
Protein-thiol substitution or protein dethiolation by thiol/disulfide exchange reactions: the...
10
proteins STRUCTURE FUNCTION BIOINFORMATICS Protein–thiol substitution or protein dethiolation by thiol/disulfide exchange reactions: The albumin model Domenico Summa, 1 Ottavia Spiga, 2 Andrea Bernini, 2 Vincenzo Venditti, 2 Raffaella Priora, 1 Simona Frosali, 1 Antonios Margaritis, 1 Danila Di Giuseppe, 1 Neri Niccolai, 2 and Paolo Di Simplicio 1 * 1 Department of Neuroscience, Pharmacology Unit, University of Siena, 53100 Siena, Italy 2 Department of Molecular Biology, University of Siena, 53100 Siena, Italy INTRODUCTION Protein S-thiolation, formation of protein–thiol mixed disulfide, occurs in all biological environments by two mechanisms involving thiol/disulfide (SH/SS) exchange reactions: XSSX1Alb-SH$XSH1 Alb-SS-X [1] and/or sulfenic acid formation, Alb-SOH1XSH?Alb- SS-X1H 2 O [2]. It protects the SH enzymes from irreversible oxida- tion to sulfinic or sulfonic acids caused by ROS, modulates cell sig- naling, and by reaction [1] buffers disulfide excesses and regenerates thiols (namely GSH). 1–6 Protein dethiolation, the reverse of reaction [1], is different in plasma than in the cell environment. In plasma, it is not supported by enzymes (glutaredoxin) and the GSH regenerating system (hexose monophosphate shunt), 1 and the thiolation/dethiolation process is almost exclusively sustained by albumin. Plasma albumin has a theoretical concentration of 0.6 mM, only one SH group, and several disulfide bridges. The cysteinyl residue, located in a well-conserved region in position 34 in all mammalian species, has a low pK a (about 5–7) 7–9 because of a salt bridge, with His39 that stabilizes the thiolate anion. It is well-known that albu- min SH is not well-exposed, which impedes development of its high potential reactivity related to its low pK a . Indeed, the reaction rate of albumin towards SH reagents (DTNB) is lower than that of thiol ABSTRACT Dethiolation experiments of thiolated albumin with thionitrobenzoic acid and thiols (glutathione, cysteine, homocysteine) were carried out to understand the role of albumin in plasma distribution of thiols and disul- fide species by thiol/disulfide (SH/SS) exchange reac- tions. During these experiments we observed that thiolated albumin underwent thiol substitution (Alb- SS-X1RSH$Alb-SS-R1XSH) or dethiolation (Alb- SS-X1XSH$Alb-SH1XSSX), depending on the dif- ferent pK a values of thiols involved in protein–thiol mixed disulfides (Alb-SS-X). It appeared in these reac- tions that the compound with lower pK a in mixed disulfide was a good leaving group and that the pK a differences dictated the kind of reaction (substitution or dethiolation). Thionitrobenzoic acid, bound to albu- min by mixed disulfide (Alb-TNB), underwent rapid substitution after thiol addition, forming the corre- sponding Alb-SS-X (peaks at 0.25–1 min). In turn, Alb-SS-X were dethiolated by the excess nonprotein SH groups because of the lower pK a value in mixed disul- fide with respect to that of other thiols. Dethiolation of Alb-SS-X was accompanied by formation of XSSX and Alb-SH up to equilibrium levels at 35 min, which were different for each thiol. Structures by molecular simu- lation of thiolated albumin, carried out for under- standing the role of sulfur exposure in mixed disulfides in dethiolation process, evidenced that the sulfur expo- sure is important for the rate but not for determining the kind of reaction (substitution or dethiolation). Our data underline the contribution of SH/SS exchanges to determine levels of various thiols as reduced and oxi- dized species in human plasma. Proteins 2007; 69:369–378. V V C 2007 Wiley-Liss, Inc. Key words: protein dethiolation; protein–thiol sub- stitution; SH/SS exchange reactions; albumin; bio- informatics; protein–thiol mixed disulfides. Abbreviations: Alb, albumin; AlbHIS, albumin with unprotonated histidine in position 39; AlbHIS1, albumin with protonated histidine in position 39; Alb-SH, mercaptoalbumin; Alb-SOH, protein sulfenic acid; Alb-SS-C, cysteine–protein mixed disulfide; Alb-SS-H, homocysteine-protein mixed disulfide; Alb-SS-X, Alb-SS-R, protein–thiol mixed disulfide; Alb-TNB, mixed disulfide between thionitrobenzoic acid and albumin; BSA, bovine serum albumin; CSH, reduced cysteine; CSSC, cystine; CSSH, heterodimer disulfide between cysteine and homocysteine; DTNB, 5,5 0 - dithiobis(2-nitrobenzoic acid); DTT, dithiothreitol; GSH, glutathione and reduced glutathione; GSSG, glutathione disulfide; His39, histidine in position 39; HSA, human serum albumin; HSH, reduced homocysteine; HSSG, heterodimer disulfide between homocysteine and glutathione; HSSH, homocystine; mBrB, monobromobimane; MD simulation, molecular dynamic simulation; ROS, reactive oxygen species; RSH, XSH, reduced low molecular weight thiols; SH/SS, thiol/disul- fide; tALB, total albumin; TCA, trichloroacetic acid; tGSH, total glutathione; tHcy, total homocys- teine; TNB, thionitrobenzoic acid; XSSX, homodimer non protein disulfide. Grant sponsors: SIGMA-TAU, PRIN, PAR. *Correspondence to: Paolo Di Simplicio. E-mail: [email protected] Received 20 September 2006; Revised 13 February 2007; Accepted 22 March 2007 Published online 2 July 2007 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/prot.21532 V V C 2007 WILEY-LISS, INC. PROTEINS 369
Transcript of Protein-thiol substitution or protein dethiolation by thiol/disulfide exchange reactions: the...

proteinsSTRUCTURE O FUNCTION O BIOINFORMATICS
Protein–thiol substitution or proteindethiolation by thiol/disulfide exchangereactions: The albumin modelDomenico Summa,1 Ottavia Spiga,2 Andrea Bernini,2 Vincenzo Venditti,2
Raffaella Priora,1 Simona Frosali,1 Antonios Margaritis,1 Danila Di Giuseppe,1
Neri Niccolai,2 and Paolo Di Simplicio1*1Department of Neuroscience, Pharmacology Unit, University of Siena, 53100 Siena, Italy
2Department of Molecular Biology, University of Siena, 53100 Siena, Italy
INTRODUCTION
Protein S-thiolation, formation of protein–thiol mixed disulfide,
occurs in all biological environments by two mechanisms involving
thiol/disulfide (SH/SS) exchange reactions: XSSX1Alb-SH$XSH1Alb-SS-X [1] and/or sulfenic acid formation, Alb-SOH1XSH?Alb-
SS-X1H2O [2]. It protects the SH enzymes from irreversible oxida-
tion to sulfinic or sulfonic acids caused by ROS, modulates cell sig-
naling, and by reaction [1] buffers disulfide excesses and regenerates
thiols (namely GSH).1–6
Protein dethiolation, the reverse of reaction [1], is different in
plasma than in the cell environment. In plasma, it is not supported
by enzymes (glutaredoxin) and the GSH regenerating system (hexose
monophosphate shunt),1 and the thiolation/dethiolation process is
almost exclusively sustained by albumin.
Plasma albumin has a theoretical concentration of 0.6 mM, only
one SH group, and several disulfide bridges. The cysteinyl residue,
located in a well-conserved region in position 34 in all mammalian
species, has a low pKa (about 5–7)7–9 because of a salt bridge, with
His39 that stabilizes the thiolate anion. It is well-known that albu-
min SH is not well-exposed, which impedes development of its high
potential reactivity related to its low pKa. Indeed, the reaction rate
of albumin towards SH reagents (DTNB) is lower than that of thiol
ABSTRACT
Dethiolation experiments of thiolated albumin with
thionitrobenzoic acid and thiols (glutathione, cysteine,
homocysteine) were carried out to understand the role
of albumin in plasma distribution of thiols and disul-
fide species by thiol/disulfide (SH/SS) exchange reac-
tions. During these experiments we observed that
thiolated albumin underwent thiol substitution (Alb-
SS-X1RSH$Alb-SS-R1XSH) or dethiolation (Alb-
SS-X1XSH$Alb-SH1XSSX), depending on the dif-
ferent pKa values of thiols involved in protein–thiol
mixed disulfides (Alb-SS-X). It appeared in these reac-
tions that the compound with lower pKa in mixed
disulfide was a good leaving group and that the pKa
differences dictated the kind of reaction (substitution
or dethiolation). Thionitrobenzoic acid, bound to albu-
min by mixed disulfide (Alb-TNB), underwent rapid
substitution after thiol addition, forming the corre-
sponding Alb-SS-X (peaks at 0.25–1 min). In turn,
Alb-SS-X were dethiolated by the excess nonprotein SH
groups because of the lower pKa value in mixed disul-
fide with respect to that of other thiols. Dethiolation of
Alb-SS-X was accompanied by formation of XSSX and
Alb-SH up to equilibrium levels at 35 min, which were
different for each thiol. Structures by molecular simu-
lation of thiolated albumin, carried out for under-
standing the role of sulfur exposure in mixed disulfides
in dethiolation process, evidenced that the sulfur expo-
sure is important for the rate but not for determining
the kind of reaction (substitution or dethiolation). Our
data underline the contribution of SH/SS exchanges to
determine levels of various thiols as reduced and oxi-
dized species in human plasma.
Proteins 2007; 69:369–378.VVC 2007 Wiley-Liss, Inc.
Key words: protein dethiolation; protein–thiol sub-
stitution; SH/SS exchange reactions; albumin; bio-
informatics; protein–thiol mixed disulfides.
Abbreviations: Alb, albumin; AlbHIS, albumin with unprotonated histidine in position 39;
AlbHIS1, albumin with protonated histidine in position 39; Alb-SH, mercaptoalbumin; Alb-SOH,
protein sulfenic acid; Alb-SS-C, cysteine–protein mixed disulfide; Alb-SS-H, homocysteine-protein
mixed disulfide; Alb-SS-X, Alb-SS-R, protein–thiol mixed disulfide; Alb-TNB, mixed disulfide
between thionitrobenzoic acid and albumin; BSA, bovine serum albumin; CSH, reduced cysteine;
CSSC, cystine; CSSH, heterodimer disulfide between cysteine and homocysteine; DTNB, 5,50-dithiobis(2-nitrobenzoic acid); DTT, dithiothreitol; GSH, glutathione and reduced glutathione;
GSSG, glutathione disulfide; His39, histidine in position 39; HSA, human serum albumin; HSH,
reduced homocysteine; HSSG, heterodimer disulfide between homocysteine and glutathione;
HSSH, homocystine; mBrB, monobromobimane; MD simulation, molecular dynamic simulation;
ROS, reactive oxygen species; RSH, XSH, reduced low molecular weight thiols; SH/SS, thiol/disul-
fide; tALB, total albumin; TCA, trichloroacetic acid; tGSH, total glutathione; tHcy, total homocys-
teine; TNB, thionitrobenzoic acid; XSSX, homodimer non protein disulfide.
Grant sponsors: SIGMA-TAU, PRIN, PAR.
*Correspondence to: Paolo Di Simplicio. E-mail: [email protected]
Received 20 September 2006; Revised 13 February 2007; Accepted 22 March 2007
Published online 2 July 2007 in Wiley InterScience (www.interscience.wiley.com).
DOI: 10.1002/prot.21532
VVC 2007 WILEY-LISS, INC. PROTEINS 369

with higher pKa.10,11 Moreover glutathione disulfide does
not react with albumin.12 Compounds such as fatty acids
that change albumin conformation, improving Cys34 ex-
posure, lead to higher reactivity of albumin SH.10,11
In this study albumin was used to understand the influ-
ence of SH/SS exchange reactions to determine the levels
of various thiols as reduced and oxidized species in human
plasma. Plasma disulfide concentration, consisting of
XSSX and Alb-SS-X, is higher than thiol concentration
and no relationships seem to exist among all these redox
species in plasma. Alb-SS-C and CSSC are the most abun-
dant disulfides13: Alb-SS-C is about double CSSC and 10
times more abundant than other Alb-SS-Xs, such as Alb-
SS-H and Alb-SS-G.13,14 Although the plasma concentra-
tion of thiols as reduced and oxidized are considered de-
pendent upon the pro-oxidant nature of plasma (thiols
and other reducing agents are rapidly oxidized in
plasma),11,15 other factors different from oxidations must
remodel their concentration. For example, GSH and HSH
have a concentration range in normal individuals13,14 not
usually conformable to their corresponding oxidation
rates.15
Since 40% of albumin is bound as mixed disulfide, it
has the requisites for fulfilling the regulation buffer func-
tion of plasma thiols and disulfides by SH/SS exchange
reactions, as suggested by methionine load experiments.16
Two SH/SS exchange reactions are possible: Alb-
SS-X1RSH$Alb-SS-R1XSH, henceforth called thiol sub-
stitution (formation of another species of protein-mixed
disulfide) or Alb-SS-X1XSH$Alb-SH1XSSX, the pro-
tein dethiolation with Alb-SH formation.
We used a biochemical approach, a molecular simula-
tion of differently thiolated albumin, and computational
chemistry of mixed disulfides to study SH/SS exchange by
protein substitution and dethiolation of thiolated albumin
with TNB.
Our results show that albumin can be subjected to dif-
ferent metabolic fates (substitution or dethiolation). The
difference of pKa of thiols involved in protein–thiol mixed
disulfides is an important feature to determine the kind of
reaction (substitution or dethiolation) and consequently
the end products. Moreover, the rate of SH/SS exchange
reactions is higher for those Alb-SS-X having a better ex-
posure of the sulfur subjected to nucleophilic attack.
EXPERIMENTAL PROCEDURES
Reagents: L-Cysteine, DL-homocysteine, glutathione, and
BSA 96% Fr V (free of fatty acid), trichloroacetic acid,
monobromobimane, dithiothreitol, and other regents
from Sigma-Aldrich, Italy. Ellman’s reagent (5,50-dithio-bis(2-nitrobenzoic acid), DTNB) was from CALBIO-
CHEM (USA).
Instruments: UV–VIS V550 spectrophotometer JASCO
(Tokyo, Japan); Hewlett Packard HPLC series 1100 (Palo
Alto, CA) with auto sampler and fluorimetric detector.
Biochemical methods
Preparation of mercaptoalbumin
A 0.830 mM BSA solution in 0.1M phosphate buffer 10.3M NaCl 1 5 mM of DTT, pH 6.86 was prepared and
maintained at room temperature for 45 min according to
Sengupta et al.17 The albumin was then dialyzed against
0.1 M phosphate buffer until the thiols disappeared from
the buffer.
Preparation of mixed disulfide albumin with TNB
The dialysis tube was placed in a solution of 0.1M phos-
phate buffer pH 7.4 1 7.5 mM DTNB for 12 h and then
dialyzed against phosphate buffer until DTNB and TNB
detected at 326 and 410, respectively disappeared from the
buffer. The absence of Alb-SH in TNB-ALB stock and thiol
were assayed with Ellman’s reagent.
Alb-TNB reaction with thiols
Two experiments were performed. In the first experi-
ment, Alb-TNB (0.830 mM) was reacted separately with
each thiol (CSH, HSH, GSH) (15 mM) by 1:1dilution. In
the second experiment, the same amount of Alb-TNB was
reacted with a mixture (15 mM) containing equal concen-
trations of thiols. This experiment was repeated using a
3 mM overall concentration of thiols. In each experiment
the reaction was stopped at different times by precipitation
with 17% TCA (w/v) (final concentration). The albumin
pellet was then washed with 1.5% TCA (w/v) to eliminate
thiols and disulfides and used to assay Alb-SH and pro-
tein–thiol mixed disulfides.
Dethiolation of Alb-SS-C by HSH
A solution of BSA (600 lM total containing 200 lMAlb-SS-C) was reacted with a 500 lM HSH solution by
1:1 dilution. The reaction was stopped at different times
by precipitation with 17% TCA (w/v) (final concentra-
tion), and the pellet was assayed for Alb-SS-X.
Alb-SH measurement
After washing in TCA, the pellet was dissolved with
400 lL 0.1M phosphate buffer (pH 7.4). Three hundred
microliters of sample was added to phosphate buffer in a
cuvette and assayed spectrophotometrically with DTNB
(20 lL 20 mM DTNB, end of reaction 15 min).18
Alb-SS-X measurement
Alb-SS-X was assayed according to Di Giuseppe et al.13
The pellet was resuspended with 0.4 mL 2 mM DTT in
water and maintained at neutral pH with saturating
NaHCO3 for 20 min. After centrifuging, the supernatant
(100 lL) was deproteinized with 6% TCA (w/v) (final
concentration) and centrifuged again. The supernatant
D. Summa et al.
370 PROTEINS DOI 10.1002/prot

was incubated with 1 mM mBrB at neutral pH under satu-
rating conditions of solid NaHCO3 in the dark for 15 min
to derivatize soluble thiols. After centrifuging, 100 lLsupernatant was acidified with 10 lL of 37% HCl (to pH
of 3–4) and injected (injection volume 0.02 mL) into the
HPLC separation column (ChromSep C18 250 3 4 mm,
Varian, Lake Forest, CA USA). Elution conditions were:
solvent A at 0.25% (v/v) acetic acid (pH 3.09) and solvent
B methanol. The elution profile was: 0–8 min, 20% solvent
B; 8–15 min, 20–40% solvent B; 15–25 min, 40–100%
solvent B (flow rate 1.0 mL/min). Thiol derivatives were
detected fluorimetrically exciting at 390 nm and reading at
410 nm. Retention times were 6.0, 10.1, and 12.2 min for
CSH, HSH, and GSH, respectively.
Calibration curves were constructed after addition of
thiols and disulfides to phosphate buffer. The detection
limit was 2 pmol/injection.
Bioinformatics methods
Homology modeling
The 3D homology model of BSA was built from the
template crystal structure of HSA PDB ID 1AO6 using
DeepView v.3.7 SP5 software.19 Sequence alignment was
carried out with ClustalW v.1.820 with 75% of pairwise
sequence identities and 18% of similarity. The 3D model
was optimized by a 10,000 step minimization run with
GROMACS v.3.221 To validate the goodness of obtained
structures two tests were performed. The first test was to
examine the distribution of u and w angles using Rama-
chandran plot generated with PROCHECK v. 3.5.4 proce-
dure.22 The second test was to apply energy criteria using
PROSAII v. 3.2 analysis.23 In this program, the Prosa Z-
scores and energy profiles were calculated for BSA model
to assess their reliability.
Modification of residues
Four new residues were drawn using Hyperchem v.6.0
(Hypercube, FL) and their parameters and topologies were
obtained using a Dundee PRODRG2 server24 based on
GROMACS force-field gmx43a2.21
Molecular dynamic simulation
The four mutated structures were used for molecular
dynamic (MD) simulations. All simulations were carried
out using the GROMACS simulation package21 with the
GROMOS force-field (version 3.1.4). Each structure was
simulated using the following protocol: (i) the system was
minimized by 10,000 steps of conjugate gradients; 100 ps
of MD simulation in vacuum was performed to remove
bad contacts; (ii) the protein was placed at the centre of a
periodic cubic box, which was filled with SPC water mole-
cules and ions to obtain a net charge of zero; to relax the
configuration of the solvent, a steepest descent minimiza-
tion was carried out; (iii) individual MD simulation was
carried out at 300 K with a time-step of 2.0 fs for 5000 ps;
MD simulations were performed at constant temperature
and pressure (300 K and 1 atm respectively); the same pro-
tocol was used to perform MD simulation of native bovine
albumin structure with negatively charged Cys34 at differ-
ent states of protonation of His39 (positively charged
His1 and neutral His).
Atom depth calculation
The results of this simulation were analyzed using
SADIC software, calculating the atom depth of each modi-
fied residue.25
Electrostatic energy computations
The atomic partial charges of sulfur involved in disul-
fide were calculated with the semiempirical quantum me-
chanical method AM1 (HyperChem version 6) on average
structures obtained by MD simulation and on CSSH,
CSSG, and HSSG drawn and geometrically optimized by
HyperChem version 6.
RESULTS
Alb-TNB dethiolation by thiols: Alb-SS-Xsubstitution and dethiolation
Reactions of excess thiols (7.5 mM) with Alb-TNB
mixed disulfides (0.415 mM) were accompanied by rapid
massive substitution of TNB, marked by an increase in
absorption at 410 nm because of free chromophore and an
increase in Alb-SS-X concentration (Figs. 1 and 2).
Alb-SS-X formed during TNB substitution was later
consumed by dethiolation because of the excess of thiol
(Figs. 1 and 2). The dethiolation effect was an increase in
free albumin (Alb-SH). The sum of Alb-SH and Alb-SS-X
was constant over time, proving that the Alb-SH increase
was a consequence of dethiolation (Figs. 1 and 2). Further
evidence of albumin dethiolation was an increase in XSSX
concentration (not reported) similar to that of Alb-SH.
Figure 1 shows typical time-courses of Alb-SS-C dethio-
lation and increase in Alb-SH concentration during the
reaction between Alb-TNB and CSH (CSH:Alb-TNB 518:1). Peak concentration of Alb-SS-C (maximum at 0.25
min) (250 � 11 lM, n 5 3) was lower than the initial
Alb-TNB concentration (0.415 mM) and peak values
remained constant for 1 min.
Under the same experimental conditions (0.415 mM
Alb-TNB and 7.5 mM of each thiol), kinetic profiles of the
reactions of Alb-TNB with HSH and GSH, separately,
were different from those observed with CSH (Fig. 2).
Although Alb-SS-H and Alb-SS-C reached maximum con-
centrations at 0.25 min, the former had higher concentra-
tions than the latter (Alb-SS-H 5 305 � 12 lM; Alb-SS-C
5 250 � 11 lM; M � SD, n 5 3; P < 0.05; Student’s
t test). The Alb-SS-G peak was lower and later than
peaks of the first two thiols (Alb-SS-C and Alb-SS-H).
Protein Substitution or Dethiolation by SH/SS
DOI 10.1002/prot PROTEINS 371

During dethiolation, concentrations of Alb-SS-C and
Alb-SS-H reached equilibrium at different times (Fig. 2)
and Alb-SS-H concentration was higher than that of Alb-
SS-C. These differences show that CSH is a better dethio-
lating thiol than HSH. Alb-SS-G kinetics (Fig. 2) were dif-
ferent from those of Alb-SS-H and Alb-SS-C, and this dif-
ference was related to lack of equilibrium. Indeed, Alb-SS-
G concentrations continued to decrease beyond 35 min
and were lower at 45 and 60 min (not shown) than at
35 min.
Protein thiolation by XSSX
Protein thiolation of albumin by disulfides is a slow
process (see Introduction). The GSSG excess (7.5 mM)
was unable to form Alb-SS-G after 4 h of exposure to
0.415 mM albumin thiolate anion at room temperature
(data not shown), explaining the nonequilibrium of Alb-
SS-G kinetics in the previous experiment. The rate of Alb-
SS-X formation during thiolation of albumin by CSSC
and HSSH was slower than protein dethiolation by thiols
(data not shown).
Reaction of Alb-TNB with thiol mixtures
When Alb-TNB reacted with the thiol mixture, the sub-
stitution phase was characterized by competition among
thiols. The difference in Alb-SS-X concentration made it
possible to establish the rank of TNB substitution
CSH>HSH>GSH (Fig. 3).
Very complex SH/SS exchange reactions were observed
during Alb-SS-X dethiolation because the three thiols in
excess compete to dethiolate the Alb-SS-Xs formed, pro-
ducing homodimer and heterodimer disulfides, such as
CSSH, CSSG, and HSSG.
The decrease in Alb-SS-C was more marked than those
of Alb-SS-H and Alb-SS-G, the kinetic profiles of which
were less steep (Fig. 3). Alb-SS-G was faster than Alb-SS-
H, unlike in the single thiol experiment. After the first
Alb-SS-H decrease by dethiolation, Alb-SS-H showed an
increasing trend and then a decrease (Fig. 3). Alb-SS-H
Figure 1Time course of reaction of Alb-TNB with CSH. Five hundred microliters of Alb-
TNB (0.830 mM) reacted with 500 lL of CSH (15 mM) at room temperature
and the reaction was stopped by precipitation with 17% TCA (w/v) (final
concentration). Alb-SS-X concentration in the pellet was assayed by HPLC after
mBrB derivatization. After resuspension of the pellet in 400 lL of phosphate
buffer (0.1M), Alb-SH concentration was assayed spectrophotometrically with
DTNB. The figure shows the complete time-course of the experiment. In the first
phase (substitution) there was an increase in Alb-SS-C concentration, and in the
next phase (dethiolation) there was a decrease in Alb-SS-C with a consequent
increase in Alb-SH concentration. The sum of Alb-SS-X and Alb-SH was
constant, confirming dethiolation of Alb-SS-X.
Figure 2Time course of the Alb-TNB reaction with single thiols. Five hundred microliters
of Alb-TNB (0.830 mM) reacted with 500 lL thiol (15 mM) at room
temperature and the reaction was stopped at different times by precipitation
with 17% TCA (w/v) (final concentration). Alb-SS-X concentration was assayed
by HPLC after mBrB derivatization. A: the kinetic profile of Alb-SS-X over the
complete time course is shown. B: the kinetic profile of Alb-SS-X during the
dethiolation phase is shown.
D. Summa et al.
372 PROTEINS DOI 10.1002/prot

was the only Alb-SS-X with this trend, as is most evident
in reaction in which a low concentration of thiol mixture
was used [Fig. 3(B)]. Therefore, the order of percentage
decrease in Alb-SS-X calculated at 35 min with respect
to the Alb-SS-X peak at 0.25 min was Alb-SS-C > Alb-SS-
G>Alb-SS-H, whereas in experiments with single thiols
this order resulted to be Alb-SS-C (K 5 0.747 min21) >
Alb-SS-H (K 5 0.183 min21) > Alb-SS-G (K 5 0.144
min21), as indicated by the corresponding kinetic con-
stants derived by best fitting curves reported in Fig-
ure 2(B).
Dethiolation of Alb-SS-C byHSH (rethiolation)
Commercial albumin containing 200 lM Alb-SS-C was
dethiolated by a relatively low amount of HSH (500 lM),
and Alb-SS-C concentration decreased to zero in 5–10
min. The disappearance of Alb-SS-C occurred at the same
time as the increase in Alb-SS-H concentration. The Alb-
SS-H increase was the consequence of albumin thiolation
by CSSH formed during the dethiolation process (Fig. 4),
as already described by other authors.17 During rethiola-
tion by CSSH, no Alb-SS-C formation was found, as indi-
cated by other authors.17
Bioinformatics
Bioinformatics experiments were carried out to explain
the biochemical data of protein substitution and dethiola-
tion and more exactly to estimate the susceptibility of sul-
fur atoms (in Alb-SS-X) to be subjected to nucleophilic
attack of thiolate anions (as entering group). The sequence
alignment between human and bovine albumin showed a
good match (Fig. 5), with high similarity in structure. The
good stereochemistry of BSA homology model, built with
DeepView software v.3.7,19 was confirmed by using PRO-
CHECK v.3.2.4.22 Indeed the main chain conformations
for 92% of amino acid residues were in the favored or
allowed regions of the Ramachandran plot, as confirmed
by G factor score (20.21), which indicates a good quality
of stereochemical parameters. The homology model of
BSA, built with DeepView software v.3.7,19 was validated
Figure 3Comparison of Alb-SS-X time course in Alb-TNB reaction with a mixture of
HSH, CSH, and GSH. Five hundred microliters of Alb-TNB (0.830 mM)
reacted with 500 lL of thiol mixture 15 mM (A) and 3 mM (B) total at room
temperature. The reaction was stopped at different times by precipitation with
17% TCA (w/v) (final concentration). Alb-SS-X concentration was assayed by
HPLC after mBrB derivatization. In the substitution phase, the three thiols
compete to substitute the TNB on the albumin, and the fastest reached the
highest concentration in mixed disulfide. In both concentrations of the mixture,
the order of substitution was CSH > HSH > GSH. Dethiolation of Alb-SS-X
formed in the first phase occurred in the second phase. The increase in Alb-SS-
H was more visible in B where the mixture was less concentrated.
Figure 4Dethiolation of Alb-SS-C by HSH. BSA (600 lM total, 200 lM Alb-CSH) was
reacted with 500 lM HSH and the reaction stopped by precipitation with 17%
TCA (w/v) (final concentration). After mBrB derivatization, Alb-SS-X
concentration was assayed by HPLC. The figure shows a simultaneous increase
in Alb-SS-H and decrease in Alb-SS-C. This was caused by the dethiolation of
Alb-SS-C and subsequent thiolation of HSSC on Alb-SH.
Protein Substitution or Dethiolation by SH/SS
DOI 10.1002/prot PROTEINS 373

using PROCHECK v.3.2.4.22 The main chain conforma-
tions for 92% of amino acid residues were in the favored
or allowed regions of the Ramachandran plot, and the
overall G factor was 20.21. The next step in the evaluation
is the assessment of the fold, which is restricted here to the
use of energetic profiles introduced by statistical criteria
like the information given by program PROSA II.23 The
Prosa II program was used to define interaction energies
for each residues of the model with the remainder of the
protein. The Prosa II energy profile plot derived for HSA
and BSA structures are shown in Figure 6, with the pre-
dominant area of the profile displaying negative score.
Once we had obtained the BSA structure, we modified the
Cys34 residue in thiolated Cys34 with GSH, HSH, CSH,
and reduced Cys34 using HyperChem v.6., and their pa-
rameters and topologies were obtained using Dundee
PRODRG2 server24 based on GROMACS force-field
gmx43a2.21
These new structures were used to perform a MD simu-
lation with GROMACS v.3.1.4.21 During the MD run we
have checked the stability of simulations and the possible
changes introduced by the different modified residues by
RMSD plot (data not shown).
The average structures obtained from the MD simula-
tion were analyzed with SADIC software25 to obtain the
depth of sulfur involved in the mixed disulfide bond
(Table I). Comparing the depth of the two sulfur atoms
for each mixed disulfide, we observed that of albumin
Figure 5BSA and HSA alignment. ClustalW pairwise alignment of BSA and HSA aminoacid sequences using user-defined parameters of gap open and extension penalties with
BLOSUM62 matrix. The two sequences show 75% of sequence identities and 18% of similarity. The free cysteine is highlighted in grey. It is possible to note the highly
aligned region (*) around free cysteine.
D. Summa et al.
374 PROTEINS DOI 10.1002/prot

(SG) was deeper than that of the other thiols, except Alb-
SS-G. The coefficient rank of atom depths (Table I) did
not explain why TNB, whose S had better exposure (SG2
5 0.965) than that of albumin (SG 5 0.573), was always
substituted, as well as the coefficient rank did not explain
why Alb-SS-G was dethiolated despite the lower exposure
of S atom of GSH (SG2 5 0.265) with respect to that of
albumin (SG 5 0.486).
To evaluate the kinetics of thiolation of albumin Cys34
thiolate anion, we performed two MD simulations using
protonated and neutral His39, on the basis of their ability
to form a salt bridge as suggested by other authors.12
Comparison of the two structures, obtained by simulation,
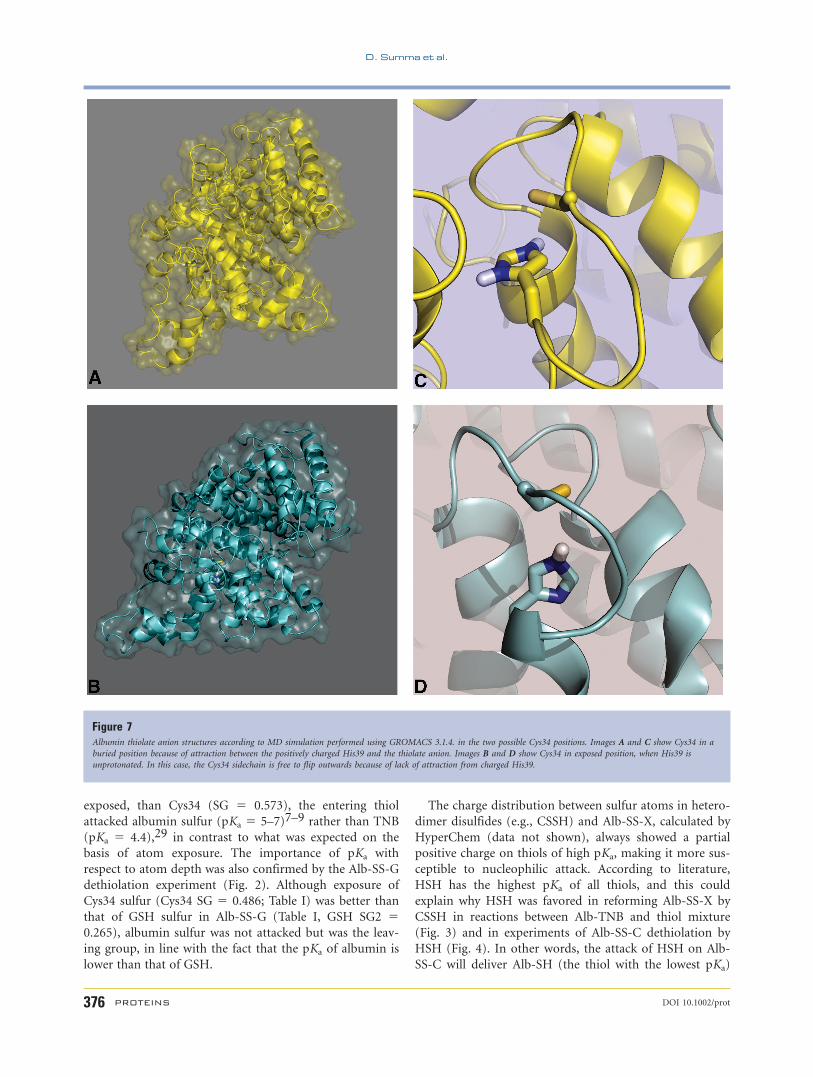
showed that Cys may be buried [Fig. 7(A,C)] or exposed
[Fig. 7(B,D)] depending on whether it formed a salt bridge
with charged histidine. Table II shows the depth of sulfur
obtained by SADIC calculations25 in these two conforma-
tions. In the presence of protonated histidine, the expo-
sure of the Cys34 thiolate anion is practically zero and in
this condition thiolation by XSSX is very low or absent
when GSSG is the thiolating agent.
The partial charge distribution of sulfur engaged in
mixed disulfides, calculated by HyperChem v.6, showed
that the sulfur having lower pKa, according to the litera-
ture, had greater average electronic density.
DISCUSSION
Experiments carried out to understand to what extent
SH/SS exchange reactions generate the normal distribution
of thiol redox species in human plasma have given a new
contribution concerning mechanisms and rates of ex-
change reactions involving albumin. Depending on pKa of
the thiol involved in mixed disulfides and in SH/SS ex-
change reactions, the albumin may be subjected to thiol
substitution (generation of another Alb-SS-X) or dethiola-
tion (formation of Alb-SH).
Albumin has proven to be an appropriate model to an-
swer basic questions about the role of pKa and sulfur ex-
posure in nonenzymatic SH/SS exchange reactions. The
free albumin cysteine has low reactivity because of its spa-
tial position. In addition, Cys34 may be buried, as con-
firmed by the structures obtained from MD simulation
with protonated His39 [Fig. 7(A,C)]. Protonated His39,
located under the protein surface, attracts the thiolate
anion in, reducing its accessibility. Indeed, atom depths of
the two structures showed no exposure of Cys34 in the
presence of protonated His39 (Table II, SG 5 0.000), and
very low exposure with unprotonated His39 (Table II, SG
5 0.005). However, when Cys34 residue is modified into
adducts (Alb-SS-X) there is a change in albumin confor-
mation and a consequent increase in protein reactivity
because of better exposure of the disulfide bridges with
respect to the original Alb-SH, as confirmed by the atom
depth coefficients calculated by SADIC (compare depth
coefficients of sulfur in Tables I and II).
The depth of sulfur in BSA thiolated with different thi-
ols and TNB explains and sustains our experimental
results, which showed that dethiolation was faster than
thiolation.
In the reaction between Alb-TNB and various thiols
(CSH, HSH, and GSH), the pKa range of which was 8.3–
9.5,26–28 TNB substitution was always observed and the
rate rank of substitution (CSH>HSH>GSH) was inversely
related to the mass and charge order as indicated by peak
height in experiments with thiol mixture (Fig. 3).
The difference in pKa values of thiols engaged in mixed
disulfides is more important than atom depth for deter-
mining the nucleophilic attack and leaving groups because
comparison of atom depth coefficients and biochemical
data did not suggest a relationship between sulfur expo-
sure and leaving groups. For example, although the atom
depth coefficient of TNB sulfur engaged in mixed disulfide
(SG2 5 0.965; Table I) was higher, and therefore better
Figure 6Prosa II energy profiles. Differences in Z-scores calculated between the BSA
model and the crystallographic structure (HSA). The values have been obtained
using the PROSA II program. The plot shows a good fit of pseudoenergy for the
two structures. The plot does not show a large increase of pseudoenergy for the
free cysteine region. In x-axis: residue numbers, y-axis: Z-scores (Prosa energy).
Table IAtom Depth Coefficient of the Different Alb-SS-X Calculated Using
SADIC Software
Alb-SS-X
Atom depth
SG SG2
Alb-SS-C 0.518 0.819Alb-SS-G 0.486 0.265Alb-SS-H 0.295 0.664Alb-TNB 0.573 0.965
Cys34 sulfur is indicated as SG, and the sulfur of the thiol bound to protein by
disulfide bond is indicated SG2.
The sulfur that underwent nucleophilic attack is in bold.
Protein Substitution or Dethiolation by SH/SS
DOI 10.1002/prot PROTEINS 375
exposed, than Cys34 (SG 5 0.573), the entering thiol
attacked albumin sulfur (pKa 5 5–7)7–9 rather than TNB
(pKa 5 4.4),29 in contrast to what was expected on the
basis of atom exposure. The importance of pKa with
respect to atom depth was also confirmed by the Alb-SS-G
dethiolation experiment (Fig. 2). Although exposure of
Cys34 sulfur (Cys34 SG 5 0.486; Table I) was better than
that of GSH sulfur in Alb-SS-G (Table I, GSH SG2 50.265), albumin sulfur was not attacked but was the leav-
ing group, in line with the fact that the pKa of albumin is
lower than that of GSH.
The charge distribution between sulfur atoms in hetero-
dimer disulfides (e.g., CSSH) and Alb-SS-X, calculated by
HyperChem (data not shown), always showed a partial
positive charge on thiols of high pKa, making it more sus-
ceptible to nucleophilic attack. According to literature,
HSH has the highest pKa of all thiols, and this could
explain why HSH was favored in reforming Alb-SS-X by
CSSH in reactions between Alb-TNB and thiol mixture
(Fig. 3) and in experiments of Alb-SS-C dethiolation by
HSH (Fig. 4). In other words, the attack of HSH on Alb-
SS-C will deliver Alb-SH (the thiol with the lowest pKa)
Figure 7Albumin thiolate anion structures according to MD simulation performed using GROMACS 3.1.4. in the two possible Cys34 positions. Images A and C show Cys34 in a
buried position because of attraction between the positively charged His39 and the thiolate anion. Images B and D show Cys34 in exposed position, when His39 is
unprotonated. In this case, the Cys34 sidechain is free to flip outwards because of lack of attraction from charged His39.
D. Summa et al.
376 PROTEINS DOI 10.1002/prot

and CSSH. In turn, CSSH may be subjected to the attack
by the thiolate anion of BSA,17 as we also found. All these
concepts relative to protein substitution and dethiolation
are summarized in Figure 8.
Another notion not in contrast with the previous one
emerged in the present study. During Alb-SS-X dethiola-
tion by excesses of thiol, the sulfur of Alb-SS-X having the
best exposure exhibited the highest dethiolation rate. In
fact, a direct relationship between dethiolation rate,
deduced from comparison of kinetic traces and atom ex-
posure of sulfur undergoing nucleophilic attack was
found. Thus, the increasing order of atom depth coeffi-
cients (CSH>HSH>GSH, SG2 in Table I) was related to
the increasing order of dethiolation rate (Alb-SS-C>Alb-
SS-H>Alb-SS-G) (Fig. 2), and for example, CSH, the faster
dethiolating agent showed the highest atom exposure
(SG2 5 0.819; Table I).
The results of the present studies explain the possible
contribution of SH/SS exchange reactions to determine
levels of thiol redox species in plasma. For example, differ-
ences in HSH and GSH concentrations in human plasma,
with ratios of reduced/total thiols of 0.02 HSH/tHcy and
0.3 GSH/tGSH, respectively,13 can not be linked to oxida-
tion rates, which are nearly equal in plasma,15 but more
plausibly to activation of SH/SS exchange reactions. The
comparison of dethiolation processes from peaks of Alb-
SS-H and Alb-SS-G (Fig. 2) emphasized the possible role
exerted by these reactions. Since GSH is a bad dethiolating
agent compared to HSH, it does not detach Alb-SS-X with
the same efficiency as HSH, and its plasma concentration
is destined to be higher than HSH, as indicated by corre-
sponding plasma levels of normal individuals.13,14,16 In a
more complex situation, such as plasma, GSH concentra-
tions would therefore be less engaged than HSH in dethio-
lating Alb-SS-X.
CONCLUSION
This study demonstrated that the albumin may be sub-
jected to different metabolic pathways via thiol substitu-
tion or protein dethiolation, by virtue of pKa of thiols
involved in albumin-thiol mixed disulfides. The nucleo-
philic reaction of the thiolate anion of the entering group
attacks the mixed disulfide at level of the more positive
sulfur, namely that with higher pKa, whereas the thiol of
the mixed disulfide with the lower pKa is released from the
intermediate ternary complex.
This study has also evidenced that the rate magnitude
of dethiolation process, on albumin, will depend on the
exposure of the more positive sulfur engaged as Alb-SS-X
and features of mass and charge of the entering group.
These results confirmed that SH/SS exchange reactions
have a relevant role in changing the redox state of thiols
elicited by plasma oxidations in living organisms.
REFERENCES
1. Di Simplicio P, Frosali S, Priora R, Summa D, Di Simplicio FC, Di
Giuseppe D, Di Stefano A. Biochemical and biological aspects of
protein thiolation in cells and plasma. Antioxid Redox Signal
2005;7:951–963.
2. Kim SO, Merchant K, Nudelman R, Beyer WF, Jr, Keng T, DeAn-
gelo J, Hausladen A, Stamler JS. OxyR: a molecular code for redox-
related signaling. Cell 2002;109:383–396.
3. Klatt P, Lamas S. Regulation of protein function by S-glutathiola-
tion in response to oxidative and nitrosative stress. Eur J Biochem
2000;267:4928–4944.
4. Ward NE, Stewart JR, Ioannides CG, O’Brian CA. Oxidant-induced
S-glutathiolation inactivates protein kinase C-a PKC-a: a potential
mechanism of PKC isozyme regulation. Biochemistry 2000;39:
10319–10329.
5. Hurd TR, Filipovska A, Costa NJ, Dahm CC, Murphy MP. Disul-
phide formation on mitochondrial protein thiols. Biochem Soc
Trans 2005;33:1390–1393.
Table IIAtom Depth Coefficient Obtained From the Structure of Albumin With
Protonated and Unprotonated His39
pH
Atom depth Res depth
SG ND1 NE2 CYS HIS
XlbHIS 0.005 0.055 0.033 1.439 0.763AlbHIS1 0.000 0.007 0.002 0.771 0.265
The depth coefficient of Cys34 sulfur is indicated SG, and the depth coefficient of
the complete amino acid (Res) is indicated CYS and HIS.
ND1 and NE2 are the His63 nitrogens.
Figure 8Scheme of reactions during the Alb-TNB reaction with excess thiol. A:
substitution reaction of TNB on albumin and consequent formation of Alb-SS-
X. Low molecular weight thiolate anion nucleophilically attacks the positively
charged albumin sulfur and TNB leaves the system as a thiolate anion due to its
low pKa (4.4) relative to albumin (pKa 5–7). B: the excess thiol nucleophilically
attacks the positively charged thiol sulfur of Alb-SS-X, dethiolating the albumin
that leaves the system as an albumin thiolate anion. The leaving group is the
albumin, as it has a lower pKa than thiols (range 8.3–9.2). Positively charged
sulfur has a higher pKa in the mixed disulfide.
Protein Substitution or Dethiolation by SH/SS
DOI 10.1002/prot PROTEINS 377

6. Reddy S, Jones AD, Cross CE, Wong PS-Y, Van Der Vliet A. Inacti-
vation of creatine kinase by S-glutathionylation of the active-site
cysteine residue. Biochem J 2000;347:821–827.
7. Lewis SD, Misra DC, Shafer JA. Determination of interactive thiol
ionizations in bovine serum albumin, glutathione, and other thiols
by potentiometric difference titration. Biochemistry 1980;19:6129–
6137.
8. Narazaki R, Hamada M, Harada K, Otagiri M. Covalent binding
between bucillamine derivatives and human serum albumin. Pharm
Res 1996;13:1317–1321.
9. Pedersen AO, Jacobsen J. Reactivity of the thiol group in human
and bovine albumin at pH 3–9, as measured by exchange with 2,20-dithiodipyridine. Eur J Biochem 1980;106:291–295.
10. Di Simplicio P, Ignesti G, Pirisino R. Relationship between SH
group reactivity and concentration of bovine serum albumin and
rat plasma. Pharmacol Res Commun 1985;17:913–925.
11. Gryzunov YA, Arroyo A, Vigne JL, Zhao Q, Tyurin VA, Hubel CA,
Gandley RE, Vladimirov YA, Taylor RN, Kagan VE. Binding of fatty
acids facilitates oxidation of cysteine-34 and converts copper-albu-
min complexes from antioxidants to prooxidants. Arch Biochem
Biophys 2003;413:53–66.
12. Christodoulou J, Sadler PJ, Tucker A. 1H NMR of albumin in
human blood plasma: drug binding and redox reactions at Cys34.
FEBS letter 1995;376:1–5.
13. Di Giuseppe D, Frosali S, Priora R, Di Simplicio Cherubini F, Buo-
nocore G, Cellesi C, Capecchi PL, Pasini F, Lazzerini PE, Jakubow-
ski H, Di Simplicio P. The effects of age and hyperhomocysteinemia
on the redox forms of plasma thiols. J Lab Clin Med 2004;144:235–
245.
14. Ueland PM, Mansoor MA, Guttormsen AB, Muller F, Aukrust P,
Refsum H, Svardal AM. Reduced, oxidized and protein-bound
forms of homocysteine and other aminothiols in plasma comprise
the redox thiol status—a possible element of the extracellular anti-
oxidant defense system. J Nutr 1996;126:1281S–1284S.
15. Sengupta S, Wehbe C, Majors AK, Ketterer ME, DiBello PM, Jacob-
sen DW. Relative roles of albumin and ceruloplasmin in the forma-
tion of homocystine, homocysteine-cysteine-mixed disulfide, and
cystine in circulation. J Biol Chem 2001;276:46896–46904.
16. Mansoor MA, Svardal AM, Schneede J, Ueland PM. Dynamic rela-
tion between reduced, oxidized, and protein-bound homocysteine
and other thiol components in plasma during methionine loading
in healthy men. Clin Chem 1992;38:1316–1321.
17. Sengupta S, Chen H, Togawa T, DiBello P, Majors AL, Budy B, Ket-
terer ME, Jacobsen DW. Albumin thiolate anion is an intermediate
in the formation of albumin-S-S-homocysteine. J Biol Chem 2001;
276:30111–30117.
18. Ellman CL. Tissue sulfhydryl groups. Arch Biochem Biophys 1959;
82:70–77.
19. Guex N, Peitsch MC. SWISS-MODEL and the Swiss-PdbViewer: an
environment for comparative protein modeling. Electrophoresis
1997;18:2714–2723.
20. Thompson JD, Higgins DG, Gibson TJ. Improving the sensitivity of
progressive multiple sequence alignment through sequence weight-
ing, position-specific gap penalties and weight matrix choice.
Nucleic Acids Res 1994;22:4673–4680.
21. Van Der Spoel D, Lindahl E, Hess B, Groenhof G, Mark AE,
Berendsen HJ. GROMACS: fast, flexible, and free. J Comput Chem
2005;26:1701–1718.
22. Laskowski RA, MacArthur MW, Moss DS, Thornton JM. PRO-
CHECK: a program to check the stereochemical quality of protein
structures J Appl Cryst 1993;26:283–291.
23. Sippl MJ. Recognition of errors in three-dimensional structures of
proteins. Proteins 1993;17:355–362.
24. Schuettelkopf W, van Aalten DMF. PRODRG—a tool for high-
throughput crystallography of protein-ligand complexes. Acta Crys-
tallogr 2004;60:1355–1363.
25. Varrazzo D, Bernini A, Spiga O, Ciutti A, Chiellini S, Venditti V,
Bracci L, Niccolai N. Three-dimensional computation of atom
depth in complex molecular structures. Bioinformatics 2005;21:
2856–60.
26. Benesch RE, Benesch R. The acid strength of the -SH group in cys-
teine and related compounds. J Am Chem Soc 1955;77:587–5881.
27. Friedman M, Cavins JF, Wall JS. Relative nucleophilic reactivities of
amino groups and mercaptide ions in addition reactions with a,b-
unsaturated compounds. J Am Chem Soc 1965;87:3672–3682.
28. Winterbourne CC, Metodiewa D. Reactivity of biologically impor-
tant thiol compounds with superoxide and hydrogen peroxide. Free
Radic Biol Med 1999;27:322–328.
29. Torchinsky YM. Sulfur in proteins. New York: Pergamon Press;
1981. p 59.
D. Summa et al.
378 PROTEINS DOI 10.1002/prot




















