inorganic phosphate and plant growth
promotion by cold tolerant mutants of Pseudomonas fluorescens.
Microbiological Research, 158(2): 163-168.
Katznelson, H. and Bose, B., 1959. Metabolic activity and phosphate dissolving
capability of bacterial isolates from wheat roots rhizosphere and rhizosphere
and non rhizosphere soil. Candian Journal of Microbiology, 5: 79-85
Khan, A.A., Jilani, G., Akhtar, M.S., Naqvi, S.M.S. and Rasheed, M., 2009. Phosphorus solubilizing bacteria: occurrence, mechanisms and their role in crop
production. J. Agric. Biol. Sci., 1(1): 48-58.
Kim, K.Y., Jordan, D. and McDonald, G.A., 1998. Enterobacter agglomerans, phosphate solubilizing bacteria, and microbial activity in soil: effect of carbon
source. Soil Biol. Biochem., 30: 995–1003.
Kucey, R.M.M., 1984. Phosphate solubilizing bacteria and fungi in various cultivated
and virgin Alberta soils. Canadian Journal of Soil Science, 63: 671-679.

xii
Kulanthaivel, S., Kiruthesswari, R., Revathi, V., Devi, N.K.A. and Nagendran, N.A.,
2006. Production of auxin by wild and mutant strains of Azospirillum. Asian
Journal of Microbiology, Biotechnology and Environmental Science, 8(2):239-
241.
Kumar, A., Bhargava, P. and Rai, L.C., 2010. Isolation and molecular characterization
of phosphate solubilizing Enterobacter and Exiguobacterium species from
paddy fields of Eastern Uttar Pradesh, India. African Journal of Microbiology
Research, 4(9): 820-829.
Kumar, H.D. and Dhruv Kumar, 1990. Nature of UV resistance in the cyanobacterium
Nostoc lincleia. Current Science, 59(8):412-414.
Kumar, V., Behl, R.K. and Narula, N., 2001. Establishment of phosphate solubilizing
strains of Azotobacter chroococcum in the rhizosphere and their effect on wheat
cultivars under green house conditions. Microbiol. Res., 156: 87-93.
Kumari, A., Kapoor, K.K., Kundu, B.S. and Mehta, R.K., 2008. Identification of
organic acids produced during rice straw decomposition and their role in rock
phosphate solubilization. Plant Soil Environ., 54: 72-77.
Lapeyrie, F., Ranger, J. and Vairelles, D., 1991. Phosphate solubilizing activity of ectomycorrhizal fungi in vitro. Can. J. Bot., 69: 342-346.
Lawrence, R.C., Fryer, T.F., and Reiter, B., 1967. Rapid method for quantitative
estimation of microbial lipases. Nature (Lond.), 213: 1264–1265.
Lee , W.H., 1995. Application of microbial lipase enzyme. Applied and Environmental
Microbiology, 5 : 257-269.
Lee, M., Breckenridge, C. and Knowles, R., 1970. Effect of some culture conditions on
the production of indole- 3-acetic acid and gibberellins like substances by
Azotobacter vinelandii. Can. J. Microbiol., 16:1325-1330.
Lianghua, T., Liming, X., Min, S. and Huaying, G., 2007. Purification and Application
of a Lipase from Penicillium expansum PED-03. Applied Biochemistry and
Biotechnology, 142(2): 194-199.
Lima, V.M.G., Krieger, N., Sarquis, M.I.M., Mitchell, D.A., Ramos, L.P. and Fontana,
J.D., 2003. Effect of nitrogen and carbon sources on lipase production by
Penicillium aurantiogriseum. Food Technol Biotechnol., 41(2): 105–10.
Lin, T.F., Huang, H.I., Shen, F.T., Young, C.C., 2006. The protons of gluconic acid are the major factor responsible for the dissolution of tricalcium phosphate by
Burkholderia cepacia CC-Al74. Bioresource Technology, 97: 957–960.

xiii
Luthen, H., Claussen, M. and Bottger, M., 1999. Growth: progress in auxin research.
Cell Biol. Physiol. Progress in Bot., 60: 315-340.
Macrae, A.R. and Hammond, R.C., 1985. The realm of microbial lipases in
biotechnology. Biotechnological Genetic Eng., 3 : 193-219.
Mahadik, N.D., Bastawde, K.B., Puntambekar, U.S., Khire, J.M. and Gokhale, D.V.,
2004. Production of acidic lipase by a mutant of Aspergillus niger NCIM 1207
in submerged fermentation. Process Biochemistry, 39: 2031–2034.
Mahadik, N.D., Puntambekar, U.S., Bastawde, K.B., Khire, J.M. and Gokhale, D.
2002. Production of acid lipase by Aspergillus niger in solid state fermentation.
Process Biochem., 38: 715-721.
Mala J.G.S., Kamini, N.R., Puvanakrishnan, R., 2001. Strain improvement of
Aspergillus niger for enhanced lipase production. J. Gen. Appl. Microbiol., 47:
181–186.
Manjugupta and Bhriguvansji, S.S., 1977. Dissolution of phosphates as influenced by
soil moisture legumes and phosphate solubilizing microorganisms. Journal of
Indian Society of Soil Science, 45(3): 613-614.
Maor, R., Haskin, S., Kedmi, H.L. and Sharon, A., 2004. In Planta Production of Indole-3-Acetic Acid by Colletotrichum gloeosporioides f. sp. aeschynomene,
Applied and Environmental Microbiology, 70(3): 1852–1854
Maria Guineth, T.B., Valencia Plata, S.A., Castillo, J.B. and Nieto, P.M., 2000.
Isolation of Enterobacteria, Azotobacter sp and Pseudomonas sp Producers of
indole – 3 –acetic acid and siderophores from Colombian rice rhizosphere.
Revista latinoamericana de Microbiologia., 42 : 171 – 176.
Martin, M., 1990. Lipase catalyzed alcoholysis of sunflower oil. Journal American Oil
Chemist’s Society, 67 :168-170.
Mba, C.C., 1996. Rock phosphate solubilizing Streptosporangium isolates from casts of
tropical earthworm resources. Conservation and Recycling, 17(3): 211-217
Mehboob, A., Stal, L.J. and Hasnain, S., 2010. Production of Indole-3-Acetic Acid by
the Cyanobacterium Arthrospira platensis Strain MMG-9. J. Microbiol.
Biotechnol., 20(9): 1259–1265.
Metha, Y.R. and Bhide, V.P. 1970. Solubilization of TCP by some soil fungi. Indian
Journal of Microbiology, 39:161-164.

xiv
Miller, S.H., Browne, P., Combaret, C.P., Meynet, E.C., Morrissey, J.P. and Gara, F.O.,
2010. Biochemical and genomic comparison of inorganic phosphate
solubilization in Pseudomonas species. Environmental Microbiology Reports,
2(3): 403-411.
Minoda, Y. and Ota, Y., 1980. Microbial utilization of renewable resources on agro-
industry microbial technology, JSPS-NRCT, 1: 73.
Mittal, V., Singh, O., Nayyar, H., Kaur, J. and Tewari, R., 2008. Stimulatory effect of
phosphate-solubilizing fungal strains (Aspergillus awamori and Penicillium
citrinum) on the yield of chickpea (Cicer arietinum L. cv. GPF2), Soil Biology
and Biochemistry, 40: 718–727.
Mohamed S.M., Abdel-Fattah, N.K. and El-Shahed, A.S., 1985. Studies on the lipolytic
activity of some local isolates of Aspergillus. Proc. Egypt. Bot. Soc. 4 (lsmaillia
conf.). pp. 68-80.
Moore, T.S., 1989. Biochemistry and physiology of plant 2nd edn. New York:
Springer-Verlag Inc., 285.
Muralidhar, R.V., Chirumamila, R.R., Marchant, R. and Nigam, P., 2001. A response surface approach for the comparison of lipase production by Candida cylindracea using two different carbon sources. Biochemical Engineering
Journal, 9:17–23
Nahas, E., 1988. Control of lipase production by Rhizopus oligosporus under various growth conditions. Journal of Genetic Microbiology, 134: 227-233.
Nakashima, T., Fukudo, H., Kyotani, S. and Morikawa, H., 1988 Culture conditions for
intracellular lipase production by Rhizopus chinensis and its immobilisation
within biomass support particles, J. Ferment. Technol., 66: 441–448.
Narsian, V. and Patel, H.H., 2000. Aspergillus aculeatus as a rock phosphate
solubilizer. Soil Biology and Biochemistry, 32: 559–565.
Narula, N., Debel, A., Gransee, A. and Merbach, W., 2007. Phosphate solubilizing Microorganisms: Mechanism of solubilization as influenced by pH, type of
organic acid and sugars. Asian Journal of Microbiol. Biotech., 9(2): 397-404
Nautiyal, C.S., Bhadauria, S., Kumar, P., Lal, H., Mond, L. R. and Verma, D., 2000. Stress induced phosphate solubilization in bacteria isolated from alkaline soils.
FEMS Microbiological Letters, 182: 291-296.

xv
Nawani, N. and Kaur, J., 2007. Studies on lipolytic isoenzymes from a thermophilic
Bacillus sp.: Production, purification and biochemical characterization, Enzyme
and Microbial Technology, 40: 881–887.
Nobuhiro, T. and Omar, I.C., 1977. Isolation and identification of alkaline lipase
Producing microorganism. Agricultural Biological Chemistry, 51 :181-187.
Normanly, J., 1997. Auxin metabolism. Physiol. Plantarum, 100: 431-442.
Normanly, J., Cohen, J.D. and Fink, G.R., 1983. Arabidopsis thaliana auxotrophs
reveal a tryptophan independent biosynthetic pathway for Indole 3-acetic acid.
Proc. Natl. Acad. Sci., 90: 10355-10359
Ogundero, V.W., 1981. Thermophilic fungi from Nigerian palm. Mycologia., 73: 198-
203.
Ohta, Y., Ikeda, K. and Ueda, S. 1968. Production of phosphatase by Aspergillus
awamori var. kawachii in a low phosphate medium. Applied Microbiology,
16(7): 973-980.
Oso, B.A., 1978. The lipase activity of Talaromyces emersonii. Can Journal of
Botony., 56 : 1840-1843.
Ostwal, K.P. and Bhide, V.P., 1972. Solubilization of TCP by soil Pseudomonas.
Indian Journal of Experimental Biology, 10:153-154.
Ouzari H., Khsairi A., Raddadi N., Jaoua L., Hassen A., Zarrouk M., Daffonchio D. and Boudabous A., 2008. Diversity of auxin-producing bacteria associated to
Pseudomonas savastanoi-induced olive knots. Journal of Basic Microbiology,
48: 1-8.
Padmapriya V. and Anand. N., 2010. Evaluation of some industrially important
enzymes in filamentous cyanobacteria. ARPN Journal of Agricultural and
Biological Science, 5(5): 86-97.
Pal, N., Das, S. and Kundu, A.K., 1978. Influence of culture and nutritional conditions
on the production of lipases by submerged culture of Aspergillus niger. J.
Ferment. Technol., 56: 593–598.
Pandey, A., Palni, L.M.S., Mulkalwar, P. and Nadeem, M., 2002. Effect of temperature
on solubilization of tricalcium phosphate by Pseudomonas corrugata. J. Sci. Ind. Res., 61:457–460.

xvi
Pany, B.K., Sarangi, C.R. and Behera, A.K., 1998. Response of wetland rice to
combined application of phosphobacterium and Inorganic phosphate in Altisol.
Indian Journal of Agricultural Science, 6: 330-332.
Papaparaskevas, D., Christakopoulos, P., Kekos, D. and Macris, B.J., 1992. Optimizing
production of extracellular lipase from Rhodotorula glutinis. Biotechnol Lett.,
14:397– 402.
Park, K.H., Lee, C.Y. and Son, H.J., 2009. Mechanism of insoluble phosphate
solubilization by Pseudomonas fluorescens RAF15 isolated from ginseng
rhizosphere and its plant growth-promoting activities. Letters in Applied
Microbiology, 49(2): 222–228.
Parks, E.J., Olson, G.J., Brinckman, F.E. and Baldi, F., 1990. Characterization by high performance liquid chromatography (HPLC) of the solubilization of phosphorus
in iron nore by a fungus. J. Ind. Microbiol., 5: 183–190.
Parmar, V.S., Bished, K.S., Singh, A. and Jha, A., 1996. The realm of microbial
lipases in biotechnology. Proc. Ind. Acad. Science, 108 :575-583.
Patten, C.L. and Glick, B.R., 2002. Role of Pseudomonas putida indole acetic acid in
development of the host plant root system. Applied and Environmental Microbiology, 68(8): 3795–3801.
Paul, F.O.W. and Carles, S.Z.Y., 1992. Lipase production by immobilized Candida
rugosa cells. Applied Microbial Biotechnology, 37 : 737-741.
Pawar, V.C. and Thaker, V.S., 2009. Acid phosphatase and invertase activities of
Aspergillus niger. Mycoscience, 50:323–330.
Pedregosa, A.M., Pinto, F., Monistrol, I.F. and Laborda, F., 1991. Regulation of acid
and alkaline phosphatases of Cladosporium cucumerinum by inorganic
phosphate. Mycol. Res., 95: 720–724.
Peix, A., Rivas, R., Mateos, P.F., Molina, E.M., Barrueco, C.R. and Velazquez, E.,
2003. Pseudomonas rhizosphaerae sp. nov., a novel species that actively
solubilizes phosphate in vitro. Int. J. Syst. Evol. Microbiol., 53: 2067-2072.
Pereira-Meirelles, F.V., Rocha-Leao, M.P.M. and Sant-Anna, G.L., 1997. A stable
lipase Candida lipolytica from: Cultivation condition and crude enzyme
characteristics. Applied Biochemistry and Biotechnology, 63:73-85.
Perez, E., Sulbarian, M., Ball, M.M. and Yarzabal, L.A., 2007. Isolation and
characterization of mineral phosphate solubilizing bacteria naturally colonising

xvii
a limonitic crust in the south-eastern Venezuela region. Soil Biol. Biochem., 36:
2905-2914.
Petrovic, S.E., Srinjar, M., Becarevic, A., Vujicic, I.F. and Banka, L., 1990. Effect various carbon sources on microbial lipase biosynthesis. Biotechnological
Letters, 12: 299-304.
Pogaku, P., Suresh, A., Srinivas, P. and Reddy, S.R., 2010. Optimization of lipase
production by Staphylococcussp. Lp12. African Journal of Biotechnology,
9(6):882-886.
Pokorny, D., Friedrich, J. and Cimerman, A., 1994. Effect of nutritional factors on
lipase biosynthesis by Aspergillus niger. Biotechnol. Lett., 16:363–66.
Ponmurugan, P. and Gopi, C., 2006. Invitro production of growth regulators and
Phosphatase activity by phosphate solubilizing bacteria, African Journal of
Biotechnology, 5(4): 348 – 350.
Posorske. L.H. 1984. Industrial scale application of enzyme to the fats and oil industry.
Journal American Oil Chemist’s Society, 61 ; 1758-1760.
Prada, P.D., Curtze, J.L. and Brenchley, J., 1996. Production of two extracellular
alkaline phosphatases by a psychrophilic Arthrobacter strain. Applied and Environmental Microbiology, 62(10): 3732–3738.
Pradhan, N. and Sukla, L.B., 2005. Solubilization of inorganic phosphate by fungi
isolated from agriculture soil. African Journal of Biotechnology, 5:850-854.
Prijambada, I.D., Widada, J., Kabirun, S. and Widianto, D., 2009. Secretion of organic
acids by phosphate solubilizing bacteria isolated from Oxisols. J. Tanah Trop.,
14(3): 245-251.
Puente, M.E., Bashan, Y., Li, C.Y. and Lebsky, V.K., 2004. Microbial populations and
activities in the rhizoplane of rock-weathering desert plants, I: root colonization
and weathering of igneous rocks. Plant Biology, 6: 629–642.
Puruchothaman, C.S., 1994. Studies on phosphate solubilizing and phosphatase
producing bacteria in Veil lake, Ph.D Thesis, Kerala University,
Thiruvananthapuram.
Radman, M., 1999. Enzyme of evolutionary change. Nature, 401: 866 – 868.
Rajankar, P.N., Tambekar, D.H., and Wate, S.R., 2007. Study of phosphate
solubilization efficiencies of fungi and bacteria isolated from saline belt of
Purna river basin. Res. J. Agric. Biol. Sci., 3: 701-703.

xviii
Rani, C. and Panneerselvam, A., 2009. Influence of environmental and nutritional
parameters on lipase production. ARPN Journal of Agricultural and Biological
Science. 4(5):39-43.
Rao, A.V., Venkaterwarlu, B. and Kaul, P., 1982. Isolation of phosphate solubilizing
soil actinomycetes. Current Science, 51(23): 1117-1118.
Rapp, P., 1995. Production, regulation, and some properties of lipase activity from
Fusarium oxysporum f.sp. vasinfectum. Enzyme Microb. Technol., 17:832–38.
Ray, N., Ray, L. and Rimani, B.N.S., 1999. Isolation and identification of Alkaline
thermo stable lipase producing microorganism, cultural conditions, nutritional
requirements and some reports of crude enzyme. Indian Journal of
Experimental Biology, 37: 818-824.
Reddy, M.S., Kumar, S., Babita, K. and Reddy, M.S., 2002. Biosolubilization of poorly
soluble rock phosphates by Aspergillus tubingensis and Aspergillus niger.
Bioresource Technology, 84: 187–189.
Relwani, L., Krishna, P. and Sudhakara Reddy, M., 2008. Effect of carbon and
nitrogen sources on phosphate solubilization by a wild-type strain and UV-
induced mutants of Aspergillus tubingensis. Curr. Microbiol., 57(5):401-406.
Reyes, I., Baziramakenga, R., Bernier, L. and Antoun, H., 2001. Solubilization of
Phosphate rocks and minerals by a acid type strain and two UV-induced
mutants of Pencillium rugulosum. Soil Biology and Biochemistry, 33, 1741 –
1747.
Reyes, I., Bernier, L., Simard, R., Tanguay, P.H., Antoun and H., 1999. Characteristics of phosphate solubilization by an isolate of a tropical Penicillium rugulosum
and two UV-induced mutants. FEMS Microbiology Ecology, 28: 291–295.
Richard G. and Tigerstrom, V., 1984. Production of two phosphatases by lysobacter
enzymogenes and purification and characterization of the extracellular enzyme.
Applied and Environmental Microbiology, 47(4): 693-698.
Richardson, A., Hadobas, P.A. and Hayes, J.E., 2000. Acid phosphomonoesterase and phytase activities of wheat (Triticum aestivum L.) roots and utilization of
organic phosphorus substrates by seedlings grown in sterile culture. Plant, Cell
and Environment, 23: 397–405.
Rifaat, H.M., El-mahalawy, A.A., El-menofy, H.A. and Donia, S.A., 2010. Production,
optimization and partial purification of lipase from Fusarium oxysporum.
Journal of Applied Sciences in Environmental Sanitation, 5(1): 39-53.

xix
Rodriguez, H. and Fraga, R., 1999. Phosphate solubilizing bacteria and their role in
plant growth promotion. Biotechnology Advances, 17: 319–339.
Rodriguez, H. and Reynaldo, F., 1999. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv., 17:319–339.
Rodriguez, J.A., Mateos, J.C., Nungaray, J., Gonzalez, V., Bhagnagar, T., Roussos, S.,
Cordova, J. and Baratti, J., 2006. Improving lipase production by nutrient
source modification using Rhizopus homothallicus cultured in solid state
fermentation. Process Biochemistry, 41: 2264–2269.
Rosas, S.B., Andres, J.A., Rovera, M. and Correa, N.S., 2006. Phosphate-solubilizing
Pseudomonas putida can influence the rhizobia–legume symbiosis. Soil Biology
& Biochemistry, 38: 3502–3505.
Sagoe, C.I., Ando, T., Kouno, K. and Nagaoka, T., 1997. Effect of organic-acid
treatment of phosphate rocks on the phosphorus availability to Italian ryegrass.
Soil Sci. Plant Nutr., 43: 1067-1072.
Saha, N. and Biswas, S., 2009. Mineral phosphate solubilizing bacterial community in
agro-ecosystem. African Journal of Biotechnology, 8(24): 6863-6870.
Sahu, S.N. and Jana, B.B., 2000. Enhancement of the fertilizer value of rock phosphate engineered through phosphate-solubilizing bacteria. Ecological Engineering,
15: 27–39.
Salihu, A., Alam, M.Z., Karim, M.I.A. and Salleh, M.H., 2011. Suitability of using
palm oil mill effluent as a medium for lipase production. African Journal of
Biotechnology, 10(11): 2044-2052.
Salleh, A.B., Musani, R., Basri, M., Ampon, K., Yunus, W.M.Z. and Razak, C.N.A.,
1993. Extra and intracellular lipases from a thermophilic R. oryzae and factors
affecting their production. Can. J. Microbiol., 39: 978–981.
Samad, M.Y.A., Salleh, A.B., Razak, C.N.K., Ampar, K., Yunus, W.M.Z.W. and
Basri, M., 1990. A lipase from a newly isolated thermophilic Rhizopus
rhizopodiformis. Journal of Microbial Biotechnology, 6: 390-394.
Samiran, B., Rakhi, P., Chandan, S. and Dominic, S., 2010. Stress induced phosphate
solubilization by Arthrobacter sp. and Bacillus sp. isolated from tomato
rhizosphere. Australian Journal of Crop Science, 4(6): 378-383.
Sarapatka, B., Dudova and Krskova, M., 2004. Effect of pH and phosphate supply on
acid phosphatase activity in cereal roots. Biologia. Bratislava, 59: 127-131.

xx
Sarwar, M. and Frankenberger, W.T. 1994. Tryptophan dependent biosynthesis of
auxins in soil. Plant and Soil., 160: 97-104.
Satyendra Kumar, Khyodano Kikon, Ashutosh Upadhyay, Shamsher S. Kanwar, Reena Gupta, 2005. Production, purification and characterization of lipase from
Thermophilic and alkaliphlic, Bacillus coagulans. Protein Expression and
Purification, 41: 32-44.
Scott, T.K., 1972. Auxins and roots. Annu. Rev. Plant Physiol., 23: 235-258.
Semenova, I.N., Egorov, S.N. and Egorov, N.S., 1986. Formation of secreted acid
phosphatase during the growth of Saccharomyces cerevisiae yeasts on different
sources of carbon and nitrogen nutrition. Mikrobiologia., 55(5):796-9.
Sen, A. and Paul, N.B., 1957. Solubilizaiton of phosphates by some common soil
bacteria. Current Science, 22: 222-223.
Seshadri, S., 1995. Phosphate solubilizing fungi from Thanjavur district, South India,
Ph.D. Thesis, Bharathidasan University, Tiruchirappalli.
Seshadri, S., Ignacimuthu, S. and Lakshminarasimhan, C. 2004. Effect of nitrogen and
carbon sources on the inorganic phosphate solubilization by different
Aspergillus niger strains. Chemical Engineering Communications, 191(8):
1043-1052.
Seshadri, S., Muthukumarasamy, R., Laksminarasimhan and Igncimuthu, S., 2000.
solubilization of inorganic phosphate by Azospirillum hakioracferans. Current
Science, 79(5): 565-567.
Sethi and Subba Rao, N.S., 1968. Phosphate solubilization of fungi associated with
legume root nodules. Canadian Journal of Microbiology, 13: 743-753.
Sevagi, E., Donmez, G., Takac, S., 2007. Isolation of Lipase producing Bacillus
sp.from olive mill waste water and improving its enzyme activity. Journal of
Hazardous Materials, 149: 720-724.
Shahab, S. and Ahmed, N., 2008. Effect of various parameters on the efficiency of zinc
phosphate solubilization by indigenous bacterial isolates. African Journal of
Biotechnology, 7(10): 1543-1549.
Shahab, S., Ahmed, N. and Khan, N.S., 2009. Indole acetic acid production and enhanced plant growth promotion by indigenous PSBs. African Journal of
Agricultural Research, 4(11): 1312-1316.

xxi
Sharan, A., Shikha and Darmwal, N.S., 2008. Efficient phosphorus solubilization by
mutant strain of Xanthomonas campestris using different carbon, nitrogen and
phosphorus sources. World Journal of Microbiology and Biotechnology, 24(12):
3087-3090.
Shekhar, C., Bhadawia, S., Kumar, P., Lal, H., Mondal, R. and Verma, D., 2000.
Stress induced phosphate solubilization in bacteria isolated from alkaline soils.
FEMS Microbiology Letters, 182(2): 2991-2996.
Shimada, Y., Sugihara, A., Nagao, T. and Tominaga, Y., 1992. Induction of
Geotrichum candidum lipase by long-chain fatty acids. J. Fermet. Bioeng., 74:
77– 80.
Silva, W.O.B., Mitidieri, S. and Schrank, M.H. 2005. Vainstein Production and
extraction of an extracellular lipase from the entomopathogenic fungus
Metarhizium anisopliae. Process Biochemistry, 40: 321–326.
Singal, R., Gupta, R. and Saxena, R.K., 1994. Rock phosphate solubilization under
alkaline conditions by Aspergillus japonicus and A. foetidus. Folia
Microbiologica, 39(1): 33-36.
Singh, H.P., Pareek, R.P. and Singh, T.A., 1984. Solubilization of rock phosphate by phosphate solubilizer in broth. Current science., 53: 1212-1213.
Singleton, T. and Sainsburg, B., 1988. Dictionary of Microbiology and molecular
biology. John wiley and Sons, Singapore, pp 1019.
Smith, R.F., Blasi, D. and Dayton, S.L., 1973. Phosphatase activity among Candida species and other yeasts isolated from clinical material. Applied Microbiology,
26: 364-367.
Son, H.J., Park, G.T., Cha, M.S. and Heo, M.S., 2006. Solubilization of insoluble inorganic phosphates by a novel salt- and pH-tolerant Pantoea agglomerans R-
42 isolated from soybean rhizosphere. Bioresources Technology, 97: 204–210.
Sperber, J.I. 1957, Solubilization of mineral Phosphate by soil bacteria. Nature, 18: 994
– 995.
Spicers, G.A., and McGill, W.B., 1979. Effects of phosphorus addition and energy supply on acid phosphatase production and activity in soils. Soil Biology and
Biochemistry, 11: 3-8.
Sridevi, M., and Mallaiah, K.V. 2007. Production of Indole Acetic Acid by Rhizobium
isolates from Sesbania species. African Journal of Microbiology Research, 1(7):
125-128.

xxii
Srinorakutara, T., Cumkhunthod, P., Suttikul, S.,Imprasittichai, W., Mouthung, B. and
Wangpila, M., 2008. Strain improvement of ethanol fermenting yeast using
random mutagenesis technique. Thailand Journal of Biotechnology, 8(1): 120-
123.
Srividya, S., Soumya, S. and Pooja, K., 2009. Influence of environmental factors and
salinity on phosphate solubilization by a newly isolated Aspergillus niger F7
from agricultural soil. African Journal of Biotechnology, 8(9): 1864-1870.
Sudha, M. and Purushothaman, C.S., 2000. Effect of orthophosphate enrichment on the
phosphatase activity of Staphylococcus. In:40th Annual Conference AMI,
Bhubaneswar, India, pp-107.
Sudhansupal, 1999. Selection of acid tolerant strains of phosphate solubilizing bacteria
in soil of Uttar Pradesh, Himalayas. Indian Journal of Agricultural Science,
69(8): 5787-82.
Sugiura, T., Ota, Y. and Minodi, Y., 1975. Effect of fatty acids, lipase activators,
Phospholipids and related substances on the lipase production by Candida
paralipolytica. Agricultural Biological Chemistry, 39:1489-1494.
Susumu, S.O. and Yashio, T., 1975. The comparison of the properties of two lipase from Penicillium cyclopium westring. Agricultural Biology Chemistry, 39:
1063-1070.
Suzuki, T., Mushiga, Y., Yamane, T. and Shimizu, S. 1988. Mass production of lipase
by fed- batch culture of Pseudomonas fluorescens. Applied Microbiology and
Biotechnology, 27: 417-422.
Tabatabai, M.A. and Bremner, J.M., 1969. Use of para nitrophenyl phosphate for assay
of soil phosphatase activity. Soil biology and Biochemistry, 1: 301-307.
Taiwo, L.B., and Oso, B.A., 1997. The influence of some pesticides on soil microbial
flora in relation to changes in nutrient level, rock phosphate solubilization and P
release under laboratory conditions. Agriculture, Ecosystems and Environment,
65: 59-68.
Tarafdar, J.C., Rao, A.V. and Praveen Kumar, 1992. Effects of different phosphatase
producing fungi on growth and nutrition of Mung beans in an arid soil. Biology
and Fertility of Soil, 13: 35-38.
Thakker, J., Varshanarishan and Patel, H.H., 1993. Inorganic phosphate solubilization
by certain soil bacteria. Indian Journal of Experimental Biology, 31: 743-746.

xxiii
Tien, T.M., Gaskins, M.H. and Hubbell, D.H., 1979. Plant growth substances produced
by Azospirillum brasilense and their effect on growth of pearl millet Pennisetum
americanum (L.) Appl. Environ.Microbiol., 37: 1016-1024.
Toro, M., Azcon, R. and Barea, J., 1997. Improvement of arbuscular mycorrhizae
development by inoculation of soil with phosphate-solubilizing rhizobacteria to
improve rock phosphate bioavailability (32P) and nutrient cycling. Applied and
Environmental Microbiology, 4408-4412.
Tripura, C., Sashidhar, B. and Podile, A.R., 2007. Ethyl methanesulfonate mutagenesis-
enhanced mineral phosphate solubilization by groundnut-associated Serratia
marcescens GPS-5. Current Microbiology, 54: 79-84.
Tsavkelova, E.A., Cherdyntseva, T.A. and Netrusov, A.I., 2005. Auxin Production by
Bacteria Associated with Orchid Roots. Microbiology, 74,(1): 46–53.
Tsavkelova, E.A., Cherdyntseva, T.A., Botina, S.G., Netrusov, A.I., 2007. Bacteria
associated with orchid roots and microbial production of auxin. Microbiological
Research, 162: 69-76.
Tsekova, K. and Galabova, D., 2003. Phosphatase production and activity in copper(II)
accumulating Rhizopus delemar. Enzymeand Microbial Technology, 33(7): 926–931.
Tudzynski, B. and Sharon, A., 2002. Biosynthesis, biological role and application of
fungal phytohormones, In H. Osiewacz (ed.), The Mycota. A comprehensive
treatise on fungi as experimental systems for basic and applied research, vol. X.
Industrial applications. Springer-Verlag, Berlin, Germany. p. 183–211.
Valerie L., Louis, W.I. and Comeau, Z. 1992. Continuous synthesis of ester by cell-
bound fungal lipases in an organic solvent. Applied Microbial Biotechnology,
37: 732-736.
Vanitha, S., 2002. Lipase production by Aspergillus niger growing on coconut kernel. Master of Philosophy in Microbiology Thesis, Bharathidasan university,
Tiruchirappalli.
Varshanarishan, Jungu, T. and Patel, H.H., 1995. Mineral phosphate solubilization by
Aspergillus aculeatus, Indian Journal of Experimental Biology, 33: 91 – 93.
Vassilev, N. and Vassileve, M., 2003. Biotechnological solubilization of rock phosphate on media containing agro-industrial wastes. Appl. Microbiol.
Biotechnol., 61: 435–440.

xxiv
Vassilev, N., Fenice, M. and Federici, F., 1996. Rock phosphate solubilization with
gluconic acid produced by immobilized Penicillium variabile P16. Biotechnol.
Tech., 10: 585–588.
Vassilev, N., Vassileve, M., Bravo,V., Serrano, M.F. and Nikolaeva, I., 2007.
Simultaneous phytase production and rock phosphate solubilization by
Aspergillus niger grown on dry olive wastes. Industrial Crops and Products,
26: 332–336.
Vassileva, N., Vassileve, M. and Azcon, R., 1997. Solubilization of rock phosphate by
immobilized Aspergillus niger. Bioresource Technology, 59: 1–4.
Vassileve, M., Azcon, R., Barea, J.M. and Vassilev, N., 1998. Application of an
encapsulated filamentous fungus in solubilization of inorganic phosphate.
Journal of Biotechnology, 63: 67-72.
Vassileve, M., Vassilev, N., Fenice, M. and Federici, F., 2001. Immobilized cell
technology applied in solubilization of insoluble inorganic (rock) phosphate and
P plant acquisition. Bioresource Technol., 79: 263-271.
Vazquez, P., Holguin, G., Puente, M.E., Lopez, A. and Bachan, Y., 2000. Phosphate
solubilizing microorganisms associated with the rhizosphere of mangroves in
semi acid coastal lagoon. Biology and Fertility of Soil, 30(5-6): 48.
Venis, M.A. and Napier, R.M., 1991. Auxin receptors: recent developments. Plant
Growth Regul., 10: 329 340.
Venkateswarlu, B., Rao, A.V. and Raina, P., 1984. Valuation of P solubilization by
microorganisms isolated from arid soils. J. Indian Soc. Soil Sci., 32: 273-277.
Vijila, K., 2000. Estimation of IAA production in nitrogen fixing microorganisms.
Practical manual-microbial interaction in soil. Tamil Nadu Agricultural
University, Coimbatore. pp 38-39.
Vivekkumar, Punia, S., Lakshminarayana, K. and Neeru, N., 1999. Effect of phosphate
solubilizing analogue resistant mutants of agriculture Azotobacter chroococcum
on sorghum. Indian Journal of Agricultural Science, 69(3): 198-200.
Vyas, P., Rahi, P., Chauhan, A. and Gulati, A., 2007. Phosphate solubilization
potential and stress tolerance of Eupenicillium parvum from tea soil.
Mycological Research, 11: 931 - 938.
Wagner, K.U., Masepohl, B. and Pistorius, E.K., 1995. The cyanobacterium
synechococcus sp. strain PCC 7942 contains a second alkaline phosphatase
encoded by pho. V. Microbiol., 141: 3049- 3058.

xxv
Waller, W. and Comaeu, L.C., 1990. A noval strain of Candida species producing an
acid resistant lipase. Rev. Fr. Corps Gras. 37: 289- 296.
Wani, P.A., Saghir Khan, M. and Zaidi, A., 2007. Synergistic effects of the inoculation with nitrogen-fixing and phosphate-solubilizing rhizobacteria on the
performance of field-grown chickpea. Journal of Plant Nutrition and Soil
Science, 170(2): 283–287.
Warner, M., Linfield, dennis, J.O’ brien, Samuel, S., Robert, Barauskas, A., 1984.
Lipid- lipase interactions. Fat splitting with lipase from Candida rugosa.
Journal American Oil Chemist’s Society, 61: 1067-1071.
Watanabe, N., Yasuhide, O., Yasuji, M. and Koichi, Y., 1977. Isolation and
Identification of alkaline lipase producing microorganism, cultural conditions.
Dept of Agricultural Biological Chemistry, 41(8): 1355-1358.
Wen Hsiung L., Peppu, A.Z.T. and Arima, K.I., 1972. Purification and general
Properties of the lipase of thermophilic fungus Humicola lamuginosa.
Agricultural Biological Chemistry, 26: 73-79.
Wen Hsiung L., Teruhiko, B. and Kei, A., 1973. Substrate specificity and mode of action of lipase of thermophilic fungus Humicola lamuginosa s-38.
Agricultural Biological Chemistry, 37 : 1349-1355.
Whitelaw, M.A., Harden, T.J. and Helyar, K.R., 1999. Phosphate solubilization in solution culture by the soil fungus Penicillium radicum. Soil Biology &
Biochemistry, 31: 655-665.
Wyss, M., Pasamontes, L., Remy, R., Kohler, J., Kusznir, E., Gadient, M., Muller, F. and Van loon, A.G.M., 1998. Comparison of the thermostability properties of
three acid phosphatases from molds: Aspergillus fumigatus Phytase, A. niger
Phytase, and A. niger pH 2.5 Acid Phosphatase, Applied and Environmental
Microbiology, 64(11): 4446–4451.
Yurekli, F., Geckil, H. and Topcuoglu, F., 2003. The synthesis of indole-3-acetic acid
by the industrially important white-rot fungus Lentinus sajor-caju under
different culture conditions. Mycol. Res., 107(3): 305–309.

LIST OF PUBLICATIONS
1. Strain Improvement of Phosphate Solubilizing Fungal Strains (2010).
Journal of Ecobiotechnology, 2(5): 65-70.
2. Strain Improvement of Pseudomonas sp for the production of Lipase
(2010). Journal of Experimental Sciences, 1(11): 01-03.

Journal of Ecobiotechnology 2/5: 65-70, 2010 ISSN 2077-0464 http://journal-ecobiotechnology.com/
Strain Improvement of Phosphate Solubilizing Fungal Strains Department of Botany, Jamal Mohammed College, (Autonomous), Trichy-620020
*Corresponding author, Email: [email protected], [email protected], Tel.: ( 0431) 2331235, 2331935, Fax : ( 0431 ) 2331135
Phosphate solubilizing fungi Mutagenesis Enhancement Mutant strains
A laboratory study was conducted to isolate, investigate and enhance the phosphate solubilization efficiency of certain isolated fungal strains viz., Aspergillus niger, Aspergillus fumigatus, Penicillium sp. These fungal strains were isolated from the rhizosphere of paddy fields in Tamilnadu, India and screened for phosphate solubilization. The enhancement of phosphate solubilization of these strains was studied through three types of mutagenesis: UV, sodium azide and ethyl methane sulphonate mutagenesis. The mutant ANuv60 exhibited 127% increased efficiency for phosphate solubilization compared to the wild strains. Similarly, the phosphate solubilization by sodium azide mutant (ANsa120) was 133% higher and EMS mutant (ANems120) was 135% higher than the wild strain. The efficient strains were further studied for the effect of carbon and nitrogen source on phosphate solubilization. Significance of the findings is discussed in detail.
SA: Sodium azide, EMS: Ethyl methane sulphonate, UV: Ultra Violet, ANsa: Sodium azide treated Aspergillus niger, AFsa: Sodium azide treated Aspergillus fumigatus, PEsa: Sodium azide treated Penicillium. ANems: Ethyl methane sulphonate treated Aspergillus niger, AFems: Ethyl methane sulphonate treated Aspergillus fumigatus, PEems: Ethyl methane sulphonate treated Penicillium, ANuv: Ultra Violet treated Aspergillus niger, AFuv: Ultra Violet treated Aspergillus fumigatus and PEuv: Ultra Violet treated Penicillium. 1. Introduction
Phosphorus is called �the key of life� as it is directly involved in most life processes. Next to nitrogen it is invariably classified as one at the macronutrients and is an important key element in frequency of use as fertilizer. It serves as a primary energy source for microbial oxidation. It is a constituent substance in life processes. Soil can not give high yields if it is deficient in phosphate. Soil contains both organic and inorganic forms of phosphorus. The organic forms of the element in soil are compounds of phytins, phospholipids and nucleic acids that come mainly by the way of decaying vegetation.
Plants take phosphate in the form of soluble orthophosphate ions but due to the presence of Ca, Mg, K, Na, Al and Fe ions in soil, the soluble orthophosphate is converted to insoluble form. Because of this process plants utilize very little amount of phosphate, even though phosphatic fertilizers are added (Vivek kumar et al., 1999; Rosas et al., 2006; Jayasinghearachchi and Seneviratne, 2006). Soil microorganisms are reported to be initially associated in the cycling of phosphorus. But they also participate in solubilization of inorganic phosphorus and in the mineralization of organic phosphorus (Agnihothri, 1970; Ostwal and Bhide, 1972).
A large fraction of microbial population can dissolve insoluble inorganic phosphorus known to
occur in the soil (Hayman, 1975). Various microorganisms were reported to solubilize different types of insoluble phosphates. Bacteria such as Achromobacter sp, Agrobacterium sp, Fungi like Aspergillus niger, A. flavus, A. fumigatious, Penicillium sp, Rhizopus sp etc are the important phosphate solubilizers present in the soil.
Different mechanisms have been suggested for the solubilization of inorganic phosphorus by phosphate solubilizers. The production of organic acids in the microenvironment around the root or in the culture media is considered to be the important cause of phosphate solubilization (Sperber, 1957; Bardiya and Gaur, 1972). Achal et al. (2007) reported the phenotypic mutants of Aspergillus tubingensis were obtained by UV irradiation and phosphate solubilization ability of these mutants were studied and compared with wild type strains. However, not much work has been carried on phosphate solubilization by mutated Aspergillus and Penicillium species. Hence the present study has been undertaken to study reveals the efficacy of phosphate solubilization by mutated strains isolated from paddy soils.
2. Materials and Methods Microorganisms and isolation
The fungal strains were isolated from rhizosphere soils of Paddy field, Mannachanallur,

Rajeshkumar Jayaraman and M.H.Muhammad Ilyas Journal of Ecobiotechnology 2/5: 65-70, 2010
one of the largest paddy producing taluk located in Tamil Nadu State, India. The soil samples were screened for phosphate solubilizing fungi in Pikovskaya�s agar medium by conventional dilution plate count method. The fungal strains showing halo formation around their colonies were isolated and grown on Sabouraud agar slants at 27ºC for 3 days and kept in the refrigerator at 40C until further use (Ellaiah et al., 2002). Identification
The fungal cultures were identified based on the colony morphology and spore structure (Pradhan and Sukla, 2005) Random mutational studies Mutagenesis by UV
72 hrs old fungal cultures were scraped off from agar slants and suspended in 5ml sterile distilled water and then diluted with 45 ml of sterile distilled water containing Tween 80 (1:4000). Sterile glass beads were added to the suspension and kept on rotary shaker for 30 min to break the hyphal mycelium. The suspension was filtered to remove the mycelium. The spore suspension was prepared in phosphate buffer (pH7.0) containing 106 spores per ml. Five ml quantities of the spore suspension were transferred aseptically into sterile petri dishes and exposed to UV light (2600 Ao) at a distance of 15cm away from the center of the Germicidal lamp for various time intervals (10, 20, 30, 40, 50, 60, 70 and 80 min). The suspension was agitated by gently rotating the plates in between the time intervals. (Ellaiah et al., 2002).
The UV exposed spore suspensions were stored in dark for overnight to avoid photo reactivation. After overnight incubation, irradiated spore suspensions were serially diluted by using phosphate buffer (pH 7.0) and plated on Sabouraud�s Dextrose agar medium. The plates were incubated for 5 days at 27°C. The colonies were selected on the basis of their morphological characters and were given the code numbers ANuv30, ANuv40, ANuv50, ANuv60, AFuv30, AFuv40, AFuv50, AFuv60, PEuv30, PEuv40, PEuv50 and PEuv60. Mutagenesis by Chemical
Chemical mutagenesis was performed using Sodium azide and Ethyl methane sulphonate (EMS) for the strain improvement of phosphate solubilizers. Spore suspensions of fungal strains were prepared by using phosphate buffer pH 7.0 as described earlier (Ellaiah et al., 2002). To 9 ml of each spore suspension, 1 ml of sterile solution of Sodium azide (250 g ml-1 in phosphate buffer) was added. Similar procedure was adopted for Ethyl methane sulphonate (EMS) (150 g ml-1 in
phosphate buffer). The reaction was allowed to proceed. Control tube was also kept without any chemical mutagen. Samples were withdrawn from the reaction mixture at an interval of 30, 60, 90, 120 and 150 min. and centrifuged for 10 min. at 5000 rpm. The cells were washed three times with sterile distilled water and again re-suspended in 10 ml sterile buffer. The samples were serially diluted in the same buffer and plated on Sabouraud�s Dextrose agar medium. The selected sodium azide treated mutants were given the code numbers ANsa30, ANsa60, ANsa90, ANsa120, AFsa30, AFsa60, AFsa90, AFsa120, PEsa60, PEsa90, PEsa120 and PEsa150.
The EMS treated mutants were given the code numbers ANems30, ANems60, ANems90, ANems120,AFems30, AFems60, AFems90, AFems120,PEems60, PEems90, PEems120 and PEems150 (Bapiraju et al., 2004). Submerged fermentation Inoculation
Pure fungal cultures from agar slants were inoculated to Sabouraud�s Dextrose agar plates and incubated for four days at 27oC. After sufficient growth, 8 mm discs (~106 spores/ml) were cut from the plates using sterile cork borer and were inoculated into 100 ml Pikovskaya�s broth (Narsian et al., 1995). The flasks were incubated at 27oC for 9days. Control Pikovskaya�s broth medium was also kept and the experiment was conducted in triplicate set. Phosphate estimation
Broth culture medium was withdrawn aseptically at three days interval from each flask and centrifuged at 10,000 rpm for 20 minutes (Dave and Patel, 1999). The supernatant was estimated for phosphate content by chlorostannous reduced molybdophosphoric blue colour method (Seshadri, 1995). Effect of different carbon sources
Effect of different carbon sources on phosphate solubilization efficiency of the isolated strains was also studied. The carbon source used was maltose, sucrose and mannitol instead of glucose. Efficient phosphate solubilizing fungal strains were inoculated in to broth media with various carbon sources and incubated. After incubation, phosphate solubilization assay was carried out (Narsian and Patel, 2000). Effect of different nitrogen sources
To study the effect of different nitrogen sources on phosphate solubilization efficiency of the fungal strains, the nitrogen source ammonium sulphate was replaced by potassium nitrate, sodium nitrate and urea. Efficient phosphate solubilizing
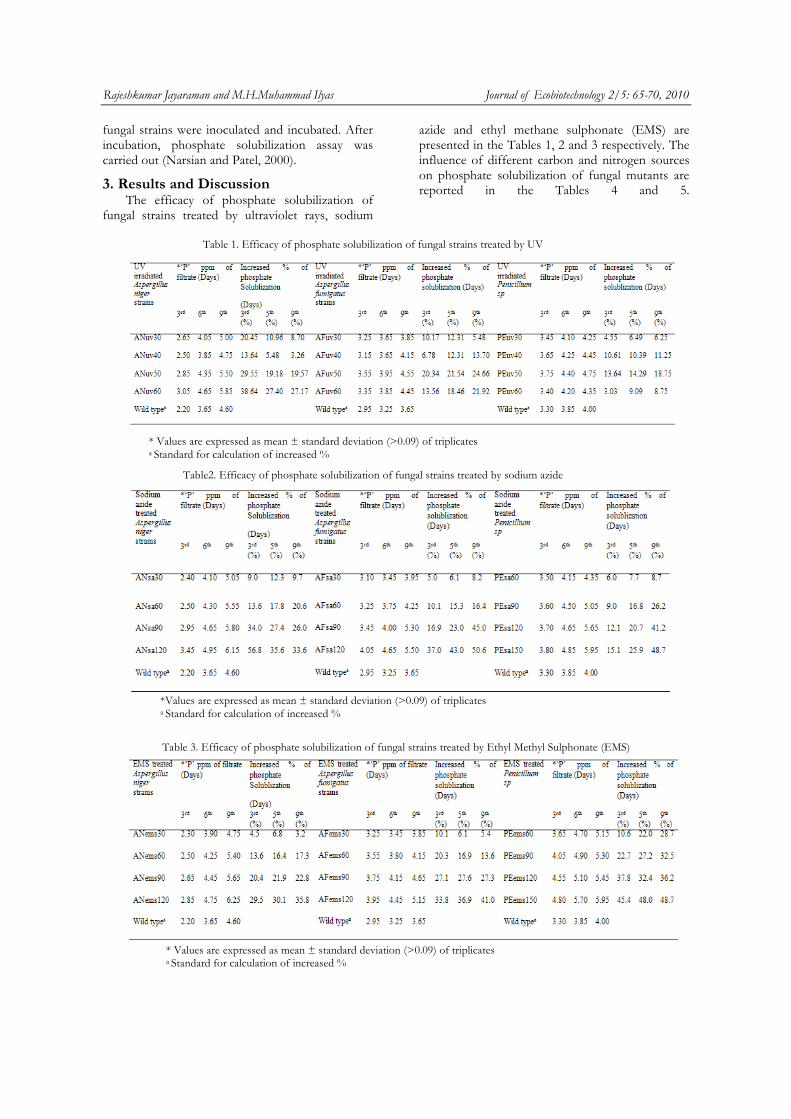
Rajeshkumar Jayaraman and M.H.Muhammad Ilyas Journal of Ecobiotechnology 2/5: 65-70, 2010
fungal strains were inoculated and incubated. After incubation, phosphate solubilization assay was carried out (Narsian and Patel, 2000).
3. Results and Discussion The efficacy of phosphate solubilization of
fungal strains treated by ultraviolet rays, sodium
azide and ethyl methane sulphonate (EMS) are presented in the Tables 1, 2 and 3 respectively. The influence of different carbon and nitrogen sources on phosphate solubilization of fungal mutants are reported in the Tables 4 and 5.
Table 1. Efficacy of phosphate solubilization of fungal strains treated by UV
* Values are expressed as mean ± standard deviation (>0.09) of triplicates a Standard for calculation of increased %
Table2. Efficacy of phosphate solubilization of fungal strains treated by sodium azide
*Values are expressed as mean ± standard deviation (>0.09) of triplicates a Standard for calculation of increased %
Table 3. Efficacy of phosphate solubilization of fungal strains treated by Ethyl Methyl Sulphonate (EMS)
* Values are expressed as mean ± standard deviation (>0.09) of triplicates
a Standard for calculation of increased %
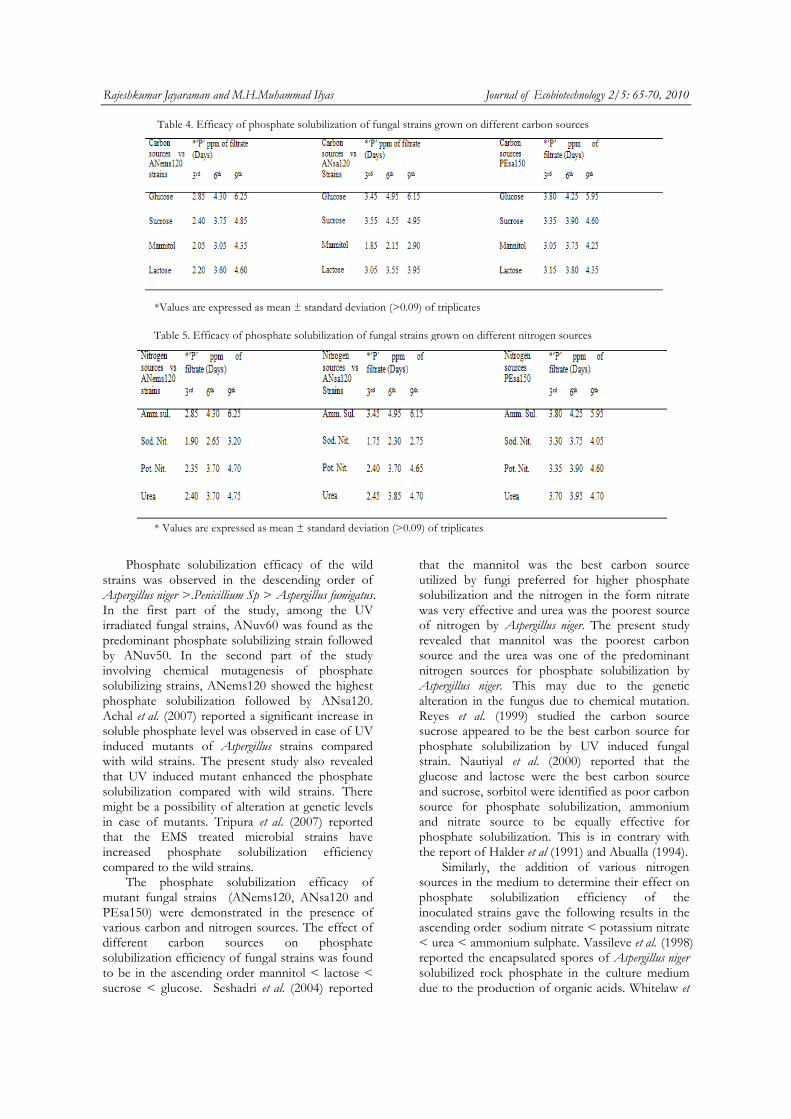
Rajeshkumar Jayaraman and M.H.Muhammad Ilyas Journal of Ecobiotechnology 2/5: 65-70, 2010
Table 4. Efficacy of phosphate solubilization of fungal strains grown on different carbon sources
*Values are expressed as mean ± standard deviation (>0.09) of triplicates
Table 5. Efficacy of phosphate solubilization of fungal strains grown on different nitrogen sources
* Values are expressed as mean ± standard deviation (>0.09) of triplicates Phosphate solubilization efficacy of the wild
strains was observed in the descending order of Aspergillus niger >.Penicillium Sp > Aspergillus fumigatus. In the first part of the study, among the UV irradiated fungal strains, ANuv60 was found as the predominant phosphate solubilizing strain followed by ANuv50. In the second part of the study involving chemical mutagenesis of phosphate solubilizing strains, ANems120 showed the highest phosphate solubilization followed by ANsa120. Achal et al. (2007) reported a significant increase in soluble phosphate level was observed in case of UV induced mutants of Aspergillus strains compared with wild strains. The present study also revealed that UV induced mutant enhanced the phosphate solubilization compared with wild strains. There might be a possibility of alteration at genetic levels in case of mutants. Tripura et al. (2007) reported that the EMS treated microbial strains have increased phosphate solubilization efficiency compared to the wild strains.
The phosphate solubilization efficacy of mutant fungal strains (ANems120, ANsa120 and PEsa150) were demonstrated in the presence of various carbon and nitrogen sources. The effect of different carbon sources on phosphate solubilization efficiency of fungal strains was found to be in the ascending order mannitol < lactose < sucrose < glucose. Seshadri et al. (2004) reported
that the mannitol was the best carbon source utilized by fungi preferred for higher phosphate solubilization and the nitrogen in the form nitrate was very effective and urea was the poorest source of nitrogen by Aspergillus niger. The present study revealed that mannitol was the poorest carbon source and the urea was one of the predominant nitrogen sources for phosphate solubilization by Aspergillus niger. This may due to the genetic alteration in the fungus due to chemical mutation. Reyes et al. (1999) studied the carbon source sucrose appeared to be the best carbon source for phosphate solubilization by UV induced fungal strain. Nautiyal et al. (2000) reported that the glucose and lactose were the best carbon source and sucrose, sorbitol were identified as poor carbon source for phosphate solubilization, ammonium and nitrate source to be equally effective for phosphate solubilization. This is in contrary with the report of Halder et al (1991) and Abualla (1994).
Similarly, the addition of various nitrogen sources in the medium to determine their effect on phosphate solubilization efficiency of the inoculated strains gave the following results in the ascending order sodium nitrate < potassium nitrate < urea < ammonium sulphate. Vassileve et al. (1998) reported the encapsulated spores of Aspergillus niger solubilized rock phosphate in the culture medium due to the production of organic acids. Whitelaw et

Rajeshkumar Jayaraman and M.H.Muhammad Ilyas Journal of Ecobiotechnology 2/5: 65-70, 2010
al. (1999) reported that the Penicillium strain solubilized the insoluble phosphate in the culture medium containing ammonium or nitrate as sole source of nitrogen. Vassileve et al. (2007) compared the phosphate solubilization efficacy of Aspergillus niger by using corn steep liquor, ammonium sulphate and yeast extract as nitrogen source.
The present study also indicates, the solubilization efficacy of the fungal strain Aspergillus niger treated with sodium azide (ANsa120) and Penicillium sp treated with EMS (PEems150) were about 1.5-fold higher when compared to wild strain. The chemical mutagenic agents such as sodium azide, ethyl methane sulphonate may alter the gene sequence by altering the base pairs. The azide ion alters the structure of cytosine such that it forms hydrogen bonds with adenine, rather than guanine. This produces a cytosine to thymine transition. Ethyl methane sulphonate is a strong mutagenic agent. It alkylates N7 of Guanine and severely alters the base pairing. 4. Conclusion
Many soil microorganisms are able to solubilize the unavailable forms of phosphate through their metabolic activities. The present study concluded that the treatment of physical and chemical mutagenic agents in fungal strains increased the phosphate solubilization efficacy of the fungal strains. This study also strengthened the idea that addition of carbon and nitrogen sources favors the phosphate solubilization to a certain extent. Acknowledgements
The authors greatly acknowledge the management and the principal of Jamal Mohammed College, Trichy, Tamil Nadu, India for providing the facility. The authors also acknowledge the support rendered by Dr. N.Sengottaian, Head, Department of Botany and Microbiology, Urumu Dhanalakshmi College, Trichy, Tamil Nadu, India. We also thank Mr. P. Malaiarasa Pandian and Miss. Sangeetha Menon for helpful discussion.
References Abd-Alla, M.H., 1994. Phosphatases and the
utilization of organic P by Rhizobium leguminosarum biovar viceae. Biology and fertility of soils 18, 294-296.
Achal, V., Savant V.V., Reddy, M.S., 2007. Phosphate solubilization by a wild type strain and UV-induced mutants of Aspergillus thubingensis. Soil Biology & Biochemistry 39, 695�699.
Agnihotri, V.P., 1970. Solubilization of insoluble phosphate by some soil fungi isolated from nursery seed beds. Canadian Journal of Microbiology 16, 877-880.
Dave, A., Patel, H.H., 1999. Inorganic phosphate solubilizing soil Pseudomonads. Indian Journal of Microbiology 39, 161-164.
Bapiraju, K.V.V.S.N., Sujatha, P., Ramana, T., 2004. Mutation induced enhanced biosynthesis of lipase. African Journal of Biotechnology 3, 618-621.
Bardiya, M.C., Gaur, A.C., 1972. Rock phosphate dissolution by bacteria. Indian Journal of Microbiology 12, 269-271.
Elliaiah, P., Prabhakar, T., Ramakrishna, B., Thaer Taleb, A., Adinarayana, K., 2002. Strain improvement of Aspergillus niger for the production of lipase. Indian Journal of Microbiology 42, 151-153.
Halder, A.K., Mishra, A.K., Chakrabartty, P.K., 1991. Solubilization of inorganic phosphates by Bradyrhizobium. Indian Journal of Experimental Biology 29, 28-31.
Hayman, D.S., 1975. Phosphorus cycling by soil microorganisms and plant roots In: soil microbiology. Butterworths, London, pp. 67-91.
Jayasinghearachchi, H.S., Seneviratne, G., 2006. Fungal solubilization of rock phosphate is enhanced by forming fungal�rhizobial biofilms. Soil Biology & Biochemistry 38, 405�408.
Nautiyal, C.S., Bhadauria, S., Kumar, P., Lal, H., Mond, L. R., Verma, D., 2000. Stress induced phosphate solubilization in bacteria isolated from alkaline soils. FEMS Microbiological Letters 182, 291-296.
Narsian, V., Jungu, T., Patel, H.H., 1995. Mineral phosphate solubilization by Aspergillus aculeatus. Indian Journal of Experimental Biology 33, 91-93.
Narsian, V., and Patel, H.H., 2000. Aspergillus aculeatus as a rock phosphate solubilizer. Soil Biology & Biochemistry 32, 559�565.
Pradhan, N., Sukla, L.B., 2005. Solubilization of inorganic phosphate by fungi isolated from agriculture soil. African Journal of Biotechnology 5, 850-854
Ostwal, K.P., Bhide, V.P., 1972. Solubilization of TCP by soil Pseudomonas. Indian Journal of Experimental Biology 10, 153-154.
Reyes, I., Bernier, L., Simard, R.R., Tanguary, P., Antoun, H., 1999. Characteristics of phosphate solubilization by an isolate of a tropical Penicillium rugulosum and two UV induced mutants. FEMS Microbiology Ecology 28, 291-295.
Rosas, S.B., Andres, J.A., Rovera, M., Correa, N.S., 2006. Phosphate-solubilizing Pseudomonas putida

Rajeshkumar Jayaraman and M.H.Muhammad Ilyas Journal of Ecobiotechnology 2/5: 65-70, 2010
can influence the rhizobia�legume symbiosis. Soil Biology & Biochemistry 38, 3502�3505.
Seshadri, S., 1995. Phosphate solubilizing fungi from Thanjavur district. South India.,Ph.D., thesis, Bharathidasan University.
Seshadri, S., Ignacimuthu, S., Lakshminarasimhan, C. 2004. Effect of nitrogen and carbon sources on the inorganic phosphate solubilization by different Aspergillus niger strains. Chemical Engineering Communications 191(8): 1043-1052.
Sperber, J.I., 1957. Solubilization of mineral phosphate by soil bacteria., Nature 180, 994-95.
Tripura, C., Sashidhar, B., Podile, A.R., 2007. Ethyl methanesulfonate mutagenesis-enhanced mineral phosphate solubilization by groundnut-associated Serratia marcescens GPS-5. Current Microbiology 54, 79-84.
Vassilev, N., Vassileva, M., Bravo,V., Serrano, M.F., Nikolaeva,I., 2007. Simultaneous phytase production and rock phosphate solubilization by Aspergillus niger grown on dry olive wastes. Industrial Crops and Products 26, 332�336.
Vassileva, M., Azcon, R., Barea, J.M., Vassilev, N., 1998. Application of an encapsulated filamentous fungus in solubilization of inorganic phosphate. Journal of Biotechnology 63, 67-72.
Vivekkumar, Punia, S., Lakshminarayana, K., Narula, N., 1999. Effect of phosphate solubilizing analogue resistant mutants of agriculture Azotobacter chroococcum on sorghum. Indian Journal of Agricultural Science 69, 198-200.
Whitelaw, M.A ., Harden, T.J., Helyar, K.R., 1999. Phosphate solubilization in solution culture by the soil fungus Penicillium radicum. Soil Biology & Biochemistry 31, 655-665.

Regular Article
Strain Improvement of sp for the Production of Lipase
Rajeshkumar Jayaraman and M.H. Muhammad Ilyas*
Department of Botany, Jamal Mohammed College, Trichy-620020
ABSTRACT: sp. capable of extracellular lipase production was isolated from rhizosphere soil of paddy fields in Tamilnadu, India. The isolated strain was mutated using ultra-violet rays and chemical agents like Sodium Azide and EMS. The study indicated that the lipase activity of the mutant strain using chemical mutagen was 2-fold higher than the wild strain.
Key words: ; Lipase; Mutagenesis; UV; Sodium Azide; EMS Abbreviation: SA: Sodium azide, EMS: Ethyl methane sulphonate, UV: Ultra Violet, PSsa: Sodium azide treated sp.PSems: Ethyl methane sulphonate treated sp. PSuv: Ultra Violet treated sp.
Introduction
Lipase enzyme plays a vital role in many food, dairy, leather, paper, pharmaceutical, detergent, textile and cosmetic industries. Lipases are secreted by microorganisms like bacteria, yeasts, molds and a few protozoa. The production of lipase by microorganisms depends largely on the species, strains and culture conditions. Microbial lipases are diverse in their enzymatic properties, substrate specificity and are usually more thermo stable than animal or plant lipases (Vanitha, 2002). Research on lipase production has intensified in recent years due to its potential application in various industrial processes. There are several reports on isolation of lipase producing microorganisms and the effect of nutritional factors on their growth and lipase production (Kamini ., 1997; Gisela ., 1991; Sangilyandi and Gunasekaran, 1996). Jette and Ziomek (1994) determined the lipase activity by a Rhodamine-Triglyceride-Agarose assay. Several species of have been reported to produce this enzyme (Suziki 1988; Narasaki 1968; Sugiura 1977 and Sugiura and Oikawa, 1977). However, no report is available on lipase production by mutated species. Therefore, the present study has been undertaken to study the efficacy of lipase production after inducing mutation in
strains isolated from rhizophere soil of the paddy fields.
Materials and Methods Microorganism
strain isolated from rhizosphere soils of paddy fields in Mannachanallur, one of the largest paddy producing area located in Trichy district of Tamil Nadu, India. The strain was sub-cultured on nutrient agar slant and incubated at 37ºC for 48hrs. This strain was stored at 4ºC and used as the parent (Wild strain) culture.
Mutagenesis by UV
The 24 hr old nutrient broth of the parent culture was diluted with phosphate buffer (pH 7.0) to contain 108 cells/ml. The culture was spread plated on nutrient agar plates. Then the plates were exposed for various time intervals (1 to 10 min) to UV light (2600 Ao) at a distance of 15cm (Kumar and Dhruv, 1990). After the stipulated time interval, each set of plates were covered with black paper to avoid photo reactivation and incubated at 37ºC for 2 days. The unexposed plate containing the wild strain served as positive control. On incubation, the resistant strains were selected based on colony development in nutrient agar plates and given code names as PSuv 1, PSuv 2, PSuv 3 and PSuv 4 .These strains were transferred to nutrient agar slants and further screened for lipase activity.
Mutagenesis by Chemical
Sodium azide and ethyl methane sulphonate (EMS) were the chemical mutagens used for mutagenesis of the parent culture,
sp. The bacterial suspension was diluted in similar way as that for U.V mutagenesis. To nine ml of bacterial suspension, one ml of sterile solution of sodium azide (50 g ml-1 in phosphate buffer) was added. Similar method was adopted for ethyl methane sulphonate (50 g ml-1 in phosphate buffer). The reaction was allowed to proceed. Samples were withdrawn from the reaction mixture at intervals of 30, 60, 90, 120, 150 and 180 min and immediately centrifuged for 10 min at 5000 rpm. The pellet was washed three times with sterile phosphate buffer and resuspended in 10 ml of the same buffer (pH 7.0). The samples were serially diluted using the same buffer and plated on nutrient agar. The plates were incubated at 37ºC for two days. The selected sodium azide treated mutants were designated as PSsa30, PSsa60, PSsa90 and PSsa120, whereas ethyl methane sulphonate treated mutants were designated as PSems60, PSems90, PSems120 and PSems150.
Submerged fermentation
18 hr old culture broth of wild type and mutant strains of sp. was used for this study. 0.5 ml of each strain (O.D
= 1.0) was inoculated into 100ml Czapek-Dox broth containing 1% olive oil as inducer. Sucrose and 10% olive oil as the carbon source. The flasks were incubated at 37°C for 96hrs. Uninoculated Czapekdox broth served as control. Each experiment was done in triplicates.
Lipase assay
Culture broth was withdrawn aseptically at three days interval from each flask and centrifuged at 3000 rpm for 15 minutes and the supernatant was collected and used for lipase activity (Ray .,1999). Lipase activity in culture broth was determined by titrimetry using olive oil as substrate (Vanitha, 2002).
Results and discussion
Lipase activity of sp treated by ultraviolet rays, sodium azide and ethyl methane sulphonate are presented in the Table 1, 2 and 3 respectively. The results indicated that lipase activity was more in sucrose medium than in olive oil medium after 96 hrs of UV Sodium azide and EMS treatments. The enzyme activity was found to be higher in mutant strains than the wild type (Table 1, 2 and 3). Efficacy of lipase activity by the mutant strains was observed in the descending order of EMS mutants > SA mutants > UV mutants. In the first part of the study, among the UV irradiated bacterial strains, PSuv3 was found as the predominant lipase producing strain followed by PSuv4. In the second part of the study involving chemical mutagenesis, PSems120 showed the highest lipase activity followed by PSems150.
The lipase activity considerably increased depending on nutritional conditions. Similar observations were made by Lawrence (1967) in (Suzuki 1988). Ray (1999) reported the lipase activity increased in isolated bacterial strains by chemical mutagens. Ellaiah (2002) reported that isolated fungal strain produced lipase with physical and chemical mutagens and the mutant has higher lipase activity than the wild strain. The increase in the production of lipase by the mutagens both physical and chemical may be due to the alternation of genotype of the micro organisms. Moreover, they may alter the gene sequence

(Freifelder, 1990; Radman, 1999). The azide ion alters the structure of cytosine such that it forms hydrogen bonds with adenine, rather than guanine. This produces a cytosine to thymine transition. Ethyl methane sulphonate is a strong mutagenic agent. It alkylates N7 of
Guanine and severely alters the base pairing. The present study also indicated that the strain of sp treated with EMS produced 2-fold (PSems120) higher amount of lipase compared to wild strain.
Table 1. Lipase activity of sp treated by ultraviolet rays
UV treated sp strains Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of Lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
PSuv1 2.22±0.01 0.23±0.02 5.71 29.31
PSuv2 2.19±0.09 0.33±0.08 4.28 91.95
PSuv3 2.49±0.07 0.38±0.03 18.57 118.39
PSuv4 2.29±0.02 0.34±0.05 9.04 92.52
Wild typea 2.10±0.08 0.17±0.06
*Values are expressed as mean ± standard deviation of triplicatesaStandard for calculation of increased %
Table 2. Lipase activity of sp treated by sodium azide
Sodium azide treated sp strains
Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
PSsa30 2.28±0.04 0.28±0.11 8.57 58.05
PSsa60 2.93±0.03 0.35±0.02 39.52 103.45
PSsa90 3.23±0.05 0.39±0.08 53.80 124.14
PSsa120 3.04±0.09 0.39±0.04 44.76 121.26
Wild typea 2.10±0.06 0.17±0.06
*Values are expressed as mean ± standard deviation of triplicates aStandard for calculation of increased %
Table 3. Lipase activity of sp treated by EMS
EMS treated sp strains Lipase activity after 96 hrs(Unit/g of substrate)*
Increased % of lipase activity
Sucrose medium
Olive oil medium
Sucrose medium
Olive oil medium
PSems60 2.42±0.10 0.34±0.06 9.50 54.55
PSems90 3.53±0.05 0.47±0.09 59.72 113.64
PSems120 4.45±0.07 0.61±0.05 101.36 177.27
PSems150 4.12±0.04 0.52±0.05 86.43 136.36
Wild typea 2.21±0.09 0.22±0.12
*Values are expressed as mean ± standard deviation of triplicates aStandard for calculation of increased %
Lipases are enzymes that catalyze the hydrolysis of triglycerides to glycerol and fatty acids Microbial lipases are relatively stable and are capable of catalyzing a variety of reactions: they are potentially of importance for divorce industrial applications. The present study concluded the addition of chemical agents in bacterial strains may produce the higher quantity of lipase enzyme.
Acknowledgements The authors greatly acknowledge the management and the
principal of Jamal Mohammed College, Trichy, Tamil Nadu, India for providing the facility. The authors also acknowledge the support rendered by Dr. N.Sengottaian, Head, Department of Botany and Microbiology, Urumu Dhanalakshmi College, Trichy, Tamil Nadu, India. We also thank Mr. P. Malaiarasa pandian and Miss. Sangeetha menon for helpful discussion.
References Elliaiah, P, Prabhakar, T, Ramakrishna, B, Thaer Taleb, A,
Adinarayana, K, (2002) Strain improvement of
for the production of lipase. Indian Journal of Microbiology 42:151-153.
Freifelder D (1990) Microbial Genetics. Narosa Publishing House, New Delhi, India. pp-191-210.
Gisela M, Dellamora O, Reneta CM, William LR, Andrea PD (1997) Activity and stability of lipase in hydrophobic media. Biotechnology Applied Biochemistry 26: 31-37.
Jette JF, Ziomek E (1994) Determination of lipase activity by a Rhodamine-Triglyceride-Agarose Assay. Analytical Biochemistry 219: 256-260.
Kamini NR, Mala JGS, Kumar PR (1997) Production and characterization of an extracellular lipase from .Indian Journal of Microbiology 37: 85-89.
Kumar D, Kumar HD (1990) Nature of UV resistance in the cyanobacterium N . Current Science 59: 412-414.
Lawrence RC, Fryer TF, Reiter B (1967) The production and characterization of lipases from a microbial and Pseudomonoid. Journal of General Microbiology 48: 401-418.

Ray N, Ray L, Srimani BN (1999) Isolation and identification of Alkaline thermostable lipase producing microorganism, cultural Conditions, nutritional requirements and some reports of crude Enzyme. Indian Journal of Experimental Biology 37: 818-824.
Sangilyandi G, Gunasekharan P (1996) Extracellular lipase producing from an oil mill refinery effluent. Indian
Journal of Microbiology 36: 109-110. Sugiura M, Oikawa T (1977) Physicochemical properties of a lipase
from . Biochemistry Biophysics Acta 489: 262-268.
Sugiura M, Oikawa T, Hirano K, Inukai T (1977) Purification, crystallization and properties of triacylglycerol lipase from
Biochemistry Biophysics Acta 489: 353-358.
Suzuki T, Mushiga Y, Yamane T, Shimizu S (1988) Mass production of lipase by fed- batch culture of .Applied Microbiology and Biotechnology 27: 417-422.
Sztajer H, Maliszweska I, Wieczorek J (1988) Production of exogenous lipases by bacteria, fungi and actinomycetes. Enzyme Microbial technology 10: 492-497.
Vanitha S (2002) Lipase production by growing on coconut kernel. Master of Philosophy in microbiology thesis Bharathidasan University. pp- 46-53.
Radman M, (1999) Enzymes of evolutionary change. Nature 401: 866-868.




















